In this report, the knowledge base for being able to estimate migration routes for wild postsmolt of Atlantic salmon is assessed, and whether these overlap with three planned areas for offshore fish farming. The knowledge about migration routes of post and what factors that can influence the post smolt's migration choice comes mainly from experiments with marking and recapture of smolt. There is data both from the recapture of fish marked with external tags, and telemetry experiments with acoustic marking and registration on listening buoys. Also, from pelagic trawling in the sea, there is data from over 9,000 catches of postsmolt over the years, which can give a picture of how the postsmolt is distributed in the ocean in different time periods. Such data have been used to develop an individual migration model for the Norwegian postmolt that is presented in this report. The model estimates the likely migration route for postsmolt from different parts of southern and central Norway and quantifies the likely overlap between the planned aquaculture areas and the post smolt's migration.
Vurdering av kunnskapsgrunnlaget for bestemmelse av vandringsruter for vill postsmolt av atlantisk laks i Norge
— Potensielle interaksjoner med planlagte områder for oppdrett til havs
Rapportserie:
Rapport fra havforskningen 2024-20
ISSN: 1893-4536
Publisert: 23.05.2024
Oppdatert: 01.12.2025
Prosjektnr: 15976
Oppdragsgiver(e): Nærings- og Fiskeridepartementet
Referanse: 24/185-2
Forskningsgruppe(r):
Populasjonsgenetikk
,
Smittespredning og sykdom
Tema:
Havbruk til havs,
Lakselus
Program:
Marine prosesser og menneskelig påvirkning,
Miljøeffekter av akvakultur
Approved by:
Research Director(s):
Geir Lasse Taranger
Program leader(s):
Frode Vikebø og Mari Skuggedal Myksvoll
English summary
Sammendrag
I denne rapporten vurderes kunnskapsgrunnlaget for å kunne estimere vandringsruter for vill postsmolt av atlantisk laks, og om disse overlapper med tre planlagte områder for oppdrett til havs. Kunnskapen om vandringsveier til postsmolt ut i havet og hva som kan påvirker postsmoltens vandringsvalg kommer i hovedsak fra forsøk med merking og gjenfangst av smolt. Det foreligger data både fra gjenfangst av fisk merket med ytre merker, og telemetriforsøk med akustisk merking og registrering på lyttebøyer. Fra pelagisk tråling i havet foreligger det data fra over 9 000 fangster av postsmolt gjennom årene, som kan gi et bilde på hvordan postsmolten fordeler seg i havet i ulike tidsperioder. Slike data er brukt til å utvikle en individuell vandringsmodell for norsk postmolt som presenteres i denne rapporten. Modellen estimerer sannsynlig vandringsrute for postsmolt fra ulike deler av Sør- og Midt-Norge og kvantifiserer sannsynlig overlapp mellom de planlagte havbruksområdene og postsmoltens vandring.
1 - Innledning
I forbindelse med at det nå åpnes opp for havbruk til havs (HTH) i tre områder utenfor kysten har det fra flere hold blitt pekt på risiko for potensiell påvirkning på utvandrende vill postsmolt av atlantisk laks fra elvene forbi disse områdene. Dette gjelder både fra norske lakseelver og også fra elver lenger sør i Europa. Dersom HTH medfører økte utslipp av lakselus eller annen smitte, vil dette kunne medføre redusert overlevelse/økt dødelighet på utvandrende postsmolt dersom de beveger seg gjennom HTH-områdene eller smittefeltene nedstrøms. Modellsimuleringer av utslipp av lakseluslarver fra HTH-områdene viser at spredningen av luselarver kan ha stor geografisk utstrekning (Ådlandsvik 2019). Dersom oppdrettsaktiviteten på kysten opprettholdes på nåværende nivå, vil dette kunne resultere i en form for «dobbeltsmitte» for postsmolt fra norske elver, dvs. økt dødelighet som følge av smitte fra kystnært oppdrett, og ytterligere påført dødelighet gjennom smitte fra HTH-områder. Det er derfor viktig, i den grad det er mulig, å vurdere hvordan vandringsrutene for postsmolt overlapper med modellerte smittefelt fra HTH-områder.
Mens det gjennom overvåkningsprogrammer som lakselusovervåkningen NALO estimeres påvirkning på vill postsmolt av laks og aure i fjord- og kyststrøk, vil det være mer komplisert å estimere effekten av smitte fra oppdrettsanlegg plassert langt fra kysten. Vurdering av slike effekter vil derfor nødvendigvis i stor grad måtte baseres på modeller. Innsamling av konkrete data fra postsmolt i disse regionene og i havet utenfor vil være svært dyrt og vanskelig å gjennomføre da post-smolten utenfor kysten sprer seg utover et større geografisk område enn den gjør i fjordene. Presisjonen i modellene vil derfor være avgjørende for hvorvidt det faktisk er mulig å gjøre en realistisk konsekvensvurdering i henhold til kriteriene i Forskrift om konsekvensutredninger av 22. juni 2017 nr. 854 (KU-forskriften). Modeller må bygges på eksisterende kunnskap og valideres. Konkret kunnskap om postsmoltens vandring ut til beiteområdene i havet er begrenset. Gjennom telemetriforsøk med merket fisk, tråling og modellering av vandring i NALO-programmet har man noe kunnskap om hvordan postsmolten vandrer gjennom fjorder og ut til grunnlinjen. Fra trålfangster over mange år har man også noe kunnskap om hvordan postsmolten er fordelt i havet, men kunnskapen om vandringrutene videre ut til beiteområdene er i hovedsak ukjent. Med hensyn til laksens vandring i havet stammer det meste vi har av kunnskap fra trålfangster av postsmolt og større laks, til dels med genetisk informasjon som angir hvilken region de kommer fra (Gilbey mfl. 2021), og merking av vinterstøinger med ulike typer av datalagringsmerker (Rikardsen mfl. 2021, Strøm mfl. 2018).
I denne rapporten ser vi på det eksisterende datagrunnlaget for kartlegging av vandringsruter for postsmolt og vurderer hvordan det kan benyttes i vurderingene av potensielle konsekvenser av de planlagte HTH-områdene for vill postsmolt på vandring ut i havet. Vi gjennomgår eksisterende datasett over postsmoltfangster nær disse områdene, og i den grad det er tilgjengelig ut fra genetiske data, ser vi på hvilke bestander/lakseregioner som er registrert. Vi presenterer også en vandringsmodell som beskriver vandring av postsmolt fra ulike deler av landet i forhold til de planlagte HTH-områdene. Vi gjør også en vurdering av hvilke tilnærminger som kan benyttes for å øke kunnskapen om postsmoltvandring og ser på fordeler og ulemper ved ulike metoder.
2 - Metodikk og tilnærming
2.1 - Kilder til kunnskap om fordeling og vandring til postsmolt
Som nevnt ovenfor er det i hovedsak tre kilder til kunnskap om postsmoltens vandring ut til beiteområdene i havet.
-
Merking av smolt i elvene. Enten med akustiske merker og registrering på lyttebøyer plassert i sjøen, eller gjenfangst i sjøen av fisk med ytre merke. Det er gjennomført mange slike forsøk i ulike regioner av landet, og disse forsøkene har gitt oss kunnskap om svømmehastighet, oppholdstid i munningsområder, og vandring i fjord og kyststrøk. Noen av disse studiene har også relatert vandringsmønster til fysiske forhold som tidevannssyklus, strøm, temperatur og salinitet. Potensielt kan kunnskap fra disse studiene om hva som påvirker postsmoltens vandringsvalg kunne bidra til økt forståelse av hvilke vandringsvalg postsmolten gjør også utenfor kysten. Se nærmere drøfting nedenfor.
-
Fangst av postsmolt på kysten og i havet. Trålfangster av postsmolt i havområdene er blitt registrert gjennom mange år. Til dels er dette tilfeldige bifangster, i hovedsak fra forskningstokt rettet mot kartlegging av andre pelagiske fiskearter, men det foreligger også noen registreringer av fangst av postsmolt fra kommersielle fiskefartøy. Det har også vært gjennomført målrettete tokt for å kartlegge utbredelsen av postsmolt i Norskehavet (Holm mfl.2000). Fordeling av de registrerte fangstene av postsmolt i Nord-atlanteren, over 9000 postsmolt gjennom årene, ble oppsummert i en artikkel fra Gilbey mfl. (2021).
-
Utvikling av individbaserte vandringsmodeller som benytter eksisterende kunnskap om vandringshastighet og adferd ut fra telemetriforsøk, strømmodeller, og andre parametere som kan påvirke postsmoltens adferd (som f.eks. temperatur, salinitet, mengde byttedyr) til å beskrive observert fordeling av postsmolt fra ulike regioner.
Nedenfor beskriver vi kunnskapsstatus innenfor disse tre kategoriene før vi oppsummerer og ser nærmere på hvordan denne kunnskapen kan forbedres ytterligere.
3 - Kunnskapsstatus basert på eksisterende data
3.1 - Merkeforsøk
Merking av smolt med ulike merketyper har vært gjennomført i mange vassdrag og for noen vassdrag foreligger det også lange tidsserier fra slik merking. Tidlige merkestudier baserte seg på passive ytre merker, eller såkalte snutemerker (magnetisk metalltråd i nesen på fisken) og gjenfangst av fisken, enten når den kom tilbake til elva der den var merket, eller fra gjenfangster i sjøen og havet. Deteksjon av snutemerker krever at fisken blir scannet med en detektor, og merket må opereres ut og avleses med lupe, mens ytre merker som Carlin-merker og Floy-merker kan avleses uten at fisken avlives.
Registering av gjenfangster i sjøen av slike merker har bidratt med noe kunnskap om vandringsruter, men i hovedsak har det gitt informasjon om beiteområder i havet hvor det har foregått fiske. Mange år med slike merkeforsøk ble oppsummert i en rapport fra en workshop i ICES (WKLustre, Ó Maoiléidigh mfl. 2018). Selv om disse merkeforsøkene har økt kunnskapsgrunnlaget, er antall gjenfangster fra de ulike vassdragene hvor det ble merket smolt relativt lavt i forhold til antallet fisk som ble merket. De fleste av gjenfangstene i tidligere perioder var fra havfiske etter laks ved Færøyene og ved Grønland, det vil si at det i hovedsak er voksen laks som er fanget. Samlet sett gir datasettene fra disse merkeforsøkene med passive merker derfor svært begrenset informasjon som kan bidra til kartlegging av kystnær vandring av postsmolt.
Merking av smolt med akustiske merker, med tilhørende nettverk av lyttebøyer i elv, munning og fjord har vært benyttet i en rekke vassdrag i Norge (se Thorstad mfl. 2004). Slike merker kan i prinsippet gi svært presis informasjon om vandring av postsmolt dersom nettverket av lyttebøyer er omfattende og avstanden mellom lyttebøyene er liten. Slike merker gir mulighet til å studere fiskens adferd i forhold til variasjon i miljøparametre som tidevann, temperatur, salinitet og strøm, og generell kunnskap om hvilke faktorer som påvirker fiskens vandringsvalg kan utledes og modelleres.
Felles for disse merkeforsøkene er at de i all hovedsak er gjennomført inne i fjordene, og derfor har begrenset relevans for postsmoltens vandring utenfor grunnlinjen (men se Rodger mfl. 2024). Man kan anta at kunnskap om hvilke miljøforhold som påvirker vandringsvalgene i fjordene ikke er direkte overførbart til vandring ut i havet. I havet, hvor variasjon i miljøparametere skjer i en større geografisk skala, og hvor postsmolten ikke har kontakt med bunnstrukturer, er det sannsynligvis andre forhold som påvirker vandringen. Det er f.eks. vist for laksearter i Stillehavet at jordmagnetisme påvirker vandringsretningen til postsmolt (Putman 2015), og det er ikke usannsynlig at det samme gjelder for Atlantisk laks.
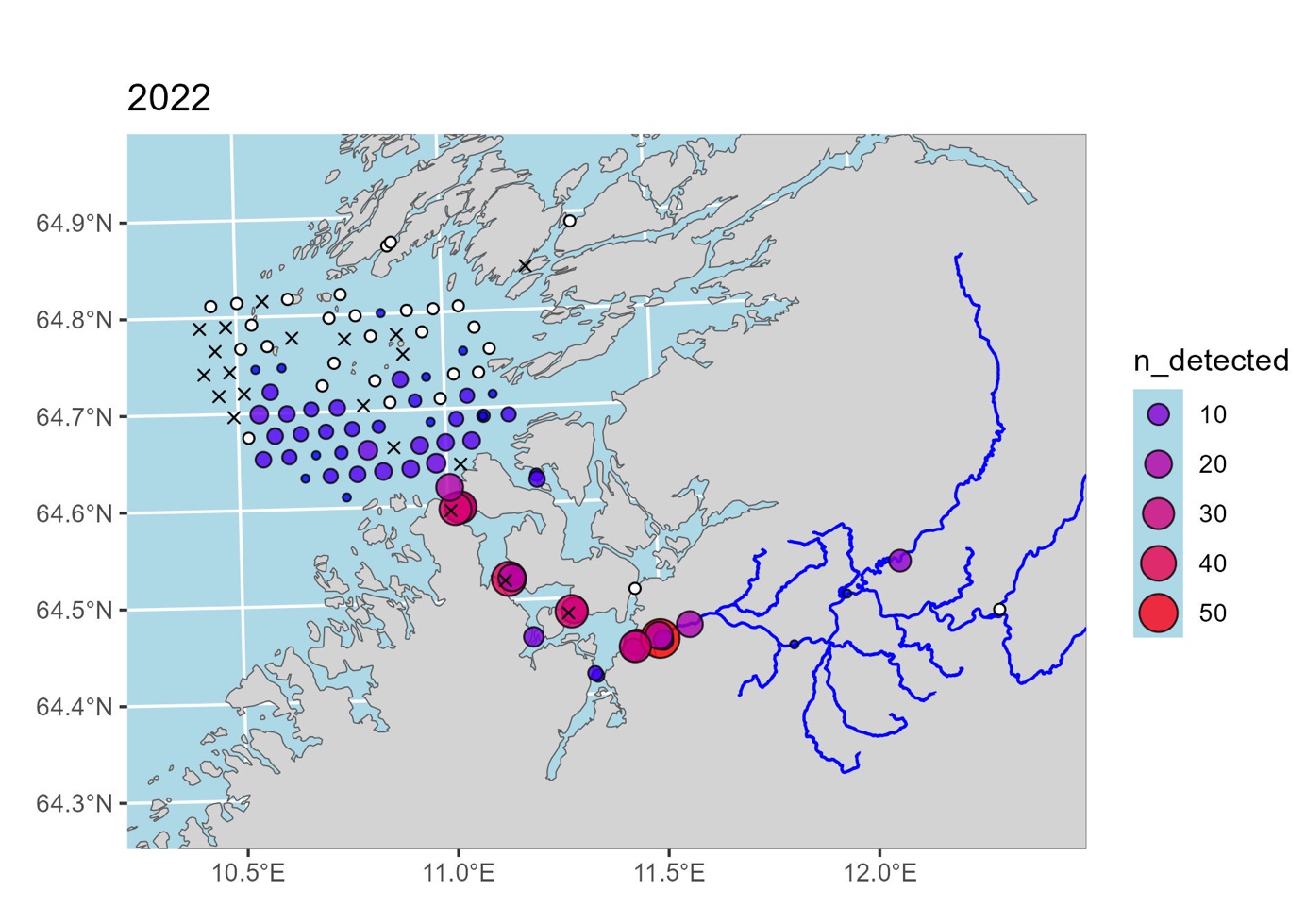
Av telemetriforsøk med akustisk merking utført i Norge er sannsynligvis forsøkene med merking av smolt i Namsen og kartlegging av vandringen ut fra elva, gjennom fjorden og videre ut mot havet sør for Vikna av mest relevans. Dette forsøket ble utført av NALO-programmet og følger postsmolten lenger ut mot havet enn andre telemetriforsøk vi kjenner til i Norge. Resultatene fra merking og registrering av merker på lyttebøyer i 2022 og 2023 er fortsatt under bearbeiding, men de foreløpige resultatene fra registreringen er vist i figur 1 og figur 2.
I 2022 ble det merket 215 smolt fanget Namsen med smoltfelle eller ved elektrofiske. Av disse ble 145 fisk registrert på lyttebøyer utenfor elva, mens 79 fisk ble registrert på lyttebøyene utenfor fjorden. Registreringene utenfor fjorden var hovedsakelig fordelt på de sørlige lyttebøyene, noe som tyder på at postsmolten ikke vandret nordover gjennom sundene i Vikna, men hadde retning mer direkte vestover.
I 2023 ble 174 smolt fanget i smoltruse og merket i Namsen. Det ble plassert ut totalt 89 lyttebøyer i elv, elvemunning, indre fjord og utenfor fjorden. Det ble registrert 102 fisk på lyttebøyer utenfor elva, og av disse ble 32 fisk registrert på de ytre lyttebøyene (se figur 2). Igjen viser fordelingen av registreringer at fisken ser ut til å velge en vestlig rute når den kommer ut av fjorden.
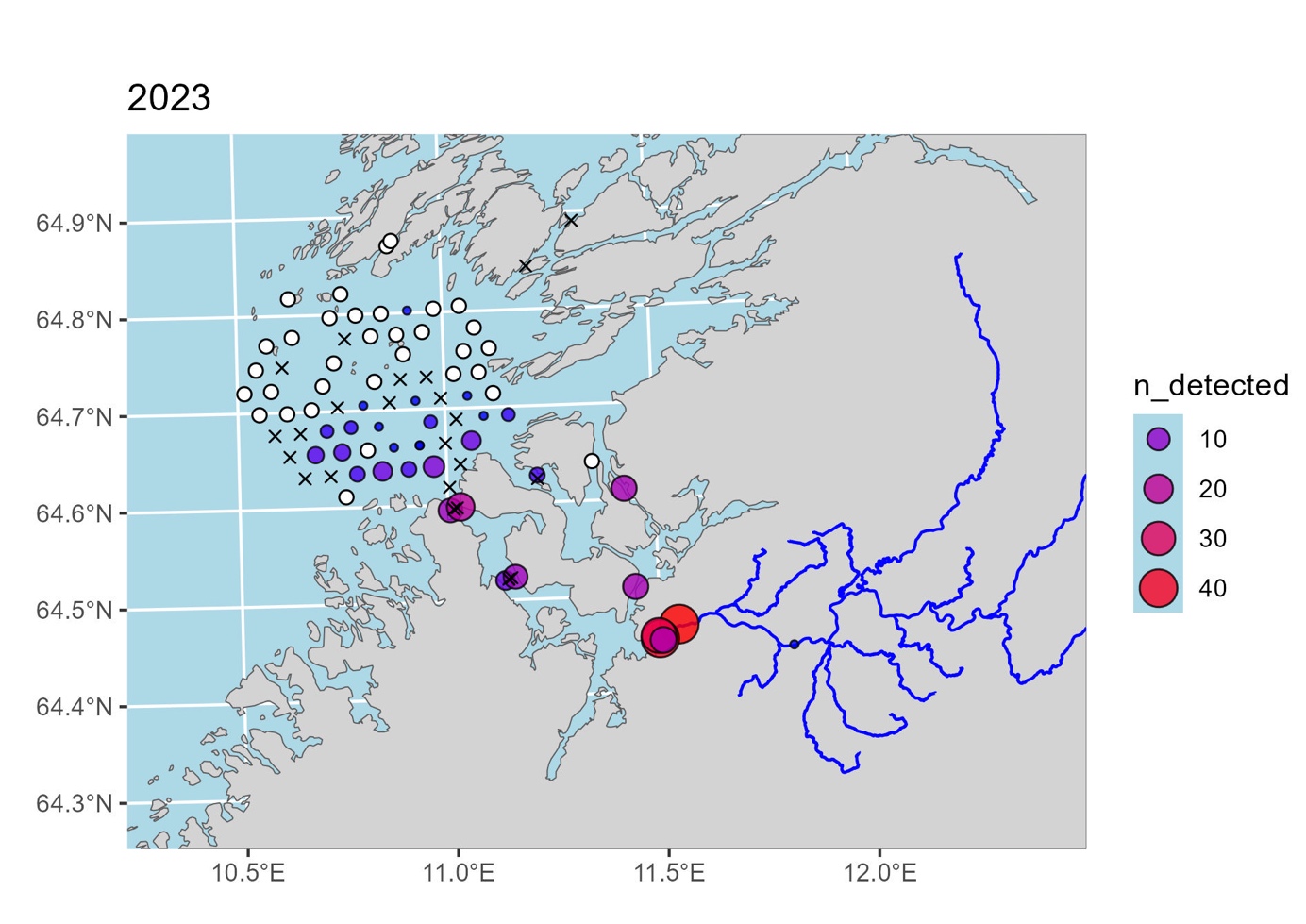
Samlet sett viser disse to telemetriforsøkene at postsmolt fra Namsen ser ut til å velge en vestlig rute ut av fjorden, og ikke følger kysten nordover mot Vikna. Dette er foreløpige data og analyser av dette datasettet pågår fortsatt. I den videre analysen vil man blant annet se på hvordan strømmen har vært i området i den aktuelle utvandringsperioden, og salinitet- og temperaturgradienter. Andre telemetriforsøk kan tyde på at salinitetgradienter og temperatur kan være en parameter som påvirker vandringsvalg hos postsmolt på vei ut av fjorden (Jensen mfl. 2022). Når resultatene fra disse analysene forligger vil man ha noe mer kunnskap om laksens vandring i mer åpne områder ut mot havet.
Selv om kunnskapen om miljøvariabler og postsmoltvandring har økt gjennom slike forsøk, kan ikke nødvendigvis denne kunnskapen benyttes til å forutsi vandringsruter i havet. Det er ikke gjennomført telemetriforsøk i Norge som har kartlagt vandring utenfor kysten, men slike forsøk er gjennomført på Island. Totalt 598 klekkerismolt ble merket med DST-merker (Data Storage Tags) som registrerte dyp og temperatur (Gudjonsson mfl. 2015). Smolten ble satt ut i 2005 og 2006 i elva Kidafellsa sørvest på Island. I alt sju individer ble gjenfanget i elva i 2006 og 2007 etter ett år i havet, og data lagret i merkene ble brukt til å estimere vandringsruter i havet. Resultatene viste hvordan postsmolten vekslet mellom å bruke Irmingerhavet som beiteområdet om sommeren, mens de trakk østover mot Irland i vintersesongen. Så langt, er dette det eneste datasettet av denne typen som har gitt kunnskap om en full vandringssyklus fra postsmolt, via beiting i havet, og tilbake til elva som gyteklar voksen laks. Men datasettet har klare begrensninger fordi posisjoner er estimert kun fra temperatur og dybdedata, noe som gir stor usikkerhet i posisjonsbestemmelsen. Og gjenfangstraten var svært lav, med kun sju individer av 698 tilbake til elva.
3.2 - Trålfangster av postsmolt
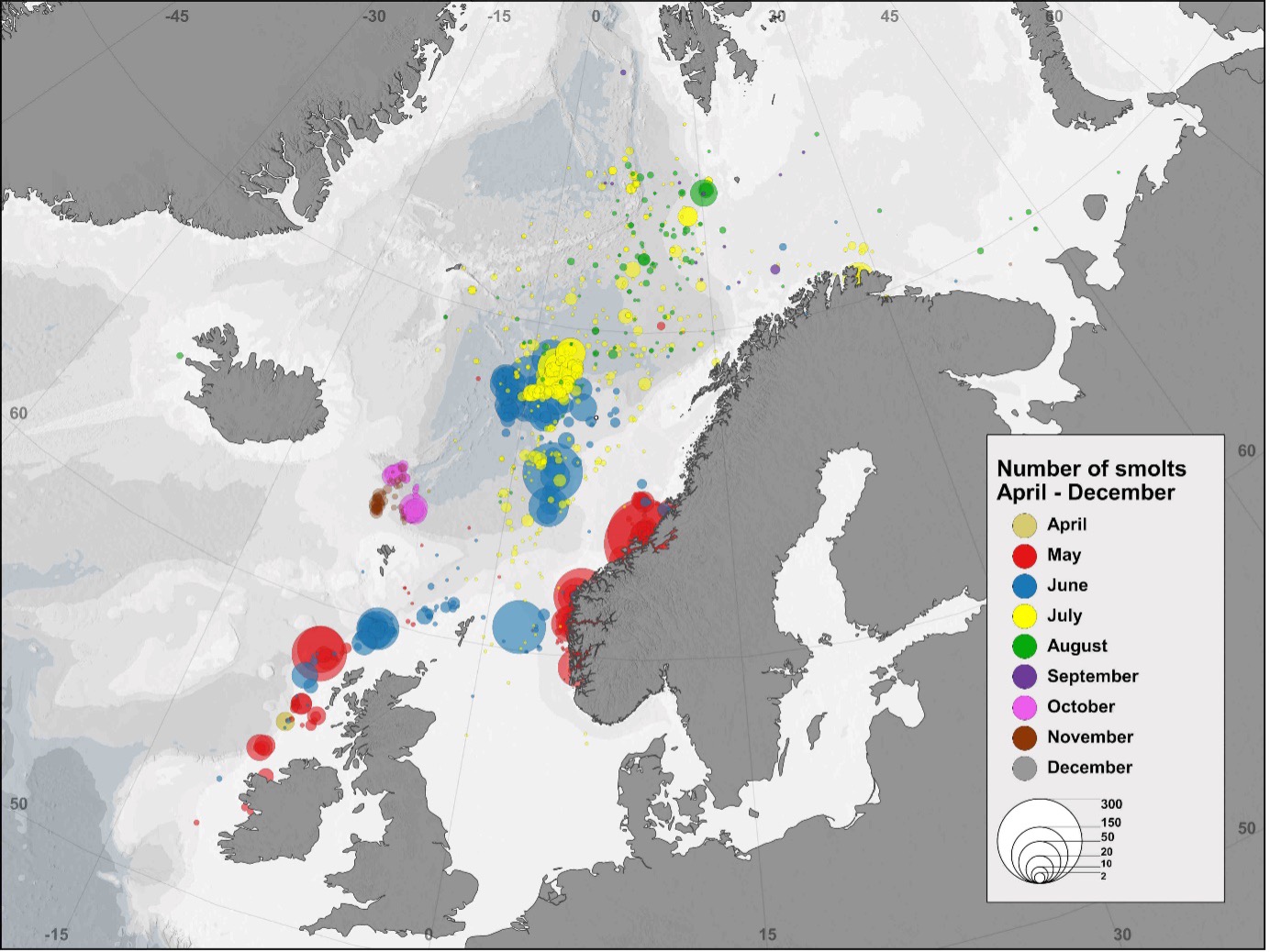
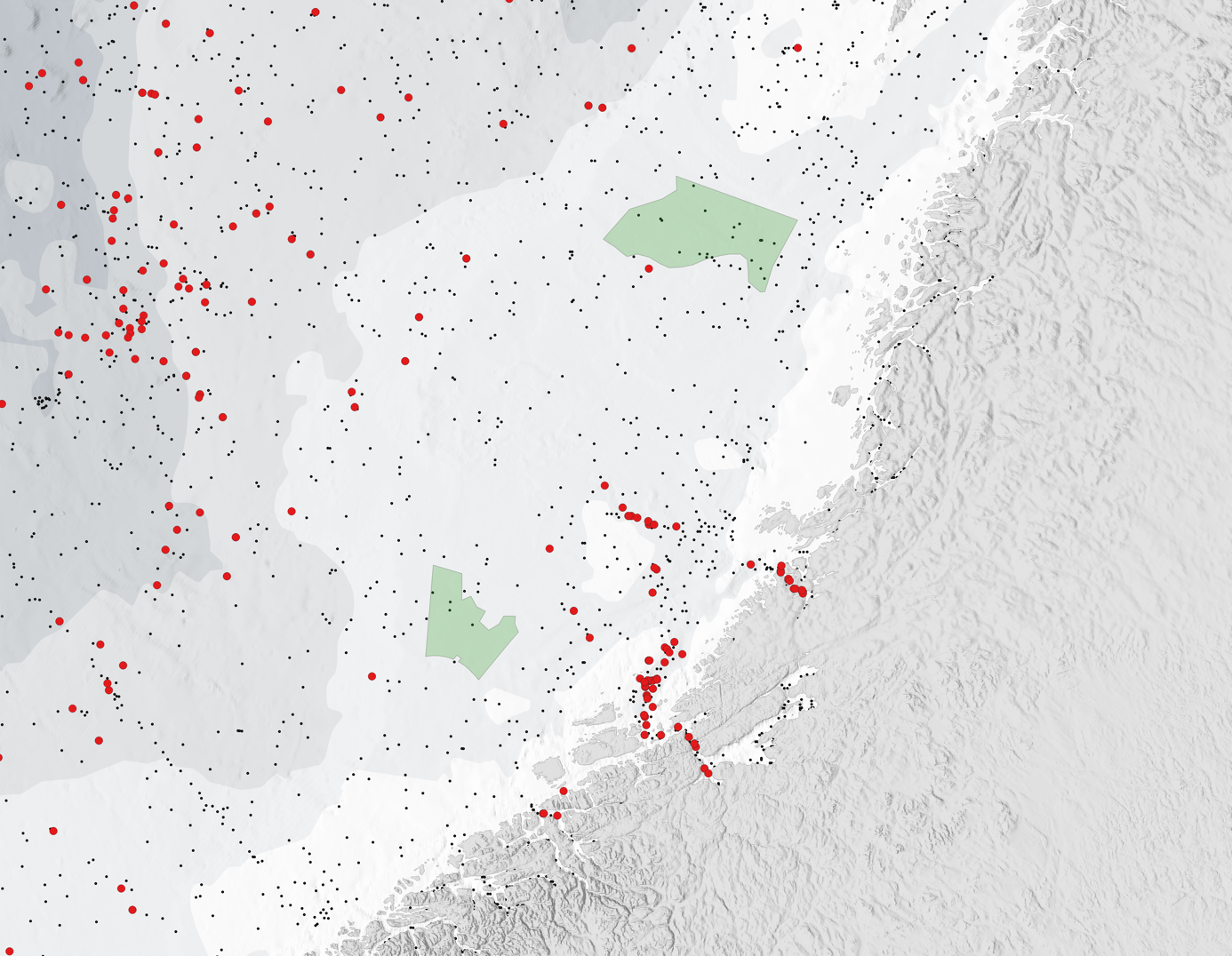
Mens forskningsinnsats og kunnskapsoppbygging om ferskvannsfasen av laksens livssyklus og vandring i fjordene på vei ut fra elvene har vært gjennomført gjennom svært lang tid, er det først i nyere tid at kunnskapen om laksens beitevandringer i havet har blitt utviklet. Laksens beiteområder i havet ved Færøyene og Grønland har vært kjent, og det har vært (Færøyene) og er (Vest-Grønland) et kommersielt fiske i disse områdene. Også langs Norskekysten har det vært et havfiske med line etter tilbakevandrende laks på vei til vassdragene og samlet har disse fiskeriene gitt noe informasjon om beiteområdene for stor laks i havet. Kunnskapen om hvordan postsmolten vandrer ut i havet har vært mer begrenset, men registreringer av tilfeldige trålfangster og målrettet tråling etter postsmolt fra 1990 tallet har økt kunnskapen betraktelig om geografisk utbredelse i Norskehavet (Holst mfl. 1993, Shelton mfl. 1997, Holm mfl. 2000, Gilbey mfl. 2021). Postsmoltfangster gjennom årene registrert i databasen ved Havforskningsinstituttet og samarbeidende institusjoner ble analysert og presentert i en vitenskapelig artikkel som oppsummerte data fra over 9000 fangster av postsmolt (Gilbey mfl. 2021). Figur 3 viser fordelingen av disse fangstene, i postsmoltperioden fra april – desember. Det generelle bildet er at mens postsmoltfangstene i i mai i hovedsak er tatt nært kystene, er det størst konsentrasjon i et område i Norskehavet ved Vøringplatået i juni-juli, før postsmolten trekker lengre nord i Norskehavet utover i juli og august. Det er få registreringer i perioden september til desember, i hovedsak fordi det er få pelagiske forskningstokt i denne perioden.
For en del av disse fangstene foreligger det genetiske prøver som har blitt analysert og sammenlignet med en genetisk database over Europeiske laksebestander utviklet i EU-prosjektet SALSEA-Merge (Gilbey mfl. 2017). Dette gir en mulighet for å si noe om vandringsrutene til postsmolt fra ulike regioner i Europa. Analysene utført av Gilbey mfl. (2021) viste at forekomsten av norsk postsmolt ved Vøringplatået i juni-juli var noe mindre enn det man kunne forvente ut fra estimater av smoltproduksjon i ulike Europeiske regioner, og at hovedmengden postsmolt her hadde opprinnelse fra mer sørlige bestander, i hovedsak fra Storbritannia og Irland. Dette kan tyde på at hovedmengden av norsk postsmolt fra Sør-Norge vandrer nordover nærmere Norskekysten, noe som potensielt gjør at de kan komme innenfor influensområdet til HTH-områdene.
Kartet i figur 4 viser postsmoltfangster registrert i nærheten av HTH-område 2, Norskerenna Sør. Det er få fangster registrert utenfor Sørvest-Norge, selv om det gjennom årene er foretatt mange pelagiske trålhal. Dette skyldes sannsynligvis at den pelagiske trålingen i hovedsak har foregått etter juni, og dermed utenfor den mest aktuelle perioden for postsmoltvandring. Genetisk tilordning foreligger for noen av fangstene, og denne viser at individene nærmest kysten er postsmolt med opprinnelse i Sørvest-Norge, men det er også noen individer med mer sørlig opprinnelse i omtrent samme avstand fra kysten som HTH-område 2.
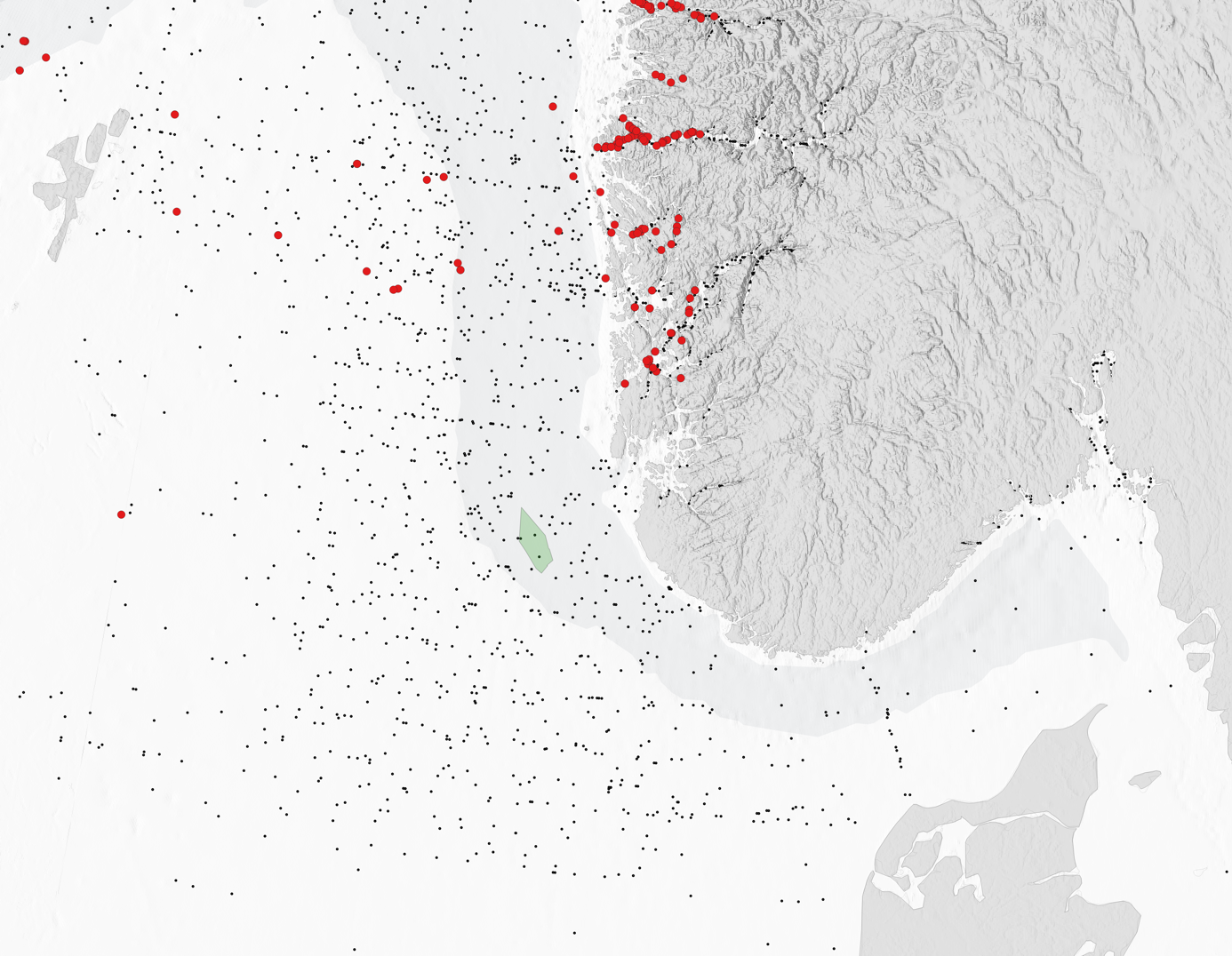
Kartet i figur 5 viser postsmoltfangster registrert i nærheten av HTH-område 11 Frøyabanken og HTH-område 5 Trænabanken. For postsmoltene hvor det foreligger genetisk tilordning er alle individer i og utenfor Trondheimsfjorden og Namsfjorden av norsk opprinnelse. Ett individ rett vest for HTH-område 5 Trænabanken er av sørlig opprinnelse (Storbritannia/Irland). Det er få fangster nært kysten, men fordelingen nordover fra Trondheimsfjorden kan tyde på en relativ kystnær initiell vandring nordover for postsmolt fra denne fjorden, noe som potensielt kan medføre kontakt med HTH-område 5.
4 - Fra direkte observasjoner til modeller
Gjenfangster av merket fisk og trålfangster av laks utenfor kysten, til dels med genetisk tilordning til region, har gitt oss direkte observasjoner av hvor postsmolt fra ulike elver og regioner befinner seg på et gitt tidspunkt. Slike observasjoner kan anvendes i individbaserte vandringsmodeller, hvor man kan undersøke hvordan en rekke potensielle påvirkningsfaktorer vil kunne endre fiskens vandring, og også identifisere hvilke parametere i modellen som gir en romlig og temporær fordeling av postsmolten som samsvarer mest med de direkte observasjonene. Vi har utviklet en individbasert vandringsmodell som har gjort det mulig å simulere hvorvidt vandringsrutene for utvandrende norsk postsmolt av vill Atlantisk laks (Salmo salar) overlapper med de foreslåtte HTH-områdene, og kvantifiserer forsøksvis andelen av postsmolt fra ulike deler av landet som vi passere gjennom HTH-områdene. I det følgende beskriver vi hvordan modellen er bygd opp, og resultater fra disse simuleringene.
4.1 - Metode
Den individbaserte modellen er en numerisk modell med romlig oppløsning som simulerer post-smoltens vandring gjennom havet, og der miljøforhold og valgte mekanismer påvirker post-smoltens vandringsadferd. Miljøforholdene i havet simuleres av den oseanografiske modellen ROMS (Regional Ocean Modeling System, Shchepetkin og McWilliams 2005) som har en geografisk oppløsning på 4 km horisontalt og 10 m vertikalt. De årlige kjøringene er hindcast-kjøringer (gjenskapte historiske oseanografiske forhold) t.o.m 2011 (Lien mfl. 2013).
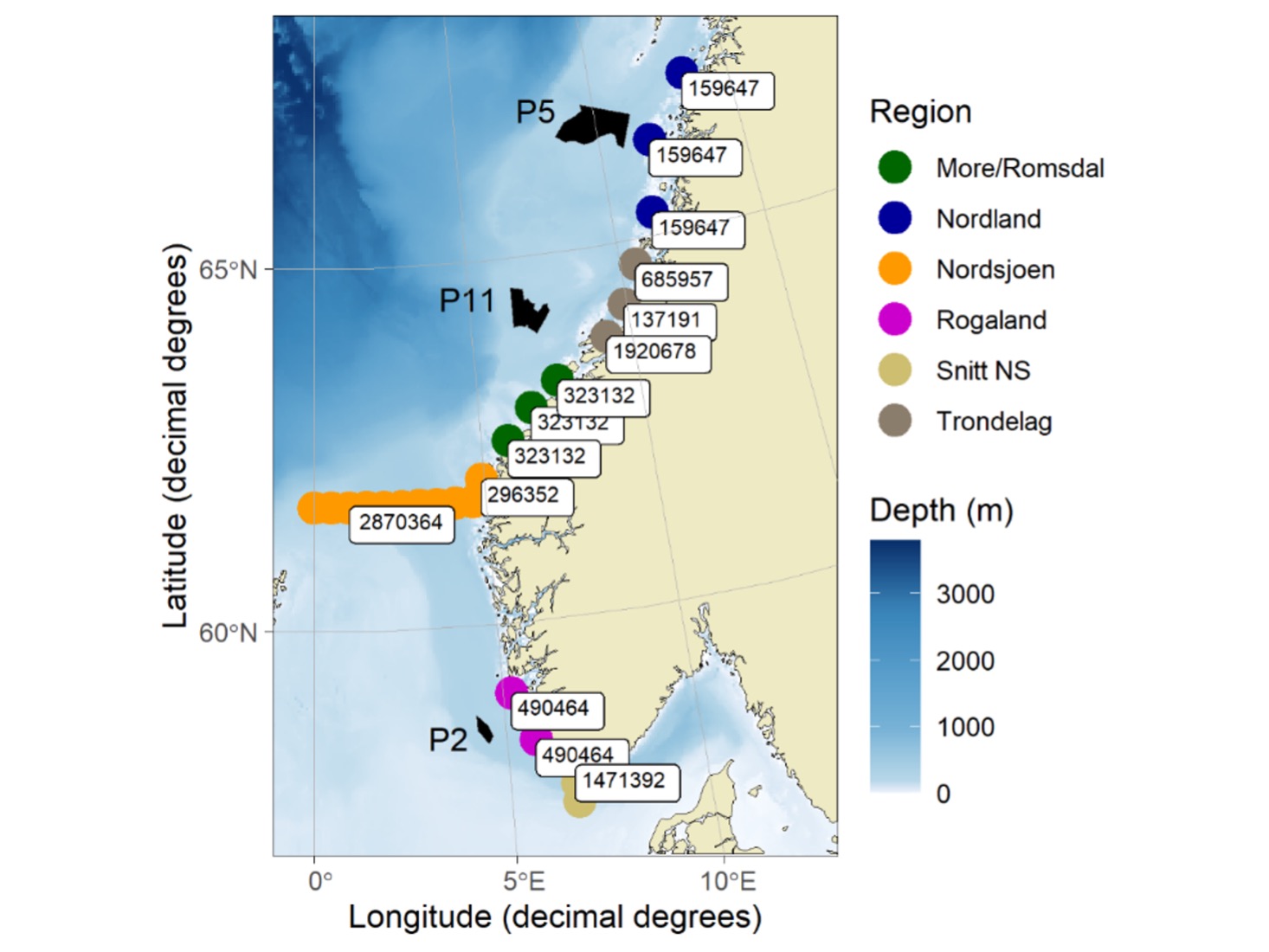
I simuleringene som ble gjennomført for dette formålet starter post-smolten vandringen ved grunnlinjen utenfor kysten. Den første vandringen gjennom fjorder blir dermed ikke simulert i denne modellen. Postsmolten blir initiert ved noen sentrale lokasjoner fra sørspissen av Norge til sentral/nordlige deler av Nordland (Figur 6). Det totale antallet postsmolt pr fylke er hentet fra Nilsen mfl. (2017), og antallet individer blir omtrentlig fordelt på utslippslokasjonene i henhold til antallet og størrelsen på elvene som utslippslokasjonene representerer. I modellen blir for eksempel 70% av post-smolten fra Trøndelag sluppet utenfor Trondheimsfjorden. Modellen benytter «superindivider», der ett simulert individ representerer 600 individer. De valgte utslippslokasjonen er i nærheten av de foreslåtte HTH-områdene. For også å kunne vurdere hvorvidt postsmolt som kommer fra elver lengre borte fra HTH-områdene overlapper med disse, ble det initiert postsmolt langs to ulike tverrsnitt i Nordsjøen. Bakgrunnen for dette var å redusere avstanden fra startposisjonen (dvs. elvemunningen) til HTH-områdene, da usikkerheten knyttet til simulert vandring øker med tid og avstand. For å vurdere overlappet mellom postsmolt fra Sør-Norge og de to nordligste HTH-områdene ble postsmolt initiert i nordlige Nordsjøen (61˚ 42’ N, 0˚ - 4˚ 36’ Ø), og antallet postsmolt representerer produksjonen i alle fylker med elver som renner ut i Nordsjøen. For å vurdere det sørligste HTH-området ble postsmolt initiert langs et tverrsnitt ved sørspissen av Norge (57˚ 30’-58˚ 00’ N, 6˚ 30’ Ø), der antallet postsmolt representer produksjonen i fylkene fra Østfold til Agder. Datoen individene blir initiert i modellen følger en normalfordeling med standardavvik på 10 dager, der gjennomsnittlig utvandringstidspunkt er senere på året ved høyere breddegrad (Vollset mfl. 2021). Størrelsen på individene ved initiering i modellen er normalfordelt med forventning på 14,0 ± 1,0 cm og 30 ± 7 g. Post-smolten har i modellen ingen interaksjon med byttedyr eller predatorer og en fast lengdevekst på 0,12 cm pr dag.
Fra et utvalg av 10 migrasjonsadferder (søk mot optimal temperatur, høyere byttedyrskonsentrasjoner, optimal saltholdighet, dypere farvann, vandring med havstrømmer eller en gitt himmelretning, eller en kombinasjon av disse faktorene) var en migrasjonsadferd uttrykt som en kombinasjon av aktivt søk etter havområder med bunndyp på over 800m, svømming med havstrømmer (negativ rheotaxis) og en tilfeldighetskomponent best til å gjenskape utbredelsen av postsmolt fanget i Norskehavet som ble genetisk identifisert til å stamme fra sør- og Vest-Norge (ca fra Telemark til Troms, Gilbey mfl. 2021). Den tilfeldige komponenten skaper naturlig variasjon mellom individer knyttet til individuell adferd eller indre motivasjon. I tillegg til postsmolten aktive vandringsadferd vil den være utsatt for passiv horisontal drift med havstrømmene. Vandringsmønsteret til postsmolt i kystnære farvann blir diskutert mer i detalj under «Sammenhengen mellom miljøforhold i havet og vandringsmønsteret til postsmolt i kystnære farvann». Hvor fort postsmolt forflytter seg i havet foreligger det begrenset kunnskap om, men det varierer trolig i tid og rom (Thorstad mfl. 2012). I modellen ble det kjørt tre ulike sett med simuleringer, der vandringshastigheten var satt til henholdsvis 1, 1,5 eller 2,0 kroppslengde i sekundet (Tabell S1 a-c). Hvert sett av kjøringer var med gjenskapte oseanografiske forhold for årene 2001-2011. Ved å kjøre simuleringene for flere år blir det inkludert mellomårlig variasjon i de oseanografiske forholdene, noe som gir et mer fullstendig bilde av hvordan f.eks. variasjonen i havstrømmene påvirker postsmoltens vandringsruter.
Fra simuleringene blir det produsert kart over fordelingen av postsmolt fra ulike regioner i løpet av perioden mai-juli. Videre blir prosentandelen av postsmolt fra hver av region som vandrer gjennom de ulike HTH-områdene beregnet. Til slutt blir generelle utvandringsruter diskutert.
4.2 - Resultat av simuleringer
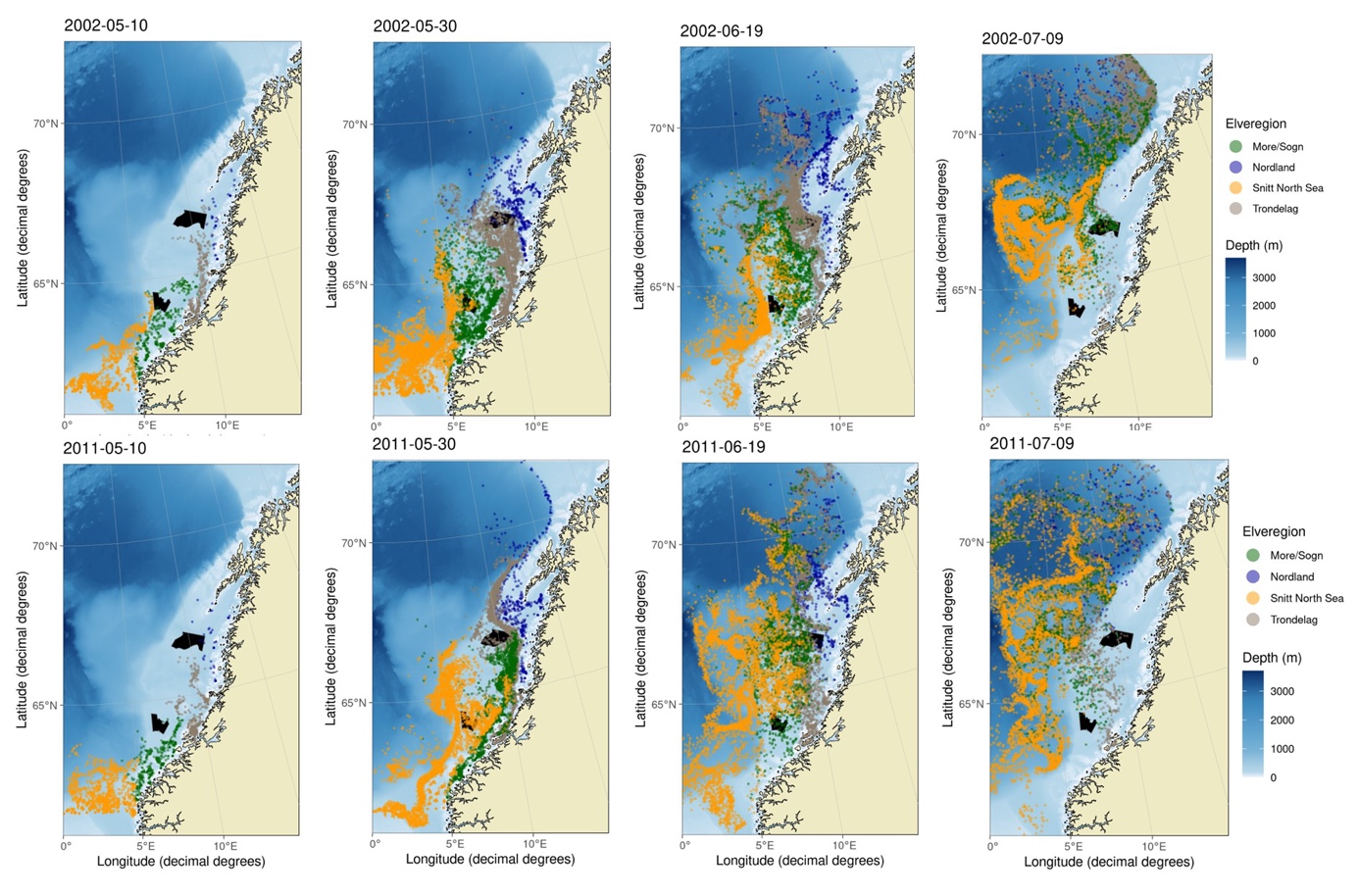
P05 - Simuleringene viser at P5 ligger plassert innenfor vandringsrutene til postsmolt fra Trøndelag, og delvis i innenfor vandringrutene til postsmolt fra Nordland (Figur 7). For postsmolt fra Trøndelag og Nordland vandret henholdsvis i snitt 30.1% (min-max= 11,8 - 52.0%) og 7,8 % (min-max= 2.2 - 18.7 %) innenfor P5 (Tabell S1). Postsmolt fra Nordsjøen og Møre og Romsdal vandret i liten grad gjennom det nordligste HTH området (P5), da deres vandringruter i all hovedsak går lengre vest i Norskehavet. Regional variasjon i havstrømmer har større innvirkning på post-smoltens vandringsruter enn postsmoltens svømmehastighet. De fleste årene vil postsmolt fra Trøndelag ha vandringsruter som går spredt gjennom P5 og lengre vest i havet, men noen år kan vandringsrutene i hovedsak gå øst for P5. For fisk fra Nordland går vandringsrutene i hovedsak øst for P5, selv om en mindre andel av post-smolten kan trekke lengre vest og dermed vandre gjennom P5. En høyere svømmehastighet førte i hovedsak til en noe lavere andel av postsmolt fra Trøndelag og en høyere andel fra Nordland innenfor P5.
P11 - Vandringsrutene i havet for postsmolt fra Nordland og Trøndelag gikk i all hovedsak nord for HTH-område P11, selv om 0,1-2,6 % av postsmolten fra Trøndelag svømte gjennom dette området i enkelte år (Figur 7). For post-smolten fra Møre og Romsdal og Nordsjøen svømte i snitt henholdsvis 22,5% (min-max= 4,8-33,6%) og 6,8% (min-max= 0,1-30,4%) gjennom P11 (Tabell S1, Figur 7). Regional variasjon i både retning og hastighet på havstrømmene gir høy mellomårlig variasjon i simulert andel av postsmolten som vandrer gjennom P11, spesielt for postsmolten som kommer fra områder lengre sør i Nordsjøen. Med økende svømmehastighet var det en økende andel fisk fra Møre og Romsdal og en synkende andel fisk fra Nordsjøen som svømte gjennom P11.
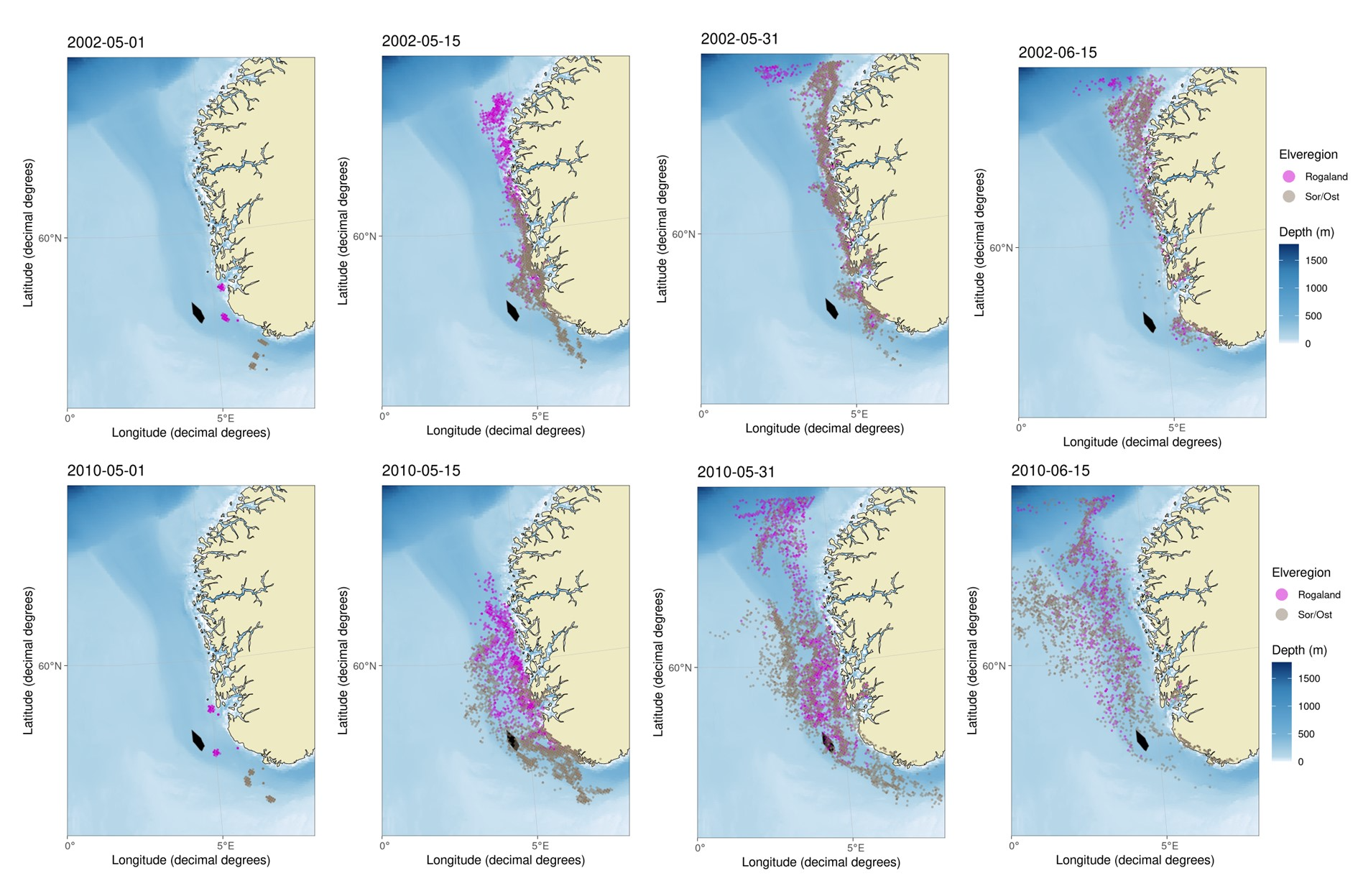
P02 - Vandringsrutene for postsmolten i Nordsjøen skjer i all hovedsak innenfor Norskerenna (Figur 8). For postsmolten fra Rogaland svømte i gjennomsnitt 8,6% (min-max= 1,6-27,7%) innenfor P2 (Tabell S1). For post-smolten initiert ved sydspissen av Norge svømte i gjennomsnitt 17.3% (min-max= 8,5-25,4%) innenfor P2. I hovedsak vandrer det meste av den simulerte postsmolten nærmere kysten enn P2, men i noen år kan havstrømmene trekke postsmolten lengre ut fra kysten slik at andelen som svømmer gjennom P2 øker (Figur 8). En høyere svømmehastighet førte i hovedsak til en lavere andel av den simulerte postsmolten innenfor P2 (Tabell S1).
Svømmehastigheten har stor innvirkning på når post-smolten vandrer gjennom HTH-områdene. Ved en svømmehastighet på 2 kroppslengder i sekundet vil det aller meste av postsmolten passere HTH-områdene P5 og P11 innen utgangen av juni. Ved 1,0 og 1,5 kroppslengder vil derimot mesteparten av postsmolten ha passert HTH-områdene innen henholdsvis midten og slutten av juli. For det sørligste området, P2, vil det meste av postsmolten ha passert tidlig i juni.
5 - Diskusjon
5.1 - Sammenhengen mellom miljøforhold i havet og vandringsmønsteret til postsmolt i kystnære farvann
Det har blitt utført flere studier av utvandrende postsmolt i fjord- og kystnære farvann der formålet har vært å estimere vandringshastighet, utvandringsruter og hvordan oseanografiske forhold påvirker svømmeadferd (f.eks. Lacroix mfl. 2004, Davidsen mfl. 2009, Newton mfl. 2021, Jensen mfl. 2022). Disse studiene har i hovedsak registrert vandringsadferden til postsmolt merket med akustiske merker, der merkene har blitt detektert av lyttebøyer forankret til en fast lokasjon eller til en liten båt. Studiene har blitt utført nært land slik at undersøkelsene i liten grad begrenses av værforhold, samt at naturlige geografiske avgrensninger (som fjordmunningene) øker sannsynligheten for å detektere utvandrende post-smolten. For perioden etter fjord- og kystvandringen er det svært få studier av vandringsruter og for norsk postsmolt har det ikke blitt utført studier av vandring i havområdet mellom kysten og ut til åpne havområder som Norskehavet (men se ovenfor). Kunnskap om vandringshastighet og drivkrefter i perioden postsmolten kan vandre forbi HTH-områdene er dermed dårlig kjent.
Post-smolten vandrer raskt gjennom fjorder og har en sterk motivasjon til å oppsøke åpne havområder (Thorstad mfl. 2012). Noen studier argumenterer for en sammenheng mellom post-smoltens vandringsretning eller vandringshastighet, og gradienter av temperatur eller saltholdighet i havet (Hedger mfl. 2008, Booker mfl. 2008, Burke mfl. 2014), men post-smoltens vandring kan tenkes å være påvirket av stor-skala miljøforhold som er korrelert med temperatur og saltholdighet (Jensen mfl. 2022).
Det er sannsynlig at post-smolt i åpne farvann svømmer med havstrømmene og dermed får flyttet seg raskt over store avstander (e.g. Booker mfl. 2008, Mork mfl. 2012). Enkelte studier av post-smoltens adferd i fjorder har derimot ikke funnet en sammenheng mellom strømretning og postsmoltens vandringsmønster (Økland mfl. 2004, Thorstad mfl. 2004, Newton mfl. 2021), selv om de fleste post-smoltene forlater fjordene når tidevannet går ut av fjorden (Lacroix mfl. 2004, Davidsen mfl. 2009). Modellsimuleringen har vist at skotsk postsmolt i østlige Nordsjøen ikke vil ende opp i Norskehavet i løpet av sommeren hvis de følger havstrømmene, og andre mekanismer enn havstrømmer påvirker trolig den skotske postsmoltens vandringsretning mens den er i Nordsjøen (Ounsley mfl. 2020).
I modellsimuleringene er det lagt til grunn at post-smolten svømmer bort fra kontinentalsokkelen og mot havområder med større bunndyp. I simuleringer der denne adferden er inkludert, sammen med å vandre i samme retning som havstrømmen, er modellen i stand til å gjenskape observert utbredelse av norsk fisk i Norskehavet (Utne and Mousing, in review). Dette betyr ikke dermed at postsmolten er i stand til å registrere bunndyp direkte, men heller at abiotiske eller biologiske mikrostrukturer i sjøen gjør det mulig for postsmolten å orientere seg mot dypere farvann. En slik adferd støttes av det lave antallet postsmolt som har blitt fanget over den norske kontinentalsokkelen sammenlignet med utenfor kontinentalsokkelen (Holm mfl. 2000, Gilbey mfl. 2021).
Flere studier har argumentert for at laksens vandring i havet er påvirket av geomagnetiske felt, der laksen har en viss kjennskap til sin geografiske posisjon og tid på året (e.g. Bracis og Anderson 2012, Byron og Burke 2014, Burke mfl. 2014). Simuleringen av vandringsmønsteret til norsk postsmolt gir ikke holdepunkter for at navigasjon i henhold til geomagnetisme er viktig for norsk post-smolt (Utne and Mousing, in review), selv om det ikke kan utelukkes at en slik adferd også kan påvirke på vandringsadferden.
Ulike studier har rapportert vandringshastigheter for postsmolt som har variert mellom 0.4-3.0 kroppslengder i sekundet (Thorstad mfl. 2012). For studier der man følger post-smolten med båt er det risiko for å skremme fisken og dermed overestimere svømmehastigheten. Studier med faste lyttebøyer og mange individer gir troverdige estimater på svømmehastigheter, og slike studier har estimert en retningsbestemt svømmehastighet på under 1.5 kroppslengder i sekundet (Lilly mfl. 2023, Chaput mfl.2019). Tidligere modeller av vandring til norsk laks har benyttet en hastighet på 1 kl/sek (Johnsen mfl. 2021) og 1.5 kl/sek (Mork mfl. 2012).
5.2 - Simulerte vandringsruter i forhold til planlagte HTH-områder
Det er høy usikkerhet rundt både hvordan post-smoltens migrasjonsadferd og vandringshastighet påvirkes av miljøforholdene, jfr diskusjonen i avsnittet over. Dersom andre oseanografiske eller biologiske faktorer påvirker fiskens vandring enn det som er lagt til grunn i simuleringene kan de reelle vandringsrutene avvike fra modellresultatene. Dette gjelder særlig vandringsrutene i Nordsjøen i tilknytning til HTH-område P2, som er et område der man ikke har hatt fangster av postsmolt utenfor grunnlinjen. Dermed kan man ikke uten videre validere de simulerte vandringsrutene i dette området.
En viktig påvirkningsfaktor på post-smoltens geografiske utbredelse er havstrømmene. Det er sterke strømmer over kontinentalsokkelen og overflatestrømmen utenfor Norskekysten ved 62-63˚ N er f.eks. 0.3 m s -1 i gjennomsnitt (Orvik mfl. 2001), noe som tilsvarer omtrent 2 kroppslengder s -1 for postsmolten. Under slike forhold vil havstrømmene ha stor påvirkning på postsmoltens geografiske utbredelse uavhengig av postsmoltens svømmeadferd. Da havstrømmene raskt kan føre postsmolten vestover mot Norskehavet eller østover mot Norskekysten, og gitt den store mellomårlige variasjonen for disse strømmene, er det svært sannsynlig at postsmoltens vandringsruter i det minste i perioder også inkluderer HTH-områdene.
Modellsimuleringene viser at det i størst grad er postsmolt fra Trøndelag som overlapper med HTH-området P5, men også postsmolt fra Møre og Romsdal vil kunne vandre gjennom P5. I enkelte år vandrer over 50% av post-smolten fra Trøndelag gjennom P5. Trøndelag er fylket med størst produksjon av smolt i Norge og i henhold til Nilsen mfl. (2017) kommer ~27% av all norsk postsmolt fra dette fylket. Postsmolt fra Nordland og fra Nordsjøen vil i svært liten grad ha vandringsruter som overlapper geografisk med P5. Vandringsrutene for postsmolt fra Nordsjøen går nesten utelukkende vest for P5.
Simuleringene viser at P11 ligger sentralt plassert i forhold til vandringsrutene til fisk fra Møre og Romsdal, de fleste årene vil vandringsrutene til postsmolt fra denne regionen overlappe med P11. Modellresultatene viser at i enkelte år vil over 30% av postsmolten fra elver i Møre og Romsdal og 27% av postsmolten fra Nordsjøen vandre gjennom HTH-område P11, mens det er lite geografisk overlapp mellom P11 og utbredelsen til postsmolt fra elver i Trøndelag og Nordland. I henhold til Nilsen mfl. (2017) kommer ~10% av all norsk postsmolt fra Møre og Romsdal. Enkelte år kan vandringsrutene til postsmolten fra Møre og Romsdal være tettere opp mot Norskekysten, og i slike år vil en liten andel av postsmolten vandre gjennom P11. Simuleringene viser derimot at i disse årene kan en høyere andel av postsmolten fra Nordsjøen svømme så langt øst at de passerer gjennom P11.
Det er i den forbindelse viktig å påpeke at andelen av postsmolt fra Nordsjøen som svømmer gjennom HTH-områdene vil avhenge av hvor kystnært denne postsmolten vandrer. Det har ikke vært systematisk gjenfangst av norsk eller utenlandsk postsmolt i Nordsjøen og de foretrukne vandringsrutene i dette området er i stor grad ukjent. I simuleringene ble postsmolt sluppet ut langs et tverrsnitt for å ta høyde for manglende kunnskap om hvor langt fra kysten postsmolten vandrer i Nordsjøen. Den estimerte andelen av postsmolten fra disse tverrsnittene som vandrer innenfor HTH-områdene påvirkes derimot av hvor postsmolten initieres. I simuleringene vandret i gjennomsnitt 17.3% av postsmolten fra sydspissen av Norge innenfor P2, og dette var i hovedsak individene som var initiert lengst fra land. Det er dermed særskilt høy usikkerhet i vandringsrutene for postsmolt som er initiert i tverrsnittene. Hvis vandringsrutene går nærmere kysten enn der post-smolten ble initiert vil en mindre andel av postsmolten vandre innenfor P2-området.
I henhold til modellresultatene vil tidsperioden som postsmolten vandrer gjennom HTH-områdene strekke seg fra tidlig mai til utgangen av juni ved en vandringshastighet på 2 kr.l. sek -1 , og fra midten av mai til utgangen av juli ved en vandringshastighet på 1 kroppslengde sek -1 . I tillegg til den benyttede vandringshastigheten, er disse resultatene sensitive for bl.a. tidspunktet postsmolten utvandrer fra elvene. Gitt det lave antallet postsmolt som fanges under tråltokt over norske kontinentalsokkelen i juli (Holm mfl. 2000, Gilbey mfl. 2021) er det sannsynlig at de fleste post-smoltene har passert HTH-områdene innen midten av juli.
5.3 - Hvordan kan kunnskapen økes og mer detaljerte modeller utvikles?
For utvikling av modeller er valideringsdata viktige. Som beskrevet ovenfor foreligger det noe slike data, fra telemetriforsøk og postsmoltfangster med genetisk bestemt opprinnelse, men antallet fisk med kjent opprinnelse som kan benyttes til å kalibrere modellen for postsmoltvandring i havet er foreløpig begrenset. Det ville derfor vært nyttig og viktig å forbedre slike datasett gjennom mer målrettet fangst av postsmolt i aktuelle områder for HTH, og telemetriforsøk innrettet mot kartlegging av miljøvariablers betydning for postsmoltens vandringsvalg i havet. Mer aktuelle og relevante data fra telemetri kan innhentes på flere måter. Merking av smolt med akustiske merker kombinert med lyttebøyer plassert i HTH-områdene ville kunne gitt mer presis informasjon om hvilke laksebestander som vandrer gjennom disse områdene. Eventuelt kunne dette også vært kombinert med autonome farkoster som patruljerer mulige vandringsruter for postsmolt og fanger opp signaler fra akustisk merket smolt. Slike forsøk med kombinasjon av ulike metoder for deteksjon har vært gjennomført i Canada hvor det nå gjennomføres en omfattende kartlegging av vandringsruter for både postsmolt og større laks i forbindelse med utvidelse av petroleumsvirksomhet nordøst for Newfoundland ( https://www.canada.ca/en/fisheries-oceans/news/2021/06/fisheries-and-oceans-canada-collaborate-on-new-research-to-study-atlantic-salmon-migration.html ).
Forsøk med DST-merking av postsmolt, slik som det som ble utført på Island, kan også gi viktig informasjon som kan benyttes for å forbedre modellene. Siden slik merking forutsetter gjenfangst av fisken for å få data er det imidlertid få vassdrag i Norge hvor slik merking er aktuelt. Etneelva i Vestland, hvor Havforskningsinstituttet har en oppgangsfelle som fanger bortimot all oppvandrende laks er det mest egnete vassdraget for et slikt forsøk.
Felles for metodene som baserer seg på merking av smolt er at de er kostnadskrevende og kun gir data for enkeltbestander. Det er også rimelig å anta at en relativt liten andel av den merkede fisken vil bli registrert på lyttebøyer plassert langt fra kysten på grunn av «fortynningseffekten» og dødelighet på postsmolten som følge av predasjon eller andre årsaker. Men denne metoden gir presis informasjon i områder som er dekket av lyttebøyer. Trålfangst og genetisk tilordning til region/vassdrag av individer vil kunne gi et bredere grunnlag for vurderinger siden mange ulike bestander og regioner kan være representert i fangsten. Det pågår nå et arbeid med å utvikle en mer presis genetisk baseline for Europeiske laksebestander, som vil kunne øke presisjonen i tilordning til region/vassdrag betydelig i forhold til det som har vært mulig hittil. Resultater av dette arbeidet forventes å foreligge høsten 2024.
6 - Konklusjoner
Modellsimuleringene gir kvantifiserte anslag for hvor stor andel av postsmolten som vandrer gjennom HTH-områdene. En mer realistisk tilnærming for å vurdere potensielle effekter på vill postsmolt ville vært å også vurdere hvor mye av postsmolten som vandrer gjennom smitteområder nedstrøms HTH-områdene. Geografisk utstrekning av disse smitteområdene vil kunne varierer med intensiteten på produksjonen av oppdrettslaks i HTH-områdene, infeksjonsstatus til oppdrettslaksen i anleggene, temperatur siden det har en effekt på utviklingstid hos lakseluslarver, og naturlige variasjoner i strømmønstre gjennom utvandringsperioden til postsmolt. Det foreligger foreløpig ikke data som gjør det mulig å foreta en slik kvantisering av postsmoltens bevegelse gjennom influensområdene. Det vil først foreligge etter at det er gjennomført flere simuleringer av smittespredning ved ulike produksjonsnivåer og miljøbetingelser. Vi går ut fra at slike simuleringer vil bli gjennomført i arbeidet med den overordnete konsekvensvurderingen og at nye estimater for smitte til vill utvandrende postsmolt dermed kan utarbeides ved anvendelse av smittemodellen som benyttes i NALO. Basert på eksistrerende modellsimuleringen er det sannsynlig at postsmolt fra store deler av Sør-Norge vil kunne bevege seg gjennom eller i influensområdet til de foreslåtte HTH-områdene i tidlig fase av utvandringen. Det er nødvendig å innhente flere data for viderutvikling og validering av vandringsmodellene. Vurdering av konsekvens av påvirkning fra HTH-områdene på utenlandsk og norsk postsmolt vil være svært usikker inntil det foreligger et bedre datagrunnlag.
I denne rapporten har vi oppsummert kunnskapsstatus med hensyn på postsmoltvandring ut i havet, og hvordan det eksisterende datagrunnlaget og vandringsmodellen utviklet ved Havforskningsinstituttet kan benyttes til å estimere i hvilken grad postsmolt fra Norge og andre land vil kunne bevege seg gjennom HTH-områdene. Analysene viser klart at det eksisterende kunnskapsgrunnlaget er svært begrenset og at modellens prediksjoner av vandring og fordeling av postsmolt er usikker. Vi har pekt på hvilke typer studier som vil kunne bidra til økt kunnskap og mer presise modeller og vil anbefale at det gjennomføres slike undersøkelser slik at forutsetningene for konsekvensvurderingen slik den er definert i forskriften kan oppfylles. Som eksempelet fra Canada viser, kreves det omfattende innsats og datainnsamling over tid for å oppnå et presist bilde av hvordan laksen vandrer i havet.
7 - Referanser
Booker, D.J., Wells, N.C., Smith, I.P., 2008. Modelling the trajectories of migrating Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 65, 352-361.
Bracis, C., Anderson, J.J., 2012. An investigation of the geomagnetic imprinting hypothesis for salmon. Fish. Oceanogr. 21, 170-181.
Burke, B.J., Anderson, J.J., Baptista, A.M., 2014. Evidence for multiple navigational sensory capabilities of Chinook salmon. Aquatic Biology 20, 77-90.
Byron, C.J., Burke, B.J., 2014. Salmon ocean migration models suggest a variety of population-specific strategies. Rev. Fish Biol. Fish. 24, 737-756.
Chaput, G., Carr, J., Daniels, J., Tinker, S., Jonsen, I., Whoriskey, F., 2019. Atlantic salmon (Salmo salar) smolt and early post-smolt migration and survival inferred from multi-year and multi-stock acoustic telemetry studies in the Gulf of St. Lawrence, northwest Atlantic. ICES J. Mar. Sci. 76, 1107-1121.
Davidsen, J.G., Rikardsen, A.H., Halttunen, E., Thorstad, E.B., Okland, F., Letcher, B.H., Skardhamar, J., Næsje, T.F., 2009. Migratory behaviour and survival rates of wild northern Atlantic salmon (Salmo salar) post-smolts: effects of environmental factors. J. Fish Biol. 75, 1700-1718.
Gilbey, J., J. Coughlan, V. Wennevik, P. Prodöhl, J. R. Stevens, C. Garcia de Leaniz, D. Ensing, E. Cauwelier, C. Cherbonnel, S. Consuegra, M. W. Coulson, T. F. Cross, W. Crozier, E. Dillane, J. S. Ellis, E. García-Vázquez, A. M. Griffiths, S. Gudjonsson, K. Hindar, S. Karlsson, D. Knox, G. Machado-Schiaffino, D. Meldrup, E. Eg Nielsen, K. Ólafsson, C. R. Primmer, S. Prusov, L. Stradmeyer, J.-P. Vähä, A. J. Veselov, L. M. I. Webster, P. McGinnity, and E. Verspoor. 2017. A microsatellite baseline for genetic stock identification of European Atlantic salmon (Salmo salar L.). ICES Journal of Marine Science 75(2): 662-674.
Gilbey, J., Utne, K.R., Wennevik, V., Beck, A.C., Kausrud, K., Hindar, K., de Leaniz, C.G., Cherbonnel, C., Coughlan, J., Cross, T.F., Dillane, E., Ensing, D., Garcia-Vazquez, E., Hole, L.R., Holm, M., Holst, J.C., Jacobsen, J.A., Jensen, A.J., Karlsson, S., Maoileidigh, N.O., Mork, K.A., Nielsen, E.E., Nottestad, L., Primmer, C.R., Prodohl, P., Prusov, S., Stevens, J.R., Thomas, K., Whelan, K., McGinnity, P., Verspoor, E., 2021. The early marine distribution of Atlantic salmon in the North-east Atlantic: A genetically informed stock-specific synthesis. Fish Fish. 22, 1274-1306.
Guðjónsson, S., S. M. Einarsson, I. R. Jónsson, and J. Guðbrandsson. 2015. Marine feeding areas and vertical movements of Atlantic salmon (Salmo salar) as inferred from recoveries of data storage tags. Canadian Journal of Fisheries and Aquatic Sciences 72:1087-1098.
Hedger, R.D., Martin, F., Hatin, D., Caron, F., Whoriskey, F.G., Dodson, J.J., 2008. Active migration of wild Atlantic salmon (Salmo salar) smolt through a coastal embayment. Mar. Ecol. Prog. Ser. 355, 235-246.
Holm, M., Holst, J.C., Hansen, L.P., 2000. Spatial and temporal distribution of post-smolts of Atlantic salmon (Salmo salar L.) in the Norwegian Sea and adjacent areas. ICES J. Mar. Sci. 57, 955-964.
Holst, J. C., F. Nilsen, K. Hodneland, and A. Nylund. 1993. Observations of the biology and parasites of postsmolt Atlantic salmon, Salmo salar, from the Norwegian sea. Journal of Fish Biology 42:962-966.
Jensen, J.L.A., Strom, J.F., Nikolopoulos, A., Primicerio, R., Skarohamar, J., Atencio, B.J., Strand, J.E.T., Bjorn, P.A., Bohn, T., 2022. Micro- and macro-habitat selection of Atlantic salmon, Salmo salar, post-smolts in relation to marine environmental cues. ICES J. Mar. Sci. 79, 1394-1407.
Johnsen, I.A., Harvey, A., Sævik, P.N., Sandvik, A.D., Ugedal, O., Adlandsvik, B., Wennevik, V., Glover, K.A., Karlsen, O., 2021. Salmon lice-induced mortality of Atlantic salmon during post-smolt migration in Norway. ICES J. Mar. Sci. 78, 142-154.
Lacroix, G.L., McCurdy, P., Knox, D., 2004. Migration of Atlantic salmon postsmolts in relation to habitat use in a coastal system. Trans. Am. Fish. Soc. 133, 1455-1471.
Lien, V.S., Gusdal, Y., Albretsen, J., Melsom, A., Vikebø, F.B., 2013. Evaluation of a Nordic Seas 4 km numerical ocean model hindcast archive (SVIM), 1960-2011. Fisken Havet 7, 80pp.
Lilly, J., Honkanen, H.H., Rodger, J.R., del Villar, D., Boylan, P., Green, A., Pereiro, D., Wilkie, L., Kennedy, R., Barkley, A., Rosell, R., Maoiléidigh, N.O., O'Neill, R., Waters, C., Cotter, D., Bailey, D., Roche, W., McGill, R., Barry, J., Beck, S.V., Henderson, J., Parke, D., Whoriskey, F.G., Shields, B., Ramsden, P., Walton, S., Fletcher, M., Whelan, K., Bean, C.W., Elliott, S., Bowman, A., Adams, C.E., 2023. Migration patterns and navigation cues of Atlantic salmon post-smolts migrating from 12 rivers through the coastal zones around the Irish Sea. J. Fish Biol.
Mork, K.A., Gilbey, J., Hansen, L.P., Jensen, A.J., Jacobsen, J.A., Holm, M., Holst, J.C., Maoileidigh, N.O., Vikebø, F., McGinnity, P., Melle, W., Thomas, K., Verspoor, E., Wennevik, V., 2012. Modelling the migration of post-smolt Atlantic salmon (Salmo salar) in the Northeast Atlantic. ICES J. Mar. Sci. 69, 1616-1624.
Newton, M., Barry, J., Lothian, A., Main, R., Honkanen, H., McKelvey, S., Thompson, P., Davies, I., Brockie, N., Stephen, A., Murray, R.O., Gardiner, R., Campbell, L., Stainer, P., Adams, C., 2021. Counterintuitive active directional swimming behaviour by Atlantic salmon during seaward migration in the coastal zone. ICES J. Mar. Sci. 78, 1730-1743.
Nilsen, F., Ellingsen, I., Finstad, B., Jansen, P.A., Karlsen, Ø., Kristoffersen, A.B., Sandvik, A.D., Sægrov, H., Ugedal, O., Vollset, K.S., Myksvoll., M., 2017. Vurdering av lakselusindusert villfiskdødelighet per produksjonsområde i 2016 og 2017. Rapport fra ekspertgruppe for vurdering av lusepåvirkning (in Norwegian).
Ó Maoiléidigh, N., White, J., Hansen, L. P., Jacobsen, J. A., Potter, T., Russell, I., Reddin, D., et al. 2018. Fifty years of marine tag recoveries from Atlantic salmon. ICES Coopera- tive Research Report No. 343. 121 pp. http://doi.org/10.17895/ices.pub.4542
Ounsley, J.P., Gallego, A., Morris, D.J., Armstrong, J.D., 2020. Regional variation in directed swimming by Atlantic salmon smolts leaving Scottish waters for their oceanic feeding grounds-a modelling study. ICES J. Mar. Sci. 77, 315-325.
Orvik, K.A., Skagseth, O., Mork, M., 2001. Atlantic inflow to the Nordic Seas: current structure and volume fluxes from moored current meters, VM-ADCP and SeaSoar-CTD observations, 1995-1999. Deep-Sea Research Part I-Oceanographic Research Papers 48, 937-957.
Putman, N. F. 2015. Inherited Magnetic Maps in Salmon and the Role of Geomagnetic Change. Integrative and Comparative Biology 55:396-405.
Rikardsen, A. H., D. Righton, J. F. Strøm, E. B. Thorstad, P. Gargan, T. Sheehan, F. Økland, C. M. Chittenden, R. D. Hedger, T. F. Næsje, M. Renkawitz, J. Sturlaugsson, P. Caballero, H. Baktoft, J. G. Davidsen, E. Halttunen, S. Wright, B. Finstad, and K. Aarestrup. 2021. Redefining the oceanic distribution of Atlantic salmon. Scientific Reports11:12266.
Rodger, J. R., J. Lilly, H. M. Honkanen, D. del Villar, R. Kennedy, N. Ó. Maoiléidigh, P. Boylan, R. Rosell, D. J. Morris, R. O'Neill, C. Waters, D. Cotter, L. Wilkie, A. Barkley, A. Green, S. V. Beck, J. Ribbens, J. Henderson, D. Parke, A. Kettle-White, L. Ballantyne, S. Marshall, P. Hopper, N. Gauld, J. D. Godfrey, L. E. Chapman, J. Thorburn, A. Drumm, F. Whoriskey, B. Shields, P. Ramsden, J. Barry, M. Milane, W. Roche, J. D. Armstrong, A. Wells, S. Walton, M. Fletcher, D. M. Bailey, B. Whyte, R. McGill, M. Bilsby, K. Whelan, C. W. Bean, and C. E. Adams 2024. Inshore and offshore marine migration pathways of Atlantic salmon post-smolts from multiple rivers in Scotland, England, Northern Ireland, and Ireland. Journal of Fish Biology n/a.
https://doi.org/10.1111/jfb.15760
Shchepetkin, A.F., McWilliams, J.C., 2005. The regional oceanic modeling system (ROMS): a split-explicit, free-surface, topography-following-coordinate oceanic model. Ocean Model. Online 9, 347-404.
Shelton, R. G. J., W. R. Turrell, A. Macdonald, I. S. McLaren, and N. T. Nicoll. 1997. Records of post-smolt Atlantic salmon, Salmo salar L., in the Faroe-Shetland Channel in June 1996. Fisheries Research 31:159-162.
Strøm, J. F., E. B. Thorstad, R. D. Hedger, and A. H. Rikardsen. 2018. Revealing the full ocean migration of individual Atlantic salmon. Animal Biotelemetry 6:2.
Thorstad, E.B., Okland, F., Finstad, B., Sivertsgard, R., Bjorn, P.A., McKinley, R.S., 2004. Migration speeds and orientation of Atlantic salmon and sea trout post-smolts in a Norwegian fjord system. Environ. Biol. Fishes 71, 305-311.
Thorstad, E.B., Whoriskey, F., Uglem, I., Moore, A., Rikardsen, A.H., Finstad, B., 2012. A critical life stage of the Atlantic salmon Salmo salar: behaviour and survival during the smolt and initial post-smolt migration. J. Fish Biol. 81, 500-542.
Utne, K.R., Mousing, E.A., in review. Regional differences in salmon post-smolt migration routes can explain variation in individual growth at sea.
Vollset, K.W., Lennox, R.J., Lamberg, A., Skaala, O., Sandvik, A.D., Saegrov, H., Kvingedal, E., Kristensen, T., Jensen, A.J., Haraldstad, T., Barlaup, B.T., Ugedal, O., 2021. Predicting the nationwide outmigration timing of Atlantic salmon (Salmo salar) smolts along 12 degrees of latitude in Norway. Divers. Distrib. 27, 1383-1392.
Økland, F., Thorstad, E.B., Finstad, B., Sivertsgård, R., Plantalech, N., Jepsen, N., McKinley, R.S., 2006. Swimming speeds and orientation of wild Atlantic salmon post-smolts during the first stage of the marine migration. Fish. Manage. Ecol. 13, 271-274.
Ådlandsvik, B. 2019. Havbruk til havs – smittespredning. Rapport fra Havforskningen 2019-58.
8 - Tabeller
Tabell S1a. Simulert andel post-smolt fra som vandrer gjennom HTH-områdene P5 og P11 for hvert av årene 2001-2011 ved en svømmehastighet på 1.0 kroppslengde s-1.
| Område overlapp | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | Gj.snitt |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nordsjøen - P11 | 7,3 % | 27,2 % | 10,1 % | 6,6 % | 2,7 % | 9,7 % | 7,0 % | 0,9 % | 6,3 % | 7,4 % | 17,4 % | 9,3 % |
| Møre/Sogn - P11 | 7,4 % | 19,5 % | 14,4 % | 22,8 % | 22,3 % | 24,6 % | 11,0 % | 25,2 % | 22,0 % | 28,1 % | 5,1 % | 18,4 % |
| Trøndelag - P11 | 0,3 % | 0,0 % | 0,0 % | 0,2 % | 0,1 % | 0,3 % | 0,7 % | 0,0 % | 1,3 % | 0,3 % | 0,1 % | 0,3 % |
| Nordland - P11 | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % |
| Nordsjøen - P5 | 1,3 % | 3,0 % | 0,5 % | 0,1 % | 0,2 % | 2,4 % | 0,3 % | 0,0 % | 0,1 % | 0,3 % | 3,1 % | 1,0 % |
| Møre/Sogn - P5 | 23,3 % | 24,1 % | 22,9 % | 7,7 % | 14,2 % | 22,8 % | 19,4 % | 0,6 % | 3,0 % | 5,0 % | 19,4 % | 14,8 % |
| Trøndelag - P5 | 24,5 % | 58,1 % | 51,0 % | 31,6 % | 31,3 % | 26,2 % | 23,5 % | 35,5 % | 23,2 % | 34,4 % | 12,7 % | 32,0 % |
| Nordland - P5 | 0,9 % | 3,1 % | 0,6 % | 10,0 % | 2,4 % | 3,3 % | 16,5 % | 2,2 % | 9,2 % | 2,0 % | 5,0 % | 5,0 % |
| Rogaland - P2 | 4,3 % | 6,7 % | 2,5 % | 16,6 % | 11,6 % | 6,9 % | 9,7 % | 20,9 % | 18,6 % | 34,7 % | 7,1 % | 12,7 % |
| Snitt Sør - P2 | 30,3 % | 5,4 % | 17,9 % | 26,0 % | 28,9 % | 14,0 % | 10,9 % | 31,3 % | 36,8 % | 18,2 % | 19,7 % | 21,8 % |
Tabell S1b. Simulert andel post-smolt fra som vandrer gjennom HTH-områdene P5 og P11 for hvert av årene 2001-2011 ved en svømmehastighet på 1,5 kroppslengde s-1.
| Område overlapp | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | Gj.snitt |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nordsjøen - P11 | 6,1 % | 30,4 % | 6,9 % | 3,7 % | 1,4 % | 3,5 % | 5,3 % | 0,1 % | 5,4 % | 3,7 % | 8,7 % | 6,8 % |
| Møre/Sogn - P11 | 13,6 % | 27,9 % | 22,4 % | 21,9 % | 24,6 % | 33,6 % | 15,9 % | 24,7 % | 25,3 % | 33,1 % | 4,8 % | 22,5 % |
| Trøndelag - P11 | 1,3 % | 0,0 % | 0,1 % | 0,2 % | 0,3 % | 0,4 % | 1,2 % | 0,1 % | 2,6 % | 1,1 % | 0,2 % | 0,7 % |
| Nordland - P11 | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % |
| Nordsjøen - P5 | 0,0 % | 0,2 % | 0,0 % | 0,1 % | 0,0 % | 0,5 % | 0,1 % | 0,0 % | 0,1 % | 0,0 % | 1,2 % | 0,2 % |
| Møre/Sogn - P5 | 8,4 % | 7,8 % | 8,8 % | 2,5 % | 5,5 % | 13,2 % | 8,2 % | 0,0 % | 2,8 % | 0,7 % | 22,6 % | 7,3 % |
| Trøndelag - P5 | 35,9 % | 51,7 % | 52,0 % | 29,5 % | 27,1 % | 26,5 % | 18,1 % | 22,5 % | 11,8 % | 27,6 % | 28,3 % | 30,1 % |
| Nordland - P5 | 4,1 % | 4,9 % | 2,2 % | 11,4 % | 5,0 % | 8,0 % | 18,7 % | 8,6 % | 15,2 % | 2,6 % | 5,2 % | 7,8 % |
| Rogaland - P2 | 3,0 % | 2,8 % | 1,6 % | 15,5 % | 6,6 % | 2,7 % | 6,4 % | 11,7 % | 12,9 % | 27,7 % | 3,8 % | 8,6 % |
| Snitt Sør - P2 | 20,7 % | 8,5 % | 11,7 % | 22,5 % | 24,3 % | 10,5 % | 11,6 % | 21,6 % | 25,9 % | 22,0 % | 11,4 % | 17,3 % |
Tabell S1c. Simulert andel post-smolt fra som vandrer gjennom HTH-områdene P5 og P11 for hvert av årene 2001-2011 ved en svømmehastighet på 2,0 kroppslengde s-1.
| Område overlapp | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | Gj.snitt |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nordsjøen - P11 | 5,0 % | 24,3 % | 5,0 % | 1,8 % | 0,8 % | 2,3 % | 3,6 % | 0,0 % | 4,5 % | 0,9 % | 9,3 % | 5,2 % |
| Møre/Sogn - P11 | 19,1 % | 33,5 % | 25,1 % | 19,4 % | 24,0 % | 36,0 % | 16,7 % | 24,9 % | 23,8 % | 31,5 % | 7,1 % | 23,7 % |
| Trøndelag - P11 | 1,4 % | 0,0 % | 0,0 % | 0,4 % | 0,4 % | 0,5 % | 1,4 % | 0,2 % | 2,8 % | 1,2 % | 0,4 % | 0,8 % |
| Nordland - P11 | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % |
| Nordsjøen - P5 | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,0 % | 0,2 % | 0,1 % | 0,0 % | 0,0 % | 0,0 % | 0,5 % | 0,1 % |
| Møre/Sogn - P5 | 5,7 % | 2,5 % | 2,4 % | 1,7 % | 2,0 % | 5,1 % | 5,7 % | 0,1 % | 1,7 % | 0,1 % | 13,7 % | 3,7 % |
| Trøndelag - P5 | 39,8 % | 38,5 % | 39,7 % | 24,3 % | 20,9 % | 23,8 % | 18,5 % | 18,2 % | 9,4 % | 22,9 % | 39,1 % | 26,8 % |
| Nordland - P5 | 5,2 % | 7,1 % | 4,7 % | 12,6 % | 10,0 % | 11,9 % | 22,9 % | 15,2 % | 17,7 % | 4,9 % | 4,7 % | 10,6 % |
| Rogaland - P2 | 2,04 % | 1,33 % | 0,71 % | 13,42 % | 2,76 % | 1,33 % | 4,08 % | 5,92 % | 8,47 % | 24,85 % | 2,65 % | 6,1 % |
| Snitt Sør - P2 | 14,01 % | 10,99 % | 6,84 % | 20,15 % | 18,74 % | 6,50 % | 10,81 % | 15,93 % | 18,80 % | 23,72 % | 5,53 % | 13,8 % |