The Institute of Marine Research (IMR) and the Norwegian Veterinary Institute (NVI) have evaluated if a spatial re-structuring of salmon aquaculture farm locations in Norway can decrease the infection of salmon lice, and virus (pancreas disease and Infectious salmon anaemia) among farming locations in production zone 3 (PZ3). Using biophysical models and network analyses to evaluate transport of lice and virus between the sites, the potential infection between sites can be decreased if the number of sites were to be reduced. For example, the analysis shows that by consolidating the number of farm-locations from 135 to 100, the infection of lice between localities can be reduced by 46%, and 30% for virus. Statistical models from NVI using empirical data from the salmon farms in PZ3, corroborate these findings that fewer farming sites can reduce the infection risk between sites. However, further studies on impact of a re-structuring on wild salmonid populations and dispersal of organic waste have to be implemented in order to recommend specific new siting structures.
Endret lokalitetsstruktur i produksjonsområde 3
— vurdert virkning på spredning av lakselus, pankreassykdom og infektiøs lakseanemi
Rapportserie:
Rapport fra havforskningen 2020-12
ISSN: 1893-4536
Publisert: 14.05.2020
Prosjektnr: 14272-09
Oppdragsgiver(e): Nærings- og Fiskeridepartementet
Referanse: 19/2159-9
Forskningsgruppe(r):
Oseanografi og klima
,
Smittespredning og sykdom
Tema:
Lakselus
Program:
Miljøeffekter av akvakultur
Godkjent av:
Forskningsdirektør(er):
Geir Lasse Taranger
Programleder(e):
Terje Svåsand
English summary
Sammendrag
Havforskningsinstituttet (HI) og Veterinærinstituttet (VI) fikk en felles bestilling av Nærings- og fiskeridepartementet om å analysere effektene av ulike scenarioer for en ny lokalitetsstruktur i produksjonsområde 3 som kan gi mindre spredning av lakselus, ILA og PD mellom lokalitetene. Det er antatt at mindre smitte mellom lokalitetene vil gjøre det lettere å drive oppdrett av laks, med lave luseverdier, bedre fiskehelse og fiskevelferd, og dermed øke bærekraften. Ved hjelp av biofysisk modellering og nettverksanalyse av spredning av lakselus, PD og ILA mellom lokaliteter er det anslått at flytting av produksjon fra særlig smittespredende lokaliteter til mindre smittespredende lokaliteter, kan redusere den totale smitten mellom lokalitetene. Analysene viser at ved å fjerne tilfeldige lokaliteter hvor man flytter biomassen til de resterende lokalitetene vil redusere smittepresset, men ikke like effektivt som ved en strategisk flytting av biomasse fra de “verste” til de beste lokalitetene med tanke på smittespredning.
I et av scenariene som er testet med Havforskningsinstituttet sine modeller er det indikert at smitten mellom lokalitetene kan reduseres med 46% for lakselus og 30% for virus, ved å redusere fra dagens 135 lokaliteter ned til 100 matfisklokaliteter – dette uten å redusere den totale produksjonen i produksjonsområdet.
På tilsvarende måte indikerer Veterinærinstituttet sin lusemodell at en ved tilfeldig fjerning av halvparten av alle lokaliteter og refordeling av biomassen til andre lokaliteter i PO3 vil få rundt regnet ca. 20% færre voksne hunnlus, 20% færre andre mobile lus, og 20% færre behandlinger i hele området. Denne effekten blir større, dersom de lokalitetene som lukkes er strategisk valgt, dvs. at de er valgt på bakgrunn av hvor mye de bidrar til spredning av lus basert på HI sin nettverksmodell. Det synes dermed å være et stort potensial for å redusere smitten mellom lokaliteter ved å redusere antall lokaliteter i PO3 og samtidig opprettholde produksjonen.
Imidlertid er det behov for mer omfattende analyser og utredning av ny lokalitetsstruktur, der en tar hensyn til en rekke andre faktorer som bl.a. om lokalitetene tåler økt biomasse med tanke på organisk belastning, selskapsstruktur, muligheter for sonevise utsett, samt påvirkning på vill laksefisk før en kan anbefale konkret ny lokalitetsstruktur i PO3.
1 - Oppsummering
Havforskningsinstituttet (HI) og Veterinærinstituttet (VI) har fått i oppdrag av Nærings- og fiskeridepartementet å analysere ulike scenarioer for en mulig ny lokalitetsstruktur i produksjonsområde 3 (PO3) som kan gi mindre spredning av lakselus, ILA og PD mellom lokalitetene. Det er antatt at mindre smitte mellom lokalitetene vil gjøre det lettere å drive oppdrett av laks med lite lus, bedre fiskehelse og fiskevelferd, og dermed øke bærekraften. Analysene skal være basert på at reduksjon av total smitte innad i PO3 ved omstrukturering ikke skal gi vesentlig økt eksport av smitte til PO4, samt at maksimal tillat biomasse (MTB) i PO3 ikke skal reduseres fra dagens nivå. Departementet ønsket også at næringsaktørene selv skulle bidra med erfaringsbasert kunnskap inn mot forslag til scenarioer for analyse av lokalitetsomstrukturering. Det var også lagt til grunn at HI og VI leverer omforente tolkninger av resultatene.
Biofysisk modellering og nettverksanalyse utført av HI viser at flytting av biomasse fra særlig smittespredende lokaliteter (dvs. lokaliteter som bidrar mye til smittespredning i nettverket) til mindre smittespredende lokaliteter, kan gi betydelig reduksjon i den totale smittespredningen mellom lokalitetene.
For eksempel vil et scenario der man reduserer antallet matfisklokaliteter fra 135 til 100 ved å flytte 24% av MTB i PO3 (dvs. ca. 176.000 tonn) til de beste lokalitetene kunne gi en potensiell smittereduksjon av lakselus mellom lokaliteter på 46%, og en reduksjon av virus-smitte med opptil 30%. I dette scenarioet økte eksporten av lus fra PO3 til PO4 med 11%.
Det er også vist at en tilfeldig reduksjon i antall lokaliteter i PO3 med omdistribuering av biomassen (dvs. MTB) til de resterende lokalitetene gir en reduksjon i smittepress i området, men i mindre grad enn hvis en flytter biomassen fra de lokalitetene som bidrar mest med smittespredning til de som bidrar minst. Effekter av å innføre nullutslippsteknologi som lukkede merder på et økende antall lokaliteter og teknologi som reduserer innkommende smitte som eks. snorkelmerder eller tubenot er også analysert.
Videre viser HI sine analyser at en omfordeling av produksjonskapasitet til færre og mindre smittespredende lokaliteter kan gi et potensial for vekst i produksjon, uten å øke den totale smitten mellom lokaliteter i forhold til dagens situasjon.
VI har ved hjelp av en stokastisk populasjonsmodell analysert hvordan mengden av voksne hunnlus, andre mobile lus og antall behandlinger påvirkes av å fjerne lokaliteter og flytte biomassen til andre lokaliteter. VI har kjørt scenarioer der en tilfeldig har fjernet 20, 30, 40 eller 50% av lokalitetene og redistribuert biomassen til andre lokaliteter innen PO3. Disse analysene viser at det oppnås en reduksjon i antall voksne hunnlus, andre mobile stadier av lus og antall behandlinger, og at denne effekten øker med andelen av lokaliteter som slås sammen. Ved å fjerne 50% av lokalitetene oppnås en reduksjon på ca. 20% i mengden lus og antall behandlinger reduseres med ca. 20%.
VI har etterfølgende kjørt simuleringer med scenarioer der de lokalitetene som er tatt ut (fjernet) er de som av nettverksanalysene til HI har blitt utpekt som å være viktigst i et smittespredningsperspektiv. Resultatene av disse analyser viser at effekten av å ta bort lokaliteter forsterkes dersom de lokalitetene som tas bort er de som bidrar mest til smittespredning. Derved understøtter VI sine analyser de resultatene som HI har funnet.
VI har også analysert effekten av å ta bort de samme lokalitetene på spredningen av PD. Resultater fra disse analysene tilsier at det også vil ha en smittebegrensende effekt på spredningen av PD dersom en tar bort disse lokalitetene og fordeler biomassen deres på andre lokaliteter i PO3 med tilsvarende produksjonshistorikk.
Samlet sett viser både modellene til Havforskningsinstituttet og Veterinærinstituttet at en reduksjon av antallet lokaliteter i PO3 der en omfordeler biomassen (dvs. MTB) til de resterende lokalitetene kan gi en betydelig reduksjon i lusesmitte. Analysene tyder også på at en samtidig får redusert smittepotensialet av virussykdom (PD og ILA) mellom lokaliteter. Videre viser både modellene til HI og VI at en oppnår større reduksjon i lusesmitte ved å fjerne de lokalitetene som bidrar mest til smittespredning av lakselus i PO3, og samtidig fordele biomassen fra de fjernede lokalitetene til de lokalitetene som kommer best ut i nettverksanalysen, sammenlignet med tilfeldig fjerning av lokaliteter.
Nettverksanalysen viser også hvordan lokaliteter utgjør større eller mindre smitteklynger i PO3, der smittespredningspotensialet er større innen klyngen enn mellom klyngene. Dette kan være et viktig vitenskapelig kunnskapsgrunnlag for mer planmessige sonevise utsett og brakkleggingsområder med tanke på både virus og lakselus. Nettverksanalysen og smittespredningsmodellene som er benyttet her er et godt egnede verktøy for å undersøke hvordan restrukturering av lokalitetene kan redusere smitte mellom klyngene, og evaluere effekten av ulike biosikkerhetstiltak som f.eks. vaksinasjon mot PD.
På grunn av ordlyden i oppdraget fra Nærings- og fiskeridepartementet er nettverksanalysen gjennomført her designet for å redusere det totale smittenettverket mellom alle lokalitetene i PO3, og ikke spesifikt for å redusere smitte mellom klyngene. Det er antatt at redusert smitte mellom klyngene vil øke effekten av koordinert innsats innenfor klyngene, altså redusere risiko for smitte inn i en klynge etter koordinert brakklegging med påfølgende utsett av fisk, og var blant analysene som ble etterspurt fra næringsaktørene.
Det bør bemerkes at analysene i denne rapporten er gjennomført uten å ta hensyn til belastning på villfisk, ulike andre biosikkerhetstiltak, mulig økt organisk belasting på lokalitetene med foreslått økt produksjon, utfordringer knyttet til eierskap av de ulike tillatelsene eller konkrete tilpasninger til sonevise utsett. De foreliggende analysene må derfor ikke forstås som konkrete forslag til relokalisering, men viser potensialet i strategisk restrukturering av lokaliteter i PO3 både når det gjelder lus og virussmitte.
Smittespredningsmodellen fra VI har i andre sammenhenger vært brukt til å simulere hva som kan skje dersom en flytter lokaliteter til andre steder eller setter inn nye lokaliteter. Dersom det utarbeides en plan med konkrete forslag til plasseringer og størrelse på lokaliteter I PO3, vil modellen kunne simulere hvordan dette vil kunne påvirke smittespredningen ved at det lages en ny «fiktiv» lokalitetsstruktur i PO3, og simuleres smittespredning i den nye strukturen. På samme måte vil det være mulig å simulere hva som skjer dersom produksjonstiden i sjø forkortes ved økt bruk av storsmolt, dersom lusegrensen endres, eller dersom det innføres obligatorisk vaksinasjon mot PD.
Oppsummert synes det å være et stort potensial for å redusere smitten mellom lokaliteter, men vi ser behov for en mer omfattende utredning av ny lokalitetsstruktur, som tar hensyn til belastning på villfisk, spredning av organisk materiale, sonevise utsett og andre forhold nevnt over, før en konkret struktur kan anbefales. Ytterligere kommunikasjon med næringsaktørene, samt nasjonale, regionale og lokale myndigheter, er nødvendig før en kan levere konkrete forslag til en optimalisert lokalitetsstruktur i PO3.
2 - Introduksjon
2.1 - Bakgrunn
Nærings- og fiskeridepartementet sendte 23. oktober 2019 en bestilling til Veterinærinstituttet (VI) og Havforskningsinstituttet (HI) hvor en analyse av ny lokalitetsstruktur i PO 3 var etterspurt. Bestillingen etterspurte spesifikt:
- analyse av smittespredning mellom lokaliteter ved dagens struktur
beregning av smittespredning mellom omstrukturerte lokaliteter
vurdere effekten av å lukke slaktemerdene i området
Målet med bestillingen var: "Å få kunnskap om omstrukturering av lokalitetene i produksjonsområde 3 kan gi mindre smittespredning mellom lokalitetene." En forutsetning var at produksjonen i området ikke skulle være mindre enn dagens produksjon. Videre er det antatt at: "Mindre smittespredning mellom lokalitetene bidra til bedre fiskehelse og fiskevelferd, herunder mindre produksjon av lakselus".
Det ble i bestillingen bedt om å invitere oppdrettsselskapene med i prosessen for å sikre mest mulig hensiktsmessig scenariotesting, hvor erfaringsbasert kunnskap fra næringen er inkludert. Den 17. januar 2020 ble det arrangert et dialogmøte i Bergen, hvor representanter fra HI, VI, Mattilsynet, Fiskeridirektoratet og næringsaktører fra 18 oppdrettsselskap var til stede. Både VI og HI presenterte sine modellverktøy, og skisserte mulige måter å analysere effekten av restrukturering på smittespredning i PO3. Presentasjonene ble etterfulgt av innspill fra, og diskusjon med, næringsaktørene. I etterkant av dialogmøtet ble det sendt ut et nettskjema til alle næringsaktører, med mulighet for å gi innspill på spesifikke lokaliteter vedrørende flytting av MTB og innføring av teknologi.
Per 1/1-2020 var det i akvakulturregisteret oppført 149 lokaliteter i PO3, der 135 var matfisklokaliteter, 7 var stamfisk-lokaliteter, og 7 var slakteri-lokaliteter. Av disse lokalitetene hadde 130 matfisklokaliteter og 5 stamfisk-lokaliteter rapportert lusetall i perioden 2012-2019 (Figur 1).
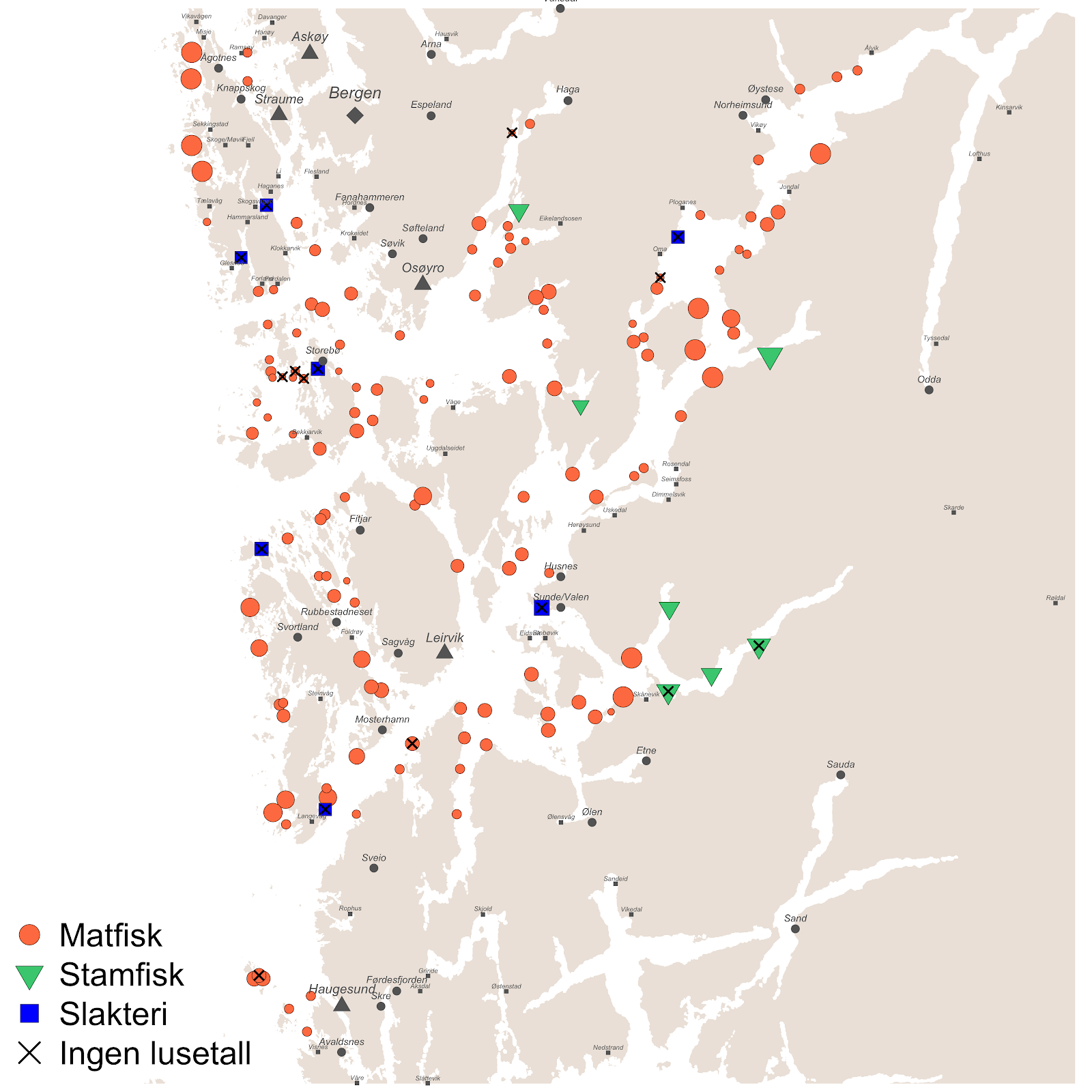
2.2 - Innspill fra næringen
Etter dialogmøtet mellom VI, HI og næringen der oppdraget, metode og foreslått framgangsmåte ble presentert, ble aktørene invitert til individuelt å komme med innspill på egnethet av lokaliteter. Innsamlingen av innspill ble administrert av HI, men utarbeidelse av spørsmålene som ble stilt til næringsaktørene, og bearbeidingen av innspillene som kom, ble utført i samarbeid mellom begge instituttene.
Innsamlingen av innspill ble primært utført ved et spørreskjema, der oppdretterne fikk anledning til å komme med kommentarer på hver enkelt lokalitet i produksjonsområdet (Figur 2).
Vi mottok innspill fra totalt 14 næringsaktører. Innspillene ble sendt inn via det utsendte nettskjemaet, men noen kom også i form av skriftlige brev. Nettskjemaet ga rom for relativt omfattende skriftlige kommentarer, men de fleste aktørene foretrakk å sende disse per brev.
Det primære formålet med å sende ut skjemaet var å få forslag til lokaliteter der oppdretterne selv vurderer at en kan øke biomassen, eventuelt også forslag til nye lokaliteter. Det var også mulighet å foreslå å legge ned lokaliteter (stenge ned anlegget). Det var en bekymring fra enkelte at selskap skulle foreslå nedstenging eller reduksjon av biomasse/lokaliteter hos andre selskap. Det skal her understrekes at ingen av næringsaktørene sendte inn slike forslag.
Vi fikk tilbakemeldinger rettet mot spesifikke lokaliteter som følger:
Økning i MTB på 32 lokaliteter (lokaliteter der en vurderte at MTB kunne økes)
10 nye lokaliteter ble foreslått
Det ble foreslått å stenge 2 lokaliteter
Det ble foreslått å innføre ny teknologi, eller videreføre ny teknologi på 6 lokaliteter
I tillegg til kommentarene for spesifikke lokaliteter, ga næringsaktørene både positive og negative tilbakemeldinger til initiativet. Det ble også gitt en god del andre faglige innspill. Flere av næringsaktørene la mye arbeid inn i tilbakemeldingene.
Forslag og kommentarer, er oppsummert her:
Næringen er generelt positivt innstilt til ny teknologi for å redusere smitte mellom lokaliteter, men de teknologiske løsningene er ikke nødvendigvis modne i dag.
Mange, men ikke alle, aktører var positive til en omstrukturering av dagens lokalitetsstruktur til en totalt sett bedre struktur.
En omstrukturering ville være kostnadskrevende. Ansatte vil få andre arbeidsreiser, infrastruktur og logistikk, ikke minst med strømforsyning, ville måtte endres.
Det er ønskelig med en analyse av effekten på villfisk av å holde en maksimumsgrense på 0,2 eller 0,1 kjønnsmodne hunnlus over lengre tidsrom i året.
Det ble av noen uttrykt skepsis til modellene som blir brukt dersom de ikke tar hensyn til driftsmønstrene (eks. vekselvise utsett og brakklegging).
Vurdering må baseres på reelle luseutslipp, ikke teoretisk MTB (dette er nå forsøkt hensyntatt også i HI sin modell).
Det er ønskelig med en ny vurdering av brakkleggingsområder.
Det var en generell oppfatning at næringsaktørene selv kjenner sine lokaliteter best.
Næringsaktørene tok også hensyn til andre miljøparametre enn bare smittepress, blant annet MOM-undersøkelser.
Næringsaktørene ønsket generelt ikke å gi innspill på andre lokaliteter enn de de selv disponerer (dette var samtlige aktørers syn, og ingen slike innspill ble gitt).
Næringsaktørene tok i innspillene i liten grad hensyn til at noen lokaliteter vil gi økt smittepress mot produksjonsområde 4. Det inngikk i oppdraget fra Nærings- og fiskeridepartementet at dette var et viktig hensyn, men det ble antakelig ikke kommunisert tilstrekkelig tydelig fra HI og VI til næringsaktørene.
3 - Materiale og metode
Modellverktøyene på HI og VI er ulike, og kan derfor ikke besvare oppgaven på samme måte. De to tilnærmingene vil gi forskjellige resultater med tolkninger som er vanskelig å sammenligne direkte. De forskjellige tilnærmingene er imidlertid også en styrke, hvor løsninger som kommer ut fordelaktige i begge tilnærminger får høyere troverdighet. Dette øker sannsynligheten for at vi finner en strukturering av oppdrettslokaliteter som er bedre enn den vi har i dag. Videre følger en kort beskrivelse av de to instituttene sine modeller.
3.1 - Modeller og nettverksanalyse utført av HI
HI benytter seg av en hydrodynamisk sirkulasjonsmodell som grunnstein for sine analyser av spredning. Denne gir en beskrivelse av det fysiske miljøet (blant annet strømhastighet, temperatur og saltholdighet) for hele produksjonsområdet. Modellen inkluderer tidevann, vindpådrag, ferskvannsavrenning fra land og utvekslingsprosesser mellom fjord og havet utenfor. For detaljert beskrivelse av havmodellen se Albretsen m.fl. (2011), og beskrivelse av modellområdet se Appendiks 1.
Videre er det brukt en såkalt «individ-basert modell» (IBM) som forsøker å gjenskape en realistisk spredning av både lakselus og virus med havstrømmene. Disse IBM-ene er basert på beste kunnskap om utslipp, utviklingshastighet, dødelighet, smittsomhet og eventuell adferd i forhold til lys og saltholdighet av lakselus.
HIs modellsystem er designet for å håndtere smittespredning i sjøvann. Verktøyet er ikke egnet for å analysere andre smitteruter, f.eks. via brønnbåt, forflytning av utstyr, mannskap eller lignende. Analysene fra HI av potensiell smitte av både lakselus, ILA og PD mellom lokaliteter baserer seg derfor utelukket på vurdering av vannbåren smitte.
For å oppnå en reduksjon i antall matfisk-lokaliteter uten å redusere produksjonen i produksjonsområdet må nødvendigvis MTB økes på enkelte lokaliteter. Det foreligger ingen overordnet vurdering av hva som er øvre grense for MTB som lar seg drive med tilfredsstillende lusetall. Innrapporterte lusetall antyder at antallet lakselus som blir produsert ved lokalitetene øker i takt med MTB opp til en produksjon på 8000 tonn (for detaljert begrunnelse se Appendiks 1). Det er antatt at mengden viruspartikler som blir sluppet fra en lokalitet ved et eventuelt utbrudd er skalert med MTB.
For å illustrere potensialet som ligger i å redusere antallet lokaliteter, uten å redusere produksjonen, har vi i våre analyser flyttet produksjonen fra enkelte lokaliteter der nettverksanalysen viste høy smittespredning basert på tidligere rapporterte lusetall og spredningsmodellen og deretter fordelt denne MTBen til lokaliteter med lav smittespredning i nettverksanalysen. Videre beregner vi det totale smittepresset mellom anlegg i produksjons-området ved en stegvis reduksjon i antall lokaliteter.
Vi har også undersøkt hvordan en omstrukturering av MTB reduserer det totale smittepresset av lakselus sammenlignet med innføring av utslippsfri teknologi med tanke på lus (eks. lukket merd) på et økende antall lokaliteter, eller en tilsvarende bruk av «snorkelmerd», skjørt, eller tube-teknologi (der man oppnår en reduksjon av smitte fra andre lokaliteter på 75%) på et økende antall lokaliteter. Disse analysene ble både utført ved “tilfeldig” innføring av slik teknologi på et økende antall lokaliteter, eller ved strategisk å innføre slik teknologi på de lokalitetene som bidrog mest med smittespredning i nettverksanalysen (merk: her er meningen med strategisk definert som den optimale innføringen av teknologi/refordelingen av MTB som ga minst total smitte i nettverksmodellen).
3.1.1 - Lakselus
For å simulere spredning av lakselusnauplier og kopepoditter i det horisontale planet brukte vi Runge-Kutta 2. ordens metode basert på strømfeltene fra havmodellen som implementert i partikkelspredningsmodellen LADIM (Lagrangian Advection and DIffusion Model, Ådlandsvik 2020, se Myksvoll et al. 2018 for lignende applikasjon). Simuleringen ble kjørt i januar 2019 og beregner smittespredning i perioden 1/3-2017 til 31/12-2018. Det er ønskelig med en lang periode for å inkludere et representativt utvalg av miljøforhold som forekommer naturlig i systemet. I partikkelspredningsmodellen slapp vi ut en partikkel fra hver lokalitet hver time i hele modelleringsperioden, som totalt ga 2.681.352 partikler. Her ble det brukt et tidssteg på to minutter og ved hvert tidssteg ble adveksjonen beregnet basert på strømmen hvor hver enkelt partikkel befant seg i med en medregnet tilfeldig horisontal diffusjon på opptil 0,2 m 2 /s.
Lakselusens adferd, dødelighet, individuell respons på miljøet og utvikling ble implementert som beskrevet i Sandvik m.fl. (2020). Videre ble infeksjonssannsynligheten til super-partiklene modellert som funksjon av alder (i døgngrader) og temperaturen i den vannmassen individet befant seg i (Skern m.fl. til fagfellevurdering). Utslippet av pelagiske lakselus fra lokalitetene ble skalert etter tidligere rapporterte lusetall (se Appendiks 1, kapittel 1 for detaljert beskrivelse)
For å beregne potensiell smitte av lakselus mellom lokaliteter tok vi utgangspunkt i de geografiske koordinatene til hver super-partikkel til enhver tid. Hvis en super-partikkel sluppet ut fra lokalitet A befant seg innenfor en radius på 160 m rundt lokalitet B ble smitteantallet fra lokalitet A til lokalitet B estimert som produktet av antall gjenværende super-partikler (som var gradvis avtagende over tid på grunn av naturlig dødelighet) og infeksjonssannsynligheten avhengig av temperatur og alder på lakselusen (Skern m.fl., 2020).
Antall lakselus sluppet ut fra lokalitet A som ble registrert smittet til lokalitet B ble så talt opp for alle anlegg og dannet en matrise med 135 x 135 elementer, hvor vert element viser smitte mellom to anlegg. Matrisen inneholdt da den totale to-veis smitten mellom hvert par av lokaliteter ble lagret, og er heretter referert til som «konnektivitetsmatrisen».
3.1.2 - Virus (PD og ILA)
Havforskningsinstituttets modellsystem for virus omhandler transport og spredning med strømsystemet og tar ikke for seg andre smitteveier. Modellen for virus er ikke publisert, men er i utgangspunktet lik modellen for lakselus, der samme strømmodell for samme tidsperiode er lagt til grunn og simuleringene bruker samme partikkeltransportmodell (LADiM). En viktig forskjell er at virus ikke kan svømme vertikalt slik som lakseluslarver og annet dyreplankton. I modellen blir derfor virus transportert med strømmen i et fast dyp (5 m) representativt for strøm nær overflaten. Andre forskjeller ligger i kildeledd, dødelighet og nødvendig smittedose.
Lakselus finnes mer eller mindre til enhver tid på alle lokaliteter og rapporteres ukentlig til Mattilsynet. Dette gjør at en kan simulere en tidsperiode med realistiske utslipp av lakseluslarver fra eksisterende lokaliteter. Selv om virusangrep av PD/SAV opptrer nærmest endemisk i PO3 (61 henholdsvis 64 påvisninger i Hordaland i 2018 og 2019), så opptrer smitte fra en lokalitet over kortere perioder på få uker. En realistisk simulering av en tidsperiode med rapporterte virusangrep er interessant for det aktuelle sykdomsutbruddet, men forteller lite om det generelle smittepotensialet fra ulike lokaliteter. Virus-simuleringene her er derfor potensielle, de kartlegger smittenettverk mellom eksisterende og eventuelt nye lokaliteter. Kildeleddet skaleres derfor med MTB ut fra forutsetning at et stort anlegg er potensielt en større smittekilde. Selv om det er urealistisk, pågår utslippene i modellen hele tiden, dette for å over tid oppsummere ulike vær- og strømsituasjoner på en representativ måte. Det absolutte nivået er mindre viktig, smittetallene brukes bare komparativt til å sammenligne ulike smitteveier.
Dødeligheten (mer presist raten hvor virus opphører å være smittsomt) er en viktig faktor for spredningsmønsteret av virus. I sammenheng med strømstyrken bestemmer dette hvor langt smitten kan spres. For en best mulig beskrivelse av smitteveier for virus er modellen parameterisert med en egen IBM for PD og ILA med dødelighet som funksjon av omgivelsestemperatur og tid, og kritiske smittedoser som gitt i Appendiks 1.
3.1.3 - Nettverksanalyser
I en stegvis prosess ble produksjonen ved én og én lokalitet lagt ned i modellen, basert på dens smitte til andre lokaliteter (i.e. summen av lakselus fra lokalitet A som ender opp i nærheten av lokalitet B , C , D , ...). Avhengig av scenario som ble testet (se seksjon 3.1.4), ble MTB på lokaliteten som ble lagt ned i modellen, fordelt på lokalitetene som hadde lavest smitte til andre anlegg og eksporterte minst lus til andre produksjonsområder (her nesten utelukkende til PO4). Konnektivitetsmatrisen ble deretter skalert i henhold til lokalitetenes “nye” MTB.
For å objektivt evaluere klynger av lokaliteter som var sterkere koblet sammen enn andre, ble det også foretatt en klyngeanalyse basert på konnektivitetsmatrisen (Rosvall & Bergstrom 2008). Det betyr på generelt grunnlag at tiltak gjort innad i en klynge vil ha større effekt på lokalitetene i klyngen den tilhører enn på lokaliteter i andre klynger.
3.1.4 - Scenarioer testet
Basert på tilbakemeldingene fra næringsaktørene og HIs modeller ble det utviklet 3 scenarioer for videre testing. Scenarioene var utviklet fra innspill fra næringen og hvilke lokaliteter som modellen vurderte som sentrale bidragsytere til det totale smittenettverket. Da næringen kun foreslo å fjerne to anlegg ble det vurdert som et scenario uten særlig effekt for det totale smittenett-verket, og er ikke presentert her.
Følgende scenarioer er presentert videre i vurderingen fra HI:
I scenario 1 ( Innspill ) fjernet vi de 35 lokalitetene som modellen identifiserte som de sterkeste bidragsyterne i smittenettverket. MTB fra de fjernede lokalitetene ble flyttet til de 32 eksisterende lokalitetene som næringen hadde foreslått som egnet for økning i MTB.
Scenario 2 ( Modell ) og 3 ( Modell+10% ) er rene modell-scenarioer. Her flyttet vi MTB fra de 35 lokalitetene som bidrog med mest smitte til andre lokaliteter i nettverket (som i scenario 1), men der den tillatte biomassen fra de flyttede lokalitetene ble fordelt til lokalitetene som bidrog med minst smitte til andre lokaliteter i modellen, samt minimerte smitte til PO4.
For å illustrere potensialet ved den nye restrukturerte lokaliseringen, la vi i tillegg til 10% økt biomasse (72.000 tonn) i scenario 3 ( Modell+10% ) .
Lokaliteter som fikk tilført biomasse fra andre lokaliteter kunne ikke overskride 8.000 tonn i noen av scenariene, begrunnelse gitt i Appendiks 1.
3.1.5 - Lukking av slaktemerder og nye lokaliteter
Det ble fra Nærings- og fiskeridepartementet bestilt spesifikt en analyse av effekten av å lukke slaktemerdene. HI gjennomførte en intervjurunde samtlige slakterier i PO3 der det fremkom følgende informasjon:
Det foreligger ikke data på tellinger av fisk i ventemerder
Det er ingen fast periodevis syklus i når de slakter fisk
Normalt har de mindre fisk i ventemerdene enn MTB
Ved mistanke om sykdom på anlegget som slaktes, eller om de er båndlagt, holdes ikke fisk i ventemerder
Det er økende bruk av bløgge/slaktebåter, samt at det tas i bruk kar på land for oppbevaring av fisk.
Altså, selv om slakteriene har oversikt over hvem de har mottatt fisk fra, er det ingen som teller lakselus på fisken i ventemerder.
Vitenskapelig forsøk har indikert at selve trengningen og pumpingen av laks over i brønnbåt kan redusere antall lakselus. Et forsøk ved Gildeskål forskningsstasjon (GIFAS) i 2014 indikerte ett ikke-signifikant tap av voksne hunn-lus på 45% (fra 0,37 voksne hunn-lus per fisk) (Reynolds, 2014). Et senere forsøk utført ved GIFAS i 2015 indikerte at det var spesielt de fast-sittende og pre-adulte stadiene som ble redusert, mens det ikke ble observert reduksjon i voksne hanner eller hunner (Reynolds, 2015). Det er derfor trolig et vesentlig tap av lakselus fra fisken hentes på anlegget til den leveres på slakteriet, men tapet kan vanskelig estimeres.
Manglende oversikt over hvor mange lus som blir sluppet fra slakte-lokalitetene gjør vår analyse høyst usikker–men pga. antatt tap av lus under transport og at slaktemerdene sjelden er fulle, ble slakteriene tilegnet en nauplieproduksjon tilsvarende halvparten av en matfisk-lokalitet med tilsvarende MTB (rundet til nærmeste 1000 tonn). Slakteriene har tillatt MTB som varierer fra 450 til 1170 tonn, som i praksis vil si at slakteriene ble tillagt en nauplieproduksjon tilsvarende en typisk matfisk-lokalitet med MTB på 500 tonn (i.e. ½ av median nauplieproduksjon ved MTB = 1000 tonn, cf. Appendiks 1; Figur A1 D). På grunn av stor usikkerhet rundt nauplieproduksjonen på slakte-lokaliteter ble det gjort separate analyser der effekten av lukkingen av slaktemerdene ble vurdert.
Næringsaktørene foreslo til sammen 10 nye lokaliteter i spørreundersøkelsen. I en separat analyse ble egnetheten av disse 10 lokalitetene vurdert. Da hensikten med denne rapporten var å evaluere effekten av et redusert antall lokaliteter, og at dette nødvendigvis innebærer færre og større lokaliteter, ble nauplieproduksjonen på disse nye lokalitetene antatt å være tilsvarende en matfisk-lokalitet med MTB på 8000 tonn (i.e. median nauplieproduksjon ved MTB = 8000 tonn, cf. Appendiks 1; Figur A1 D).
3.2 - Modeller og analyser utført av VI
Veterinærinstituttet har gjennom de siste tiårene jobbet med utvikling av statistiske modeller for å beregne hvordan smittespredning skjer mellom havbrukslokaliteter. Disse modellene bygger på historiske registreringer av lakselus og sykdomstilfeller (for eksempel PD og ILA), og spesielle forhold rundt livssyklus til lakselus og virus’ overlevelse i vann. Slike modeller er godt egnet til å simulere hva som skjer hvis man endrer strukturen i oppdrett av laksefisk, ved for eksempel å flytte lokaliteter eller ved å flytte biomasse mellom lokaliteter (såkalte hva-hvis analyser).
I modellene inkluderes produksjonshistorikk, det vil si at man bruker data rapportert til Altinn om hvor mange fisk som var til stede på enhver lokalitet til enhver tid og hvor stor fisken var. Dette er viktig, da mengden av virus eller luselarver som utskilles er avhengig av hvor stor fisken er, hvor mange de er og om lokaliteten er aktiv. I tillegg brukes sjøavstanden mellom lokaliteter, der spredning mellom to lokaliteter avtar eksponentielt med økende avstand. Veterinærinstituttet har i samarbeid med Norsk Regnesentral utviklet ulike modeller for lakselus, PD og ILA, siden disse sykdommene ikke sprer seg på den samme måten. Alle modeller baserer seg på data fra virkeligheten, og de har løpende blitt korrigert og tilpasset, slik at de på best mulig måte gjenspeiler virkeligheten. For eksempel vet vi nå at PD skyldes to separate epidemier med to forskjellige virus, og dette er inkludert i den nyeste modellen for PD.
Før vi fikk denne bestillingen, hadde Veterinærinstituttet allerede begynt å jobbe med å tilpasse modellene slik at de kan brukes til hva-hvis analyser for lakselus, PD eller ILA; eksempelvis ved å øke biomassen eller ved å samle lokaliteter tettere sammen og lage større branngater mellom dem. Dette arbeidet er kommet lengst for lakselus, som beskrevet under.
3.2.1 - Lakselus
Veterinærinstituttet har brukt en tidligere publisert modell for å simulere hvordan lusesituasjonen ville utviklet seg i PO3 under forskjellige scenarioer (Aldrin, Jansen & Stryhn, 2019) i årene 2014-2017. Modellen beregner i grove trekk produksjon av luselarver i hver lokalitet som en funksjon av hvor mange hunnlus som var på lokaliteten (rapporterte gjennomsnitt*rapportert beholdning). Spredningen beregnes etter en avstandsfunksjon, og utviklingstider fra egg til voksne hunnlus beregnes ut fra temperaturavhengige utviklingsrater estimert fra innrapporterte data.
Beholdningsdata simuleres ikke, slik at det er de innrapporterte beholdningene som angir om en lokalitet er aktiv, og hvor mange fisk som befinner seg der til enhver tid.
Fordelen med en slik tilnærming er at den bruker nokså realistiske data, og vi slipper usikkerheten ved å simulere beholdningsdata i tillegg. Denne tilnærmingen gir også et mer reelt bilde av luseutslippene enn om man benytter MTB. En mulig svakhet i VIs tilnærming er imidlertid at spredningskomponenten er avstandsbasert og ikke fanger opp vanntransport gjennom havstrømmer. Dette kan gi økt usikkerhet for enkeltlokaliteter, spesielt i områder preget av sterke strømmer som ikke jevnes ut over en lengre periode. Detaljer og parametere som er brukt, er beskrevet i Aldrin m.fl., 2019.
3.2.2 - Virus (PD og ILA)
Når det gjelder effekten av endret lokalitetsstruktur på PD, har vi, tilsvarende som for lakselus, brukt en tidligere publisert modell for å simulere hvordan PD-situasjonen i PO3 ville utviklet seg under ulike scenarioer i årene 2014-2017 (Aldrin et al., 2015). Modellen har blitt oppdatert, slik at det nå tas høyde for at spredning av PD siden 2010 skyldes to ulike epidemier med spredning av to ulike virus, kalt SAV2 og SAV3. Den oppdaterte modellen baserer seg på hvordan PD har spredd seg fra januar 2004 til august 2017. Modellen beregner sannsynligheten for at hvert enkelt anlegg innenfor 100km av en gitt PD-infisert lokalitet vil bli smittet med PD-virus. Smittespredning er som for lakselus, en funksjon av lokalitetens størrelse og avstanden til andre anlegg i sjø. En sentral størrelse i modellen er smitteraten på de ulike lokalitetene. Smitteraten angir sannsynligheten for at en kohort, som ikke er smittet, blir smittet i nærmeste fremtid. Smitteraten på en gitt lokalitet vil variere med tiden og modellen tar høyde for bidrag fra ulike smittekilder hvorav de viktigste er omkringliggende anlegg og andre kilder, inklusive tidligere kohorter på samme anlegg. Det ligger noen antakelser til grunn for modellen, blandt annet at en lokalitet som er smittet med PD forblir smittsom frem til den slaktes ut, og at tiden fra infeksjon med SAV til PD oppdages er rundt 3 måneder. Siste oppdaterte parametere samt beskrivelse av oppdateringen er ikke publisert, men følger ved i Appendiks 2. Denne modellen ble i 2012 brukt til å simulere hva som ville skje dersom SAV2 fikk lov å spre seg uten ‘stamping-out’ nord for Hustadvika. Denne simuleringen viste seg å forutsi ganske bra hva som faktisk skjedde etterpå.
Veterinærinstituttet har i samarbeid med Norsk Regnesentral også utviklet en modell som simulerer smittespredning av ILA. Denne modellen er ikke publisert ennå, og det har ikke vært mulig å rekke å anvende den til scenario-simulering slik som for PD innenfor denne bestillingen. I modellen tas høyde for genetiske avstander mellom ILA-utbrudd og for den tilfeldige introduksjonen av HPR0 til virulent HPR-del. Modellen bygger ellers på de samme prinsippene som PD-modellen.
3.2.3 - Scenarioer testet
Veterinærinstituttet har simulert hva som på et teoretisk grunnlag vil skje dersom det blir færre og større lokaliteter i PO3. Dette har vi gjort ved å fjerne noen lokaliteter mens den biomassen som var på dem fordeles på andre lokaliteter med matchende kohorter. Dette gir et grunnlag for å finne ut om endring av lokalitetsstrukturen vil påvirke smittespredningen. Veterinærinstituttet har brukt to ulike tilnærminger for å velge hvilke lokaliteter som skal «tas ut»:
A. Et tilfeldig utvalg blant de nåværende lokalitetene, der det ikke tas høyde for størrelse eller plassering. Her har vi gjort en randomisert lukking av 20, 30, 40 eller 50% av lokalitetene og overførsel av produksjonen deres til nabolokaliteter (Scenario 2-5 i Tabell 3.1). For hvert scenario, har vi gjentatt en randomisert trekning av hvilke lokaliteter som skal lukkes 10 ganger, og deretter kjørt tusen simuleringer for hver. Totalt 40.000 simuleringer.
B. Basert på nettverksanalysene til Havforskningsinstituttet (cf. 3.1.3 Nettverksanalyser), har vi tatt ut de 10, 20, 30, 40 eller 50% av lokalitetene som er identifisert av modellene som å være viktigst i et smittespredningsperspektiv (Scenario 6-10 i Tabell 3.1). For hvert scenario, har vi som i A, kjørt tusen simuleringer, totalt 5.000 simuleringer.
I tillegg har vi kjørt en simulering basert på dagens lokalitetsstruktur; et såkalt ‘baseline’ scenario. (Scenario 1 i Tabell 3.1) Denne simulering har vi også gjentatt tusen ganger.
Beholdning i lokalitetene som ble tatt ut ble fordelt på andre lokaliteter med tilsvarende tidspunkt for oppstart av generasjoner. Figur 3 viser et eksempel på en randomisert omstrukturering der 40% av lokalitetene tas vekk og biomassen fordeles på andre lokaliteter.
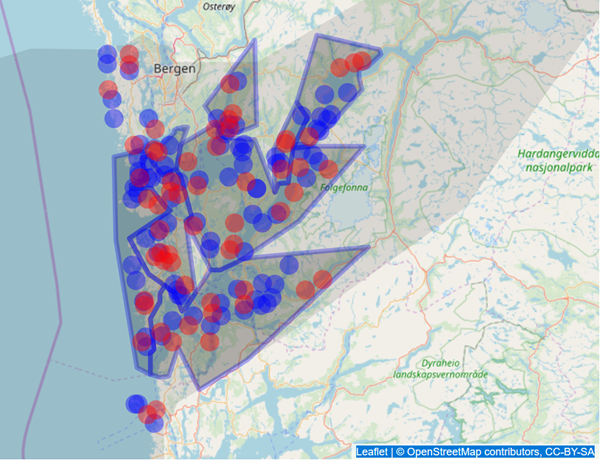
Scenario 1 og 6-10 er kjørt både for lakselus og PD, mens scenario 2-5 er kjørt bare for lakselus.
| Scenario | Navn | Beskrivelse |
| 1 | Baseline | Nåværende lokalitetsstruktur |
| 2 | VI20 | Tilfeldig lukking 20% av lokaliteter |
| 3 | VI30 | Tilfeldig lukking 30% av lokaliteter |
| 4 | VI40 | Tilfeldig lukking 40% av lokaliteter |
| 5 | VI50 | Tilfeldig lukking 50% av lokaliteter |
| 6 | HI10 | Strategisk lukking 10% av lokaliteter |
| 7 | HI20 | Strategisk lukking 20% av lokaliteter |
| 8 | HI30 | Strategisk lukking 30% av lokaliteter |
| 9 | HI40 | Strategisk lukking 40% av lokaliteter |
| 10 | HI50 | Strategisk lukking 50% av lokaliteter |
3.2.4 - Analyse av effekten av lukkede slaktemerder
I bestillingen var det ønske om å teste effekten av å lukke slaktemerder. Som beskrevet i avsnitt 3.1.5 ble det innhentet opplysninger fra samtlige slakterier i PO3. Det fremgikk av dette, at det ikke er mulig å vite hvor mange fisk som er i disse ventemerdene over en lengre periode. VI sine modeller har en oppløsning på en måned, da det er for dette tidsrommet at beholdningsdata rapporteres. For å kunne inkludere slaktemerder i analysene, ville vi være nødt til å gjøre en del antakelser om hvor mange fisk som forventes å være i slaktemerder på hver slaktelokalitet, hvor mange lus disse har og hvor lenge de står der. Resultater fra analyser basert på såpass usikre antakelser ville trolig ikke bidra i noen troverdig grad til vurderinger, og det er derfor ikke utført slike analyser.
4 - Resultat
4.1 - Analyser fra HI
4.1.1 - Lakselus med dagens lokalitetsstruktur
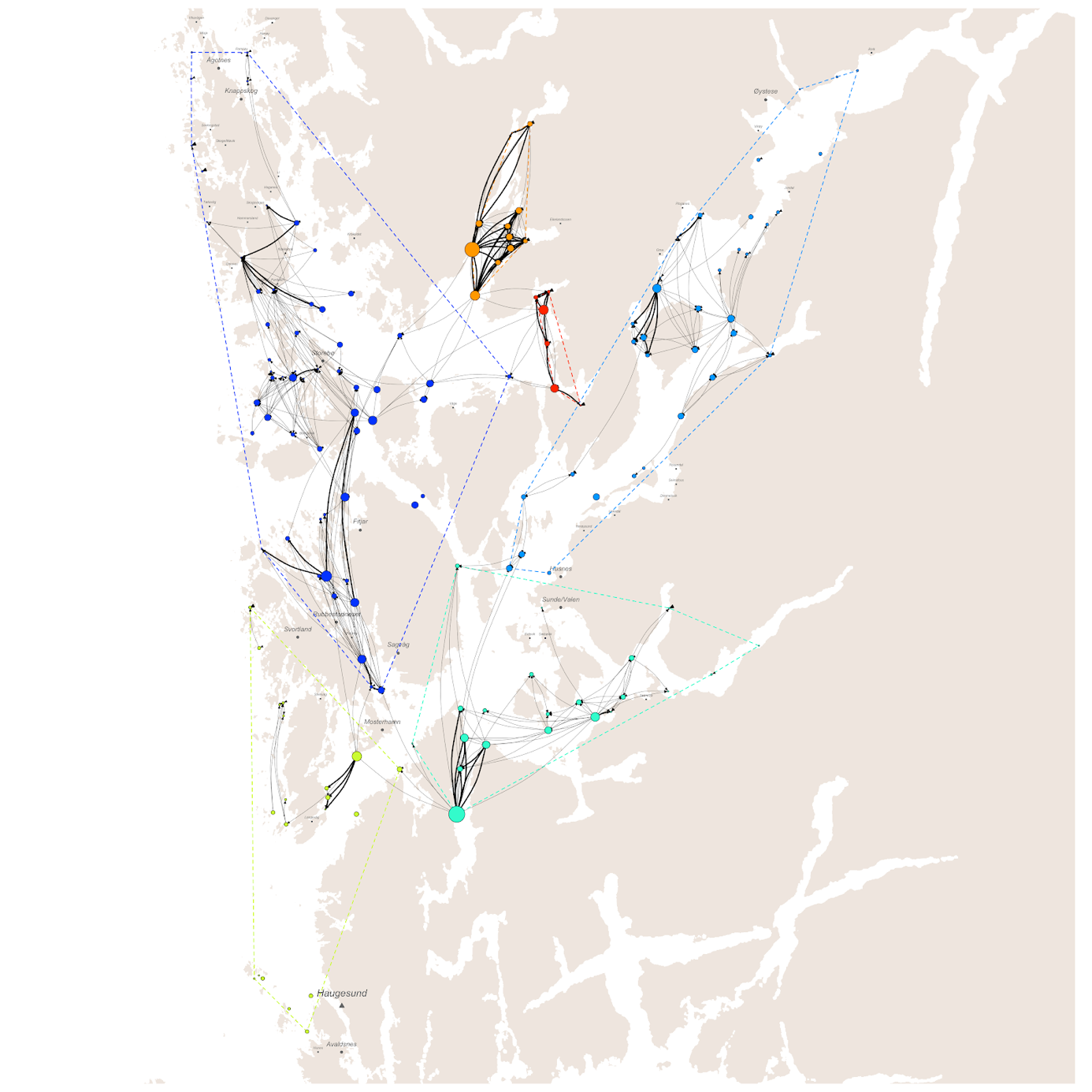
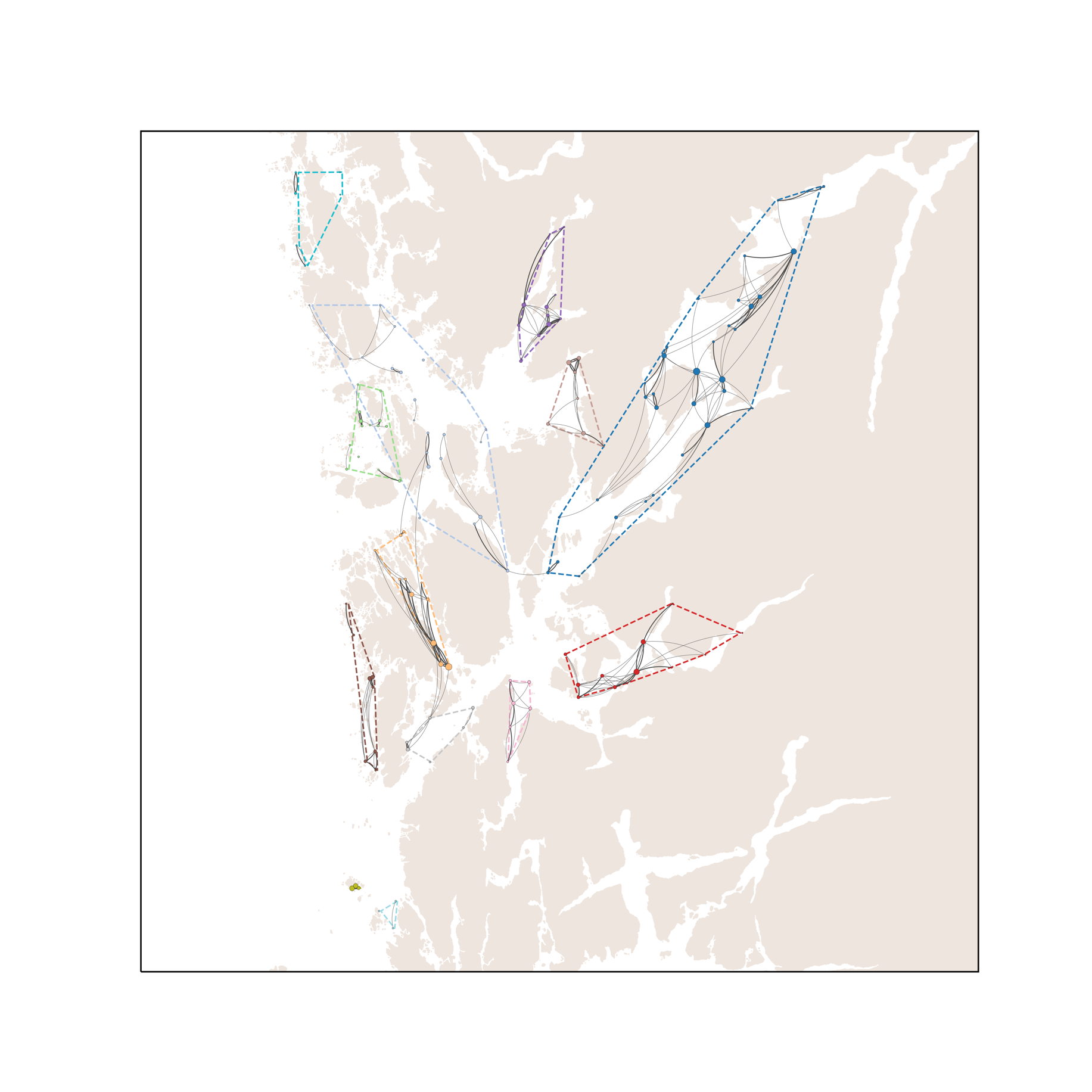
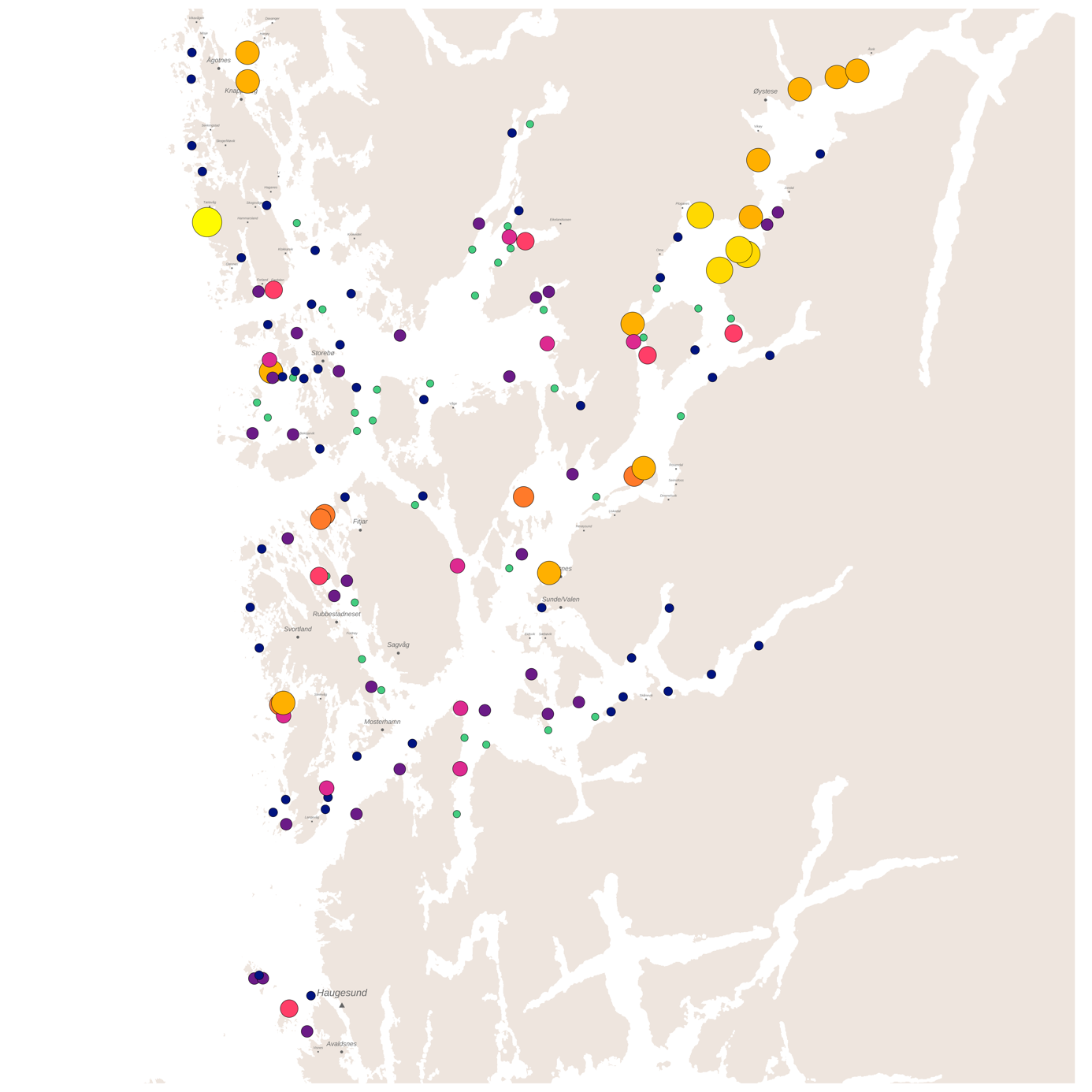
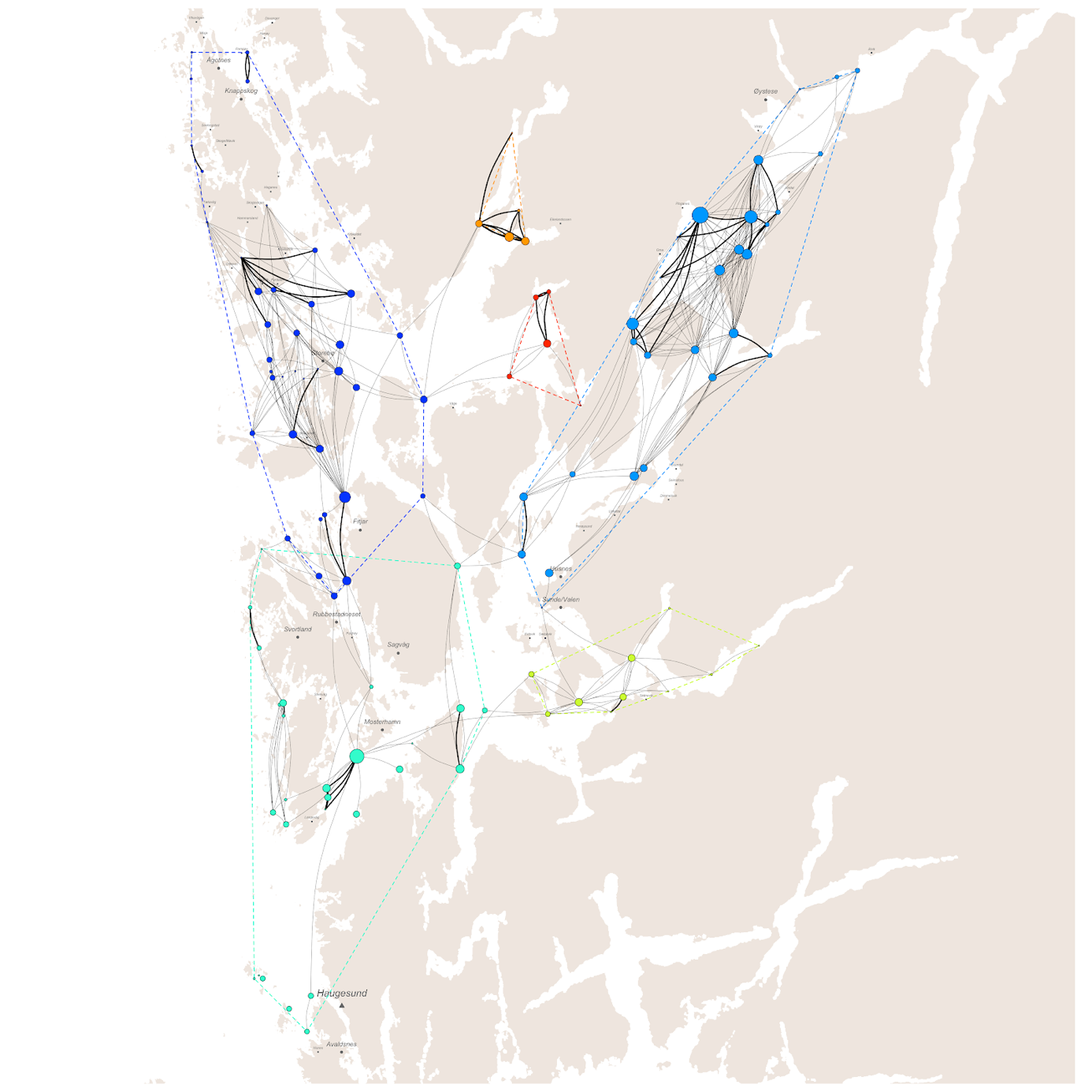
Basert på tidligere produksjon av lakselus på lokalitetene og modellert spredning med havstrømmene i løpet av de planktoniske stadiene, trer det frem et smittenettverk bestående av seks klynger av lokaliteter med en høy grad av konnektivitet innad. Den største klyngen består av 46 lokaliteter, distribuert fra Øygarden i nord til Stord og Tysnes i sør. Spredning innad i denne klyngen står for 26% av alle smittetilfellene (Figur 4).
Høyest konsentrasjon av smitte mellom lokaliteter finner vi i Fusafjorden, Samnangerfjorden, Eikelandsfjorden og Ådlandsfjorden, som stod for 8% av den totale smitten i produksjonsområdet fordelt mellom 11 lokaliteter. Lavest konsentrasjon av smitte på tvers av lokaliteter finner vi i klyngen mellom Haugesund og Bømlo som bare sto for 5% av smitten mellom 20 lokaliteter. Til sammen består smitte innad i klyngene for 80% av den totale smitten på tvers av lokaliteter, der de resterende 20% av smittetilfellene foregår på tvers av klyngene.
For hele produksjonsområdet var det stor variasjon i hvor mye lus de ulike lokalitetene smittet hverandre med. Median lusesmitte til andre anlegg var ca. 25% høyere ved enkelte lokaliteter sammenlignet med lokalitetene med lavest smitte til andre. Smittebidraget i produksjonssyklusene med laveste nauplieproduksjon var for de fleste lokaliteter ikke så ulike smittebidraget ved median (typisk) nauplieproduksjon. Derimot var det enkelte lokaliteter som bidro stort til det totale smittebildet ved enkelte produksjonssykluser (se linje for maksimum nauplieproduksjon i Figur 5).
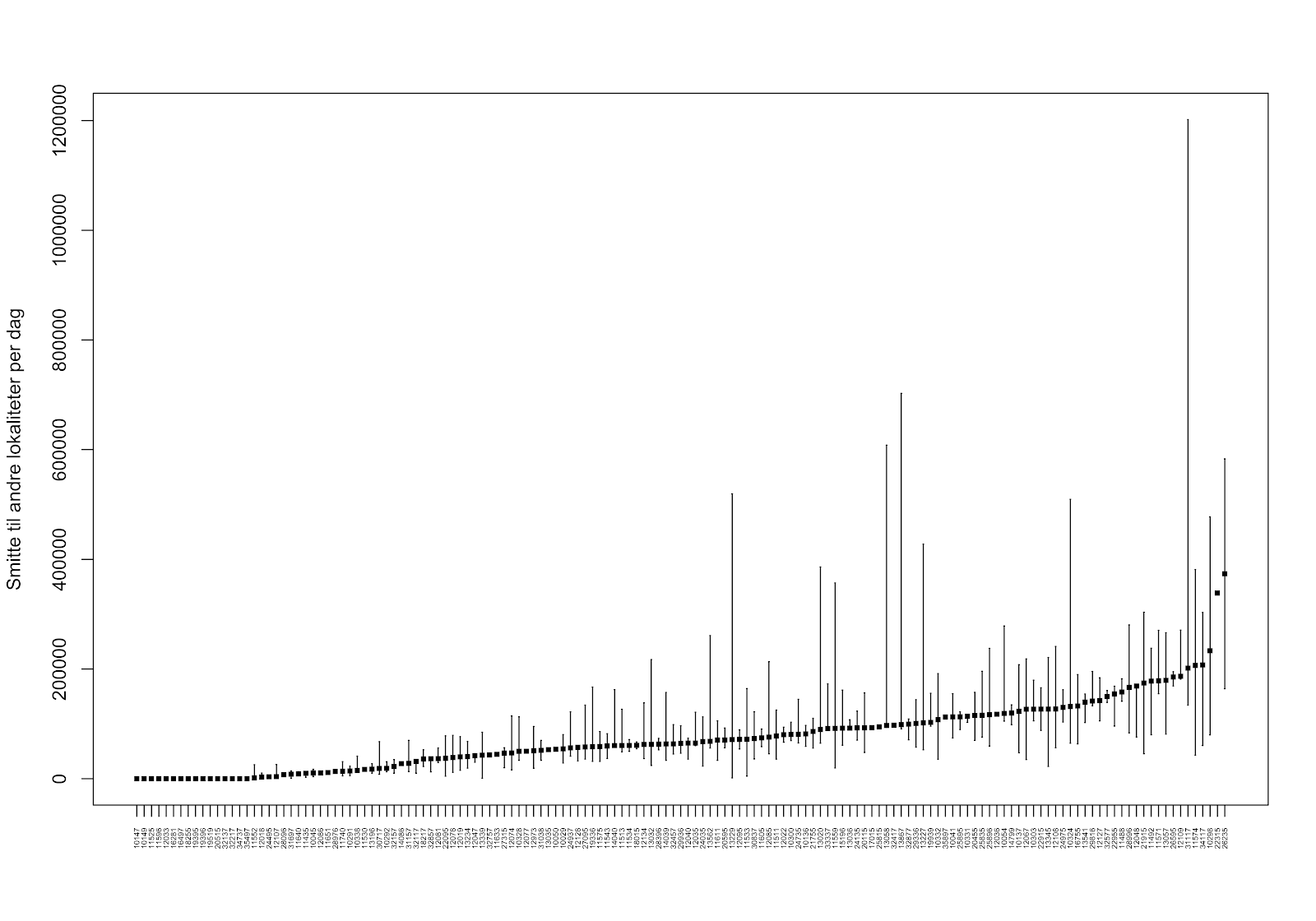
4.1.2 - Virus (PD og ILA) med dagens lokalitetsstruktur
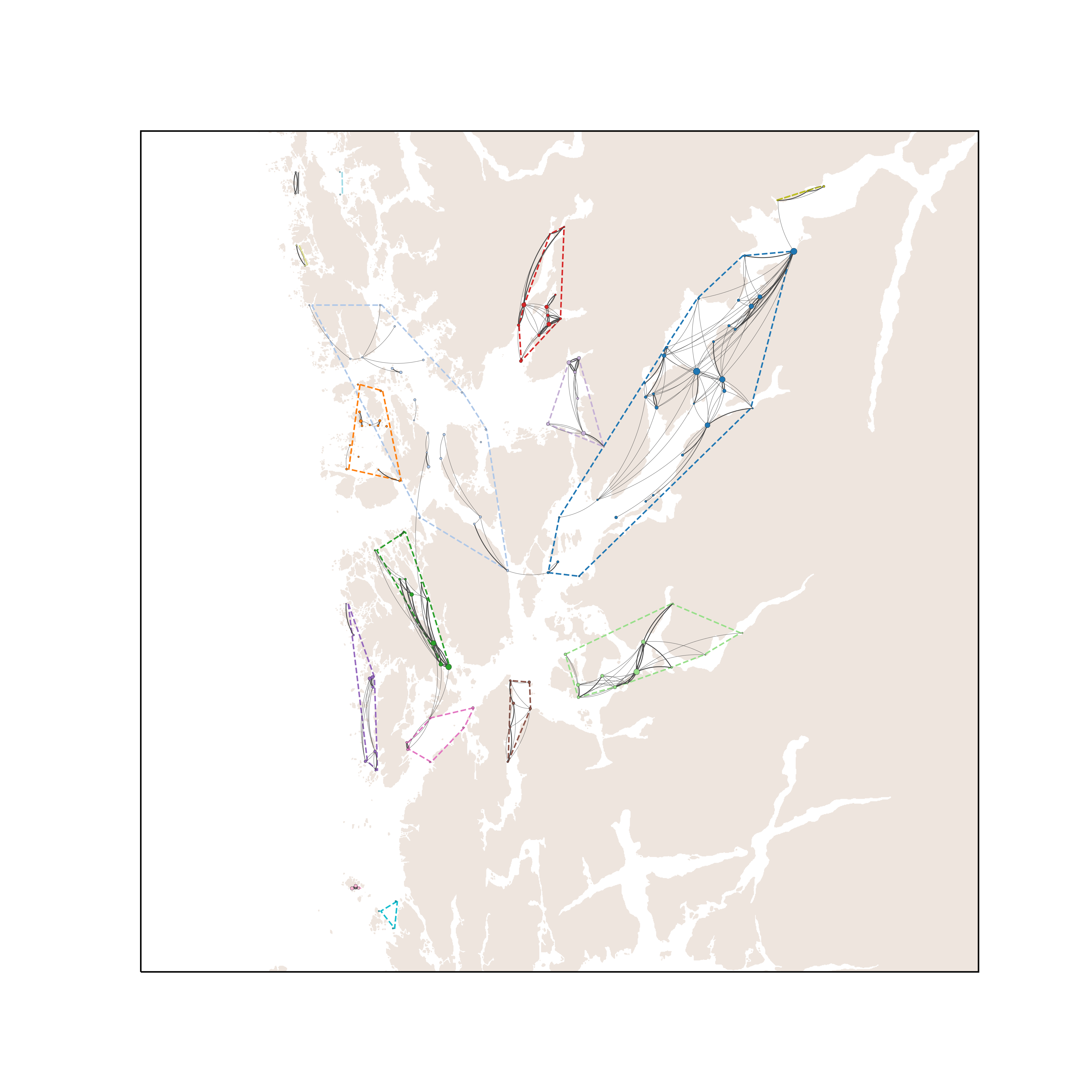
Med sammenlignbar dødelighet som for lakselus forventes ikke konnektiviteten for potensiell smitte av PD og ILA å være så veldig forskjellige. Dette bekreftes av Figur 6 og 7 under. Figur 6 viser det modellerte smittenettverket for PD. Terskelverdien for smitte (dvs. konsentrasjon av virus i vannmassene som trengs for at en lokalitet blir smittet) gir kortere smitteavstand og dermed flere klynger enn for lakselus. For PD, Figur 6, er det 13 klynger. Hovedforskjell fra lakselus (Figur 4) er at den store klyngen i nord-vest har skilt ut flere mindre klynger, Stord–Bømlo, Austevoll vest og Sotra, samt at klyngen i Ålfjorden–Skånevikfjorden er splittet i to.
Erfaringsmessig anses ILA for å ha kortere smitteavstand enn PD, selv om litteraturen kan antyde at dødeligheten av virus i sjøvann er lavere (Appendiks 1; Figur A4). Dette reproduseres i Figur 7, hvor smitteverket danner litt flere og noe mindre klynger for ILA enn for PD. De 16 klyngene framkommer ved at Ålvik-kysten inne i Hardangerfjorden skilles ut som egen klynge, og at den lille klyngen ved Sotra splittes i tre.
4.1.3 - Potensial for reduksjon i total smitte av lakselus
På grunn av den store variasjonen i smitte mellom lokalitetene forventes det et stort potensial for å redusere smitte ved å flytte produksjonen fra lokaliteter med stor smittespredningspotensiale til en lokalitet med lavere smittespredningspotensiale.
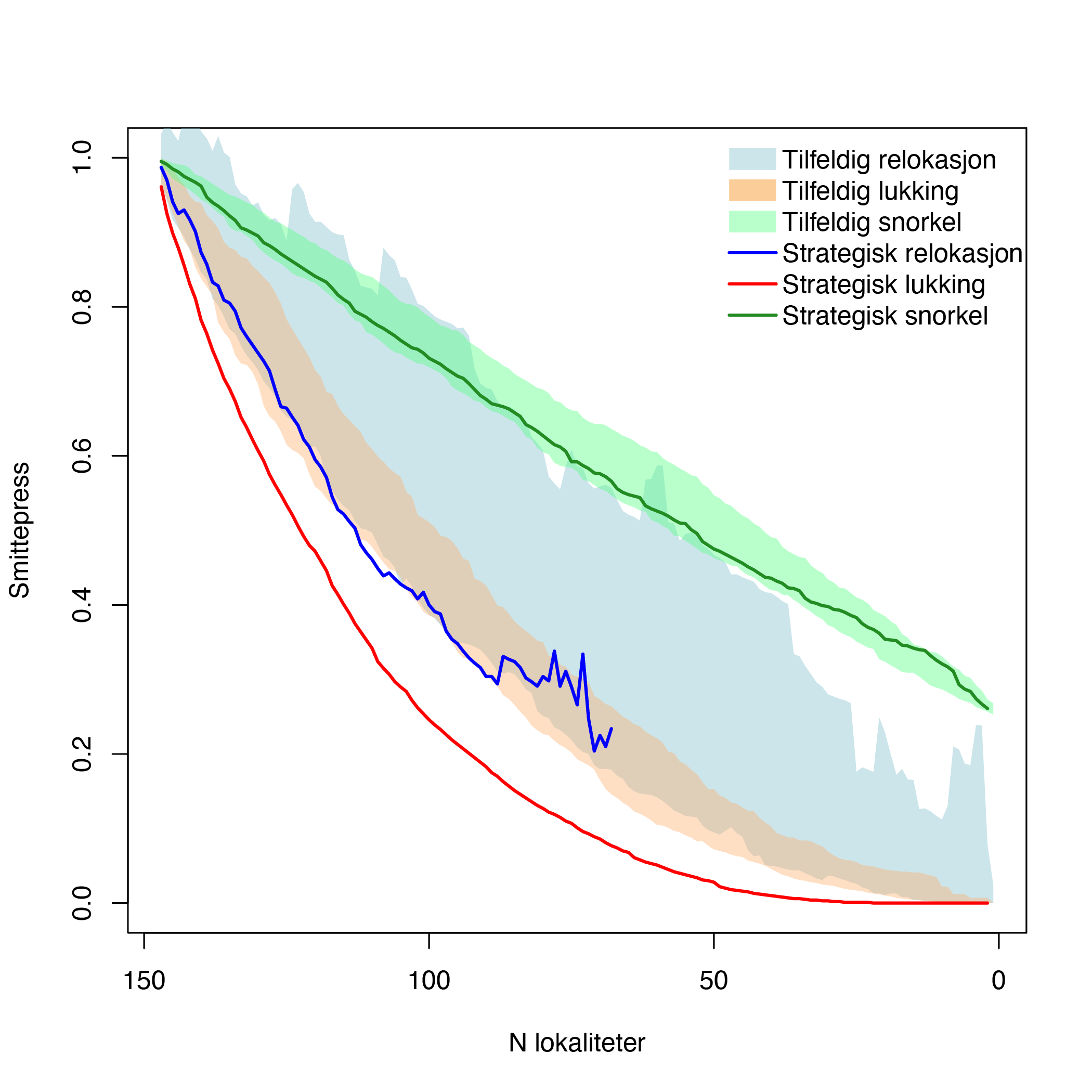
Ved å fjerne tilfeldige lokaliteter og refordele biomassen til de resterende lokalitetene får en til en viss reduksjon i smitte ettersom det blir færre lokaliteter og dermed større avstand mellom lokalitene (Figur 8). Ved å kjøre 100 ulike simuleringer med tilfeldig fjerning av lokaliteter ser en imidlertid at det er stor variasjon mellom de ulike utfallene i hvor effektiv en tilfeldig refordeling er for å redusere smittepresset (lyseblått felt i Figur 8). Derimot oppnår en raskere reduksjon i smittepresset ved strategisk å fjerne de lokalitetene som bidrar mest i smittenettverket og refordele til de beste lokalitetene (merk: her er meningen med strategisk definert som den optimale refordelingen av MTB som ga minst total smitte i nettverksmodellen, blå linje i Figur 8).
På samme måte ser en at det er mer effektivt å innføre lukket-merd teknologi på strategisk utvalgte lokaliteter som bidrar mye til smittespredning (rød linje i Figur 8), enn ved å innføre dette på tilfeldige lokaliteter (oransjefelt i Figur 8)
Tilsvarende oppnår en raskere reduksjon i smittepress ved strategisk å innføre «snorkel-merd» teknologi der man antar at innkommende smitte fra andre lokaliteter reduseres med 75% (grønn linje i Figur 8), enn ved å innføre denne på tilfeldige lokaliteter (lysegrønt felt).
4.1.4 - Scenariotesting
I scenariotestingen ble smittenettverket i de ulike scenariene vurdert opp mot dagens smittenettverk både for lakselus, PD og ILA. Tabell 4.1 under viser reduksjon i potensielt smittepress i prosent fra referanse-simuleringen med dagens lokalitetsstruktur og utslippshistorikk når det gjelder lakselus. Som nevnt i Appendiks 1 avhenger det modellerte potensielle smittepresset for virus av usikre terskelverdier. En sensitivitetsanalyse viser imidlertid at reduksjonen i potensielt smittepress er lite følsom for disse verdiene. Dette fordi det er de sterkeste smitteveiene som dominerer kvantitativt, noe som tyder på at nettverksanalysen er relativt robust når det gjelder virussmitte.
I scenario 1; Innspill , der modellen plukket ut lokalitetene som bidro med høyeste smittepress, og fordelte MTB fra disse til lokaliteter som næringen hadde foreslått økning på, oppnådde vi en total reduksjon i smitte på 9% for lus og 20–21 % reduksjon for virus, men med en økt eksport av lakselus inn til PO4 på 63% (Tabell 4.1).
En naturlig forklaring på denne økte transporten til PO4 var at mange av de foreslåtte lokalitetene lå i eksponerte, kystnære områder, som på grunn av gode strømforhold sannsynligvis vil tåle den økte organiske belastningen. De samme strømforholdene transporterer imidlertid også pelagiske lusestadier nordover og inn i PO4.
I det første av våre rene modell-scenarioer (scenario 2; Modell ), der både smitte innbyrdes i nettverket og eksport til andre produksjonsområder (dvs. fra PO3 til PO4) var minimert ved å flytte MTB fra 35 lokaliteter (som tilsvarte 24% av den totale MTB i produksjonsområdet, ≈176.000 tonn), ble det totale smittetrykket redusert med rundt 46% for lus og 29–30% for virus (Tabell 4.1). Omfordeling av MTB og smittenettvert for scenario 2 er vist i Figur 9 og 10. Eksporten av lus fra lokalitetene i PO3 til PO4 økte med 11% i scenario 2 sammenlignet med dagens struktur.
I det det andre av våre rene modell-scenarioer (scenario 3; Modell +10% ), der både smitte innbyrdes nettverket og eksport til andre produksjonsområder var minimert, og det i tillegg ble lagt til totalt 10% MTB (≈72.000 tonn) til produksjonsområdet, ble det totale smittetrykket redusert med 33% fra dagens situasjon, med tilsvarende reduksjon på 22 og 24% for PD og ILA. Flyttingen og økningen i MTB, førte til en økt eksport av lakselus inn i PO4 på 27%.
Reduksjonen i smittetrykk for de 3 scenariene var ulikt for lakselus og virus. For scenario 1 var smittepresset mest redusert for virus, men det motsatte var tilfelle i scenario 2 og 3. Den prosentvise reduksjonen i smittepress var ganske lik for PD og ILA, med litt sterkere reduksjon for ILA.
Her kan det presisere at det ikke ble foretatt en eksplisitt modellering av spredning av lus mellom lokalitetene i PO3 og PO4 i noen av scenariene, men kun spredningen inn over grensen til produksjonsområdet. Disse tallene for eksport inn i PO4 kan derfor anses som et grovt overslag.
| Scenario | Endring MTB | Endring eksport PO4 (lus) | Endring smittepress | ||
| Lus | PD | ILA | |||
| (1) Innspill | 0% | +63% | -9% | -20% | -21% |
| (2) Modell | 0% | +11% | -46% | -29% | -30% |
| (3) Modell +10% | +10% | +27% | -33% | -22% | -24% |
4.1.5 - Vurdering av lukkede slaktemerder og nye lokaliteter
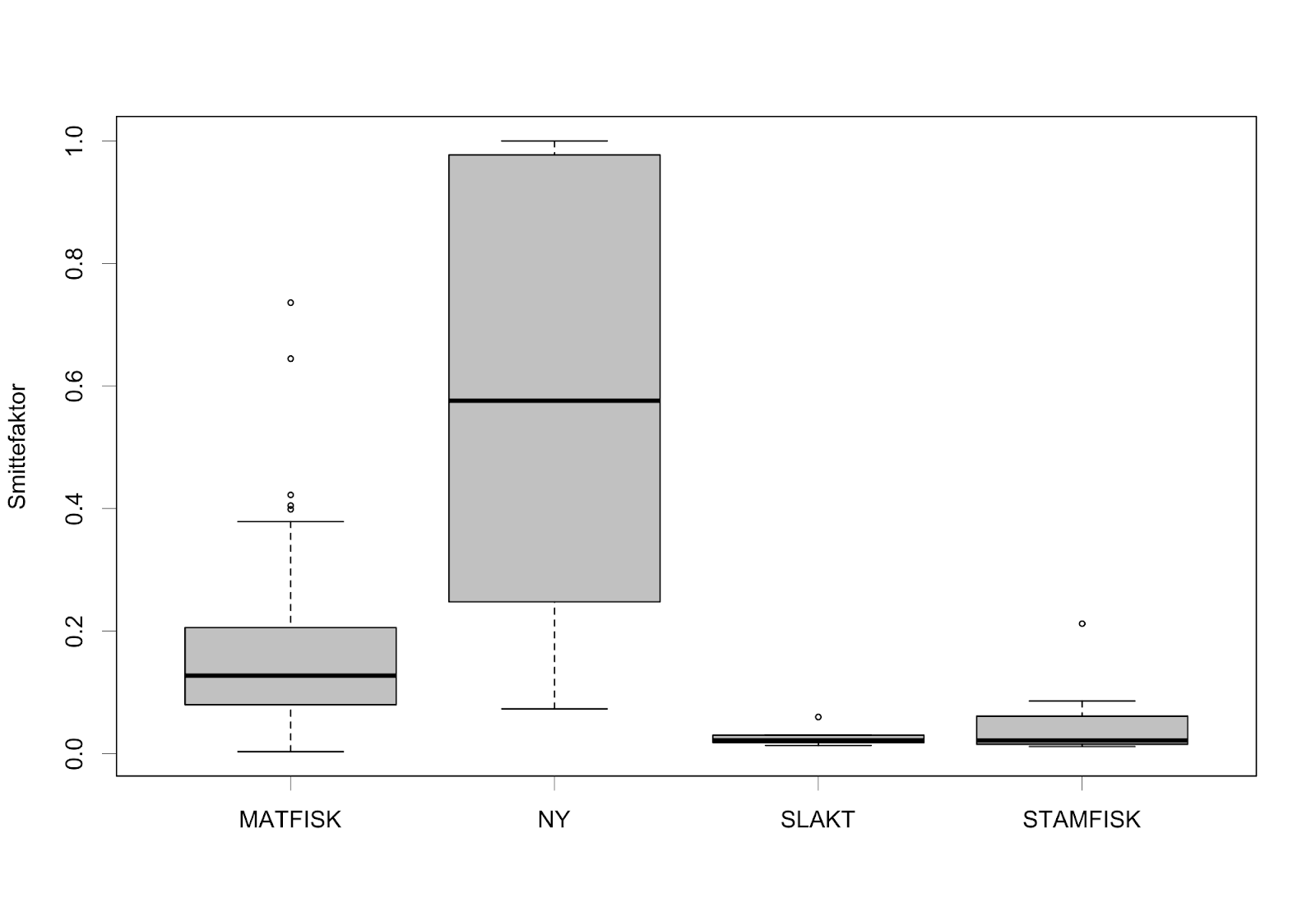
Basert på det grove overslaget i produksjon av nauplier ved slakte-lokalitetene, hadde spredning av lus fra slakterier kun en liten effekt på det totale smittebildet i PO3 i forhold til matfisk-lokalitetene (Figur 11). Lukking av alle de syv slakte-lokalitetene ville redusere den totale smitten med 8%, men merk den store usikkerheten rundt utslipp av lakselus fra slaktemerdene (se seksjon 3.1.5).
De nye lokalitetene som foreslått av aktørene i spørreundersøkelsen ga generelt mye høyere smitte til andre lokaliteter enn de allerede etablerte lokalitetene (Figur 11), der kun én av lokalitetene som ble foreslått ble vurdert som god, eller bedre enn de eksisterende basert på innbyrdes rangering av smitte til andre lokaliteter og eksport til PO4. Grunnen til denne lave egnetheten til de nye lokalitetene var grovt sett enten at de lå for nær- eller i stor grad bidro til smitte innad i klynger som allerede viste stor tetthet av smitte innad (cf. Figur 4); eller lå for nær- eller i retningsbestemte havstrømmer som førte smitte inn i PO4.
Ifølge slakteriene brukes ikke ventemerder ved slakteri ved mistanke om virus-smitte. Lukking av slaktemerder ble derfor vurdert som ikke relevant for vannbåren virussmitte.
4.2 - Resultater av analyser fra VI
Under presenteres resultatene av analyser av de scenarioene som er beskrevet i avsnitt 3.2. Dette er scenarioer som er basert på en «teoretisk» endring av strukturen i PO3, og hvor innspill fra oppdretterne ennå ikke har blitt tatt med. Analysene kan brukes til å belyse eventuelle effekter på smittepress og smittespredning dersom en reduserer antallet av lokaliteter, men fortsatt bevarer biomassen i området. Scenarioene er ikke realistiske ved det at de ikke tar høyde for hva som er praktisk mulig, men de kan peke i retning av bestemte mekanismer, for eksempel om det å fordele biomassen på færre lokaliteter vil ha noen effekt på lakselusbelastning og smittespredning mellom oppdrettslokaliteter i PO3.
4.2.1 - Scenariotesting for smittebelastning av lakselus
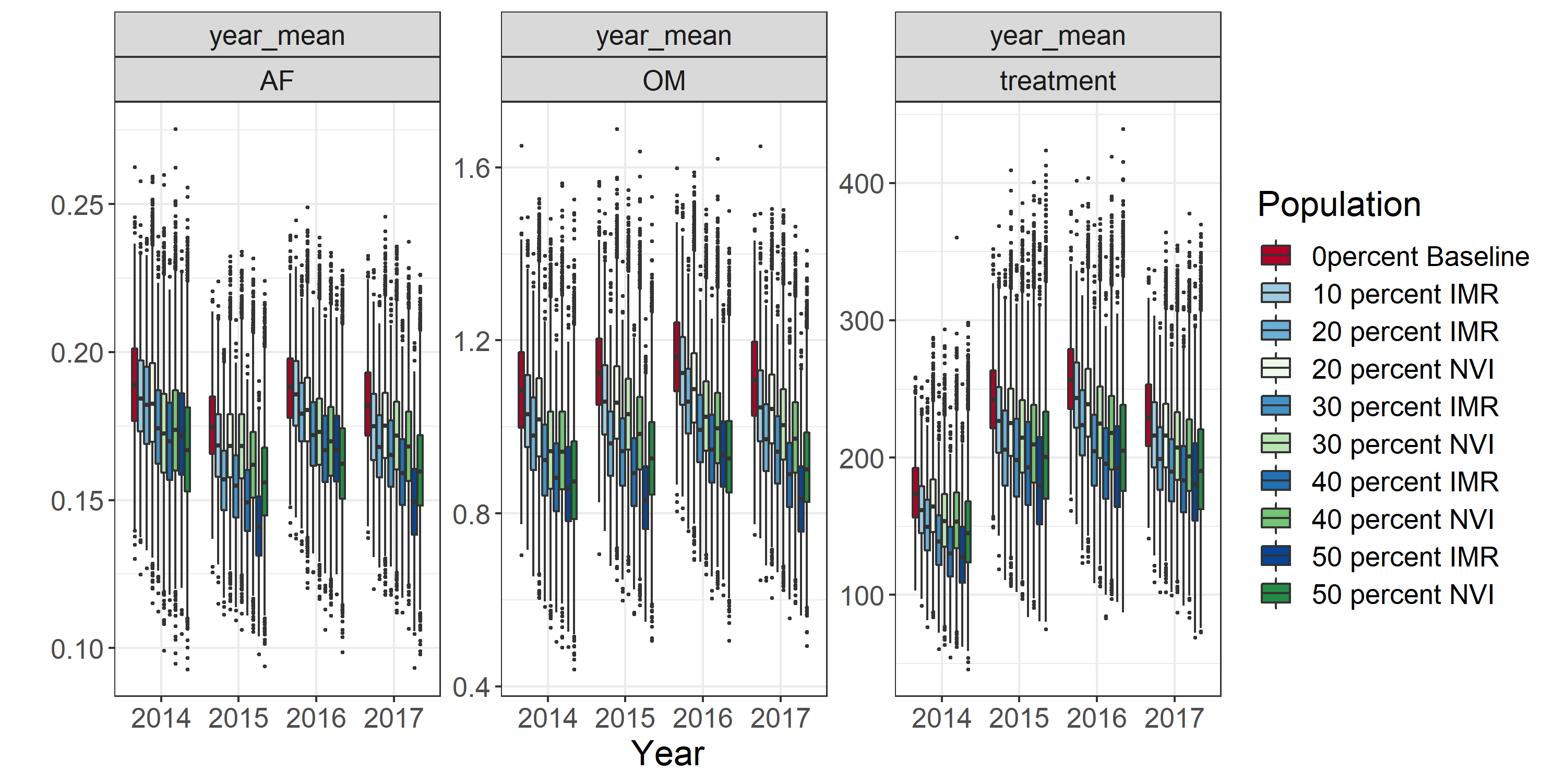
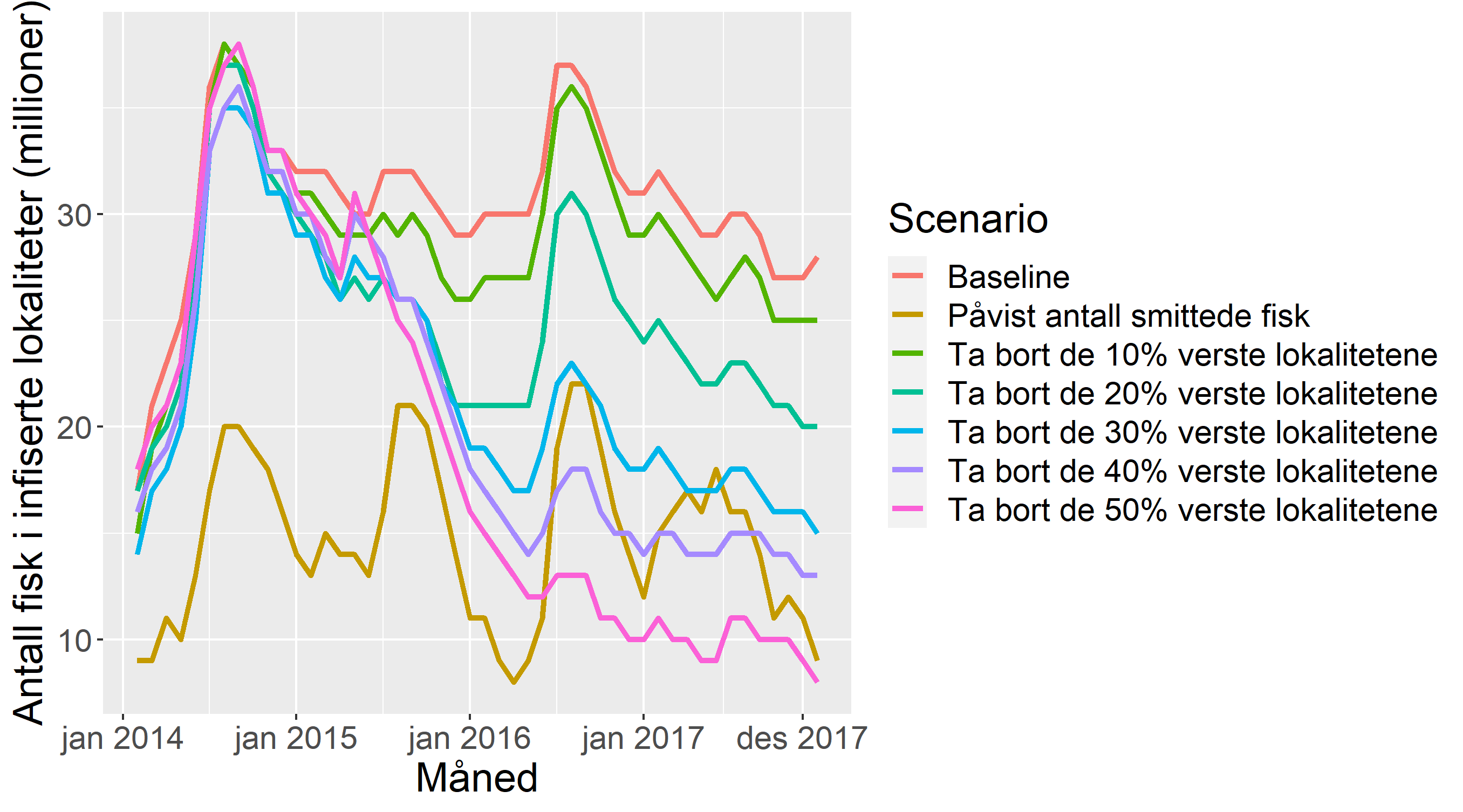
Figur 12 viser resultatene av simuleringene av scenario 1-10 for lakselus. Figuren viser at en gradvis reduksjon i antall lokaliteter gir en gradvis reduksjon både i antall lakselus og i antall behandlinger, gitt forutsetningene i modellen. Grovt regnet kan vi si at å fordele biomassen til halvparten så mange lokaliteter gir ca. 20% færre voksne hunnlus, 20% færre andre mobile lus, og 20% færre behandlinger mot lakselus. Behandlinger er skalert til antall millioner fisk som er behandlet. I 2014 er antall behandlinger betydelig lavere enn for de andre årene. Dette skyldes at modellen bruker 16 uker på å “kjøre seg opp”, og resultatene fra disse ukene rapporteres ikke. Dette fører til at antall behandlinger for 2014 telles opp for ukene 17-52, mot ukene 1-52 for de andre årene.
Alt i alt kan vi si at i den verden som beskrives av modellen, er det god effekt av å fordele biomassen ut på færre lokaliteter. Figuren viser også, at denne effekten blir større, dersom de lokalitetene som lukkes er strategisk valgt, dvs. at de er valgt på bakgrunn av hvor mye de bidrar til spredning av lus basert på HI sin nettverksmodell (de blå boksene i figuren).
4.2.2 - Scenariotesting for smittespredning av PD
Figur 13 viser resultatene av simuleringene av scenario 1 og 6-10 for PD. Figuren viser antall fisk som teoretisk sett blir smittet med PD gitt et av de simulerte scenarioene. For hvert scenario har vi startet simuleringen i januar 2014, der smittespredningen starter i alle aktive kohorter med påvist PD-smitte denne måneden. I simuleringene spres så smitten i månedlige steg til de omkringliggende lokalitetene, basert på sannsynligheter som beregnes i modellen. Når en lokalitet får status som smittet, beholdes denne statusen til lokaliteten slaktes ut.
Antall smittede fisk en gitt måned ble beregnet utifra antall lokaliteter med status som smittet, og beholdningen av fisk på disse lokalitetene. I figuren vises i tillegg til scenarioene, også beregnet antall smittede fisk basert på det faktisk observerte antall smittede lokaliteter.
Figuren viser at det i teorien oppnås færre PD-smittede fisk dersom en reduserer antall lokaliteter. Som for lakselus, ses det, at en gradvis reduksjon i antall lokaliteter gir en gradvis reduksjon i antall smittede fisk, gitt forutsetningene i modellen. Vi ser også at det observerte antall smittede fisk er noe lavere enn det som simuleres i baseline-scenarioet. Modellen vi bruker indikerer at rundt 1/3 av PD-infeksjonene ikke oppdages eller meldes, trolig fordi de ikke gir klinisk sykdom eller maskeres av annen sykdom. I tillegg viser modellen at det i gjennomsnitt går 3,6 måneder fra en lokalitet blir smittet til smitten oppdages. Man må dermed forvente at antall pågående infeksjoner i den virkelige verden er betydelig underrapportert.
Da obligatorisk screening for PD ble innført i høsten 2017 ble det da også en brå øking i antall rapporterte PD-tilfeller, noe som trolig skyldes at en del infeksjoner som ikke tidligere var oppdaget nå ble det.
Merk at vi i skrivende stund har hatt begrensede muligheter til å kvalitetssikre simuleringene for PD. Vi er blant annet usikre på hvorfor vi får den store økningen i antall tilfeller i starten av alle scenarioene, før trendene begynner å divergere etter et drøyt år. Det er imidlertid liten grunn til å tro at konklusjonene fra denne analysen blir endret, men vi velger likevel å fremheve at dette er foreløpige resultater som bør tolkes med en viss forsiktighet.
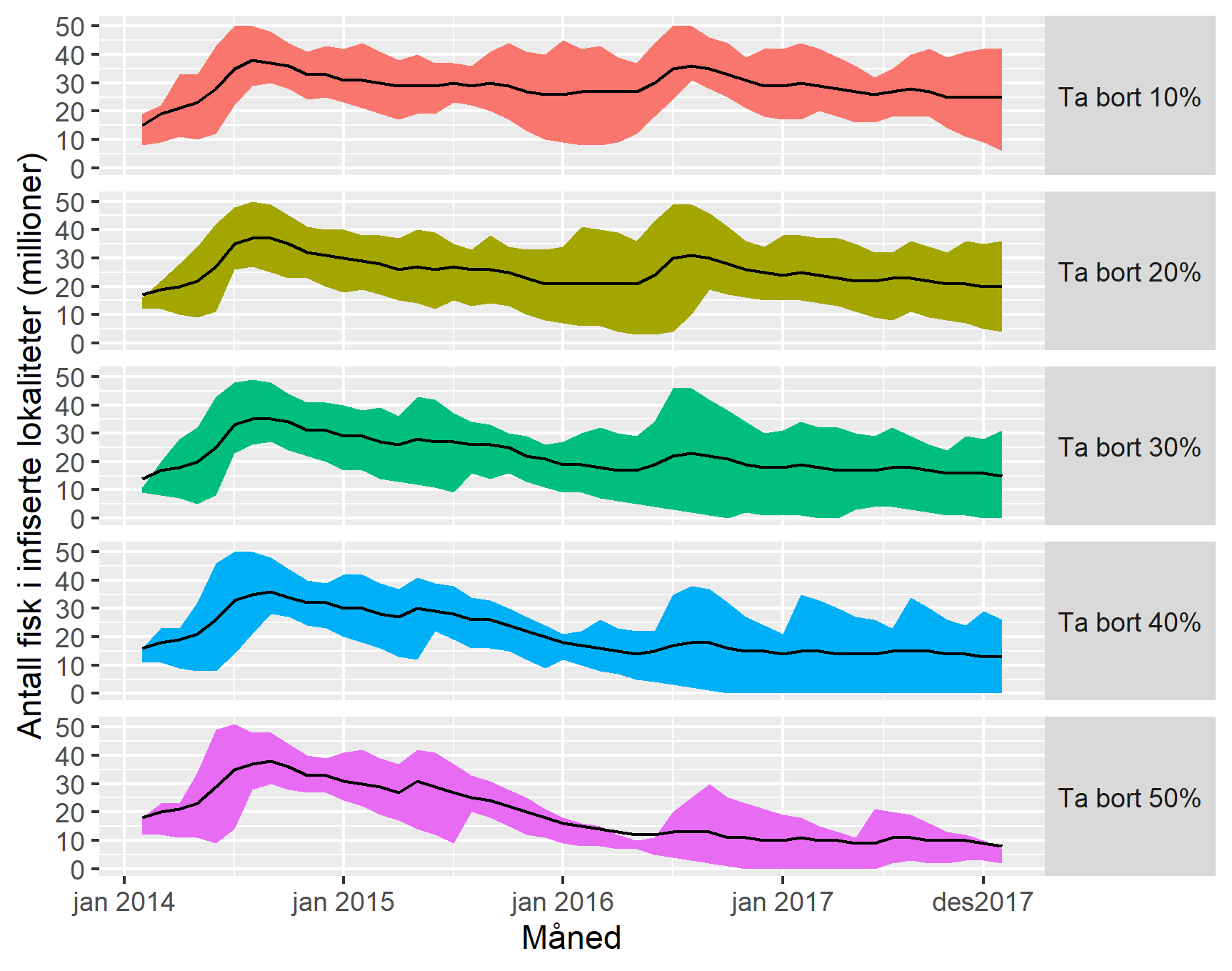
Trendlinjene i figur 13 er basert på gjennomsnittet fra 1000 simuleringer. Simuleringene bruker randomiserte sannsynligheter for smitte, og hver enkelt simulering vil variere noe. I figur 14 viser vi variasjonsbredden i simuleringene med 75% (øvre) og 25% (nedre) kvantilene fra simuleringene.
5 - Diskusjon
Aktiviteten i denne rapporten ble igangsatt av en bestilling i fra Nærings- og fiskeridepartementet, hvor en analyse av smittenettverket mellom lokaliteter i PO3, og eventuell gevinst i restrukturering ble etterspurt. Analyser gjennomført av både HI og VI viser at en kan oppnå redusert lusesmitte mellom oppdrettslokalitetene i PO3 ved å fordele samme biomasse som i dag på et redusert antall lokaliteter. Resultatene tyder på at man også vil oppnå reduksjon i smittepotensialet for både PD og ILA mellom lokalitetene ved en slik restrukturering. Det var stor variasjon i hvor stor grad lokalitetene påvirket de andre, noe som medfører at den vurderte effekten økte ved tiltak på strategiske lokaliteter i forhold til tilfeldig innsats.
Vurderingen av nettverket analysert av HI baserer seg på vannbåren spredning av lakselus, PD og ILA. Det finnes relativt mye litteratur som dokumenterer adferd, modning og smittsomhet for lakselus. For PD og ILA er litteraturen mer begrenset. HI valgte å analysere totalt 3 scenarier, som baserte seg på tilbakemeldinger i fra næringsaktørene og informasjon fra modellen. Analysene baserer seg for lakselus på tidligere produksjon av nauplier i de konkrete anleggene og drivbaner med strømmen. For virus bygger analysen på spredning, hvor vi vurderer potensielt påvirkede lokaliteter dersom det skulle forekomme utbrudd.
Analysene viser at man kan oppnå en betydelig reduksjon av smitte mellom anlegg ved å omstrukturere produksjonen. Modellen til HI viste at ved å flytte 28% av MTB i området, fra lokaliteter som i stor grad førte smitte til andre til lokaliteter som i liten grad smittet andre, ble det totale smittenettverket redusert med ~46% for lakselus, og ~30% for PD og ILA.
VI har brukt modeller som bygger på observerte lusetall og PD-tilfeller, samt virkelig produksjonshistorikk. Likevel er det viktig å holde seg for øye at resultatene er utført i en modellverden under gitte forutsetninger. Ved endringer i oppdrettspraksis i framtiden kan noen av forutsetningene bli ugyldige. Videre estimeres parametere i modellene med usikkerheter, som både kan skyldes rene tilfeldigheter og ikke-observerte variabler. Flytting eller nedlegging av lokaliteter kan endre sammensetningen av de ikke-observerte variablene i systemet, og medfører derfor noe usikkerhet. Ved å summere opp resultater fra flere tilfeldige omstruktureringer reduserer vi denne usikkerheten noe. Resultatene fra simuleringene viser en klar gradient i redusert lusebelastning når vi beveger oss mot større og færre lokaliteter. Dersom en tar ut nøkkellokaliteter identifisert av HIs nettverksmodell, vil denne effekten bli større. Effekten av å fjerne disse ‘nøkkellokalitetene’ gjenfinnes i resultatene av simuleringene for PD.
Arbeidsgruppen anser det som en stor styrke at to publiserte modeller med vidt forskjellig tilnærming identifiserer en positiv effekt med hensyn på lakselus av å omstrukturere biomassen til større og færre lokaliteter. Videre viser begge modelltilnærminger en ekstra effekt av å flytte biomasse fra nøkkellokaliteter identifisert med HIs nettverksanalyser, noe som underbygger indentifiseringen av nøkkellokalitetene.
Når det gjelder modellene for PD, så bruker HI en modell med parmeterisering som ikke er vitenskapelig publisert ennå, mens VI må kvalitetssikre sine simuleringsresultater bedre. Dette innebærer at resultatene for PD er noe mer usikre, men vi anser det likefult som sannsynlig at færre og større lokaliteter vil gi en positiv effekt på spredning av PD.
Modellene benyttet i dette arbeidet er egnet til å simulere hva som skjer med smittespredning dersom lokaliteter plasseres etter forslag fra oppdrettsaktørene. Dersom vi får inn spesifikke forslag til plassering av lokaliteter i PO3, kan både Havforskningsinstituttet og Veterinærinstituttet lage en ny «fiktiv» lokalitetsstruktur i PO3, og simulere smittespredning i den nye strukturen. En tolkning av resultatene vil belyse om en slik struktur kan være bedre eller verre enn dagens når det gjelder spredning av lakselus og PD. Dette arbeidet kan bli ganske tidskrevende, avhengig av hvor store mengder nye data må bearbeides.
Næringsaktørene har kommet med en del innspill til andre ting som bør tas hensyn til ved beslutning om en eventuell endring av lokalitetsstrukturen. For eksempel var det et ønske om å se på effekten av økt bruk av storsmolt. Dette kan simuleres ved å endre på produksjonshistorikken, slik at fisken står kortere tid i sjø, men med en høyere startvekt. Ved en slik analyse må det tas hensyn til at det kan være flere kohorter per lokalitet i samme tidsrom. Det er også mulig å simulere effekten av å endre lusegrensen på f.eks. antallet av behandlinger.
For lakselus vil vannbåren smitte være hovedkilden til smitte, men for virus kan også andre smitteveier bidra. For eksempel gjennom f.eks. via brønnbåt, forflytning av utstyr, mannskap eller lignende. Effekten en restrukturering av lokalitetene vil ha på andre smitteveier (mannskap, båter, utstyr, osv.), kan ikke vurderes med HIs hydrodynamiske modell som er benyttet her, og vurderingen fra HI i denne rapporten baserer seg utelukkende på den potensielle vannbårne spredningen mellom lokalitetene i tilfelle et sykdomsutbrudd.
For spredning av PD, har det kommet innspill om at effekt av brønnbåttrafikk bør inkluderes. En studie der det er brukt registrerte brønnbåtbevegelser til å modellere spredningen av PD i Norge i 2016, viser at analyser av brønnbåtbevegelser i stor grad kunne forutsi PD-status på anleggene (Haredasht et al. 2018). I modellen til VI som er brukt i denne rapporten, er denne typen smitte inkludert under smitte fra ‘andre kilder’. Det er trolig mulig å inkludere effekten av smittespredning med brønnbåt også i scenariosimuleringer, men det vil være en stor jobb, med usikker nytteverdi. Det ble av noen næringsaktører også nevnt muligheten for å simulere effekten, dersom det vaksineres mot PD. Vaksinasjon mot PD har god effekt på reduksjon av smittespredning, særlig etter introduksjon av en ny generasjon av vaksiner mot PD i de senere år. Smittespredningsmodellen til VI for PD egner seg bra til å simulere effekten av vaksinasjon mot PD. For å kunne gjøre dette, trengs det midlertid input i form av data på effekten av PD-vaksine på prevalens og utskillelse av virus fra vaksinerte og ikke-vaksinerte lokaliteter fra felt. Denne typen data er under inhentning i en pågående studie på VI.
Lokalitetene foreslått for økt MTB fra næringsaktørene førte i HI sin modell til økt transport av lakselus fra PO3 til PO4. Det er i etterkant vår vurdering at det burde ha blitt presisert tydeligere fra forskningsaktørene til næringsaktørene at transport fra PO3 til PO4 var et vesentlig moment i bestillingen fra Nærings- og fiskeridepartementet.
Få anlegg ble fra næringen foreslått for lukking, og en analyse basert kun på dette reduserte smitten av lakselus, PD og ILA i liten grad. Ved å en refordele biomassen fra lokaliteter som ble identifisert i nettverksanalysen til å bidra mye med smittepress til de 32 lokalitetene som næringsaktørene mente tålte økt biomasse, ble reduksjonen i smittepress begrenset og eksporten av lus til PO4 større enn dagens situasjon. Her skal det nevnes at ikke alle næringsaktørene meldte inn lokaliteter som kunne tåle økt biomasse, og tilbakemelding fra flere aktører kunne her ha gitt et annet resultat.
En rekke av lokalitetene som modellen foreslo som egnet for potensiell økning i biomasse, for å redusere smittepress av lakselus, lå i indre områder av Hardangerfjorden med relativt lav saltholdighet. Da lakselus unngår vann med lav saltholdighet, kan dette ha vært en medvirkende årsak til at de indre lokalitetene ble vurdert som potensielle for økning. Det bør bemerkes at lokalitetens bæreevne med tanke på organisk belastning er ikke vurdert her, og må analyseres nærmere.
På grunn av ordlyden i bestillingen som lå til grunn for dette arbeidet var analysene her målrettet mot å vurdere og redusere det totale smittepresset innad i PO3. Næringen uttrykte ønske om å vurdere nye driftsmønstre (eks. vekselvise utsett og brakklegging). Siden dette ikke var spesifisert i bestillingen har vi ikke gått videre med slike analyser, men vi anser nettverksanalysene som er brukt her som svært egnede for slike analyser.
Innføring av lukkede merder på slakterier vil ha en effekt, i likhet med lukking/flytting av matfisk-lokaliteter. I analyser gjennomført med HIs spredningsmodell var effekten av lukkede merder ved slakteri begrenset (kun 8%). Det bør bemerkes at vurderingen er høyst usikker da man ikke har tallfestet hvor mange lakselus slakte-lokalitetene slipper ut.
Vurderingen av de nye foreslåtte lokalitetene viste at 9 av 10 var lokalisert slik at den i stor grad vil øke smittenettverket mellom de allerede eksisterende lokalitetene. Det bør imidlertid bemerkes at dersom de nye lokalitetene skulle erstatte de gamle lokalitetene, og ikke komme i tillegg, ville dette potensielt endre vurderingen av de foreslåtte lokalitetenes egnethet.
Siden analysen av smittenettverk i HI sin modell baserer seg på vannbåren smitte, er det stor grad av samsvar i de vanligste smitterutene for lakselus, PD og ILA. Da det er antatt at virus må opptre i en viss dose for å utgjøre et smittetrykk, ble de beregnede smitteklyngene noe mindre for virus enn for lakselus, altså at nettverket for virus ble mer fragmentert. Et fragmentert nettverk anses som et ønskelig ved en eventuell omstrukturering da det reduserer spredningen av den potensielle smitten. De foreslåtte endringene i produksjonen basert på lakselus reduserte også den potensielle smitten av virus. Da scenarioene hovedsaklig var designet for å redusere lusesmitte, var reduksjonen for virus var svakere enn for lakselus.
En annen vesentlig forskjell i smitte av lakselus og virus er at lakselus driver med strømmen i flere dager før den er moden til å sette seg på en vertsfisk, mens virus er smittsomt fra det øyeblikk det skilles ut. Dette gjør egensmitte, smitte til egen lokalitet, et større problem for virussmitte enn for lus. Dersom en lokalitet med stor MTB får et virusutbrudd, vil utslakting gi et større tap enn om MTB var spredd over flere lokaliteter (Pettersen m.fl., 2015). Dette hensyn må balanseres opp mot det generelle budskapet i rapporten, at utvalgte færre, men større lokaliteter reduserer det generelle smittepresset. VIs foreløpige analyser av smittespredning for PD viser imidlertid at totalt antall fisk i lokaliteter infisert med PD blir mindre ved større og færre lokaliteter. Videre arbeid med sistnevnte analyser vil kunne belyse denne problemstillingen.
I analysene er det ikke vurdert hvorvidt et anlegg klarer å avluse like effektivt når vi øker MTB. Det er i HI sin modell lagt til grunn at anlegg som tidligere har holdt lave lusetall vil være i stand til å opprettholde de gode resultatene dersom lokalitetene får økt MTB. Det er ikke sikkert at dette vil være tilfelle for alle lokaliteter ved en faktisk økning. I mangel på bedre kunnskap er analysene gjennomført basert på antagelsen at lusetallene skaleres lineært med den økte tillatte MTB.
I analysene ble det synlig at det er stor ulikhet mellom lokaliteter både i hvor mange lakselus som blir produsert på hver lokalitet, men også i hvor stor grad lokalitetene påvirker hverandre innad i produksjonsområdet på grunn av sirkulasjonen i området. Mellomårlig variabilitet er også vesentlig.
Det er derfor lite trolig at den beregnede reduksjonen i smitte i våre analyser vil være gyldig hele tiden, men det er antatt at med tilsvarende innsats fra aktørene og effekt av tiltak som tidligere, vil smitten mellom anleggene over tid reduseres. Dersom påvirkningen var lik mellom lokalitetene, hadde tiltak på tilfeldige lokaliteter hatt like stor effekt uavhengig av hvor de ble innført. Nettopp den store variasjonen i hvordan lokalitetene påvirker hverandre, gjør at det er gevinst i strategisk og målrettet innsats hvor det vil ha størst effekt.
6 - Veien videre
Det ble fra næringen side etterspurt analyser som ikke er gjennomført i denne rapporten. Et ønske var en analyse av effekten på villfisk av å holde en maksimumsgrense på 0,2 eller 0,1 kjønnsmodne hunnlus over lengre tidsrom i året. Analyser av dette er igangsatt på HI, og vil leveres i egen rapport.
Bestillingen som lå til grunn for arbeidet etterspurte spesifikt en analyse av smittenettverket i PO3. Dette er et produksjonsområde med høy oppdrettsintensitet og mange involverte aktører. Metodene som er benyttet her for å undersøke effekten av relokalisering/omfordeling av MTB på smittenettverk, er generiske og kan relativt lett overføres til andre områder. Design av områdestruktur er enklere å implementere under utviklingen av produksjonsområder som enda ikke har like stor produksjon, f.eks. produksjonsområder lenger nord eller til havs.
Det er vår vurdering at en videre utredning om hvorledes produksjonen i PO3 bør struktureres bør inkludere analyse av bentisk bæreevne ved lokaliteter foreslått for økt MTB. Videre bør det også vurderes om den endrede strukturen vil gi økt påvirkning på de ville laksefiskene i området.
For å sikre at en anbefalt restrukturering kan gjennomføres i praksis, bør en involvere næringsaktører, samt nasjonale, regionale og lokale myndigheter.
Både HI og VI er av den oppfatning at det ligger et stort potensial for å redusere smitte mellom lokaliteter ved å restrukturere produksjonen. Dette er mest tydelig for lakselus, men vil trolig også ha betydning for smitte av PD og ILA. Tross det store potensialet kan vi ikke på grunnlag av denne studien gi en klar anbefaling om hvordan restruktureringen skal gjennomføres, men en anbefaling om å fortsette det påbegynte arbeid med noe utvidede rammer.
7 - Referanser
Albretsen J, Sperrevik AK, Sandvik AD, Asplin L (2011) NorKyst-800 Report No. 1 User Manual and technical descriptions. Fisk og Havet 2:1–48
Aldrin M, Huseby RB, Jansen PA (2015) Space–time modelling of the spread of pancreas disease (PD) within and between Norwegian marine salmonid farms. Preventive Veterinary Medicine, 121, 132-141
Aldrin M, Jansen PA, Stryhn (2019) A partly stage-structured model for the abundance of salmon lice in salmonid farms. Epidemics, 26, 9-22
Graham DA et al. (2013) Biophysical properties of the samonid alphaviruses: influence of temperature and pH on virus survival. Journal of Fish Diseases, 2007, 30, 553–543.
Gregory, A., et al. (2009), An experimental investigation on aspects of infectious salmon anaemia virus (ISAV) infection dynamics in seawater Atlantic salmon, Salmo salar L., Journal of Fish Diseases, 32, 481–489.
Haredasht SA, Tavornpanich S, Jansen MD, Lyngstad TM, Yatabe T, Brun E, Martínez-López B. (2019) A stochastic network-based model to simulate the spread of pancreas disease (PD) in the Norwegian salmon industry based on the observed vessel movements and seaway distance between marine farms. Preventive Veterinary Medicine, 167, 174-181.
Heuch, P. A., Parsons, A., & Boxaspen, K. (1995). Diel vertical migration: a possible host-finding mechanism in salmon louse (Lepeophtheirus salmonis) copepodids?. Canadian Journal of Fisheries and Aquatic Sciences, 52(4), 681-689.
Huse, I., & Holm, J. C. (1993). Vertical distribution of Atlantic salmon (Salmo salar) as a function of illumination. Journal of Fish Biology, 43, 147-156.
Jarungsriapisit, J., et al (2016). Relationship between viral dose and outcome of infection in Atlantic salmon, Salmo salar L., post-smolts bath-challenged with salmonid alphavirus subtype 3. Veterinary Res. 47:102
Jones, S. R. (2009). Controlling salmon lice on farmed salmon and implications for wild salmon. CAB Rev Perspect Ag Vet Sci Nutr Nat Res, 4, 1-13.
McGurk, M. D. (1986). Natural mortality of marine pelagic fish eggs and larvae: role of spatial patchiness. Marine ecology progress series, 34(3), 227-242
Myksvoll MS, Sandvik AD, Albretsen J, Asplin L, Johnsen IA, Karlsen Ø, Kristensen NM, Melsom A, Skardhamar J, Ådlandsvik B (2018) Evaluation of a national operational salmon lice monitoring system—From physics to fish. PLoS One 13:1–25
Pettersen, J.M., Rich, K.M., Jensen, B.B., Aunsmo, A., 2015. The economic benefitsof disease triggered early harvest: a case study of pancreas disease in farmedAtlantic salmon from Norway. Prev. Vet. Med. 121, 314–324.
Reynolds P (2014) Short-term exposure tests to elucidate handling effects. The use of freshwater to control infestations of the sea louse Lepeophtheirus salmonis on Atlantic salmon Salmo salar https://www.fhf.no/prosjekter/prosjektbasen/901006/
Reynolds P (2015) Elucidation of the effects of physical handling in removing attached sea lice from infested Atlantic salmon. GIFAS, FHF prosjekt 901006. 22 s. https://www.fhf.no/prosjekter/prosjektbasen/901006/
Rosvall M, Bergstrom CT (2008) Maps of random walks on complex networks reveal community structure. Proc Natl Acad Sci U S A 105:1118–23
Sandvik AD, Johnsen IA, Myksvoll MS, Sævik N, Skogen MD (2020) Prediction of the salmon lice infestation pressure in a Norwegian fjord.
Skern R, Sissener NH, Sandvik AD, Meier S, Sævik PN, Skogen MD, Vågseth T, Dalvin S, Skern-Mauritzen M, Bui S (til fagfellevurdering) Parasite development affect dispersal dynamics; infectivity, activity and energetic status in cohorts of salmon louse copepodids. Journal of Experimental Marine Biology and Ecology (JEMBE)
Stien A, Bjørn PA, Heuch PA, Elston DA (2005) Population dynamics of salmon lice Lepeophtheirus salmonis on Atlantic salmon and sea trout. Mar Ecol Prog Ser 290:263–275
Tapia, E et al. (2013) Assessment of the in vitro survival of the Infectious Salmon Anaemia Virus (ISAV) under different water types and temperature. Bulletin of the European Association of Fish Pathologists, 33, 3–12.
Ådlandsvik, B. (2020), LADiM documentation, https://ladim.readthedocs.io/
8 - Appendiks 1
8.1 - Beskrivelse av modellområdet
Både lakselus og virus vil transporteres mellom lokaliteter med vannet. Strømforholdene i de øvre vannmassene, hvor lakselus oppholder seg, er variable både i tid og rom. Langs kysten domineres strømmen av den nordgående kyststrømmen, men har betydelig variabilitet i form av virvler og påvirkning av vind. Drivkreftene som skaper strøm er flere, hvor både tidevann, vind, indre bølger og ferskvannsavrenning fra land påvirker vannets evne til å transportere lus med strømmen. Tidevann er den mest regelmessige drivkraften, men da den snur retning ca. hver 6. time, er den totale transporten grunnet tidevann begrenset, men det bidrar til økt spredning av smitte. Vind gir et vesentlig moment til vannet i de vertikale lag som lus oppholder seg. I de ytre delene av produksjonsområdet er det relativt åpne områder, hvor vinden kan ha alle retninger. Lenger inne i fjordene vil vinden hovedsakelig være rettet langs fjorden og gi vannet moment ut eller inn fjorden. Sterk strøm inn og ut fjorden blir generert av tetthetsforskjeller mellom vannet inne i fjordene og vannet ute på kysten. Disse strømmene inntreffer med ujevne mellomrom, ca 2 ganger i måneden, og transporterer alt passivt materiale effektivt over store avstander. PO3 har en betydelig avrenning av ferskvann fra land. Hovedavrenningen er konsentrert rundt vårsmeltingen, men Folgefonna vil ha en senere topp og bidra utover sommeren. Ferskvannsavrenningen skaper et brakkvannslag i de indre delene av området. Dette brakkvannslaget påvirker spredningen av lakselus både fordi lakselus unngår vann med lav saltholdighet, og plasserer seg dypere i område med lav saltholdighet nær overflaten, men også indirekte, både lakselus og virus, da en vertikal tetthetsprofil påvirker hvor dypt vinden klarer å dra med seg vannet.
8.2 - Estimert luseutslipp i HIs modeller
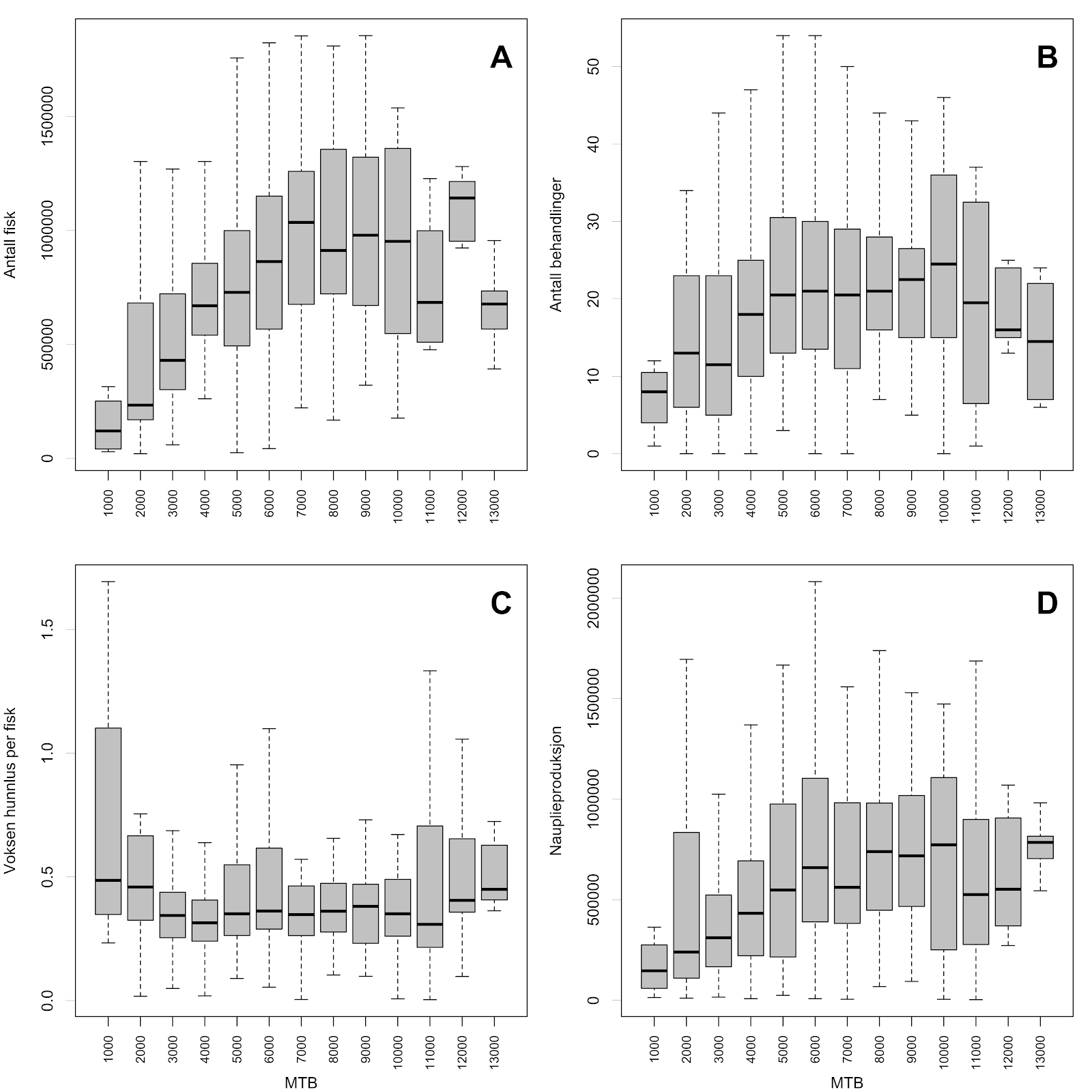
For å oppnå en reduksjon i antall matfisk-lokaliteter uten å redusere produksjons-taket i produksjonsområdet må nødvendigvis MTB økes på enkelte lokaliteter. Per i dag foreligger det ingen offisielle tall på en øvre grense på total tillatt MTB som i praksis lar seg drive med tilfredsstillende lusetall. Hvis vi ser på aggregerte lusetall fra produksjonsområder 1-7 viser tallene at antall fisk som til enhver tid står på lokalitetene varierer stort mellom lokalitetene, men samsvarer til en viss grad med MTB på lokalitetene opp til omkring 7000 tonn (Figur A1 A). Antall tiltak for redusering av lus ser også ut til å følge dette mønsteret (Figur A1 B), samtidig som gjennomsnittlig antall lus per fisk ser ut til å holde seg stabil selv om MTB øker på lokaliteten (Figur A1 C). Til syvende og sist er det antall nyklekte lakslus (nauplier) produsert på lokaliteten som avgjør den potensielle smitten til andre lokaliteter. Her gjenspeiles det samme mønsteret med økning i nauplieproduksjon opp til en MTB på 7000-8000 tonn, for deretter å flate ut ved de antatt største lokalitetene (Figur A1 D). Basert på innrapporterte lusetall kan det derfor synes som at antallet lakselus som blir produsert ved lokalitetene øker i takt med MTB opp til en produksjon på 8000 tonn. Det er også antatt at mengden viruspartikler som blir sluppet fra fisk ved et eventuelt utbrudd er skalert med biomassen. Det å anta økt utslipp av både lakselus og virus med økt biomasse synes derfor å være en rimelig antagelse.
Aggregert ukentlig statistikk fra lokaliteter i produksjonsområder 1-7, sortert etter maksimal tillatt biomasse på lokaliteten (MTB, rundet til nærmeste 1000 tonn) fra perioden 2018-2019, plottet mot: (A) antall fisk per lokalitet, (B) antall rapporterte behandlinger per lokalitet, (C) gjennomsnittlig antall voksne hunn-lus per fisk per lokalitet, og (D) estimert nauplieproduksjon per lokalitet (se seksjon 3.1). Her representerer sort strek medianen per klasse, grå boks interkvartil avstand (i.e. intervallet fra 25. til 75. persentil), og stiplet linje 1.5 ganger interkvartil avstand.
Basert på ukentlig innrapporterte antall kjønnsmoden hunn-lus per lokalitet modellerte vi nauplieproduksjon på lokalitetene som funksjon av antall fisk per lokalitet, gjennomsnittlig antall lus per fisk og observert temperatur på lokaliteten (Stien m.fl., 2005). Deretter delte vi opp hver av de lokalitetsspesifikke tidsseriene for nauplieproduksjon inn i produksjonssykluser. Her lagde vi en algoritme som delte opp serien basert på antall rapporterte fisk på lokaliteten, der hver produksjonssyklus ble startet ved maksimum antall fisk på lokaliteten (i.e. når all fisk var satt ut på lokaliteten), og avsluttet når bare 20% av antallet fisk var igjen på lokaliteten (i.e. frem til en nedslakting på 80%). Til sammen identifiserte vi 525 slike produksjonssykluser, fordelt på 135 lokaliteter som har rapportert lusetall i perioden 2012-2019.
Gjennomsnittlig varighet av hver produksjonssyklus var 428 dager (standardavvik på 133 dager), og hver lokalitet hadde alt fra én til åtte produksjonssykluser i perioden. Deretter gjorde vi en «trepunktsvurdering» av nauplieproduksjonen på alle lokalitetene. Her regnet vi ut gjennomsnittlig nauplieproduksjon for hver av disse produksjonssyklusene, og sorterte produksjonssyklusene for hver lokalitet etter hvilke syklus som hadde (1) lavest gjennomsnittlig nauplieproduksjon, (2) høyest gjennomsnittlig nauplieproduksjon, og (3) medianen over alle gjennomsnittene av produksjonssyklusene. I trepunktsvurderingen ble de tre momentene (lavest, høyest og median nauplieproduksjon) vektet likt. Denne trepunktsvurderingen av nauplieproduksjonen på hver lokalitet ble så brukt som utslippsscenarioer i spredningsmodellen i de videre analysene (Figur A2).
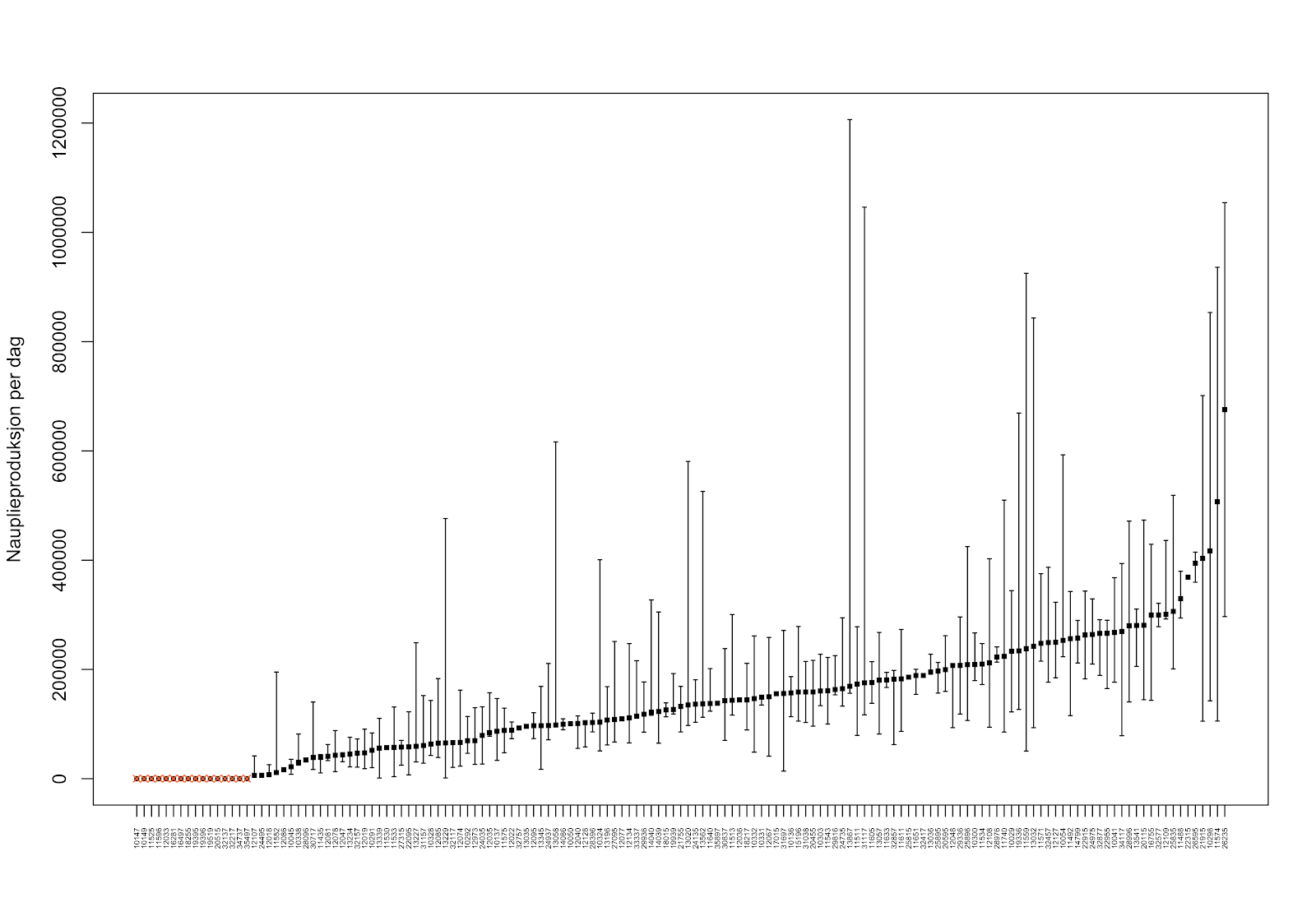
Figur A2: “Trepunktsvurdering” av nauplieproduksjon per lokalitet, basert på rapporterte lusetall i perioden 2012-2019. Her er det tatt utgangspunkt i gjennomsnittet av produksjonssyklusen som har hatt (1) den laveste nauplieproduksjon, (2) den høyeste nauplieproduksjon, og (3) median av gjennomsnittene av alle produksjonssyklusene. Lokaliteter med oransje kryss har ikke rapportert lusetall og er derfor ekskludert fra analysen.
8.3 - Parameterisering av virus i HIs spredningsmodell
Den sparsomme litteraturen på dødelighet av virus bruker halveringstid (tiden det tar før halvparten av viruset er blitt inaktivt) som mål. Hovedprinsippet er at dødeligheten øker med økende temperatur, mens andre miljøfaktorer spiller mindre rolle. Resultatene er oppsummert i Figur A3 under. For PD (blått) er verdiene hentet fra Graham m.fl., 2007. Alle data for saltvann med organisk materiale er brukt. Dataverdiene spriker stort, noe som tilsier at det er stor usikkerhet i dødeligheten. Sammenhengen er lineær, det er ikke noe å hente på et kvadratisk ledd i regresjonen. Regresjonslinjen er gitt ved:
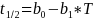
der 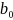 = 8.33 ,
= 8.33 , 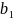 = -0.29 og
= -0.29 og 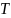 er temperaturen.
er temperaturen.
For ILA (ISAV) er datagrunnlaget enda spinklere. De grønne punktene er tatt fra Tapia m.fl., 2013. De viser relativ konstant dødelighet for lave temperaturer og deretter raskt økende. Disse observasjonene tyder på at dødeligheten for ILA virus er lavere enn for PD. Studiene som ligger til grunn, er imidlertid omstridt og gir et utilstrekkelig grunnlag for å konkludere generelt at ILA virus overlever lengre enn PD. I mangel på bedre informasjon tilpasser denne modellstudien de sparsomme observasjonene med en kvadratisk funksjon:
Der = 7,35, = 0,55 og 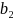 = -0,033.
= -0,033.
Modellsystemet implementerer dødeligheten som en reduksjon i smittepress hver time. Dette beregnes som 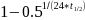 , der halveringstid er gitt i døgn. Dette er vist i Figur A4 under. Til sammenligning brukes dødelighet for lakselus som 17% per døgn, dette svarer til en halveringstid på 3,72 døgn og en dødelighet på 0,77% per time. Dette gjør at levetiden for virus er sammenlignbart og til dels lenger enn for lakselus.
, der halveringstid er gitt i døgn. Dette er vist i Figur A4 under. Til sammenligning brukes dødelighet for lakselus som 17% per døgn, dette svarer til en halveringstid på 3,72 døgn og en dødelighet på 0,77% per time. Dette gjør at levetiden for virus er sammenlignbart og til dels lenger enn for lakselus.
Erfaringsmessig regnes det likevel at lakselus er det patogen som smitter over størst avstand. Dette fordi én lakselus er nok til å infisere en fisk mens virus trenger en viss minste konsentrasjon for å være infektivt. Dette er implementert ved en terskelverdi for smittedose. Smittedosen beregnes som sum over 3x3 gridceller omkring mål-lokaliteten, et kvadrat med sidekant 480 meter. For PD, må smittedosen overstige 0,01% av nivået i kilde-kvadratet for å telle med. For ILA er denne terskelverdien satt til 1%. Disse verdiene er usikre, men litteraturen oppgir en betydelig høyere minste infeksiøse dose for ILA (Jarungsriapisit m.fl., 2016) enn for PD (Gregory m.fl., 2009).
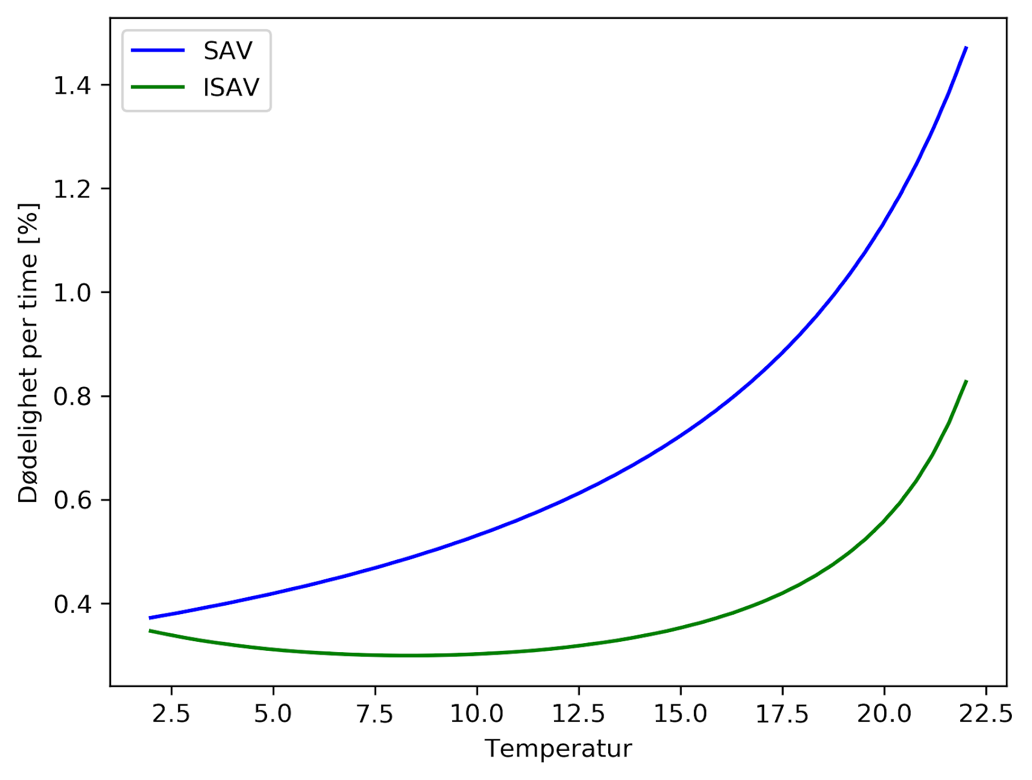
9 - Appendiks 2
9.1 - Parameterisering av virus i VIs spredningsmodell


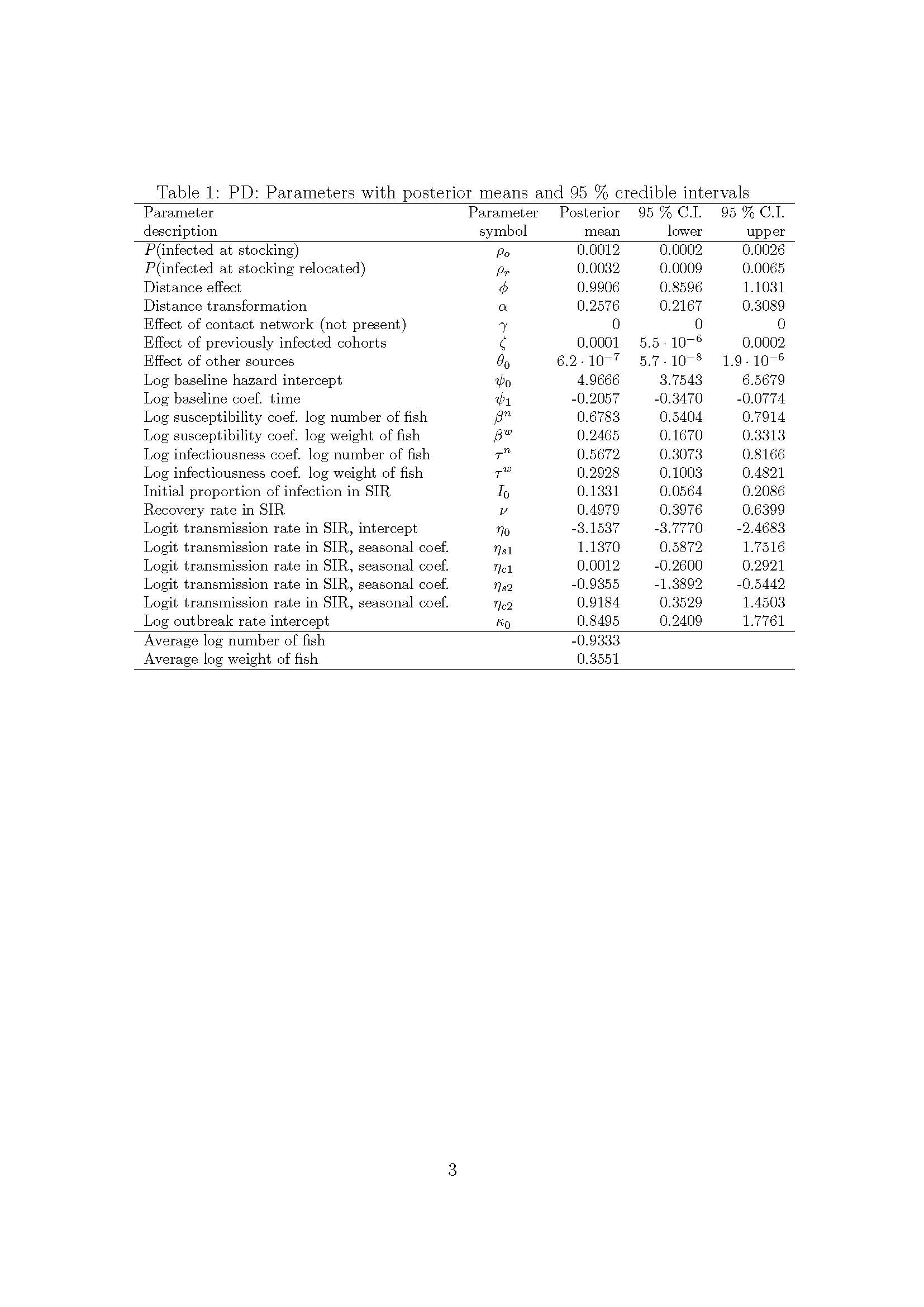
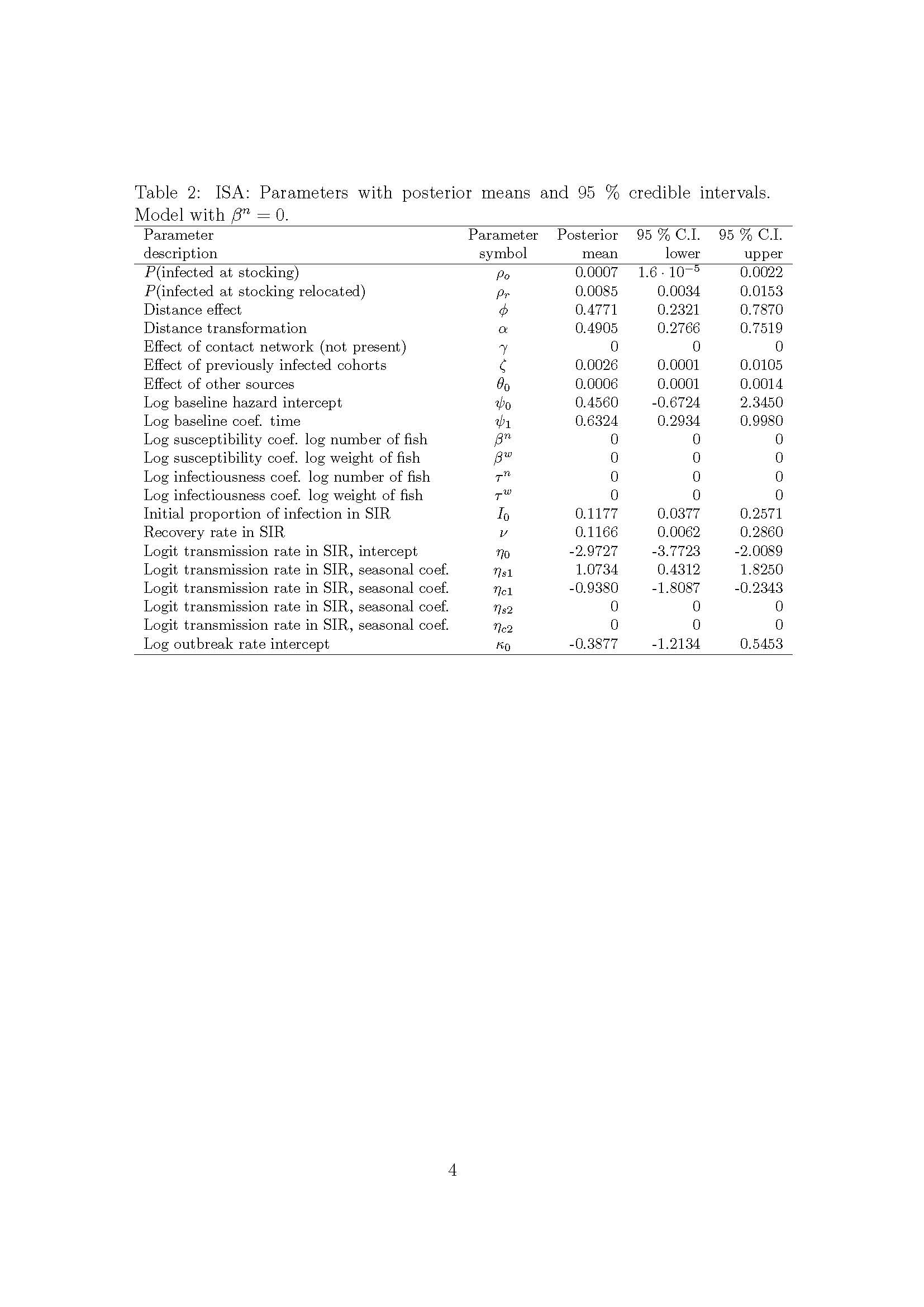
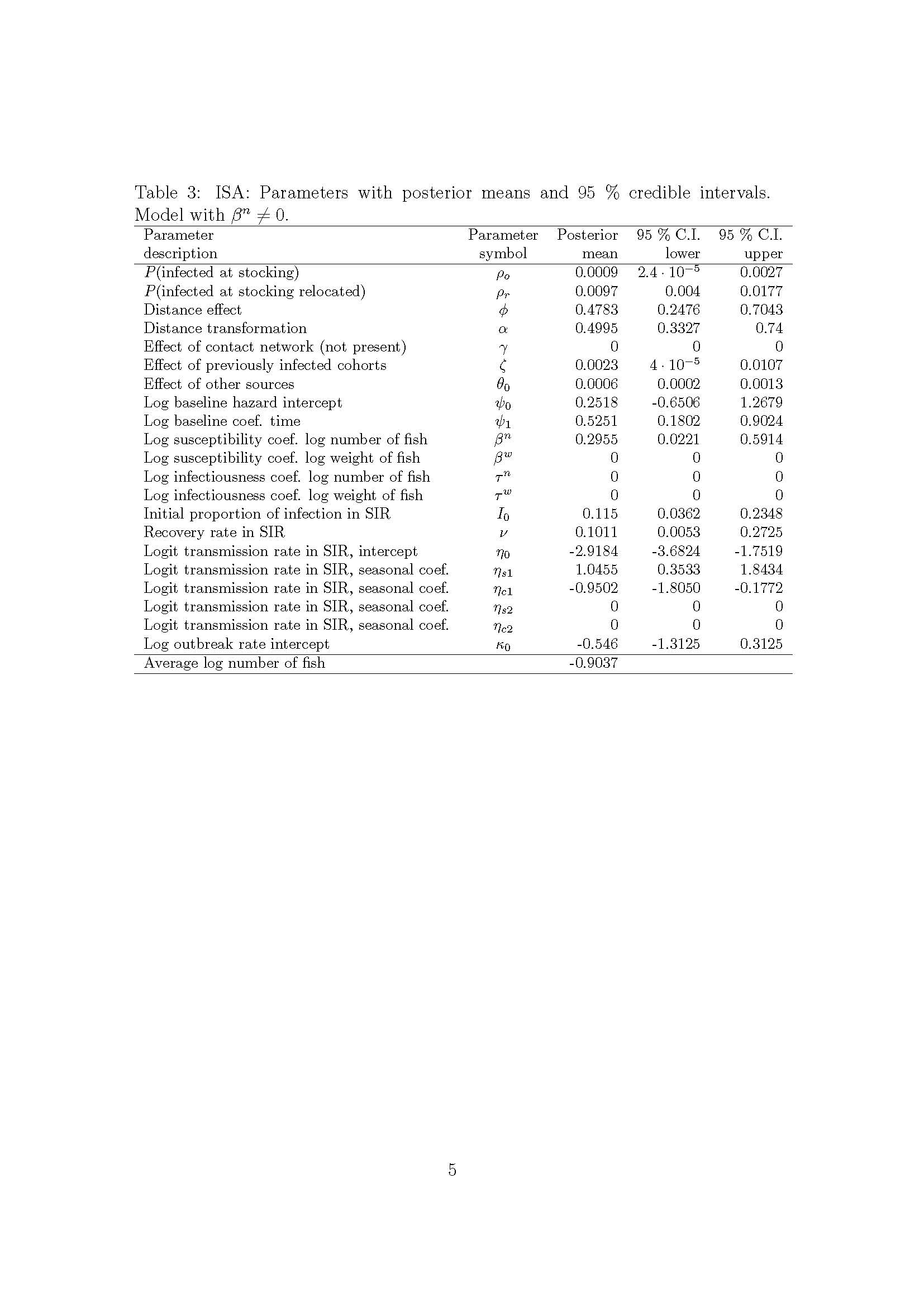

HAVFORSKNINGSINSTITUTTET
Postboks 1870 Nordnes
5817 Bergen
Tlf: 55 23 85 00
E-post: post@hi.no
www.hi.no