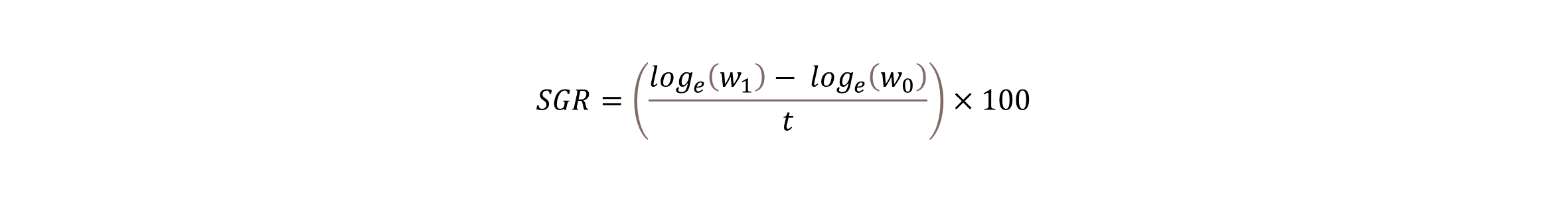Effekt på dyrevelferd av gjentatte termiske behandlinger med ulik Δt, ulik utgangstemperatur og ulik behandlingstemperatur
—
FHF - 901646 - SLUTTRAPPORT (L2.2)
Author(s):
Lars Helge Stien
, Samantha Bui
, Jonatan Nilsson
, Angelico Madaro
, Per Gunnar Fjelldal
(IMR), Merete Bjørgan Schrøder (Sintef Ocean AS), Birger Venås (Sintef Ocean AS), Malin Johansen (NCE Aquaculture), Tore Seternes (Universitetet i Tromsø), Caroline Piercey Åkesson (Pharmaq Analytiq AS), Helene Wisløff (Pharmaq Analytiq AS), Monica Fengsrud Brinchmann (Nord universitet), Deepti Manjari Patel (Nord universitet) and Martin Haugmo Iversen (Nord universitet)
Rapportserie:
Rapport fra havforskningen 2022-39ISSN: 1893-4536Publisert: 21.11.2022Prosjektnr: 15687Oppdragsgiver(e): Fiskeri- og havbruksnæringens forskningsfinansiering (FHF)Referanse: 901649 - Objektiv dokumentasjon og beste praksis for å forbedre termisk avlusing (TermVel) Forskningsgruppe(r):
DyrevelferdTema:
FiskevelferdProgram:
Fremtidens havbruk Approved by:
Research Director(s):
Geir Lasse Taranger
Program leader(s):
Rune Waagbø
Forord
Dette er sluttrapporten i arbeidspakke 2 av FHF-prosjektet «Objektiv dokumentasjon og beste praksis for å forbedre termisk avlusing (TermVel)». Denne arbeidspakken bestod av to laboratorieforsøk som ble gjennomført ved henholdsvis forskningsstasjon Matre nær Bergen i regi av Havforskingsinstituttet og ved Mørkvedbukta forskningsstasjon nær Bodø i regi av Nord universitet.
Disse forsøkene ble gjennomført med god assistanse og support ved de ovennevnte forsøksstasjonene, og forfatterne ønsker rette en stor takk til de som deltok i forsøkene.
Framdriftsforslag og gode tilbakemeldinger ble gitt av FHF sin referansegruppe og vi ønsker å takke: Remi Mathisen (Nordlaks Oppdrett AS), Kristin Ottesen (haVet AS), Tor Hugo Hestnes (LetSea AS), Tiril Slettjord (Cermaq Norway AS) og Kjell Maroni (FHF).
Sammendrag
Formålet med forsøkene i arbeidspakke 2 i TermVel-prosjektet var å avdekke effekt på dyrevelferd av gjentatt termisk behandling av laks gitt ulik utgangstemperatur, ulik behandlingstemperatur og ulik Δt (differanse mellom behandlingstemperatur og utgangstemperatur). Dette inkluderte å studere atferds-respons under behandling, skader etter behandling, fysiologiske reaksjoner og eventuelle langtidseffekter av behandling.
Arbeidspakken bestod av to parallelle studier, der det ene studiet ble utført på Havforskningsinstituttet sin forskningsstasjon i Matre og det andre på Nord universitet sin forskningsstasjon i Mørkvedbukta. Begge studiene benyttet relativt stor laks (1-2 kg) som først ble individmerket og deretter plassert i forsøkskarene i et såkalt «common-garden»-oppsett. Vanntemperaturen i forsøkskarene var 14 °C i forsøket i Matre, mens vanntemperaturen var 8 °C i forsøket ved Mørkvedbukta forskningsstasjon. To til tre uker etter merking og overføring til forsøkskarene ble fiskene sortert ut basert på individmerkene og behandlet på enten samme temperatur som i karene, eller eksponert til vann på enten 27, 30 eller 33 °C i 30 sekunder. Denne eksponeringen ble så gjentatt etter 1 måned.
Forsøkene viste at:
en av laksegruppene hadde økt dødelighet som funksjon av behandlingstemperatur.
laksen reagerte på å bli eksponert for 27 °C også ved høy utgangstemperatur (lav Δt).
graden av panikkatferd økte med økende behandlingstemperatur
i tillegg til panikkatferd kan laks også reagere på varmt vann ved å legge seg over på siden, og tilsynelatende «gi opp».
laksen responderte med «gi opp»-atferd i større grad ved 30 °C enn ved 33 °C.
eksponering til varmt vann gir risiko for øyeskader.
det ble ikke funnet noe som tyder på at eksponering til 27-33 °C i 30 sekunder gir laks langvarige vevsskader i hjertet.
I studiet i Matre var det dødelighet etter den første behandlingen, med økende dødelighet i forhold til behandlingstemperatur. Denne fiskegruppen hadde hatt relativ kort tid (to uker) til å restituere seg etter at den hadde blitt merket med utvendige og innvendige ID-merker og overført til forsøkskarene. Ved den andre eksponeringen, da fiskegruppen hadde fått restituert seg i fire uker etter første behandling, og i forsøket i Bodø, der fiskegruppen fikk lenger tid (tre uker) til å restituere seg etter merkingen, var det ikke dødelighet knyttet til behandlingene. Dette tyder på at frisk, restituert fisk, overlever eksponering til varmt vann i 30 sekunder, men at fisk som ikke er restituert, eller har andre iboende svakheter, har økende risiko for dødelighet ved økt temperatur på termisk behandling. Det er ikke avklart om skadene som ble observert på øynene til laksen etter behandling ble direkte forårsaket av det varme vannet, indirekte via panikkresponsen, eller en kombinasjon der det varme vannet svekker øynene, mens panikkresponsen førte til selve skaden. Basert på disse resultatene anbefaler vi at det utføres oppfølgingsprosjekter og studier for å avdekke:
«helsemarkører» som kan brukes av oppdrettere og fiskehelsepersonell, for å avgjøre om en fiskegruppe er egnet for termisk behandling eller ikke.
hva som avgjør om laks, reagerer med «gi opp»-atferd til termisk behandling og om denne atferden fører til mindre skader.
Hvorfor laksens øyne synes å være utsatt for skade ved termisk behandling, hvordan disse skadene påvirker fisken, og hva som eventuelt kan gjøres for å forhindre skade på øynene.
1. Innledning
Kampen mot lakselus har betydelige økonomiske og dyrevelferdsmessige konsekvenser for oppdrettsnæringen (Grefsrud mfl. 2022). Oppdretterne må følge strenge regler for å forhindre at infeksjonsnivåer blir for høye og avluse laksen før grensenivåene på 0,2 – 0,5 voksne hunnlus nås (se Forskrift om lakselusbekjempelse). Tidligere ble laksen typisk avluset ved hjelp av ulike kjemiske preparater, men pga. resistensutvikling og miljøhensyn har termisk avlusing blitt den dominerende avlusingsmetoden i industrien de siste årene (Grefsrud mfl. 2022).
Termisk avlusing består av at fisken først blir trengt og pumpet opp fra merden, eksponert for varmt sjøvann (28-34 °C) i opptil 30 sekunder, og deretter returnert til merden via vannrør og vannsklier. Termisk avlusing innebærer ingen bruk av kjemiske midler og det er derfor ingen risiko for skade av nærmiljøet. Termisk avlusing er imidlertid forbundet med fysiske skader og forhøyet dødelighet på den behandlete fisken (Grefsrud mfl. 2022). Det har også blitt vist at laks responderer med panikklignende atferd hvis de blir tatt fra 8 °C og eksponert til vann på 28 °C og høyere (Nilsson mfl. 2019), og resultater fra kontrollerte studier tyder på at det først og fremst er denne atferden som forårsaker fysiske skader på fisken og redusert velferd i etterkant av termisk behandling (Moltumyr mfl. 2021, 2002).
Det har imidlertid vært flere kritiske stemmer mot disse studiene. Noe av kritikken som har blitt reist er at fisken har vært for liten (ca. 230g i Nilsson mfl. 2019) og at forskjellen mellom temperaturen i karene (utgangstemperaturen) og behandlingstemperaturen har vært for høy for å være representativ for termisk avlusing i næringen. Retningslinjene til næringen anbefaler f.eks. en Δt (differanse mellom utgangstemperatur og behandlingstemperatur) på 20 til 22,5 °C (Nygård mfl. 2020), mot Δt = 25 °C (9 vs. 34°C) i Moltumyr mfl. (2021, 2022).
Termisk avlusing fjerner ikke de fastsittende stadiene av lakselus, bare de mobile (Brunsvik 1997, Roth 2016). Oppdretterne må derfor ofte avluse laksen sin med få ukers mellomrom (Grefsrud mfl. 2022). Basert på dette, og den overnevnte kritikken, var formålet med forsøkene i arbeidspakke 2 i TermVel-prosjektet å avdekke effekt på dyrevelferd av gjentatt termisk behandling av mellomstor laks (1-2 kg) gitt ulik utgangstemperatur, ulik behandlingstemperatur og ulik Δt. Dette inkluderte å studere atferdsrespons under behandling, skader etter behandling, fysiologiske reaksjoner og eventuelle langtidseffekter av behandling på fiskevelferd. Arbeidspakken bestod av to parallelle studier, der forsøk 1 ble utført på Havforskningsinstituttet sin forskningsstasjon i Matre og forsøk 2 på Nord universitet sin forskningsstasjon i Mørkvedbukta. I forsøk 1 var utgangstemperaturen i forsøkskarene 14 °C, mens den var 8 °C i forsøk 2.
2. Materiale og Metode
2.1 Forsøk 1, utgangstemperatur 14 °C
Forsøket ble utført på Havforskningsinstituttet sin forskningsstasjon på Matre fra 22. mars til 14. juni 2021. Forsøket var planlagt med 3 behandlinger (figur 1), men på grunn av høy dødelighet etter første behandling, ble det vurdert at det ikke var velferdsmessig forsvarlig å gjennomføre en ny behandling allerede to uker etterpå. I samråd med forsøksdyrsansvarlig på stasjonen ble det derfor bestemt å avlyse denne behandlingen, men å gjennomføre behandlingen etter 6 uker som planlagt. Det gikk derfor fire uker mellom første og andre behandling, istedenfor 2 som opprinnelig planlagt.
Figur 1: Tidslinje for forsøk 1. Fisken ble merket, målt og fordelt på forsøkskar i uke 0. Behandling etterfulgt av subsampling for fysiologi var planlagt etter 2, 4 og 6 uker, sluttsampling etter 7 uker, og deretter gikk en mindre gruppe fisk videre i ytterligere 5 uker for å observeres for mulige langtidseffekter på vekst og velferd. På grunn av uventet høy dødelighet etter første behandling ble det bestemt å ikke gjennomføre behandlingen i uke 4, det ble derfor bare gjennomført to behandlinger i forsøket, behandling B1 og behandling B2.
To uker før første behandling ble laks (~2 kg, Salmo salar) overført fra et produksjonskar, merket, målt, veid, skåret for velferd og fordelt på fem forsøkskar (Ø = 3 m, ~5,3 m3, 14 °C, figur 2).
Figur 2. Forskningsteknikere Simon Flavell og Tone Knappskog (foran fra venstre) og forsker Samantha Bui (i bakgrunnen) forbereder en ny ladning av laks for merking, måling og fordeling til forsøkskar.
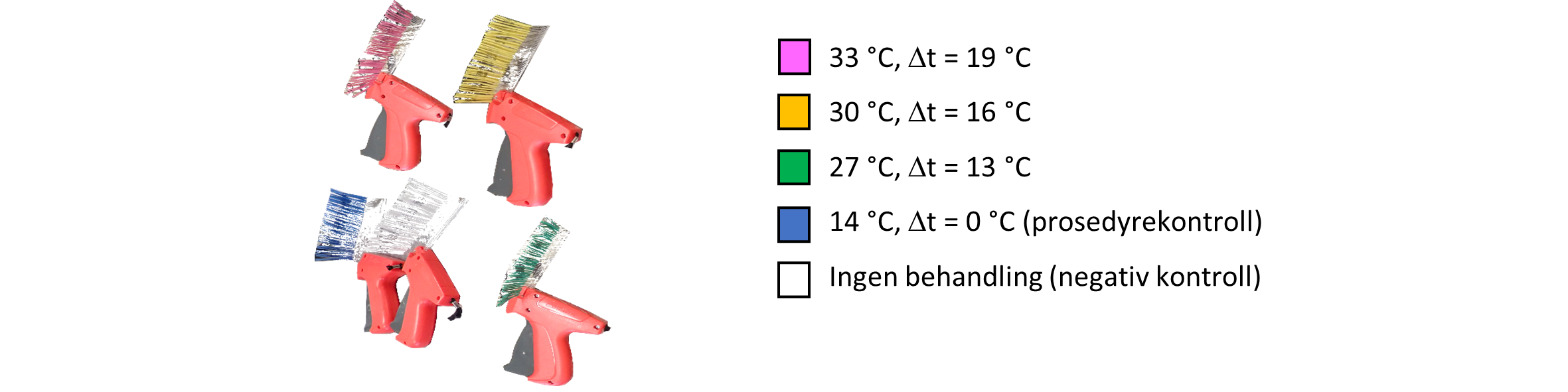
Merking og fordeling i kar ble utført blokkrandomisert for å unngå mulig bias fra at fisk overført tidlig var av en annen størrelse eller helsetilstand enn fisk som ble hentet senere fra produksjonskaret. Fisken ble merket med enten hvite (negativ kontroll), blå (prosedyrekontroll), grønne (27 °C-gruppe), gule (30 °C-gruppe) eller rosa (33 °C-gruppe) Floy T-bar merker (figur 3), i tillegg ble det satt inn PIT-merker i buken i tilfelle tap av de utvendige merkene.
Fiskene som fikk hvitt Floy-merke (120 stk.) ble overført til et eget kar, mens de andre gruppene ble fordelt likt i de fire andre karene (30 fisk fra hver gruppe) slik at det til sammen var 120 fisk i hvert kar (30 fisk/gruppe * 4 grupper = 120 fisk per kar). Ved å fordele behandlingsgruppene likt i fire kar unnvikes eventuelle bias fra at et kar tilfeldigvis skulle være mer eller mindre gunstig for fisken enn andre. Det er imidlertid en risiko for kareffekt hos negativkontrollgruppen siden disse alle var i samme kar. Gjennom hele forsøksperioden ble fisken fôret etter et standard fôrregime og holdt med kontinuerlig belysning (24L:0M). Dødelighet og atferd ble overvåket som del av standard røkteprosedyre i forbindelse med daglig sjekk av kar og overvåking av fôring.
Figur 3. Pistoler med T-bar merker (Floy-tag Inc, Seattle, USA) med ulik farge. Der rosa indikerer at denne fisken skulle behandles på 33 °C, oransje 30 °C, grønn 27 °C og blå 14 °C (samme som i forsøkskar, dvs. prosedyrekontroll). Hvit indikerer gruppen som skulle stå urørt i eget forsøkskar og ikke behandles (negativ kontroll).
To uker etter at fisken var fordelt i forsøkskarene ble den første behandlingen gjennomført. For hvert kar ble fisken først håvet og sortert i separate oppbevaringskar i henhold til deres behandlingsgruppe (figur 4). Fire fisk om gangen fra hver gruppe ble deretter eksponert enkeltvis for behandlingstemperaturvann i 30 sekunder i individuelle behandlingskammer. Etter to runder ved en temperatur ble vannet skiftet ut, og fisk ble tatt fra neste oppbevaringskar. Ved å rotere behandlingen på denne måten ble det samme vannet kun brukt på to fisk før det ble fornyet. Temperatur, oksygenmetning og totalt gasstrykk ble kontrollert og justert fortløpende. To av behandlingskamrene ble filmet fra utsiden under eksponeringene for å dokumentere fiskenes atferd i de ulike vanntemperaturene.
Figur 4. Fisk fra hvert forsøkskar ble først fordelt på 4 oppbevaringskar basert på fargen på det utvendige T-bar merket, deretter ble de enkeltvis håvet videre til behandling på enten 14, 27, 30 eller 33 °C før de ble returnert tilbake til det opprinnelige forsøkskaret.
Tjuefire timer etter første og andre behandling ble 8-12 fisk fra hver behandlingsgruppe (ikke fra negativ kontroll) avlivet med en overdose bedøvelsesmiddel. Fisken ble veid og målt, og det ble tatt blodprøver. Ved prøvetakingen etter andre behandling ble fisken i tillegg røntgenfotografert for å avdekke om det var gassoppbygging bak noen av øynene. All fisk, inkludert negative kontroller, ble målt, veid og skåret for utvendige sår og skader 5-6 dager etter andre behandling (prøvetaking uke 7, figur 1), for å dokumentere eventuelle korttidseffekter av behandlingene. I tillegg ble det tatt ut vevsprøver av gjeller, hjerte, hud, muskulatur, nyrer, hjerne og øyne fra seks fisk fra hver gruppe og sendt til histopatologisk undersøkelse. For å overvåke eventuelle langtidseffekter mellom gruppene ble 50 fisk fra hver gruppe omfordelt likt i 3 kar (~80 per kar), for å holdes i ytterligere 4 uker (figur 1). I denne perioden ble de fôret og røktet etter standard prosedyre, og ikke håndtert på noe vis. På slutten av denne perioden ble all fisk på nytt veid og målt og skåret for ytre skader.
2.2 Forsøk 2, utgangstemperatur 8 °C
Forsøket ble utført på Nord Universitet sin forskningsstasjon i Mørkvedbukta fra 28. september til 20. januar 2022 (figur 5).
Figur 5. Tidslinje for forsøk 2. Fisken ble merket, målt og fordelt på kar i uke 0. Behandling etterfulgt av subsampling for fysiologi og velferd var planlagt etter 3, og 8 uker, sluttsampling etter 9 uker, og deretter gikk en mindre gruppe fisk videre i ytterligere 7 uker for å observeres for mulige langtidseffekter på vekst og velferd.
Tre uker før første behandlingen ble laksen (~1 kg) overført fra et produksjonskar, merket på samme måte som i forsøk 1 (se over), målt, veid, skåret for ytre sår og skader og fordelt på fire forsøkskar (Ø = 3 m, ~8,0 m3, 8 °C). Negativkontrollgruppen ble overført til et eget kar, mens de andre gruppene ble fordelt likt i de fire andre karene (25 fisk fra hver gruppe) slik at det til sammen var 100 fisk i hvert kar.
Tre uker etter at fisken var fordelt i forsøkskarene ble den første behandlingen gjennomført. For hvert kar ble fisken først håvet og sortert ifølge farge på Floy-merke i fire oppbevaringskar (1 m3) med samme temperatur som i forsøkskarene. Deretter ble, en og en fisk håvet til behandlingskammeret, holdt der i 30 sekunder, og deretter returnert til det opprinnelige forsøkskaret. Vann i behandlingskamrene ble byttet mellom hver fisk. Alle behandlingskamrene ble filmet ovenfra for å overvåke laksens atferd ved de ulike temperaturene. Etter erfaring fra forsøk 1 var det et ønske å se om fisk på individ-nivå endret adferd fra første til andre termiske behandling. All fisk ble derfor individregistret inn ved hver behandling, slik at hver film fikk en ID som stemte overens med fiskens ID.
Figur 6. Behandlingskamrene som ble benyttet i forsøk 2.
Tjuefire timer etter hver behandling ble 12 fisk fra hver behandlingsgruppe (ikke fra negativ kontroll) avlivet, veid og målt, i tillegg til at det ble tatt blod- og vevsprøver.
All fisk, inkludert negative kontroller, ble undersøkt 5-6 dager etter den andre behandlingen (prøvetaking uke 9, figur 5), for å bestemme korttidseffektene av de termiske behandlingene.
Som et tillegg til forsøket, ble 25 fisk per behandlingsgruppe omfordelt likt i to vanlige kar, for å holdes i ytterligere 8 uker. Formålet var å avdekke eventuelle langtidseffekter av behandlingene. Ved slutten av denne perioden ble all fisk på nytt tatt prøver av (prøvetaking uke 16, figur 5).
2.3 Måling av vekt og lengde og skåring av ytre skader
Kroppsstørrelsen ble registrert ved gaffellengde (cm) og vekt (g). Skåringen av ytre skader ble gjort ved hjelp av LAKSVEL-skåringssystemet, som inkluderer indikatorer for hudstatus (sår, blødninger, skjelltap), hodeskade (snute, gane), øyestatus (skade, blødning, blakking), og finnestatus (hale, rygg-, bryst-, buk- og analfinner) (Nilsson mfl. 2022). I dette skåringssystemet blir hver indikator skåret fra 0-3, der 0 = sunn uskadet og avviksfri, 1 = mindre skade eller avvik, 2 = tydelig skade eller avvik med moderat betydning for fisken og 3 = alvorlig skade eller avvik som antas å ha store konsekvenser for fisken. Spesifikk vekstrate (SGR) ble beregnet etter følgende formel:
hvor w1 er sluttvekt i g, w0 er startvekt i g og t er antall dager.
2.4 Atferdsvurdering
Mens de var i det termiske behandlingsbadet ble 2 kar filmet samtidig fra utsiden for å dokumentere atferd under eksponering (forsøk 1) eller i hvert behandlingskar ovenfra (forsøk 2). Atferden ble inndelt i kategoriene «plasking», «langsomme bevegelser», «stillestående», og «på siden» (tabell 1). For hvert individ ble den dominerende atferden registrert for hvert femte-sekund i løpet av behandlingstiden på 30 sekunder. En valgte å se bort fra den første delen av de første fem-sekundene etter at fisken var satt ned i behandlingskamrene, for å unngå å registrere den umiddelbare fluktatferden fra å ha blitt håvet oppi behandlingskamrene.
Tabell 1. Kategoriseringsgrupper av atferd til fisken i behandlingsgruppene
Kategori
Beskrivelse
Plasking
Eksplosiv aktiv atferd med mye plasking, hopping, buktning, m.m.
Noe bevegelse
Små hale-bevegelser, snur seg rolig rundt i kammeret.
Ikke bevegelse
Fisken står stille, holder seg oppreist, men nesten ingen romlig forflyttelse.
På siden
Har mistet likevekten, ligger på siden tilsynelatende bevisstløs.
2.5 Vevsprøver og histologi
For å undersøke for mulige skader eller sykdommer, ble det tatt vevsprøver fra organer ved sluttsampling (ca. 1 uke etter andre behandling) hvor små vevsprøver ble tatt fra gjeller, hjerte, hud, muskulatur, nyre, hjerne og begge øyne. Innen 2 minutter etter anestesi ble vevsprøvene fiksert i en 10 % fosfatbufret formalinløsning for senere histopatologiske undersøkelser. Gjellevev ble tatt fra den andre gjellebuen. Hud- og skjelettmuskelprøver ble tatt ved tverrsnitt i sidelinjeområdet ved ryggfinnen, og inkluderte både rødt og hvitt muskelvev. Nyreprøver ble tatt fra midtnyren. Vevsprøvene ble lagret ved 4 °C inntil de ble prosessert og analysert hos Pharmaq Analytiq AS. En generell diagnostisk oppsummering ble gitt, og alle vevsprøver ble skåret 0 – 3, der 0 = ingen spesifikke funn, 0,5 = minimale funn, 1 = mild endring, 2 = moderat endring og 3 = alvorlig eller uttalt endring.
2.6 Vevsprøver av hjerte i forsøk 2
For å undersøke for eventuelle senskader på hjertet ble det i forsøk 2 også tatt vevsprøver fra hjerte 2 måneder etter siste behandling (ekstra sampling, figur 5,) av tolv fisk fra hver av gruppene. Innen 2 minutter etter anestesi ble vevsprøvene fiksert i en 10 % fosfatbufret formalinløsning for senere histopatologiske undersøkelser. Snitt av ventrikkel ble HE-farget og undersøkt for degenerasjon av muskelvevet i stratum compactum som vist i Poppe, mfl. (2021ab). Snittene ble undersøkt blindet, og mulig degenerasjon ble registrert som pluss (mulig degenerasjon) eller minus (ingen degenerasjon).
2.7 Røntgen av hode i forsøk 1
Hodene til fiskene etter behandling 2 i forsøk 1 ble røntgenfotografert for å undersøke om varmtvannet hadde ført til gassansamling bak øynene. Røntgenbildene ble tatt direkte ovenfra (Canon CXDI-410C Wireless, CANON, INC, Japan) ved hjelp av en bærbar røntgen-enhet (Hiray Plus, Model Porta 100 HF, JOB Corporation, Japan) med en avstand på 88 cm, 40 kV og 4 mAs.
2.8 Fysiologisk respons
For å undersøke hvordan termiske behandlinger påvirker konsentrasjoner av blodplasmaparametere, tok vi prøver med hepariniserte tuberkulinsprøyter, 18-24 timer etter hver behandling for å sammenligne nivåer mellom grupper. Vi fokuserte på parametere som mest sannsynlig ikke ville endre seg innenfor den korte prøvetakingstiden. I dataene fra forsøk 1 ble kortisol ekskludert pga. forsinkelser i prøvetakingen som påvirket verdiene. Dette ble justert i forsøk 2, hvor en sederte fiskene med 5 mg/L vann Aqui-S Vet (MSD Animal Health Norgem AS) før prøvetaking. Blodprøver ble tatt fra fisken umiddelbart etter inntredelse av bevisstløshet. Helblod ble plassert på is inntil sentrifugering ved 5000 g i 5 minutter. Plasmasupernatanten ble overført til nye rør og lagret ved -80 °C for senere analyser. Plasma-osmolalitet ble målt ved frysepunktbestemmelse i 20 μl delprøver med et Osmo Pro-Multi-prøvemikroosmometer (Advanced Instruments). Konsentrasjonen av plasma K+, Na+, Cl−, Ca2+ ioner, pH og glukose- og laktatmetabolitter ble analysert ved bruk av en ABL90 FLEX blodgassanalysator (Radiometer Medical ApS, Danmark). Magnesium (Mg2+) analyse ble utført ved bruk av et Fluitest Mg-XB analysesett (Biocon Diagnosemittel GmbH og Co., Tyskland), etter modifikasjoner tidligere beskrevet av Iversen mfl. (2009).
2.9 Kronisk stresstest; ACTH- og DEXtest
For å undersøke behandlingseffekten av hypothalamus – hypofyse-interrenalaksen (HPI-aksen) sin sensitivitet og tilbakekoblingssystemet, ble en stimulerings- og tilbakekoblingstest gjennomført ved bruk av henholdsvis adrenokortikotropisk hormon (ACTH) og deksametason (DEX). Testen ble utført i samsvar med tidligere studier av Pottinger og Carrick (2001), med noen mindre modifikasjoner som beskrevet av Iversen og Eliassen (2014). For denne testen ble 12 fisk per gruppe, inkludert den negative kontrollen, testet en uke etter siste behandling i begge forsøk. Fisken ble håvet fra oppbevaringskarene, bedøvet (trikainmetansulfonat, 0,1 g/L) og deretter injisert intraperitonealt med 1 mg/kg deksametason (Sigma-Aldrich) i etanol/fosfatbufret saltvann (PBS) (1 :3; 1 μg μL−1). Til slutt ble de overført til 5 beholdere (140 × 100 × 74 cm, ~1000 L). Etter 24 timer ble fisken bedøvet, og 6 fisk fra hver gruppe fikk enten en intraperitonal injeksjon av 0,5 mL/kg adrenokortikotropt hormon (ACTH, fragment 1–24; Sigma-Aldrich) ved 45 μg/μL eller 0,5 mL/kg PBS. To timer etter ACTH/PBS-administrasjonen ble fisken håvet, bedøvet (trikainmetansulfonat, 1 g/L) og tatt blodprøver fra. Disse prøvene ble så blesentrifugert for plasma.
2.10 Etikk
Forsøkene ble utført ved Havforskningsinstituttets sin forskningsstasjon Matre og Nord universitet sin forskningsstasjon i Mørkvedbukta. Begge stasjonene er autorisert for dyreforsøk av Mattilsynet, (anlegg ID: 110, og 017). Begge forsøkene er godkjent av Mattilsynet (søknads-ID: 26549 og 26550) i henhold til Forskrift om bruk av dyr i forsøk.
3. Resultater
3.1 Dødelighet
3.1.1 Forsøk 1, utgangstemperatur 14 °C
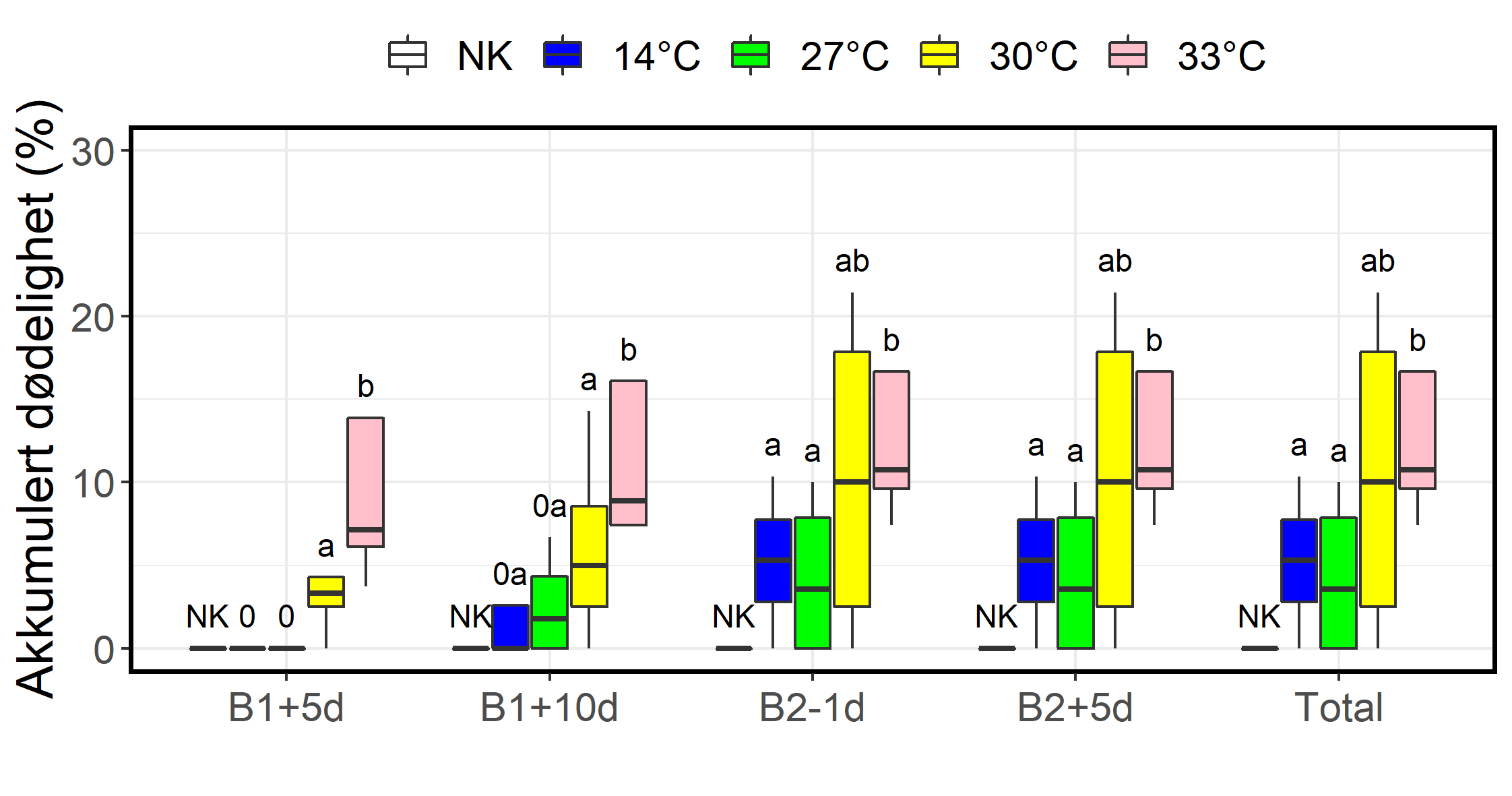
I de første dagene etter første behandling døde det ingen fisk i gruppene som hadde blitt eksponert for 14 °C (prosedyre kontroll) og 27 °C, mens det var signifikant dødelighet i gruppen som hadde blitt behandlet på 30 °C, og ytterligere høyere dødelighet i gruppen som hadde blitt behandlet på 33 °C (figur 7). Etter hvert døde det også fisk i gruppene som hadde blitt behandlet på 14 og 27 °C og forskjellen ble mindre. I motsetning til etter første behandling var det ingen dødelighet etter andre behandling, og dermed ingen endring i akkumulert dødelighet for gruppene. For den negative kontrollen, dvs. fisken som forble urørt, ble det ikke registrert noe dødelighet i løpet av eksperimentet.
Figur 7. Boksplott med akkumulert dødelighet 5 og 10 dager etter første behandling, 1 dag før andre behandling (B2-1d), 5 dager etter andre behandling og ved slutten av forsøket (Total). 0 indikerer ingen signifikant forskjell i forhold til negativ kontroll, Ulike små bokstaver indikerer at de høyere temperaturene hadde statistisk signifikant høyere dødelighet enn de lavere som testet med Fisher’s excact test og Cochran-Mantel-Haenszel test. Figur fra Bui mfl. (2022).
3.1.2 Forsøk 2, utgangstemperatur 8 °C
Totalt 11 av 400 fisk (ekskludert de som ble tatt ut for prøvetaking) døde. Disse døde hovedsakelig i løpet av dagene etter første og andre behandling, men det var ingen signifikante forskjeller mellom de ulike gruppene (tabell 2).
Tabell 2. Antall døde fisk gjennom hele forsøksperioden per kar og behandlingsgruppe. * Fjernet pga store sår på kroppen.
Gruppe
Kar 1
Kar 2
Kar 3
Kar 4
Total
Negativ kontroll
4*
4
8 °C (p. kontroll)
0
2
0
2
27 °C
0
0
0
0
30 °C
1
0
2
3
33 °C
0
2
0
2
3.2 Atferd
3.2.1 Forsøk 1, utgangstemperatur 14 °C
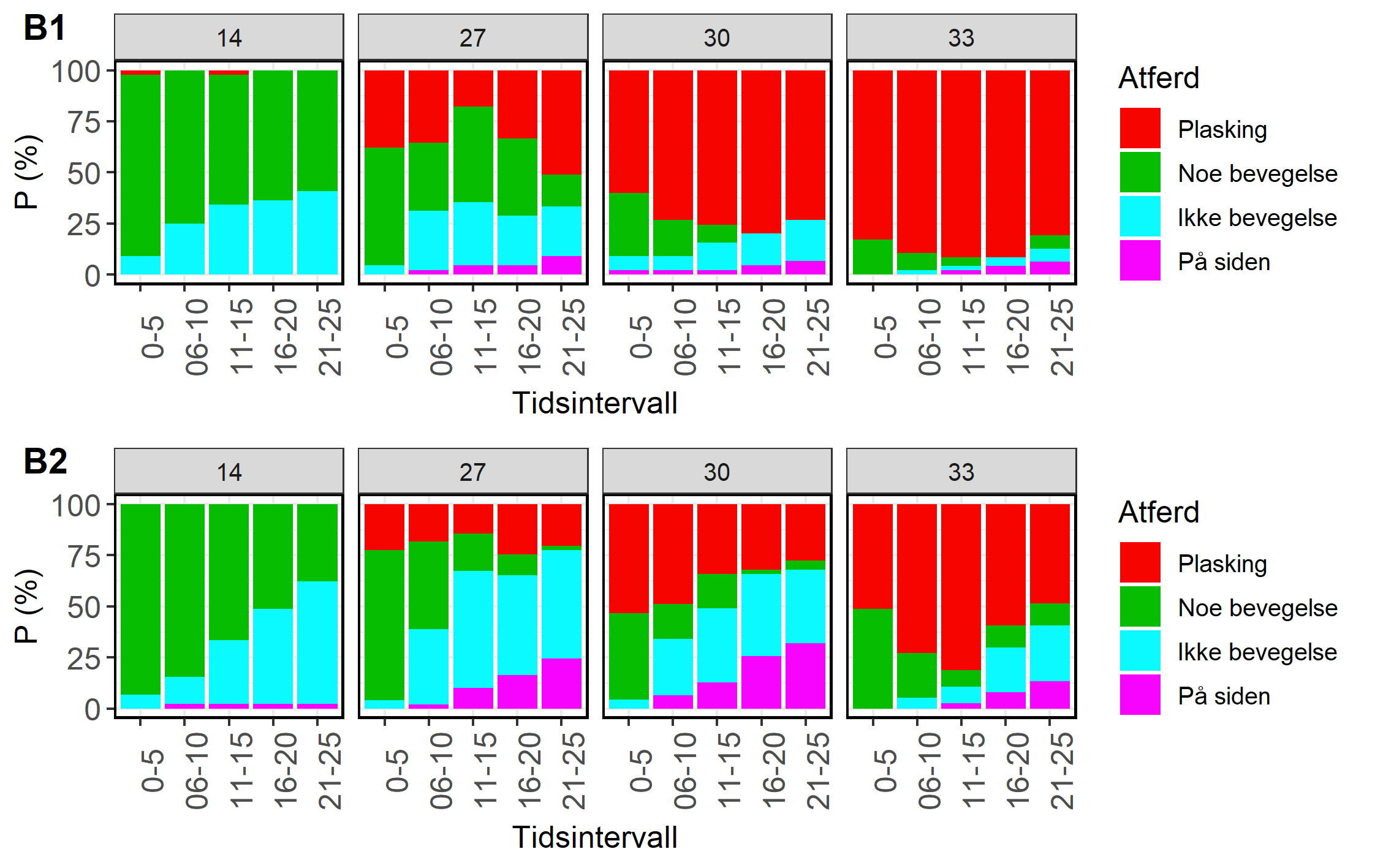
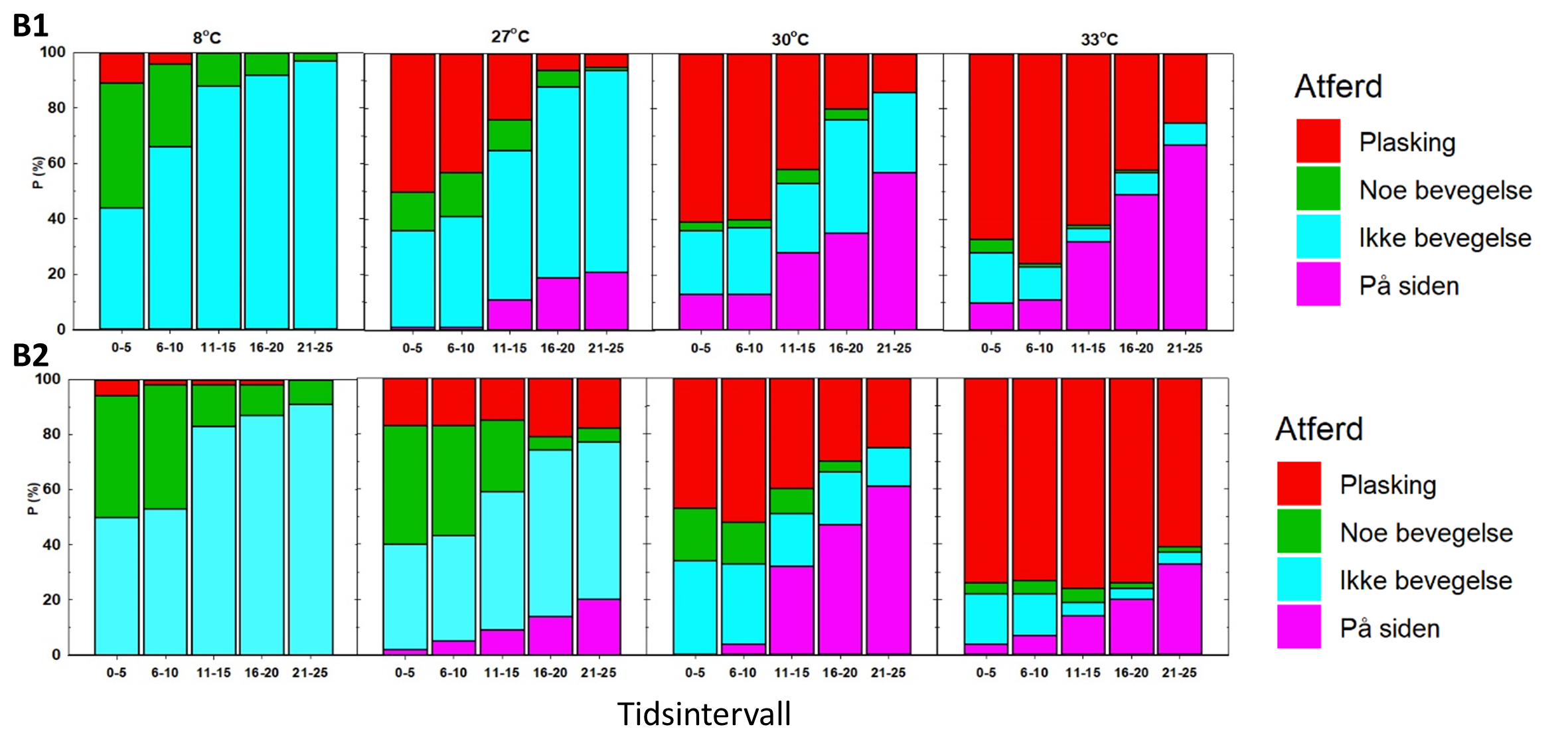
Figur 8. Observert atferd ved første behandling (B1) og andre behandling (B2). Y-aksen angir andel fisk med observert atferd (Tabell 3) for tidsrom første 5 sekunder av eksponeringen (0-5), neste fem sekunder (6-10), osv. Figuren er reprodusert fra Bui mfl. (2022).
Adferdsobservasjonene fra forsøk 1 viste en klar effekt av temperatur ved begge behandlinger (figur 8). Fisken som ble eksponert for 14 °C oppførte seg rolig i behandlingskammeret, og det ble i hovedsak kun registrert «Noe bevegelse» eller «Ikke bevegelse» gjennom hele eksponeringen (figur 8). Ved de andre temperaturene var det et økende innslag av atferd kategorisert som «plasking» med økende temperatur, altså eksplosiv aktiv atferd med mye plasking, hopping, buktning, m.m. Ved første behandling var det innslag av fisk som la seg over på siden, mens dette var betydelig vanligere ved andre behandling, særlig for fisk som ble eksponert for 27 og 30 °C (figur 8). Et typisk atferdsmønster for fisk som la seg over for siden var at de først plasket, så stod stille noen sekunder før de la seg over på siden.
3.2.2 Forsøk 2, utgangstemperatur 8 °C
På samme måte som i forsøk 1 viste atferdsobservasjonene fra forsøk 2 en klar effekt av temperatur både ved første og andre behandling (figur 9). I motsetning til i forsøk 1 økte ikke innslaget av fisk som la seg over på siden fra første til andre behandling, men også her syntes en større andel av fisken å reagere ved å legge seg over på siden ved 30 enn ved 33 °C.
Figur 9. Observert atferd ved første (B1) og andre (B2) behandling. Y-aksen angir andel fisk med observert atferd i % (Tabell 2) for tidsrom første 5 sekunder av eksponeringen (0-5), neste fem sekunder (6-10), osv.
3.3 Skåring av ytre skader
3.3.1 Forsøk 1, utgangstemperatur 14 °C
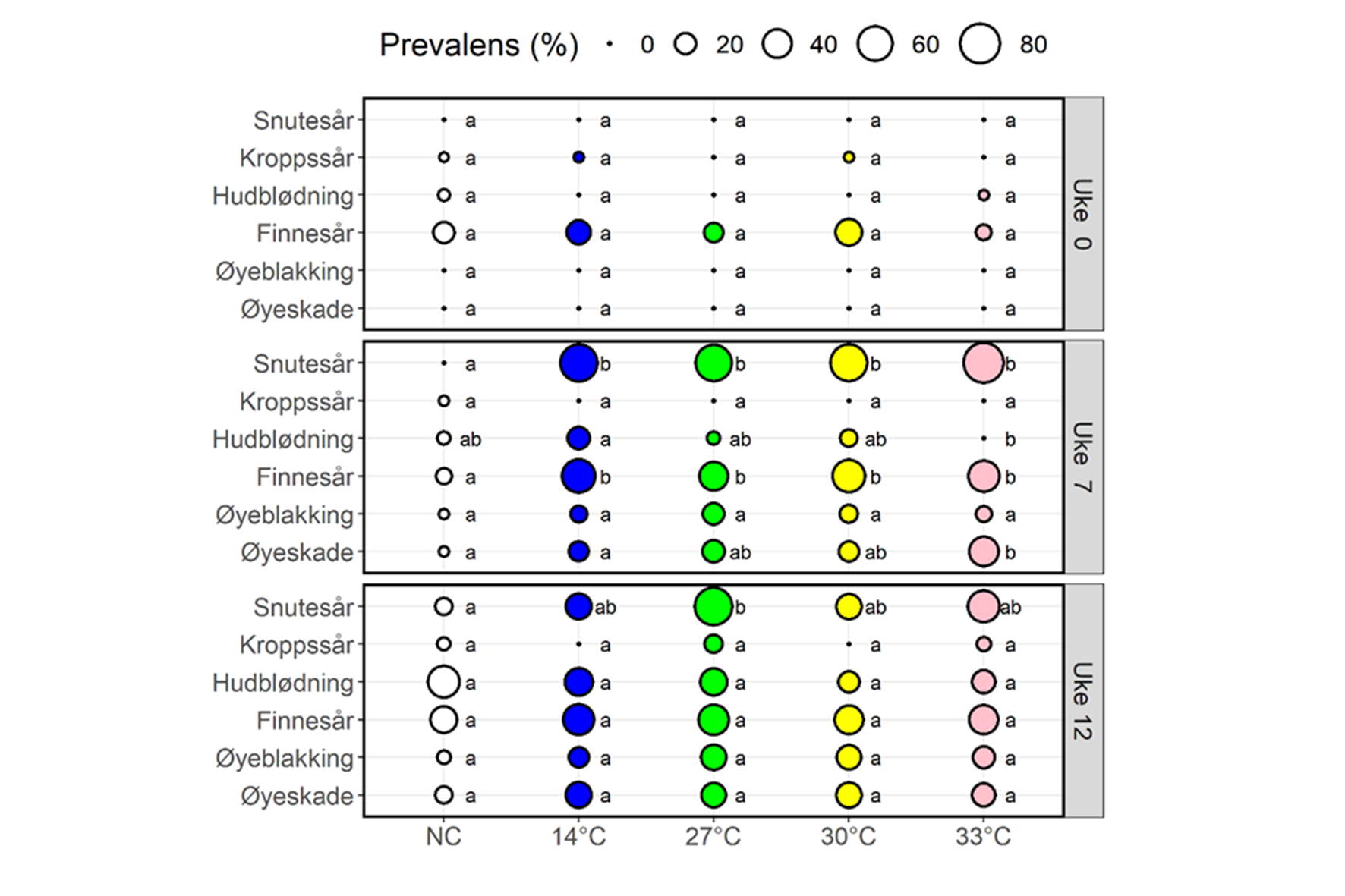
Alle gruppene hadde tilsvarende skåring av ytre skader da de ble flyttet inn i forsøksfasiliteten (Uke 0, figur 10). Skåringen en uke etter siste behandling viste en generell økning i andel fisk som ble skåret med moderate avvik (skår ≥ 2) for de fleste av indikatorene (Uke 0 vs. Uke 7, figur 10). Resultatene viste ingen tydelig forskjell mellom prosedyrekontrollgruppen som ble behandlet på 14 °C og gruppene som ble behandlet på de høyere temperaturene, bortsett fra signifikant mer øyeskade i 33 °C-gruppen, og generelt mindre skade i negativkontroll-gruppen (uke 7. figur 10), Ved den siste prøvetakingen, etter at negativkontrollgruppen også hadde blitt håndtert i forbindelse med samplingen i uke 7 og flytting til nytt kar sammen med de andre gruppene, hadde negativkontrollgruppen lignende skår som de andre gruppene (figur 10, uke 12).
Figur 10. Prevalens av forsøksfisk blant behandlingsgrupper som ble vurdert fra moderat til alvorlig (skår 2–3) for hver velferdsindikator for prøvetaking i uke 0, 7 og 12. For hver velferdsindikator indikerer samme liten bokstav at det ikke er noen statistiske forskjeller i prevalens mellom gruppene. Fra Bui mfl. (2022).
3.3.2 Forsøk 2, utgangstemperatur 8 °C
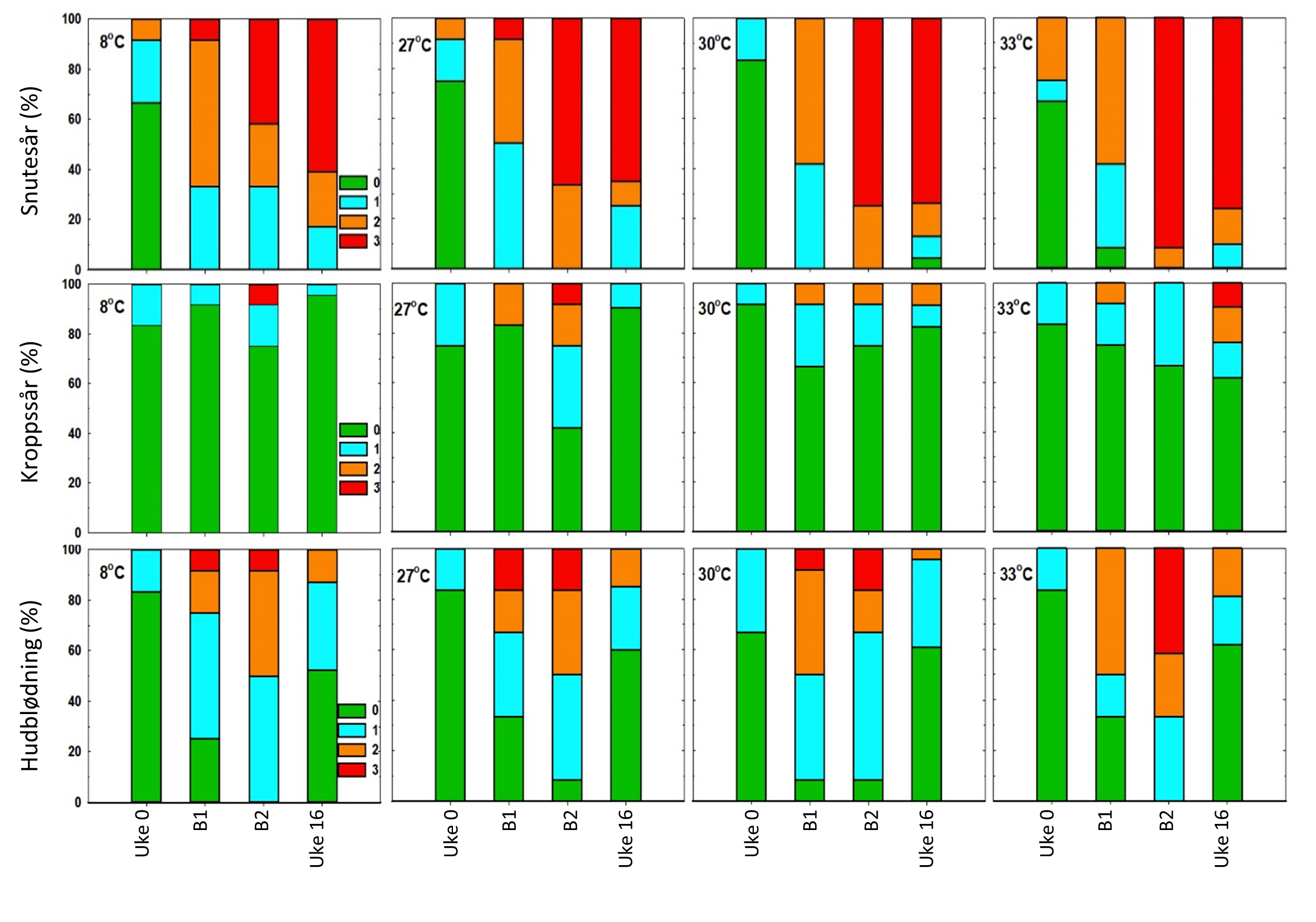
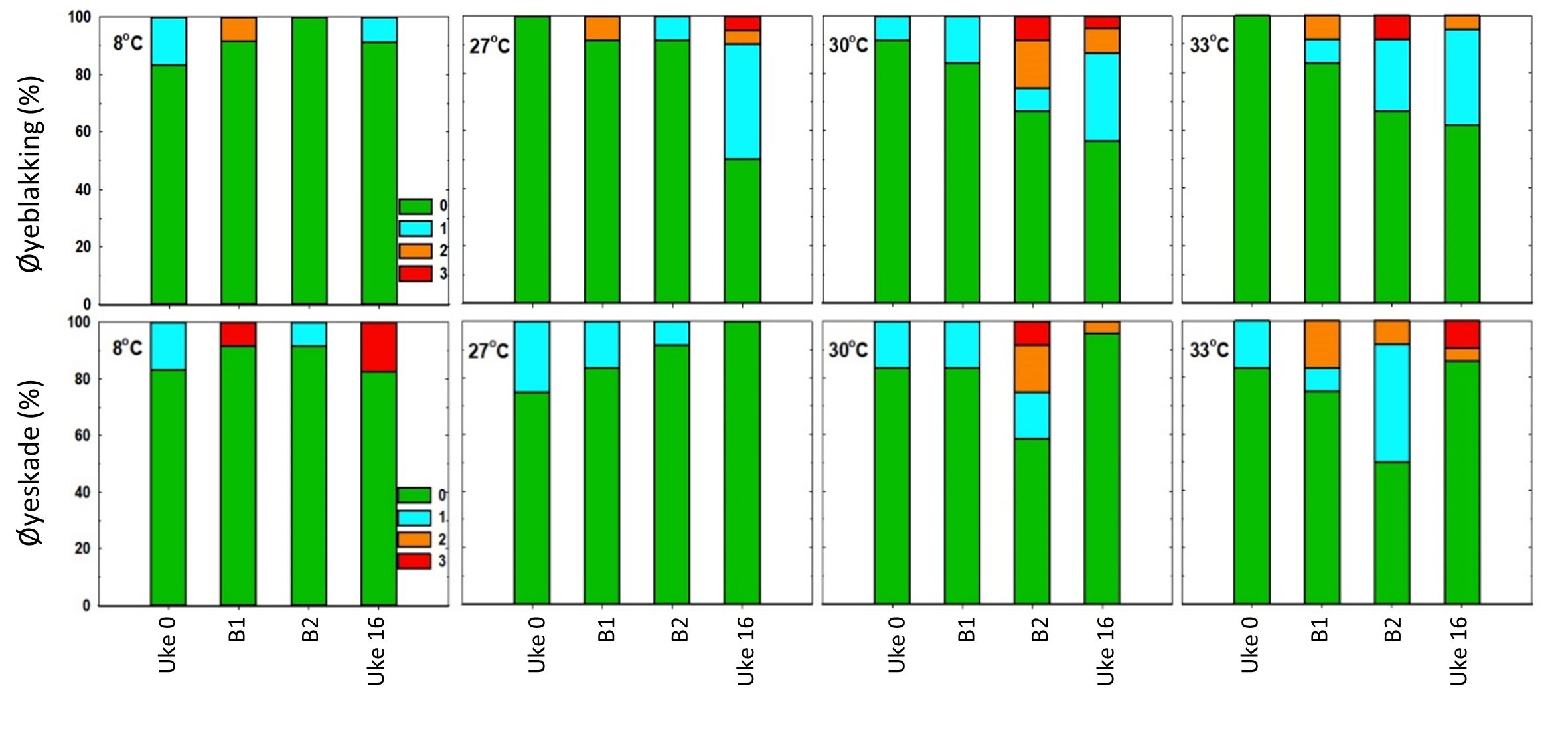
Alle gruppene hadde tilsvarende skår for ytre skader da de ble flyttet inn i forsøksfasiliteten (Uke 0, figur 11-12). Resultatene viser ingen tydelig forskjell mellom prosedyrekontrollgruppen som ble behandlet på 8 °C og gruppene som ble behandlet på de høyere temperaturene (figur 11-12). Med unntak finneskår (data ikke vist) var det imidlertid en generell økning i andel fisk som ble skåret med tydelige avvik (skår ≥ 2) for de fleste av indikatorene, inkludert hudblødninger, snuteskader og øyeblødninger. Disse indikatorene syntes også å øke i frekvens etter andre behandling (B2, figur 11-12), men det ble ikke påvist noen signifikante forskjeller mellom behandlingsgruppene, bortsett fra for en signifikant økning av hudblødninger i behandlingsgruppe 33 °C sammenliknet med de andre gruppene.
Figur 11. Prevalens av skår for velferdsindikatorer snutesår, kroppssår og hudblødning i uke 0, 24 timer etter første behandling (B1), 24 timer etter andre behandling (B2) og i uke 16 (langtidseffekter). Grønn = prosentandel med skår 0, turkis = andel med skår 1, oransje = andel med skår 2 og rød = andel med skår 3.Figur 12. Prevalens av skår for velferdsindikatorer øyeblakking og øyeskade i uke 0, 24 timer etter første behandling (B1), 24 timer etter andre behandling (B2) og i uke 16 (langtidseffekter). Grønn = prosentandel med skår 0, turkis = andel med skår 1, oransje = andel med skår 2 og rød = andel med skår 3.
3.4 Vekst
3.4.1 Forsøk 1, utgangstemperatur 14 °C
Ved prøvetakingen av all fisk en uke etter siste behandling (Uke 7) hadde alle de behandlete gruppene gått noe ned i vekt, mens negativkontrollgruppen hadde gått noe opp i vekt (tabell 3). Det var ingen signifikante forskjeller i vekst mellom gruppene som hadde blitt behandlet (tabell 3). I måneden uten forstyrrelser mellom tredje sampling i uke 7 og fjerde sampling i uke 12 begynte alle gruppene å vokse igjen (SGR7-12, tabell 3).
Tabell 3. Snitt vekt (g) og SGR for overlevende fisk til tredje (Uke 7) og fjerde sampling (uke 12). Signifikante forskjeller i vekst (GLM modell korrigert for startvekt) er indikert med ulik bokstav bak SGR-verdiene. *Ved tredje sampling ble størsteparten av fisken avlivet, bortsett fra 30 fisk som fikk gå videre i ytterligere fem uker. Antall fisk inkludert i hver snitt-beregning er angitt i kolonnene N0-7 og N7-12. Tabell fra Bui mfl. 2022.
Gruppe
Uke 0
Uke 7
SGR0-7
N0-7
Uke 7 *
Uke 12
SGR7-12
N7-12
NK
2245±37
2381±45
+0.089a
116
2505±82
2885±91
+0.403a
29
14 °C
2100±40
2055±44
÷0.040b
85
2101±80
2378±82
+0.364a
30
27 °C
2138±41
2111±49
÷0.033b
87
2315±87
2516±90
+0.224a
28
30 °C
2207±39
2162±42
÷0.039b
83
2263±65
2681±77
+0.374a
30
33 °C
2131±55
2033±54
÷0.081b
72
2240±87
2529±110
+0.330a
29
3.4.2 Forsøk 2, utgangstemperatur 8 °C
Snittvekt ved første prøvetaking (uke 0) lå mellom 1700 og 1880 g (tabell 4). Etter første (B1) og andre (B2) behandling var det en stagnasjon i veksten hos alle behandlingsgruppene. En uke etter siste behandling (uke 9) begynte tilveksten å tilta, og to måneder etter siste behandling (uke 16) lå snittvekten mellom 2830 og 3130 g. Dette var signifikant høyere (p<0,05, t-test) enn ved forsøksstart (tabell 4), men det var ingen forskjeller mellom temperaturgruppene ved de ulike prøvetakningstidspunktene.
Tabell 4. Snitt vekt (g) og SGR for overlevende fisk til tredje (uke 8) og fjerde sampling (uke 9). Signifikante forskjeller i vekst mellom negativ kontroll og behandlingsgruppene er indikert med * (p<0.05, t-test). *Ved tredje sampling ble størsteparten av fisken avlivet, bortsett fra 30 fisk som fikk gå videre i ytterligere fem uker. Antall fisk inkludert i hver snitt-beregning er angitt i kolonnene N0-8 og N9-16.
Gruppe
Uke 0
Uke 8
SGR0-8
N0-7
Uke 9 *
Uke 16
SGR9-16
N9-16
NK
1820±53
2090±44
+0.247
100
2400±30
3080±74
+0.509
30
8 °C
1880±45
1560±36
÷0.333*
64
2280±50
3130±74
+0.647
28
27 °C
1730±54
1550±48
÷0.196*
62
2160±46
2990±92
+0.664
25
30 °C
1800±53
1840±63
+0.039*
64
2060±52
2830±103
+0.648
30
33 °C
1850±57
1860±53
+0.009*
64
2200±75
2950±95
+0.599
30
3.5 Fysiologiske responser
3.5.1 Forsøk 1, utgangstemperatur 14 °C
Blodplasmamålingene viste ingen signifikant forskjell mellom behandlingsgruppene etter verken første eller andre behandling, men, med unntak av magnesium, var verdiene generelt lavere etter den andre termiske eksponeringen sammenlignet med etter den første (se Bui mfl. 2022).
3.5.2 Forsøk 2, utgangstemperatur 8 °C
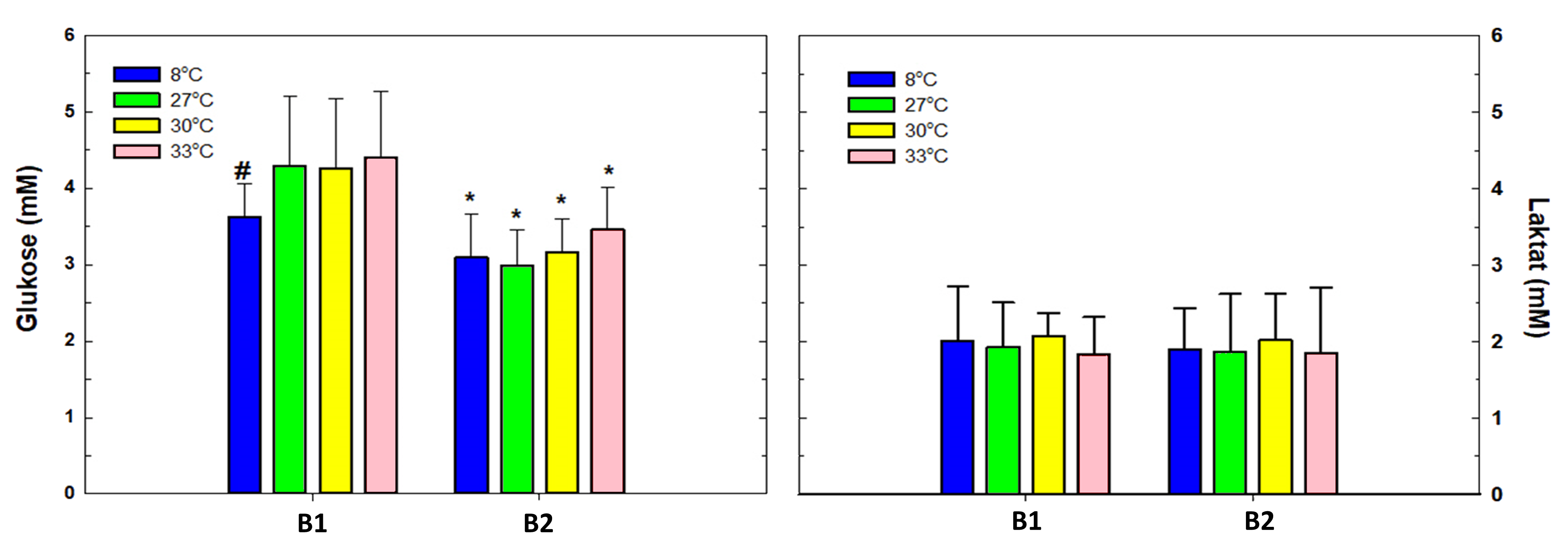
Figur 13. Laktat (mM) og glukose (mM) av helblod 24 timer etter første (B1) og andre (B2) behandling. Signifikante forskjeller i figurene innen en gruppe ved ulike prøvetakingstidspunkt ble indikert med *, og forskjeller mellom forsøksgruppene ved samme prøvetakingstidspunkt ble indikert med #. Alle resultater er uttrykt i gjennomsnitt med standardavvik (ñ ± SD). Signifikante forskjeller er basert på en ikke-parametrisk Kolmogorov-Smirnov test, og signifikantnivå p<0,05.
Resultatene av glukose viser tydelig forskjell mellom prosedyrekontrollgruppen som ble behandlet på 8 °C, og gruppene som ble behandlet på de høyere temperaturene etter første behandling, men ingen signifikante forskjeller mellom høy temperaturgruppene (figur 13). Laktatmålingene viste ingen signifikante forskjeller, hverken etter første eller andre behandling (figur 13).
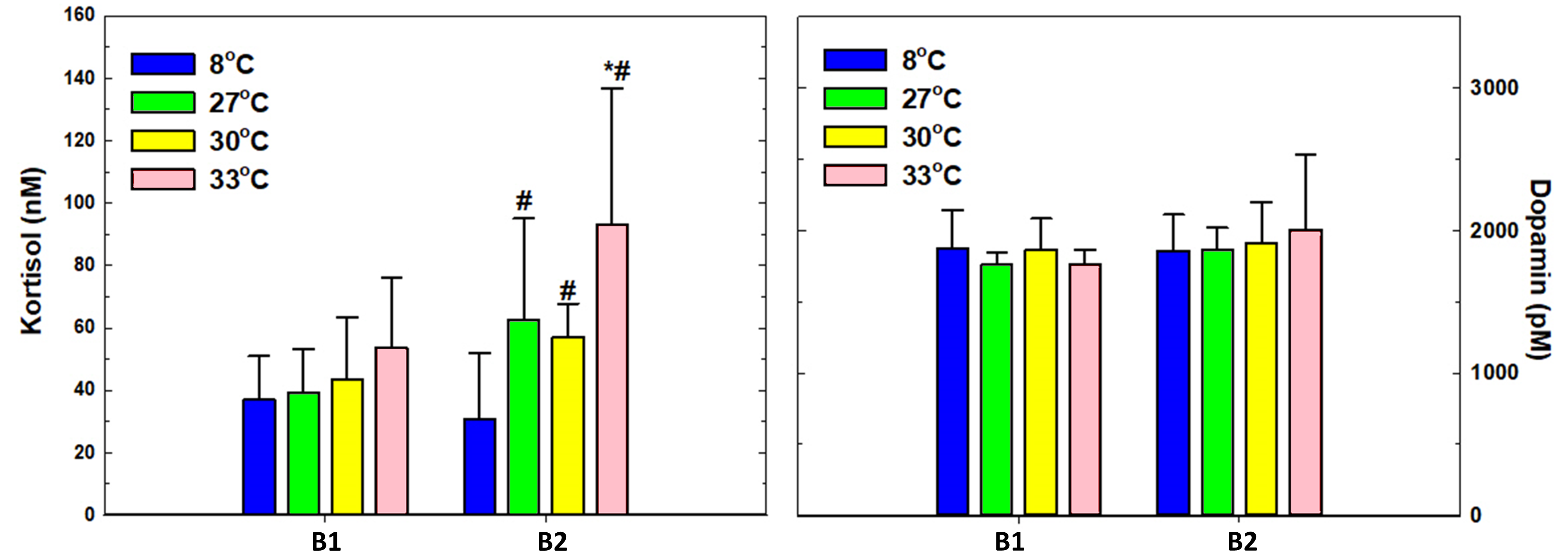
Figur 14. Blodplasmanivåene av kortisol (nM) og dopamin (pM). Signifikante forskjeller i figurene innen en gruppe ved ulike prøvetakingstidspunkt ble indikert med *, og forskjeller mellom forsøksgruppene ved samme prøvetakingstidspunkt ble indikert med #. Alle resultater er uttrykt i gjennomsnitt med standardavvik (ñ ± SD). Signifikante forskjeller er basert på en ikke-parametrisk Kolmogorov-Smirnov test, og signifikantnivå p<0,05.
Resultatene viser tydelig forskjell i plasmakortisol mellom prosedyrekontrollgruppen som ble behandlet på 8 °C, og gruppene som ble behandlet ved de høyere temperaturene ved prøvetakingstidspunkt B2. (figur 14). I alle temperaturbehandlingene ved B2 ligger plasmakortisol over de normale hvilenivåene en kan forvente hos postsmolt av denne størrelsen (13,8 – 40 nM) (Maule mfl. 1988; Pickering og Pottinger 1989; Iversen og Eliassen, 2014), og høyest var plasmakortisolnivåene i 33 °C gruppen (rosa gruppe, figur 14).
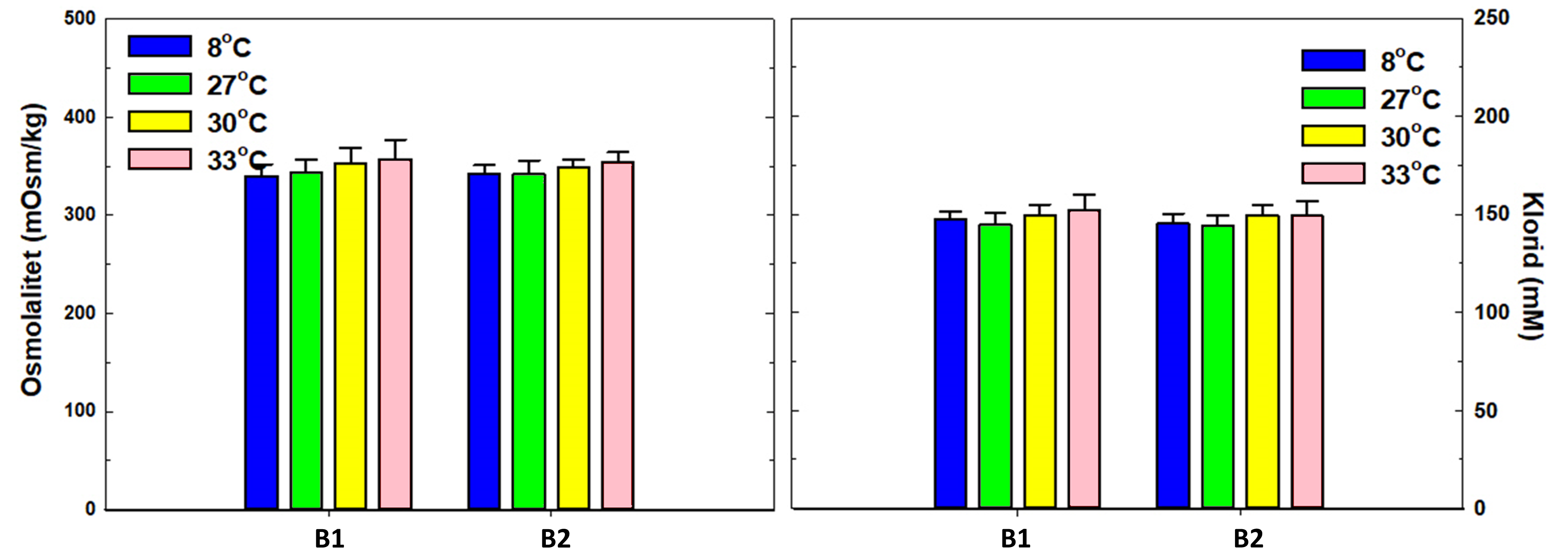
Figur 15. Blodplasmanivåene av osmolalitet (mOsm/kg) og klorid (mM). Signifikante forskjeller i figurene innen en gruppe ved ulike prøvetakingstidspunkt ble indikert med *, og forskjeller mellom forsøksgruppene ved samme prøvetakingstidspunkt ble indikert med #. Alle resultater er uttrykt i gjennomsnitt med standardavvik (ñ ± SD). Signifikante forskjeller er basert på en ikke-parametrisk Kolmogorov-Smirnov test, og signifikantnivå p<0,05.
Det var ingen effekt på osmolalitet og klorid ved noen av behandlingstemperaturene hverken etter første eller andre behandling (figur 15).
3.6 Kronisk stresstest; ACTH- og DEXtest
3.6.1 Forsøk 1, utgangstemperatur 14 °C
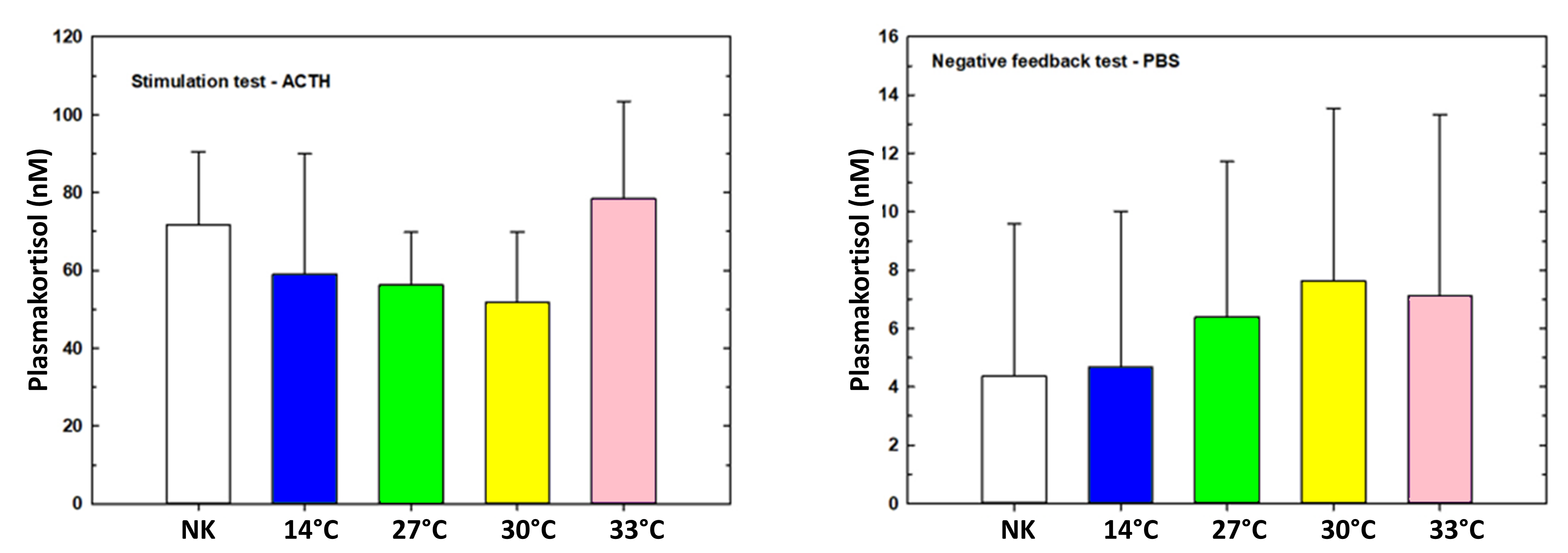
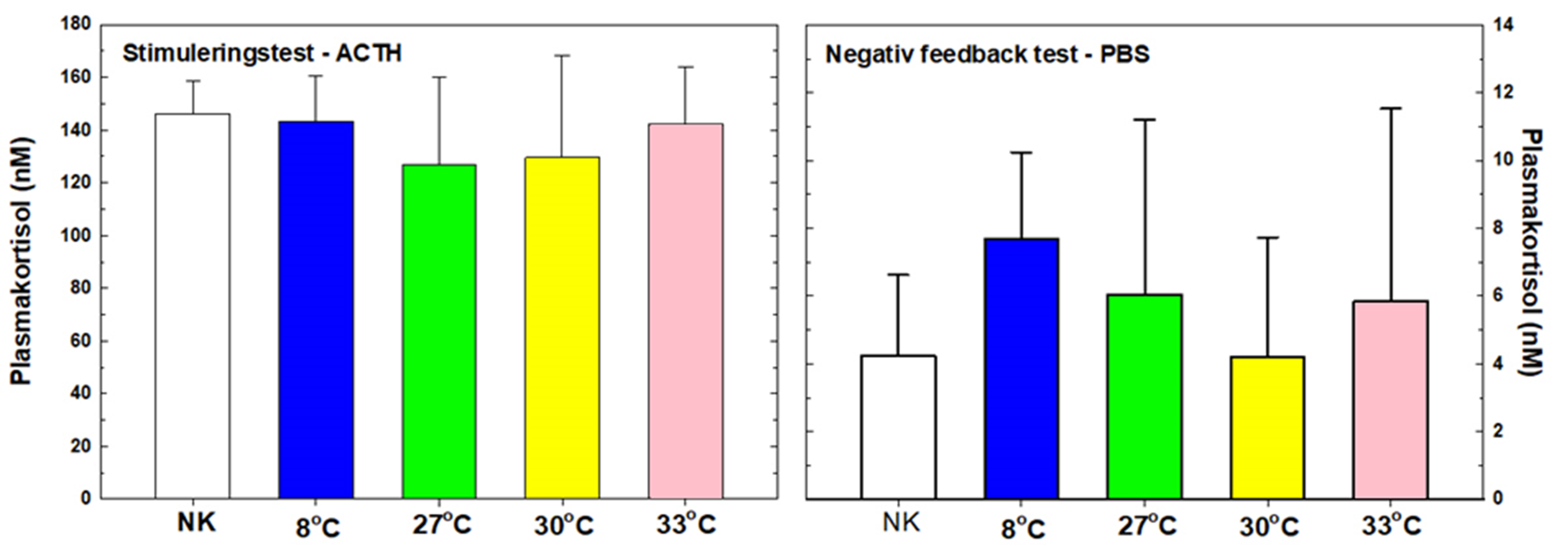
Figur 16. Stimulerings- (ACTH) og negativ tilbakekolblingstest (PBS) ved forsøk 1, utgangstemperatur 14 °C.
Det ble ikke målt noen signifikante forskjeller mellom gruppene i stimulerings-(ACTH) eller negativ tilbakekoblingstesten (PBS) ved sluttsampling (uke 7, figur 5).
3.6.2 Forsøk 2, utgangstemperatur 8 °C
Som i forsøk 1 (figur 16) viste stimulerings-(ACTH) eller negativ tilbakekoblingstesten (PBS) i forsøk 2 ingen signifikante forskjeller mellom gruppene, og det ble ikke påvist noen kronisk stressbelastning av gjentatte håndtering og temperatureksponeringer (figur 17).
Figur 17. Stimulerings- (ACTH) og negativ tilbakekolblingstest (PBS) ved forsøk 2, utgangstemperatur 8 °C.
3.7 Histologi og røntgen
3.7.1 Forsøk 1, utgangstemperatur 14 °C
Røntgenanalysen viste tegn til gassoppbygging bak øyet (figur 18) i 1 av 36 termisk behandlete fisk og ingen av de 12 samplede fiskene fra prosedyrekontrollen. Den aktuelle fisken hadde blitt behandlet på 27 °C. Øyet hadde tegn til exophthalmia, men obduksjon av øyet avdekket ingen andre tegn til skade eller infeksjon.
Figur 18. Røntgenbilde av fisk med gassoppbygging på innsiden av høyre øyet (angitt med pil). Fra Bui mfl. 2022.
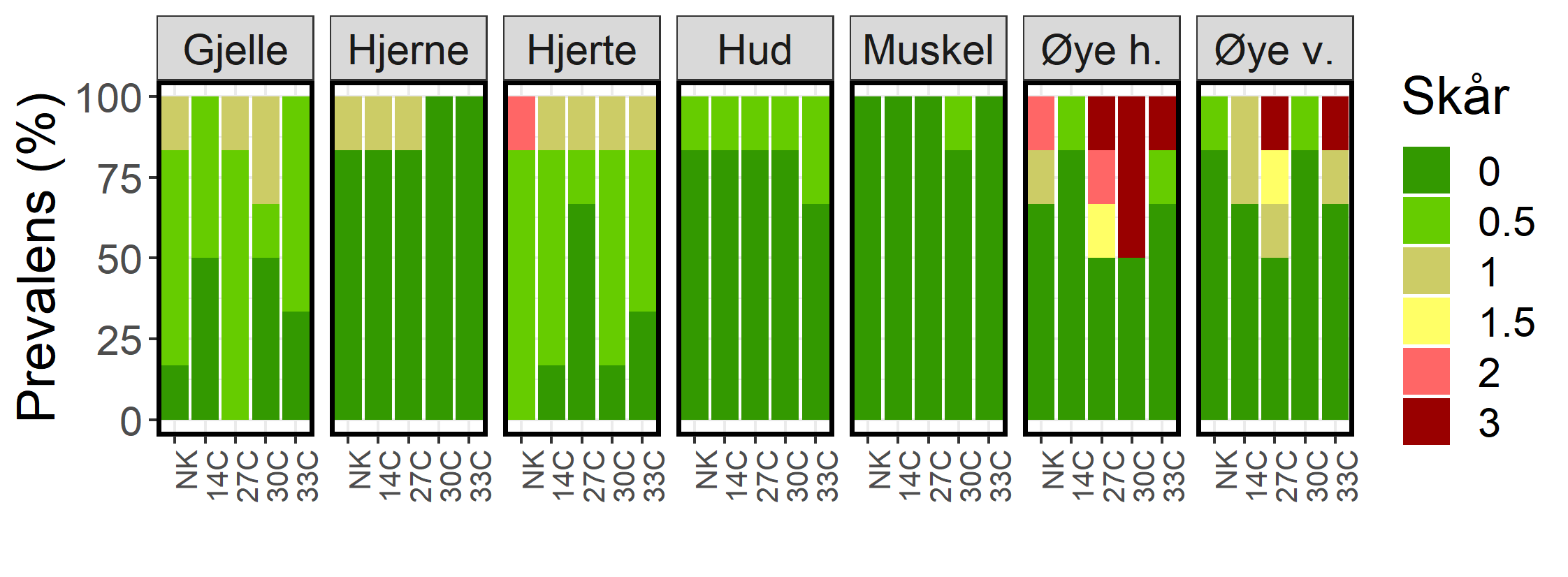
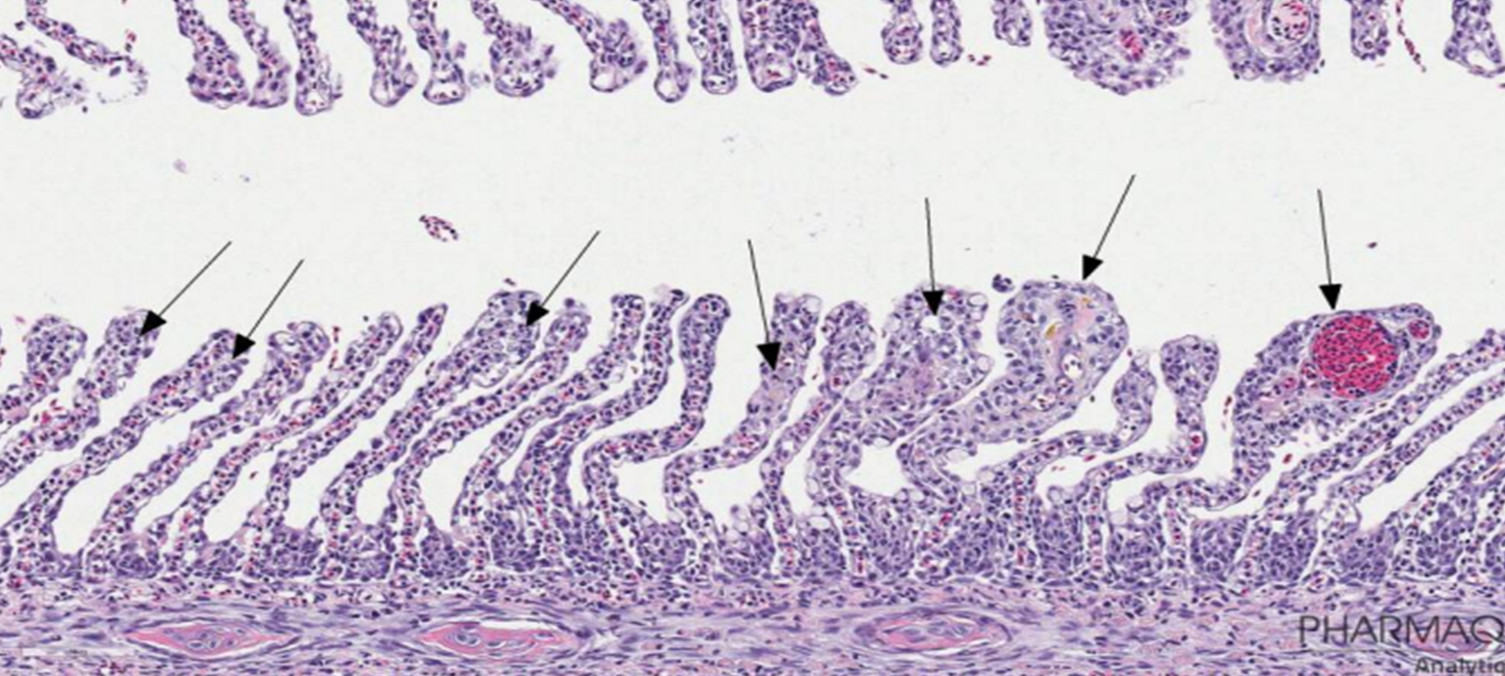
Histologisk undersøkelse av gjeller, hjerne, hjerte, hud og muskel fra 6 fisk fra hver gruppe (totalt 30 fisk) påviste kun minimale til milde funn (figur 19), bortsett fra hjertet til en negativ kontroll fisk som hadde moderat kardiomyopati (CMS). I prøvene av øynene fra de samme fiskene ble det påvist 7 tilfeller av alvorlige betennelsesforandringer (se figur 20 for eksempler). Alle disse påvisningene var fra fisk som hadde blitt eksponert for vann på 27, 30 eller 33 °C (figur 19). Det var ingen fisk som hadde alvorlige betennelsesforandringer i både venstre og høyre øye. I negativkontrollgruppen og gruppen av fisk som ble utsatt for vanntemperatur på 14 °C (prosedyre kontroll) ble det påvist milde til moderate betennelsesforandringer i 4 av totalt 24 øyne, men ingen av fiskene i denne gruppen hadde alvorlige betennelsesforandringer i øynene. Den histologiske undersøkelsen viste dermed en tydelig forskjell i risiko for skader på øynene mellom fisk eksponert for varmt vann og fisk i kontrollgruppene (p=0,035, Fisher’s excact test).
Figur 19. Histologiresultat fra prøvetaking Uke 7. Seks fisk ble analysert fra hver gruppe. Venstre og høyre øye, i tillegg til hjerne, hjerte, gjeller, hud, skjelettmuskulatur og nyre ble undersøkt og funnene beskrevet og gradert. Funnene er gradert fra 0,5-3, der "0,5" representerer minimale funn uten diagnose, "1" representerer milde funn, "2" representerer moderate funn og "3" representerer alvorlige eller uttalte funn. "0" representerer ingen spesifikke funn. Y-aksen angir frekvensen av observert skår per vev. Figur fra Bui mfl. 2022.
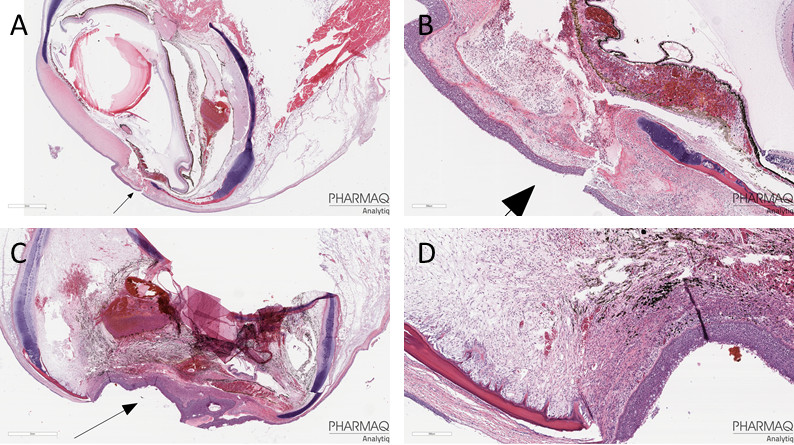
Figur 20. Representative histopatologiske funn i øyne med uttalte betennelsesforandringer (skår 3). A og B: Hornhinnebetennelse (keratitt). C og D (nærbilde): keratitt og betennelse inne i øyet (oftalmitt). Begge fiskene i disse eksemplene hadde blitt behandlet på 30 °C. Bilder: Caroline Piercey Åkesson, Pharmaq Analytiq AS.
3.7.2 Forsøk 2, utgangstemperatur 8 °C
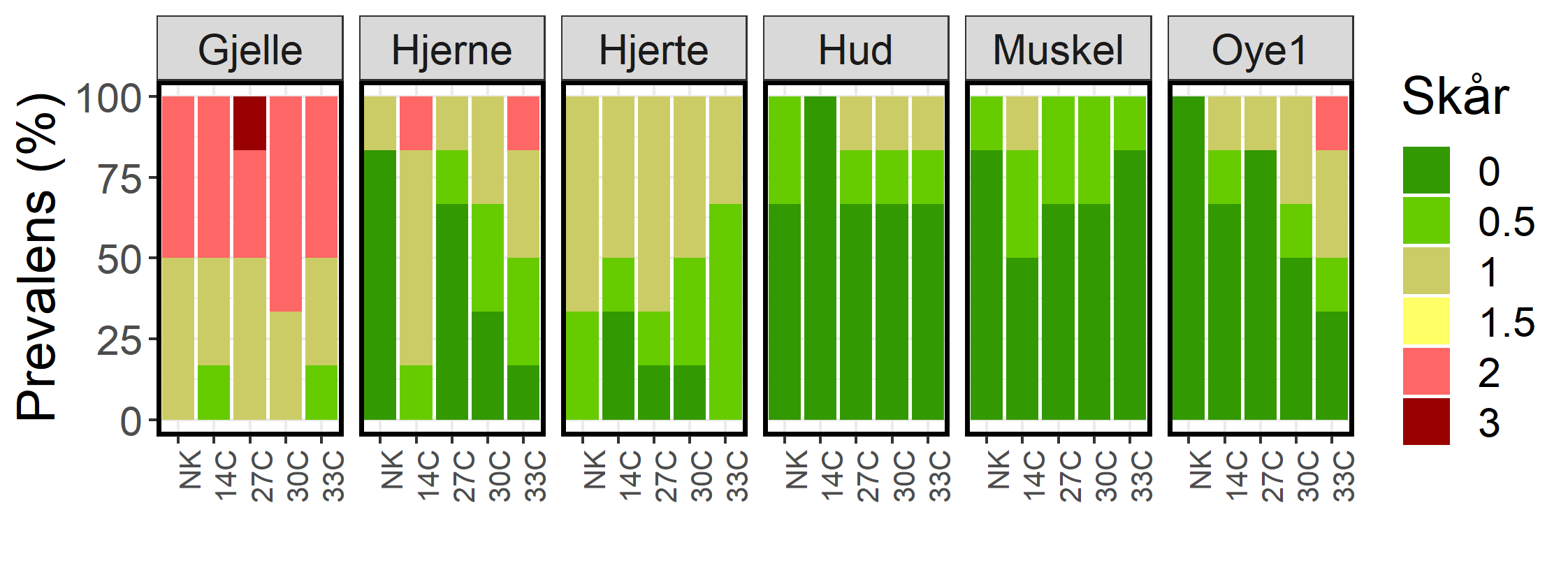
Figur 21. Histologiresultat fra prøvetaking Uke 9. Seks fisk ble analysert fra hver gruppe. Venstre øye, i tillegg til hjerne, hjerte, gjeller, hud, skjelettmuskulatur ble undersøkt og funnene beskrevet og gradert. Funnene er gradert fra 0-3, der "0" representerer ingen spesifikke funn, "0,5" representerer minimale funn uten diagnose, "1" representerer milde funn, "2" representerer moderate funn og "3" representerer alvorlige eller uttalte funn. Y-aksen angir frekvensen av observert skår per vev.
De histologiske analysene avdekket overveiende milde vevsskader, bortsett fra gjeller der det ble påvist varierende grad av bruskdeformiteter, betennelse og lamellære karskader (figur 21). Subepitelial nekrose og betennelse (figur 22), lignet mest på det man ser ved infeksjon med Paranucleospora theridion, men kan også være forårsaket av Branchiomonas cysticola og Parvicapsula pseudobranchicola (Myxosporidie), som ble påvist i inntaksvannet (Patogen). Gjelleforandringene var like fremtredende hos kontrollfisk som hos fisk eksponert for de ulike behandlingstemperaturene.
Figur 22. Gjelle med lamellære blodkarskader, subepiteliale nekroser og betennelser. Bilde: Helene Wisløff, Pharmaq Analytic AS.
3.7.3 Ekstra vevsundersøkelse av hjerte, forsøk 2
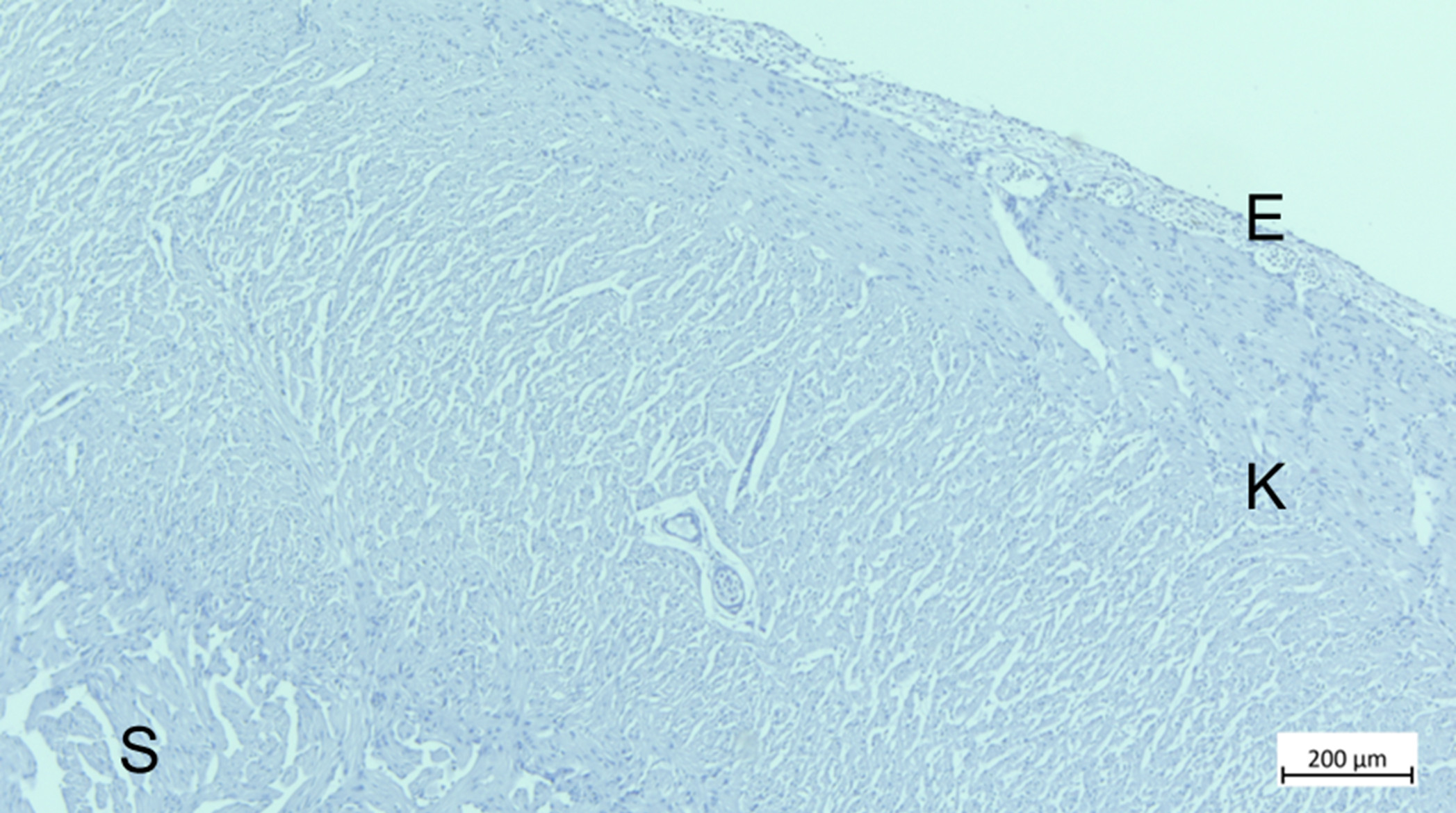
Det ble ikke påvist histopatologiske forandringer i hjertevevet tatt 2 måneder etter siste behandling, verken i hjertevev fra kontrollfisk eller i hjertevev fra termisk behandlet fisk. Hjertevevet var normalt (figur 23).
Figur 23. Histologisk snitt av hjerteventrikkel fra fisk eksponert for 33 °C to måneder etter siste termiske behandling. N Normal hjertehistologi. E = epikard, K = stratum kompaktum, S = stratum spongiosum. Bilde: Tor Seternes, UiT.
4. Diskusjon
I tidligere kontrollerte forsøk med repetert termisk behandling var det ikke forhøyet dødelighet etter behandling (Moltumyr mfl. 2022). Den forhøyede dødeligheten etter første behandling i forsøk 1 kom derfor som en overraskelse, særlig siden behandlingen hadde en lavere Δt enn i Moltumyr mfl. (2022). I Moltumyr mfl. (2022) var utgangstemperaturen 8 °C og behandlingstemperaturen 34 °C, mot utgangstemperatur på 14 °C og behandlingstemperaturer på enten 27, 30 og 33 °C i forsøk 1. Det er imidlertid godt kjent fra industridata at termisk avlusing gir risiko for høy dødelighet i tiden etter behandlingen (Overton mfl. 2019, Grefsrud mfl. 2022). Her er det viktig å ta inn over seg betydningen av ordet ‘risiko’. Risiko innebærer at en hendelse kan inntreffe, ikke at den alltid gjør det. Dette er videre illustrert med at det ikke var forhøyet dødelighet etter andre behandling i forsøk 1 og ikke etter noen av behandlingene i forsøk 2. Spørsmålet blir da, hva var det som gjorde at den første behandlingen i forsøk 1 ga forhøyet dødelighet, men ikke de andre behandlingene?
Siden behandlingene i forsøk 2 uten dødelighet fulgte tilsvarende prosedyre som behandlingene i forsøk 1, og andre behandling i forsøk 1, som også var uten dødelighet, ble utført identisk til første behandling, er det nærliggende å anta at fiskegruppen i forsøk 1 hadde en iboende svakhet før første behandling som gjorde at de ikke tålte behandlingen. For denne gruppen var det kun to uker siden de hadde blitt flyttet fra produksjonstanken til de mindre forsøkstankene (⌀ = 7 m til 3 m), og gjennomgått en innvendig (PIT) og utvendig (Floy) merking. Flytting av fisk inn i mindre kar og merking er kjente helserisikoer, særlig hvis fisken ikke får tilstrekkelig tid til å restituere seg (Espmark mfl. 2017, Vollset mfl. 2020). Til forskjell fikk fisken i forsøk 2 en uke mer til å restituere seg, og her var også forsøkskarene noe større (8,0 m3 vs 5,3m3). Fisken i Moltumyr mfl. (2022) hadde også fått lenger tid til å restituere seg (fem uker) og større forsøkskar (⌀ = 5 m). Da fisken i forsøk 1 hadde fått fire uker til å restituere seg før andre behandling, var det ikke dødelighet etter behandlingen. Dette understøtter at restitusjon og fiskens tilstand er avgjørende for utfallet av termisk avlusing.
I likhet med tidligere studier (Nilsson mfl. 2019, Moltumyr mfl. 2021, 2022) responderte laksen på det varme vannet med eksplosiv atferd med mye plasking, hopping og buktning, mens denne atferden var nesten helt fraværende hos prosedyrekontrollfisken. Dette gjaldt også når utgangstemperaturen var 14 °C og behandlingstemperaturen kun 27 °C, og dermed en svært lav Δt. En observerte også at andelen laks som viste plaskeatferd økte når temperaturen økte fra 27 til 30 °C, og ved ytterligere økning fra 30 til 33 °C. Dette ligner tidligere undersøkelser gjort av Nilsson mfl. (2019) som fant en tydelig atferdsrespons hos atlantisk laks (~230 g) eksponert for 28 °C, sammenlignet med laks utsatt for lavere temperaturer, med en ytterligere økning i atferdsrespons når laksen ble utsatt for vanntemperaturer over 34 °C. Det har blitt vist at regnbueørret (Oncorhynchus mykiss) har mekanotermiske smertereseptorer i huden og hoderegionen med en termisk terskel rundt 29 °C, og polymodale smertereseptorer med en terskel rundt 33 °C (Ashley mfl. 2007). Disse tersklene varierer innen en populasjon, og varierer sterkt mellom ulike individer (Ashley mfl. 2007). Dermed er det forventet at i en populasjon vil noen individer ikke ha den samme sterke atferdsresponsen på 27 °C som andre, og på samme måte ved 33 °C vil flere individer reagere med en sterkere fluktrespons («plasking») enn ved 30 °C. En kan imidlertid ikke utelukke at utgangstemperaturen hadde en påvirkning på adferden, men siden atferdsresponsen i forhold til absolutt behandlingstemperatur er så like mellom de to forsøkene synes Δt å spille en mindre rolle.
I tillegg til den kraftigere plaskingen var det også en del individer som reagerte med å ligge seg over på siden, typisk flere ved 30 °C enn ved 33 °C. Denne type atferd kan være tegn på manglende mestring og at fisken har gitt opp (Gismervik mfl. 2019, Nilsson mfl. 2019). I Nilsson mfl. (2019) og Gismervik mfl. (2019) mistet laksen likevekt og la seg over på siden etter ~100 s ved 36–38 °C, og undersøkelse av prøver viste alvorlig vevsskade i gjellene og hjernen. I vårt tilfelle var imidlertid eksponeringstiden bare 30s, og fisken våknet og reagerte når den ble fjernet fra behandlingskammeret. En mulig årsak til hvorfor mange fisk la seg over på siden allerede etter få sekunder i både forsøk 1 og 2 kan være at de allerede var slitne etter å ha vært håvet fra forsøkskar, trengt i et oppbevaringskar, og deretter håvet til behandlingskammeret. Trengingen og gjentatt håving av annen fisk i forsøkskar og oppbevaringskar kan ha vært en vesentlig stressfaktor for fisken som sammen med all den andre håndteringen virket utmattende på fisken.
Behandlingsgruppene mistet opptil 4,6 % av kroppsvekten sin over ~50 dager (forsøk 1) eller forble stasjonær (forsøk 2) i perioden fra forsøksstart til andre varmebehandling sammenlignet med den svake veksten av kontrollfisken som ikke gjennomgikk noen prosedyrer. Dette gjenspeiles også i kondisjonsfaktoren til behandlingsfisk, som også ble redusert i denne perioden, noe som tyder på at stressbelastningen ved gjentatt håndtering og behandling påvirket appetitt og/eller fôrkonvertering. Reduksjonen i velferdsstatus etter behandlinger kan også representere en kronisk stressor som vil undertrykke appetitten ytterligere. I henhold til Sørensen mfl. (2011) er det minst tre sannsynlige forklaringer på hvordan stress undertrykker vekst. For det første har kortisolbehandlinger (injeksjoner eller implantater) vist seg å forårsake en økning i basal metabolsk hastighet (målt som oksygenforbruk) hos fisk (Morgan og Iwama, 1996), noe som kan føre til reduksjon av mengden energi som kan brukes til vekst. For det andre kan kortisol direkte redusere cellulær vekst, ettersom [3H]-tymidin-inkorporering i DNA av cellekulturer avledet fra fisk er vist å reduseres etter kortisolbehandling (Pagniello mfl. 2002), og denne effekten er sannsynligvis mediert gjennom binding til glukokortikoidreseptorer (Lee og Bols, 1989). Og til slutt, har Barton mfl. (1987) vist at oral kortisolbehandling kan undertrykke vekst ved å endre tarmmorfologien, redusere næringsopptak fra maten og dermed begrense energien som er tilgjengelig for vekst. I tillegg til disse plausible forklaringene kan plasmakortisol ha en mer direkte effekt på veksten ettersom økning i kortisol under stress har vist seg å redusere veksthormon og insulinlignende vekstfaktorer (IGF-1) produsert av leveren, og dermed påvirke både matinntak og fôrkonverteringsraten (Peterson og Small, 2005; Salas-Leiton mfl. 2012).
At håndtering kan ha en negativ effekt på laks understrekes av at prosedyrekontrollen i forsøk 1 hadde 6,5 % dødelighet etter behandling, mot ingen dødelighet hos negativ kontroll gruppen som fikk stå i fred. Slik håndteringsrelatert dødelighet ble ikke registrert ved forsøket i Bodø. Håndtering er en nødvendig del av termisk avlusing, både i kommersiell og eksperimentell sammenheng, så skadene en observerte her er ikke nødvendigvis relatert til forsøksprosedyrene. I kommersiell produksjon vil fisken ikke bli flyttet med håv, men den vil bli trengt sammen, pumpet inn i behandlingssystemet, og transportert og presset gjennom ulike pumper, sklier og kammer der de risikerer å bli skadet. Selv om både operatører av avlusingssystemer og fiskeoppdrettere forsøker å begrense risikoen for fisken, rapporterer fiskehelsepersonell høy forekomst av skader på finner, øyne, gjeller, samt skjelltap og hudblødninger etter både termisk og mekanisk avlusing av oppdrettslaks (Sommerset mfl. 2022).
Både velferdsskårene av ytre skader og histologianalysene viste økt forekomst av øyeskade hos termisk behandlet fisk sammenlignet med kontrollfisken. Gismervik mfl. (2019) fant også økt forekomst av øyeskader, dog ved høyere temperaturer (34–38 °C) og lengre eksponeringstider (72–140 s). Tilsvarende viste Moltumyr mfl. (2021) at 3 av 39 laks behandlet ved 34 °C i 30 s hadde alvorlige øyeskader vs. 0 av 19 av kontrollfisken, og Moltumyr mfl. (2022) rapporterte at 12 % av laksen hadde alvorlige øyeskader to uker etter siste termiske behandling sammenlignet med 0 % i kontrollgruppen. Dette underbygger at eksponering for varmtvann gir risiko for skade. Det er imidlertid usikkert om det er det varme vannet i seg selv som skader øyet, om det er den eksplosive atferdsresponsen som forårsaker mekanisk skade, eller begge deler.
Poppe mfl. rapporterte i 2021 om en økning i forekomst av fisk med lesjoner i hjertet etter termisk og mekanisk lusebehandling, og oppfordret til at dette måtte følges opp med kontrollerte forsøk. Histologiundersøkelsene fra både forsøk 1 og forsøk 2, inkludert de utvidete undersøkelsene i forsøk 2, viste imidlertid ingen tegn på at prosedyrene eller det varme vannet hadde forårsaket patologiske forandringer i hjertevevet til fiskene. Dette tyder på at observasjonene til Poppe mfl. (2021ab) enten har en annen årsak, eller at forsøksoppsettet vårt ikke var tilstrekkelig tøft for fisken til å fremprovosere disse skadene. Det kan også være at dette er en type skade som opptrer bare i en liten andel av populasjonen. Hvis dette er en skade som f.eks. opptrer i 1 av 1000 fisk, er det liten sannsynlighet for at den vil opptre i vårt materiale. Imidlertid kan denne en-i-tusen fisken være akkurat det individet som helsepersonell i felt vil sende til laboratorier for videre undersøkelse. Enkeltfisken funnet med mulig gassopphopning inne i øyet i forsøk 1 kan også være et eksempel på et slikt sjeldent tilfelle, som ikke vil bli signifikant i et forsøk, men som likevel kan ha stor betydning i industrien hvor det kan være opp til 200 000 fisk i hver merd som avluses.
Resultatene av kronisk stresstest avslørte ingen forskjeller mellom noen av behandlingsgruppene og HPI-aksen var fult funksjonell i alle gruppene ved forsøksslutt. Dette tyder på at fisken ved slutten av forsøket var i stand til å håndtere det påførte stresset, og at de tidligere behandlingene ikke hadde kompromittert fiskens stressmestringsevne i alvorlig grad på lang sikt (Iversen og Eliassen, 2014).
5. Konklusjon
Resultatene fra forsøk 1 og 2 viser tydelig at laks har en kraftig atferdsrespons på varmt vann selv om de er 1-2 kg, og selv om de kommer fra en høy utgangstemperatur og har lav Δt. Fra forsøk 1 er det tydelig at laks som ikke er restituert, eller har andre iboende svakheter, har økt risiko for dødelighet med økt behandlingstemperatur, og resultatene fra forsøk 2 underbygger at hvis fisken er i god stand behøver det ikke opptre dødelighet etter termisk avlusing. Fremtidige studier bør derfor fokusere på å finne «helsemarkører» som oppdrettere og fiskehelsepersonell kan bruke til å avgjøre om en fiskegruppe er egnet for termisk avlusing eller ikke. Her kan det også være interessant å få mer klarhet i hva som gjør om en laks reagerer med «gi-opp»-atferd, og om dette er en atferd som gir mindre fysisk skade på fisk som blir termisk avluset. Det er også viktig å få klarhet i hvorfor laksens øyne synes å være utsatt for skade ved eksponering til varmt vann, hvordan disse skadene påvirker fisken, og hva som eventuelt kan gjøres for å forhindre skade på øynene.
6. Referanser
Ashley PJ, Sneddon LU, McCrohan CR (2007). Nociception in fish: stimulus–response properties of receptors on the head of trout Oncorhynchus mykiss. Brain Research 1166, 47-54.
Barton BA, Schreck CB, Barton LD (1987). Effects of chronic cortisol administration and daily acute stress on growth, physiological conditions, and stress responses in juvenile rainbow-trout. Diseases of Aquatic Organisms 2, 173-185.
Bui S, Madaro A, Nilsson J, Fjelldal PG, Iversen MH, Brinchman MF, Venås B, Schrøder MB, and Stien LH (2022). Warm water treatment increased mortality risk in salmon. Veterinary and Animal Science 17,100265.
Espmark ÅM, Kolarevic J, Åsgård T, Terjesen BF (2017). Tank size and fish management history matters in experimental design. Aquaculture Research 48, 2876-2894.
Gismervik K, Gåsnes SK, Gu J, Stien LH, Madaro A, Nilsson J (2019). Thermal injuries in Atlantic salmon in a pilot laboratory trial. Veterinary and Animal Science 8, 100081.
Grefsrud ES, Bjørn PA, Grøsvik BE, Hansen PK, Husa V, Karlsen Ø, Kvamme BO, Samuelsen OB, Sandlund N, Solberg MF, Stien LH (2022). Risikorapport norsk fiskeoppdrett 2022 - kunnskapsstatus - Effekter på miljø og dyrevelferd i norsk fiskeoppdrett, Rapport fra havforskningen 2022-13, 406 s.
Iversen M, Eliassen RA, Finstad B (2009). Potential benefit of clove oil sedation on animal welfare during salmon smolt, Salmo salar L. transport and transfer to sea. Aquaculture Research 40, 233-241.
Iversen MH, Eliassen RA (2014). The effect of allostatic load on hypothalamic-pituitary-interrenal (HPI) axis before and after secondary vaccination in Atlantic salmon postsmolts (Salmo salar L.). Fish Physiology and Biochemistry 40, 527-538.
Maule AG, Schreck CB, Bradford CS, Barton BA (1988). Physiological-effects of collecting and transporting emigrating juvenile Chinook salmon past dams on the Columbia river. Transactions of the American Fisheries Society 117, 245-261.
Moltumyr L, Gismervik K, Gu J, Gåsnes SK, Kristiansen TS, Rønnestad I, Nilsson J, Stien LH (2021). Does the thermal component of warm water treatment inflict acute lesions on Atlantic salmon (Salmo salar)? Aquaculture 532, 736048.
Moltumyr L, Nilsson J, Madaro A, Seternes T, Winger F, Rønnestad I, Stien LH (2022). Long-term welfare effects of repeated warm water treatments on Atlantic salmon (Salmo salar). Aquaculture 548, 737670.
Nilsson J, Gismervik K, Nielsen KV, Iversen MH, Noble C, Kolarevic J, Frotjold H, Nilsen K, Wilkinson E, Klakegg B, Hauge HS, Sæther PA, Kristiansen T, Stien LH. Laksvel— Standardisert operasjonell velferdsovervåking for laks i matfiskanlegg. Rapport fra havforskningen 2022-14, 40s.
Nilsson J, Moltumyr L, Madaro A, Kristiansen TS, Gåsnes SK, Mejdell CM, Gismervik K, Stien LH (2019). Sudden exposure to warm water causes instant behavioural responses indicative of nociception or pain in Atlantic salmon. Veterinary and Animal Science 8,100076.
Nygaard S, Klakegg B, Markussen Ø, Horsberg TE, Hamadi M, Persson D (2020). Tiltaksveileder kontroll med lakselus og skottelus. Lusedata.no
Overton K, Dempster T, Oppedal F, Kristiansen TS, Gismervik K, Stien LH (2019). Salmon lice treatments and salmon mortality in Norwegian aquaculture: a review. Reviews in Aquaculture 11, 1398-1417.
Pagniello KB, Bols NC, Lee LEJ (2002). Effect of corticosteroids on viability and proliferation of the rainbow trout monocyte/macrophage cell line, RTS11. Fish Shellfish Immunol 13, 199–214.
Pickering AD, Pottinger TG (1989). Stress responses and disease resistance in salmonid fish: effects of chronic elevation of plasma cortisol. Fish Physiology and Biochemistry 7, 253-258.
Poppe TT, Reed AK, Hofossæter M, Wisløff H (2021a). Novel myocardial pathology in farmed salmonids. Journal of Fish Diseases 44, 1657-1660.
Poppe TT, Wisløff H, Reed AK, Hofossæter M (2021b). Alvorlige hjerteskader på oppdrettslaks etter termisk og mekanisk behandling mot lakselus. Norsk Veterinærtidsskrift 3,150–153.
Pottinger TG, Carrick TR (2001). ACTH does not mediate divergent stress responsiveness in rainbow trout. Comparative Biochemistry and Physiology Part A-Molecular & Integrative Physiology 129, 399-404.
Salas-Leiton E, Coste O, Asensio E, Infante C, Cañavate JP, Manchado M (2012). Dexamethasone modulates expression of genes involved in the innate immune system, growth and stress and increases susceptibility to bacterial disease in Senegalese sole (Solea senegalensis Kaup, 1858). Fish & Shellfish Immunology 32, 769-778.
Sommerset I, Walde CS, Jensen BB, Wiik-Nielsen J, Bornø G, Oliveira VHSD, Haukaas A, Brun E (2022). Veterinærinstituttet, Rapport 21/2022, 209s.
Sørensen C, Bohlin LC, Øverli Ø, Nilsson GE (2011). Cortisol reduces cell proliferation in the telencephalon of rainbow trout (Oncorhynchus mykiss). Physiology & Behavior 102, 518-523.
Vollset KW, Lennox RJ, Thorstad EB, Auer S, Bär K, Larsen MH, Mahlum S, Näslund J, Stryhn H, Dohoo I (2020). Systematic review and meta-analysis of PIT tagging effects on mortality and growth of juvenile salmonids. Reviews in Fish Biology and Fisheries 30, 553-568.