The Institute of Marine Research have monitored the kelp vegetation after kelp (Laminaria hyperborea) harvesting trials in southern parts (65° 20’ N - 65° 33’ N) of Nordland county in Norway in 2013-2019. Survey stations included both harvested areas and unharvested areas (reference areas) and was performed by underwater video. In addition, kelp plants were collected for studies of plant morphology, age, growth history, epiphytes and juvenile sea urchins. Observations in previously harvested areas revealed that although kelp rapidly re-monopolizes the substratum and kelp biomass is restored within four years, the kelp epiphytes and recruit vegetation require a fallow period of six years to reach pre-harvesting levels. The abundance of sea urchins was generally low at most stations observed in Nordland in 2019 and grazing from sea urchins appear to have little effect on the recovery process after harvesting. A survey made further north in Nordland (between Herøy (66° 02’ N) and Bodø (67° 22’ N) municipalities) in 2019 showed that L. hyperborea was most abundant in the outer, western parts of the region. Density of juvenile green sea urchins (Strongylocentrotus droebachiensis) in the hapteron of collected kelp plants was higher in the northern parts of the surveyed area, indicating that this species recruitment potential increases towards the north. Growth zone analyses of kelp plants collected in pristine areas revealed that the annual stipe growth was slower in the northern parts of the surveyed area (Herøy – Bodø) than further south.
Tareundersøkelser i Nordland i 2019
Rapportserie:
Rapport fra havforskningen 2020-9
ISSN: 1893-4536
Publisert: 25.03.2020
Prosjektnr: 14914-12
Forskningsgruppe(r):
Bunnsamfunn
Tema:
Stortare
Program:
Kystøkosystemer
Godkjent av:
Forskningsdirektør(er):
Geir Huse
Programleder(e):
Jan Atle Knutsen
English summary
Sammendrag
Havforskningsinstituttet har i perioden 2013-2019 gjennomført undersøkelser i forbindelse med prøvehøsting av stortare (Laminaria hyperborea) på kyststrekningen Sømna (65° 20’ N) – Vega (65° 33’ N) i Nordland. Det ble i 2019 gjennomført oppfølgende undersøkelser av taresamfunnene på felt som ble prøvehøstet i 2013, 2014 og 2017, samt i nærliggende områder som ikke har vært tarehøstet tidligere (referanseområder). I tillegg ble det gjennomført kartlegging av tarevegetasjonen og kråkebolleforekomster i utvalgte breddegradssektorer på kyststrekningen fra Herøy (66° 02’ N) til Bodø (67° 22’ N). Tarevegetasjonen ble undersøkt ved hjelp av undervannsvideo og innsamlinger av tareplanter for studier av morfologi, alder, veksthistorikk, epifytter og kråkebollerekrutter. Undersøkelsene i de tidligere prøvehøstede områdene viser at selv om høstefeltene raskt rekoloniseres av små stortareplanter og tarebiomassen restitueres i løpet av fire år, kreves en hvileperiode på seks år for reetablering av epifyttstruktur og rekruttvegetasjon. Videoobservasjonene viser at tettheten av kråkebolle var lav på de fleste stasjoner undersøkt i Nordland i 2019, og kråkebollebeiting har hatt liten effekt på tarevegetasjonens reetablering i de tidligere prøvehøstede områdene. På kyststrekningen Herøy - Bodø er stortarevegetasjonen best utviklet i ytre, vestlige deler av de undersøkte sektorene. Tettheten av juvenile grønne kråkeboller (Strongylocentrotus droebachiensis) observert i de innsamlede tareplantenes festeorgan (hapter) økte fra sør til nord. Veksthistorikkanalyser av innsamlede tareplanter indikerer at den årlige stilktilveksten er langsommere på kyststrekningen Herøy – Bodø, enn i sørligere områder.
1 - Innledning
Det er i de senere år observert tilbakegang av grønn kråkebolle (Strongylocentrotus droebachiensis) og gjenvekst av stortare (Laminaria hyperborea) i sørlige deler av Nordland (Norderhaug & Christie 2009, Fagerli et al. 2013, 2015). Med tilbakegang for kråkeboller reetableres tareskoger som representerer nye ressurser etter flere tiår med beiting. For å utrede ressursgrunnlaget for tarehøsting i dette området har tarenæringen i perioden 2013-2017 gjennomført prøvehøstinger av stortare på kyststrekningen fra grensen mot Trøndelag til sørsiden av Vega i Nordland med bruk av tindetrål.
I forkant av prøvehøstingene har Havforskningsinstituttet gjennomført kartlegginger av de omsøkte høstefeltene for å innhente grunnlagsdata på naturtilstanden og dokumentere feltenes egnethet for høsting mhp forekomst av stortare og kråkeboller. Det er videre gjennomført årlige etterundersøkelser i et utvalg av de tidligere prøvehøstede områdene, samt i nærliggende områder som ikke har vært tarehøstet (referanseområder), for å dokumentere taresamfunnenes reetablering. Resultater fra tidligere undersøkelser gjennomført sør for Vega i Nordland i perioden 2013-2018, tyder på at de vestlige deler av dette kystområdet rommer betydelige tareforekomster og at beiting fra kråkeboller ikke har påvirket reetableringen av tare etter prøvehøstingene (Steen et al. 2015ab, 2016b, 2018ab). For å studere den videre utviklingen i taresamfunnene og kråkebollebestandene ble det sommeren 2019 gjennomført oppfølgende videoundersøkelser i et utvalg av de tidligere prøvehøstede områdene på kyststrekningen mellom Sømna (65° 20’ N) og Vega (65° 33’ N), i tillegg til at det ble samlet tareplanter på enkelte felt som ble prøvehøstet i 2013 og 2014, for studier av morfologi, alder, veksthistorikk og begroingsorganismer (epifytter). Tilsvarende undersøkelser ble også gjennomført i referanseområder som ikke har vært gjenstand for tarehøsting.
Undersøkelsene gjennomført i forbindelse med prøvehøstingene i Nordland (Steen et al. 2014a, 2015ab, 2016b, 2018ab) har gitt muligheter for å sammenligne tilstanden før og etter tarehøsting da en her har områder som ikke har vært utsatt for denne type påvirkning tidligere. De oppfølgende undersøkelser gjennomført i de tidligere prøvehøstede områdene vil bidra med informasjon om taresamfunnenes reetableringsevne og restitusjonsperiodens varighet i denne regionen som tidligere har vært mye preget av kråkebollebeiting (Sivertsen 1997, 2006).
Det er fra forvaltningshold og tarenæringen uttrykt ønske om å utrede tareressursgrunnlaget også i områdene nord for Vega i Nordland. Det er dessuten av generell interesse å få oppdatert kunnskap om tilstanden i taresamfunnene og forekomster av kråkeboller i indre og ytre kystområder i denne delen av Nordland. På bakgrunn av dette gjennomførte Havforskningsinstituttet sommeren 2018 en sektorvis kartlegging av tare og kråkebolleforekomster, vha video og innsamlede tareprøver, fra nordsiden av Vega til Lurøy (Steen et al. 2018b). For å undersøke tilstanden i taresamfunnene lengre nord i Nordland, ble det i 2019 gjennomført tilsvarende kartlegginger på kyststrekningen fra Herøy til Bodø.
2 - Metodikk
2.1 - Videoundersøkelser
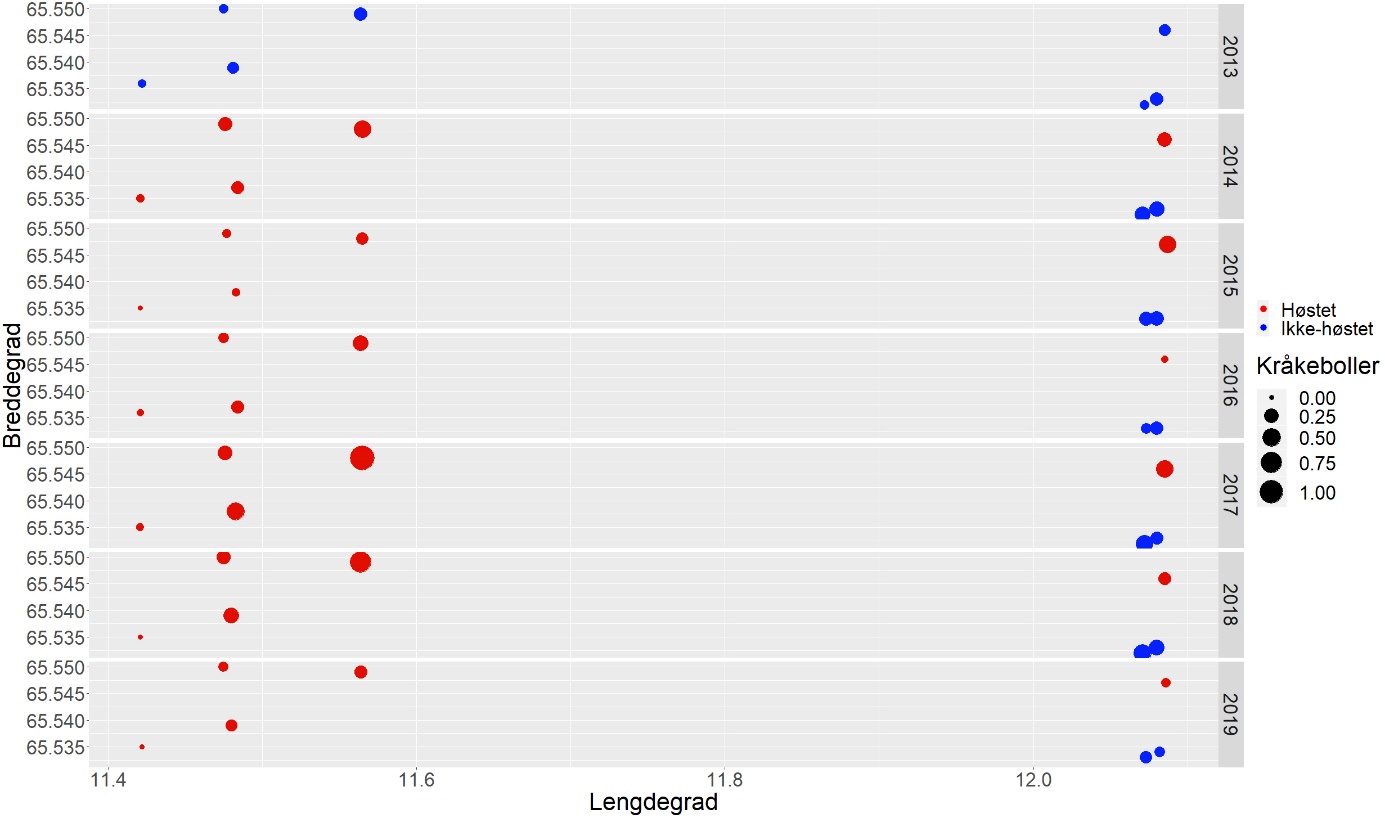
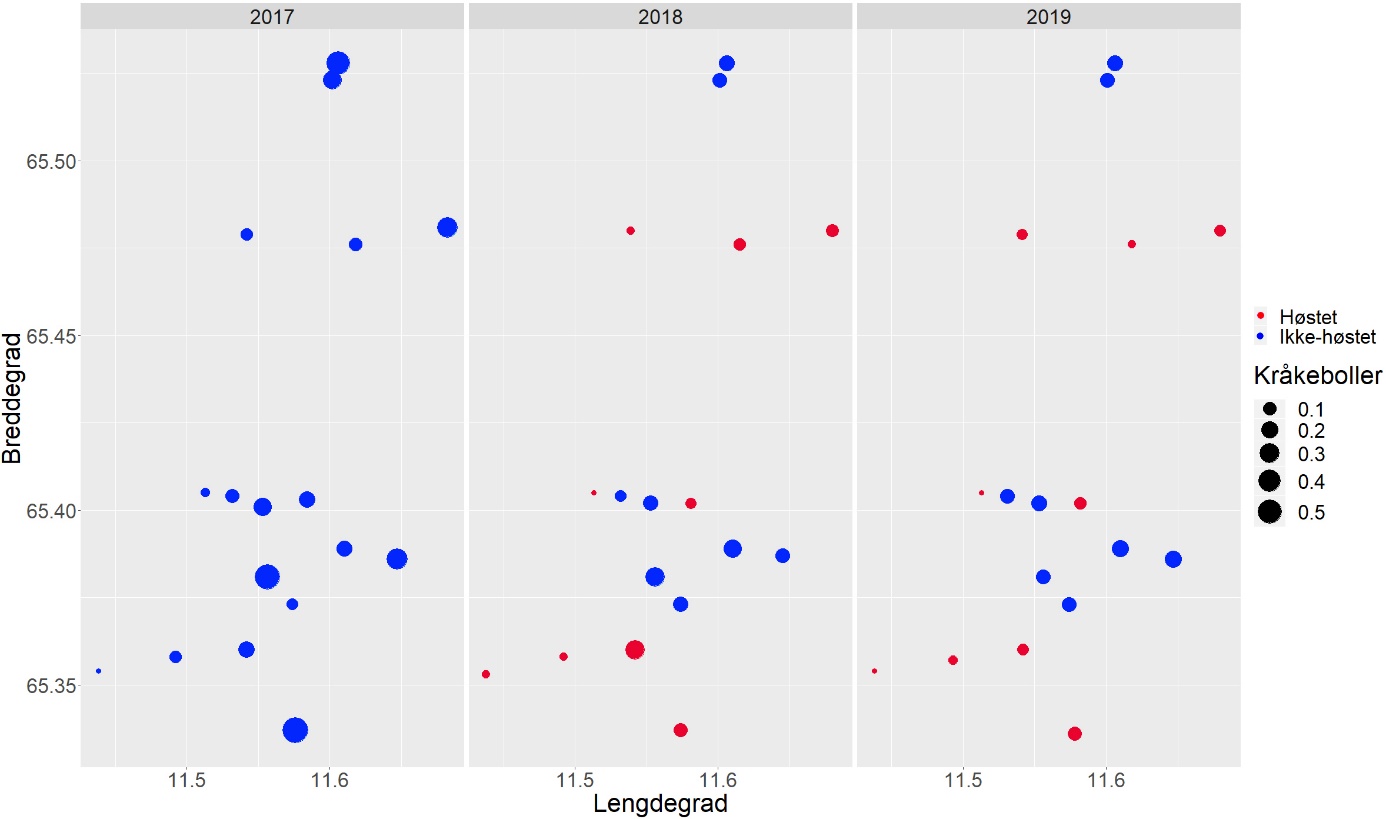
I juni 2019 ble det i området mellom Sømna (65° 20’ N) og Vega (65° 33’ N) i Nordland gjennomført oppfølgende videoundersøkelser på faste stasjoner i felt som ble prøvehøstet i 2013 (PF3), 2014 (PF5-7) og 2017 (PF19-21) og i nærliggende referanseområder som ikke har vært utsatt for tarehøsting (Fig. 1).

For å kartlegge forekomster av tare og kråkeboller lenger nord i Nordland, ble det gjennomført videoundersøkelser på utvalgte stasjoner i breddegradssektorer fra Herøy (66° 02’ N) til Bodø (67° 22’ N) i 2019 (Fig. 2). Observasjonene i de to sørligste sektorene (Herøy og Lurøy) ble sammenlignet med observasjoner gjennomført på de samme stasjonene i 2018 for å avdekke eventuelle endringer over tid. Bortsett fra de to sørligste sektorene som hadde en innbyrdes avstand på ca 20 nautiske mil (20 breddeminutter), så hadde de undersøkte sektorene på denne kyststrekningen en innbyrdes avstand på ca 10 nautiske mil (10 breddeminutter) i nord-sør retning. Stasjonene innen hver sektor ble spredt fra indre til ytre kystområder for å fange opp eventuelle geografiske variasjoner og dekke en så stor bølgeeksponeringsgradient som mulig.

Videostasjonene ble plukket på basis av ekkoloddregistreringer av dybde og bunnforhold. Det ble i juni 2019 tatt opp videofilm på 35 stasjoner på strekningen Sømna - Vega og 48 stasjoner på strekningen Herøy - Bodø. Videoobservasjonene på de enkelte stasjonene ble gjennomført med nedsenkbart undervannskamera (UVS 5080), med innebygd dybdesensor, langs faste transekter ved hjelp av forskningsfartøy (FF Fangst) med kartplotter og ekkolodd, med en gjennomsnittshastighet på ca 1 knop. For å få høyoppløselige filmopptak ble det også montert et GoPro kamera (HERO 3, Black edition) på oversiden av UVS 5080-kameraet (Fig. 3). På hvert transekt ble undervannskameraet vekselvis ført rett over og gjennom tarevegetasjonen over en distanse på ca 100-200m, der kamerapiloten justerer høyden i forhold til tarevegetasjonen og bunnen vha en monitor.

Filmopptakene ble fortløpende konvertert til PC-format (mpg) vha en Pinnacle movie box, og lagret på eksterne harddisker (dubletter). Før analyse ble videotransektene splittet opp i mindre avsnitt (for hvert minutt film) der gjennomsnittlig dyp, bunntype, tarevegetasjonens dekningsgrad (definert som andel bunnflate dekket av tarevegetasjon), kronesjikttetthet (definert som tetthet av dekkvegetasjons (eller kronesjikt) planter), kronesjikthøyde, rekruttering (definert som tettheten av undervegetasjonsplanter <25 cm) og forekomst av epifytter (begroing) på tarestilkene ble anslått. Høyden av tareplantene måles fra tareplantenes festeorgan (dvs bunnen) til tarebladet vha kameraets innebygde dybdesensor (Figur 4). Epifyttene (begroingen) på tarestilkene skaper et viktig habitat for små evertebrater som snegl og krepsdyr og som igjen er føde for fisk. Forekomst av epifytter (Fig. 5) ble rangert på en 4-trinns skala, der tarestilker uten epifytter ble gitt verdien 0, tarestilker med flekkvis forekomst av skorpeformete epifytter gitt verdien 1, tarestilker med dominans av skorpeformede epifytter gitt verdien 2, og tarestilker med dominans av tredimensjonale, voluminøse epifytter gitt verdien 3 (Steen 2019).

Det ble også gjort anslag av gjennomsnittlig biomassetetthet av stortare langs videotransektene ved å multiplisere gjennomsnittlig plantevekt med plantetthet per kvadratmeter (D). Stortareplantenes vekt ble beregnet utifra plantehøyden (dvs kronesjikthøyden) som observeres langs videotransektene ved å benytte en relasjon mellom individuell plantehøyde i meter (H) og plantevekt i kg (V) der V = 0,97H 1,57 . Den beregnede relasjonen mellom plantehøyde og plantevekt har en korrelasjonskoeffisient (R 2 ) = 0,86, og er beregnet utifra fra målinger av 1607 stortareplanter samlet inn på kyststrekningen Agder – Nordland i perioden 2010-2019. Biomassetettheten (B) kan deretter beregnes utifra formelen B = V * D i kg per kvadratmeter.

Det ble også gjennomført tellinger av kråkeboller og forekomstene ble angitt som antall registrerte kråkeboller per meter videotransekt. Det er her viktig å skille mellom ulike arter av kråkebolle, da beiteadferden vil variere fra art til art. Røde kråkeboller ( Echinus esculentus ) beiter normalt ikke tareplantene like aggressivt som grønne kråkeboller ( Strongylocentrotus droebachiensis ), men forringer habitatet ved å beite bort epifytter på tarestilkene og kan ved høye tettheter også gi beiteskader på selve tarevegetasjonen (Sjøtun et al. 2006).
For å standardisere analysene ble transektavsnitt med substrattyper uegnet for stortarevekst (mudder, sand, grus) og gjennomsnittdybder på mer enn 15 meter utelatt, da mesteparten av tarehøstingen allikevel finner sted grunnere enn 15 meters dyp (Steen 2019).
2.2 - Tareinnsamlinger
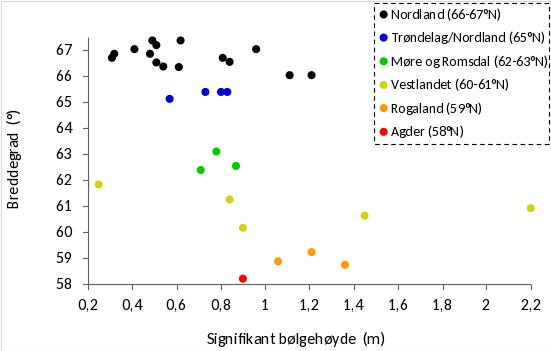
For undersøkelser av tarevegetasjonens morfologi, alder, veksthistorikk, epifyttsammensetning og forekomst av kråkebollerekrutter, ble det i juni 2019 samlet inn ca 10 tareplanter fra hver av 22 stasjoner (Fig. 1, 2) ved hjelp av tareskrape (Fig. 6). Av innsamlingsstasjonene sør av Vega er den nordligste stasjonen som ligger i felt PF3 prøvehøstet i 2013, mens de tre stasjonene sør for denne, i felt PF5, ble prøvehøstet i 2014 (Fig. 1). De tre sørligste innsamlingsstasjonene ligger innenfor Horsvær naturreservat og har ikke vært prøvehøstet tidligere (Fig. 1). Det ble også samlet inn tareplanter på stasjonene sør av Vega i forkant av prøvehøstingene i henholdsvis 2013 og 2014. De 15 stasjonene som det ble samlet inn stortare nord for Vega (strekningen Herøy – Bodø) i 2019, ligger alle i områder som ikke har vært utsatt for tarehøsting tidligere (Fig. 2). I perioden april – juni 2019 ble det også samlet inn ca 10 tareplanter fra hver av 13 referansestasjoner (i områder som er stengt for tarehøsting) på kyststrekningen Agder – Trøndelag for å bl.a undersøke geografiske variasjoner i tarevegetasjonens morfologi, alderssammensetning, veksthistorikk, samt epifytt- og kråkebolleforekomster. Dypet på innsamlingsstasjonene varierte fra 6 m til 12 m, mens midlere signifikant bølgehøyde varierte fra 0,25 m til 2,20 m.

Tarestilkenes lengde ble målt fra overgang festeorgan/stilk til overgang stilk/blad, mens tarebladenes lengde ble målt fra overgang stilk/blad til enden av blad. Begroingsorganismene (epifyttene) ble skrapt av tarestilkene og veid (våt vekt), og de dominerende arter/grupper ble rangert etter forekomst for hver enkelt tarestilk. Tareplantenes blad (lamina), stilk (stipes) og festeorgan (hapter) ble separert og veid (våt vekt) hver for seg. Forekomster av juvenile kråkeboller (rekrutter) i tareplantenes festeorgan ble registrert.
Tarestilkenes diameter ble målt i tverrsnitt tatt ca 1 cm over festeorganet, der stilkene er på sitt tykkeste. Alderen på de innsamlede tareplantene ble bestemt ved å telle vekstsoner i tverrsnittene og samtlige tverrsnitt ble avfotografert for senere målinger av vekstsonearealene og rekonstruksjon av plantenes veksthistorikk. Størrelsen (arealet) av hver enkelt vekstsone vil gi informasjon om tareplantenes vekst i et gitt år, der den ytre ringen representerer veksten i 2019, den nest ytterste ringen veksten i 2018, etc, etc (Fig. 7). Arealet på en vekstsone (a) ble estimert ved å måle bredden fra tverrsnittets sentrum til indre (i) og ytre (y) vektsonegrense langs to akser med ca 90 graders vinkel og beregnet utifra formelen a = π y 2 – π i 2 .

2.3 - Modellering av bølgeeksponering
Bølgeeksponering er en faktor som kan ha betydning for utbredelse og vekst av stortare og dens begroingsorganismer (Sjøtun & Fredriksen 1995, Sjøtun et al. 1998, Norderhaug & Christie 2011, Norderhaug et al. 2012, Pedersen et al. 2012, Bekkby et al. 2014ab, Smale et al. 2016, Steen et al. 2014, 2015ab, 2016b, 2018ab). På samtlige undersøkte stasjoner ble det derfor gjort modellberegninger av bølgeeksponering for å undersøke om i hvilken grad denne faktoren påvirker de ulike målvariablene. Modellen estimerte midlere signifikant bølgehøyde som et mål på eksponering. Bølgehøyden var representert som et statistisk langtidsmiddel. Midlere signifikant bølgehøyde er per definisjon lik gjennomsnittsverdien av den høyeste tredjedelen av alle bølger innen et gitt tidsintervall. Strøklengde ble beregnet, og vindobservasjoner og modellerte bølgehøyder til havs (kilde: Meteorologisk institutt) ble brukt for å beregne et endelig mål på bølgehøyden for en gitt lokalitet eller et geografisk område. Modellen som bygger på formelverket til Norsk Standard 9415 (2009), har innebygd fire ulike empiriske formelsett for beregning av signifikant bølgehøyde basert på vindmålinger, og alle formuleringene gir sammenliknbare resultater.
2.4 - Statistisk behandling
Resultatene er presentert som aritmetiske gjennomsnitt ± 95 % konfidensintervall, hvis ikke annet er oppgitt. For å statistisk teste effekter av tarehøsting ble det benyttet lineære sammensatte modeller («linear mixed-effect models») tilpasset med “lmer” funksjonen i “Lme4” pakken i R (Bates et al. 2015). Modellen ble repetert for ulike responsvariable som tarevegetasjonsdekning, -høyde, -tetthet, -rekruttering, -biomasse og kråkebolleforekomst for videodataene og alder, stilklengde, stilkdiameter, epifyttvekt, epifytt-tetthet og forekomster av hapterkråkeboller for innsamlede tareplanter. Felttype (høstefelt vs referansefelt) krysset med periode (før høsting vs etter høsting) som faste («fixed») faktorer, og stasjon nestet i felt som tilfeldig («random») faktor. Fordeling og varianshomogenitet av residualer ble kontrollert før kjøring av testene. Effekter av tarehøsting på de ulike responsvariablene observeres gjennom styrken på interaksjonen mellom de to faste faktorene, felttype og periode. For å teste forskjeller mellom to og to faktornivåer ble det benyttet parvise t-tester. Korrelasjoner mellom variable ble estimert vha Pearsons produkt-moment korrelasjonskoeffisient (r). T-testene og korrelasjonstestene ble programmert i R.
3 - Resultater
3.1 - Effekter av prøvehøstingen i 2013
3.1.1 - Videoundersøkelser
Reetableringen etter prøvehøstingen i 2013 har generelt vært god og gjennomsnittlig dekningsgrad av stortare har vært stabil på rundt 95 % gjennom de fire siste årene. Stortarevegetasjonens høyde, tetthet, biomasse og epifyttforekomster var i 2019 ikke signifikant forskjellig fra det som ble observert før prøvehøstingen seks år tidligere. Kråkebollebeiting har ikke påvirket reetableringen av tare etter prøvehøstingen i Nordland, og forekomstene av rød kråkebolle i 2019 var lavere enn i de to foregående år. Grønne kråkeboller er ikke observert i perioden 2013-2019.
Totalt 5.576 tonn stortare ble prøvehøstet på et felt (PF3) i Nordland i juli 2013. Fem videostasjoner i prøvehøstefeltet og to videostasjoner på et referansefelt har blitt undersøkt årlig fra før høstingen i 2013, for å studere gjenvekst av tarevegetasjon og utvikling i kråkebollebestandene. Videostasjonene dekker en stor eksponeringsgradient med variasjoner i midlere signifikant bølgehøyde fra 0,37 m i øst til 1,33 m i vest, og flest høstepåvirkede arealer (trålflater) ble observert i de vestligste delene av høstefeltet. De to videostasjonene i referansefeltet som ligger i et bølgebeskyttet område, har midlere signifikant bølgehøyde på henholdsvis 0,30 m og 0,39 m.
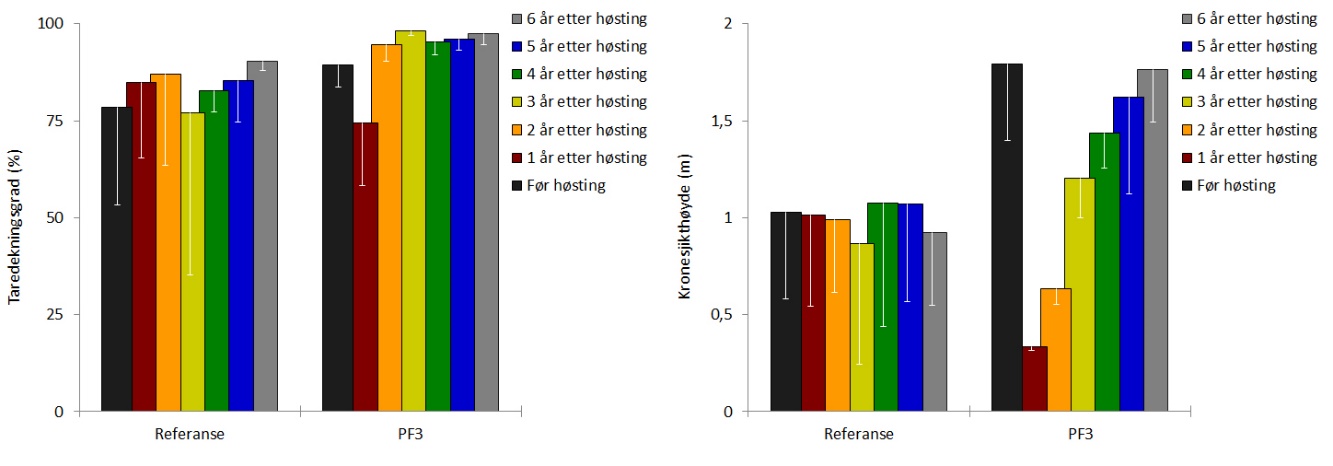
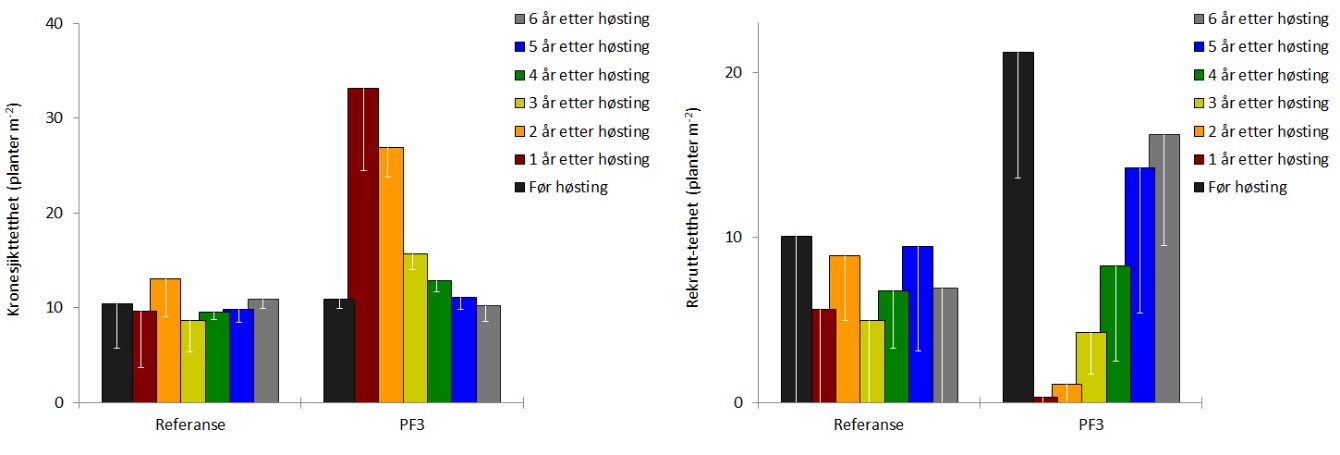
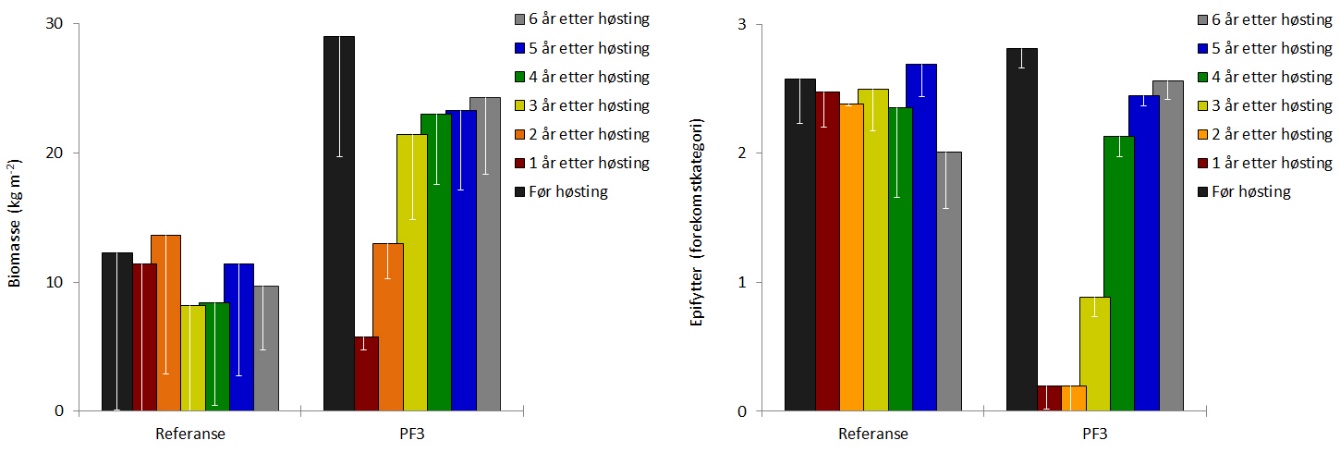
Dekningsgraden av stortare (andel bunnflate dekket av stortare) var reetablert allerede to år etter prøvehøstingen, og har siden vært stabil på rundt 95 % (Fig. 8). Gjennomsnittshøyden av tarevegetasjonen (kronesjikthøyden) observert i 2019 var ikke signifikant forskjellig (t-test, p = 0,80) fra kronesjikthøyden observert før prøvehøsting i 2013 (Fig. 8). Etter å ha økt de første årene etter høsting, har tettheten av stortareplanter i kronesjiktet på trålflatene flatet ut i de seneste årene og var i 2019 omtrent på samme nivå som observert før høsting i 2013 (Fig. 9). Tettheten av tarerekrutter i undervegetasjonen har økt i hele perioden etter prøvehøstingen, og var i 2019 ikke signifikant forskjellig (t-test, p = 0,11) fra rekrutt-tettheten observert i 2013 (Fig. 9). Gjennomsnittlig biomassetetthet av tare på høsteflatene ble i 2019 beregnet til 24 ± 6 kg tare per kvadratmeter, og var ikke signifikant forskjellig (t-test, p = 0,23) fra biomassetettheten observert før prøvehøsting i 2013 (Fig. 10). Forekomstene av epifytter på tarestilkene ble registrert på en semi-kvantitativ skala, og epifyttforekomstene på høstefeltet i 2019 var ikke signifikant forskjellig (t-test, p = 0,15) fra forekomstene registrert før prøvehøstingen seks år tidligere (Fig. 10).
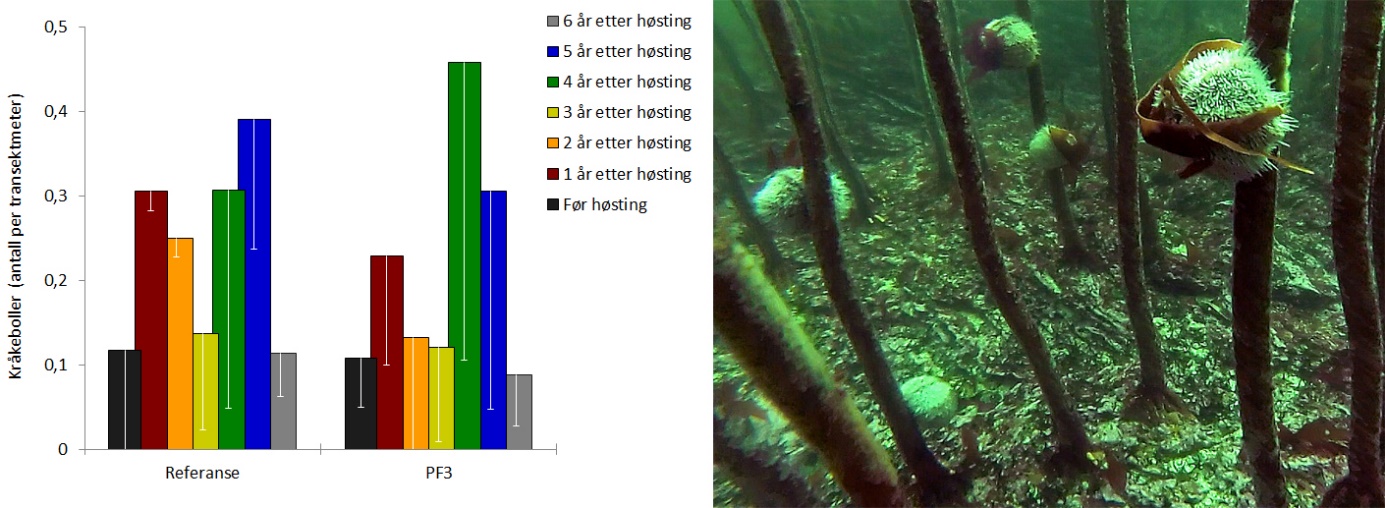
Som i tidligere år, ble det ikke observert innslag av grønn kråkebolle ( Strongylocentrotus droebachiensis ) på noen av videostasjonene i 2019. Registreringsfrekvensen av rød kråkebolle ( Echinus esculentus ) varierer fra år til år, og var i 2019 omtrent på samme nivå som i 2013, men lavere enn i 2017 og 2018 (Fig 11). I prøvehøstingsfeltet ble det i 2019 observert 81 individer av rød kråkebolle ( Echinus esculentus ) med en gjennomsnittlig registreringsfrekvens på 0,09 (± 0,06) per meter videotransekt. På referansestasjonene lå gjennomsnittlig registreringsfrekvens i 2019 på 0,11 (± 0,05) per meter videotransekt.
3.1.2 - Tareinnsamling
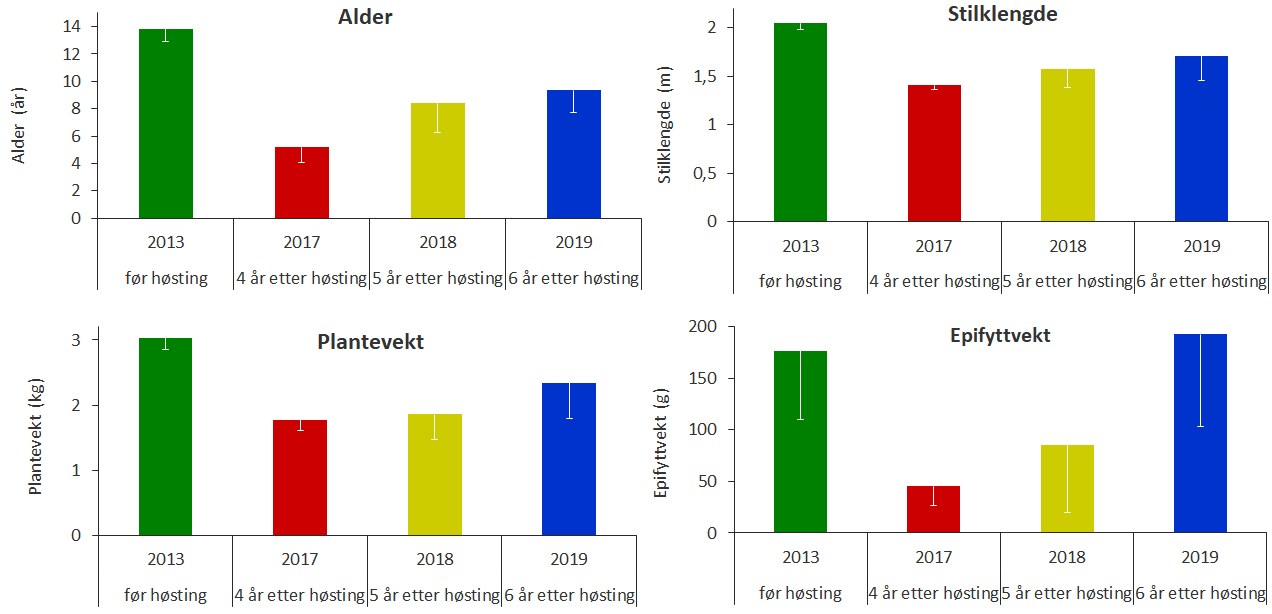
Tareplantene som ble samlet inn på en stasjon i vestlig, bølgeeksponert del av felt prøvehøstet i 2013 (PF3) hadde i 2019 en gjennomsnittlig alder på 9,4 år, og må hovedsakelig være rekruttert fra tareplanter som overlevde trålingen seks år tidligere. Tareplantene var i 2019 eldre, større og mer epifyttbevokste enn tareplanter samlet inn på samme stasjon i 2018. Selv om tareplantene samlet inn i 2019 var yngre og av mindre størrelse enn i 2013, var epifyttbiomassen på tarestilkene tilbake på samme nivå som før prøvehøstingen.
Det ble i juni 2019 samlet inn 11 tareplanter vha skrape på en stasjon i prøvehøstefelt PF3 (Fig. 1). Innsamlingsstasjonen ligger helt vest i feltet og er bølgeeksponert med midlere signifikant bølgehøyde på 1,3 meter. Tareprøvene samlet inn i 2019 ble sammenlignet med tareprøver tatt på den samme stasjon i 2013 (før prøvehøsting), 2017 og 2018 (Fig. 13).
På grunn av at det bare er gjennomført innsamling av tareplanter på en stasjon er det problematisk å teste eventuelle forskjeller fra før til etter høsting statistisk. Tareplantene samlet inn i prøvehøstefeltet hadde seks år etter høstingen en gjennomsnittlig alder på 9,4 år, noe som tilsier at vegetasjonen på trålflatene hovedsakelig er rekruttert fra tareplanter som var tilstede allerede før prøvehøstingen i 2013. Tareplantene var i 2019 eldre, større og mer epifyttbevokste enn tareplanter samlet inn på samme stasjon i 2018. Selv om tareplantene i 2019 var yngre og av mindre størrelse enn i 2013, var epifyttbiomassen på tarestilkene reetablert seks år etter prøvehøsting.
3.2 - Effekter av prøvehøstingen i 2014
3.2.1 - Videoundersøkelser
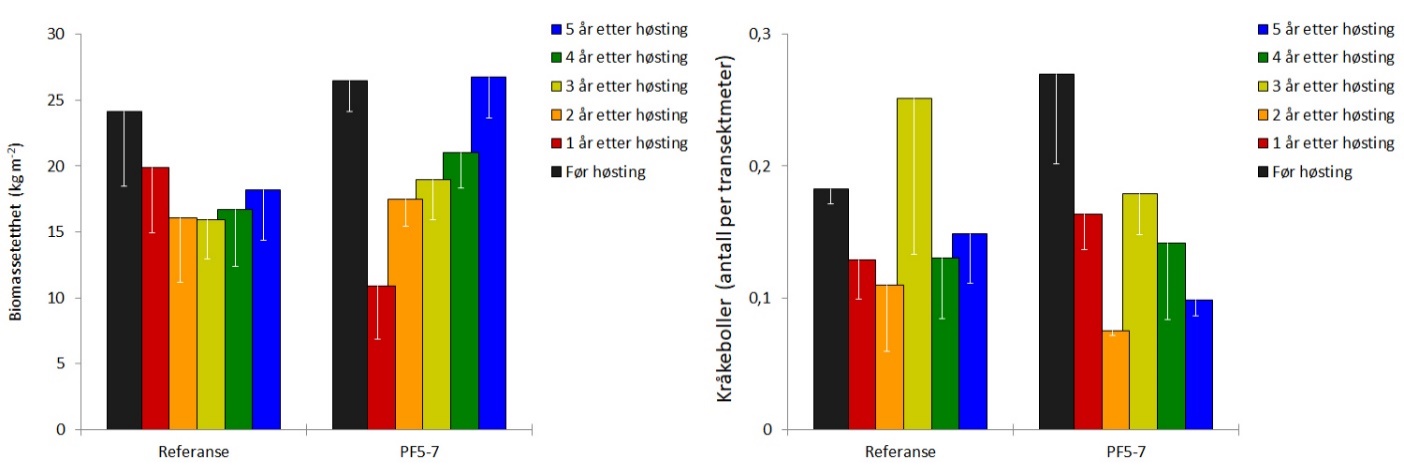
I 2014 ble det høstet 5.929 tonn stortare i tre felt (PF5-7) i Nordland. Videoobservasjoner på de tre høstefeltene og nærliggende referansefelt ble gjennomført i juni 2019, på de samme stasjonene som har vært undersøkt årlig siden 2014. Rekoloniseringen etter prøvehøstingen i 2014 har vært svært god med mer enn 95 % dekning av stortare på trålflatene allerede to år etter høsting. Den reetablerende tarevegetasjonen på trålflatene hadde i 2019 en gjennomsnittlig canopyhøyde på 1,7 m som er en økning på 0,2 m i forhold til 2018 observasjonene, og ikke signifikant forskjellig fra den gjennomsnittlige canopyhøyden (1,8 m) målt før prøvehøstingen i 2014. Forekomstene av rød kråkebolle var i 2019 på et lavt nivå, og ser ikke ut til å ha gitt synlige beiteskader på tarevegetasjonen. Grønne kråkeboller er ikke observert i perioden 2014-2019.
Totalt 5.929 tonn stortare ble høstet i tre prøvehøstefelt (PF5, PF6 og PF7) i Nordland i 2014. Det ble i juni 2019 gjennomført videoundersøkelser på stasjoner i prøvehøstefeltene PF5-7 og stasjoner i referansefelt nord i Horsvær naturreservat og sør i verdensarvområdet ved Vega (Fig. 1). Samtlige av disse videostasjonene har vært undersøkt årlig siden 2014. Den midlere signifikante bølgehøyden var 0,80 ± 0,20 m på videostasjonene i prøvehøstefeltene og 0,75 ± 0,04 m på videostasjonene i referansefeltene.
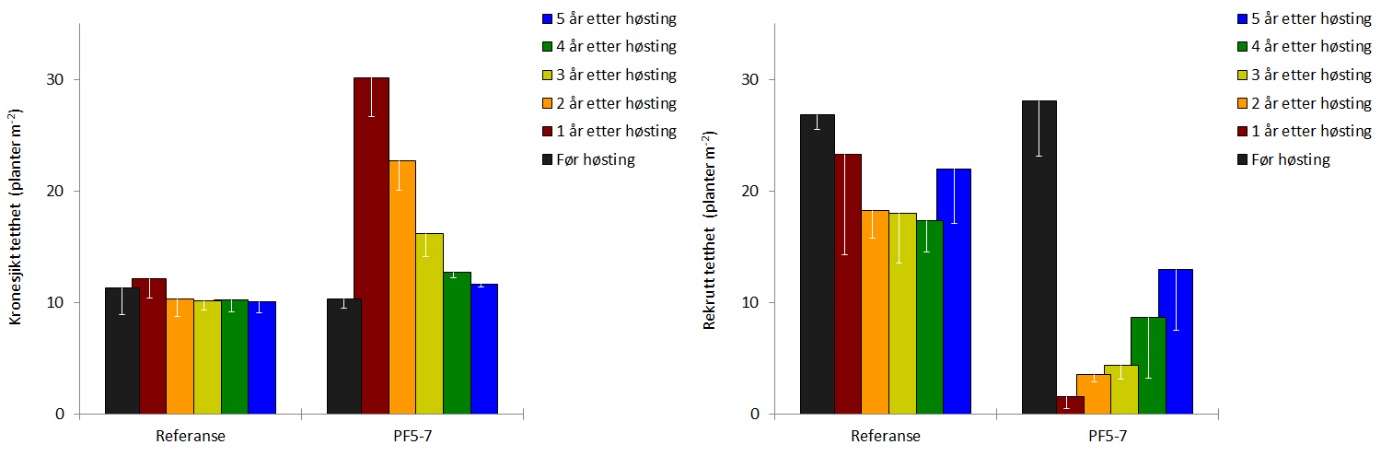
Rekoloniseringen av stortare etter prøvehøstingen i 2014 har vært svært god og den gjennomsnittlige dekningsgraden av tare (dvs % taredekket bunnflate) har vært høyere gjennom de tre siste årene, enn det som ble observert før prøvehøsting i 2014 (Fig 14, Tab. 1). Den gjennomsnittlige kronesjikthøyden av tareplanter på trålflatene hadde økt fra 1,1 ± 0,1 m i 2017 og 1,5 ± 0,2 m i 2018 til 1,7 ± 0,1 m i 2019, og er ikke lenger signifikant forskjellig fra kronesjikthøyden observert før prøvehøstingen i 2014 (Fig. 14, Tab. 1). Epifyttstrukturen på tarestilkene er i 2019 fortsatt dominert av skorpeformede typer på enkelte stasjoner (Fig. 15).
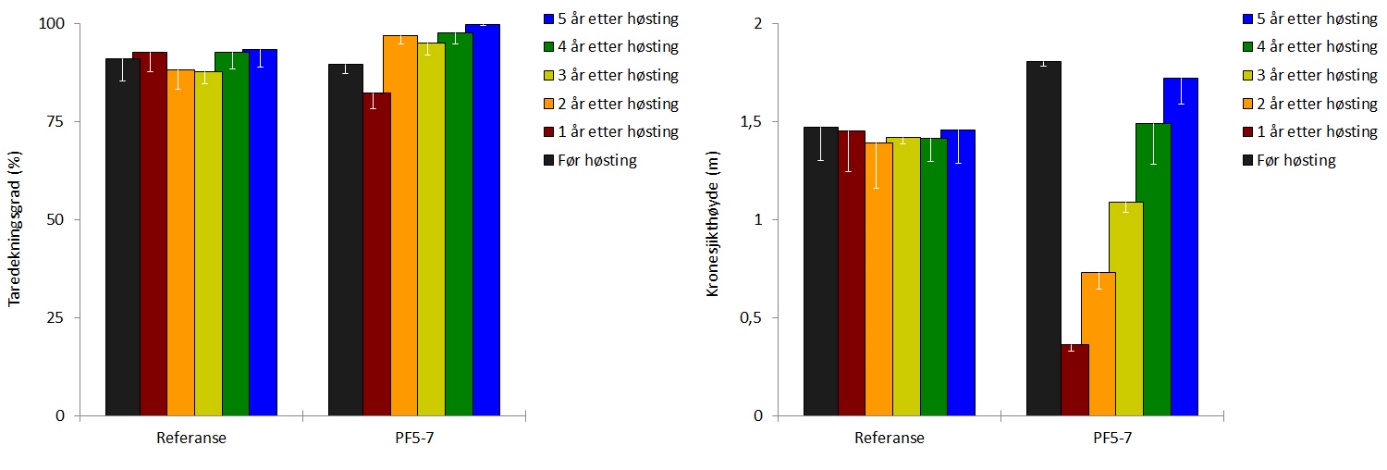

Tettheten av stortareplanter i kronesjiktet på trålflatene har avtatt de seneste årene, og er i 2019 ikke signifikant forskjellig fra tettheten av kronesjiktplanter observert før høstingen i 2014 (Fig. 16, Tab. 1). Gjennomsnittlig tetthet av tarerekrutter i undervegetasjonen i de tidligere høstede områdene ble i mai 2019 estimert til 13 ± 5 rekrutter per kvadratmeter, noe som fortsatt er signifikant lavere enn den gjennomsnittlige rekrutt-tettheten på 28±5 rekrutter per kvadratmeter som ble observert før prøvehøsting i 2014 (Fig. 16, Tab. 1).
Tabell 1. Lineære sammensatte modeller («linear mixed-effect models») for videodata med felttype (høstefelt vs referansefelt) krysset med periode (før høsting (2014) vs etter høsting (2015, 2016, 2017, 2018, 2019)) som uavhengige variabler. Modellen ble kjørt for et sett av responsvariabler (i kursiv): Taredekningsgrad, kronesjikthøyde, kronesjikt tetthet, tarerekrutt tetthet, tarebiomasse og forekomst av rød kråkebolle. Signifikante sannsynlighetsverdier (p < 0,05) for interaksjonsleddene (hvis signifikans indikerer effekt av taretråling) er understreket.
| Responsvariabel | Effekt | Estimat | t verdi | p verdi | Responsvariabel | Effekt | Estimat | t verdi | p verdi | |
| Taredekningsgrad (%) | (Skjæringspunkt) | 91,00 | 43,72 | <0,001 | Kronesjikthøyde (m) | (Skjæringspunkt) | 1,483 | 18,16 | <0,001 | |
| Periode (2014 vs 2015) | 1,667 | 0,617 | 0,518 | Periode (2014 vs 2015) | -0,050 | -0,698 | 0,488 | |||
| Periode (2014 vs 2016) | -2,500 | -0,974 | 0,333 | Periode (2014 vs 2016) | -0,100 | -1,395 | 0,167 | |||
| Periode (2014 vs 2017) | -3,167 | -1,123 | 0,221 | Periode (2014 vs 2017) | -0,067 | -0,93 | 0,355 | |||
| Periode (2014 vs 2018) | 1,667 | 0,65 | 0,518 | Periode (2014 vs 2018) | -0,067 | -0,93 | 0,355 | |||
| Periode (2014 vs 2019) | 2,667 | 1,039 | 0,302 | Periode (2014 vs 2019) | -0,05 | -0,698 | 0,488 | |||
| Felttype (referanse vs høstefelt) | 2,627 | -0,426 | 0,675 | Felttype (referanse vs høstefelt) | 0,335 | 3,298 | 0,002 | |||
| Interaksjon (2015 x høstefelt) | -9,394 | -2,945 | 0,004 | Interaksjon (2015 x høstefelt) | -1,395 | -15,66 | <0,001 | |||
| Interaksjon (2016 x høstefelt) | 9,864 | 3,092 | 0,003 | Interaksjon (2016 x høstefelt) | -0,982 | -11,02 | <0,001 | |||
| Interaksjon (2017 x høstefelt) | 8,712 | 2,731 | 0,008 | Interaksjon (2017 x høstefelt) | -0,652 | -7,312 | <0,001 | |||
| Interaksjon (2018 x høstefelt) | 6,424 | 2,014 | 0,048 | Interaksjon (2018 x høstefelt) | -0,233 | -2,619 | 0,011 | |||
| Interaksjon (2019 x høstefelt) | 7,242 | 2,271 | 0,026 | Interaksjon (2019 x høstefelt) | -0,041 | -0,459 | 0,647 | |||
| Kronesjikttetthet (per m-2) | (Skjæringspunkt) | 11,50 | 11,910 | <0,001 | Rekrutt tetthet (per m-2) | (Skjæringspunkt) | 26,83 | 9,424 | <0,001 | |
| Periode (2014 vs 2015) | 0,667 | 0,558 | 0,578 | Periode (2014 vs 2015) | -3,500 | -0,935 | 0,353 | |||
| Periode (2014 vs 2016) | -1,167 | -0,977 | 0,332 | Periode (2014 vs 2016) | -8,500 | -2,271 | 0,026 | |||
| Periode (2014 vs 2017) | -1,500 | -1,256 | 0,213 | Periode (2014 vs 2017) | -8,833 | -2,36 | 0,021 | |||
| Periode (2014 vs 2018) | -1,333 | -1,117 | 0,268 | Periode (2014 vs 2018) | -9,5 | -2,539 | 0,013 | |||
| Periode (2014 vs 2019) | -1,333 | -1,117 | 0,268 | Periode (2014 vs 2019) | -4,833 | -1,292 | 0,201 | |||
| Felttype (referanse vs høstefelt) | -1,271 | -1,042 | 0,308 | Felttype (referanse vs høstefelt) | 1,061 | 0,296 | 0,769 | |||
| Interaksjon (2015 x høstefelt) | 18,88 | 12,72 | <0,001 | Interaksjon (2015 x høstefelt) | -22,63 | -4,833 | <0,001 | |||
| Interaksjon (2016 x høstefelt) | 13,62 | 9,178 | <0,001 | Interaksjon (2016 x høstefelt) | -15,68 | -3,371 | 0,002 | |||
| Interaksjon (2017 x høstefelt) | 7,773 | 5,237 | <0,001 | Interaksjon (2017 x høstefelt) | -14,62 | -3,143 | 0,002 | |||
| Interaksjon (2018 x høstefelt) | 4,151 | 2,797 | 0,007 | Interaksjon (2018 x høstefelt) | -10,05 | -2,159 | 0,034 | |||
| Interaksjon (2019 x høstefelt) | 2,879 | 1,94 | 0,056 | Interaksjon (2019 x høstefelt) | -10,35 | -2,224 | 0,029 | |||
| Tarebiomasse (kg m-2) | (Skjæringspunkt) | 21,11 | 10,53 | <0,001 | Kråkeboller (per m-1) | (Skjæringspunkt) | 0,202 | 4,702 | <0,001 | |
| Periode (2014 vs 2015) | 0,561 | 0,292 | 0,771 | Periode (2014 vs 2015) | -0,042 | -0,85 | 0,440 | |||
| Periode (2014 vs 2016) | -4,124 | -2,142 | 0,035 | Periode (2014 vs 2016) | -0,088 | -1,802 | 0,076 | |||
| Periode (2014 vs 2017) | -4,269 | -2,217 | 0,023 | Periode (2014 vs 2017) | 0,053 | 1,088 | 0,28 | |||
| Periode (2014 vs 2018) | -3,124 | -1,937 | 0,109 | Periode (2014 vs 2018) | -0,073 | -1,496 | 0,139 | |||
| Periode (2014 vs 2019) | -3,730 | -1,937 | 0,056 | Periode (2014 vs 2019) | -0,055 | -1,122 | 0,265 | |||
| Felttype (referanse vs høstefelt) | 4,095 | 1,583 | 0,123 | Felttype (referanse vs høstefelt) | 0,075 | 1,401 | 0,167 | |||
| Interaksjon (2015 x høstefelt) | -20,81 | -8,694 | <0,001 | Interaksjon (2015 x høstefelt) | -0,076 | -1,241 | 0,219 | |||
| Interaksjon (2016 x høstefelt) | -8,108 | -3,388 | 0,001 | Interaksjon (2016 x høstefelt) | -0,113 | -1,862 | 0,066 | |||
| Interaksjon (2017 x høstefelt) | -3,541 | -1,48 | 0,143 | Interaksjon (2017 x høstefelt) | -1,171 | -2,799 | 0,007 | |||
| Interaksjon (2018 x høstefelt) | 1,31 | 0,547 | 0,586 | Interaksjon (2018 x høstefelt) | -0,068 | -1,109 | 0,271 | |||
| Interaksjon (2019 x høstefelt) | 4,739 | 1,98 | 0,051 | Interaksjon (2019 x høstefelt) | -0,124 | -2,036 | 0,045 |
Gjennomsnittlig biomassetetthet av stortare ble i 2019 estimert til 27±3 kg per kvadratmeter, og er ikke signifikant forskjellig fra biomassetettheten estimert før prøvehøstingen i 2014 (Fig. 17, Tab. 1). Registreringsfrekvensen av rød kråkebolle på stasjonene i prøvehøstefeltene var i gjennomsnitt 0,10 ± 0,01 individer per transektmeter i 2019, mot en registreringsfrekvens på 0,27±0,07 kråkeboller per transektmeter før prøvehøsting i 2014 (Fig. 17). Grønn kråkebolle har ikke vært observert på noen av videostasjonene, verken i prøvehøstefeltene eller i referanseområdene, i perioden 2014-2019.
3.2.2 - Tareinnsamlinger
Det ble samlet inn tareplanter på tre stasjoner i prøvehøstefelt PF5 og tre stasjoner i et referansefelt nord i Horsvær naturreservat i juni 2019. Tareplantene på stasjonene i prøvehøstefeltet hadde fem år etter høstingen en gjennomsnittlig alder på 5,4 år og var betydelig yngre enn planter samlet inn på de samme lokalitetene før prøvehøstingen i 2014. Epifyttbiomassen hos tareplantene samlet inn på prøvehøstingsstasjonene i 2019 var ca 30 %, av tilsvarende hos tareplanter samlet inn rett før prøvehøstingen i 2014. Analyser av tarestilkenes vekstsoner viser at stilktilveksten har vært høyere på prøvehøstingsstasjonene enn på referansestasjonene i de senere år.
Det ble i juni 2019 samlet inn stortare på tre stasjoner i prøvehøstefelt PF5 og på tre stasjoner i et referanseområde nord i Horsvær naturreservat i Nordland (Fig. 1) som ble sammenlignet med tareprøver samlet inn i forkant av prøvehøstingen i august 2014 og fire år etter prøvehøstingen i juli 2018. Innsamlingsstasjonene ligger alle innenfor en bølgeeksponeringskategori som varierer fra 0,6 m – 0,8 m midlere signifikant bølgehøyde og med dybder fra 6 m – 9 m. Tareplantene samlet inn i 2018 og 2019 var både yngre og av mindre størrelse i forhold til tareplantene samlet før prøvehøsting i 2014, både i høstefelt og referansefelt (Fig. 18, Tab. 2). Rundt 1/3 av tareplantene samlet inn i prøvehøstefeltet i 2019 var 6 år eller eldre, og må derfor være rekruttert fra tareplanter som var tilstede allerede før prøvehøstingen i 2014. Den gjennomsnittlige epifyttbiomassen på stasjonene i prøvehøstefeltet var i 2019 nesten tre ganger høyere enn i 2018, men likevel ikke mer enn ca 30 % av den gjennomsnittlige epifyttbiomassen på tarestilkene samlet inn før prøvehøstingen i 2014 (Fig 18).
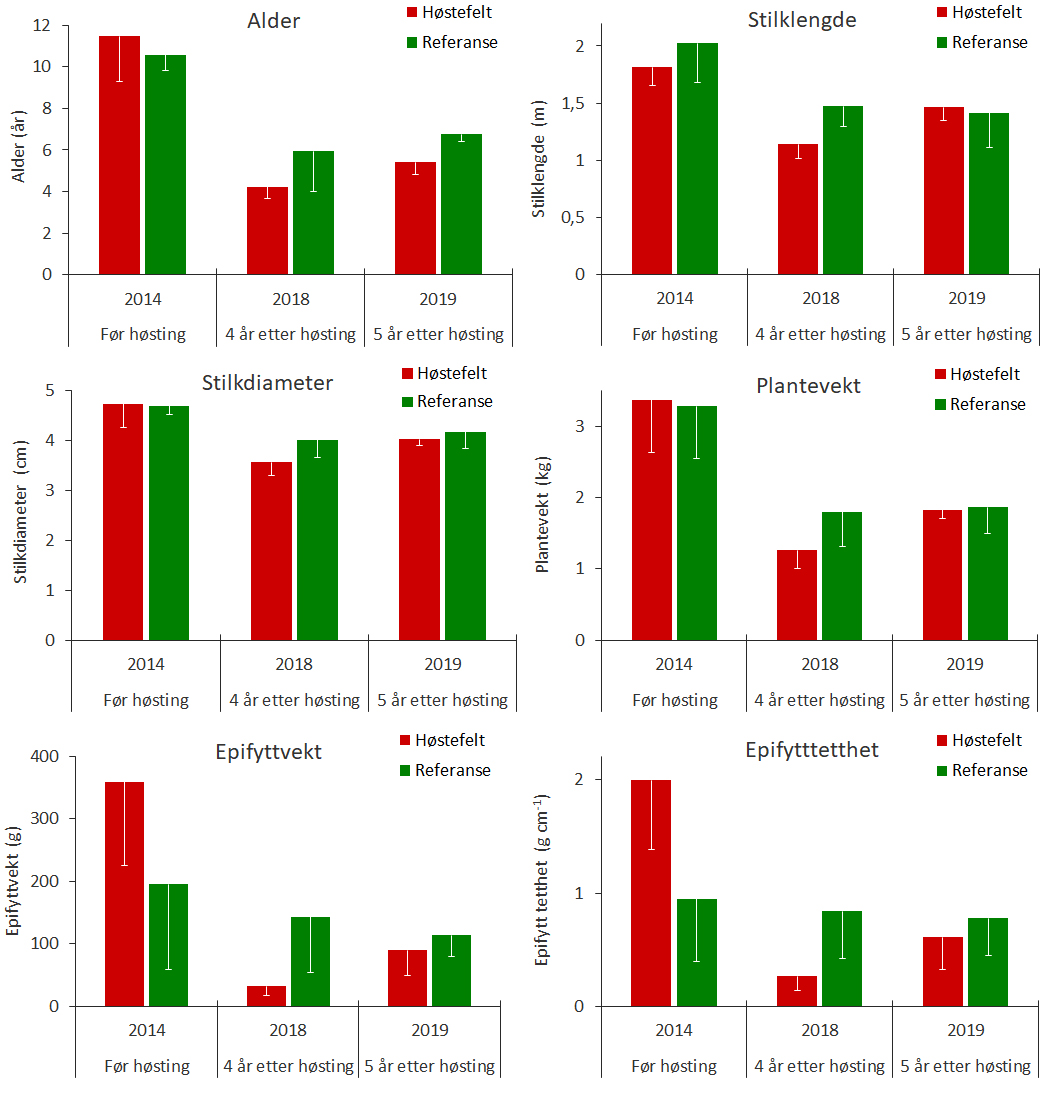
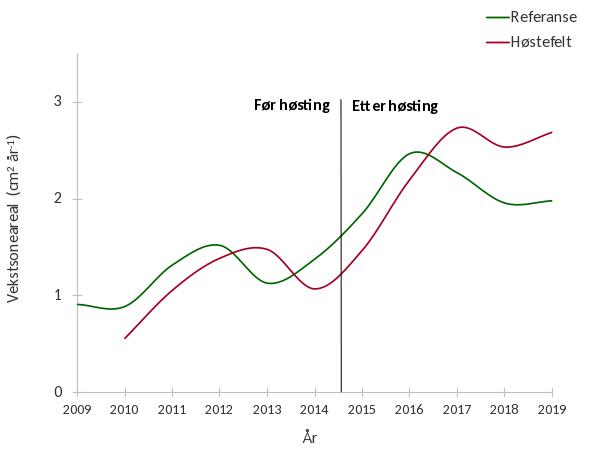
Vekstsoneanalyser av planter samlet inn i 2019 viste at tykkelsestilveksten av tarestilker økte i årene etter høstingen i 2014, både i prøvehøstefeltet og referansefeltet (Fig. 19). Den gjennomsnittlige årlige tykkelsestilveksten har imidlertid vært høyere for tarestilker samlet inn i prøvehøstefeltet enn i referansefeltet i perioden 2017-2019.
Tabell 2. Lineære sammensatte modeller («linear mixed-effect models») for tareinnsamlingsdata med felttype (høstefelt vs referansefelt) krysset med periode (før høsting vs etter høsting) som uavhengige variabler. Modellen ble repeterende kjørt for et sett av responsvariabler (i kursiv): Alder, stilklengde, stilkdiameter, plantevekt, epifyttvekt og epifytt tetthet (per cm tarestilk). Signifikante sannsynlighetsverdier (p < 0,05) for interaksjonsleddene (hvis signifikans indikerer effekt av taretråling) er understreket.
| Responsvariabel | Effekt | Estimat | t verdi | p verdi | Responsvariabel | Effekt | Estimat | t verdi | p verdi | |
| Alder (år) | (Skjæringspunkt) | 10,57 | 22,09 | <0,001 | Stilklengde (cm) | (Skjæringspunkt) | 202,3 | 22,33 | <0,001 | |
| Periode (2014 vs 2018) | -4,633 | -9,863 | <0,001 | Periode (2014 vs 2018) | -55,33 | -8,737 | <0,001 | |||
| Periode (2014 vs 2019) | -3,8 | -8,089 | <0,001 | Periode (2014 vs 2019) | -60,93 | -9,621 | <0,001 | |||
| Felttype (referanse vs høstefelt) | 0,9 | 1,33 | 0,3217 | Felttype (referanse vs høstefelt) | -21 | -1,639 | 0,155 | |||
| Interaksjon (2018 x høstefelt) | -2,6 | -3,914 | <0,001 | Interaksjon (2018 x høstefelt) | -12,07 | -1,347 | 0,18 | |||
| Interaksjon (2019 x høstefelt) | -2,267 | -3,412 | <0,001 | Interaksjon (2019 x høstefelt) | 25,77 | 2,877 | 0,005 | |||
| Stilkdiameter (cm) | (Skjæringspunkt) | 4,697 | 31,74 | <0,001 | Stilkvekt (g) | (Skjæringspunkt) | 2280 | 13,64 | <0,001 | |
| Periode (2014 vs 2018) | -0,680 | -6,800 | <0,001 | Periode (2014 vs 2018) | -1117 | -11,4 | <0,001 | |||
| Periode (2014 vs 2019) | -0,517 | -5,167 | <0,001 | Periode (2014 vs 2019) | -977,3 | -10,17 | <0,001 | |||
| høstefelt (referanse vs høstefelt) | 0,045 | 0,215 | 0,837 | høstefelt (referanse vs høstefelt) | -0,267 | -0,001 | 0,999 | |||
| Interaksjon (2018 x høstefelt) | -0,493 | -3,488 | <0,001 | Interaksjon (2018 x høstefelt) | -370,9 | -2,674 | 0,008 | |||
| Interaksjon (2019 x høstefelt) | -0,192 | -1,355 | 0,177 | Interaksjon (2019 x høstefelt) | -21,4 | -0,155 | 0,877 | |||
| Epifyttvekt (g) | (Skjæringspunkt) | 195,5 | 5,817 | <0,001 | Epifytt tetthet (g cm-1) | (Skjæringspunkt) | 0,948 | 5,938 | <0,001 | |
| Periode (2014 vs 2018) | -52,20 | -1,324 | 0,188 | Periode (2014 vs 2018) | -0,110 | -0,503 | 0,616 | |||
| Periode (2014 vs 2019) | -82,21 | -2,082 | 0,040 | Periode (2014 vs 2019) | -0,171 | -0,784 | 0,435 | |||
| høstefelt (referanse vs høstefelt) | 163,6 | 3,443 | 0,004 | høstefelt (referanse vs høstefelt) | 1,045 | 4,629 | <0,001 | |||
| Interaksjon (2018 x høstefelt) | -275,0 | -4,935 | <0,001 | Interaksjon (2019 x høstefelt) | -1,617 | -5,231 | <0,001 | |||
| Interaksjon (2019 x høstefelt) | -218,7 | -3,359 | 0,001 | Interaksjon (2019 x høstefelt) | -1,21 | -3,917 | <0,001 |
Epifytter (begroingsorganismer) på innsamlede stortarestilker ble bestemt til slekt/gruppe og rangert etter forekomst. Epifyttsamfunnene på tarestilkene ble dominert av tråd- og bladformede rødalger både før og etter høsting (Fig. 21). Av de 22 slektene/gruppene som ble registrert på prøvehøstefeltet i 2014 ble 16 også registrert i 2019, mens det ble registrert 2 slekter/grupper i 2019 som ikke ble registrert i 2014. På prøvehøstefeltet var den relative forekomsten til enkelte av epifyttene som f.eks draugfjær ( Ptilota ) var noe lavere i 2019 enn i 2014, mens den relative forekomsten av f.eks søl ( Palmaria ) var noe høyere i 2019 enn i 2014. Den relative forekomsten av epifyttiske dyr (hovedsakelig mosdyr (bryozoer)) var lavere i 2019, enn i 2014, mens forekomstene av eikeving (Phycodrys) var høyere i 2019, enn i 2014, både i prøvehøstefeltet og referansefeltet.
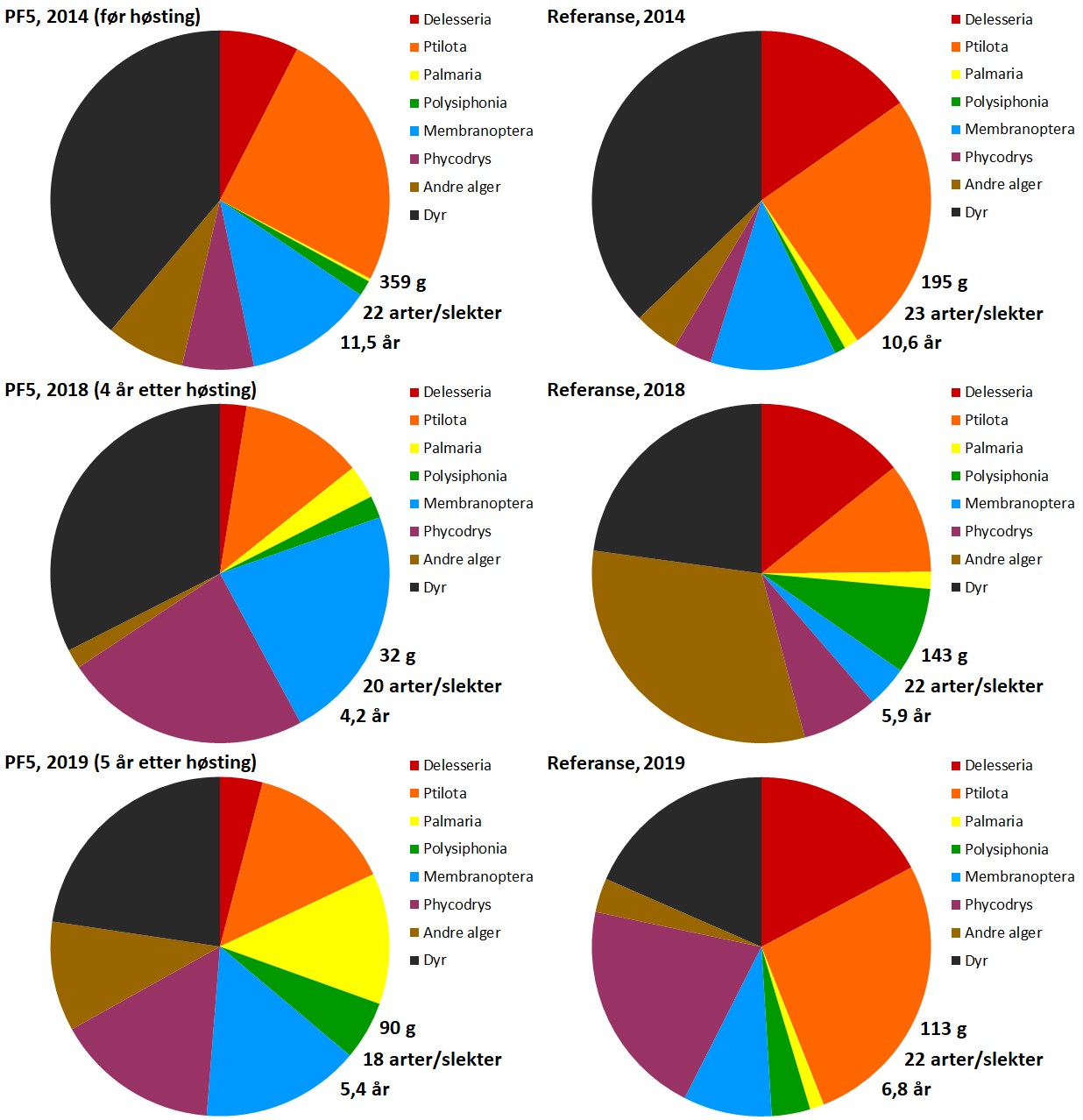
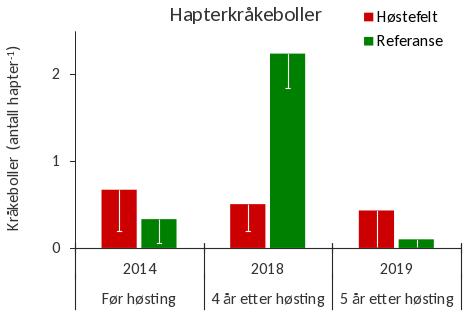
Juvenile individer av grønn kråkebolle ( S. droebachiensis ) ble registrert i tareplantenes festeorgan (hapter) på samtlige av innsamlingsstasjonene i 2019 (Fig. 21). Det ble i gjennomsnitt registrert 0,4 kråkeboller per tarehapter på stasjonene i prøvehøstefeltet, mens gjennomsnittet var 0,1 kråkeboller per tarehapter på stasjoner i referanseområdet. Mens forekomstene av hapterkråkeboller på prøvehøstefeltet har vært stabile, ble det i 2019 observert en nedgang i forekomstene av hapterkråkeboller på referansefeltet etter den kraftige økningen fra 2014 til 2018 (Fig. 21).
3.3 - Effekter av prøvehøstingen i 2017
3.3.1 - Videoundersøkelser
Det ble til sammen høstet 12.837 tonn stortare fordelt på fire felt (PF18-21) i Nordland i 2017. Samtlige prøvehøstfelt ble undersøkt vha video før høsting i 2017 og videotransektene på tre av prøvehøstefeltene (PF19-21) ble fulgt opp med nye undersøkelser i 2018 og 2019. Rekoloniseringen av tare på felt som ble prøvehøstet i 2017 har vært meget god, med en gjennomsnittlig dekningsgrad på ca 90 % allerede året etter høsting. Tarevegetasjons kronesjikthøyde ble i 2019 målt til 72 cm, som er en dobling fra året før, og ca 40 % av canopyhøyden målt før høsting i 2017. Forekomstene av rød kråkebolle var moderate og ser ut til å ha hatt liten effekt på reetableringen av tare på høsteflatene. Grønne kråkeboller ble ikke observert på noen av videostasjonene, verken før eller etter prøvehøstingen.
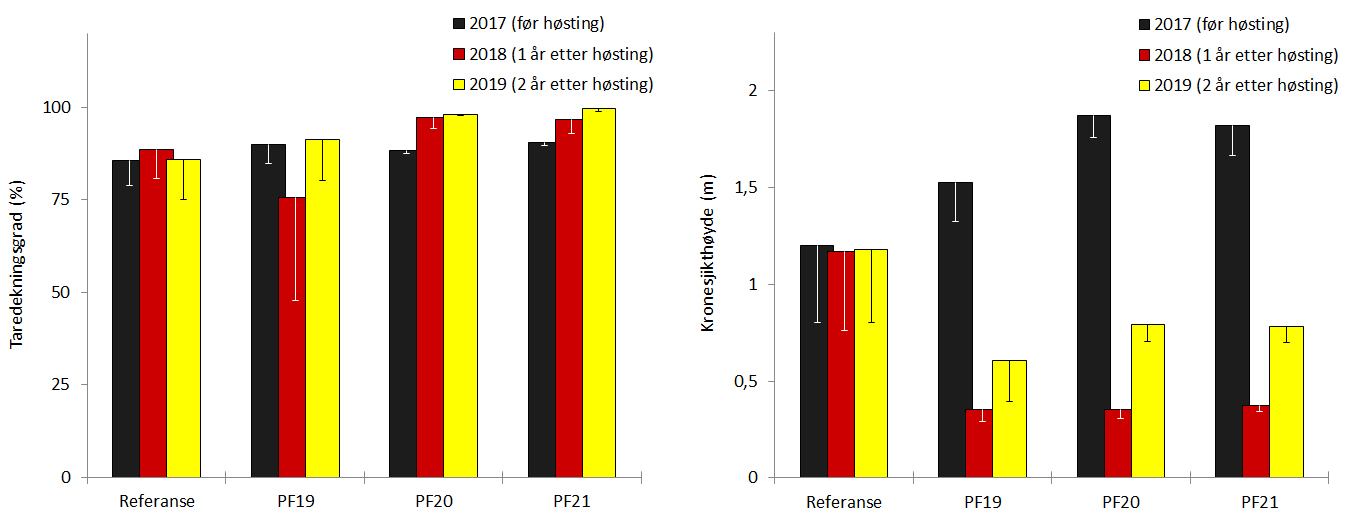
Det ble til sammen høstet 12.837 tonn stortare fordelt på fire felt (PF18-21) i Nordland i 2017 (Tab. 5). Tre av høstefeltene (PF19-21), der det til sammen ble høstet 9.427 tonn stortare i 2017, er fulgt opp med nye undersøkelser 2018 og 2019, langs de samme videotransektene som ble undersøkt før prøvehøstingen i 2017. Observasjonene fra prøvehøstefeltene ble også sammenlignet med tilsvarende observasjoner gjennomført i nærliggende referanseområder nordvest i Horsvær naturreservat og sør i verdensarvområdet ved Vega (Fig. 1). Den midlere signifikante bølgehøyden var 0,84 ± 0,13 m for videostasjonene i prøvehøstefeltene og 0,76 ± 0,11 m for videostasjonene i referansefeltene.
Det ble registrert trålspor langs samtlige videotransekt som ble undersøkt på prøvehøstefeltene i 2019 (Fig. 22). Gjennomsnittlig dekningsgrad av tare observert på høsteflatene i 2019 var 96 ± 5 % (Fig. 23). Den gjennomsnittlige kronesjikthøyden av den reetablerende tarevegetasjonen hadde økt fra 0,36 ± 0,01 m i 2018 til 0,73 ± 0,12 m i 2019. Vegetasjonshøyden på høsteflatene var i 2019 ca 40 % av vegetasjonshøyden observert før prøvehøsting to år tidligere (Fig. 23). Epifyttbegroingen på tarestilkene er to år etter prøvehøsting begrenset til flekkvis forekomst av skorpeformer (Fig. 22).

Det har ikke vært registrert innslag av grønn kråkebolle på videostasjonene undersøkt i perioden 2017-2019. Det ble i 2019 observert moderate forekomster av rød kråkebolle på stasjoner i prøvehøstefeltene (PF19-21) med en gjennomsnittlig registreringsfrekvens 0,03 (± 0,01) individer per transektmeter (Fig. 24). Registreringsfrekvensen av rød kråkebolle på de samme stasjonene før prøvehøstingen i 2017 var i gjennomsnitt 0,13 (± 0,05) individer per transektmeter.
3.4 - Kartlegging av tareforekomster på kyststrekningen Herøy – Bodø i 2019
3.4.1 - Videoundersøkelser
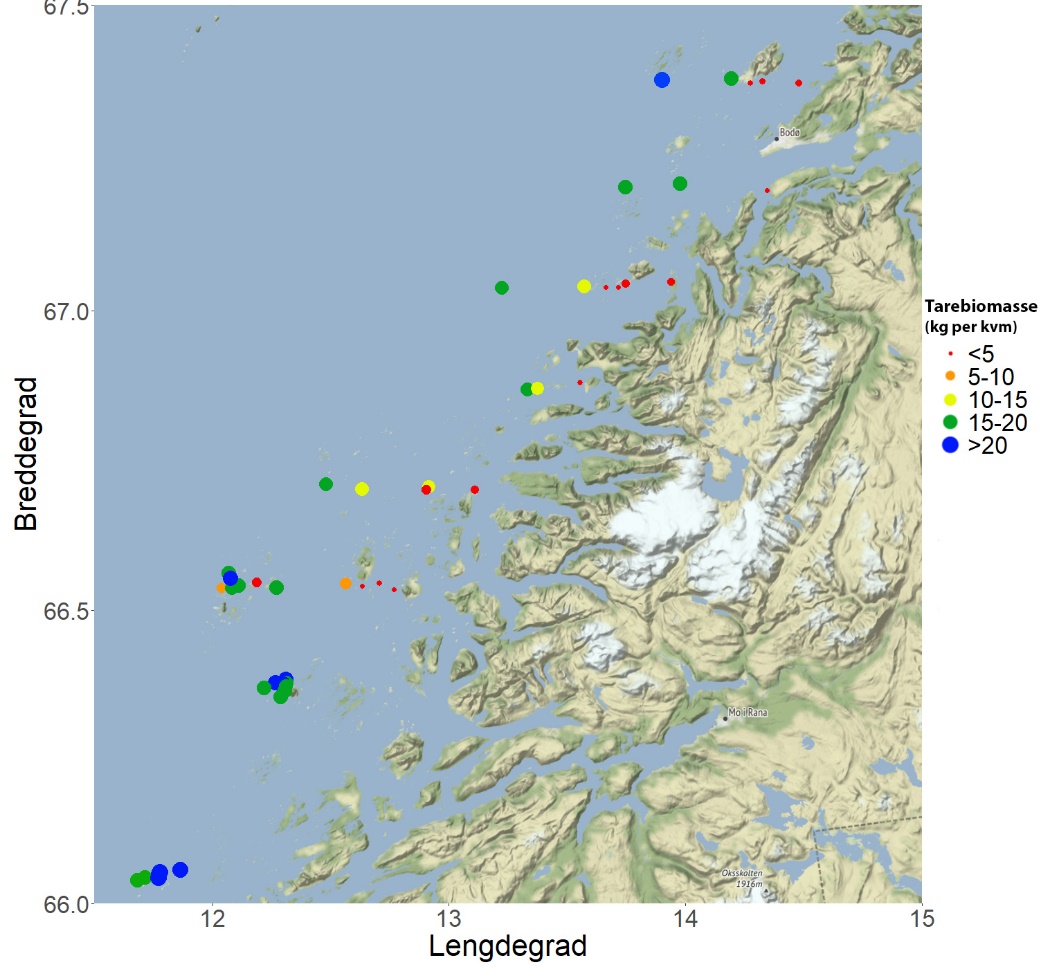
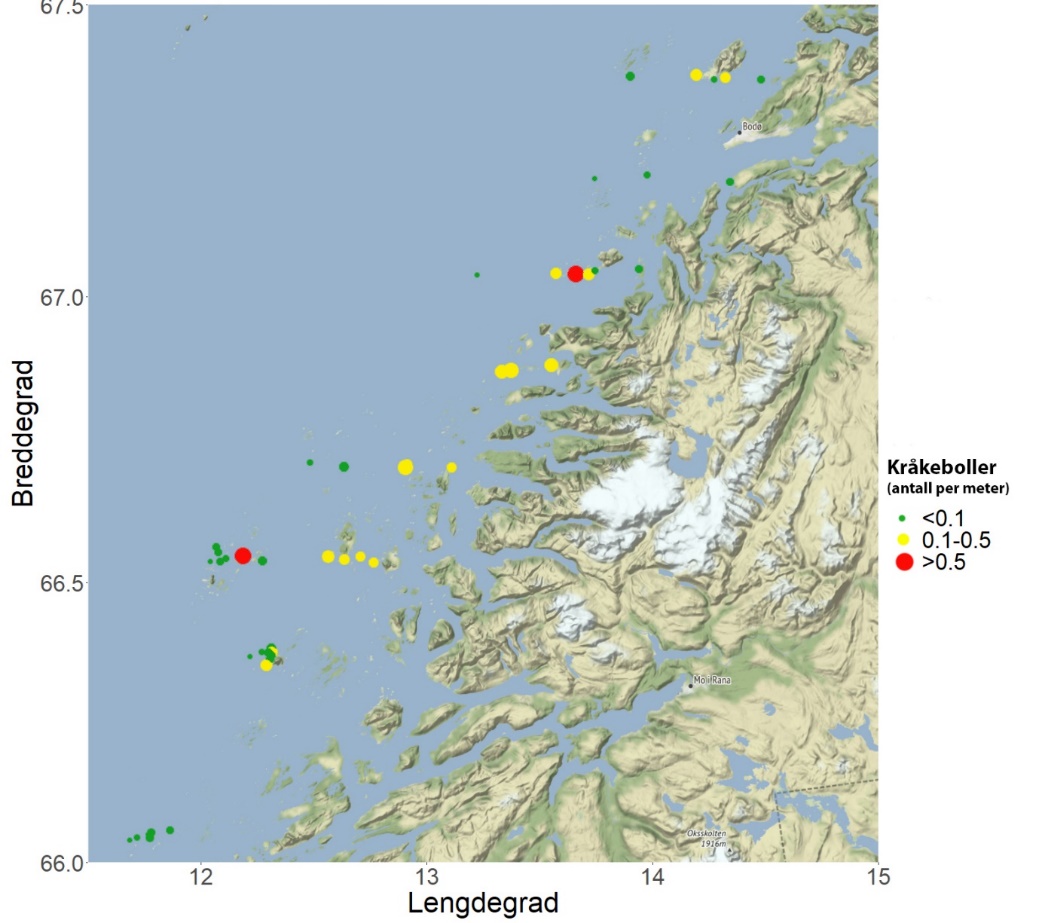
Det ble i juni 2019 gjennomført videoundersøkelser på 48 stasjoner fordelt på 8 breddgradsektorer i området fra Herøy til Bodø i Nordland for å kartlegge forekomster av tare og kråkeboller. Stortare ble registrert på 47 av 48 stasjoner og arten dominerte bunnvegetasjonen på de vestlige og mest bølgeeksponerte videostasjoner i samtlige sektorer. Sukkertare dominerte bunnvegetasjonen på østlige, mer bølgebeskyttede stasjoner. På enkelte stasjoner var det også innslag av butare og draugtare. Den gjennomsnittlige beregnede biomassetettheten av stortare varierte fra 0 – 24 kg per kvadratmeter, og økte fra østlige til vestlige deler av sektorene. Grønn kråkebolle ble ikke observert på noen av videostasjonene. Forekomsten av rød kråkebolle var moderat på de fleste stasjoner, med en gjennomsnittlig registreringsfrekvens på 0,12 individer per transektmeter. På stasjonene i de to sørligste sektorene som også ble undersøkt i 2018, ble det ikke registrert endringer i kråkebolleforekomstene fra 2018 til 2019.
Det ble gjennomført videokartlegging på 49 stasjoner fordelt på 8 breddgradsektorer i området fra Herøy (66° 02’ N) til Bodø (67° 22’ N) i Nordland i juni 2019 (Fig. 2). Bunnen på de fleste videostasjonene var dominert av fjell, mens enkelte av videostasjonene i østlige deler av sektorene hadde høyere innslag av bløtbunn. Gjennomsnittsdypet på stasjonene varierte fra 7 m til 13 m, mens midlere signifikant bølgehøyde varierte fra 0,18 m til 1,53 m, og økte fra østlige til vestlige deler av sektorene.
Det ble registrert stortare ( Laminaria hyperborea ) langs 47 av 48 videotransekt (Fig 25, 26), og forekomstene (dekningsgraden) var positivt korrelert med midlere signifikant bølgehøyde (r = 0,43, p = 0,002). Det ble registrert sukkertare ( Saccharina latissima ) langs 22 av 48 videotransekt (Fig. 26, 27), og forekomstene var negativt korrelert med bølgehøyde (r = -0,36, p = 0,011). Spredte innslag av butare ( Alaria esculenta ) ble registrert på 29 av 48 stasjoner (Fig. 25, 27), men forekomstene var ikke signifikant korrelert med bølgehøyde (r = 0,27, p = 0,070). Draugtare ( Saccorhiza polyschides ), som er en art med sørlig utbredelse i Norge, ble kun registrert på fire stasjoner i den tredje sørligste breddegradssektoren (66° 32’ N) ved Lurøy – Træna. Gjennomsnittlig estimert biomassetetthet av stortare varierte fra 0 – 24 kg per kvadratmeter, og økte fra østlige til vestlige deler av sektorene (Fig. 28). Stortarebiomassen var positivt korrelert med signifikant midlere bølgehøyde (r = 0,52, p < 0,001).

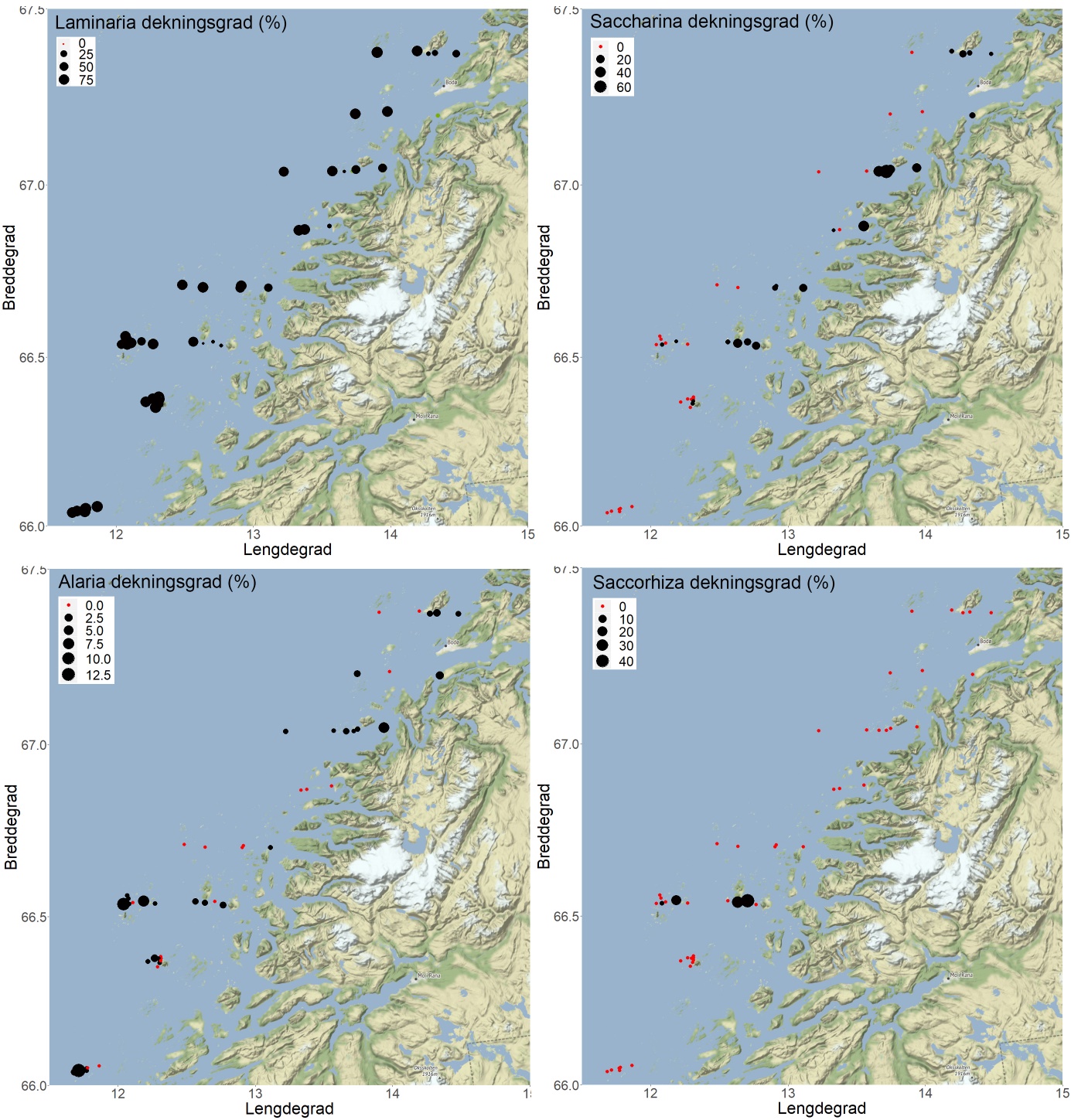

Det ble ikke registrert grønn kråkebolle ( Strongylocentrotus droebachiensis ) på videostasjonene undersøkt på strekningen Herøy – Bodø i 2019. Det ble derimot registrert 863 individer av rød kråkebolle ( Echinus esculentus ) langs en total transektdistanse på 7,3 km. Registreringsfrekvensen av kråkeboller varierte fra 0.00 til 0.54 individer per transektmeter, med en gjennomsnittlig registreringsfrekvens på 0,12 (± 0,04) individer per transektmeter (Fig. 29). Kråkebolleforekomstene var negativt korrelert med signifikant midlere bølgehøyde (r = -0,31, p = 0,004). Ni av videostasjonene i de to sørligste sektorene ble undersøkt også i 2018, og på disse ble det ikke registrert endringer i kråkebolleforekomstene fra 2018 til 2019 (t-test, p = 0,88).
3.4.2 - Tareinnsamlinger
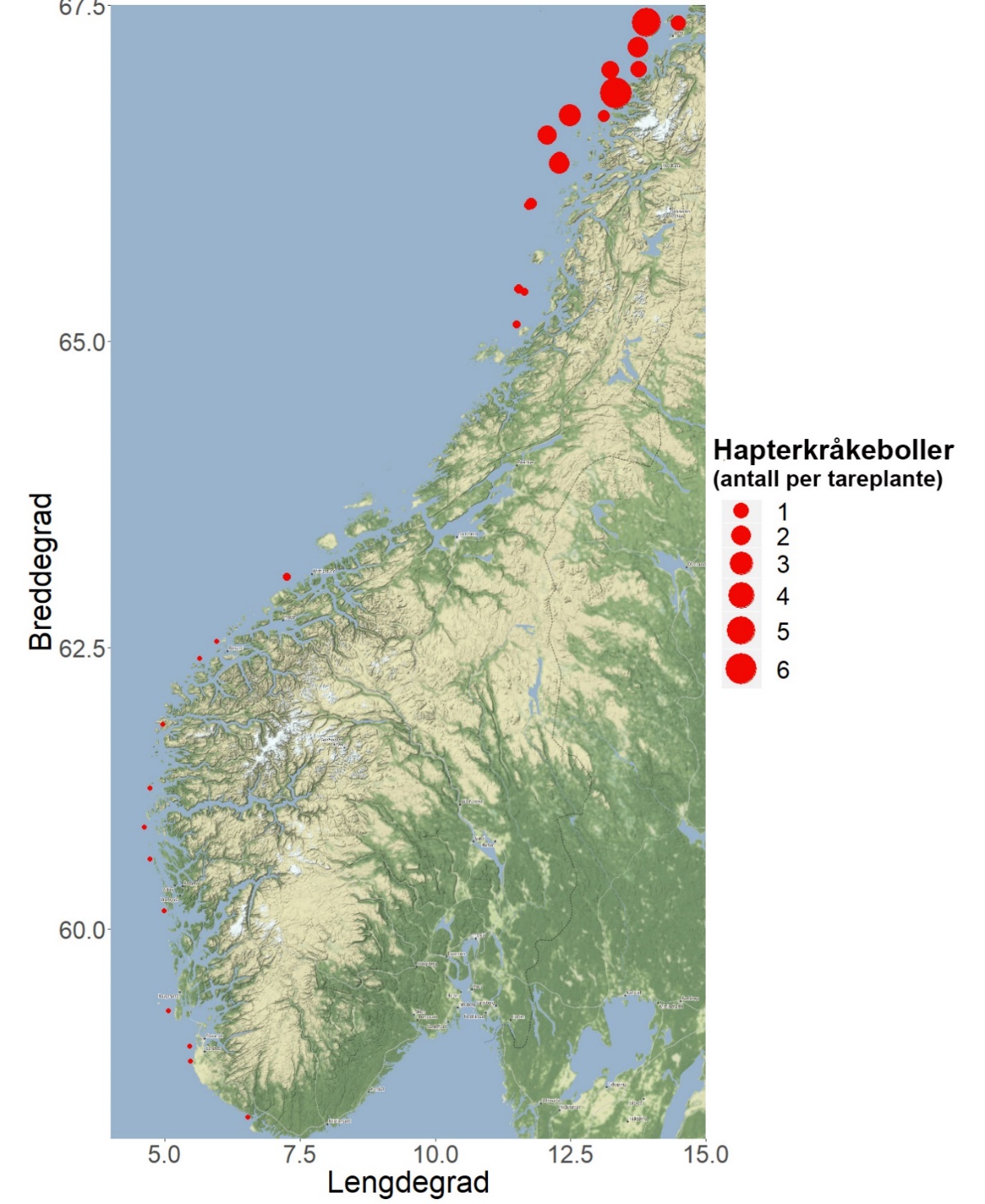
Stortareplanter ble samlet inn på totalt 15 stasjoner på kyststrekningen Herøy – Bodø 2019, og sammenlignet med tilsvarende innsamlinger gjort i referanseområder på strekningen Flekkefjord – Brønnøysund. Tareplantenes gjennomsnittsalder økte med bølgeeksponering på innsamlingsstasjonene i Nordland, og var i bølgeeksponerte områder høyere enn hos tareplanter samlet inn på stasjoner med tilsvarende bølgeeksponering lenger sør. Veksthistorikkanalyser indikerer at den årlige stilktilveksten er tregere hos tareplanter på strekningen Herøy – Bodø, enn lenger sør i Nordland og på vestlandskysten. Tettheten av juvenile grønne kråkeboller (Strongylocentrotus droebachiensis) observert i de innsamlede tareplantenes festeorgan (hapter) var betydelig høyere på stasjoner nord for Lurøy i Nordland, enn på stasjoner lenger sør.
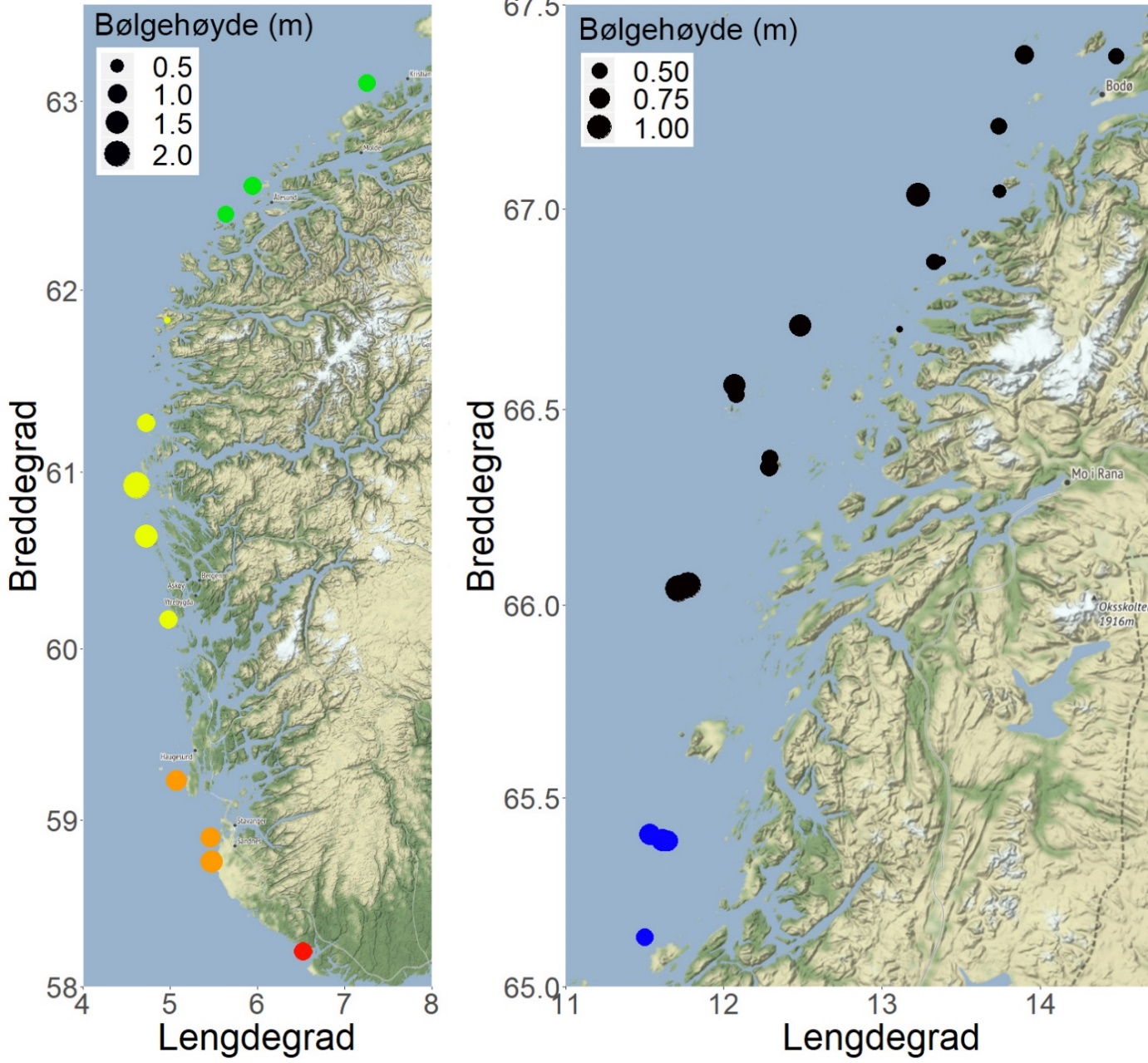
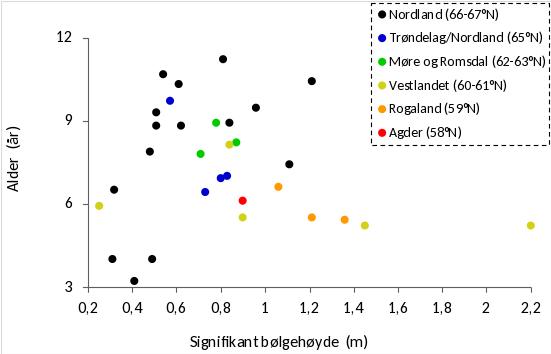
Det ble i juni 2019 samlet inn stortareplanter vha skrape på 15 stasjoner på kyststrekningen Herøy (66° 02’ N) - Bodø (67° 22’ N) for undersøkelse av morfologi, alder, veksthistorikk, epifytter og hapterkråkeboller. Det ble også gjort sammenligninger med tareplanter samlet inn i trålfrie referanseområder på kyststrekningen fra Flekkefjord i Agder til Horsvær sør i Nordland 2019 (Fig. 30). Tareprøvene på strekningen Agder – Møre og Romsdal (12 stasjoner) ble samlet inn i april 2019, mens tareprøvene rundt fylkesgrensen mellom Trøndelag og Nordland (4 stasjoner) ble samlet inn i juni 2019. Innsamlingsstasjonene dekker en bred bølgeeksponeringsgradient fra midlere signifikant bølgehøyde 0,25 m til 2,20 m (Fig. 31). Tareplantenes gjennomsnittsalder økte med grad av bølgeeksponering i Nordland, men ikke for stasjonene lenger sør (Fig. 32). På stasjoner med lav bølgeeksponering det en tendens til yngre gjennomsnittsalder for stortareplanter samlet inn i nordlig del av Nordland enn lenger sør. På mer eksponerte stasjoner var tareplantenes gjennomsnittsalder høyere i nordlige deler av Nordland (66-67°N) enn lenger sør.
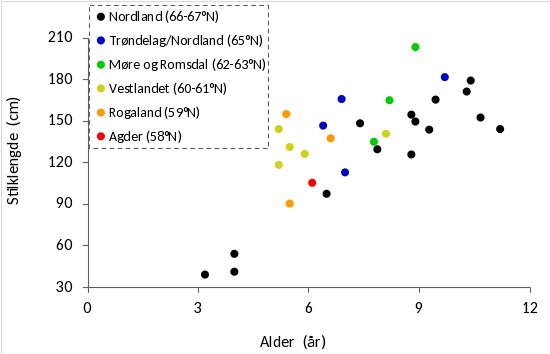
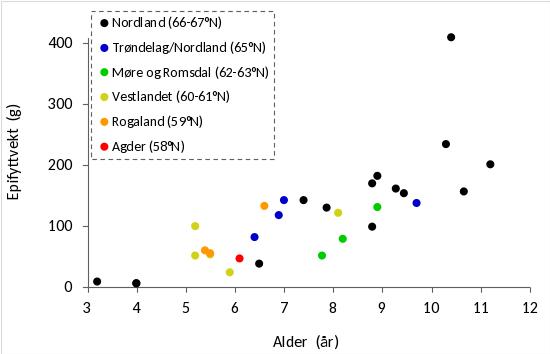
Tareplantenes gjennomsnittlige stilklengde på stasjonene varierte fra 40 cm til 200 cm og økte med den gjennomsnittlige alderen på stasjonene (Fig. 33). På stasjoner med tilnærmet lik gjennomsnittsalder var det en tendens til noe kortere stilklengder i Nordland (66-67°N), enn på stasjoner lenger sør. Den gjennomsnittlige epifyttbiomassen på stasjonene økte med tareplantenes gjennomsnittlige alder (Fig. 34).
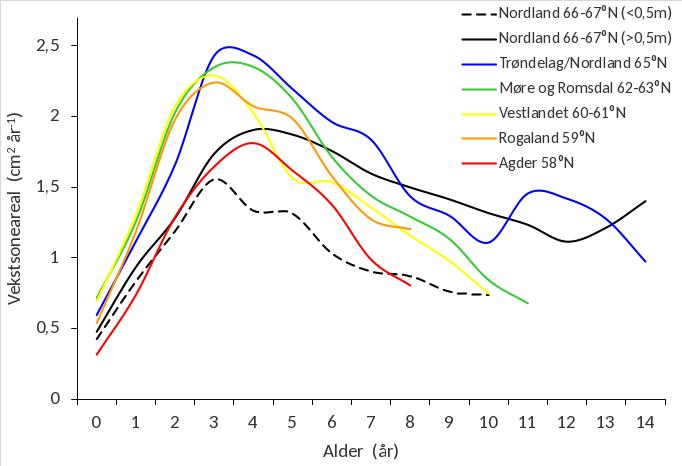
Størrelsen (arealet) av tarestilkenes vekstsoner gir informasjon om tykkelsestilveksten i et gitt år. Gjennomsnittlig årlig tykkelsestilvekst per aldersgruppe av stortare i ulike regioner er vist i figur 35. De nordligste områdene i Nordland (66-67°N) inkluderer flere bølgebeskyttede stasjoner enn regionene lenger sør (Fig. 31). For denne regionen er det derfor beregnet separate veksthistorikklinjer for eksponerte stasjoner med midlere signifikant bølgehøyde >0,5 m (som er sammenlignbare med regionene lenger sør) og mer beskyttede stasjoner med midlere signifikant bølgehøyde <0,5 m. Tykkelsestilveksten varierte med tarestilkenes alder og nådde et maksimum når plantene var ca 3-4 år gamle, for deretter å gradvis avta med økende alder. Den årlige tilvekstraten for de aller yngste aldersgruppene (0 - 2 år) var høyest for tareplanter samlet inn på kyststrekningen Rogaland - Møre og Romsdal. For eldre aldersgrupper (>3 år) var den årlige stilktilveksten høyere hos planter fra grenseområdet mellom Trøndelag/Nordland, enn planter fra Møre og Romsdal, som igjen hadde høyere tilvekst enn planter fra Rogaland og Vestlandet. For stortare i eksponert sektor hadde plantene samlet inn i Agder den laveste gjennomsnittlige årlige stilktilveksten av alle regioner gjennom mesteparten av livsløpet.
Med unntak av tareplantene fra Agder, hadde plantene samlet inn i den nordlige delen av Nordland (66-67°N), det laveste og seneste vekstmaksimum av samtlige regioner. For eldre aldersgrupper (>6 år) hadde imidlertid plantene fra Nordland (66-67°N), sammen med plantene fra grenseområdet mellom Trøndelag/Nordland, høyere årlig stilktilvekst enn de sørligere regionene. Den årlige tykkelsestilveksten var lavere på bølgebeskyttede, enn bølgeeksponerte stasjoner i Nordland for alle aldersgrupper, og vekstforskjellene mellom de to eksponeringskategoriene var størst for eldre (>3 år) aldersgrupper.
Forekomster av juvenile individer av grønn kråkebolle ( S. droebachiensis ) i tareplantenes festeorgan (hapter) økte fra sør til nord (Fig. 36). Det ble observert hapterkråkeboller på samtlige 15 undersøkte stasjoner på kyststrekningen Herøy – Bodø i 2019, med et gjennomsnitt på 2 individer per tareplante. Til sammenligning så hadde plantene samlet inn på stasjonene rundt grensen ved Trøndelag/Nordland et gjennomsnitt på 0,1 hapterkråkeboller per plante. På kyststrekningen mellom Flekkefjord i Agder og Giske i Møre og Romsdal ble det ikke observert en eneste hapterkråkebolle i 2019.
4 - Diskusjon
Etter at det ble rapportert om tilbakegang av kråkeboller og gjenvekst av tare i sørlige deler av Nordland (Norderhaug & Christie 2009), har tarenæringen og forvaltningen vist interesse for å utrede potensialet for en fremtidig kommersiell utnyttelse av tareressursene i denne regionen. I den forbindelse har det blitt gjennomført prøvehøsting av stortare ( Laminaria hyperborea ) i avgrensede områder på kyststrekningen mellom Trøndelagsgrensen og Vega i perioden 2013-2017. Tareskogene langs denne delen av Norskekysten har inntil nylig vært beitet ned av grønne kråkeboller ( Strongylocentrotus droebachiensis) og kan være spesielt sårbare overfor inngrep. Det har derfor vært viktig å følge utviklingen av taresamfunnene etter prøvehøstingene i dette området over tid.
Videoundersøkelser gjennomført etter prøvehøstingene i Nordland har demonstrert en rask rekolonisering av stortare på høstefeltene gjennom hele perioden (Steen et al. 2014ab, 2015ab, 2016b, 2018ab). På felt som ble prøvehøstet i 2017, ble det allerede året etter observert en gjennomsnittlig dekningsgrad av stortare på ca 90 %, og vegetasjonshøyden doblet seg fra 2018 til 2019. Utviklingen i den tidlige reetableringsfasen etter prøvehøstingen av tare ser dermed ut til å følge omtrent samme mønster som tidligere observert i Nord-Trøndelag (Steen et al. 2014b, 2016a). Veksten av stortare på feltene som ble prøvehøstet i Nordland i 2013 og 2014 har vært god med en jevn økning av vegetasjonshøyden år for år, mens tarevegetasjonens høyde i uhøstede referanseområder varierte lite fra år til år. På feltet som ble prøvehøstet i 2014 var tarevegetasjonens kronesjikthøyde og biomasse tilbake på samme nivå i 2019 som før prøvehøstingen fem år tidligere.
Tareplantenes epifytter er viktig habitat for små snegl, krepsdyr og andre mobile evertebrater og har stor betydning for tareskogens totale biomangfold og produksjon (Christie et al. 1998, 2003, 2007, Norderhaug et al. 2003, Teagle et al. 2017). På felt som ble prøvehøstet i 2014 er epifyttforekomstene fortsatt ikke reetablert fem år etter høsting. På feltet som ble prøvehøstet seks år tidligere, i 2013, ser derimot epifyttforekomstene ut til å være reetablert per 2019. I en studie gjennomført ved Smøla i Møre og Romsdal fant Christie et al. (1998) at forekomst og artsmangfold av epifytter på tarestilkene ikke var reetablert 6 år etter tarehøsting. Den lengre restitusjonsperioden for epifyttene utenfor Smøla kan f.eks skyldes at de innsamlede tareplantene 6 år etter høsting var yngre her (~7 år), enn tareplantene samlet inn 6 år etter høsting i Nordland (~9 år). Epifyttforekomstene er sterkt korrelert med tarevegetasjonens alder (Steen et al. 2015b, 2016b, 2018ab) og vil i sørlig del av Nordland kreve en hvileperiode på seks år mellom påfølgende høstinger (som tilsvarer en høstesyklus på syv år) for å reetableres. Restituering av tarevegetasjonens aldersstruktur ser ut til å kreve lengre tid, da plantene samlet inn i 2019 fortsatt var yngre enn det som ble observert før prøvehøstingene både fem og seks år tidligere.
Den gjennomsnittlige alderen observert hos innsamlede tareplanter på høstefeltene var høyere enn antall år som er gått siden prøvehøstingene ble gjennomført og indikerer at mye av rekrutteringen har vært fra planter som var tilstede allerede på høstetidspunktet. Tidligere undersøkelser fra Nord-Trøndelag og Nordland har også vist at den reetablerende tarevegetasjonen etter førstegangshøstinger i stor grad ble rekruttert fra planter som overlevde taretrålens passering (Steen et al. 2014, 2016a, 2018ab). Videoobservasjonene tyder på at tettheten av tarerekrutter i undervegetasjonen på høsteflatene hadde økt siden 2018, og var i 2019 ikke signifikant forskjellig fra det som ble observert før prøvehøstingen i 2013. På feltene som ble prøvehøstet fem år tidligere, i 2014, var imidlertid tettheten av tarerekrutter i undervegetasjonen per 2019 fortsatt ikke reetablert. En reetablering av tarerekruttforekomstene vil derfor kreve en hvileperiode på 6 år mellom påfølgende høstinger (dvs en høstesyklus på 7 år) En høstesyklus som er for kort til at rekrutteringsvegetasjonen utvikles vil kunne medføre at tarevegetasjonens reetablering må skje fra mindre utviklede livsstadier (kimplanter, sporer, etc) og gi langsommere gjenvekst på sikt, enn hva tilfellet var etter førstegangshøstingen da tettheten av synlige tarerekrutter i undervegetasjonen var høy. For hard og hyppig beskatning av tareressursene vil kunne gi skadelige effekter på taresamfunnene over tid (Buschmann et al. 2014, Mac Monagail et al. 2017), og det som ser ut til å være en bærekraftig høsting mhp reetablering av tarevegetasjonen etter en høsterunde, kan derfor vise seg mindre bærekraftig ved gjentatte høstinger av det samme området.
Vekstsoneanalyser av innsamlede tareplanter viser at stilktilveksten økte i årene etter prøvehøsting. Stilktilveksten varierer med tareplantenes alder (Steen et al . 2015b, 2016b) og vekstøkningen etter prøvehøstingen skyldes sannsynligvis bedre lystilgang for et sett med yngre og mer vekstpotente tareplanter. Økning i stilktilveksten i perioden 2014-2019 ble også registrert hos tareplanter samlet inn på referansestasjoner som ikke har vært gjenstand for tarehøsting og kan skyldes et naturlig generasjonsskifte da plantene samlet inn i 2019 var klart yngre enn plantene samlet inn i 2014. Tareinnsamlingene før prøvehøstingen i 2014 ble gjort i slutten av august, mens øvrige innsamlinger i Nordland er gjort på forsommeren. Sesongvariasjoner mellom de ulike innsamlingsperiodene vil også kunne ha påvirket resultatene, da både tareblad og epifytter utvikler seg i løpet av sommeren (Andersen 2007, Pedersen et al. 2014).
Det er fra forvaltning og tarenæring tidligere uttrykt ønske om å utrede tareressursgrunnlaget i områdene nord for Vega i Nordland. Det er dessuten av generell interesse å få oppdatert kunnskap om tilstanden i taresamfunnene og forekomster av kråkeboller i indre og ytre kyststrøk av dette området. Havforskningsinstituttet har derfor i 2018 (Steen et al. 2018b) og 2019 foretatt kartlegginger av tare og kråkebolleforekomster på kyststrekningen mellom Vega og Bodø.
Videoundersøkelsene som i 2019 ble gjennomført på strekningen Herøy – Bodø viste at dekningsgrad og biomassetetthet av stortare økte med økende grad av bølgeeksponering. Stortare dominerte algevegetasjonen på de vestlige stasjoner i samtlige sektorer, mens sukkertare ( Saccharina latissima ) var mer utbredt i østlige, bølgebeskyttede områder. Det ble også registrert spredte innslag av arter som butare ( Alaria esculenta ) og draugtare ( Saccorhiza polyschides ), men forekomstene av disse tareartene viste ingen signifikant trend vis a vis bølgeeksponering. Draugtare har en sørlig utbredelse i Norge, og er tidligere ikke registrert nord for 66° 22’ (Brattegard & Holthe 1997, Steen 2018b, www.artsdatabanken.no ). Draugtare ble i 2019 registrert på fire stasjoner lokalisert langs breddegradsektoren 66° 32’ i Træna og Lurøy, som dermed representerer de nordligste registreringer for denne arten i Norge.
Alderen for stortareplanter samlet inn i Nordland i 2019 varierte fra 2 – 14 år, og gjennomsnittsalderen økte med innsamlingsstasjonenes grad av bølgeeksponering. På stasjoner med moderat til høy bølgeeksponering var gjennomsnittsalderen for innsamlede tareplanter høyere på strekningen Herøy – Bodø, enn for stasjoner lenger sør i Nordland og langs Vestlandskysten. På enkelte av de bølgebeskyttede innsamlingsstasjonene i Nordland hadde imidlertid tareplantene lav gjennomsnittsalder (3-4 år), noe som kan bety at tarevegetasjonen her er i en gjenvekstfase etter tidligere å ha vært nedbeitet av kråkeboller. Det var også en tendens til at tareplantene samlet inn mellom Herøy og Bodø i Nordland var noe kortere enn tareplanter samlet inn lenger sør når prøvetakingsstasjoner der tareplantene hadde tilnærmet lik gjennomsnittsalder sammenlignes. Dette harmonerer med tidligere observasjoner av at stilkstørrelsene hos stortare reduseres fra midt Norge (63°N - 65°N) og nordover for sammenlignbare aldersgrupper (Rinde & Sjøtun 2005).
Størrelsen av stortarestilkenes vekstsoner gir informasjon om tykkelsestilveksten i et gitt år, og kan brukes til å rekonstruere plantenes veksthistorikk gjennom hele livsløpet. Vekstsoneanalyser av tarestilkenes tverrsnitt viste at tykkelsestilveksten varierte med tarestilkenes alder og nådde et maksimum når plantene var ca 3-4 år gamle, for deretter å gradvis avta med økende alder. Tidligere studier i Nordland har vist at tykkelsestilveksten hos stortare øker med grad av bølgeeksponering (Steen et. al. 2015b), noe som også ble registrert i 2019. Den årlige tilvekstraten for de yngste aldersgruppene var lavere for tareplanter samlet inn på kyststrekningen Herøy-Bodø, enn for tareplanter samlet inn lenger sør i Nordland og på vestlandskysten. Dette kan være indikasjoner på at stortarevegetasjonen har en langsommere tilvekst i nordlige områder, og at gjenveksten etter en eventuell tarehøsting dermed vil kreve mer tid her enn i områdene lenger sør. Regionale forskjeller i alder, størrelse og vekst av stortare langs norskekysten har tidligere blitt relatert til geografiske variasjoner i temperatur og lysforhold (Rinde & Sjøtun 2005), men regionale genetiske variasjoner hos stortare (Evankow et al. 2019) kan også spille en rolle. En skal heller ikke se bort ifra at variasjoner i innsamlingsperiode mellom de ulike regioner kan ha bidratt noe til de geografiske forskjellene som ble observert. Tareprøvene på kyststrekningen Agder – Møre og Romsdal ble samlet inn tidligere på året (april) enn tareprøvene fra Trøndelag – Nordland (juni), og kan derfor være noe underestimert mhp det siste års stilktilvekst, men dog ikke tidligere års stilktilvekst.
Tap av tareskog som følge av beiting av kråkeboller er et fenomen som er rapportert fra flere kystområder rundt om i verden (Filbee-Dexter & Scheibling 2014, 2017, Ling et al. 2015). Langs store deler av kysten i Nord-Norge ble tareskogen beitet ned av grønn kråkebolle ( S. droebachiensis ) tidlig på 1970-tallet (Sivertsen & Bjørge 1980, Sivertsen 1982, 1997, Sakshaug & Sjøtun 2002). I de senere år er det imidlertid observert lite grønn kråkebolle i sørlige deler av Nordland (Fagerli et al. 2013, 2015, Norderhaug & Christie 2009, Steen et al. 2014, 2015ab, 2016b, 2018ab). Selv om forekomstene av grønn kråkebolle har gått tilbake, er det fortsatt observert høye populasjonstettheter av denne arten i områder rundt Vega (Rinde et al. 2014) og i Brønnøysund (Steen et al. 2016b).
I 2019 ble det til sammen gjennomført undersøkelser langs 83 videotransekt i Nordland, innenfor et område som strekker seg fra Horsvær i Sømna til Landego/Helligvær i Bodø. Langs disse videotransektene ble det ikke registrert et eneste individ av grønn kråkebolle, mens det til sammen ble registrert 1541 individer av rød kråkebolle ( E. esculentus ). Det ble derimot gjort observasjoner av små juvenile individer av grønn kråkebolle i de innsamlede tareplantenes festeorgan (hapter) på 21 av 22 innsamlingsstasjoner i Nordland i 2019. Forekomstene av hapterkråkeboller var høyest på de nordligste innsamlingsstasjonene (fra Lurøy til Bodø), og registreringsfrekvensen var her mer enn 10 ganger høyere enn på innsamlingsstasjoner lenger sør i Nordland. Høyere forekomster av juvenile grønne kråkeboller i festeorganene hos stortare kan være indikasjoner på at arten har bedre rekrutteringsvilkår i nordlige områder, og dermed utgjør en større trussel for taresamfunnene her, enn i sørlige områder.
Rød kråkebolle utgjør normalt ikke samme trussel overfor tareskogene som grønn kråkebolle, og opptrer vanligvis i lavere tettheter. I deler av Sør-Trøndelag, bl.a øst av Frøya og sør på Fosen-halvøya, er det i de senere år observert høye tettheter av rød kråkebolle som sannsynligvis har gitt beiteskader på tarevegetasjonen og ført til stenging av enkelte tarehøstefelt i disse områdene (Steen 2011, 2013, 2014, 2016, 2018). Forekomstene av røde kråkeboller ( Echinus esculentus) var lav på de fleste videostasjoner undersøkt i Nordland i 2019 med en gjennomsnittlig registreringsfrekvens på ca 0,1 individ per transektmeter. Det ble i 2019 ikke registrert noen økning i kråkebolleforekomstene fra tidligere år og det er heller ingen indikasjoner på at prøvehøstingene av tare i Nordland har ført til økte forekomster av kråkeboller. Den gode rekoloniseringen av tare på høsteflatene i Nordland indikerer også at kråkebollebeiting i liten grad har påvirket vegetasjonsutviklingen etter prøvehøstingene i denne regionen.
5 - Konklusjoner
Undersøkelser etter prøvehøstinger av stortare på kyststrekningen fra Trøndelagsgrensen til Vega i Nordland i perioden 2013 - 2019 har demonstrert en rask rekolonisering av stortare på høsteflatene. Mens reetableringen av stortarebiomassen tok fire år, tok reetableringen av tarevegetasjonens rekrutt- og epifyttforekomster seks år. Kråkebollebeiting ser ikke ut til å ha påvirket reetableringen av tare etter prøvehøstingene i Nordland.
På bakgrunn av resultatene fra prøvehøstingsstudiene anser Havforskningsinstituttet det som forsvarlig at man åpner de ytre, vestlige deler (dvs områdene vest for lengdegrad 11 ° 45’, med unntak av verne- og referanseområder) av strekningen mellom Trøndelagsgrensen og Vega (sør for breddegrad 65 ° 33’) i Nordland for kommersiell tarehøsting (Steen et al. 2018b). De indre, østlige kystområdene (øst for lengdegrad 11° 45’) på strekningen mellom Trøndelagsgrensen og Vega vurderes som lite egnet for høsting i industriell målestokk da stortarevegetasjonen her er lite utviklet både med hensyn på utbredelse og størrelse (Steen et al. 2018ab). For å gi tareøkosystemene mulighet til å reetablere seg mellom høstingene i henhold til målsetning om økosystembasert forvaltning, bør feltene i sørlig del av Nordland ha en hvileperiode på minst seks år mellom påfølgende høstinger, dvs en høstesyklus på minst syv år. Vekstsoneanalyser tyder også på at den årlige stilveksten for eldre tareplanter er høyere i Nordland, enn for tilsvarende aldersgrupper lenger sør. En høstesyklus på 7 år kan derfor gi et bedre ressursutbytte i Nordland, enn hva tilfellet ville vært for tarehøstefeltene lenger sør langs vestlandskysten.
Høstefeltene bør inndeles etter et mønster som sikrer at nabofelt ikke høstes i påfølgende sesonger slik at tarevegetasjonen ikke reduseres over større sammenhengende arealer i løpet av kort tid. En alternativ forvaltningsstrategi som kombinerer best mulig ressursutnyttelse samtidig som tareskogens økosystemfunksjonalitet opprettholdes, kan være å avsette definerte områder med godt ressursgrunnlag til tarehøsting med en høstesyklus som optimaliserer ressursutbyttet («tareindustriområder») samtidig som høsting ikke tillates i andre områder der tareskogen får stå urørt i en naturtilstand («tareurskogsområder»). Uansett hvilken forvaltningsstrategi man velger så bør omfanget av tarehøstingsaktiviteten rapporteres og tilstanden på høstefeltene overvåkes regelmessig.
En kartlegging av tareressursene og kråkebolleforekomstene på kyststrekningen fra Herøy til Bodø indikerer et variabelt ressurspotensial for stortarehøsting, begrenset til de ytre, vestlige kystområdene. For å få en bedre oversikt over utbredelsen og stabiliteten av tareforekomstene i denne delen av Nordland bør det imidlertid gjennomføres mer detaljerte kartlegginger over tid. Analyser av innsamlede stortareplanter på kyststrekningen Herøy - Bodø tyder på at stilktilveksten er langsommere og at grønne kråkeboller har bedre rekrutteringsvilkår her, enn i områder lenger sør. En eventuell framtidig kommersiell høsting av tareressursene i denne regionen vil derfor kunne kreve en annen forvaltningsmodell enn i sørligere kyststrøk, og utformingen av denne bør baseres på kunnskap fra lokale prøvehøstingsstudier.
Det er videre behov for å beregne størrelsen på de utnyttbare tareressursene i denne delen av landet. Uttaket av stortare bør reguleres innenfor bærekraftige rammer basert på kvantitativ kunnskap om hvor store tareressurser som er tilgjengelige i hvert høstefelt. Det bør også beregnes hvor store områder som fortsatt er beitet ned av kråkeboller og hvordan de romlige endringene for beiting og tarebiomasse skjer over tid.
6 - Referanser
Andersen G. 2007. Kelp associated floral epiphytes - Productivity and community structure in relation to wave exposure and season. Master of Science Thesis. University of Oslo
Artsdatabanken. Artskart. https://artskart.artsdatabanken.no/app/ .
Bates DM, Maechler M, Bolker B, Walker S. 2015. Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software, 67 (1), 1-48. <doi:10.18637/jss.v067.i01>.
Bekkby T, Rinde E, Gundersen G, Norderhaug KM, Gitmark JK, Christie H. 2014a. Length, strength and water flow: relative importance of wave and current exposure on morphology in kelp Laminaria hyperborea . Marine Ecology Progress Series. 506: 61–70.
Bekkby T, Angeltveit G, Gundersen H, Tveiten L, Norderhaug KM. 2014b. Red sea urchins ( Echinus esculentus ) and water flow influence epiphytic macroalgae density. Marine Biology Research. DOI: 10.1080/17451000.2014.943239.
Brattegard T, Holthe T. 2001. Distribution of marine, benthic macroorganisms in Norway. A tabulated catalogue. Preliminary edition. Directorate for nature management. Research Report 2001(3) Trondheim: 394 pp.
Buschmann AH, Prescott S, Potin P, Faugeron S, Vasquez JA, Camus C, Infante J, Hernandez-Gonzalez MC, Gutierrez A, Varela DA. 2014. The status of kelp exploitation and marine agronomy, with emphasis on Macrocystis pyrifera , in Chile. Advances in Botanical Research. 71: 161–188.
Christie H, Fredriksen S, Rinde E. 1998. Regrowth of kelp and colonization of epiphyte and fauna community after kelp trawling at the coast of Norway. Hydrobiologia 375 (376): 49–58.
Christie H, Jørgensen, NM, Norderhaug KM, Waage-Nielsen E. 2003. Species distribution and habitat exploitation of fauna associated with kelp ( Laminaria hyperborea ) along the Norwegian coast. Journal of the Marine Biological Association of the United Kingdom 83: 687–699.
Christie H, Jørgensen NM, Norderhaug KM. 2007. Bushy or smooth, high or low; importance of habitat architecture and vertical position for distribution of fauna on kelp. Journal of Sea Research 58: 198–208.
Evankow A, Christie H, Hancke K, Brysting AK, Junge C, Fredriksen S, Taulow J. 2019. Genetic heterogeneity of two bioeconomically important kelp species along the Norwegian coast. Conservation Genetics 20: 615-628.
Fagerli CW, Norderhaug KM, Christie H. 2013. Lack of sea urchin settlement may explain kelp forest recovery in overgrazed areas in Norway. Marine Ecology Progress Series. 488: 119–132.
Fagerli CW, Stadniczeñko SG, Pedersen MF, Christie H, Fredriksen S, Norderhaug KM. 2015. Population dynamics of Strongylocentrotus droebachiensis in kelp forests and barren grounds in Norway. Marine Biology. 162: 1215–1226.
Filbee-Dexter K, Scheibling RE. 2014. Sea urchin barrens as alternative stable states of collapsed kelp ecosystems. Marine Ecology Progress Series. 495: 1–25.
Filbee-Dexter K, Scheibling RE. 2017. The present is the key to the past: linking regime shifts in kelp beds to the distribution of deep-living sea urchins. Ecology. 98(1): 253–264.
Ling SD, Scheibling RE, Rassweiler A, Johnson CR, Shears N, Connell SD, Salomon AK, Norderhaug KM, Pérez-Matus A, Hernández JC, Clemente S, Blamey LK, Hereu B, Ballesteros E, Sala E, Garrabou J, Cebrian E, Zabala1 M, Fujita D, Johnson LE. 2015. Global regime shift dynamics of catastrophic sea urchin overgrazing. Philosophical Transactions Royal Society B. 370: 20130269.
Mac Monagail M, Cornish L, Morrison L, Araújo R, Critchley AT. 2017. Sustainable harvesting of wild seaweed resources. European Journal of Phycology. 52: 371-390.
Norderhaug KM, Fredriksen S, Nygaard K. 2003. Trophic importance of Laminaria hyperborea to kelp forest consumers and the importance of bacterial degradation to food quality. Marine Ecology Progress Series 255: 135–144.
Norderhaug KM., Christie H. 2009. Sea urchin grazing and kelp re-vegetation in the NE Atlantic. Marine Biology Research 5: 515-528.
Norderhaug KM., Christie H. 2011. Secondary production in a Laminaria hyperborea kelp forest and variation according to wave exposure. Estuarine, Coastal and Shelf Science 95: 135-144.
Norderhaug KM, Christie H, Andersen GS, Bekkby T. 2012. Does the diversity of kelp forest macrofauna increase with wave exposure? Journal of Sea Research 69: 36–42.
Norsk Standard 9415. 2009. Flytende oppdrettsanlegg. Krav til utforming, dimensjonering, utførelse, installasjon og drift. ICS 65.150; 67.260.
Pedersen MF, Nejrup LB, Fredriksen S, Christie H, Norderhaug KM. 2012. Effects of wave exposure on population structure, demography, biomass and productivity of the kelp Laminaria hyperborea . Marine Ecology Progress Series. 451: 45–60.
Pedersen MF, Nejrup LB, Pedersen TM, Fredriksen S. 2014. Sub-canopy light conditions only allow low annual net productivity of epiphytic algae on kelp Laminaria hyperborea . Marine Ecology Progress Series. 516: 163-176.
Rinde E, Sjøtun K. 2005. Demographic variation in the kelp Laminaria hyperborea along a latitudinal gradient. Marine Biology, 146: 1051–1062.
Rinde E, Christie H, Fagerli CW, Bekkby T, Gundersen H, Norderhaug KM, Hjermann DØ. 2014. The Influence of Physical Factors on Kelp and Sea Urchin Distribution in Previously and Still Grazed Areas in the NE Atlantic. PLoS ONE 9(6): e100222. https://doi.org/10.1371/journal.pone.0100222 .
Sakshaug E, Sjøtun K. 2002. Nedbeiting av tareskog i Norge. Rapport fra arbeidsgruppe nedsatt av Fiskeridepartementet og Miljøverndepartementet. 1-47.
Sivertsen K, Bjørge A. 1980. Reduksjon av tareskogen på Helgelandskysten. Fisken og Havet 4: 1-9.
Sivertsen K. 1982. Utbredelse og variasjon i kråkebollebeiting av tareskogen på vestkysten av Norge. - NF-Rapport nr. 7/82. Nordlandsforskning. 1-31.
Sivertsen K. 1997. Geographic and environmental factors affecting the distribution of kelp beds and barren grounds and changes in biota associated with kelp reduction at sites along the Norwegian coast. Canadian Journal of Fisheries and Aquatic Sciences. 54: 2872–2887.
Sivertsen K. 2006. Overgrazing of kelp beds along the coast of Norway. Journal of Applied Phycology 18: 599-610.
Sjøtun K, Fredriksen S. 1995. Growth allocation in Laminaria hyperborea (Laminariales, Phaeophyceae) in relation to age and wave exposure. Marine Ecology Progress Series 126: 213-222.
Sjøtun K, Fredriksen S, Rueness J. 1998. Effect of canopy biomass and wave exposure on growth in Laminaria hyperborea (Laminariaceae: Phaeophyta). European Journal of Phycology 33: 337-343.
Sjøtun K, Christie H, Fosså JH. 2006. Effects of kelp recruitment and sea urchin grazing on stability in kelp forest ( Laminaria hyperborea ). Marine Biology Research 2:24-32.
Smale DA, Burrows MT, Moore P, O’Connor N, Hawkins SJ. 2013. Threats and knowledge gaps for ecosystem services provided by kelp forests: a northeast Atlantic perspective. Ecology and Evolution 3: 4016–4038.
Smale DA, Vance T. 2015. Climate-driven shifts in species distributions may exacerbate the impacts of storm disturbances on northeast Atlantic kelp forests. Marine and Freshwater Research 67 (1): 65-74.
Smale DA, Burrows MT, Evans AJ, King N, Sayer MDJ, Yunnie ALE, Moore P. 2016. Linking environmental variables with regional scale variability in ecological structure and standing stock of carbon within UK kelp forests. Marine Ecology Progress Series 542: 79–95.
Steen H. 2011. Undersøkelser av A-felt for tarehøsting i Møre og Romsdal og Sør-Trøndelag i 2011. Rapport fra Havforskningen Nr. 13-2011.
Steen H. 2013. Undersøkelser av C-felt for tarehøsting i Møre og Romsdal og Sør-Trøndelag i 2013. Rapport fra Havforskningen Nr. 26-2013.
Steen H. 2014. Undersøkelser av D-felt for tarehøsting i Møre og Romsdal og Sør-Trøndelag i 2014. Rapport fra Havforskningen Nr. 24-2014.
Steen H. 2016. Undersøkelser av A-høstefelt for tare i Sør-Trøndelag og Nord-Trøndelag i 2016. Rapport fra Havforskningen. Nr. 31-2016.
Steen H. 2018. Tilstandsvurdering av C-felt for tarehøsting i Sør-Trøndelag og Nord-Trøndelag i 2018. Nr. 32-2018.
Steen H, Moy FE, Bodvin T. 2014a. Undersøkelser av stortarehøsting i Nord-Trøndelag og Nordland i 2013. Fisken og Havet 2014-3.
Steen H, Moy FE, Bodvin T. 2014b. Prøvehøsting av stortare i Nord-Trøndelag – Gjenvekstundersøkelser 2010-2014. Rapport fra Havforskningen Nr. 37-2014.
Steen H, Husa V, Moy FE, Bodvin T, Øverbø Hansen H, Sannæs H, Bosgraaf S. 2015a. Undersøkelser av stortarehøsting i Nordland i 2014. Rapport fra Havforskningen Nr. 1-2015.
Steen H, Bodvin T, Moy FE, Sannes H, Øverbø Hansen H. 2015b. Undersøkelser av stortarehøsting i Nordland i 2015. Rapport fra Havforskningen Nr. 26-2015.
Steen H, Moy FE, Bodvin T, Husa V. 2016a. Regrowth after kelp harvesting in Nord-Trøndelag, Norway. ICES Journal of Marine Science. 73(10): 2708–2720.
Steen H, Bodvin T, Moy FE, Gustad E, Øverbø Hansen H, Jelmert A, Baardsen P. 2016b. Effekter av stortarehøsting i Nordland i 2016. Rapport fra Havforskningen Nr. 38-2016.
Steen H, Norderhaug KM, Moy FE. 2018a. Tareundersøkelser i Nordland i 2017. Rapport fra Havforskningen Nr. 9-2018.
Steen H, Norderhaug KM, Moy FE. 2018b. Tareundersøkelser i Nordland i 2018. Rapport fra Havforskningen Nr. 44-2018.
Teagle H, Hawkins SJ, Moore PJ, Smale DA. 2017. The role of kelp species as biogenic habitat formers in coastal marine ecosystems. Journal of Experimental Marine Biology and Ecology. 492: 81-98.
7 - Appendix
Appendix 1. Posisjoner (WGS84 i grader,desimaler) på innsamlingsstasjoner for stortare på kyststrekningen Flekkefjord – Bodø i april - juni 2019.
| Midlere | ||||||||
| Innsamlings | Dyp | bølgehøyde | Planter | |||||
| Fylke | Høstestatus | Dato | Kl. | Breddegrad | Lengdegrad | (m) | (m) | (antall) |
| Agder | Ikke høstet | 30.04.2019 | 10:10 | 58,2146°N | 6,5296°Ø | 9 | 0,9 | 10 |
| Rogaland | Ikke høstet | 29.04.2019 | 10:15 | 58,7555°N | 5,47291°Ø | 8 | 1,36 | 10 |
| Rogaland | Ikke høstet | 28.04.2019 | 17:45 | 58,8959°N | 5,4629°Ø | 9 | 1,06 | 10 |
| Rogaland | Ikke høstet | 28.04.2019 | 08:45 | 59,2340°N | 5,06461°Ø | 8 | 1,21 | 10 |
| Vestlandet | Ikke høstet | 27.04.2019 | 09:30 | 60,168°N | 4,98433°Ø | 9 | 0,9 | 10 |
| Vestlandet | Ikke høstet | 26.04.2019 | 15:20 | 60,6385°N | 4,72458°Ø | 8 | 1,45 | 10 |
| Vestlandet | Ikke høstet | 26.04.2019 | 09:40 | 60,9289°N | 4,6149°Ø | 8 | 2,2 | 10 |
| Vestlandet | Ikke høstet | 24.04.2019 | 18:55 | 61,2726°N | 4,72726°Ø | 7 | 0,84 | 10 |
| Vestlandet | Ikke høstet | 23.04.2019 | 15:00 | 61,8348°N | 4,9625°Ø | 9 | 0,25 | 10 |
| Møre og Romsdal | Ikke høstet | 20.04.2019 | 16:30 | 62,405°N | 5,64366°Ø | 7 | 0,71 | 10 |
| Møre og Romsdal | Ikke høstet | 20.04.2019 | 13:00 | 62,5531°N | 5,95116°Ø | 10 | 0,87 | 10 |
| Møre og Romsdal | Ikke høstet | 19.04.2019 | 17:15 | 63,0975°N | 7,25291°Ø | 10 | 0,78 | 10 |
| Trøndelag | Ikke høstet | 07.06.2019 | 17:45 | 65,1322°N | 11,4984°Ø | 8 | 0,57 | 10 |
| Nordland | Ikke høstet | 09.06.2019 | 17:50 | 65,3866°N | 11,6452°Ø | 8 | 0,73 | 10 |
| Nordland | Ikke høstet | 08.06.2019 | 15:30 | 65,3889°N | 11,6102°Ø | 9 | 0,83 | 10 |
| Nordland | Ikke høstet | 08.06.2019 | 14:10 | 65,4046°N | 11,5345°Ø | 8 | 0,8 | 10 |
| Nordland | Høstet i 2014 | 09.06.2019 | 16:50 | 65,4197°N | 11,7154°Ø | 7 | 0,63 | 10 |
| Nordland | Høstet i 2014 | 09.06.2019 | 16:00 | 65,4257°N | 11,687°Ø | 7 | 0,57 | 10 |
| Nordland | Høstet i 2014 | 09.06.2019 | 15:30 | 65,4321°N | 11,6760°Ø | 7 | 0,66 | 10 |
| Nordland | Høstet i 2013 | 11.06.2019 | 12:50 | 65,536°N | 11,4223°Ø | 9 | 1,35 | 11 |
| Nordland | Ikke høstet | 12.06.2019 | 11:00 | 66,0444°N | 11,7146°Ø | 9 | 1,21 | 11 |
| Nordland | Ikke høstet | 12.06.2019 | 11:30 | 66,0522°N | 11,7777°Ø | 10 | 1,11 | 10 |
| Nordland | Ikke høstet | 13.06.2019 | 13:00 | 66,3536°N | 12,2883°Ø | 8 | 0,61 | 10 |
| Nordland | Ikke høstet | 13.06.2019 | 12:30 | 66,3752°N | 12,2957°Ø | 8 | 0,54 | 12 |
| Nordland | Ikke høstet | 15.06.2019 | 13:15 | 66,5370°N | 12,0825°Ø | 9 | 0,51 | 10 |
| Nordland | Ikke høstet | 15.06.2019 | 14:00 | 66,5615°N | 12,0658°Ø | 10 | 0,84 | 10 |
| Nordland | Ikke høstet | 16.06.2019 | 15:30 | 66,7015°N | 13,1102°Ø | 7 | 0,31 | 12 |
| Nordland | Ikke høstet | 16.06.2019 | 10:15 | 66,7105°N | 12,4804°Ø | 10 | 0,81 | 10 |
| Nordland | Ikke høstet | 17.06.2019 | 12:45 | 66,8692°N | 13,3302°Ø | 9 | 0,48 | 8 |
| Nordland | Ikke høstet | 17.06.2019 | 13:10 | 66,8713°N | 13,3769°Ø | 8 | 0,32 | 12 |
| Nordland | Ikke høstet | 18.06.2019 | 10:30 | 67,0363°N | 13,2250°Ø | 9 | 0,96 | 9 |
| Nordland | Ikke høstet | 18.06.2019 | 14:15 | 67,0441°N | 13,7467°Ø | 7 | 0,41 | 10 |
| Nordland | Ikke høstet | 20.06.2019 | 10:45 | 67,2034°N | 13,7407°Ø | 12 | 0,51 | 11 |
| Nordland | Ikke høstet | 19.06.2019 | 09:45 | 67,3744°N | 14,4802°Ø | 6 | 0,49 | 10 |
| Nordland | Ikke høstet | 19.06.2019 | 14:00 | 67,3791°N | 13,9013°Ø | 10 | 0,62 | 10 |
Appendix 2. Start og stopposisjoner (WGS84 i grader,desimaler) for videotransekt filmet i juni 2019 på strekningen Sømna - Vega i Nordland.
| Transekt | Transekt | Transekt | Transekt | Gj.sn | Midlere | |||||
| Tarehøstet | Start | Start | Stopp | Stopp | Dyp | bølgehøyde | ||||
| Felt | Stasjon | (år) | Dato | Kl. | Breddegrad | Lengdegrad | Breddegrad | Lengdegrad | (m) | (m) |
| PF3 | A | 2013 | 11.09.2019 | 12:25 | 65,5352°N | 11,0382°Ø | 65,5368°N | 11,0396°Ø | 11 | 1,35 |
| PF3 | B | 2013 | 11.09.2019 | 13:15 | 65,55°N | 11,0418°Ø | 65,5483°N | 11,0433°Ø | 9 | 1,20 |
| PF3 | C | 2013 | 11.09.2019 | 13:38 | 65,5393°N | 11,0468°Ø | 65,5373°N | 11,0494°Ø | 10 | 1,19 |
| PF3 | D | 2013 | 11.09.2019 | 14:15 | 65,5492°N | 11,0641°Ø | 65,5479°N | 11,0647°Ø | 10 | 1,01 |
| PF3 | I | 2013 | 09.06.2019 | 08:52 | 65,547°N | 12,0029°Ø | 65,5459°N | 12,0013°Ø | 8 | 0,36 |
| PF5 | F | 2014 | 09.06.2019 | 16:33 | 65,4194°N | 11,0804°Ø | 65,4203°N | 11,0830°Ø | 8 | 0,63 |
| PF5 | G | 2014 | 09.06.2019 | 15:42 | 65,4255°N | 11,0685°Ø | 65,426°N | 11,0721°Ø | 10 | 0,57 |
| PF5 | H | 2014 | 09.06.2019 | 15:08 | 65,4317°N | 11,0769°Ø | 65,4331°N | 11,0734°Ø | 9 | 0,66 |
| PF6 | D | 2014 | 09.06.2019 | 11:53 | 65,4855°N | 11,0546°Ø | 65,4862°N | 11,0583°Ø | 9 | 0,68 |
| PF6 | E | 2014 | 09.06.2019 | 12:18 | 65,4922°N | 11,0635°Ø | 65,4940°N | 11,0639°Ø | 7 | 0,60 |
| PF6 | F | 2014 | 11.06.2019 | 10:15 | 65,4974°N | 11,0502°Ø | 65,4994°N | 11,0662°Ø | 9 | 0,87 |
| PF6 | G | 2014 | 11.06.2019 | 10:00 | 65,4857°N | 11,0608°Ø | 65,4864°N | 11,0608°Ø | 14 | 1,04 |
| PF7 | A | 2014 | 11.06.2019 | 11:55 | 65,5287°N | 11,4602°Ø | 65,5298°N | 11,4580°Ø | 10 | 1,06 |
| PF7 | B | 2014 | 11.06.2019 | 11:26 | 65,5177°N | 11,4642°Ø | 65,5185°N | 11,4614°Ø | 12 | 1,20 |
| PF7 | C | 2014 | 11.06.2019 | 10:49 | 65,5227°N | 11,5210°Ø | 65,5229°N | 11,5181°Ø | 12 | 1,10 |
| PF7 | D | 2014 | 09.06.2019 | 12:55 | 65,5186°N | 11,6229°Ø | 65,5191°N | 11,6261°Ø | 7 | 0,60 |
| PF19 | A | 2017 | 08.06.2019 | 09:33 | 65,3363°N | 11,5784°Ø | 65,3369°N | 11,5744°Ø | 7 | 0,85 |
| PF19 | B | 2017 | 08.06.2019 | 11:24 | 65,3599πN | 11,5418°Ø | 65,3621°N | 11,5418°Ø | 8 | 0,89 |
| PF19 | C | 2017 | 08.06.2019 | 10:55 | 65,3570πN | 11,4925°Ø | 65,3586°N | 11,4915°Ø | 7 | 0,94 |
| PF19 | D | 2017 | 08.06.2019 | 10:23 | 65,3541πN | 11,4378°Ø | 65,3551°N | 11,4404°Ø | 10 | 1,15 |
| PF20 | A | 2017 | 08.06.2019 | 13:24 | 65,4046πN | 11,5126°Ø | 65,4059°N | 11,5161°Ø | 9 | 0,89 |
| PF20 | B | 2017 | 08.06.2019 | 14:39 | 65,4022πN | 11,5816°Ø | 65,4036°N | 11,5837°Ø | 11 | 0,78 |
| PF21 | A | 2017 | 09.06.2019 | 11:26 | 65,4803πN | 11,6799°Ø | 65,4814°N | 11,6820°Ø | 7 | 0,49 |
| PF21 | B | 2017 | 09.06.2019 | 14:16 | 65,4755πN | 11,6179°Ø | 65,4762°N | 11,6065°Ø | 9 | 0,73 |
| PF21 | C | 2017 | 11.09.2019 | 09:45 | 65,4792πN | 11,5412°Ø | 65,4804°N | 11,5395°Ø | 11 | 0,93 |
| Ref2 | C | Ikke høstet | 08.06.2019 | 12:02 | 65,3697πN | 11,5735°Ø | 65,3725°N | 11,5729°Ø | 9 | 0,85 |
| Ref2 | D | Ikke høstet | 08.06.2019 | 12:35 | 65,3807πN | 11,554°Ø | 65,3818°N | 11,5552°Ø | 9 | 0,85 |
| Ref3 | A | Ikke høstet | 09.06.2019 | 09:32 | 65,5333πN | 12,0729°Ø | 65,5320°N | 12,0711°Ø | 8 | 0,39 |
| Ref3 | B | Ikke høstet | 09.06.2019 | 09:11 | 65,5335πN | 12,0817°Ø | 65,5327°N | 12,0803°Ø | 6 | 0,30 |
| Ref5 | A | Ikke høstet | 08.06.2019 | 13:52 | 65,4036πN | 11,5314°Ø | 65,4047°N | 11,5344°Ø | 8 | 0,85 |
| Ref5 | B | Ikke høstet | 08.06.2019 | 14:20 | 65,4019πN | 11,5525°Ø | 65,4028°N | 11,5556°Ø | 9 | 0,65 |
| Ref6 | C | Ikke høstet | 08.06.2019 | 15:17 | 65,3885πN | 11,6100°Ø | 65,3899°N | 11,6127°Ø | 11 | 0,83 |
| Ref6 | D | Ikke høstet | 09.06.2019 | 17:32 | 65,3860πN | 11,6474°Ø | 65,3869°N | 11,6443°Ø | 10 | 0,73 |
| Ref7 | A | Ikke høstet | 09.06.2019 | 13:27 | 65,5276πN | 11,6056°Ø | 65,5277°N | 11,6086°Ø | 8 | 0,77 |
| Ref7 | B | Ikke høstet | 09.06.2019 | 13:16 | 65,5234πN | 11,6012°Ø | 65,5239°N | 11,6021°Ø | 13 | 0,66 |
Appendix 3. Start og stopposisjoner (WGS84 i grader,desimaler) for videotransekt filmet i 2019 på kyststrekningen Herøy - Bodø i Nordland.
| Transekt | Transekt | Transekt | Transekt | Gj.sn | Midlere | ||
| Start | Start | Stopp | Stopp | Dyp | bølgehøyde | ||
| Dato | Kl. | Breddegrad | Lengdegrad | Breddegrad | Lengdegrad | (m) | (m) |
| 12.06.2019 | 10:03 | 66,0399°N | 11,0667°Ø | 66,0383°N | 11,0674°Ø | 11 | 1,53 |
| 12.06.2019 | 10:33 | 66,0447°N | 11,0818°Ø | 66,0436°N | 11,0777°Ø | 10 | 1,19 |
| 12.06.2019 | 09:09 | 66,0499°N | 11,0715°Ø | 66,0485°N | 11,0694°Ø | 12 | 1,08 |
| 12.06.2019 | 09:26 | 66,0435°N | 11,0732°Ø | 66,042°N | 11,0713°Ø | 10 | 0,97 |
| 12.06.2019 | 08:51 | 66,0528°N | 11,0785°Ø | 66,0517°N | 11,0766°Ø | 11 | 1,10 |
| 12.06.2019 | 12:04 | 66,0578°N | 11,0963°Ø | 66,0570°N | 11,0941°Ø | 11 | 0,96 |
| 13.06.2019 | 10:35 | 66,3833°N | 12,0274°Ø | 66,3823°N | 12,0269°Ø | 10 | 0,37 |
| 13.06.2019 | 10:13 | 66,3766°N | 12,0284°Ø | 66,3759°N | 12,0253°Ø | 13 | 0,42 |
| 13.06.2019 | 12:10 | 66,3756°N | 12,0304°Ø | 66,3737°N | 12,0261°Ø | 10 | 0,55 |
| 13.06.2019 | 11:50 | 66,3766°N | 12,0176°Ø | 66,3759°N | 12,031°Ø | 11 | 0,71 |
| 13.06.2019 | 11:17 | 66,3678°N | 12,0168°Ø | 66,3693°N | 12,0185°Ø | 12 | 0,65 |
| 13.06.2019 | 09:27 | 66,3635°N | 12,0231°Ø | 66,3629°N | 12,0249°Ø | 11 | 0,30 |
| 13.06.2019 | 09:57 | 66,3707°N | 12,3104°Ø | 66,3705°N | 12,3068°Ø | 12 | 0,39 |
| 13.06.2019 | 08:56 | 66,3530°N | 12,2878°Ø | 66,3538°N | 12,2892°Ø | 12 | 0,61 |
| 15.06.2019 | 12:35 | 66,5367°N | 12,0397°Ø | 66,5361°N | 12,0368°Ø | 10 | 0,94 |
| 15.06.2019 | 11:30 | 66,5372°N | 12,0833°Ø | 66,5364°N | 12,0806°Ø | 11 | 0,51 |
| 15.06.2019 | 10:58 | 66,5405°N | 12,1085°Ø | 66,5409°N | 12,1116°Ø | 13 | 0,42 |
| 15.06.2019 | 10:17 | 66,5459°N | 12,1873°Ø | 66,5481°N | 12,1875°Ø | 10 | 0,65 |
| 15.06.2019 | 09:31 | 66,5380°N | 12,2695°Ø | 66,5392°N | 12,2730°Ø | 8 | 0,61 |
| 15.06.2019 | 17:30 | 66,545°N | 12,5626°Ø | 66,5439°N | 12,5618°Ø | 12 | 0,50 |
| 15.06.2019 | 16:55 | 66,5400°N | 12,6335°Ø | 66,5404°N | 12,6303°Ø | 12 | 0,38 |
| 15.06.2019 | 16:24 | 66,5453°N | 12,706°Ø | 66,5457°N | 12,7036°Ø | 10 | 0,42 |
| 15.06.2019 | 15:49 | 66,5339°N | 12,7668°Ø | 66,5350°N | 12,7675°Ø | 10 | 0,47 |
| 15.06.2013 | 13:42 | 66,5620°N | 12,0665°Ø | 66,5610°N | 12,0647°Ø | 12 | 0,85 |
| 15.06.2013 | 12:00 | 66,5531°N | 12,0755°Ø | 66,5512°N | 12,0738°Ø | 12 | 0,52 |
| 16.06.2019 | 09:54 | 66,7108°N | 12,4816°Ø | 66,7102°N | 12,4784°Ø | 14 | 0,81 |
| 16.06.2019 | 11:25 | 66,7033°N | 12,6338°Ø | 66,7025°N | 12,6322°Ø | 14 | 0,54 |
| 16.06.2019 | 13:24 | 66,7079°N | 12,9128°Ø | 66,7077°N | 12,9102°Ø | 11 | 0,59 |
| 16.06.2019 | 13:40 | 66,7019°N | 12,9046°Ø | 66,7017°N | 12,9020°Ø | 14 | 0,54 |
| 16.06.2019 | 15:09 | 66,7021°N | 13,1100°Ø | 66,7007°N | 13,1103°Ø | 10 | 0,30 |
| 17.06.2019 | 12:18 | 66,8685°N | 13,3319°Ø | 66,8693°N | 13,3297°Ø | 11 | 0,48 |
| 17.06.2019 | 11:46 | 66,8712°N | 13,3739°Ø | 66,8711°N | 13,3785°Ø | 11 | 0,33 |
| 17.06.2019 | 10:36 | 66,8806°N | 13,5536°Ø | 66,8811°N | 13,5555°Ø | 13 | 0,34 |
| 17.06.2019 | 09:48 | 66,877°N | 13,6294°Ø | 66,8783°N | 13,6304°Ø | 7 | 0,21 |
| 18.06.2019 | 10:03 | 67,0369°N | 13,2233°Ø | 67,0359°N | 13,2270°Ø | 11 | 0,96 |
| 18.06.2019 | 11:58 | 67,0403°N | 13,5731°Ø | 67,0407°N | 13,5768°Ø | 12 | 0,78 |
| 18.06.2019 | 12:49 | 67,0393°N | 13,6624°Ø | 67,0407°N | 13,6631°Ø | 12 | 0,31 |
| 18.06.2019 | 13:28 | 67,0391°N | 13,7170°Ø | 67,0407°N | 13,7189°Ø | 10 | 0,38 |
| 18.06.2019 | 13:58 | 67,0435°N | 13,7472°Ø | 67,0450°N | 13,7466°Ø | 8 | 0,40 |
| 18.06.2019 | 15:26 | 67,0478°N | 13,9385°Ø | 67,0488°N | 13,9404°Ø | 7 | 0,18 |
| 20.06.2019 | 10:14 | 67,2032°N | 13,7437°Ø | 67,2033°N | 13,7396°Ø | 13 | 0,51 |
| 20.06.2019 | 12:23 | 67,2091°N | 13,9774°Ø | 67,2102°N | 13,9773°Ø | 11 | 0,38 |
| 20.06.2019 | 14:36 | 67,2013°N | 14,3510°Ø | 67,2033°N | 14,3492°Ø | 10 | 0,30 |
| 20.06.2019 | 14:13 | 67,1981°N | 14,3438°Ø | 67,1986°N | 14,3454°Ø | 13 | 0,34 |
| 19.06.2019 | 13:45 | 67,3784°N | 13,9007°Ø | 67,3799°N | 13,9020°Ø | 11 | 0,62 |
| 19.06.2019 | 11:54 | 67,3809°N | 14,1930°Ø | 67,3808°N | 14,1970°Ø | 10 | 0,52 |
| 19.06.2019 | 11:09 | 67,3729°N | 14,2738°Ø | 67,3735°N | 14,2756°Ø | 11 | 0,33 |
| 19.06.2019 | 10:37 | 67,3764°N | 14,3226°Ø | 67,3776°N | 14,3267°Ø | 9 | 0,37 |
| 19.06.2019 | 09:26 | 67,3734°N | 14,4794°Ø | 67,3754°N | 14,4812°Ø | 7 | 0,48 |