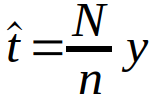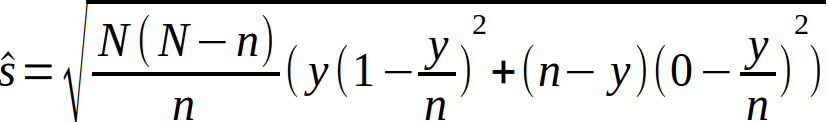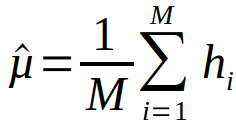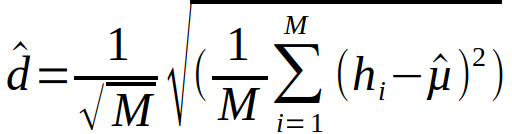Genetic analysis of an egg-survey covering cod spawning grounds
We have analysed the composition of species in egg samples from the 2022 Northeast Arctic Cod spawning survey, with both genetic and microscopic methods. The purpose of the analysis was to assure the quality of the microscopic methods that are used in the standard annual execution of the survey.
We have been particularly interessted in any confusion between cod and haddock, as these cannot reliably be separated microscopically. We did find a lower abundance of haddock-eggs than what was expected based on catches of spawning haddock, and results from other surveys in close spatial and temporal proximity. A few egg samples also had a large abundance of eggs for which species could not be genetically determined.
However, the vast majority of samples was determined genetically, with very few uncertain eggs, and we found generally very good agreement between genetic and microscopic methods for determination of species.
Sammendrag
Vi har bestemt artssammensetning i eggprøver fra skreitoktets eggsurvey i 2022 med genetisk analyse. Formålet har vært å kvalitetssikre den mikroskopiske artsbestemmelsen som inngår i den faste skreitoktprotokollen.
Vi har særlig vært interessert i forvekslingsfaren mellom torsk og hyse, ettersom disse ikke lar seg skille ved mikroskopi. Vi fant en lavere mengde hyseegg enn det som var ventet ut i fra fangster av gytende hyse og nær samtidige tokt i nær overlappende område. Noen få håvtrekk hadde også store mengder egg som ikke lot seg sikkert bestemme genetisk.
Det klare flertall av håvtrekk var imidlertid genetisk bestemt med få usikre egg, og vi fant i hovedsak svært godt samsvar mellom artsbestemmelse gjort ved hjelp av henholdsvis mikroskop og genetikk.
1 - Bakgrunn
På det årlige skreitoktet inngår systematisk sampling av egg med håvtrekk som en del av den faste overvåkingen. Egg blir sortert ombord med stereomikroskop, og egg som er klassifisert som gadoide egg med diameter 1.2-1.6 mm blir i videre databehandling antatt å være torskeegg. Det blir også observert andre gadoide fisk i området, og noe innblandig av andre arter kan ikke utelukkes. Etter skreitoktet i 2022 gjennomførte vi derfor en genetisk analyse for å undersøke innblanding av andre arter i håvtrekkene. Prøvene ble fiksert på sprit etter normal opparbeiding ombord, og senere genetisk analysert på laboratorium ved Havforskingsinstituttet sin stasjon i Flødevigen. Vi håper med det å avdekke eventuell innblanding av andre arter enn torsk (Gadus morhua). 2022 var et år med et moderat innsig av skrei til Lofoten og Vesterålen (Fuglebakk & Thorsen 2022). Vi regner derfor mulighetene for innblanding av andre gadoide egg som høy sammenlignet med tidligere år, og det er slikt sett et år som er godt egnet for å ettergå artsbestemmelsesprotokollen som blir brukt på toktet.
2 - Metode
Egg ble samlet inn ombord på Johan Hjort under ordinært skreitokt i 2022. Håvtrekk utføres på skreitoktet i et fast rutenett med forskjellig tetthet i tre forskjellige strata, som vist i figur 1. Håvtrekk utføres med en T-80 egghåv med 375 µm maskevidde. Denne trekkes vertikalt gjennom de øverste 100 m av vannmassene med en vinsjhastighet på om lag 0,5 m/s. Når bunndypet er grunnere enn 105 m trekkes håven fra 5 m over bunn og opp til overflaten.
Figur 1: Håvtrekk på skreitoktet 2022. Fargelagt etter områdestratifisering.
Protokoll for prøveopparbeiding og eggklassifisering ombord er beskrevet i interne kvalitetsdokument (Overvåkingstokt - skreitokt) og i de årlige toktrapportene (Fuglebakk & Thorsen 2022 ). I korthet var prosedyren som følger: Når håvtrekkene kom ombord ble prøven vurdert og deretter delt med planktondeler ut fra ønsket om at delprøven skulle inneholde mellom 100 og 200 egg (se vedlegg 1). Delprøven ble så opparbeidet levende under stereomikroskop; alle eggene ble plukket ut, deretter fotografert og til sist konservert på 96 % etanol for genetisk analyse. Bildene ble analysert ved hjelp av spesialutviklet billedanalyse programvare hvor hvert egg ble diametermålt samt type- og stadiebestemt. Alle gadoid lignende egg med diameter mellom 1.2 og 1.6 mm ble klassifisert som torskeegg.
De innsamlede eggprøvene ble tatt videre til genetisk analyse ved Lab Flødevigen. Utvalg av prøvene fra de ulike ønskede håvtrekk ble plukket ut tilfeldig og satt opp til DNA-ekstraksjon i 96-brønns plateformat på Hamilton Star robot. Det ble valgt ut 11 egg fra hver håv, bortsett fra håver med mer enn 500 egg, hvor 93 egg med utvalgt for genetisk analyse. For håver med mindre enn 11 egg ble alle egg genetisk analysert. Hver plate fikk to negative kontroller randomisert for å kvalitetssikre plateidentitet og orientering av platen i videre analyser. Kjemien og metoden for ekstraksjonen er levert av Omega Biotek (omegabiotek.com) og baseres på Mag-Bind® Blood and Tissue DNA HDQ.
Mag-Bind-teknologien isolerer DNA ut fra enzymatisk nedbrutte celler i løsning ved at små magnetiske kuler binder til seg DNA. Dette renses videre og separeres til slutt fra kulene og gir et DNA-ekstrakt. Konsentrasjon av DNA-ekstraktene ble videre målt ved hjelp av en fluorometrisk metode på en ThemoFisher Fluoroskan Microplate Fluorometer med Quan IT kit dsDNA High Sensitivity (0.2-50ng) (Ref Thermofisher Scientific)
Prøvene normaliseres til en konsentrasjon på <= 5 ng/µl for videre tilberedning av DNA-bibliotek for sekvensering. Den aktuelle sekvensen er en universell mini barcode 295 bp sekvens av det mitokondrielle COI (cytochrome c oxidase underenhet 1) utviklet for artsidentifisering av fisk av Sultana et al. (2018).
Bibliotekprepareringen skjer gjennom en todelt prosess, med to runder PCR. Ved PCR1 oppkopieres COI-fragmentet, og ved PCR2 merkes hver individuelle prøve med en barkode (i 5), samtidig som den får en indekskode (i 7) som representerer platen. Slik kan hver enkelt prøve identifiseres etter sekvensering. Biblioteksprepareringen gjøres med egne barkoder og indekser, som ikke er del av et kommersielt kit, men som tidligere er utviklet fra protokoll beskrevet i Campell et al. (2015).
Hvert DNA-bibliotek til sekvensering for artsidentifisering er sammensatt av fragmenter fra 16 plater, det vil si maksimum 1504 individuelle prøver og i tillegg 32 negative kontroller.
Biblioteket ble kvalitetskontrollert ved kjøring på en Agilent Bioanalyzer, som gir en profil av biblioteket ved gel elektroforese, og konsentrasjonsmålt på Qubit fluorometer (ThermoFisher Scienific). Sekvenseringen av biblioteket ble utført på en Illumina MiSeq, ved såkalt «paired end sequencing» med et MiSeq Reagent Kit v3 (150 bp) med bruk av 5% PhiX som kontroll.
Rådata fra sekvenseringen i fastq format ble behandlet i R (R Core Team 2021) med modulen DADA2 (Callahan et al. 2016). Kvalitetssikrede sekvenser ble deretter sammenliknet med en database bestående av COI sekvenser for gadoide fisk. Sammenlikningen ble gjort med R modulen rBLAST (Hahsler 2019). For suksessfull artsbestemmelse ble det forutsatt at et segment har minst 250 sammenhengde basepar som er sammenstilt med 100% likhet med et segment i databasen, og at dette forekommer for kun en enkelt art.
Genetisk analyse ble utført på et tilfeldig valgt sub-sample av de prøvene som var sortert ombord ved hjelp av mikroskopi. Noe forskjell i antall torskeegg identifisert med de to metodene må derfor påregnes som følge av tilfeldige samplingfeil. Vi kan estimere total antall torskeegg i mikroskopi-sorterte prøver i et håvtrekk ved:
hvor y er antall egg identifisert som torskeegg i genetisk analyse. n er antall egg i sub-sample til genetisk analyse, og N er totalt antall egg sortert ombord. Forventet samplingfeil er gitt ved standardfeilen:
Vi lar u betegne antall torske-egg identifisert ved mikroskopi, og regner ut absolutt forskjell mellom identifiseringsmetodene som:
For å få et innblikk i praktisk betydning av forskjeller i artsidentifiseringsmetodikk, kan vi estimere gjennomsnittlige håvfangster for romlige strata på bakgrunn av begge metodene. Vi behandler håvtrekkene som et tilfedig utvalg i hvert strata og regner gjennomsnittlig eggmengde pr håv i et strata som:
hvor hi er antall torskeegg i håvtrekk i, estimert enten fra artssortering med mikroskopi eller genetikk. M er totalt antall håvtrekk i strata. Hvert enkelt håvtrekk dekker en forsvinnende liten andel av det totale arealet i hvert strata, og standardfeilen til gjennomsnittlig eggmentde kan estimeres som:
Vi ser da vekk fra samplingfeil som skriver seg fra sub-sampling av hvert håvtrekk (Se Delingsfaktor i Vedlegg 1).
3 - Resultater
3.1 - Artsbestemmelse
Grunnet naturlig variasjon i genfragmentet som ble analysert er det en viss risiko for feilklassifisering, og en viss risiko for at vi finner varianter av en art som ikke er belagt i databasen, og derfor ikke kan artsbestemmes. Som en kontroll på metodikken har vi derfor analysert sekvenslikhet mellom de forskjellige artene i databasen. Disse er oppsummert i Tabell 1. Med den genetiske variasjonen som er belagt i databasen er det kun mellom hyse og hvitting det er en viss forvekslingsfare med kriteriene vi bruker for genetisk bestemmelse. Den reelle variasjonen er naturlig nok større, men statistikken i tabell 1 viser likevel at torskesekvensene vi har analysert ikke kan forveksles med de 681 sekvensene fra 9 ulike arter torskefisk (Gadiformes) som vi har i databasen. Særlig ser vi at forvekslingsfaren mellom torsk og hyse er lav, med ingen like sekvenser funnet for 473*194 sammenlignede sekvenspar. Det er av interesse, fordi vi på forhånd regnet hyse som den viktigste forvekslingsarten i mikroskopiundersøkelsene.
Tabell 1: Maksimal sekvenslikhet for sekvenspar lengre enn eller lik 250 basepar, mellom arter i databasen. Antall sekvenser for hver art er angitt i parentes under artsnavn i kolonneoverskrifter.
Torsk
(473)
Hyse
(194)
Sei
(152)
Lyr
(34)
Hvitting
(112)
Øyepål
(16)
Brosme
(27)
Kolmule
(109)
Sypike
(37)
Lysing
(307)
Torsk
(Gadus morhua)
100,0
98,1
95,0
94,6
96,5
86,1
92,9
91,9
90,5
89,2
Hyse
(Melanogrammus aeglefinus)
100,0
97,8
97,4
100,0
90,7
95,3
91,5
91,5
93,9
Sei
(Pollachius virens)
100,0
97,6
95,4
86,3
93,2
92,2
90,2
91,3
Lyr
(Pollachius pollachius)
100,0
95,4
90,3
92,9
91,6
91,6
90,2
Hvitting
(Merlangius merlangus)
100,0
85,8
94,1
91,0
90,0
89,6
Øyepål
(Trisopterus esmarkii)
100,0
90,4
99,6
95,9
Brosme
(Brosme brosme)
100,0
91,5
92,9
90,9
Kolmule
(Micromesistius poutassou)
100,0
96,6
89,9
Sypike
(Trisopterus minutus)
100,0
89,2
Lysing (Merluccius merluccius)
100,0
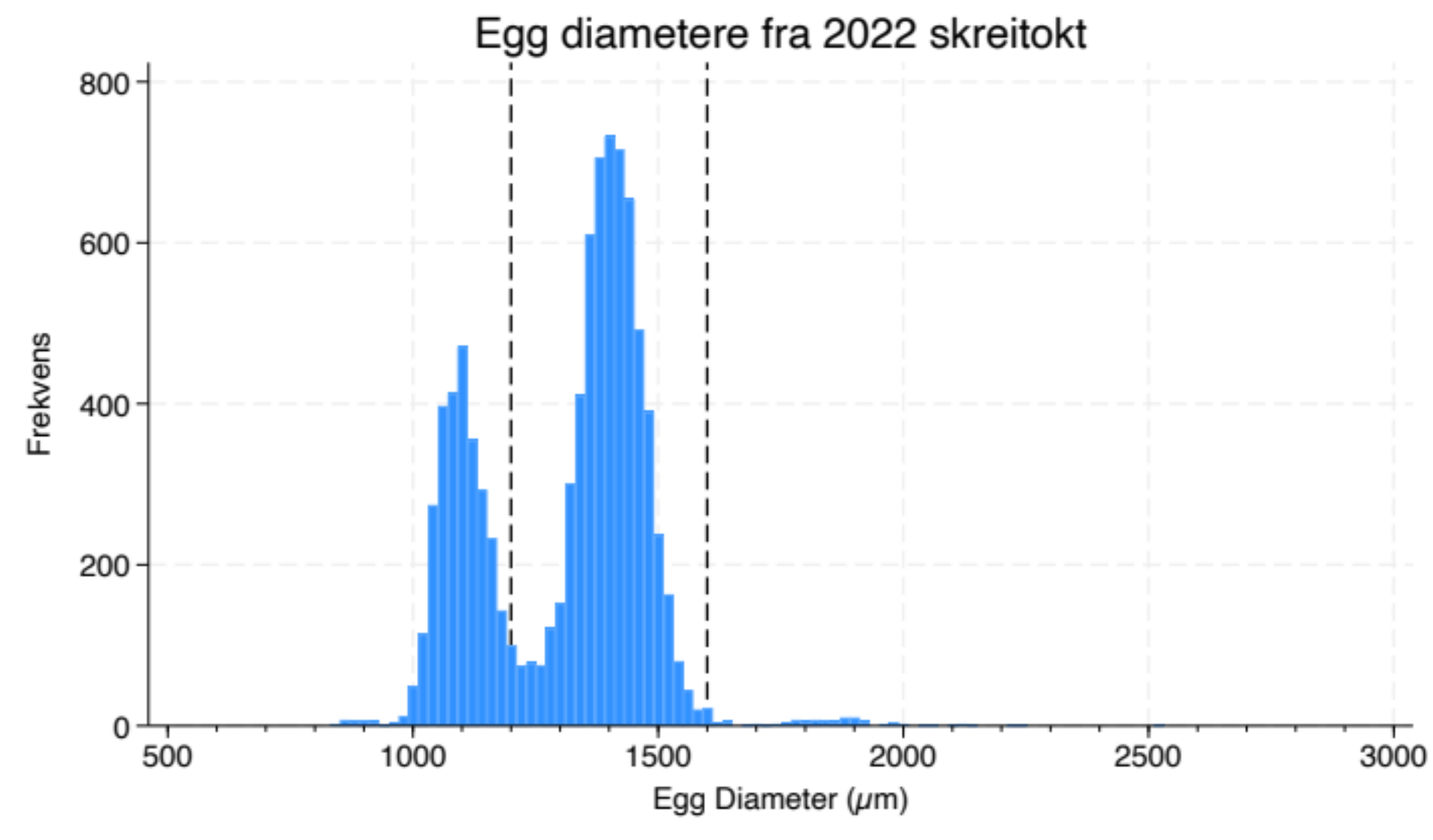
De fleste eggene lot seg genetisk identifisere med protokollen beskrevet over. 373 av 1468 analyserte egg hadde ikke 100% treff på sekvens i databasen. Av disse hadde 321 mer enn 98% sekvenslikhet med øyepålindivider (Trisopterus esmarkii) i databasen. Disse uidentifiserte eggene er utelatt fra videre analyse. 834 egg ble identifisert som torskeegg, og kun 11 ble identifisert som hyse (Melanogrammus aeglefinus), som er den arten som vi på forhånd regnet som den som det er størst forvekslingsfare med ved mikroskopisk bestemmelse. Det lave antallet hyse-egg var et noe overraskende funn, da det ble funnet en del gytende hunn-hyse ved tråling under det samme toktet, og i overlappende toktområde på umiddelbart påfølgende tokt (se vedlegg 1 til Johannesen et al. 2022). Av de øvrige eggene var de fleste identifisert som sei (Pollachius virens, 247 egg) . For disse regner vi med liten forvekslingsfare med torskeegg ved mikropskiopisk bestemmelse ettersom diameteren typisk er noe mindre. Russel (1976) oppgir diameteren for seiegg til å være mellom 1.03 og 1.22 mm og torskeegg fra 1.16 til 1.89 mm. Imidlertid er vår erfaring fra mange eggtokt helt tilbake til 70-årene at skrei og kysttorsk i dette området gyter egg med diametere innenfor et mer snevert intervall, typisk mellom 1.2-1.6 mm. En viss overlapp med sei kan det imidlertid være, noe som også indikeres når man ser på størrelses fordelingen av egg fra toktet (Figur 2).
Figur 2: Eggdiametere fra skreitoktet i 2022. Vertikale stiplede linjer angir intervallet fra 1200 – 1600 µm som vi regner som nedre og øvre grense for torskeegg.
3.2 - Sammenligning
Figur 3 viser sammenligning av total estimert antall egg for hvert håvtrekk basert på enten artsidentifisering ved mikroskopi eller genetikk. Det overordnede bildet er et godt samsvar, foruten stasjonene i Austnesfjorden. Vi kan også merke oss at forskjell i eggmengde mellom håvtrekk later til å være vesentlig større enn forskjellen mellom artsbestemmelsesmetodene for samme håvtrekk. Dette indikerer at totalestimater fra toktet ikke vil være veldig sensitiv til hvilken artsbestemmelsesmetodikk som benyttes.
Figur 3: Antall torskeegg i håv. Estimert fra mikroskopi ombord og fra genetikk av subsample
Som vist i tabell 2 er også gjennomsnittlig absolutt forskjell mellom antall torskeegg identifisert i samme størrelsesorden som forventet samplingfeil. Det tyder på at forskjellene er mindre enn eksperimentet er egnet til å demonstrere. Også her ser vi imidlertid et tydelig unntak for stasjonene i Austnesfjorden.
Tabell 2: gjennomsnittlig standardfeil og absolutt forskjell mellom artsbestemmelsesmetoder
strata
Gjennomsnitlig standardfeil (s) genetikkestimat
Gjennomsnittlig absolutt forskjell (r)
Ytre Nord
6 egg
6 egg
Ytre Syd
14 egg
10 egg
Vestfjorden
11 egg
10 egg
Austnesfjorden
14 egg
38 egg
For å vise praktisk betydning av hvilken artsbestemmelsesmetodikk som blir brukt estimerer vi gjennomsnitlig eggmengde for hvert romlig stratum og bergener standardfeil. Denne beregningen inngår i beregning av eggindeksen, slik den er formulert i vedlegg til Fuglebakk & Thorsen (2022). Sammenligningen er vist i Figur 4.
Figur 4: Gjennomsnittlig eggmengde +/- standardfeil for hhv artsbestemmelse ved mikroskopi og genetikk
Vi ser at forskjellen mellom artsbestemmelsesmetodikk er mindre enn omtrentlig ett standardavvik for alle områdene, igjen med unntak av Austnesfjorden. For en total eggmengdeindeks må det tas hensyn til at håvtrekkene ikke er utført med lik tetthet i de forskjellige områdene (Figur 2). Siden Austnesfjorden utgjør et svært lite areal bidrar den lite eggmengdeindeks for hele toktområde. En arealvektet bergening for hele toktområdet gir et gjennomsnitt på 53 +/- 5 torskeegg for genetikk, og 54 +/- 5 egg for mikroskopi. Vi regner at håven sampler et areal på ca 2 m2, så dette tilsvarer mellom 20 og 30 egg pr. m2.
3.3 - Austnesfjorden
Avvikene i Austnesfjorden skriver seg fra håvtrekkene med stasjonsnummer (CTD-nummer) 261, 262, 263, 264, og 265. Disse utgjør de fleste og innerste stasjonene i Austnesfjorden. I tabell 3 er det angitt noen nøkkeltall om prøveutvalget.
Tabell 3: Nøkkeltall om prøveutvalg for utvalgte stasjoner i Austnesfjorden.
stasjonsid
Torsk genetikk
Torsk mikroskopi
N
n
261
6
48
219
93
262
1
35
125
93
263
1
77
265
93
264
0
43
91
11
265
1
86
120
11
De fleste av disse stasjonene har altså blitt svært intensivt samplet for genetisk analyse, og det er ikke sannsynlig at forskjellene skyldes samplingfeil, i alle fall ikke for stasjonene 261, 262 eller 263. Figur 4 viser at artssammensetningen i disse prøvene i hovedsak er sei og ukjente arter. Eggene som her ikke er identifisert med 100% sekvenslikhet har 99% sekvenslikhet med øyepål. Vi forventer imidlertid liten grad av forveksling med torsk for både sei og øyepål. Det er mulig at avvikene kan forklares med forvekslingsarter som ikke er beskrevet i databasen vi har brukt for genetisk analyse. Dette kan ettergåes senere etterhvert som databasen utvides med nye data. Austnesfjorden har spesielle forhold sammenlignet med toktområdet forøvrig, og vi finner ofte relativt store mengder med egg i sene utviklingsstadier der. Det kan tilsi at tilknyttede gyteplasser er godt egnet for arter og bestander som har best oppvekstvilkår nært kysten.
Figur 5: Genetisk artsbestemmelse for utvalgte stasjoner i austnesfjorden. sekvenser med <100% sekvenslikhet med art i database er vist som Ukjent.
4 - Konklusjon
Genetisk analyse av skreitoktets eggsurvey i 2022 viser at torskeegg er dominerende i håvtrekkene også i et år med et relativt lite innsig av gytetorsk fra Barentshavet. Analysen bekrefter også i hovedsak at protokollen for å identifisere torskeegg med mikroskopi fungerer tilfredstillende for beregning av eggmengdeindekser. Vi har ikke en fullgod forklaring på at en del stasjoner i Austnesfjorden avviker fra dette bildet (Figur 3 og 4), men dette har lite praktisk betydning for eggmengdebestemmelser for toktområdet som helhet. Vi var også litt overrasket over at vi ikke fant mer hyseegg, ettersom vi gytende hyse er observert i trålhal i Vestfjorden på denne tiden. Vi har heller ikke en fullgod forklaring på det, men har kontrollert mot et rikt utvalg av hysesekvenser i genetikkdatabasen.
Feil i artsbestemmelse later til å være av en størrelsesorden som er mindre enn samplingfeil (Figur 4, Tabell 2), og slik ikke av betydning for fortolkning av indeksen. Vi føler oss betrygget på at artssortering med mikroskopi er en kostnadseffektiv metode for å opparbeide en pålitelig eggmengdeindeks. Artsbestemmelse med mikroskopi er likevel prisgitt forholdene i havet, slik som mengde av forvekslingsarter i området, og deres gytetidspunkt. Begge disse forholdene kan være gjenstand for endring over tid. Eksperimentet kan derfor med fordel gjentas om noen år, og mulighetene for å gjennomføre rutinemessig genetisk analyse bør fortløpende vurderes mot kostnad.
Sultana, Sharmin, Md. Eaqub Ali, M.A. Motalib Hossain, Asing, Nina Naquiah, og I.S.M. Zaidul. Universal Mini COI Barcode for the Identification of Fish Species in Processed Products’. Food Research International 105 (March 2018): 19–28.
(https://doi.org/10.1016/j.foodres.2017.10.065)
Campbell, Nathan R., Stephanie A. Harmon, og Shawn R. Narum. ‘Genotyping‐in‐Thousands by Sequencing (GT‐seq): A Cost Effective SNP Genotyping Method Based on Custom Amplicon Sequencing’. Molecular Ecology Resources 15, no. 4 (July 2015): 855–67.
(https://doi.org/10.1111/1755-0998.12357)
R Core Team (2021). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
(https://www.R-project.org/)
Callahan, B., McMurdie, P., Rosen, M. et al. DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods 13, 581–583 (2016).
(https://doi.org/10.1038/nmeth.3869)
Hahsler M, Nagar A (2019). rBLAST: R Interface for the Basic Local Alignment Search Tool. R package version 0.99.2.
(https://github.com/mhahsler/rBLAST)
Russel, F. S.1976. The Eggs and Planktonic Stages of British Marine Fishes, Academic Press, London - New York - San Francisco.
6 - Vedlegg 1 – Oversikt over eggprøver fra skreitoktet i 2022
Tabellen viser oversikt over håvtrekk på ordinært tokt (stasjoner fra snitt er utelatt). Torske-egg er antall egg bestemt til å være torsk ved mikroskopi. Total egg er totalt antall egg i prøven som ble undersøkt. Delingsfaktor angir hvor stor andel denne prøven utgjør en av alle eggene i håvtrekket. Betegnelsene u og N refererer til variable i avsnittet «Metode».
Stratum
Stasjons-nummer
Bredde-grad
Lengde-grad
Delings-faktor
Torske-egg (u)
Total egg (N)
Austnesfjorden
258
68.189
14.605
1
22
45
259
68.222
14.632
2
102
173
260
68.234
14.705
1
48
150
261
68.285
14.711
4
48
219
262
68.329
14.742
8
35
125
263
68.304
14.728
8
77
265
264
68.252
14.694
4
43
91
Vestfjorden
225
67.334
11.626
1
25
92
226
67.266
11.860
1
0
1
227
67.312
11.936
1
2
4
228
67.397
11.949
1
26
85
229
67.383
12.106
1
5
7
230
67.463
12.156
1
45
80
231
67.382
12.485
1
1
4
232
67.476
12.463
1
0
0
233
67.559
12.476
1
3
4
234
67.524
12.703
1
0
0
235
67.609
12.704
1
1
1
236
67.675
12.804
1
124
137
237
67.638
13.005
1
2
2
238
67.724
13.005
1
12
16
239
67.807
13.013
1
176
193
240
67.731
13.236
1
4
8
241
67.877
13.043
2
110
154
242
67.822
13.234
1
44
56
243
67.913
13.218
2
89
133
244
67.817
13.507
1
105
115
245
67.985
13.240
2
180
241
246
67.909
13.490
2
61
94
247
68.001
13.474
1
73
109
248
67.910
13.745
2
112
137
249
67.996
13.743
1
1
1
250
68.074
13.741
4
109
174
251
67.986
14.028
1
2
6
252
68.072
14.034
1
29
46
253
68.148
14.017
8
187
199
254
68.073
14.297
1
3
7
255
68.151
14.300
1
38
49
256
68.113
14.513
1
11
18
257
68.176
14.489
1
90
127
Ytre Nord
157
69.711
17.756
1
32
38
158
69.840
17.332
1
21
31
159
69.989
16.847
1
51
56
160
69.813
16.826
1
31
33
161
69.676
17.276
1
46
61
162
69.629
16.842
1
32
34
163
69.622
16.282
1
18
28
164
69.435
16.294
1
3
6
165
69.460
15.644
1
4
4
166
69.346
16.004
1
13
17
167
69.284
15.632
1
11
14
168
69.295
15.038
1
19
30
169
69.127
14.999
1
20
25
170
69.129
14.434
1
43
51
171
68.917
14.533
1
53
57
172
68.984
13.778
1
20
23
173
68.696
14.114
1
107
113
Ytre Syd
174
68.900
13.492
1
64
79
175
68.737
13.446
2
214
225
176
68.510
14.144
2
106
115
177
68.582
13.379
4
83
85
178
68.678
13.089
1
145
156
179
68.664
12.599
2
72
104
180
68.495
13.112
2
81
88
181
68.349
13.557
1
56
60
182
68.357
13.126
1
143
151
183
68.530
12.604
1
31
51
184
68.560
12.125
1
56
68
185
68.398
12.604
2
87
94
186
68.215
13.100
2
196
199
187
68.273
12.585
4
122
125
188
68.421
12.139
2
68
95
189
68.453
11.663
1
41
44
190
68.287
12.149
2
114
128
191
68.143
12.581
1
141
163
192
68.004
12.601
4
149
150
193
68.163
12.125
1
118
140
194
68.325
11.641
1
83
122
195
68.207
11.610
1
20
123
196
68.038
12.116
4
101
107
197
67.826
12.602
2
130
136
198
67.800
12.166
2
120
134
199
67.630
12.150
1
95
122
200
67.902
12.121
2
64
103
201
68.081
11.587
1
90
119
202
68.269
11.033
1
40
68
203
68.156
10.977
2
81
182
204
67.950
11.595
2
132
190
205
67.816
11.600
1
12
128
206
68.029
10.972
1
22
149
207
67.902
10.967
2
28
101
208
67.651
11.581
1
133
182
209
67.483
11.552
2
85
121
210
67.780
10.942
1
10
120
211
67.980
10.358
1
6
65
212
67.864
10.317
1
1
7
213
67.632
10.989
1
0
8
214
67.318
11.526
1
0
2
215
67.518
10.948
1
2
10
216
67.742
10.296
1
0
1
217
67.629
10.249
1
1
2
218
67.401
10.902
1
5
102
219
67.148
11.516
1
0
17
220
67.283
10.883
1
0
0
221
67.510
10.229
1
1
6
222
67.584
9.639
1
1
1
223
67.383
10.209
1
0
1
224
67.165
10.845
1
0
1
7 - Vedlegg 2 – Genetisk analyse av eggprøver fra skreitoktet 2022
Tabellen viser resultat av genetikk utført på ordinære håvtrekk i skreitoktet. Noen få stasjoner fra snitt ble også analysert, men er ikke inkludert i tabellen. Betegnelsen n refererer til variable i avsnittet «Metode».