The report is a hearing document sent to the Ministry of Petroleum and Energy regarding a proposed program for environmental assessment connected to extraction of minerals on the Norwegian continental shelf areas. In the hearing document, Institute of Marine Research emphasize the extensive lack of knowledge, both with respect to biodiversity and ecological aspects, but also regarding the mining technology for extraction of minerals and metals from the seabed. This lack of knowledge represents an uncertainty that need to be visualised in the environmental assessment, e.g. through a risk assessment. Institute of Marine Research also addresses that different ongoing initiatives for building a knowledge base should have been finalized before initiation of the present hearing. A more thoroughly summary of the report content is given in the first chapter.
Innspill til høring - Forslag til program for konsekvensutredning for mineralvirksomhet på norsk sokkel
Rapportserie:
Rapport fra havforskningen 2021-23
ISSN: 1893-4536
Publisert: 27.05.2021
Prosjektnr: 14384-01
Tema:
Gruveavfall
Program:
Nordsjøen
Godkjent av:
Forskningsdirektør(er):
Geir Huse
Programleder(e):
Henning Wehde
English summary
Sammendrag
Rapporten er et høringssvar sendt til Olje- og Energidepartementet i forbindelse med et forslag til konsekvensutredningsprogram for mineralvirksomhet på norsk kontinentalsokkel. I høringssvaret påpeker Havforskningsinstituttet at det er stor mangel på kunnskap, både med hensyn til biomangfold og økologiske forhold, men også når det gjelder teknologien som vil anvendes for utvinning av mineraler og metaller på havbunnen. Denne kunnskapsmangelen representerer en usikkerhet som bør synliggjøres i konsekvensutredningen, blant annet gjennom en risikoanalyse. Havforskningsinstituttet påpeker også at ulike initiativ når det gjelder kunnskapsinnhenting burde vært ferdigstilt før gjennomføringen av høringen. En grundigere oppsummering av rapportens innhold er gitt i første kapittel.
1 - Oppsummering
En konsekvensutredning etter havbunnsmineralloven for åpning av undersøkelse og utvinning av havbunnsmineraler på norsk sokkel bør være forankret i økosystembasert forvaltning etter prinsipper som blant annet er nedfelt i den internasjonale havbunnsmyndigheten (ISA) sine anbefalinger. Dette vil innbefatte etablering av et solid kunnskapsgrunnlag gjennom baselinjestudier, samt vurdere tiltak for å ivareta økosystemenes integritet, biodiversitet og evne til restitusjon etter eventuelle inngrep. Økosystemelementer som dynamikk, utbredelse, skalering, artsmangfold, sårbarhet, utveksling mellom populasjoner og genetiske ressurser vil være sentrale her, og føre-var-prinsippet bør anvendes ved kunnskapsmangel.
Det er stor kunnskapsmangel om økosystemene i utredningsområdet, både i vannmassene og spesielt i de dypere bunnområdene. Store dyp i kombinasjon med kompleksitet i bunnområder og vannmasser tilsier at utredningsområdet kan inneholde en rekke nye og uoppdagede arter. Toleranse for ulike påvirkninger er i stor grad ukjent for dyptlevende arter. Innhenting av ny kunnskap vil ta lang tid, blant annet fordi utredningsområdet er svært stort, og identifisering av arter fra bunnprøver vil være meget tid- og ressurskrevende. Usikkerheten som denne kunnskapsmangelen representerer, bør vektlegges og få en vesentlig plass i konsekvensutredningen, og en risikoanalyse for biologisk mangfold og økosystemtjenester bør gjennomføres etter nye metoder som synliggjør denne kunnskapsmangelen.
Ut fra økosystembasert forvaltning og ISA sine anbefalinger bør konsekvensutredningen også innbefatte elementer som vurderer vern av områder: 1) med unikt biologisk mangfold, 2) som representerer alle typer habitater innen utredningsområdet, 3) som omfatter alle typer biofysiske landskaper, 4) som sikrer konnektivitet mellom verneområder, 5) som omfatter flere verneområder på forskjellige steder i utredningsområdet der alle typer habitater er representert, og 6) som hver for seg er av en størrelse slik at de kan fungere som selvstendige enheter.
Spredning av mineralpartikler fra overflateenheter for mottak og avvanning av malm eller fra utvinningsroboter på bunnen medfører at et hydrodynamisk modellverktøy bør etableres før konsekvensutredningen gjennomføres. Dette er viktig fordi påvirkningsområdet for slike utslipp kan omfatte store arealer ut over området der selve utvinningen vil foregå. Videre kan vannmassene utenfor 200-mils økonomisk sone være under internasjonal jurisdiksjon, og påvirkninger her bør derfor utredes med hensyn til internasjonale konvensjoner, reguleringer og anbefalinger, for eksempel ut fra OSPAR, UNCLOS og ISA.
En konsekvensutredning bør ta for seg ulike effekter av partikkelutslipp, slik som opptak i næringskjedene og bioakkumulering med hensyn til metaller, kjemikalier og mikroplast i relasjon til mattrygghet. I hvilken grad partikkelstørrelse kommer inn under EU sitt regelverk om nanopartikler bør også vurderes.
Det foregår både et nasjonalt og internasjonalt fiskeri i utredningsområdet. Effekter på dette og på sjøpattedyr som benytter området til beiting eller reproduksjon, bør inngå i konsekvensutredningen.
Håndtering av avfall fra prosessering av malm på land bør også inngå i en konsekvensutredning.
Vi har under arbeidet med kvalitetssikring av dette høringsdokumenter blitt oppmerksomme på at det pågår flere andre prosesser inn mot utvinning av havbunnsmineraler på norsk kontinentalsokkel og i dyphavet, både hos Havforskningsinstituttet og andre organer. Det er til dels overlappende bestillinger fra ulike etater, og rekkefølgen på bestillingene fremstår lite hensiktsmessig for en god prosess da kunnskapsbidrag ser ut til å komme etter denne høringsrunden. Dette er omhandlet i et eget brev. Viser også til Melding til Stortinget om marint vern (Meld. St. 29, 2020-2021) som nettopp ble publisert og som er relevant for konsekvensutredningen.

Bilder fra 460 – 1000 m dyp, Storegga og utenfor Mørebanken. (Kilde: Mareano/Havforskningsinstituttet, høsttoktet 2013)
2 - Overordnede kommentarer
Vi viser til Forslag til program for konsekvensutredning etter havbunnsmineralloven i forbindelse med Åpningsprosess for undersøkelse og utvinning av havbunnsmineraler på norsk kontinentalsokkel. Utvinning av metaller fra havbunnen utenfor nasjonalstatenes jurisdiksjon i det som er kalt «Området», er underlagt FN sitt organ UNCLOS. Det er økende interesse for slik metallutvinning, både i Stillehavet, Atlanterhavet og det Indiske hav (Levin m.fl. 2020). «International Seabed Authority» (ISA) er opprettet av UNCLOS for å etablere et rammeverk med hensyn til regulering av mineralvirksomhet på havbunnen i «Området». ISA-rammeverket er stadig under utvikling ut fra ny kunnskap gjennom arbeidet i den tekniske og rådgivende komitéen i ISA («Legal and Technical Commission», LTC), og dette bør være svært relevant også når det gjelder utvinning av havbunnsmineraler innenfor nasjonal jurisdiksjon. Dette rammeverket omfatter regler, prosedyrer og anbefalinger som i stadig større grad de siste årene har tatt opp i seg prinsippet om økosystembasert forvaltning (EBM) (Guilhon m.fl. 2020), og som trolig vil bli viktig for internasjonal anerkjennelse av aktivitet knyttet til mineralutvinning på havbunnen i fremtiden. Forslaget til program for konsekvensutredning, heretter benevnt «Høringsdokumentet», nevner ISA flere ganger, men det går ikke klart frem i hvilken grad anbefalinger fra ISA er innarbeidet i den kommende konsekvensutredningen.
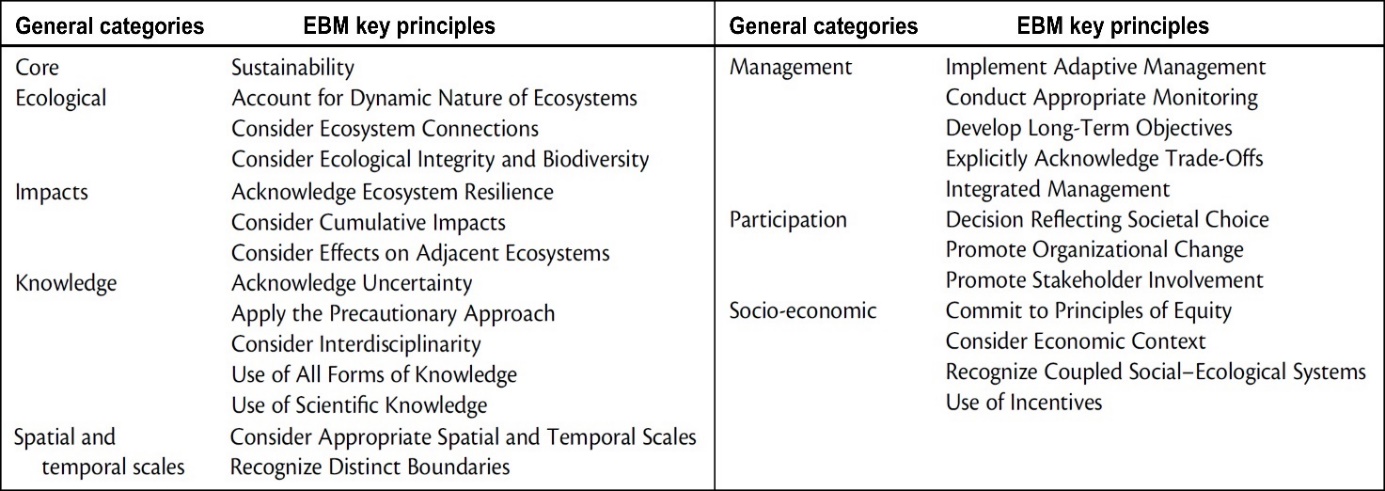
Økosystembasert forvaltning bygger på en rekke prinsipper både for samfunn og miljø (Tabell 1), og er en helhetlig forvaltning med mål å oppnå balanse mellom bevaring, bærekraftig bruk, og en rettferdig og lik fordeling av goder og økosystemtjenester som naturen kan tilby. Dette innebærer at for å bevare et økosystem sin integritet og sitt biologiske mangfold, vil en grundig forståelse av et økosystem sin funksjon være nødvendig forut for enhver planlagt påvirkning i økosystemet. Som høringsdokumentet påpeker omfatter havområdene som nå vurderes åpnet, store arealer med svært ulik karakter, fra sokkelområder med kontinentalskråning til dyphav med undersjøiske topper, sletter og midthavsrygger, der alle disse har sin egen områdespesifikke og karakteristiske fauna. Høringsdokumentet viser til at kunnskapsgrunnlaget er best på sokkelområdene, og at det vil skje en oppdatering av kunnskapsstatusen rettet mot økosystemer assosiert med habitater der havbunnsmineraler er påvist i de dypere delene av havområdet, inkludert strømforhold i vannsøylen. Ideelt sett burde denne kunnskapsoppdateringen vært på plass før planprogrammet for konsekvensutredningen ble sendt ut på høring.
Tabell 1. Fordeling av prinsipper for økosystembasert forvaltning (EBM) i generelle kategorier slik disse er angitt i Guilhon m.fl. (2020).
Ut fra det store arealet som planlegges åpnet, vil likevel kunnskap om økosystemene med hensyn til dynamikk, utbredelse, skalering, artsmangfold, sårbarhet, utveksling mellom populasjoner og genetiske ressurser være svært fragmentarisk uten at det gjennomføres betydelig mer omfattende undersøkelser i tid og rom. Kunnskapsmangelen medfører en stor usikkerhet, noe også høringsdokumentet angir vil bli ivaretatt i konsekvensutredningen. Ut fra økosystembasert forvaltning kan denne usikkerheten i tillegg tas høyde for gjennom risikovurderinger og anvendelse av føre-var-prinsippet. I anbefalingene fra ISA pekes det blant annet på behovet for omfattende baselinjestudier før arealer åpnes for mineralvirksomhet og behov for uberørte og representative referanseområder som ikke vil utsettes for påvirkninger fra mineralutvinning på havbunnen (ISA 2020). Havforskningsinstituttet vil derfor anbefale en omfattende heving av kunnskapsgrunnlaget ut over det som er tilgjengelig i dag, spesielt i de dypere delene utenfor de grunne sokkelområdene. Videre vil vi anbefale at konsekvensutredningen gjennomfører en risikoanalyse for biologisk mangfold og økosystemtjenester som synliggjør kunnskapsmangelen, for eksempel etter nye metoder anvendt i Havforskningsinstituttets risikoanalyse for fiskeoppdrett (Grefsrud m.fl. 2019).
Ifølge UNCLOS vil vannsøylen utenfor grensene til 200-mils-sonen være under internasjonal jurisdiksjon. I høringsdokumentet er betydelige deler av arealet som er foreslått åpnet for mineralutvinning, omfattet av dette. Siden mineralutvinning på havbunnen vanskelig kan skje uten partikkelutslipp til vannmassene, bør dette inngå i konsekvensutredningen med tanke på internasjonale konvensjoner, reguleringer og anbefalinger, for eksempel ut fra OSPAR, UNCLOS og ISA. Det bør tidlig i prosessen etableres modellverktøy for å kunne simulere spredning av partikler ved utslipp fra overflateenheter, da dette vil være vesentlig for å vurdere økologiske effekter av et utslipp.
Høringsdokumentet viser videre til at utvinning av havbunnsmineraler er en helt ny virksomhet, og at det ikke foregår slik utvinning i dag. Det innebærer at metoder og teknologi for realistisk gjennomføring av slik virksomhet ikke er fremskaffet eller utprøvd, hverken på havbunnen eller om bord på tilknyttede skip eller plattformer. Dette gjør det vanskelig å vurdere miljøeffekter av en slik aktivitet. Blant annet vil omfanget av kjemikaliebruk eller i hvilken grad avløpsvann filtreres og renses for partikler, være bestemmende for mulige miljøeffekter. Ved bruk av kjemikalier bør disse klassifiseres i fargekoder slik som offshoreindustrien i oljenæringen er underlagt.
I forbindelse med konsekvensutredningen viser høringsdokumentet til at det skal det gjennomføres en teknologistudie for hele verdikjeden. At endelig utforming av teknologi og metoder mangler vil likevel medføre en stor usikkerhet i hvordan ulike aspekter av utvinningen bør konsekvensutredes. En konsekvensutredning bør derfor ut fra den nevnte usikkerheten favne over et vidt spekter av scenarier når det gjelder metoder og teknologi, både for innhenting av malm på bunnen, transport av malm til overflateenhet og prosessering av malm før transport til land. Konsekvensutredningen bør også omfatte en rekke scenarier med hensyn til aktuelle kjemikalier samt avløpsvannets beskaffenhet med hensyn til størrelser, mengder og typer av partikler, inkludert mulige effekter av avløpsvannet når det slippes på ulike dyp. I tillegg bør konsekvensutredningen utrede behandling av avfall etter at malmen er prosessert på land.
Vi vil i det følgende presentere aktuelle forhold vedrørende hydrodynamikk samt ulike aspekter ved økosystemene på bunnen (det bentiske systemet) og i de frie vannmassene (det pelagiske økosystemet). Fra disse vil vi beskrive momenter vi mener er viktige for en konsekvensutredning knyttet til leting og utvinning av havbunnsmineraler.
3 - Hydrodynamiske forhold og partikkelspredning
3.1 - Hydrodynamisk karakterisering og måleprogram
Havstrømmene i Norskehavet er komplekse og sammensatte (figur 1). Vannmassenes bevegelser er essensielle for spredning av partikler og materiale som frigjøres både nær bunnen og i vannmassene fra punktet der avløpsvannet fra overflateplattformen vil slippes ut. Angående oppdatering av kunnskapsstatus for havsirkulasjon må det etableres et måleprogram for innsamling av oseanografiske data som har nødvendig romlig og tidsoppløsning for å få en adekvat karakteristikk av havmiljøet i hele utredningsområdet. Måleprogrammet må dekke de hydrodynamiske prosesser ved bunn, vannkolonnen og overflaten og inneholde fysiske og kjemiske parametere som sannsynligvis vil bli berørt av utslipp og gruveaktivitet. Dette inkluderer trykk, strømretning og hastighet, temperatur, saltinnhold, oksygen, turbiditet, optiske forhold (lys, partikkelkonsentrasjon og sammensetning, pH, etc.).

Figur 1: Simulert overflatestrøm i Norskehavet for 19. mars 2021 kl. 13:00. Datakilde: OSCAR / Earth & Space Research (https://www.esr.org/research/oscar/ og https://earth.nullschool.net).
Det må tas hensyn til at utredningsområdet er i skjæringspunktet mellom flere havbunnsbassenger med forskjellige vannmasser (for eksempel atlantisk vann, arktisk vann, intermediært vann, dypvann og bunnvann) samt at den grove topografien gir kompliserte strømforhold. For å få en god romlig miljøbeskrivelse av området vil dette kreve flere målestasjoner eller rigger, spesielt i områder med store horisontale gradienter. Man kan for eksempel følge anbefalingene fra World Ocean Circulation Experiment (WOCE) angående målestrategi. Målinger av turbulens er viktige for å bestemme spredning av partikler, og slike målinger sammen med strømmålinger må dekke flere tidevannsykluser. Målinger av de fysiske, kjemiske oseanografiske parameterne må også beskrive de sesongmessige variasjonene. Innsamling av data bør følge den beste tilgjengelige metodikk og internasjonale standarder, for eksempel slik dette er gitt av UNESCO gjennom Intergovernmental Oceanographic Commission (IOC) og International Oceanographic Data and Information Exchange (IODE), eller Ocean Best Practices System (OBPS).
3.2 - Simulering av partikkelspredning
Det er særlig viktig at det etableres en sirkulasjon- og spredningsmodell som har tilstrekkelig romlig oppløsning for å gjøre sensitivitetsanalyse av effektene av mineralutvinning (simuleringsforsøk av partikkelspredning) i aktuelle utvinningsområder. Et slikt modellverktøy må kunne benyttes til simulering av spredning av finpartikler både fra produksjonsutslipp i vannmassene og fysisk utvinningsaktivitet på havbunnen, og modellen må ta hensyn til bunntopografi og strømmønster i ulike dyp innenfor store nok arealer til å få realistiske anslag for partikkelspredning. Dette innebærer at det etableres en hydrodynamisk modell for vannbevegelser som kobles til en spredningsmodell hvor partiklenes størrelse, form og sammensetning inngår. Det må benyttes realistiske randverdier for modellområdet, og den hydrodynamiske modellen må kunne simulere både horisontale og vertikale bevegelser i alle vannlag.
I høringsdokumentet henvises det til modellering av partikkelspredning der flokkulering kan være en viktig faktor (Spearman m.fl. 2020). I dette arbeidet ble vannhastigheter på 0,03-0,3 m/s benyttet, men det er episoder med kraftig strøm som er viktig med hensyn til partikkelspredning. For eksempel er det målt opp mot 0,76 m/s ved Schulz bank i området mellom Mohns- og Knipovitsj-ryggen (Hanz m.fl. 2020). Modellene må valideres, og en metode for dette kan for eksempel være in situ hydrografiske data (Lopes m.fl. 2019) eller bruk av metallisotoper (Wiederhold 2015). Fastsittende hardbunnsfauna som filtrerer vannmassene for føde, for eksempel svamper, tar opp metaller og kan derfor benyttes som bioindikatorer (Hansen m.fl. 1995, Orani m.fl. 2018).
Det er sannsynlig at både selve utvinningen av mineraler på bunnen og særlig oveflateenheter for mottak og avvanning av malm, vil produsere en finfraksjon av partikler hvor en betydelig andel kan være av størrelse < 20 µm. Det er blant annet funnet at barittpartikler på mellom 10 og 20 µm størrelse fra oljeboring i Nordsjøen sedimenterer flere hundrede km unna, i Norskerenna (Lepland m.fl. 2000). Resuspensjon av finpartikler ved episodisk høye strømhastigheter er også et moment som bør utredes med hensyn til spredning av partikler, og slike episoder er påvist å skje jevnlig i området ved Shultz Bank (Hanz m.fl. 2020). Konsentrering av malm gjennom avvanning som innebærer ulik grad av filtrering og eventuelt bruk av kjemikalier, kan representere både kjemisk, finpartikulær og nanopartikulær forurensing. Figurer i høringsdokumentet og Levin m.fl. (2020) antyder at utslipp fra overflateenhet kan skje fritt i vannmassene. Dette gir mulighet for et svært stort spredningsområde både vertikalt og horisontalt, særlig hvis utslippet har mye finfraksjon. En simulering av et partikkelutslipp nær Azorene viste at partikler med størrelse 100 µm sluppet ut i 200 m dyp kan bruke opptil 40 dager på å nå bunnen (Lopez m.fl. 2019). Dette illustrerer tidsaspektet ved spredning av partikler ved utslipp i vannsøylen, der mindre partikler i en finfraksjon vil holde seg svevende i vannsøyler i betydelig lengre tid. En viktig del av den kommende konsekvensutredningen bør derfor være å evaluere om relevante modell- og simuleringsverktøy kan gi realistiske estimater for spredning, akkumulering og resuspensjon av finpartikler, både i vannmassene og ved bunnen, slik at man har det beste grunnlaget for å bergene den arealmessige påvirkning og risiko som omfatter alle aspekter ved en mineralutvinning på havbunnen. Det vil være utilstrekkelig bare å utrede lokale forhold, selv om aktiviteten rundt mineralutvinning vil være lokalt avgrenset, når virkningene kan ha mye større geografisk utbredelse.
3.3 - Mulig utslipp av nanomaterialer
Når blandingen av malm og sjøvann blir avvannet før transport til landbasert opparbeiding, er det i høy grad sannsynlig at den finest oppmalte malmfraksjonen slippes tilbake i havet med prosessvannet. Det bør derfor vurderes om dette utslippet defineres som nanomateriale. Selv om det finnes ulike definisjoner på nanomaterialer, vil «nano» i prinsippet omfatte størrelsesområdet 1-1000 nanometer. Ifølge EU defineres et nanomateriale som:
«et naturlig, tilfeldig oppstått eller fremstilt materiale, som består av partikler i ubundet tilstand eller som et aggregat eller som et agglomerat, og hvor minst 50 % av partiklene i den antallsmessige størrelsesfordelingen i en eller flere eksterne dimensjoner ligger i størrelsesintervallet 1-100 nm. I særlige tilfeller, og hvor hensynet til miljø, helse, sikkerhet eller konkurranseevne berettiger det, kan terskelen for den antallsmessige størrelsesfordeling på 50 % erstattes av en terskel på mellom 1 og 50 %.» (EU 2011/696).
Denne definisjonen betrakter også agglomerater av nanopartikler, dannet for eksempel ved hjelp av flokkuleringsmidler eller sjøvann, som nanopartikler. Selv om en lav vektprosentandel av massen består av nanopartikler kan det utgjøre 50 % eller mer av totalmengden med hensyn til partikkelantall (figur 2).
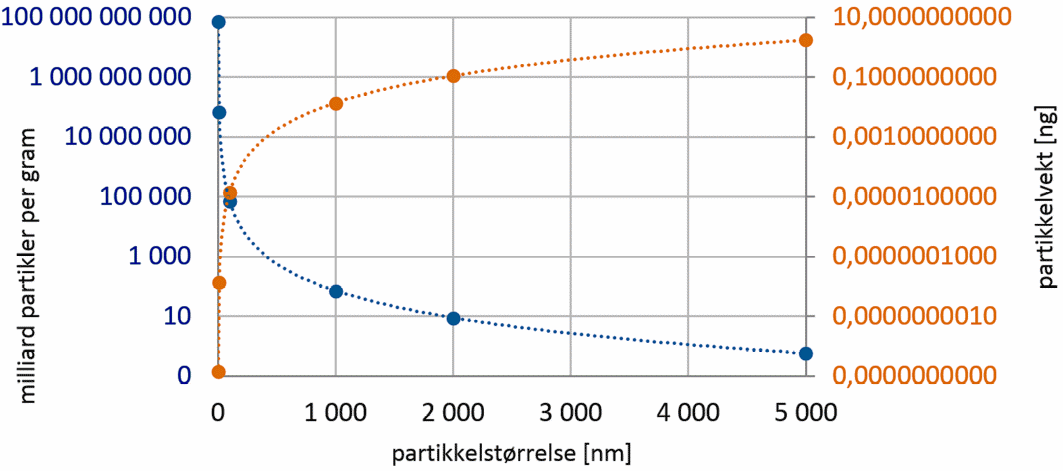
Figur 2. Sammenheng mellom størrelse, vekt og antall for kvartspartikler. Blå: Partikkelantall pr. gram i milliarder. Oransje: vekt pr. partikkel i nanogram (sfæriske partikler med en tetthet av 2,65 g/cm3). Merk: Y-aksene er log-skalert. (Havforskningsinstituttet 2020).
Som vi har belyst i andre sammenhenger (Havforskningsinstituttet 2020), har effekter av nanomaterialer i både miljø, mat og mennesker fått økende fokus de siste årene på grunn av de unike egenskapene slike materialer har. Dette gjenspeiles i både særegne fysisk-kjemiske egenskaper og toksisitet til nanomaterialer. Utslipp og effekter av utilsiktete nanomaterialer produsert i forbindelse med brytings-, oppmalings- og anrikningsprosesser gjennom mineralutvinning på havbunnen bør derfor undersøkes og dokumenteres slik at dette kan risikovurderes på en meningsfull måte. Det er viktig å påpeke at EU har oppdatert REACH regelverket (2006/1907) når det gjelder stoffer i nanoform og utslippskarakterisering (EU 2018/1881).
3.4 - Mulig utslipp av mikroplast
Når brutt malm skal transporteres fra havbunnen til overflaten for videre opparbeiding på et skip eller en plattform, skal det sannsynligvis skje ved å pumpe en malm/sjøvann-dispersjon gjennom rørledninger. Det er i høy grad sannsynlig at et slikt rørsystem vil bestå av plast. Transport av malmpartikler gjennom plastrør vil kunne føre til slitasje på innsiden av røret, som fører til dannelse av mikro- og (potensielt) nanoplast. Slitasje på rørsystemet er ikke nevnt som mulig forurensningskilde og bør inngå i en konsekvensutredning. Slik slitasje er et kjent problem i lakseoppdrett, der fôr og rensesvamper transporteres med høyt trykk gjennom plastrør (Gomiero m.fl. 2020). Oppdrettsindustrien har allerede begynt å se etter alternativer og utreder konsekvenser for dette. Mengden av plast som kan dannes ved mineralutvinning på havbunnen og rørslitasje bør beregnes. Det bør også vurderes om det er andre kilder til plastforurensing som kan oppstå i teknologien for utvinning av havbunnsmineraler.
4 - Det pelagiske økosystemet
Havforskningsinstituttet har fått en bestilling fra Oljedirektoratet med tittel «Åpningsprosess for utforskning og utvinning av havbunnsmineraler på norsk kontinentalsokkel. Grunnlagsstudie: Pelagisk økosystem », og som skal leveres i løpet av juni 2021. Denne bestillingen omfatter en rapport på kunnskapsgrunnlaget for det pelagiske økosystemet i området som vurderes åpnet for undersøkelser og utvinning av havbunnsmineraler, og den ville vært viktig for innspill til den pågående høringen om «Forslag til program for konsekvensutredning etter havbunnsmineralloven».
Utredningsområdet er heterogent med hensyn til bunntopografi (dyp) og vannmassetyper (for eksempel arktisk og atlantisk vann), samtidig som det finnes store hydrografiske forskjeller vertikalt gjennom vannsøylen, særlig med hensyn til temperatur. Disse forholdene har stor betydning for hvilke arter som forekommer i et gitt geografisk område, og det vil være klare forskjeller langs aksene nord-syd og øst-vest. Videre vil det være klare gradienter både når det gjelder sesongmessig utvikling i planktonsamfunnet for primær- og sekundærprodusenter og med hensyn til forekomster, mengder og biogeografisk sammensetning av arter. Et eksempel er Mohns-ryggen som strekker seg nordøstover fra Jan Mayen og separerer Grønlandsbassenget fra Lofotenbassenget. Mohns-ryggen markerer samtidig et viktig skille i havklima mellom dominans av arktisk vann i vest versus varmere atlantisk vann i øst, og hver av disse vannmassene er karakterisert ved egne typiske planktonarter. Vi vil i det følgende nevne en del viktige grupper og arter av dyreplankton (zooplankton) for de ulike vannmassene og dypene, som har interesse for utredningsområdet for mineralleting og utvinning på havbunnen.
4.1 - Mesozooplankton
-
Mesozooplankton i Norskehavet er representert ved en rekke forskjellige grupper og arter, hvor hoppekreps (copepoder) vurderes å være blant de aller viktigste med hensyn til biomasse og antall (Melle m.fl. 2004). Hoppekreps i slekten Calanus er blant de viktigste beiterene på planteplankton i Norskehavet og Grønlandshavet, og utgjør en betydelig biomasse. Calanus-artene spiller i tillegg en nøkkelrolle i økosystemet som sentralt bindeledd i næringsnettet mellom planteplankton og høyere trofiske nivåer som for eksempel fisk, fugl og sjøpattedyr (Melle m.fl. 2014).
Calanus finmarchicus (raudåte) forekommer i særlig store mengder i den relativt varme atlantiske vannmassen sentralt og øst i Norskehavet (Strand m.fl. 2020, Melle m.fl. 2014, Conover 1988). Årlig produksjon av C. finmarchicus i Norskehavet er anslått å være omtrent 190-290 millioner tonn våtvekt (Hjøllo m.fl. 2012, Skjoldal m.fl. 2004). Dens mye større slektning, Calanus hyperboreus, har derimot sin kjerneutbredelse i Grønlandshavet (Hirche 1997) hvor den har en tilsvarende økologisk funksjon som C. finmarchicus har i Norskehavet. En tredje slektning, Calanus glacialis, er knyttet til arktiske vannmasser, særlig assosiert med kontinentalsokkelområdene i nord, og finnes også spredt i kaldtvannspåvirkede områder rundt Jan Mayen (Fossum m.fl. 2012). For utredningsområdet angitt i høringsdokumentet så vurderes forekomsten av denne arten å være av liten betydning sammenlignet med sine nære slektninger nevnt ovenfor.
Calanus-artene foretar store sesongmessige vertikalvandringer og tilbringer sommeren i overflatelagene, men vinteren på dypt vann (Østvedt 1955, Halvorsen m.fl. 2003). I Norskehavet, som i stor grad overlapper med utredningsområdet, vandrer Calanus finmarchicus (raudåte) opp fra de dypere deler av vannsøylen (500-1000 m) til overflatelagene tidlig om våren for å beite på våroppblomstringen av planteplankton og reprodusere (Hirche 1996, Bagøien m.fl. 2012). I Grønlandsgyren og i Vest-Spitsbergen-strømmen (mellom Bjørnøya og Mohns-ryggen), overvintrer flere generasjoner av C. hyperboreus på dybder mellom 500 m og 3000 m (Hirche 1997).
4.2 - Makrozooplankton
-
Pelagiske amfipoder (Hyperiidae) er en svært viktig økosystemkomponent i store deler av utredningsområdet, og særlig arter i slekten Themisto. Disse er viktige predatorer på annet dyreplankton, spesielt på hoppekreps som C. finmarchicus. Themisto libellula er tradisjonelt antatt å være en arktisk assosiert art som er viktig i de øverste vannlag fra overflaten til 300 m dyp. Den er en viktig fødeorganisme for blant annet Grønlandssel, torsk og polartorsk i områder i nærheten av iskanten og i arktiske vannmasser i det nordlige Atlanterhavet. Svermer av T. libellula er imidlertid også er observert på betydelige dyp i Norskehavet (Aarbakke 2007) og ned mot 1200 m i Grønlandshavet (Vinogradov 1999). Lignende observasjoner av Themisto abyssorum (atlantisk boreal art) er gjort på Vøringplatået på 800 meters dyp like over bunnen (Vinogradov, 1999), assosiert med typiske atlantiske vannmasser i de øvre vannlag, men hvor det er langt kaldere vannmasser som dominerer over bunnen.
-
Krill (Euphausiidae) er en annen sentral økosystemkomponent i utredningsområdet med 3-6 arter, men hvor Meganyctiphanes norvegica, Thysanoessa inermis og Thysanoessa longicaudata er de viktigste artene ut fra antall og biomasse. Storkrillen, M. norvegica, er den største og sannsynligvis viktigste krillarten. Den er svært tallrik langs kontinentalskråningene rundt Norskhavsbassenget, særlig i øst, og følger Atlanterhavsstrømmen vest og nord for Svalbard. Den finnes også i de sentrale deler av Norskehavsbassengene, og i området rundt Jan Mayen. M. norvegica er lyssensitiv, og foretar døgnlige vertikale vandringer fra overflaten om natten ned til 500 m eller dypere på dagtid (Knutsen & Serigstad 2001). En annen viktig krillart, Thysanoessa inermis, er særlig knyttet til kontinentalsokkelen rundt Svalbard og Jan Mayen, men den er også regelmessig til stede i norske kystfarvann. Voksne individer er registrert så dypt som 700 m over kontinentalskråningen i det østlige Norskehavet. Også denne arten er lyssensitiv og foretar døgnlige vertikalvandringer, men dens vertikale fordeling i vannsøylen avhenger av årstid og generelle fysiske forhold, og kan variere for området over kontinentalsokkelen versus dyphavet. Den tredje arten, Thysanoess logicaudata, er i hovedsak knyttet til atlantisk-påvirkede vannmasser der overvintring skjer på dypt vann i Norskehavet med en sesongmessig oppvandring til overflatelagene om våren.
-
Pelagiske reker representert ved slektene Sergestes, Hymenodora og Pasiphaea er tallrike og har sitt kjerneområde i de mesopelagiske vannmassene i Norskehavet, på dyp mellom 200 og 1000 m. Larver og yngre individer av enkelte rekearter kan være tallrike også høyere opp i vannsøylen. Alle artene i disse slektene er lyssensitive og foretar døgnlige vertikalvandringer. Dybdeintervallet de vandrer over er trolig noe mindre enn det vi kjenner til for storkrill. Disse rekene er predatorer på annet dyreplankton og er derfor viktige aktører i det pelagiske økosystemet, blant annet ved å bringe organisk materiale fra de øvre vannlag til de dypere delene av vannsøylen. De er trolig også en viktig fødeorganisme for fisk, blekksprut og maneter i de mesopelagiske vannmassene.
-
Maneter er en fellesbenevnelse for en heterogen gruppe organismer bestående av Cnidaria (nesledyr) og Ctenophora (kammaneter). Ulike typer nesledyr og kammaneter finnes fra overflaten til de største havdyp. Arter som er mest aktuelle i forbindelse med området interessant for mineralvirksomhet i Norskehavet, er flerårige og samtidig tallrike i de mesopelagiske vannmassene (200-1000 m). Kronemaneten, Periphylla periphylla, er en nøkkelart i denne sammenheng. Arten Atolla wyvillei, som er betydelig mindre av størrelse enn P. periphylla, er tidvis svært tallrik og har delvis overlappende dybdeutbredelse med sistnevnte.
-
Pelagiske blekkspruter forekommer i utredningsområdet, og den dominerende arten, Gonatus fabricii, er vidt utbredt og tallrik fra Nordpolen til de sørlige områdene av Norskehavet (Arkhipkin & Bjørke 1999). Den er et viktig byttedyr for blant annet fisk, flere hvalarter og sel. De yngste og minste individene er hovedsakelig lokalisert i de øvre 80 m av vannsøylen og vandrer til dypere vann på mellom 400 og 1200 m ved økende alder og størrelse (Bjørke & Gjøsæter 1998).
4.3 - Mineralutvinning og det pelagiske økosystemet
Manglende metodikk og teknologi for utvinning, vask, og oppkonsentrering av malm gjør det vanskelig å evaluere effekter av mineralutvinning på havbunnen. Prosessene nevnt ovenfor kan resultere i at betydelige mengder «restavfall» må tilbakeføres til sjø i åpent hav. Det kan også resultere i generering av sedimentskyer ved bunnen og partikkelskyer oppe i vannmassene (Figur 3). Dette innebærer at store vannvolum kan bli påvirket av partikler med tilhørende innhold av metaller og prosesskjemikalier. Mulige effekter av dette er listet i figur 3. Effektene bør utredes, og det pelagiske økosystemet bør inngå i baselinjestudier for å vurdere risiko ved mineralutvinning på havbunnen og utslipp fra overflateenheter etter prosessering og avvanning (Drazen m.fl. 2020).
Utslipp av partikler i øvre vannlag med tilhørende økt turbiditet har blitt vurdert å kunne ha en signifikant negativ effekt på produksjon av planteplankton (Chan & Anderson 1981, Christiansen m.fl. 2020). Lysskygging fra økt turbiditet kan medføre redusert produksjon på grunn av mindre lys til fotosyntesen. På den annen side kan tilførsel av uorganiske næringsstoffer og sporstoffer potensielt bidra til økt primærproduksjon. I sistnevnte tilfelle er det imidlertid ikke gitt at økt produksjon vil føre til en gunstig sammensetning og fordeling av algetyper for dyreplanktonet. Det er velkjent at særlig i kystområder kan lysregimet i vannsøylen ha en betydelig strukturerende effekt på den akvatiske næringskjeden (Aksnes m.fl. 2009). Økt turbiditet ved utslipp av mineralpartikler kan også ha konsekvenser for dyr som benytter synet til jakt, eller for selv å unngå predatorer, for eksempel fisk og krepsdyr med velutviklede fasettøyne som krill, amfipoder og pelagiske reker. Andre predatorer som maneter er ikke avhengig av synet for å fange et bytte, og kan under slike forhold få et konkurransefortrinn (Eiane m.fl. 1999).
Finpartikler med opprinnelse i utslipp fra overflateenheter ved mineralvirksomhet til havs kan bidra til tetting av munnapparat, lukt og respiratoriske overflater og organer hos dyreplankton. Dette kan ha særlig betydning for en rekke ulike arter, for eksempel kan organismer som filtrerer vannet med hensyn til fôropptak, være utsatt for finpartikler opp til flere titalls kilometer fra utslippsområdet (Drazen m.fl. 2020). En slik påvirkning kan gjelde for flere ulike planktongrupper, eksempelvis salper (kappedyr), appendikularier (halesekkdyr) og krepsdyr, men finpartikler kan også feste seg på gelatinøse planktonorganismer som maneter og kammaneter.
Generelt kan finpartikler og kjemikalier antas å påvirke fødeopptak samt kjemosensorisk adferd og luktspor som er relevant i forhold beiting og reproduksjon (Kiørboe & Bagøien 2005). Det er påvist at hoppekreps, inkludert C. finmarchicus , spiser mineralpartikler (Farkas m.fl. 2017). Dyreplankton kan på denne måten få i seg uorganiske mineralpartikler gjennom fôrinntak, enten ved passiv filtrering eller aktiv seleksjon av partikler (Paffenhöfer m.fl. 1982), noe som kan være i direkte i konkurranse med opptak av naturlig føde og påvirke reproduksjonen (Paffenhöfer 1972). Hoppekreps som spiser metallholdige finpartikler fra utslipp, kan i tillegg være en vektor for metaller direkte inn i næringskjeden.
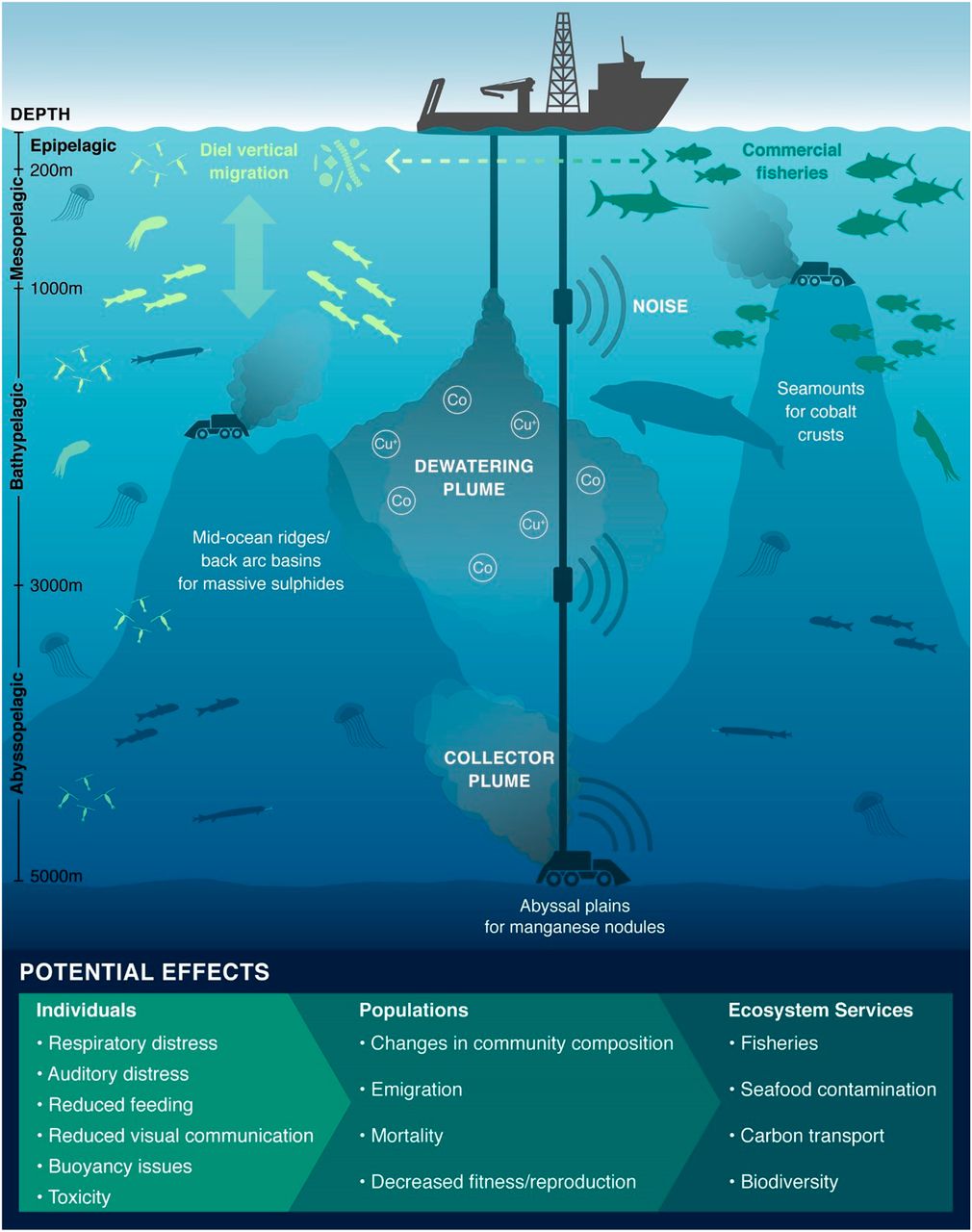
Figur 3. Partikkelutslipp og støy fra mineralvirksomhet i åpent hav kan ha en rekke effekter på pelagiske organismer. Figuren er hentet fra Drazen m.fl. (2020).
Bioakkumulering av uønskede giftkomponenter som tungmetaller (Kadiene m.fl. 2019), kan gi akutte eller kroniske effekter (Hook & Fisher 2001, Fuchida m.fl. 2017). Høye konsentrasjoner av biotilgjengelige metaller som frigjøres med utslippene i vannsøylen, vil kunne skade de omkringliggende planktonsamfunnene, og kan for en del organismer resultere i forhøyet dødelighet (Fuchida m.fl. 2017), hemming av vekst, feilutvikling eller lavere reproduksjonshastigheter (Hook & Fisher 2001). Høyere trofiske nivåer kan være særlig utsatt på grunn av bioakkumulering i næringskjeden, og området som påvirkes kan lett utvides gjennom delvis eksponering av plankton som foretar vertikale eller horisontale migrasjoner.
Utvinning av mineraler ved bruk av steinknusere og pumpesystemer, nær eller på selve havbunnen, kan få betydelige konsekvenser for eksempelvis hyperbenthos og meroplankton (Christansen m.fl. 2020). Hyperbenthos er bunnlevende organismer som også svømmer i vannlaget like over bunnen, og meroplankton er organismer med livsstadier knyttet til bunnen. Begge disse er potensielt sårbare dyregrupper i vannmassene nær bunnen, og forhøyet dødelighet av disse kan på sikt bidra til tap av biodiversitet og redusert fødetilgang for andre komponenter i økosystemet.
En rekke planktonorganismer er svært vare for både naturlig lys fra himmellegemene, men også for kunstig lys. Dette fordi egenprodusert lys (bioluminescens) som blant annet benyttes til kommunikasjon eller fangst av bytte, kan forstyrres i betydelig grad og resultere i adferdsendringer som kan påvirke overlevelse og muligens reproduksjon. Kunstige lyskilder nær overflaten eller på dypere vann kan påvirke døgnlige vertikalvandringer og kanskje forstyrre fôropptak, svekke unnvikelse fra predatorer og påvirke reproduksjonsadferd. Negative effekter av lys kan være tydeligere i miljøer med lave naturlige lysnivåer, som på større dyp. Hvis kunstige lyskilder er koblet til et pumpeinntak på en utvinningsrobot, vil organismer som tiltrekkes av lys risikere økt dødelighet. I litteraturen er det videre rapportert eksempler på at visse krepsdyr som eksponeres for kunstig lys risikerer å få permanente skader på fasettøynene (Attramadal m.fl. 1985, Herring m.fl. 1999).
5 - Klimaeffekter
Havforsuring som følge av økt innhold av CO2 i atmosfæren, kan endre metaller sin løselighet i havet. Med redusert pH minker hydroxid- og karbonationer, og disse endringene vil påvirke fordeling, frigjøring og tilgjengelighet av metaller som danner sterke kompleks med disse ionene (Millero m.fl. 2009). Marine organismer trenger spormetaller for å vokse, for eksempel jern, mangan og zink. En endring i tilgjengelighet av disse vil kunne påvirke biologisk produksjon. Havforsuring kan også føre til endring i hvordan metall binder eller adsorberes til organisk stoff (danner kompleks med organiske ligander), og kan endre på frigjøring og toksisitet av ulike tungmetaller slik som kvikksølv, bly, kadmium, nikkel og kobolt. Modeller viser at for eksempel fri ionekonsentrasjon av bly, aluminium og kvikksølv vil mer enn dobles i et fremtidig klima scenario med oppvarming og økt havforsuring (Stockdale m.fl. 2016). Metylkvikksølv er den organiske formen av kvikksølv som er giftig for levende organismer. Organisk bundet kobber og kadmium vil minke med 10 % ved økt havforsuring, mens organisk bundet kobolt og nikkel vil øke. Det trengs fortsatt informasjon om hvilken effekt disse endringer vil ha på marine økosystemer, men det er sannsynlig at klimaendringer og påfølgende havforsuring vil endre metallsyklusene i havet (Hoffmann m.fl. 2012), noe som kan gi en betydelig effekt på marine mikroorganismer (Stockdale m.fl. 2016). Konsekvensutredningen bør derfor vurdere fremtidig påvirkninger av mineralutvinning på havbunnen i et klimamessig perspektiv, der metallbelastning på marine organismer er ventet å øke.
6 - Mikrobielle forhold
Hydrotermale områder huser komplekse mikrobielle samfunn bestående av bakterier, arker, encellede eukaryoter og virus, inkludert mange arter som direkte eller indirekte er involvert i biokjemiske sykluser som svovel, jern og mangansyklusene (Van Dover 2019). Den mikrobielle delen av økosystemet påvirker dermed mange prosesser som blant annet fører til utfelling av metaller og svovelforbindelser i disse områdene. I dyphavet er kjemotrofe bakterier de viktigste primærprodusentene av organisk karbon, og er derfor sammen med synkende partikler (marin snø) viktige næringskilder for andre organismer. Filtrerende og gressende organismer assosiert med aktive hydrotermale områder, som for eksempel skjell, børstemark og krepsdyr, er direkte avhengig av disse mikrobielle samfunnene (Orcutt m.fl. 2020). Det er færre studier som fokuserer på ikke-aktive områder, og det er dermed mer usikkerhet rundt hvordan de mikrobielle samfunnsstrukturene er der, og deres rolle i næringsnettet (Orcutt m.fl. 2020, Van Dover 2019). Flere studier tyder imidlertid på at det mikrobielle mangfoldet assosiert med ikke-aktive svovelfelt er stort og komplekst, og involvert i essensielle kjemiske prosesser på havbunnen (Van Dover 2019 og assosierte referanser).
6.1 - Aktive områder
Mikrobielle samfunn i aktive hydrotermale områder har blitt ansett for å være særlig sårbare i forbindelse med gruvedrift (Orcutt m.fl. 2020). Olsen m.fl. (2016) påpeker at alle hydrotermale områder har sine egne karakteristika, og det bør gjøres steds-spesifikke analyser på hvert område. Slike studier er viktige med tanke på potensiell rekolonisering av området fra nærliggende områder i etterkant. Denne problemstillingen kan også være aktuell med tanke på mikroorganismer. Studier av hydrotermale områder blant annet i Stillehavet fant store forskjeller både i sammensetningen av mikrober og deres funksjonelle potensiale ved geografisk spredte områder (Fortunato m.fl. 2018). Mest sannsynlig reflekterer dette de abiotiske forholdene på de ulike lokasjonene, som pH og temperatur, tilgang på oksygen, metan, hydrogensulfid, etc., samt hvordan vann og hydrotermale væsker blandes (Fortunato m.fl. 2018). Det er derfor naturlig å anta at hvert område har et distinkt mikrobielt samfunn som vil variere med geografisk spredning og de lokale kjemiske og fysiske forholdene.
6.2 - Ikke-aktive områder
Ikke-aktive områder med svovelavsetninger er generelt mindre undersøkt enn aktive områder når det gjelder mikrobielt mangfold, metabolsk potensiale og aktivitet. De er imidlertid mest sannsynlig like mangfoldige som aktive områder (Van Dover, 2019 og assosierte referanser), og aktuelle utvinningsområder bør derfor vurderes individuelt.
Det bør avklares hvor motstandsdyktige de mikrobielle samfunnene i manganskorper og sulfidforekomster er med hensyn til mulig slamdeponering på havbunnen. Disse samfunnene er tilpasset sakte nedfall av marin snø og utfellinger fra hydrotermale områder over lang tid, men ikke nødvendigvis store mengder slam over korte perioder. To mulige konsekvenser av deponering av slam på havbunnen som potensielt kan ha stor påvirkning på det mikrobielle samfunnet er utslipp av tungmetaller og toksiske substanser, samt hypoxia (Christiansen m.fl. 2020, Olsen m.fl. 2016).
Effekten av utslipp av tungmetaller på de mikrobielle miljøene bør undersøkes nærmere. Bakterier og arker kan være sårbare når de utsettes for tungmetaller, og store utslipp av disse kan ha drastisk påvirkning på sammensetningen og mangfoldet i de mikrobielle samfunnene. Stimulering av mikrobesamfunn som metaboliserer tungmetaller kan også potensielt påvirke de kjemiske syklusene, og øke utfellingen av tungmetaller i disse områdene (Van Dover 2019). Tungmetaller kan også akkumuleres av bakterier og arker, enten intracellulært, som utfellinger rundt cellene eller som utfellinger på biofilmer (Chen m.fl. 1995, Jroundi m.fl. 2020). Bakterier og arker utgjør første steg i næringskjeden, både som mat for dyreplankton og gjennom gressing av biofilmer av for eksempel krepsdyr. Det er dermed en fare for at økt tilgjengelighet av tungmetaller i miljøet vil føre til økt akkumulering av disse oppover i næringskjeden. De mulige konsekvensene av økte mengder tungmetaller med opprinnelse i sulfider og manganskorper bør derfor undersøkes nærmere.
Gruvedrift som etterlater fordypninger i havbunnen i kombinasjon med deponering av store mengder slam, kan også føre til oksygenfrie soner på havbunnen (Christiansen m.fl. 2020, Orcutt m.fl. 2020). Et oksygenfritt miljø kan fremme vekst av anaerobe bakterier og arker og midlertidig fortrenge aerobe organismer. Områder med mye strømninger vil sannsynligvis gå forholdsvis raskt tilbake til «normale» tilstander, men det bør tas høyde for at dette kan ta lengre tid på enkelte lokasjoner. Anaerobe «lommer» på havbunnen vil videre ha en negativ effekt på bentiske dyr som lever på bunnen eller i sedimentene. Hver enkelt lokasjon bør derfor vurderes individuelt, og det bør tas høyde for hvor sannsynlig det er at anaerobe lommer vil etableres.
7 - Sårbare bunnhabitater
Verneområder og regionale forvaltningsplaner vil bli en viktig del av forvaltningen av mineralutvinning i Norskehavet. For å begrense det negative avtrykket av gruvedrift regionalt arbeider den internasjonale havbunnsmyndigheten (ISA) i sine områder med at det settes opp nettverk av verneområder hvor gruvedrift ikke vil bli tillatt (ISA 2011, ISA 2012). Plasseringen av verneområder skal være vitenskapelig begrunnet og være basert på fysisk, geokjemisk, biologisk og økologisk informasjon (Lodge m.fl. 2014, Dunn m.fl. 2018, Taboada m.fl. 2018, Simon-Lledo m.fl. 2019). Nettverket av verneområder skal:
1. Verne 100 % av alle kjente områder med unikt biologisk mangfold (som korallrev, varme havkilder) eller viktig økosystemfunksjon (som gyteplasser).
2. Verne 30-50 % av alle forskjellige typer habitat i området (leveområder som for eksempel spredningsrygger, undervannsfjell, dyphavssletter, etc.).
3. Representere alle biofysiske landskap likt (dyp, temperatur, oksygen, etc.).
4. Sikre at sammenhengen (konnektiviteten) mellom verneområder blir opprettholdt (at verneområdene ligger tilstrekkelig tett for at pelagiske stadier som for eksempel larver skal kunne drive mellom dem).
5. Sikre god replikasjon (at alle habitattyper er inkludert i vernet på flere forskjellige plasser).
6. Sikre at alle verneområdene er så store at de også kan fungere som selvstendige enheter.
Det er bred enighet i det internasjonale forskningsmiljøet og forvaltningsorganet ISA at et nettverk av vernede områder er essensielt for å bevare det biologiske mangfoldet og funksjonen i de marine økosystemene både under og etter mineralutvinning. Det er også vist at det å etablere verneområder tidlig og før lisenser for leting blir lyst ut, er viktig. Etablering av verneområder mellom allerede utdelte lisensområder vil ikke gi tilstrekkelig beskyttelse til det marine miljøet slik dette er vurdert for den midtatlantiske rygg i sørlige Atlanterhavet og Clarion Clipperton Fracture Zone (CCFZ) i Stillehavet. Det pågående arbeidet i Norskehavet for å identifisere særlig verdifulle områder (SVO) vil gi viktig informasjon som kan trekkes inn i vurderingene av mulig plassering av verneområder i norske havområder. Men det må skje en utstrakt kartlegging av biodiversitet, økosystemfunksjon, taksonomi og konnektivitet før man har data av god nok kvalitet til at en slik vurdering kan bli foretatt. Biodiversitetsstudier er et meget tidskrevende arbeid, særlig i områder som er lite undersøkt, og hvor man kan forvente å finne mange nye og ubeskrevne arter. I snitt vil det ta 5 år å beskrive en art. I CCFZ har dette arbeidet pågått i mange år allerede, det er blitt foretatt mange månedslange tokt årlig, og det er fortsatt mange nye arter å finne. Norskehavet kan ha noen flere forskjellige geomorfologiske habitatstyper enn CCFZ, og i tillegg vil de atlantiske og arktiske vannmassene by på forskjellige arter. Det er derfor forventet at man vil finne minst like mange nye arter her som i CCFZ. Forslaget til program for konsekvensutredning viser tydelig til at bare de grunnere, mer kystnære delene av utredningsområdet er relativt godt kjent, mens de dypere delene av utredningsområdet er lite undersøkt. Dette bør særlig vektlegges i konsekvensutredningen.
7.1 - Særlig verdifulle og sårbare områder
Særlig verdifulle og sårbare områder (SVOer) er områder med en vesentlig betydning for det marinbiologiske mangfoldet og produksjonen, med ringvirkning også utenfor seg selv, og de er en viktig del av forvaltningsplanene i norske havområder (Meld. St. 2019-2020). SVOer blir identifisert ved hjelp av forhåndsdefinerte kriterier som for eksempel EBSA-kriterier (Ecologically and Biologcally Significant Areas), og baserer seg på strikte vitenskapelige undersøkelser. Det pågår i 2020-2021 en revisjon av SVOer i Norske havområder som med all sannsynlighet vil medføre at de dype bassengene i Norskehavet og midthavsryggene, det vil si Jan Mayen-ryggen, Jan Mayen bruddsonen, Mohns-ryggen og Knipovich-ryggen, vil bli kategorisert som særlig verdifullt og sårbare områder i de nye forvaltningsplanene. De inneholder unike og sjeldne habitater, fungerer som livshistorisk viktige områder for en rekke arter, inneholder truede og nedadgående arter samt sårbare arter med lav restitusjonsevne. Detaljerte vitenskapelige undersøkelser med AUVer og ROVer har blitt utført på noen få plasser langs Mohns-ryggen og Jan Mayen-ryggen. Resterende deler av midthavsryggene i Norskehavet er meget dårlig kartlagt. Storskala systematisk taksonomisk kartlegging av det bentiske biomangfoldet i de dype bassengene av Norskehavet og den sedimentære havbunnen som strekker seg fra midthavsryggen til de dype bassengene, er ikke blitt foretatt siden den Norske Nordhavekspedisjonen i 1876-1878 som resulterte i en rekke meget godt kjente vitenskapelige publikasjoner. Undersøkelser av midthavsryggene de siste 20 årene inkluderer funn og beskrivelser av fauna rundt flere aktive varme havkilder, som Loki’s Castle, Ægirs Kilde og Fåvne, samt utdødde kilder som Copper Hill og Mohns Treasure, med mye endemisk fauna og mikroorganismer (Kongsrud & Rapp 2012, Kongsrud m.fl. 2017, Ramirez-Llodra m.fl. 2020, Steen m.fl. 2016, Tandberg m.fl. 2012, 2013, 2018). Den komplekse topografien over ryggene påvirker de hydrografiske strømningene, og lokalt vil det oppstå vertikal strøm, brytende bølger og blanding av vannlag (McClain m.fl. 2010, Etnoyer 2010, Schlacher m.fl. 2014; White m.fl. 2007). Dette fører ofte til tette samfunn av filtrerende dyr som bygger svampskoger og korallhager (Fossum m.fl. 2012, Burgos m.fl. 2020, Meyer m.fl. 2019, Roberts m.fl. 2018). De siterte arbeidene er viktige, for de peker på hvilken type biologisk liv vi kan forvente å finne ved fremtidige undersøkelser, men det må understrekes at kun en brøkdel av området er blitt undersøkt.
Eksisterende undersøkelser av punktlokaliteter samt modelleringsarbeid som indikerer en vid forekomst av varme havkilder, svamp og korallskoger, er grunnen til at midthavsryggene er å betrakte som særlig verdifulle og sårbare områder. Svampskoger og korallhager er av OSPAR kommisjonen vurdert som truet (OSPAR 2008), og står på den norske rødlisten for naturtyper (Artsdatabanken 2018). Varme havkilder er også på den norske rødlisten. Både svamper og koraller er saktevoksende arter som kan bli svært gamle, men har dårlig evne til å restituere seg (Doughty m.fl. 2014, Lacharité & Metaxas 2013, Mortensen & Buhl-Mortensen 2005, Roberts m.fl. 2009). Filtrerende arter kan være meget sensitive for forhøyede mengder suspenderte partikler i vannet (Fang m.fl. 2018, Kutti m.fl. 2015, Scanes m.fl. 2018). Mange arter ved varme og kalde kilder samt i sulfidhaugene ved utdødde kilder har meget lav til ingen restitusjonsevne hvis habitatet de er avhengig av blir ødelagt eller tatt bort (Ramirez-Llodra m.fl. 2020). Svampskoger og korallskoger spiller en svært viktig rolle for karbonsyklusen på bunn (Cathalot m.fl. 2015, Kutti m.fl. 2013), er levested for en rekke andre arter fra mikroorganismer til fisk, og er ofte assosiert med et kraftig forhøyet biomangfold (Klitgaard 1995, Freiwald m.fl. 2012). Det er behov for økt kunnskap om økosystemfunksjon, også for andre bunnlevende samfunn i relevante områder hvor det kan bli aktuelt med gruvedrift eller leting etter mineraler, samt forstå den rollen nøkkelarter spiller i økosystemene. For eksempel kan noen koraller eller svamparter være viktig for juvenil fisk.
7.2 - Effekter av partikler på organismer
Det er blitt vist at forskjellige organismer responderer veldig ulikt på eksponering for partikler og tungmetaller. Det er derfor viktig at det blir gjennomført effektstudier for å undersøke toleranse hos alle relevante pelagiske og bunnlevende nøkkelarter med hensyn til mineralpartikler og tungmetaller. Det er ikke mulig å bruke sensitivitetsdata for andre arter for å regulere utslipp, eller for å forutsi konsekvenser av utslipp. Det er også kjent at forskjellige livsstadier kan oppvise ulik sensitivitet og at sesong kan påvirke utfallet av sensitivitetsstudier. Dette innebærer at for sensitivitetsstudier med samme art, for samme tungmetall og ved samme konsentrasjoner, kan utfallet variere betraktelig ved to forskjellige tidspunkter. Andre faktorer som man vet påvirker arters sensitivitet, er temperatur, pH og trykk. Det er derfor viktig at effektene av temperatur og pH blir undersøkt, og at eksponeringsstudier blir gjennomført under relevant trykk. Toleransedata er imidlertid svært viktig å fremskaffe, og slike studier bør bli prioritert tidlig i prosessen. Eksperimenter utført under forskjellig temperatur og pH er viktig, for det er forventet at arters sensitivitet til tungmetaller vil øke i fremtiden. De dype delene av Norskehavet er et område hvor både pH og temperatur er under rask forandring.
8 - Bunnfisk
Havforskningsinstituttet ved faggruppe «Dyphavsarter og bruskfisk» har bestandsansvar for følgende arter/grupper: blåkveite, vanlig uer, snabeluer, lange, blålange, brosme, vassild og 22 arter av bruskfisk (herunder haier, skater og havmus). Mange av artene har en sterk tilknytning til Eggakanten, og en konsekvensutredning bør gjøre rede for hvor stor sikkerhetsmargin som er nødvendig for å sikre at effekter av mineralutvinningen ikke når Eggakanten. De fleste av disse artene lever lenge, hvilket gjør dem ekstra sårbare for menneskelig påvirkning. I Norge har vanlig uer, blålange og mange av bruskfiskartene i området rødlistestatus som sterk truet (for eksempel brugde, pigghå, storskate og håbrann), truet eller sårbar. For mange av de andre artene har vi ikke nok data til å si noe om tilstanden.
Figur 4 viser utbredelsesområde og gyteområde for utvalgte dyphavsarter. Ved sammenligning med utredningsområdet ser vi at artene har utbredelse nært eller i utredningsområdet. Deler av yngleområdet for snabeluer overlapper med østlige deler av utredningsområdet. Vi ser også at mange av artene har gyteområder som ikke overlapper med utredningsområdet, men som ligger veldig nært. Det er et sterkt behov for å utrede hvorvidt de negative effektene av mineralutvinning vil spres til disse områdene. For de datafattige bestandene, herunder mange bruskfisk, har vi ikke nok data til å si noe om utbredelsesområde/gyteområde, hvilket gjør det viktig å følge «føre-var»-prinsippet. Arter som vandrer over store områder, som for eksempel brugde og håbrann, vil potensielt være ekstra sårbare.
8.1 - Tap av leveområde
Utvinning av dyphavsmineraler innebærer fjerning av store mengder substrat. Bunndyrfauna vil mest sannsynlig ikke overleve gruveprosessen på en lokasjon. Av fisk vil arter med en sterk kobling mot bunnen, for eksempel skater, være mest utsatt, selv om de direkte effektene sannsynligvis er betydelig mindre en for fastsittende organismer. Eventuelle egg/eggkapsler fra bruskfisk og andre fisk som gyter langs bunnen vil kunne bli ødelagt, men det er uvisst hvor utstrakt gyting er på de aktuelle dyp. Endring av bunndyrfauna forårsaket av permanent fjerning av substrat vil kunne forstyrre koblingene mellom benthopelagiske samfunn (hyperbenthos og meroplankton) og bunndyrsamfunn.
8.2 - Nedslamming
Oppdrift er viktig både for overlevelse og spredning av egg. Partikler i sediment-suspensjoner kan i varierende grad feste seg til egg og redusere oppdriften (Page 2014a). Endringer i oppdrift kan føre til økt eggpredasjon langs bunnen (Westerberg m.fl. 1996) og til klekking under suboptimale forhold (Zeldis m.fl. 1995, Moser m.fl. 1997, Murdock m.fl. 1990). Fiskelarver er generelt mindre tolerante mot suspenderte sedimenter enn egg av samme art, og eksponering for partikler kan resultere i redusert vekst og kvelning ved at gjellene går tett (Morgan m.fl. 1983). Vanlig uer og snabeluer vil være ekstra sårbare ettersom de føder levende larver som vil påvirkes fra første dag. Suspenderte sedimenter kan også påvirke voksen fisk ved å redusere synligheten av pelagiske byttedyr og tette gjeller med tilhørende fysiologisk stress og reduksjon i vekst og reproduktiv kondisjon (Page 2014b, 2014c). Unnvikelsesadferd er påvist for blant annet torsk selv ved lave sediment-suspensjoner (Westerberg m.fl. 1996).
Hoppekrepsen Calanus er en viktig del av dietten for mange fiskelarver. Calanus overvintrer i dypet i Norskehavet og vil derfor være utsatt for effektene av planlagt mineralvirksomhet. Ulike stadier av Calanus utgjør en viktig del av dietten til larvene for arter som blåkveite (Simonsen m.fl. 2006), vanlig uer og snabeluer (Anderson 1994, González m.fl. 2001). Som nevnt ovenfor i kapitlet om det pelagiske økosystemet kan Calanus spise mineralpartikler og derved være en vektor for bioakkumulering av metaller og kjemikalier knyttet til mineralutvinningen. Effekter av mineralvirksomhet på arter av Calanus kan derfor potensielt ha negative effekter for tidlige livsstadier hos flere bunnlevende arter.
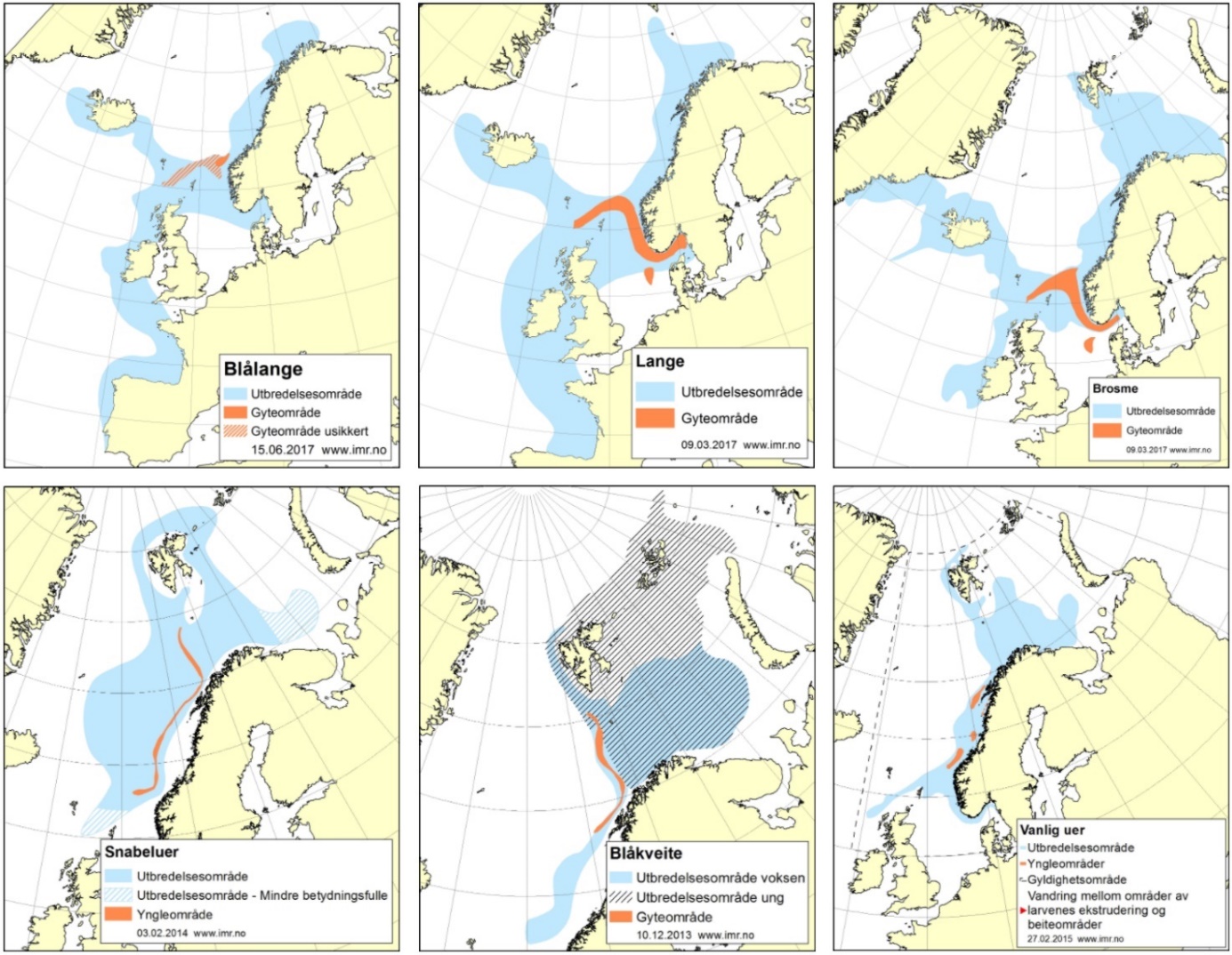
Figur 4: Utbredelses- og gyte/yngelområder for aktuelle dyphavsarter. Ytterligere informasjon er tilgjengelige på https://www.hi.no/hi/temasider/arter/.
8.3 - Frigjøring av partikler, tungmetaller og kjemikalier
Ifølge en systematisk gjennomgang av Elbeshti (2018) kan tungmetaller påvirke vekst, utvikling, reproduksjon og overlevelse av fisk ved å påvirke fysiologiske, biokjemiske, metabolske, systemiske og genetiske funksjoner. Effektene kan variere mellom arter, alder, kjønn, størrelse, fôrvalg og habitat for fisk, vannets fysiske og kjemiske egenskaper, og med det aktuelle tungmetallets egenskaper. Tungmetaller akkumuleres over tid, og ettersom mange dyphavsarter blir svært gamle, vil utslipp av tungmetaller være ekstra problematisk. Allerede i dag eksisterer det kostholdsråd (advarsler) mot å spise ulike typer sjømat på grunn av høye nivåer av tungmetaller og andre uønskede stoffer. Advarsler mot å spise fisk og annen sjømat er ofte knyttet til forurensing fra industriell virksomhet i fjorder, men også ute i Norskehavet på Sklinnadjupet er det uønsket høyt og uforklarlig innhold av kvikksølv i kveite (Nilsen m.fl. 2016). Med tanke på mattrygghet må det utredes i hvilken grad mineralutvinning i Norskehavet kan øke nivåer av tungmetaller i organismer som allerede kan ha høye verdier fra naturlige kilder. Overskridelser av grenseverdier leder til advarsler mot å spise den aktuelle fisken, forbud mot eksport og kan også lede til stenging av fiskeområder, jfr. Atlantisk kveite ved Sklinnadjupet i 2017 basert på rapporten fra Nilsen m.fl. (2016).
Utvinningen av sulfider har potensial til å generere svovelsyre på havbunnen (Bilenker m.fl. 2016). Eksperimenter indikerer at produksjonen av syrer fra slik gruvedrift ikke overstiger bufferkapasiteten til sjøvannet. Det er imidlertid ikke klart om effekten av havforsuring på grunn av klimaendringer kan forsterkes lokalt ved utslipp av syre gjennom gruvedrift (Christiansen m.fl. 2020).
8.4 - Støy, rystelser og bruk av lys
Menneskeskapt støy kan forstyrre den naturlige bruken av lyd, enten ved å maskere biologisk relevante lyder, eller ved å utløse falske svar. Siden lydutbredelse under vann (særlig ved lave frekvenser) når veldig langt, kan støy fra gruvedrift bevege seg over store avstander.
En studie av effekten av undervannskanoner påviste økt svømmeaktivitet og redusert spiseaktivitet hos blåkveite, uer og torskefisk (Normandeau Associates Inc. 2012). En slik følsomhet for lyd kan potensielt også påvirke reproduksjon hos uer-arter ettersom observasjoner fra tokt viser at uer kan slippe larvene prematurt, noe som sannsynligvis skyldes stress (pers.komm. Hannes Höffle, HI).
Mange fiskearter samles på konsentrerte områder for å gyte, hvor de viser en kompleks reproduktiv oppførsel med visuelle og akustiske elementer (de Jong m.fl. 2020, Hawkins & Picciulin 2019, Amorim m.fl. 2015). Menneskeskapt støy kan kamuflere disse lydene og dermed ha en negativ innvirkning på reproduksjonen. Lydproduserende fisk vil sannsynligvis være spesielt sårbar for effekten av menneskeskapt lyd. Lydproduserende strukturer finnes hos mange gadoider som lange, blålange og brosme som alle gyter i nærheten av utredningsområdet. Det er også påvist hos en rekke uer-arter av slekten Sebastes, og menneskeskapt støy kan derfor være et problem for snabeluer hvor reproduksjon foregår i områder som overlapper med østlige deler av utredningsområdet.
Informasjon om hørselsevnene til bruskfisk er begrenset til bare noen få av hundrevis av eksisterende arter. Vår generelle forståelse antyder et relativt smalt hørselsområde med forholdsvis dårlig følsomhet, spesielt sammenlignet med mange beinfisker. Denne mangelen på kunnskap gjør det vanskelig å evaluere de potensielle effektene som kan være forbundet med eksponering for menneskeskapt støy (Casper & Popper 2010).
Kunstig lys kan påvirke fiskenes atferdsmessige respons i betydelig grad, spesielt for dypvannsarter som er tilpasset et liv i mørket. Kunstig lys kan føre til økt dødelighet ved å tiltrekke fisk til utvinningsområdene. Kunstig lys har vist seg å påvirke spiseadferd og stimeadferd, romlig fordeling, predasjonsrisiko, migrasjon og reproduksjon (Nightingale m.fl. 2006). Lys vil tiltrekke seg stimfisk og større predatorfisk som jakter visuelt, og økt overlevelse av predator-arter kan gi unaturlige «top-down»-reguleringer av fiskepopulasjoner (Becker m.fl. 2013).
9 - Pelagisk fisk
Flere pelagiske fiskearter benytter Norskehavet og farvannet rundt Jan Mayen som beiteområde (figur 5). Dette gjelder blant annet viktige kommersielle arter som norsk vårgytende sild, kolmule, makrell og lodde. Disse bestandene har en vid utbredelse som delvis overlapper med utredningsområdet for mineralutvinning, og de er gjenstand for et flernasjonalt fiskeri. Artene beiter på ulike typer plankton, der blant annet raudåte er en nøkkelart.
I høringsdokumentet hevdes det at siden de aktuelle områdene for mineralvirksomhet ligger langt fra kysten, vil aktivitet knyttet til kartlegging og utvinning ikke påvirke andre aktiviteter «som fiskerier, skipsfart, petroleumsvirksomhet og fornybar energiproduksjon». En slik antakelse overser at det er en betydelig flernasjonal fiskeriaktivitet både i norsk økonomisk sone (NØS), internasjonalt farvann i Norskehavet utenfor NØS, i fiskevernsonen rundt Svalbard og i Jan Mayen-sonen.
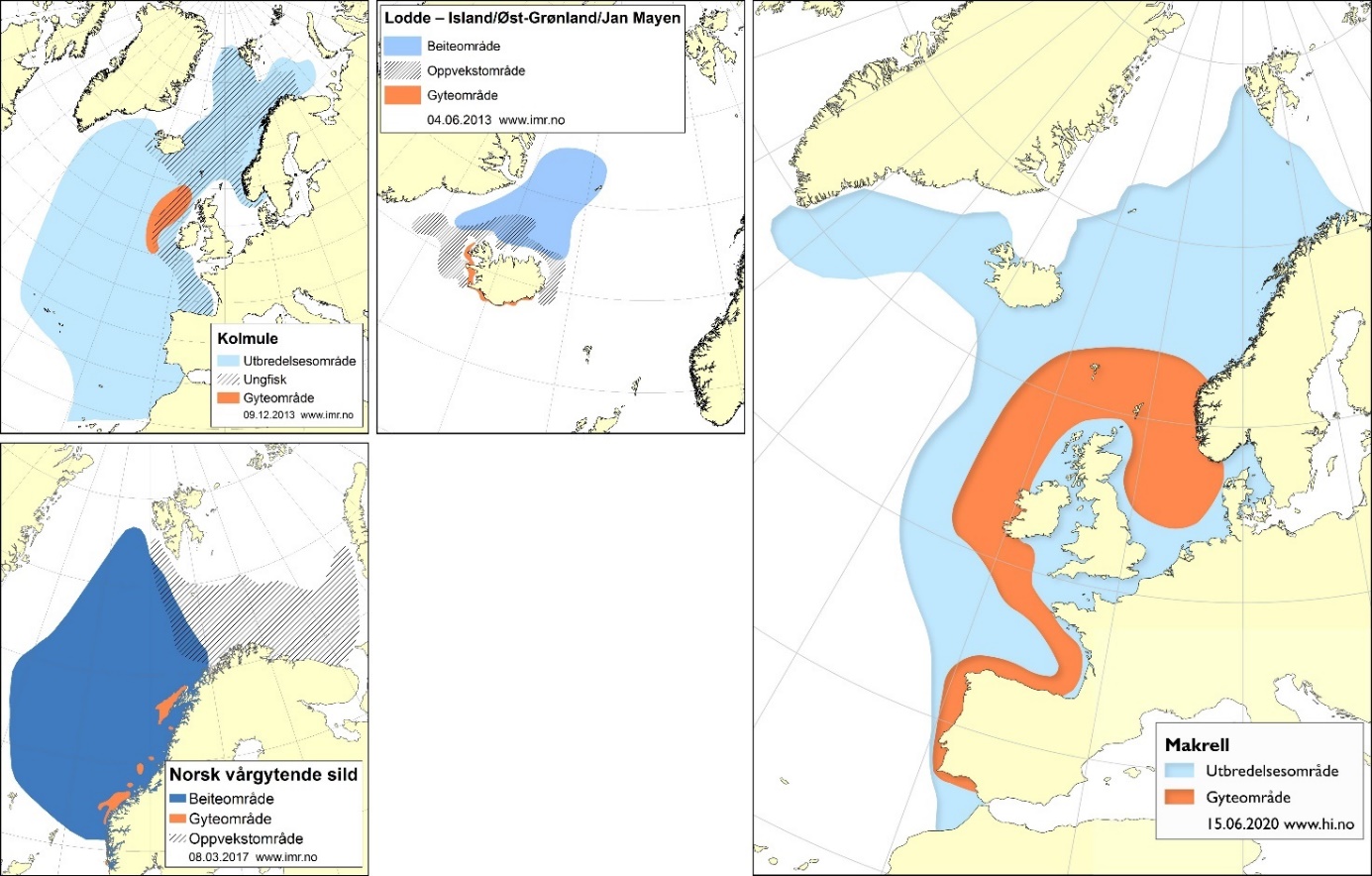
Figur 5: Utbredelses- og gyte/yngelområder for aktuelle pelagiske arter. Ytterligere informasjon er tilgjengelige på https://www.hi.no/hi/temasider/arter/.
Det finnes også ressurser av ulike arter med mesopelagisk fisk i Norskehavet, men størrelsen av disse bestandene er ikke kjent. To viktige arter er laksesild og lysprikkfisk, og registreringene av mesopelagisk fisk skjer ofte fra 200 til 1000 m dyp. Artene beiter på plankton der raudåte er et viktig byttedyr. Den økologiske betydningen av mesopelagisk fisk i Norskehavet er ikke fullt ut forstått, blant annet på grunn av datamangel. Nordlig lysprikkfisk fra Norskehavet hadde konsentrasjoner av kadmium over grenseverdien som gjelder for muskelkjøtt av fisk til mat (Wiech m.fl. 2018). Tilsvarende ble det funnet at konsentrasjonen av bly i liten laksetobis fra Norskehavet var over grenseverdien som gjelder for muskelkjøtt av fisk til mat.
Mineralutvinning på havbunnen kan medføre utslipp av mineralpartikler i vannsøylen som gjennom opptak i plankton kan bli tilgjengelig for pelagiske og mesopelagiske fiskearter. Utslipp av metallholdige partikler med eventuelle kjemikalier fra overflateenheter knyttet til mineralutvinningen kan spres over større områder. Bioakkumulering av metaller og andre uønskede stoffer som følge av slike utslipp bør derfor være en viktig del av en konsekvensutredning.
10 - Sjøpattedyr
10.1 - Bardehvaler
Sommertelletokter for vågehval innenfor utredningsområdet over perioden 1995-2018 har vist relativt høy forekomst av vågehval, finnhval og knølhval rundt Mohns-ryggen (Øien 2009, Leonard & Øien 2020a, 2020b, NAMMCO 2021). Alle disse artene beiter primært pelagisk på krepsdyr (typisk krill) og småfisk som lodde, sild og polartorsk. Mohns-ryggen forbinder områder med høy tetthet av bardehvaler i Danmarksstredet, vestkysten av Spitsbergen og lenger øst og nord. Området ser dermed ut til å være et gjennomreiseområde, men kan også i seg selv være et viktig beiteområde. Observasjoner av både finn- og knølhval i perioden 1995-2001 antyder også en alternativ vandringsrute fra området ved Jan Mayen til eggakanten inn mot Barentshavet via Vøringutstikkeren og den nordlige kanten av Vøringplatået (Øien 2009). Vågehval og finnhval er i tillegg observert i relativt store tettheter i områdene rundt Knipovich-ryggen (Øien 2009, Leonard & Øien 2020a, 2020b, NAMMCO 2021).
I perioden 2014-2018 ble det også gjort flere observasjoner av blåhval i dette området (Leonard & Øien, 2020b). Blåhvalen beiter nesten utelukkende på dyreplankton, og er fortsatt på et historisk lavt nivå etter hvalfangsten med en reproduserende bestand på rundt 1000 individer i Nordøst Atlanteren som helhet, med hovedtyngden i fordeling vest for Island (Pike m.fl. 2009).
Den kritisk truede Spitsbergenbestanden av grønlandshval har historisk sett forekommet i utredningsområdet. På grunn av havisens tilbaketrekking er dette imidlertid blitt svært sjeldent de siste tiårene. Den vestlige delen av Framstredet er et viktig leveområde for grønlandshval både sommer og vinter (Kovacs m.fl. 2020). Veldig aktiv vokalisering («synging») av grønlandshval i dette området om vinteren tyder på at det er et viktig parringsområde for Spitsbergenbestanden (Ahonen m.fl. 2017, Boertmann m.fl. 2021).
10.2 - Tannhvaler
De mest utbredte tannhvalarter innen utredningsområdet er spermhval, nebbhval og spekkhoggere (Øien 2009, Leonard & Øien 2020a, 2020b). De første to artene er dypdykkere. Spermhvalen observeres relativt ofte i de sørlige og østlige deler av utredningsområdet langs Vøringsutstikkeren, sørlige kanten av Vøringplatået, rundt Mohns- og Knipovich-ryggene samt ved eggakanten inn mot Barentshavet. Noen observasjoner er også gjort rundt Jan Mayen. Nebbhval er primært blitt observert rundt Jan Mayen og langs Vøringutstikkeren. Historisk har nebbhvalen hatt høy forekomst ved eggakanten vest for Svalbard og på Knipovich-ryggen (Øien & Hartvedt 2011), og den er også sett her i nyere tid (Storrie m.fl. 2018).
Diettdata for spermhval i Nordøstatlanten finnes bare fra hvalfangsten (Martin & Clarke 1986, Christensen m.fl. 1992) og fra strandete dyr (Santos m.fl. 1999, Santos m.fl. 2002). Blekkspruten Gonatus fabricii dominerer mageinnholdet hos spermhvaler, men også fisk (hovedsakelig rognkjeks) er funnet i spermhval fanget utenfor Østgrønland (Martin & Clarke 1986). Hos nebbhval i Nordøstatlanten dominerer Gonatus (Fernandez m.fl. 2014). I tillegg er det observert inntak av bunntilknyttet fisk som uer, blåkveite, havmus, lange og pigghå samt bentiske organismer som sjøpølser, sjøstjerner og reker ( Benjaminsen & Christensen 1979, Reeves m.fl. 1993, Whitehead m.fl. 2021). Kunnskapen om diett hviler imidlertid på et veldig begrenset datagrunnlag. Lokalisering av kalvingsområdene er heller ikke kjent for nebbhvalen. Det er relativt høy forekomst av spekkhogger i den sørvestlige delen av utredningsområdet rundt Jan Mayen, langs Vøringutstikkeren og i området rundt Mohns-ryggen (Leonard & Øien 2020a, 2000b). En del observasjoner er også gjort ut forbi eggakanten vest for Spitsbergen (Storrie m.fl. 2018). Spekkhoggerne i Norskehavet menes å beite hovedsakelig på pelagisk fisk som sild og makrell (Jourdain m.fl. 2019).
10.3 - Seler
Isutbredelsen rundt Jan Mayen har trukket seg vestover, og kaste- og hårfellingsområdene hos bestandene av grønlandssel og klappmyss i Grønlandshavet er nå utenfor utredningsområdet. Satellittmerkingsdata fra 1990 tallet viser imidlertid at voksne grønlandsseler krysser den nordlige delen av utredningsområdet på vei til og fra Barentshavet. Også Grønlandssel fra Barentshavet forekommer av og til i den nordlige delen av utredningsområdet (Nordøy m.fl. 2008). Grønlandssel beiter primært på krill og amfipoder fra vår til høst og på småfisk som lodde og polartorsk om vinteren (Haug m.fl. 2004).
Klappmyss er klassifisert som sterkt truet i Norsk rødliste. I motsetning til grønlandssel er klappmyss en dypdykkende art som kan gå ned til flere hundre meter under beiting (Folkow & Blix 1999, Folkow m.fl. 2010). Beiteområdene er langs kontinentalskråninger og undervannsrygger i hele Norskehavet og til dels lenger sør og nord (Folkow m.fl. 1996, Folkow & Blix 1999, Folkow m.fl. 2010). Studier av beiteintensitet antyder at Vøringutstikkeren, Knipovich-ryggen og overgangen til Mohns-ryggen kan være blant de viktigste beiteområder for bestanden av klappmyss fra Grønlandshavet (Vacquie-Garcia m.fl. 2017). Dietten hos klappmyss nær iskanten kan bestå av blekkspruten Gonatus, polartorsk, amfipoder og lodde (Haug m.fl. 2007), men fisk som uer, blåkveite, ulkefisker, torsk og sild er også funnet i magene hos klappmyss andre steder enn i utredningsområdet (Hammill & Stenson 2000, Haug m.fl. 2007, Enoksen m.fl. 2017). Registrering av vibrasjoner ved hjelp av værhårene er sannsynligvis viktig for at seler skal finne bytte, i tillegg til registrering av lyder og undervannslys i form av bioluminescens.
10.4 - Sårbarhet for menneskelig aktivitet
Eventuelle påvirkninger av mineralutvinningsaktiviteter på sjøpattedyr vil avhenge av omfang i tid og rom, og anvendt teknologi. Siden dette er en ny type marin aktivitet der disse forholdene i stor grad er ukjent, bør konsekvensutredningen presentere et vidt spekter av scenarier som belyser mulige påvirkninger for sjøpattedyrene i utredningsområdet. Siden det både er betydelig kunnskapsmangel om sjøpattedyrenes fordeling og aktivitet i utredningsområdet og uklart hvilke forstyrrelser mineralvirksomheten kan medføre, må man i stor utstrekking bygge vurderinger fra generell biologisk kunnskap og erfaringer fra andre typer av menneskeskapte forstyrrelser til havs.
Dessverre er den generelle økologiske kunnskapen vedrørende områdebruken, dietten og biologien til mange av de relevante sjøpattedyrene svært sparsom. Dette gjelder særlig de dypdykkende artene klappmyss, spermhval og nebbhval, som er blant de vanskeligste å studere, men som også er artene som er tettest knyttet til utredningsområdet. Alle disse artene har allerede betydelige utfordringer. Dette gjelder spesielt bestanden av klappmyss i Grønlandshavet som er på et historisk lavmål (Øigård m.fl. 2014). Lite er kjent om klappmyssens reaksjoner på forskjellige forstyrrelser, men eksperimenter med mellomfrekvente militære sonarer kan tyde på at menneskeskapt lyd kan forstyrre dykkeadferden (Kvadsheim m.fl. 2010). Dette gjelder kanskje i enda større grad for nebbhval, som har vist seg å respondere med en voldsom fluktrespons på militære sonarer (Miller m.fl. 2015). Spermhval ser imidlertid ut til å være mindre følsom overfor lav og mellomfrekvent menneskeskapt støy enn nebbhval, vågehval og spekkhogger (Miller m.fl. 2012, Kvadsheim m.fl. 2020).
Spekkhogger er veldig sensitiv mot visse typer av støy, men kan i andre tilfeller beite i områder med mye aktivitet av fiskebåter og andre fartøyer (Kvadsheim m.fl. 2020). Generelt har man funnet at hvalers responser på menneskeskapt støy varierer i forhold til viktigheten av hvalens aktivitet i et område. Det er mindre sannsynlig at hvalene avbryter beiting på grunn av støy enn at de for eksempel velger en annen rute for forflytting mellom områder (Kvadsheim m.fl. 2020). Selv om hvalene velger å bli i områder med støy, har målinger av hormonnivåer vist at de kan ha økt stressnivå i støyende omgivelser (Rolland m.fl. 2012).
Bardehvaler kommuniserer ved hjelp av mellom- og lavfrekvente lyder og må ofte modifisere enten frekvens, hyppighet eller styrke på sine lydsignaler for å kompensere for menneskeskapt støy (Kvadsheim m.fl. 2020). Vågehval synes å være en spesiell sensitiv art, som ofte kan unngå det eksponerte område også etter at aktiviteten er opphørt (Kvadsheim m.fl. 2020, Miller m.fl. 2012). Blåhvalen er den eneste bardehvalen som «synger» noe særlig om sommeren under sitt beiteopphold i Norskehavet (Ahonen m.fl. 2017). Dette kan tyde på at lydkommunikasjon er særlig viktig for denne arten, også under beiting.
Høye byttedyrkonsentrasjoner er viktig for alle sjøpattedyrarter, ikke minst de store bardehvalene som foretar lange vandringer til yngle- og parringsområder i mindre produktive farvann nær ekvator. Dette har tidligere vært antatt å gjelde stort sett alle bardehvaler, men i de senere år er det dokumentert at en del finnhvaler og knølhvaler kan bli igjen i de nordiske hav utover vinteren (Gunnlaugsson & Vikingsson 2014, Magnusdottir m.fl. 2012). Lite er kjent om antall, fordeling og adferd på denne tid av året. Det er imidlertid dokumentert noe som ligner parringssang for begge arter (Magnusdottir m.fl. 2012, Ahonen m.fl. 2017). Tidlige studier av knølhvalfostre under hvalfangstperioden i norske farvann antydet til og med at det kunne forekomme kalving i nordlige farvann (Ingebrigtsen 1929). Dette er veldig grunnleggende aspekter ved biologien som er ukjent for en del viktige arter i utredningsområdet, og som dermed øker usikkerheten på vurderinger av effekter av økt menneskelig aktivitet her.
11 - Referanser
Ahonen, H., Stafford; K.M., De Steur, L., Lydersen, C., Wiig, Ø. & Kovacs, K. (2017). The underwater soundscape in western Fram Strait: Breeding ground of Spitsbergen’s endangered bowhead whales. Marine Pollution Bulletin 123: 97-112. https://doi.org/10.1016/j.marpolbul.2017.09.019
Aksnes, D.L., Dupont, N., Staby, A., Fiksen, Ø., Kaartvedt, S. & Aure, J. (2009). Coastal water darkening and implications for mesopelagic regime shifts in Norwegian fjords. Marine Ecology Progress Series 387: 39-49. https://doi.org/10.3354/meps08120
Amorim, M.C.P., Vasconcelos, R. & Fonseca, P. J. (2015). Fish sounds and mate choice. In : Ladich F. (ed) Sound Communication in Fishes . Animal Signals and Communication 4. 1-33. Springer, Vienna. https://doi.org/10.1007/978-3-7091-1846-7_1
Anderson, J.T. (1994). Feeding ecology and condition of larval and pelagic juvenile redfish Sebastes spp.. Marine Ecology Progress Series 104: 201-226. https://doi.org/10.3354/MEPS104211
Arkhipkin, A.I. & Bjørke, H. (1999). Ontogenetic changes in morphometric and reproductive indices of the squid Gonatus fabricii (Oegopsida, Gonatidae) in the Norwegian Sea. Polar Biology 22: 357-3б5. https://doi.org/10.1007/s003000050429
Artsdatabanken (2018). Norsk rødliste for naturtyper 2018. 26.10.2020. https://www.artsdatabanken.no/rodlistefornaturtyper
Attramadal, Y.G, Fosså, J.H. & Nilsson, H.L. (1985). Changes in behaviour and eye-morphology of Boreomysis megalops G.O. Sars (Crustacea: Mysidacea) following exposure to short periods of artificial and natural daylight. Journal of Experimental Marine Biology and Ecology 85: 135-148. https://doi.org/10.1016/0022-0981(85)90139-X
Bagøien, E., Melle, W. & Kaartvedt, S. (2012). Seasonal development of mixed layer depths, nutrients, chlorophyll and Calanus finmarchicus in the Norwegian Sea – A basin-scale habitat comparison. Progress in Oceanography 103: 58-79. https://doi.org/10.1016/j.pocean.2012.04.014
Becker, A., Whitfield, A.K., Cowley, P.D. & Järnegren J. (2013). Potential effects of artificial light associated with anthropogenic infrastructure on the abundance and foraging behavior of estuary‐associated fishes. Journal of Applied Ecology 50: 43-50. https://doi.org/10.1111/1365-2664.12024
Benjaminsen, T. & Christensen, I. (1979). The natural history of the bottlenose whale, Hyperoodon ampullatus (Forster). In : Winn, H.E. & Olla, B.L. (eds.). Behavior of Marine Animals. Springer, Boston, MA, USA. Pp 143-164. https://doi.org/10.1007/978-1-4684-2985-5_5
Bilenker, L.D., Romano G.Y. & McKibben, M.A. (2016). Kinetics of sulfide mineral oxidation in seawater: Implications for acid generation during in situ mining of seafloor hydrothermal vent deposits. Applied Geochemistry 75: 20-31. https://doi.org/10.1016/j.apgeochem.2016.10.010
Bjørke, H. & Gjøsæter, H. (1998). Who eats the larger Gonatus fabricii (Lichtenstein) in the Norwegian Sea? ICES CM 1998/M:10 (in mimeo). https://imr.brage.unit.no/imr-xmlui/bitstream/handle/11250/100387/M10-1998.pdf?sequence=3&isAllowed=y
Boertmann, D., Blockley, D. & Mosbech, A. (2020). Greenland Sea – an updated strategic environmental impact assessment of petroleum activities. 2nd revised edition. Scientific report from DCE – Danish Centre for Environment and Energy No. 375, 380 pp. http://dce2.au.dk/pub/SR375.pdf
Burgos J, Buhl-Mortensen, L., Buhl-Mortensen, P., Ólafsdóttir, S.H., Steingrund, P., Ragnarsson, S.Á. & Skagseth, Ø. (2020). Predicting the distribution of Indicator Taxa of Vulnerable Marine Ecosystems in the Arctic and Sub-arctic Waters of the Nordic Seas. Frontiers in Marine Science 7: 131. https://doi.org/10.3389/fmars.2020.00131
Casper, B. & Popper, A.N. (2010). Anthropogenic noise: Is this an issue for elasmobranch fishes? The Journal of the Acoustical Society of America 127: 1753. https://doi.org/10.1121/1.3383688
Cathalot, C., Van Oevelen, D., Cox, T.J.S., Kutti, T., Lavaleye, M., Duineveld, G. & Meysman, F.J.R. (2015). Cold-water coral reefs and adjacent sponge grounds: hotspots of benthic respiration and organic carbon cycling in the deep sea. Frontiers in Marine Science 2: 37. https://doi.org/10.3389/fmars.2015.00037
Chan, A.T. & Anderson, G.C. (1981). Environmental investigation of the effects of deep-sea mining on marine phytoplankton and primary productivity in the tropical Eastern North Pacific Ocean. Marine Mining 3: 121-149.
Chen, J.H., Lion, L.W., Ghiorse, W.C. & Shuler, M.L. (1995). Mobilization of adsorbed cadmium and lead in aquifer material by bacterial extracellular polymers. Water Research 29: 421-430. https://doi.org/10.1016/0043-1354(94)00184-9
Christensen, I., Haug, T. & Øien, N. (1992). Seasonal distribution, exploitation and present abundance of stocks of large baleen whales (Mysticeti) and sperm whales ( Physeter Macrocepahlus ) in Norwegian and adjacent waters. ICES Journal of Marine Science 49: 341-355. https://doi.org/10.1093/icesjms/49.3.341
Christiansen, B., Denda, A. & Christiansen, S. (2020). Potential effects of deep seabed mining on pelagic and benthopelagic biota. Marine Policy 114: 103442. https://doi.org/10.1016/j.marpol.2019.02.014
Conover, R.J. (1988). Comparative life histories in the genera Calanus and Neocalanus in high latitudes of the northern hemisphere. Hydrobiologia 167: 127-142. https://doi.org/10.1007/BF00026299
de Jong, K., Forland, T.N., Amorim, M.C.P. Rieucau, G., Slabbekoorn, H. & Sivle, L.D. (2020). Predicting the effects of anthropogenic noise on fish reproduction. Reviews in Fish Biology and Fisheries 30: 1-24. https://doi.org/10.1007/s11160-020-09598-9
Doughty, C.L., Quattrini, A.M. & Cordes, E.E. (2014). Insights into the population dynamics of the deep-sea coral genus Paramuricea in the Gulf of Mexico . Deep-Sea Res. Part II: Topical Studies in Oceanography 99: 71-82. http://doi.org/10.1016/j.dsr2.2013.05.023
Drazen, J.C., Smith, C.R., Gjerde, K.M., Haddock, S.H.D., Carter, G.S., Choy, C.A., Clark, M.R., Dutrieux, P., Goetze, E., Hauton, C., Hatta, M., Koslow, J.A., Leitner, A.B., Pacini, A., Perelman, J.N., Peacock, T., Sutton, T.T., Watling, L. & Yamamoto, H. (2020). Midwater ecosystems must be considered when evaluating environmental risks of deep-sea mining. PNAS 117: 17455-17460. www.pnas.org/cgi/doi/10.1073/pnas.2011914117
Dunn, D., Van Dover, C.L., Etter, R.J., Smith, C.R., Levin, L.A., Morato, T., Colaco, A., Dale, A., Gebruk, A.V., Gjerde, K., Halpin, P.N., Howell, K.L., Johnson, D., Perez, J.A., Ribeiro, M.C., Stuckas, H. & Weaver, P. (2018). A strategy for the conservation of biodiversity on mid-ocean ridges from deep-sea mining. Science Advances 4: eaar4313. https://doi.org/10.1126/sciadv.aar4313
Eiane, K., Aksnes, D.L., Bagøien, E. & Kaartvedt S. (1999). Fish or jellies - a question of visibility? Limnology and Oceanography 44: 1352-1357. https://doi.org/10.4319/lo.1999.44.5.1352
Elbesheti, R.T.A., Elderwish, N.M., Abdelali, K.M.K. & Tastan, Y. (2018). Effects of Heavy Metals on Fish. Menba Journal of Fisheries Faculty 4: 36-47. https://dergipark.org.tr/en/download/article-file/506762
Enoksen, S, Haug, T., Lindstrøm, U. & Nilssen, K.T. (2016). Recent summer diet of hooded Cystophora cristata and harp Pagophilus groenlandicus seals in the drift ice of the Greenland Sea. Polar Biology 40: 931-937. https://doi.org/10.1007/s00300-016-2002-2
Etnoyer, P. (2010). Deep-sea corals on seamounts. Oceanography 23: 128-129. https://doi.org/10.5670/oceanog.2010.91
EU 2011/696: Kommissionens henstilling af 18. oktober 2011 om definitionen af nanomaterialer (EØS-relevant tekst). https://eur-lex.europa.eu/legal-content/DA/TXT/?uri=CELEX%3A32011H0696
EU 2018/1881: Kommisjonsforordning 2018/1881 av 3. desember 2018 om endring av vedlegg I, III, VI, VII, VIII, IX, X, XI og XII til europaparlaments- og rådsforordning (EF) nr. 1907/2006 om registrering, vurdering og godkjenning av samt begrensninger for kjemikalier (REACH) for å ta hensyn til nanoformer av stoffer. https://lovdata.no/static/NLX3/32018r1881.pdf
Fang, J.K.H., Rooks, C.A., Krogness, C.M., Kutti, T., Hoffmann F. & Bannister, R.J. (2018). Impact of particulate sediment, bentonite and barite (oil-drilling waste) on net fluxes of oxygen and nitrogen in Arctic-boreal sponges. Environmental Pollution 238: 948-958. https://doi.org/10.1016/j.envpol.2017.11.092
Farkas, J., Altin, D., Hammer, K.M., Hellstrøm, K.C., Booth, A.M. & Hansen, B.H. (2017). Characterisation of fine-grained tailings from a marble processing plant and their acute effects on the copepod Calanus finmarchicus . Chemosphere 169: 700-708. https://doi.org/10.1016/j.chemosphere.2016.11.118
Fernández, R., Pierce, G.J., MacLeod, C.D., Brownlow, A., Reid, R.J., Rogan, E., Addink, M., Deaville, R., Jepson, P.D. & Santos, M.B. (2014). Strandings of northern bottlenose whales, Hyperoodon ampullatus , in the north-east Atlantic: seasonality and diet. Journal of the Marine Biological Association of the United Kingdom 94: 1109-1116. https://doi.org/10.1017/S002531541300180X
Freiwald, A., Beuck, L. & Wisshak, M. (2012). Korallenriffe im kalten Wasser des Nordatlantiks – Entstehung, Artenvielfalt und Gefährdung. In : Beck, E. (ed.) Die Vielfalt des Lebens. Wiley-VCH, Weinheim, Germany. Pp. 89-96.
Folkow, L.P., Mårtensson, P.E. & Blix, A.S. (1996). Annual distribution of hooded seals ( Cystophora cristata ) in the Greenland and Norwegian seas. Polar Biology 16: 179-189. https://doi.org/10.1007/BF02329206
Folkow, L.P. & Blix, A.S. (1999). Diving behaviour of hooded seals ( Cystophora cristata ) in the Greenland and Norwegian Seas. Polar Biology 22: 61-74. https://doi.org/10.1007/s003000050391
Folkow, L.P., Nordøy, E.S. & Blix, A.S. (2010). Remarkable development of diving performance and migrations of hooded seals ( Cystophora cristata ) during their first year of life. Polar Biology 33: 433-441. https://doi.org/10.1007/s00300-009-0718-y
Fortunato, C.S., Larson, B., Butterfield, D.A. & Huber, J.A. (2018). Spatially distinct, temporally stable microbial populations mediate biogeochemical cycling at and below the seafloor in hydrothermal vent fluids. Environmental Microbiology 20: 769-784. https://doi.org/10.1111/1462-2920.14011
Fossum, P., Mork, K.A., Tverberg, V., Boitsov, S., Heldal, H.E., Bagøien, E., Rønning, J., Slotte, A., Vollen, T., Wienerroither, R., Dahle, G., Buhl-Mortensen, P., Øien, N., Øigård, T.A. & Fauchald, P. (2012). Beskrivelse av miljø og levende marine ressurser i havområdene ved Jan Mayen. Rapport fra Havforskningen 16-2012. 119 pp. https://www.hi.no/resources/publikasjoner/rapport-fra-havforskningen/2012/hi-rapp_16-2012_rap.pdf
Fuchida, S., Yokoyama, A., Fukuchi, R., Ishibashi, J.-i., Kawagucci, S., Kawachi, M. & Koshikawa, H. (2017). Leaching of metals and metalloids from hydrothermal ore particulates and their effects on marine phytoplankton. ACS Omega 2: 3175-3182. https://doi.org/10.1021/acsomega.7b00081
Grefsrud, E.S., Svåsand, T., Glover, K., Husa, V., Hansen, P.K., Samuelsen, O., Sandlund, N. & Stien, L.H. (2019). Risikorapport norsk fiskeoppdrett 2019. Fisken og Havet 2019-5. 115 pp. https://www.hi.no/hi/nettrapporter/fisken-og-havet-2019-5#sec-11
Gomiero, A., Haave, M., Kögel, T., Bjorøy, Ø., Gjessing, M., Berg Lea, T., Horve, E., Martins, C. & Olafsen, T. (2020). Tracking of Plastic emissions from aquaculture industry. NORCE Report n. 4/2020 https://norceresearch.brage.unit.no/norceresearch-xmlui/handle/11250/2649891
González, C., Bruno, I. & Paz, X. (2001). Food and feeding of deep-sea redfish ( Sebastes mentella Travin) in the North Atlantic. NAFO Scientific Council Studies 10: 89-101. https://archive.nafo.int/open/studies/s33/gonzalez.pdf
Guilhon, M., Montserrat, F. & Turra, A. (2020). Recognition of ecosystem-based management principles in key documents of the seabed mining regime: implications and further recommendations. ICES Journal of Marine Science : fsaa229. https://doi.org/10.1093/icesjms/fsaa229
Gunnlaugsson, T.H. & Vikingsson, G.A. (2014). Winter occurrence of whales in waters around Iceland. Paper presented to the IWC Scientific Committee. IWC SC/65b/RMP06 (in mimeo). (available at https://archive.iwc.int/pages/search.php?search=!collection158&bc_from=themes)
Halvorsen, E., Tande, K.S., Edvardsen, A., Slagstad, D. & Pedersen, O.P. (2003). Habitat selection of overwintering Calanus finmarchicus in the NE Norwegian Sea and shelfwaters off Northern Norway in 2000-02. Fisheries Oceanography 12: 339-351. https://doi.org/10.1046/j.1365-2419.2003.00255.x
Hammill, M.O. & Stenson G.B. (2000). Estimated prey consumption by harp seals ( Phoca groenlandica ), hooded seals ( Cystophora cristata ), grey seals ( Halichoerus grypus ) and harbour seals ( Phoca vitulina ) in Atlantic Canada. Journal of Northwest Atlantic Fishery Science 26: 1-23. https://doi.org/10.2960/J.v26.a1
Hansen, I.V., Weeks, J.M. & Depledge, M.H. (1995). Accumulation of copper, zinc, cadmium and chromium by the marine sponges Halichondria panicea Pallas and the implications for biomonitoring. Marine Pollution Bulletin 31: 133-138. https://doi.org/10.1016/0025-326X(94)00228-2
Orani, A.M., Barats, A., Vassileva, E. & Thomas, O.P. (2018). Marine sponges as a powerful tool for trace elements biomonitoring studies in coastal environment. Marine Pollution Bulletin 131. 633-645. https://doi.org/10.1016/j.marpolbul.2018.04.073
Hanz, U., Roberts, E.M., Duineveld, G., Davies, A., van Haren, H., Rapp, H.T., Reichart, G.-J. & Mienis, F. (2021). Long-term observations reveal environmental conditions and food supply mechanisms at an Arctic deep‐sea sponge ground. Journal of Geophysical Research: Oceans 126: e2020JC016776. https://doi.org/10.1029/2020JC016776
Haug, T., Nilssen, K.T. & Lindblom, L. (2004). Feeding habits of harp and hooded seals in drift ice waters along the east coast of Greenland in summer and winter. Polar Research 23: 35-42. https://doi.org/10.3402/polar.v23i1.6264
Haug, T., Nilssen, K.T., Lindblom, L. & Lindstrøm, U. (2007). Diets of hooded seals in coastal and drift ice waters along the east coast of Greenland. Marine Biology Research 3: 123-133. https://doi.org/10.1080/17451000701358531
Havforskningsinstituttet (2020). Høringsuttalelse vedrørende revidering av tillatelsen til Sydvaranger Drift AS. Ref. nr. 20/02773. https://www.hi.no/resources/20-02773-Rapport-om-horingsuttalelse-Sydvaranger-11122020.pdf
Hawkins, A.D. & Picciulin, M. (2019). The importance of underwater sounds to gadoid fishes. The Journal of the Acoustical Society of America 146: 3536-3551. https://doi.org/10.1121/1.5134683
Herring, P.J., Gaten, E. & Shelton, P.M.J. (1999). Are vent shrimps blinded by science? Nature 398: 116. https://doi.org/10.1038/18142
Hirche, H.-J. (1996). Diapause in the marine copepod, Calanus finmarchicus - A review. Ophelia 44: 129-143. https://doi.org/10.1080/00785326.1995.10429843
Hirche, H.-J. (1997) Life cycle of the copepod Calanus hyperboreus in the Greenland Sea. Marine Biology 128: 607-618. https://doi.org/10.1007/s002270050127
Hjøllo, S.S., Huse, G., Skogen, M.D. & Melle, W. (2012). Modelling secondary production in the Norwegian Sea with a fully coupled physical/primary production/individual-based Calanus finmarchicus model system. Marine Biology Research 8: 508-526. https://doi.org/10.1080/17451000.2011.642805
Hoffmann, L.J., Breitbarth, E., Boyd, P.W. & Keith A. Hunter (2012). Influence of ocean warming and acidification on trace metal biogeochemistry. Marine Ecology Progress Series 470: 191-205. https://doi.org/10.3354/meps10082
Hook, S.E. & Fisher, N.S. (2001). Reproductive toxicity of metals in calanoid copepods. Marine Biology 138: 1131-1140. https://doi.org/10.1007/s002270000533
Ingebrigtsen A. (1929). Whales caught in the North Atlantic and other seas. Rapports et Procès-verbaux des Réunions, Conseil international pour L’Exploration de la Mer 56: 1-26.
ISA-International Seabed Authority. (2011). Environmental Management Plan for the Clarion-Clipperton Zone. ISBA/17/LTC/7. https://isa.org.jm/files/files/documents/isba-17ltc-7_0.pdf
ISA—International Seabed Authority. (2012). Decision of the Council relating to an environmental management plan for the Clarion-Clipperton Zone. ISBA/18/C/22. https://isa.org.jm/files/files/documents/isba-18c-22_0.pdf
ISA-International Seabed Authority. (2020). Recommendations for the guidance of contractors for the assessment of the possible environmental impacts arising from exploration for marine minerals in the area. ISBA/25/LTC/6/Rev.1. https://isa.org.jm/files/files/documents/26ltc-6-rev1-en_0.pdf
Jourdain, E., Ugarte, F., Vikingsson, G., Samarra, F., Ferguson, S., Lawson, J., Vongraven, D. & Desportes, G. (2019). North Atlantic killer whale Orcinus orca populations: a review of current knowledge and threats to conservation. Mammal Review 49: 384-400. https://doi.org/10.1111/mam.12168
Jroundi, F., Martinez-Ruiz, F., Merroun, M.L. & Gonzalez-Muñoz, M.T. (2020). Exploring bacterial community composition in Mediterranean deep-sea sediments and their role in heavy metal accumulation. Science of The Total Environment 712: 135660. https://doi.org/10.1016/j.scitotenv.2019.135660
Kadiene, E.U., Ouddane, B., Hwang, J.-S. & Souissi, S. (2019) Bioaccumulation of metals in calanoid copepods by oral intake. Scientific Reports 9: 9492. https://doi.org/10.1038/s41598-019-45987-2
Kiørboe, T. & Bagøien, E. (2005). Motility patterns and mate encounter rates in planktonic copepods. Limnology & Oceanography 50: 1999-2007. https://doi.org/10.4319/lo.2005.50.6.1999
Klitgaard, A.B. (1995). The fauna associated with outer shelf and upper slope sponges (Porifera, Demospongiae) at the Faroe Islands, northeastern Atlantic. Sarsia 80: 1-22. https://doi.org/10.1080/00364827.1995.10413574
Knutsen, T. & Serigstad, B. (2001). Potential implications on the pelagic fish and zooplankton community of artificially induced deep-water releases of oil and gas during DeepSpill_2000 - an innovative acoustic approach. Fisken og Havet nr.14. 37 pp. https://www.hi.no/resources/publikasjoner/fisken-og-havet/74739/biology_report.pdf
Kongsrud, J.A., Eilertsen, M.H., Alvestad, T., Kongshavn, K. & Rapp, H.T. (2017). New species of Ampharetidae (Annelida: Polychaeta) from the Arctic Loki Castle vent field. Deep-Sea Res. Part II: Topical Studies in Oceanography 137: 232-245. https://doi.org/10.1016/j.dsr2.2016.08.015
Kongsrud J.A. & Rapp, H.T. (2012). Nicomache (Loxochona) lokii sp. nov. (Annelida: Polychaeta: Maldanidae) from the Loki’s Castle vent field: an important structure builder in an Arctic vent system. Polar Biology 35: 161-170. https://doi.org/10.1007/s00300-011-1048-4
Kovacs, K.M., Lydersen, C., Vacquiè-Garcia, J., Shpak, O., Glazov, D. & Heide-Jørgensen, M.P. (2020). The endangered Spitsbergen bowhead whales’ secrets revealed after hundreds of years in hiding. Biology Letters 16: 20200148. http://dx.doi.org/10.1098/rsbl.2020.0148
Kutti, T., Bannister, R.J. & Fosså, J.H. (2013). Community structure and ecological function of deep-water sponge grounds in the Traenadypet MPA-Northern Norwegian continental shelf. Continental Shelf Ressearch 69: 21-30. https://doi.org/10.1016/j.csr.2013.09.011
Kutti, T., Bannister, R.J., Fosså, J.H., Krogness, C.M., Tjensvoll, I. & Søvik, G. (2015). Metabolic responses of the deep-water sponge Geodia barretti to suspended tailings, drill cuttings and bottom sediment. Journal of Experimental Marine Biology and Ecology 473: 64-72. https://doi.org/10.1016/j.jembe.2015.07.017
Kvadsheim, P.H., Sevaldsen, E.M., Folkow, L. & Blix, A.S. (2010). Behavioural and physiological responses of hooded seals ( Cystophora cristata ) to 1 to 7 kHz sonar signals. Aquatic Mammals 36: 239-247. https://doi.org/10.1578/AM.36.3.2010.239
Lacharité, M. & Metaxas, A. (2013). Early life history of deep-water gorgonian corals may limit their abundance. PLoS ONE 8: e65394. https://doi.org/10.1371/journal.pone.0065394
Leonard, D.M. & Øien, N.I. (2020a). Estimated abundances of cetaceans species in the Northeast Atlantic from two multiyear surveys conducted by Norwegian vessels between 2002-2013. NAMMCO Scientific Publications 11. https://doi.org/10.7557/3.4695
Leonard, D.M. & Øien, N.I. (2020b). Estimated abundances of cetacean species in the Northeast Atlantic from Norwegian shipboard surveys conducted in 2014-2018. NAMMCO Scientific Publications 11. https://doi.org/10.7557/3.4694
Lepland, A., Sæther, O. & Thorsnes, T. (2000). Accumulation of barium in recent Skagerrak sediments: sources and distribution controls. Marine Geology 163: 13-26. https://doi.org/10.1016/S0025-3227(99)00104-8
Levin, L.A., Amon, D.J. & Lily, H. (2020). Challenges to the sustainability of deep-seabed mining. Nature Sustainability 3: 784-794. https://doi.org/10.1038/s41893-020-0558-x
Lodge, M., Johnson, D., Le Gurun, G., Wangler, M., Weaver, P. & Gunn, V. (2014). Seabed mining: International Seabed Authority environmental management plan for the Clarion Clipperton Zone, a partnership approach. Marine Policy 49: 66-72. https://doi.org/10.1016/j.marpol.2014.04.006
Lopes, C.L., Bastos, L., Caetano, M., Martins, I., Santos, M.M. & Iglesias, I. (2019). Development of physical modelling tools in support of risk scenarios: A new framework focused on deep-sea mining. Science of The Total Environment 650: 2294-2306. https://doi.org/10.1016/j.scitotenv.2018.09.351
Magnúsdóttir, E.E., Rasmussen, M.H., Lammers, M.O. & Svavarsson, J. (2014). Humpback whale songs during winter in subarctic waters. Polar Biology 37: 427-433. https://doi.org/10.1007/s00300-014-1448-3
Martin, A.R. & Clarke, M.R. (1986). The diet of sperm whales ( Physeter macrocaphalus ) captured between Iceland and Greenland . Journal of the Marine Biological Association of the United Kingdom 66: 779-790. https://doi.org/10.1017/S0025315400048426
McClain, C.R., Lundsten, L., Barry, J. & DeVogelaere, A. (2010). Assemblage structure, but not diversity or density, change with depth on a northeast Paci fi c seamount. Marine Ecology 31: 14-25. https://doi.org/10.1111/j.1439-0485.2010.00367.x
Melle, W., Ellertsen, B. & Skjoldal, H.R. (2004). Zooplankton: The link to higher trophic levels. . In: Skjoldal, H.R. (Ed.). The Norwegian Sea Ecosystem. Tapir Academic Press, Trondheim, pp. 137-202.
Melle, W., Runge, J., Head, E., Plourde, S., Castellani, C., Licandro, P., Pierson, J., Jonasdottir, S., Johnson, C., Broms, C., Debes, H., Falkenhaug, T., Gaard, E., Gislason, A., Heath, M., Niehoff, B., Nielsen, T.G., Pepin, P., Stenevik, E.K. & Chust, G. (2014). The North Atlantic Ocean as habitat for Calanus finmarchicus : Environmental factors and life history traits. Progress in Oceanography 129: 244-284. https://doi.org/10.1016/j.pocean.2014.04.026
Meyer, H.K., Roberts, E.M., Rapp, H.T., Davies, A.J. (2019). Spatial patterns of arctic sponge ground fauna and demersal fish are detectable in autonomous undervater (AUV) imagery. Deep Sea Research Part I: Oceanographic Research Papers 153: 103137. https://doi.org/10.1016/j.dsr.2019.103137
Miller, P.J.O., Kvadsheim, P.H., Lam, F.P.A., Wensveen, P.J., Antunes, R., Alves, A.C., Visser, F., Kleivane, L., Tyack, P.L. & Sivle, L.D. (2012). The severity of behavioral changes observed during experimental exposures of killer ( Orcinus orca ), long-finned pilot ( Globicephala melas ), and sperm ( Physeter macrocephalus ) whales to naval sonar. Aquatic Mammals 38: 362-401. https://doi.org/10.1578/am.38.4.2012.362
Miller, P.J.O., Kvadsheim, P.H., Lam, F.P.A., Tyack, P.L., Cure, C., DeRuiter, S.L., Kleivane, L., Sivle, L.D., Van Ijsselmuide, S.P., Visser, F., Wensveen, P.J., Von Benda-Beckmann, A.M., Martín López, L.M., Narazaki, T. & Hooker, S.K. (2015). First indications that northern bottlenose whales are sensitive to behavioural disturbance from anthropogenic noise. Royal Society Open Science 2: 140484. https://doi.org/10.1098/rsos.140484
Millero, F.J., Woosley, R., DiTrolio, B. & Waters, J. (2009). Effect of ocean acidification on the speciation of metals in seawater. Oceanography 22: 72-85. https://doi.org/10.5670/oceanog.2009.98.
Morgan, R.P., Rasin, J.R. & Noe, L.A. (1983). Sediment effect on eggs and larvae of striped bass and white perch. Transactions of the American Fisheries Society 112: 220-224. https://doi.org/10.1577/1548-8659(1983)112<220:SEOEAL>2.0.CO;2
Mortensen, P.B. & Buhl-Mortensen, L. (2005). Morphology and growth of the deep-water gorgonians Primnoa resedaeformisand Paragorgia arborea . Marine Biology 147: 775-788. https://doi.org/10.1007/s00227-005-1604-y
Moser, H.G., Lo, N.C.H. & Smith, P.E. (1997). Vertical distribution of Pacific hake eggs in relation to stage of development and temperature. California Cooperative Oceanic Fisheries Investigations Reports 38: 120-126.
Murdoch, R.C., Guo, R. & McCrone, A. (1990). Distribution of Hoki ( Macruronus novaezealandiae ) eggs and larvae in relation to hydrography in Cook Strait, September 1987. New Zealand Journal of Marine and Freshwater Research 24: 529-539. https://doi.org/10.1080/00288330.1990.9516444
NAMMCO (2021). Common minke whale. https://nammco.no/topics/common-minke-whale
Nightingale, B., Longcore, T. & Simenstad, C.A. (2006). Artificial night lighting and fishes. In : Rich, C., Longcore, T. (Eds.). Ecological Consequences of Artificial Night Lighting . Island Press, Washington (DC), USA. pp. 257–276.
Nilsen, B.M., Nedreaas, K. &Måge, A. (2016). Kartlegging av fremmedstoffer i Atlantisk kveite ( Hippoglossus hippoglossus ). Sluttrapport for programmet «Miljøgifter i fisk og fiskevarer» 2013-2015. NIFES, ISBN: 978-82-91065-43-4. https://www.mattilsynet.no/mat_og_vann/uonskede_stofferimaten/miljogifter/miljogifter_i_fisk_2016__atlantisk_kveite.26145-1/binary/Milj%C3%B8gifter%20i%20fisk%202016%20%E2%80%93%20atlantisk%20kveite
Normandeau Associates, Inc. (2012). Effects of noise on fish, fisheries, and invertebrates in the U.S. Atlantic and Arctic from energy industry sound-generating activities . A workshop report for the U.S. Departement of the Interior, Bureau of Ocean Energy Management . Contract # M11PC00031. 72 pp. plus Appendices. https://www.cbd.int/doc/meetings/mar/mcbem-2014-01/other/mcbem-2014-01-submission-boem-04-en.pdf
Nordøy, E.S., Folkow, L.P., Potelov, V., Prischemikhin, V. & Blix, A.S. (2008). Seasonal distribution and dive behaviour of harp seals ( Pagophilus groenlandicus ) of the White Sea-Barents Sea stock. Polar Biology 31: 1119. https://doi.org/10.1007/s00300-008-0453-9
Olsen, B.R., Økland, I. E., Thorseth, I.H., Pedersen, R.B., & Rapp, H.T. (2016). Environmental challenges related to offshore mining and gas hydrate extraction. Miljødirektoratet Rapport M-532 . 28 pp. https://www.miljodirektoratet.no/globalassets/publikasjoner/m532/m532.pdf
Orcutt, B.N., Bradley, J.A., Brazelton, W.J., Estes, E.R., Goordial, J.M., Huber, J.A., Jones, R.M., Mahmoudi, N., Marlow, J.J., Murdock, S. & Pachiadaki, M. (2020). Impacts of deep-sea mining on microbial ecosystem services. Limnology and Oceanography 65: 1489–1510. https://doi.org/10.1002/lno.11403
OSPAR (2008). Descriptions of Habitats on the OSPAR List of Threatened and/or Declining Species and Habitats. OSPAR Agreement 2008-07. https://www.ospar.org/documents?d=32797
Paffenhöfer, G.-A., Strickler, J.R., Alcaraz, M. (1982) Suspension-Feeding by Herbivorous Calanoid Copepods: A Cinematographic Study. Marine Biology 67: 193-199. https://doi.org/10.1007/BF00401285
Page, M.(2014a). Effects of total suspended solids on marine fish: Eggs and larvae on the Chatham Rise. Prepared for Chatham Rock Phosphate Ltd. NIWA Client Report No WLG2012-61: 22 pp. https://www.epa.govt.nz/assets/FileAPI/proposal/EEZ000006/Applicants-proposal-documents/f2819f56f0/EEZ000006-Appendix27-Page-2014a-TSS-Eggs-and-Larvae.pdf
Page, M. (2014b). Effects of total suspended solids on marine fish: Pelagic, demersal and bottom fish species avoidance of TSS on the Chatham Rise. Prepared for Chatham Rock Phosphate. NIWA Client Report No WLG2014-7: 22 pp https://www.epa.govt.nz/assets/FileAPI/proposal/EEZ000006/Applicants-proposal-documents/5f52d0848a/EEZ000006-Appendix28-Page-2014b-TSS-and-Fish.pdf
Page, M. (2014c). Statement of Evidence of Mike Page for CRP ltd. 28 August 2014. Paragraph 30. https://www.epa.govt.nz/assets/FileAPI/proposal/EEZ000006/Evidence/fba56a8e25/EEZ000006-13AppEvidence-Page-Mike-Effects-of-sediment-plume-on-fish-fish-eggs-and-larvae.pdf
Pike, D.G., Víkingsson, G.A., Gunnlaugsson ,T. & Øien, N. (2009). A note on the distribution and abundance of blue whales ( Balaenoptera musculus ) in the Central and Northeast North Atlantic. NAMMCO Scientific Publication 7: 19-29. https://doi.org/10.7557/3.2703
Ramirez-Llodra, E., Hilario, A., Paulsen, E., Costa, C.V., Bakken, T., Johnsen, G. & Rapp, H.T. (2020). Benthic communities on the Mohn’s Treasure Mound: Implications for management of seabed mining on the Arctic Mid-Ocean Ridge. Frontiers in Marine Science 7 : 490. https://doi.org/10.3389/fmars.2020.00490
Reeves, R.R., Mitchell, E. & Whitehead, H. (1993). Status of the northern bottlenose whale, Hyperoodon ampullatus . Canadian Field-Naturalist 107: 490-508.
Roberts, E.M., Mienis, F., Rapp, H.T., Hanz, U., Meyer, H.K. & Davies, A.J. (2018). Oceanographic setting and short-timescale environmental variability at an Arctic seamount sponge ground. Deep Sea Research Part I: Oceanographic Research Papers 138: 98-113. https://doi.org/10.1016/j.dsr.2018.06.007
Roberts, J.M., Wheeler, A.J., Freiwald, A. & Cairns, S.D. (2009). Cold-water Corals: The Biology and Geology of Deep-sea Coral Habitats . Cambridge University Press, Cambridge, UK. 334 pp.
Rolland, R. Parks, S. Hunt, K., Castellote, M., Corkeron, P.J., Nowacek, D.P., Wasser, S.K. & Kraus, S.D. (2012). Evidence that ship noise increases stress in right whales. Proceedings of the Royal Society B 279: 2363-2368. https://doi.org/10.1098/rspb.2011.2429
Santos, M.B., Pierce, G.J., Boyle, P.R., Reid, R.J., Ross, H.M., Patterson, I.A.P., Kinze, C.C., Tougaard, S., Lick, R., Piatkowski, U. & Hernández García, V. (1999). Stomach contents of sperm whales Physeter macrocephalus stranded in the North Sea 1990-1996. Marine Ecology Progress Series 183: 281-294. https://doi.org/10.3354/meps183281
Santos, M.B., Pierce, G.J., García Hartmann, M., Smeenk, C., Addink, M.J., Kuiken, T., Reid, R.J., Patterson, I. A.P., Lordan, C., Rogan, E. & Mente, E. (2002). Additional notes on stomach contents of sperm whales Physeter macrocephalus stranded in the NE Atlantic. Journal of the Marine Biological Association of the United Kingdom 82: 501-507. https://doi.org/10.1017/S0025315402005787
Scanes, E., Kutti, T., Fang, J.K.H., Johnston, E.L., Ross, P.M. & Bannister, R.J. (2018). mine waste and acute warming induce energetic stress in the deep-sea sponge Geodia atlantica and coral Primnoa resedeaformis ; results from a mesocosm study. Frontiers in Marine Science 5: 129. https://doi.org/10.3389/fmars.2018.00129
Schlacher, T., Baco, A.R., Rowden, A.A., O'Hara, T., Clark, M.R., Kelley, C. & Dower, J.F. (2014). Seamount benthos in a cobalt-rich crust region of the central Pacific: conservation challenges for future seabed mining. Diversity and Distributions 20: 491-502. https://doi.org/10.1111/ddi.12142
Simon-Lledo, E., Bett, B.J., Huvemme, V.A.I., Schoening, T., Benoist, N.M.A. & Jones, D.O.B. (2019). Ecology of a polymetallic nodule occurrence gradient: implications for deep-sea mining. Limnology and Oceanography 64: 1883-1894. https://doi.org/10.1002/lno.11157
Simonsen, C.S., Munk, P. Folkvord, A. & Pedersen, S.A. (2006). Feeding ecology of Greenland halibut and sandeel larvae off West Greenland. Marine Biology 149: 937:952. https://doi.org/10.1007/s00227-005-0172-5
Skjoldal, H.R., Dalpadado, P. & Dommasnes, A. (2004). Food webs and trophic interactions. In : Skjoldal, H.R. (Ed.). The Norwegian Sea Ecosystem . Tapir Academic Press, Trondheim, pp. 447-506.
Spearman, J., Taylor, J., Crossouard, N., Cooper, A., Turnbull, M., Manning, A., Lee. M. & Murton, B. (2020). Measurement and modelling of deep sea sediment plumes and implications for deep sea mining. Nature Research, Scientific Reports 10: 5075. https://doi.org/10.1038/s41598-020-61837-y
Steen, I.H., Dahle, H., Stokke, R., Roalkvam, I., Daae, L., Rapp, H.T., Pedersen, R.B. & Thorseth, I.H. (2016). Novel barite chimneys at the Loki’s Castle vent field shed light on key factors shaping microbial communities and functions in hydrothermal systems. Frontiers in Microbiology 6: 1510 https://doi.org/10.3389/fmicb.2015.01510
Paffenhöfer, G.-A. (1972). The effects of suspended 'red mud' on mortality, body weight, and growth of the marine planktonic copepod, Calanus helgolandicus . Water, Air, and Soil Pollution 1: 314-321.
Stockdale, A., Tipping, E., Lofts, S. & Mortimer, R.J.G. (2016). Effect of ocean acidification on organic and inorganic speciation of trace metals. Environmental Science & Technology 50: 1906-1913. https://doi.org/10.1021/acs.est.5b05624
Storrie, L., Lydersen, C., Andersen, M., Wynn, R.B. & Kovacs, K.M. (2018). Determining the species assemblage and habitat use of cetaceans in the Svalbard Archipelago, based on observations from 2002 to 2014. Polar Research 37. https://doi.org/10.1080/17518369.2018.1463065
Strand, E., Bagøien, E., Edwards, M., Broms, C. & Klevjer, T. (2020). Spatial distributions and seasonality of four Calanus species in the Northeast Atlantic. Progress in Oceanography 185: 102344. https://doi.org/10.1016/j.pocean.2020.102344
Taboada, S., Riesgo, A., Wiklund, H., Paterson, G.L.J., Koutsouveli, V., Santodomingo, N., Dale, A.C., Smith, C.R., Jones, D.O.B., Dahlgren, T.G. & Glover, A.G. (2018). Implications of population connectivity studies for the design of marine protected areas in the deep sea: An example of a demosponge from the Clarion-Clipperton Zone. Molecular Ecology 27: 4657-4679. https://doi.org/10.1111/mec.14888 .
Tandberg, A.H.S., Rapp, H.T., Schander, C., Vader, W., Sweetman, A.K. & Berge, J. (2012). Exitomelita sigynae gen. et sp. nov.: a new amphipod from the Arctic Loki Castle vent field with potential gill ectosymbionts. Polar Biology 35: 705-716. https://doi.org/10.1007/s00300-011-1115-x
Tandberg, A.H.S., Rapp, H.T., Schander, C. & Vader, W. (2013). A new species of Exitomelita (Amphipoda: Melitidae) from a deep-water wood fall in the northern Norwegian Sea . Journal of Natural History 47: 25-28. https://doi.org/10.1080/00222933.2012.725778
Tandberg, A.H.S., Vader, W., Olsen, B.R. & Rapp, H.T. (2018). Monoculodes bousfieldi sp.n. from the Arctic hydrothermal vent Loki’s Castle. Marine Biodiversity 48: 927-937. https://doi.org/10.1007/s12526-018-0869-6
Vacquie-Garcia, J., Lydersen, C., Biuw, M., Haug, T., Fedak, M.A. & Kovacs, K.M. (2017). Hooded seal Cystophora cristata foraging areas in the Northeast Atlantic Ocean - investigated using three complementary methods. PLoS ONE 12: e0187889. https://doi.org/10.1371/journal. pone.0187889
Van Dover, C.L. (2019). Inactive sulfide ecosystems in the deep sea: A review. Frontiers in Marine Science 6: 461. https://doi.org/10.3389/fmars.2019.00461
White M, Bashmachnikov I, Arístegui J, Martins AR 2007. Physical processes and seamount productivity. In : Pitcher, T.J., Morato, T., Hart P.J.B., Clark, M.R., Haggan, N., Santos, R.S. (Eds.) Seamounts: ecology, fisheries & conservation . Fish and Aquatic Resources Series 12: pp. 65-84. https://doi.org/10.1002/9780470691953.ch4
Vinogradov, G.M. (1999). Deep-sea near-bottom swarms of pelagic amphipods Themisto : observations from submersibles. Sarsia 84: 465-467. https://doi.org/10.1080/00364827.1999.10807352
Westerberg, H., Rönnbäck, P. & Frimansson, H. (1996). Effects of suspended sediments on cod egg and larvae and on the behaviour of adult herring and cod. ICES CM 1996/E:26 . (in mimeo). https://www.ices.dk/sites/pub/CM%20Doccuments/1996/E/1996_E26.pdf
Whitehead, H., Reeves, R., Feyrer, L. & Brownell Jr., R.L. (2021). Hyperoodon ampullatus . The IUCN Red List of Threatened Species 2021: e.T10707A50357742. https://www.iucnredlist.org/species/10707/50357742
Wiech, M., Duinker, A. & Sanden, M. (2018). Kartlegging av fremmedstoffer i mesopelagiske arter fra norske farvann - Mesopelagisk fisk fra Norskehavet og Sognefjorden. Rapport fra Havforskningen 36-2018. 26 pp. https://www.hi.no/resources/publikasjoner/rapport-fra-havforskningen/2018/kartlegging_mesopelagisk161018_final.pdf
Wiederhold, J.G. (2015). Metal stable isotope signatures as tracers in environmental geochemistry. Environmental Science & Technology 49: 2606-2624. https://doi.org/10.1021/es504683e
Zeldis, J.R., Grimes, P.J. & Ingerson, J.K.V. (1995). Ascent rates, vertical-distribution, and a thermal history model of development of orange roughy, Hoplostethus atlanticus , eggs in the water column. Fishery Bulletin 93: 373-385. https://spo.nmfs.noaa.gov/sites/default/files/pdf-content/1995/932/zeldis.pdf
Øien, N. (2009). Distribution and abundance of large whales in Norwegian and adjacent waters based on ship surveys 1995-2001. NAMMCO Scientific Publications 7: 31-47. https://doi.org/10.7557/3.2704
Øien, N. & Hartvedt, S. (2011). Northern bottlenose whales Hyperoodon ampullatus in Norwegian and adjacent waters. Paper presented to the IWC Scientific Committee, May 2011, Cambridge. IWC SC/63/SM1. (in mimeo). (available at https://www.researchgate.net/profile/Nils-Oien)
Øigård, T.A., Haug, T. & Nilssen, K.T. (2014). Current status of hooded seals in the Greenland Sea. Victims of climate change and predation? Biological Conservation 172: 29-36. https://doi.org/10.1016/j.biocon.2014.02.007
Østvedt, O.J. (1955). Zooplankton investigations from weather-ship M in the Norwegian Sea 1948-49. Hvalrådets Skrifter 40: 1-93.
Aarbakke, O.N.S. (2007). Bio-acoustic investigations of sound scattering layers overlaying hydrothermal vents in the Arctic Ocean . Master of Science thesis in Marine Biology, University of Bergen, Department of Biology. 105 pp.