[Text]
Havbruk til havs – Fysiske miljøbetingelser og økosystempåvirkning
Rapportserie:
Rapport fra havforskningen 2019-41
ISSN: 1893-4536
Publisert: 16.12.2019
Prosjektnr: 15311
Oppdragsgiver(e): Fiskeridirektoratet
Tema:
Havbruk til havs
Program:
Marine Processes and Human Impact
Approved by:
Research Director(s):
Geir Lasse Taranger
Program leader(s):
Frode Vikebø
English summary
Sammendrag
Nærings- og fiskeridepartementet ber om kunnskapsinnspill for å vurdere egnede lokaliteter for havbruk til havs. Fiskeridirektoratet har i den sammenheng bedt Havforskningsinstituttet om å bidra på fire områder; i) Krav til miljø som gir god fiskevelferd for laks, ii) Smitte til og fra anlegg, iii) Fysiske miljøbetingelser, og iv) Økosystempåvirkning. I denne rapporten adresserer Havforskningsinstituttet de to siste pkt. listet over. Ettersom en rekke ulike rapporter tidligere har beskrevet sårbare habitater, arter og stadier vil vi her spesielt vektlegge en kvantitativ beskrivelse av det fysiske og dynamiske miljø på kysten ut til øvre del av kontinentalskråningen av betydning for fiskevelferd, belastning på havbruksrelaterte installasjoner og miljøpåvirkning. Havbruk til havs vil oppleve andre miljøbetingelser enn i skjermede farvann i fjorder og nær kyst. Som grunnlag for å vurdere beskaffenhet er det spesielt strøm, bølger og temperatur som vurderes som de miljøparametere som kan være begrensende i forhold til anleggenes utforming (belastning) samt fiskevelferd. I tillegg vil havbruk til havs kunne påvirke sårbare organismer i nærmiljø gjennom organisk belastning og i avstand gjennom spredning med omliggende strøm. Forstyrrelser av gyte- eller beitevandring gjennom støy eller kjemiske signaler er også noe som kan påvirke det lokale økosystem selv om grunnlaget for å vurdere dette er mindre kjent. Det er naturlig å skille mellom arter som er på vandring og stasjonære arter ettersom de sistnevnte vil kunne være nær områder påvirket av havbruk til havs deler av eller hele tiden. Imidlertid vil miljøbelastning avta med avstand fra anlegg som funksjon av fortynning og strøm. Avhengig av artenes sensitivitet for miljøforandring er likevel anledningen til å justere vandringsrute større på sokkelen enn i fjordene. Det fysiske miljø på sokkelen domineres av den Norske kyststrøm, kantstrøm på kontinentalskråningen og de sesongmessige svingningene. Observerte og modellerte strømhastigheter tilsier at et område mellom kyststrømmen og kantstrømmen fra rundt 64 – 67 °N har maksimalstrøm under laksens vedvarende svømmekapasitet som rapportert i Hvas et al. (2019). Merk at nær kyststrøm eller kantstrøm kan det opptre hyppige virvelavleiringer med sterke strømhendelser. I Barentshavet er det også moderate strømhastigheter, men til forskjell fra områder lengre sør er minimumstemperaturene betydelig lavere og dette vil resultere i at laksen får lavere tålegrense for strøm. Sør og sørvest for kyststrømmen i Nordsjøen er det også moderate strømhastigheter og i motsetning til Barentshavet er temperaturene her fordelaktige for ytelse og det forventes ikke at de skal påvirke laksens svømmeevne i særlig grad. Generelt viser havmodellen at områder nord og vest for Lofoten har lave temperaturer som svekker laksens ytelse og dermed tåleevne for strøm. Fiskebestander som gyter på Norskekysten er oppsummert i rapportene KILO og KINO samt tilgjengelig i http://www.imr.no/geodata/geodataHI.html. Gyting i det aktuelle området foregår som regel tett på kyst eller på sokkelkanten. Men etter gyting driver egg og nyklekkede fiskelarver nordover i et bredt belte i og langs kyststrømmen. Det frarådes derfor å legge større havbruksinstallasjoner på eller umiddelbart nedstrøms av viktige gytefelt langs kyst. Kunnskapen om vandringsrutene til postsmolt fra Norske lakseelver og ut til beiteområdene i havet er begrenset. Det samme er kunnskapen om hvordan større laks eventuelt benytter kystnære områder for beiting, og vandringsrutene til kjønnsmoden laks som returnerer til elvene for å gyte. Det gjør det vanskelig å vurdere eventuell påvirkning av havbruk til havs. Dersom effekt fra havbruk til havs på vandrende laks skal kunne vurderes uten stor usikkerhet må kunnskapsgrunnlaget bedres. Ved utpeking av lokaliteter som er egnet for havbruk til havs må man ta hensyn til eksisterende anbefalinger om områder som er definert som sårbare og eller særlig verdifulle. Videre må man ta hensyn til kandidatområder for marint vern og områder som skal fungere som referanseområder for å vurdere endringer som følge av naturlig eller antropogen påvirkning. Områder tiltenkt akvakulturproduksjon på norsk sokkel bør kartlegges spesifikt for lokal artssammensetning og artsmangfold tidlig i prosessen, slik at man kan unngå å legge produksjonsanlegg over områder med sårbar og/eller funksjonelt viktige økosystem. Det er sannsynlig med sykdomshendelser i laks også på lokaliteter til havs, og disse vil på samme måte som lokaliteter i kyst og fjordstrøk utgjøre en smittespredningsfare. Store lokaliteter med mye biomasse og høyt antall fisk vil kunne slippe ut betydelige mengder parasitter. På den annen siden vil sterk strøm og store vannmasser i stor grad gjøre at det er lave konsentrasjoner av agens, og sannsynligvis spredte verter. Tett kontakt med mange ulike arter fisk øker også muligheten for at parasitter fra oppdrett kan etablere seg i «nye» verter. Det er derfor nødvendig å øke kunnskapsgrunnlaget for å vurdere smitte og smittespredning i store anlegg til havs med mye høyere biomassetetthet enn det vi har erfaring med fra kyst.
1 - Miljøforhold relevant for havbruk til havs
Myndighetene vurderer nye arealer for havbruksnæring og havbruk til havs kan være ett alternativ. I den forbindelse må det vurderes hvilke begrensinger miljøforhold kan ha med tanke på fysisk stress på installasjoner og oppdrettsfisk. I tillegg må miljøbelastning for omliggende naturtyper beskrives i tilstrekkelig grad slik at forvaltningen kan vurdere næringsutbytte mot miljøbelastning. I denne rapporten har vi sammenstilt data og kunnskap vi mener er sentrale for en slik vurdering.
1.1 - Fysisk miljø
Strømstyrke, temperaturforhold og bølger er de fysiske parametere som anses som mest relevante for vurdering av egnede områder til havs for havbruk. I denne rapporten sammenstilles derfor statistikk basert på de best tilgjengelig observerte og modellerte verdier med fokus på åpen kyst ut til 500 m dybdekontur som sammenfaller med øvre del av sokkelskråningen ut i dyphavet. Det beregnes både middelverdier og ekstremverdier. Ettersom havbruksinstallasjoner kan ha ulik vertikal utforming for å håndtere ulike fysiske og biologiske utfordringer som strømstyrke, bølgepåvirkning og lakselus presenterer vi verdier for 2, 20 og 50 m dyp. Disse antas å være representative for ulike dybdesegmenter; 2 m for de øvre 10 m, 20 m for 10-30 m og 50 m for 30-100 m. Mindre kjent, og i mindre grad tilgjengelig fra in situ målinger, er turbulens. Vi vet at turbulens påvirker blant annet interaksjoner mellom byttedyr og dens predatorer (e.g. Kristiansen et al., 2014). Men lignende kan også være relevant for eksponering av marine organismer for fôr, miljøgifter og parasitter. Imidlertid beregner havmodeller turbulent blanding. Men ettersom vi ikke har nok kunnskap om hvordan turbulens eventuelt kan påvirke havbruk, og vi har ikke sammenlignbare observasjoner for turbulens som for havstrømmer, er det ikke vektlagt i denne rapporten.
1.2 - Vandrende arter
Vi har valgt å fokusere på gytefordeling og -vandring på kyst for kommersielt relevante arter, vandring av laks fra elver og ut i dyphavet, og sjøpattedyrvandring. Gytefordeling og -vandring er godt beskrevet i rapportene KunnskapsInnhenting for Barentshavet-Lofoten-Vesterålen (KILO) (Sundby et al., 2013) og Dynamic Mapping of the North Sea Spawning – Report of the KINO Project (Sundby et al., 2017) samt tilgjengelig på http://www.imr.no/geodata/geodataHI.html.
1.3 - Stasjonære arter
Stasjonære arter som koraller og svamp er ikke i stand til å justere overlapp med havbruk til havs slik vandrende arter kan. De er prisgitt den miljøbelastning som finner sted i sitt nærområde og oppstrøms. De utgjør viktige habitater for andre marine organismer, og har viktige biologiske funksjoner. Eksempelvis står korallrevene for 30 % av all omsetning av organisk materiale som synker til bunns til tross for at de kun dekker 1 % av arealet på norsk sokkel.
1.4 - Modelleringskapasitet
Manglende evne til å observere i tid og rom kompenseres med modellverktøy. I tillegg fungerer modellene som minilaboratorier hvor vi kan teste ulike miljøforhold og studere respons i de inkluderte artene. Aktuelle modeller for å vurdere miljøpåvirkning fra havbruk til havs er i) strømmodell (havmodell) som gir en kontinuerlig beskrivelse av strøm og hydrografi i tid og rom, ii) individ-basert modell koblet til en partikkelsporingskode som flytter partikler (kan representere arter og/eller stadier) i henhold til strømmodell (eksempelvis lakselusmodellen i trafikklyssystemet), iii) NPZD-modell (også med inngangsdata fra strømmodell) som tallfester plante- og dyreplanktondynamikk og som er velegnet til å studere effekter av organisk avfall, iv) end-2-end modeller som inkluderer alt fra fysikk via bakterier til fisk og pattedyr og som er velegnet til å studere hvordan pådrag dempes eller forsterkes i næringsnettet. I forbindelse med trafikklyssystemet for å gi råd om lakselus i de 13 nasjonale lakseproduksjonsområdene benyttes en koblet strømmodell og en individbasert modell for lakselus. Strømmodellen er etablert gjennom et nasjonalt samarbeid med tunge bidrag fra HI og MET, og kjøres i dag operasjonelt av MET for hele Norskekysten med havvarsler for strøm og hydrografi med 800 m horisontaloppløsning i en rekke vertikale lag. Den nøstes også til enda høyere oppløste modellområder for mindre områder - typisk enkeltfjorder.
1.5 - Referanser
Sundby et al. (2017) Dynamic Mapping of the North Sea Spawning – Report of the KINO Project. Fisken og Havet nr. 2.
Sundby et al. (2013) KunnskapsInnhenting Barentshavet-Lofoten-Vesterålen (KILO). Fisken og Havet nr. 3.
2 - Observert og modellert strøm - og temperaturforhold
I de faste hydrografiske stasjoner (http://www.imr.no/forskning/forskningsdata/stasjoner/) – 8 stasjoner hvor det måles temperatur og saltholdighet i flere dyp minst en gang per måned med de første målingene alt fra 1935 - ser vi naturlige svingninger for ulike perioder (de lengste 60-80 år) samt en jevn temperaturøkning i kystvannet. Svingningene følger i stor grad regionale temperatursignaler fra omliggende havområder med vekslende varme og kalde perioder. Det siste 10-året er på tampen av en varmetopp. Naturlige svingninger kan derfor forbigående bidra til å dempe lokale effekter av klimaendringer på temperatur. I perioden vi har bak oss er variasjon mellom år større enn det som kan tilskrives klimaendringer, men ekstremverdiene øker fordi naturlig variabilitet kommer på toppen av klimaendringene. I spesialrapporten om hav og is fra IPCC (IPCC, 2019) pekes det på økende frekvens av marine varmebølger som følge av klimaendringer. Det betyr at statistiske vurderinger av historiske observasjoner vil ha avtagende relevans for å beskrive fremtidige marine miljøforhold.
2.1 - Havmodellen NorKyst800
Modelloppsett
Den hydrodynamiske modellen som ble brukt for å representere de fysiske forholdene til havs er ROMS (Regional Ocean Modeling System, http://myroms.org). I applikasjonen omtalt som Norkyst800 er den kjørt med en horisontaloppløsning på 800 x 800 m, en vertikaloppløsning på 35 terrengfølgende dybdenivåer og et modellområde som dekker hele Norskekysten og utover sokkelkanten. Mer informasjon om havmodellen og modelloppsett kan leses i den tekniske rapporten om NorKyst800 (Albretsen et al., 2011). Havforskningsinstituttet har over flere år bygget opp kompetanse på bruk av havmodellen ROMS. ROMS er en hydrodynamisk modell i forskningsfronten som inkluderer alle relevante fysiske prosesser. Modellen brukes i stor grad i det internasjonale havmodellmiljøet og er en av de dominerende metodikkene innenfor hav- og kystmodellering i verden.
Det er avgjørende å ha gode drivkrefter og randverdier for å gjennomføre en representativ strømmodellering. I oppsettet for å etablere et modellarkiv for årene 2006 - 2016 ble det brukt informasjon (vannstand, strøm og hydrografi) langs de åpne rendene for modellområdet fra en stor og godt dokumentert Norskehavsmodell med 4 km oppløsning (Lien et al., 2013; Lien et al., 2014). Tidevannsamplitude og -bevegelse ble hentet fra det globale tidevannsatlaset TPXO (Egbertand Erofeeva, 2002). Atmosfærepådraget ble hentet fra en egen høyoppløst simulering med WRF (Weather Research and Forecastingmodel, utviklet av NCAR/National Center of Atmospheric Research, USA) med 3 km horisontaloppløsning. Elveavrenning for alle vassdrag er basert på den hydrologiske HBV-modellen kjørt av NVE (Norges Vassdrags- og Energidirektorat, se Beldring et al., 2003) og er avgjørende for å gjenskape den ferske kyststrømmen.
Bunndypet i NorKyst800 er glattet under selve strømsimuleringen, men grunnlagsdataene er hentet fra Norge Digitalt (http://www.norgedigitalt.no) som er etablert av Statens Kartverk Sjø. Den opprinnelige oppløsning er på rundt 50 m på et irregulært gitter for hele kysten, men disse er interpolert til et 800 m gitter med en vekting i forhold til antall land- og sjøpunkter. Bunndypene for hver gitterrute som er angitt som datagrunnlag i figurene er hentet fra det interpolerte produktet, og modellens bunndyp under modellkjøringene er noe glattet for å unngå numeriske støy.
Modellresultat
Variabelen temperatur ble hentet direkte fra NorKyst800-arkivets døgnmidler, og ble her midlet over alle år for perioden 2013 –2016 for hvert av dypene 2, 20 og 50 m.
Strømhastighet ble hentet direkte fra NorKyst800-arkivets timesverdier for dypene 2, 20 og 50 m. Dette for å kunne løse opptidevannsbevegelse. Her ble året 2011 valgt ut for å representere strømhastighet da dette året er antatt å ha høy frekvens av sterk strøm på grunn av den høye NAO-indeksen det året. NAO’en (North Atlantic Oscillation, Hurrel, 2003) er en indeks som representerer den atmosfæriske trykkforskjellen mellom Island og Portugal. Positiv verdi tilsier økt frekvens av vestavind på våren, som igjen påvirker de vinddrevne havstrømmene. Gjennomsnittsverdien av strømhastigheter er basert på perioden Oktober 2010 – September 2011. Vi har også beregnet persentiler, men de er kun basert på vinterhalvåret Oktober 2010 – Mars 2011 da den vindpåvirkede strømmen er antatt å være blant de høyeste de siste 20 årene.
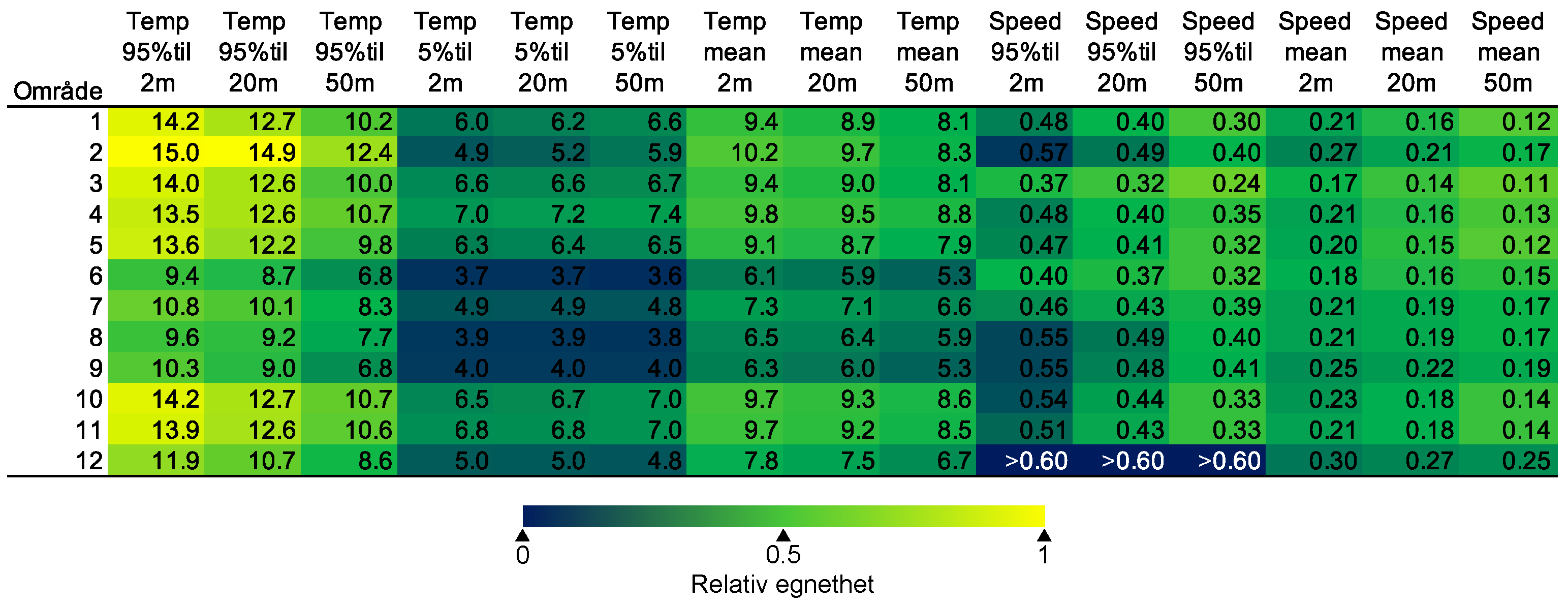
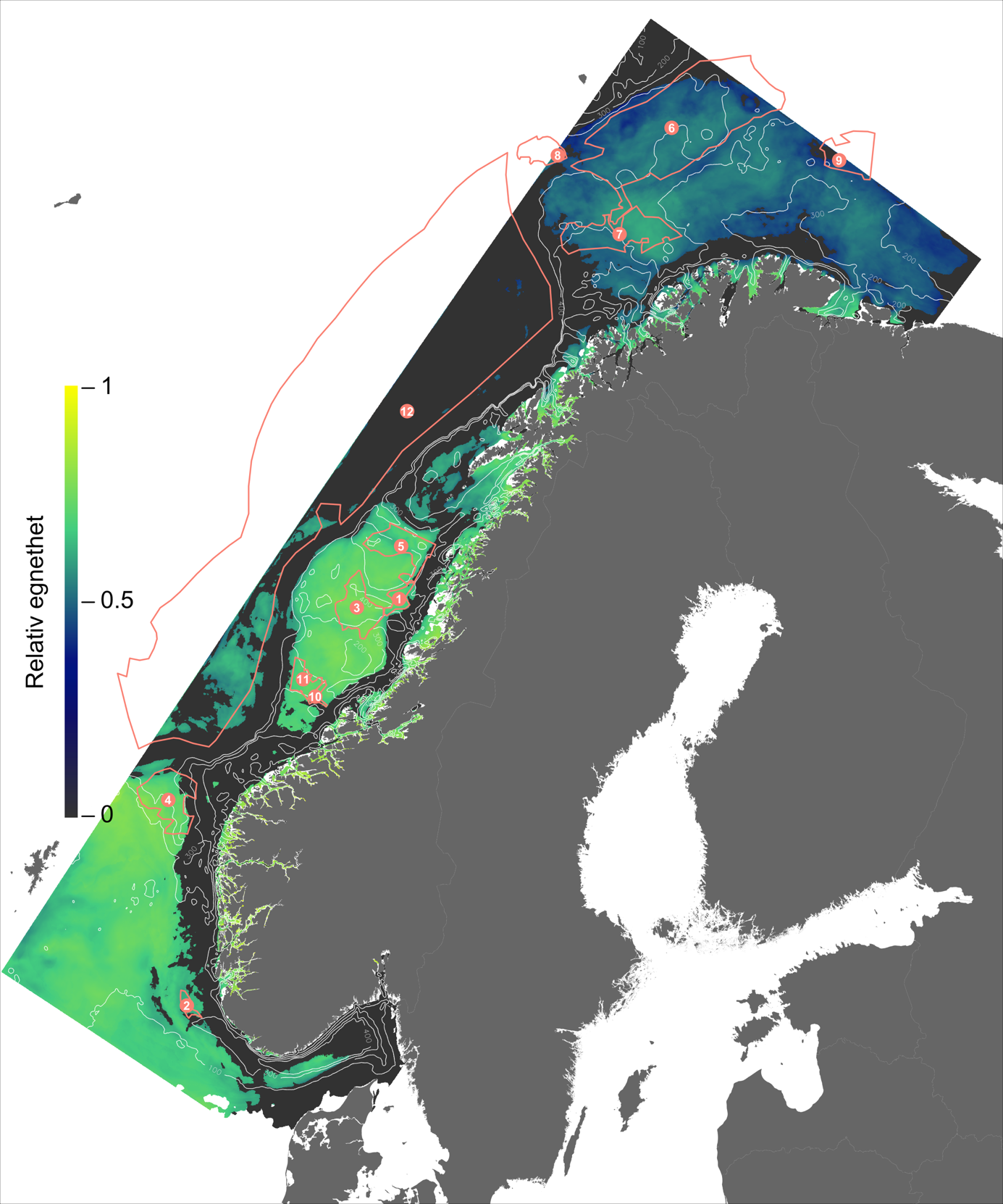
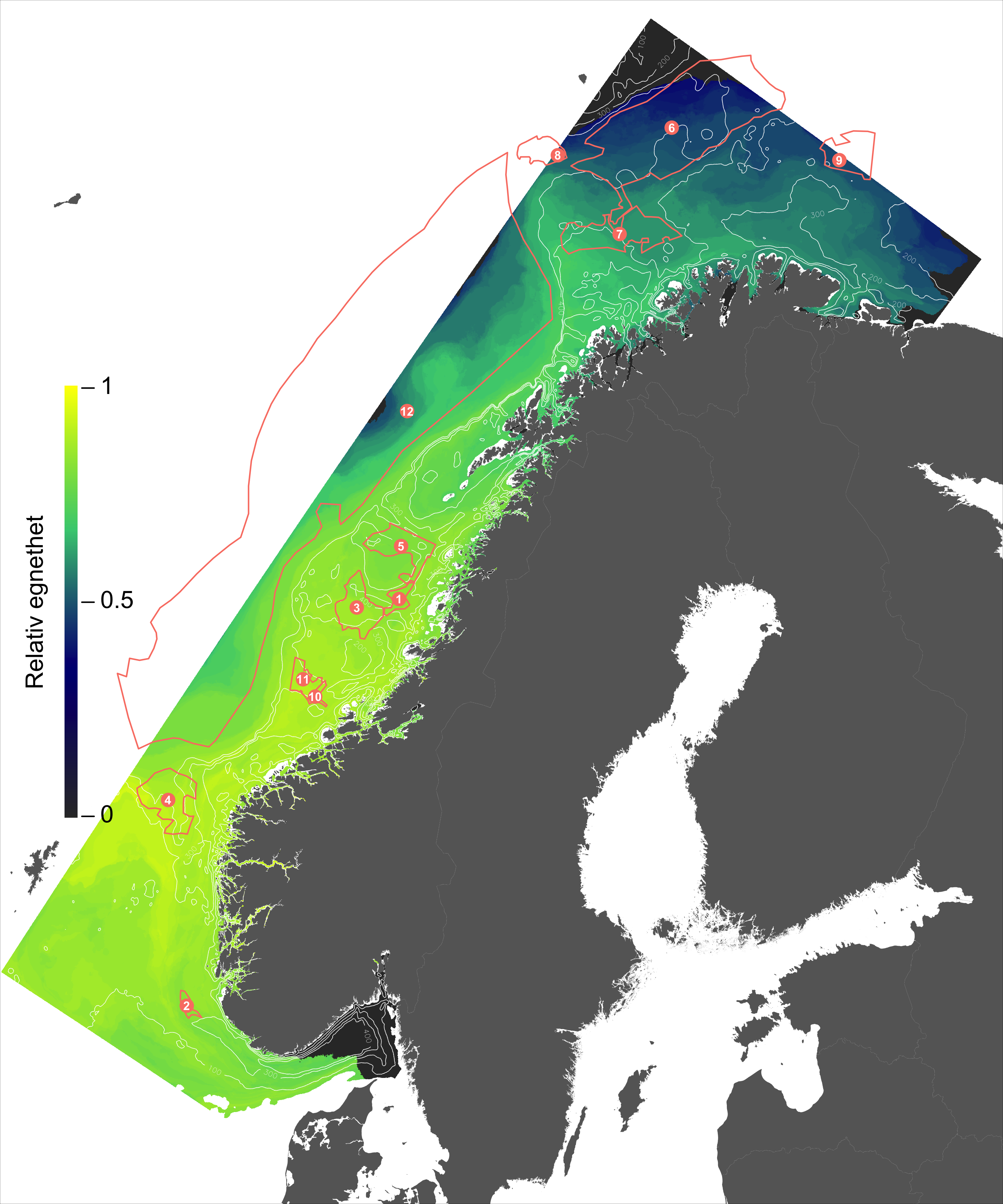
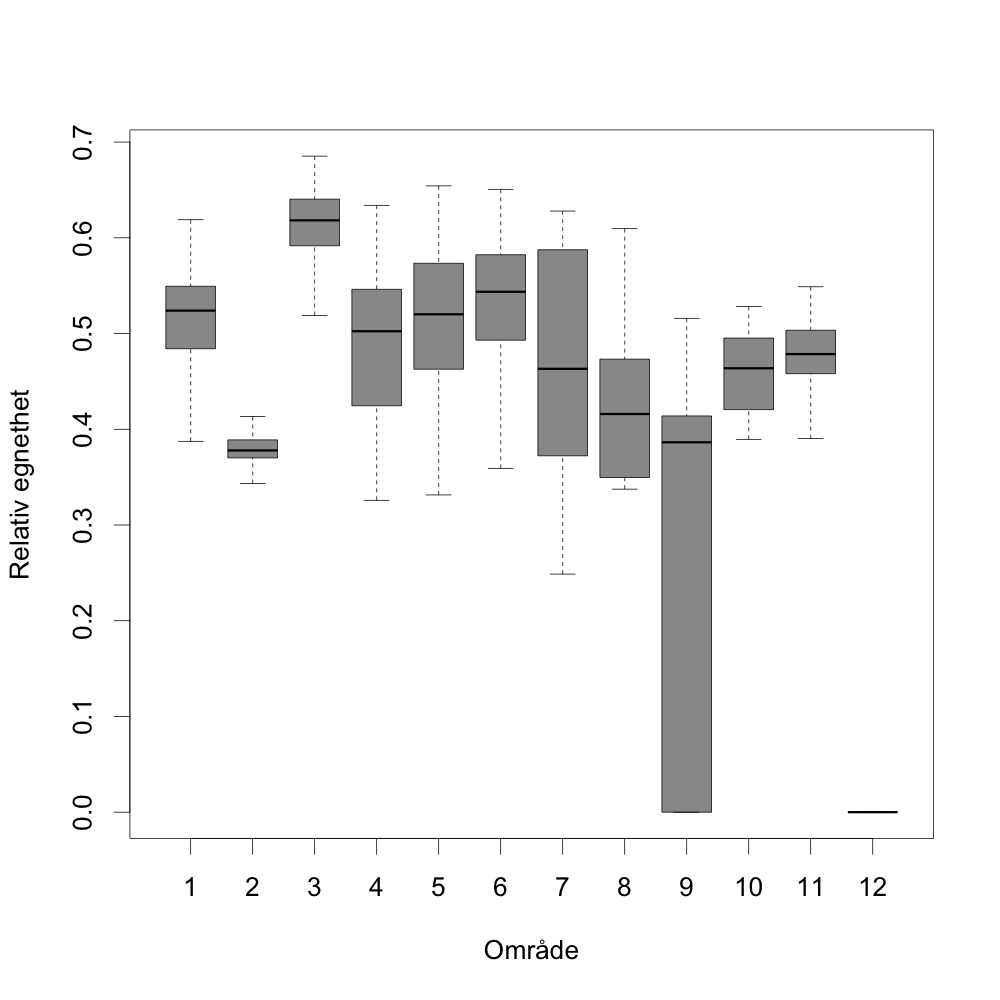
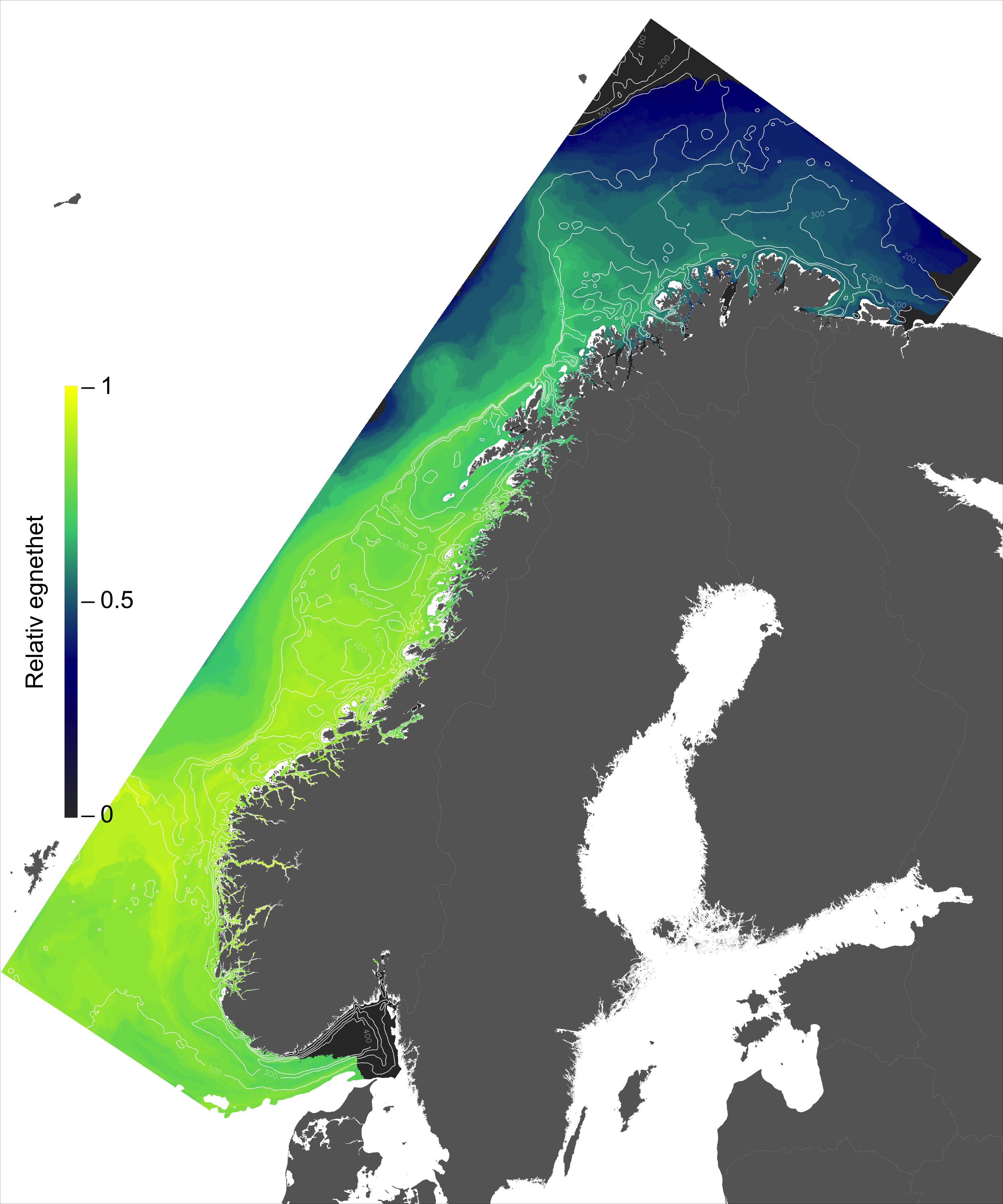
For en vurdering av egnethet av områder for havbruk til havs, basert på modellert strøm og temperatur, vurderes ulike strøm- og temperaturkarakteristikker (se Figur 2.1, 2.2 og Appendix med Figurer 9.1-9.14) i forhold til laksens svømmeevne og temperaturtoleranse. Her ble kartlagene som representerer de høyeste strømhastighetene (95 persentil i 2, 20 og 50 m) vurdert mot laksens kritiske svømmehastighet (her satt til 0,6 m/s, se Hvas et al., 2019 for definisjon av «Ucrit») og midlere strømhastighet vurdert mot laksens vedvarende svømmekapasitet, her definert som 80% av den kritiske svømmehastighet (0,8*0,6 m/s = 0,48m/s, se Hvas et al., 2019 for definisjon av «sustained swimming speed»). I tillegg vurderes temperaturpåvirkning på laksens svømmeevne, der gitterruter med temperatur på 14 °C ga høyest egnethet, og med avtagende egnethet ved høyere og lavere temperaturer (Hvas et al., 2019). Samlet relativ egnethet per gitterrute i de to figurene som oppsummerer resultatene oppgis som middelet over henholdsvis relativ egnethet for de ulike strøm- og temperaturkarakteristikker (normalisert til mellom 0 og 1, der 0 er uegnet og 1 er høy egnethet), såfremt terskelverdiene for strøm ikke brytes. Områder med modellerte strømhastigheter som overstiger kritisk svømmehastighet lik 0,6 m/s (relativ til 95 persentil av modellert strøm) eller vedvarende strømhastighet lik 0,48 m/s (relativ til modellert midlere strøm) skraveres i sort. Se Appendix 9.3 for vurdering av relativ egnethet enkeltvis for hver av de 12 foreløpige foreslåtte områdene som Fiskeridirektoratet har identifisert som mindre konfliktfull mht annen eksisterende bruk og dermed aktuell for havbruk til havs.
Hovedtrekk modellert strøm og temperatur
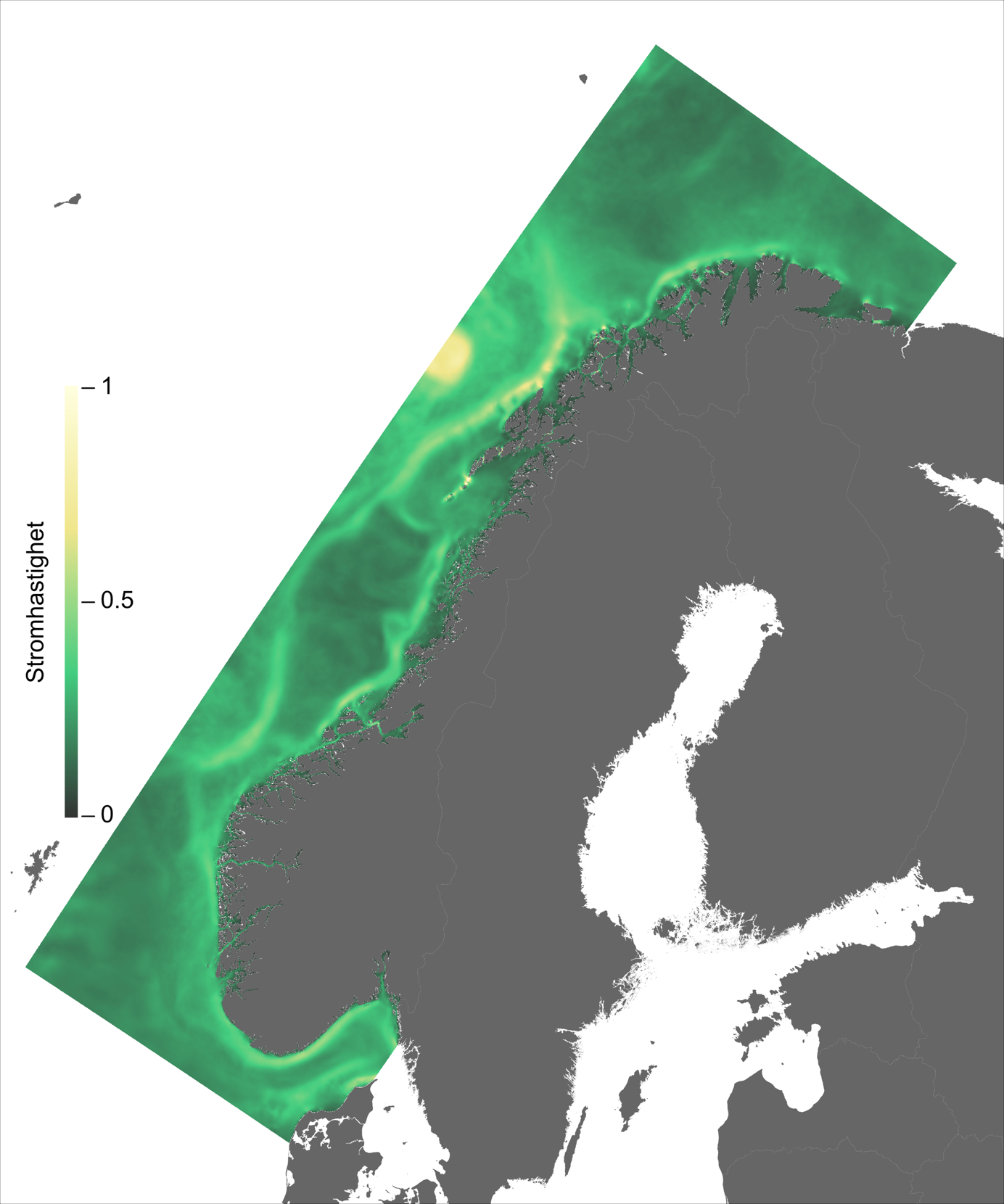
Ett av hovedtrekkene som kjennetegner strøm i modellområdet er den Norske kyststrøm, som brer seg nordover som et belte nær land helt fra Svenskegrensen til Finnmark. Bredden og styrken på kyststrømmen varierer betydelig, fra smal og konsentrert langs deler av Sørlandskysten til en bred (> 100 km om sommeren) og mer diffus strøm over Norskerenna på Sørvestlandet med en rekke hvirvler som brer seg nordover (Figur 2.1).
Ved Tampen, nord for Vikingbanken, kommer også den Norske Atlantarhavsstrømmen inn i modellområdet. Her bøyer noe av Atlanterhavsvannet av mot sørøst og inn på vestsiden av Norskerenna, men før 57 °N er allerede mesteparten av Atlanterhavsvannet på vei ut igjen på østsiden av Norskerenna, under kystrømmen. Hoveddelen av Atlanterhavsvannet beveger seg derimot nordover, styrt av kontinentalskråningen, med noen krysninger inn på sokkelen i de mange dype ravinene i sokkelkanten.
I tillegg til de tetthetsdrevne/vinddrevne strømmene kommer tidevannet som kan ha stor påvirkning på strømhastigheten i trange sund og i fjorder.
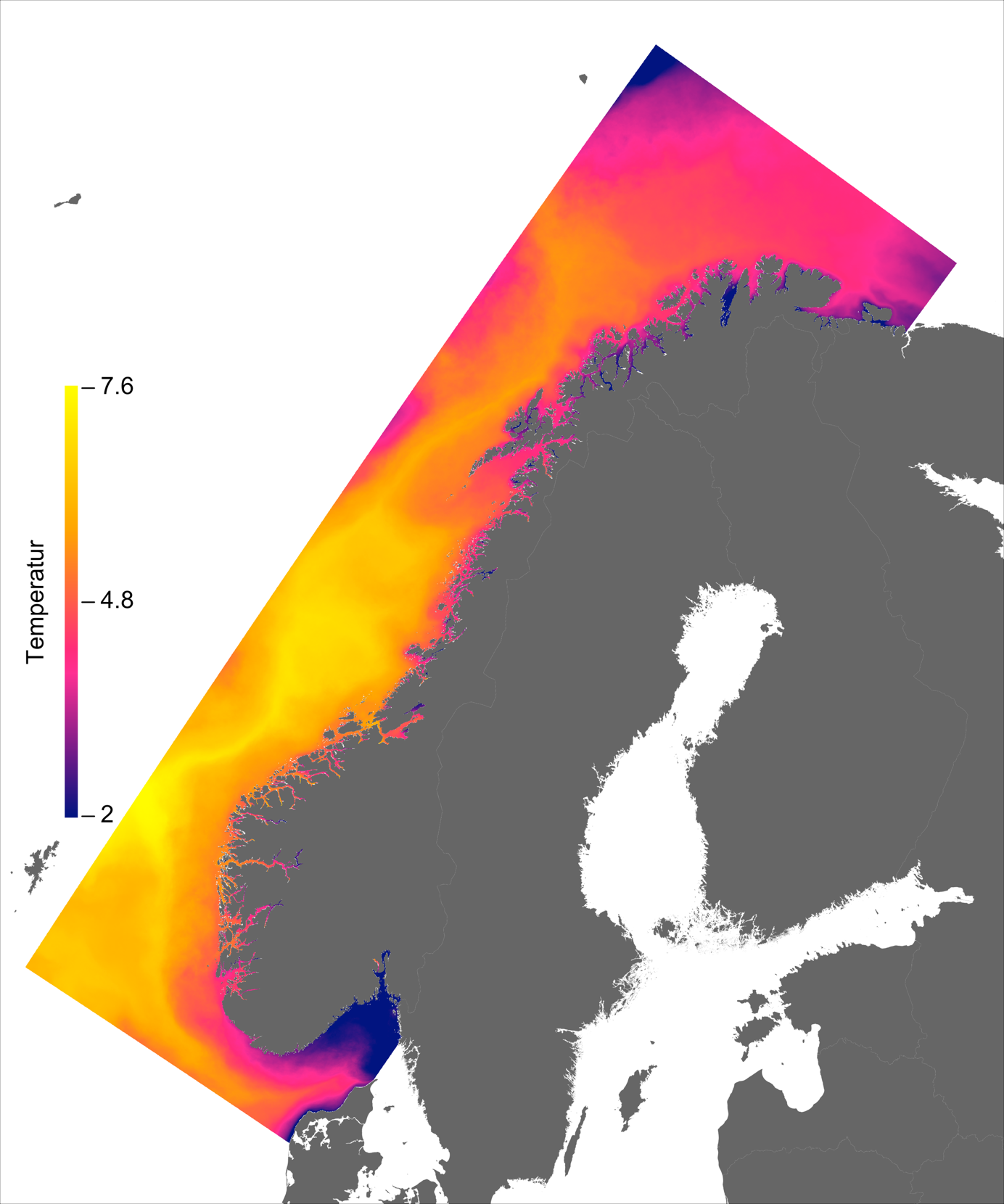
Temperaturen opptrer ikke like dynamisk som strømmene, men følger i stor grad sesongsyklusen. Kystvannet finner man stort sett i fjordene og i kyststrømmen, mens Atlanterhavsvannet ligger under kystvannet langs store deler av kysten og opptrer kun i overflaten langs kontinentalskråningen og over Norskerenna. Mens temperaturen i kystvannet varierer betydelig med sesong, fra ned mot 0 °C på det kaldeste om vinteren og opp mot 18 °C på det varmeste om sommeren, holder Atlanterhavsvannet en jevn temperatur over 8 °C gjennom hele vinteren og opp mot 14 °C om sommeren (Figur 2.2).
2.2 - Modellert strøm og temperatur i lys av fiskevelferd
Havbruk til havs vil bringe nye utfordringer for fiskehelse sammenlignet med skjermet havbruk inne i fjordene. Dette gjelder spesielt strømhastighetene i vannmassene rundt fisken i forhold til dens svømmeevne. Hvas et al. (2019) oppsummerer tålegrenser for laks av ulike størrelser, for ulike strømeksponeringsperioder, ved ulike omgivelsestemperaturer og ved ulike tilstander av laks (for eksempel eksponert for smitte). Den mest gunstige temperatur for svømmeevne er funnet til å være mellom 13 - 18 °C og alt annet påvirker svømmeevne negativt.
Ved høye temperaturer (> 22 °C) vil laksen oppleve høy dødelighet. Basert på de modellerte temperaturene kan vi slå fast at slike høye temperaturer oppleves uhyre sjeldent til havs i våre områder og høy temperatur vil dermed gi få føringer for hvor produksjonen kan finne sted. Det er imidlertid verdt å merke seg den nye spesialrapporten til IPCC om fremtidige forhold i hav og is (IPCC, 2019) som peker på økt hyppighet av marine varmebølger.
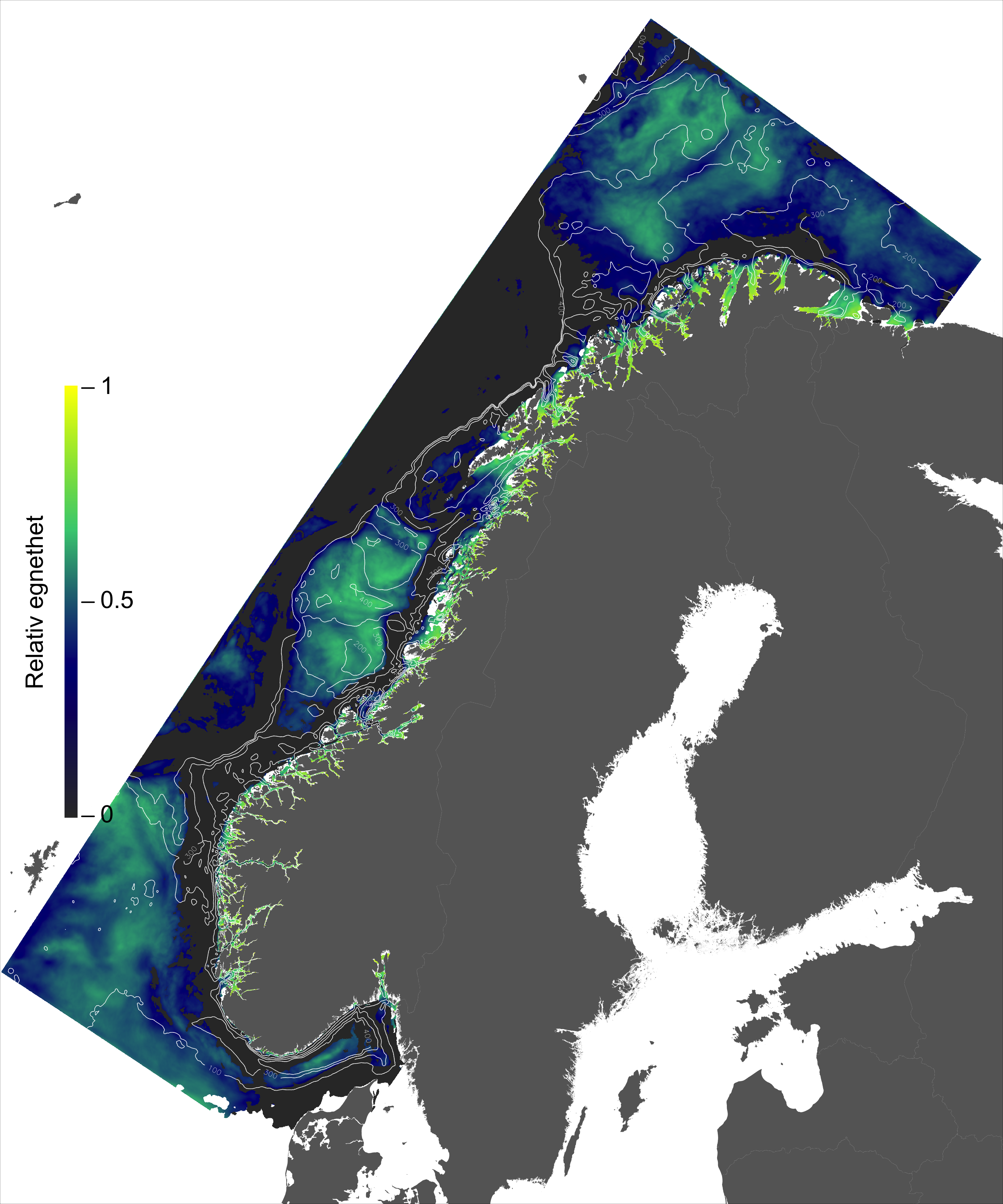
I de mer Atlanterhavspåvirkede områdene langs kontinentalskråningen vil temperatur være nær optimal året rundt, men her er også strømhastighetene høyest. De vil være over laksens vedvarende svømmeevne (> 0.8*0.6 m/s, se Hvas et al., 2019) store deler av tiden. Mer interessant fremtrer derimot havområdene som hverken domineres av kyststrømmen eller Atlanterhavsstrømmen. Fra sør til nord utpeker følgende områder seg som aktuelle basert på egnede strøm- og temperaturforhold (Figur 2.3a); sørvestlige deler av Skagerrak, store deler av Nordsjøen vest for Norskerenna men sør for Atlanterhavsstrømmen, kontinentalsokkelen langs den midt-Norske kysten mellom kyststrømmen og Atlanterhavsstrømmen omtrent fra Hitra til Træna, og store deler av Barentshavet. Merk imidlertid at Barentshavet generelt fremstår som mindre egnet sammenlignet med områdene lenger sør på grunn av de lave temperaturene med tilhørende reduksjon i laksens ytelse og dermed tåleevne for strøm (Figur 2.3b).
Strøm omtalt her er i de frie vannmasser og vi vurderer ikke skjermingseffekter på strøm innen i merder. Dette vil være avhengig av havbruksinstallasjonenes utforming.
2.3 - Observert strøm
Observert strømhastighet er en sum av ulike fenomener som tidevannsstrøm, vinddrevet strøm (Stokes drift - forflytning som følge bølgebevegelse - se for øvrig 3.1, Ekman-drift) og stokastisk drift (virvler av ulike skalaer). Både topografi og vannets tetthet påvirker realisert strøm som følge av disse prosessene og er en viktig årsak for hvorfor strømmodeller avviker fra observert strøm.
Maksimum strøm vil være når de overnevnte komponenter er maksimum og i fase (samme retning). Dette betyr at det vil være en bias i korte observasjonsserier hvor en typisk vil underestimere maksimum strøm.
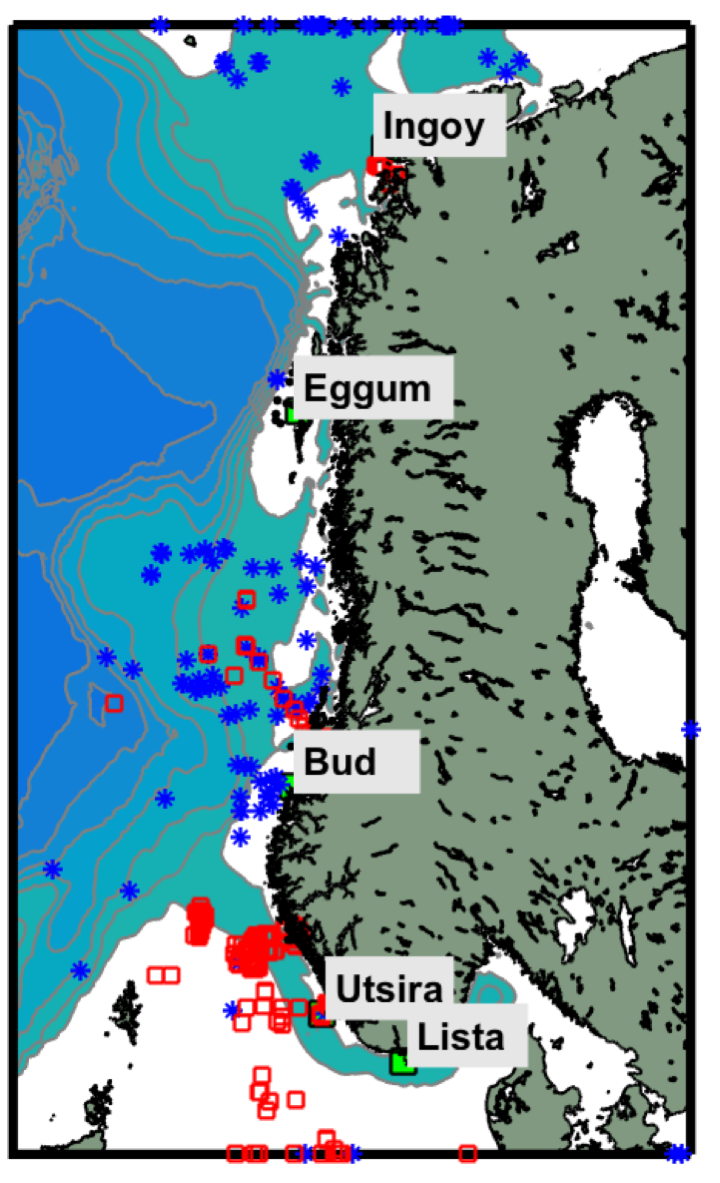
Vi har sammenstilt eksisterende observasjonsserier av strøm fra bunnforankrede rigger langs kysten (Figur 2.4). Observasjoner på den norske kontinentalsokkelen er i all hovedsak utført i forbindelse med petroleumsvirksomhet. Via kontakt med Equinor v / Einar Nygård har vi fått tilgang til strømobservasjoner utført / finansiert av oljeselskapene for perioden 1980 – 2010. I hovedsak er seriene relativt korte, kun 1-2 måneder (Figur 2.5). Merk imidlertid at i Vesterålen er det svært god datadekning med ni profilerende strømrigger som inkluderer en full år-syklus. I tillegg har vi inkludert observasjoner utført av HI i 2018 vest av Øygarden og Smøla med varighet 7-9 måneder.
Merk også at det foreligger strømmålinger på Svinøy-snittet (Orvik and Skagseth, 2005) og Fugløy-snittet (Ingvaldsen et al., 2006) men at disse ikke er inkludert her. Eksempelvis dekker strømmålinger fra Svinøysnittet kun områder utenfor sokkelkanten.
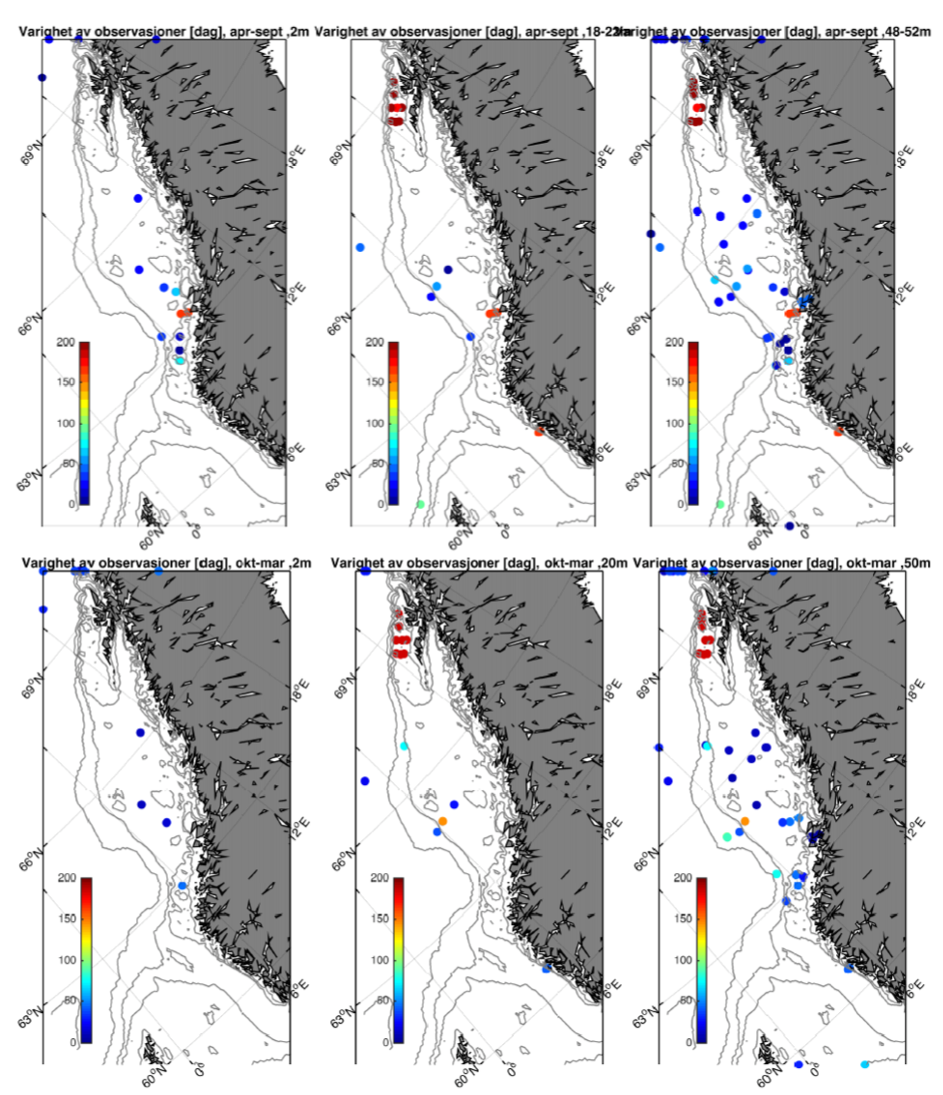
Fokuset i denne rapporten er øvre del av havet og vi har derfor valgt å fokusere på tre ulike dyp; 2, 20, og 50 m. Som et kompromiss mellom å skille mellom ulike årstider med klimatologiske forskjeller i værforhold og samtidig ha tilstrekkelig med data, har vi delt året inn i en sommerperiode (april-september) og en vinterperiode (oktober-mars). Generelt er hyppighet og styrke på lavtrykk vesentlig sterkere i vinterhalvåret og derfor forventer vi også at dette vil gjenspeiles i relativt økte strømhastigheter.
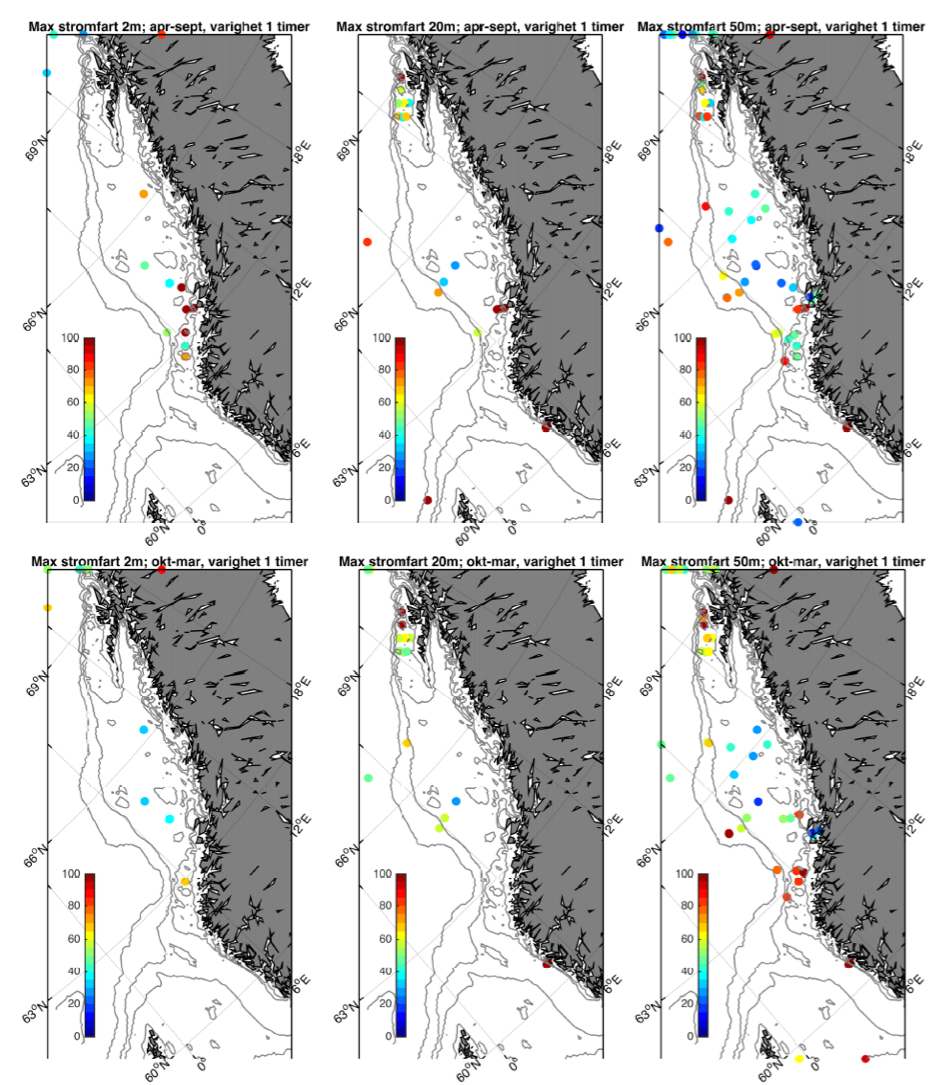
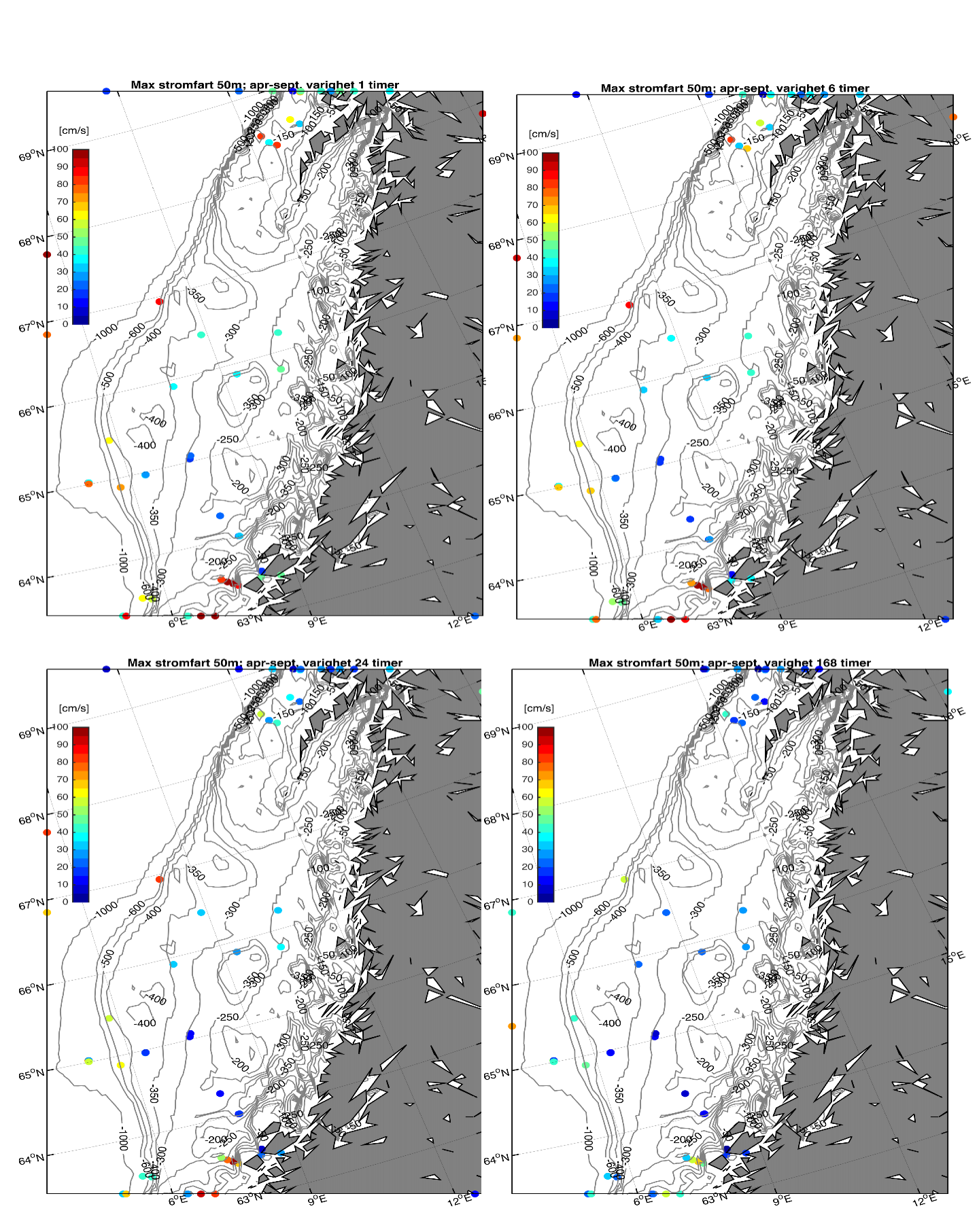
Fra Hvas et al. (2019) vet vi at laksens evne til å svømme mot strømmen er avhengig av varighet på hendelsene.Vedvarende strømeksponering er beregnet til å være 80 % av kritisk svømmehastighet. For å adressere dette presenterer vi maksimumstrøm for 1, 6, 24, og 168 timers middelverdier.
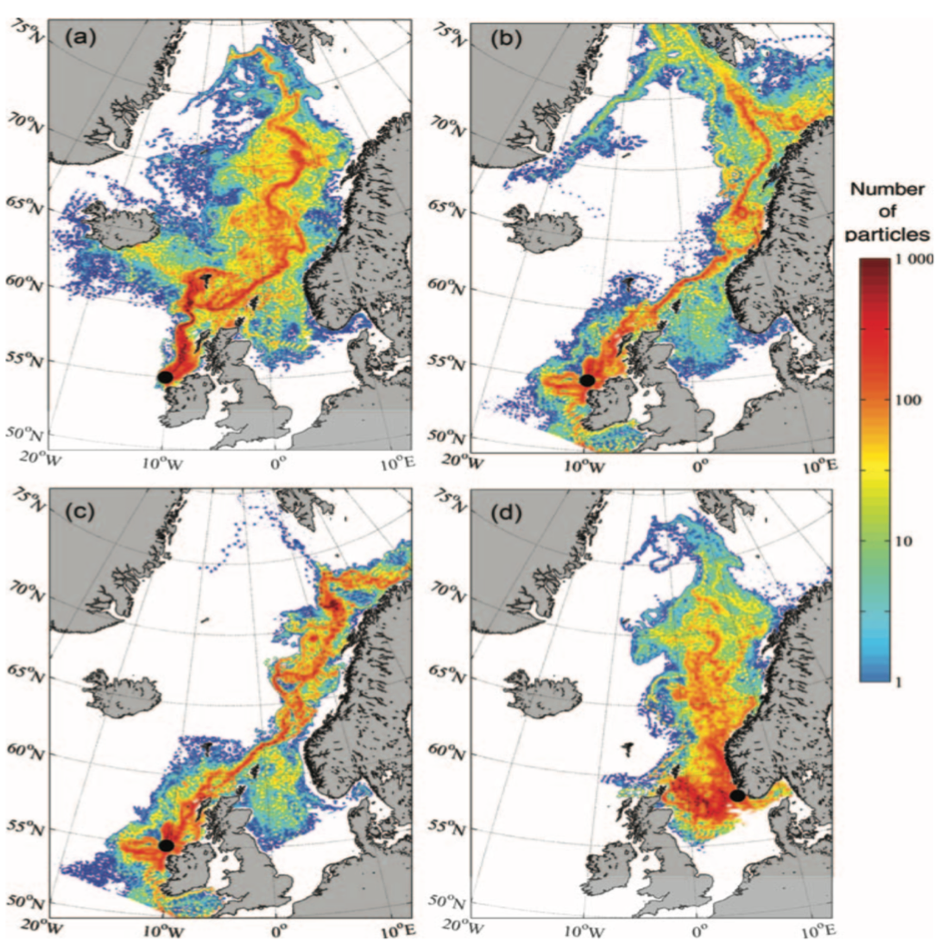
Figur 2.6 viser et generelt minimum i strømhastighet mellom kantstrømmen på sokkelkanten og kyststrømmen tettere ved kyst, både på bankene og de flatere områdene mellom omtrent 64-67 °N. Sør og nord for dette område, på kontinentalskråningen og i rennene inn mot kysten, er strømmen typisk sterkere. Generelt kan vi se at maksimal strømhastigheter mellom kantstrømmen og kyststrømmen er mindre enn det som i Hvas et al. (2019) oppgir som vedvarende svømmekapasitet, altså 0.48 m/s.
I fortsettelsen fokuserer vi på målinger på 50 m dyp som både har den beste observasjonsdekningen samtidig som usikkerheten trolig er mindre enn målinger fra 2 og 20 m.
Figurene for observert strømhastighet er vist som maksimum strømhastighet av varighet lengre enn 1, 6, 24, eller 168 timer på 50 m dyp (Figur 2.7). For alle tidsmidlene er strømhastigheten langs kontinentalskråningen, på bankene og dyprennene ved Møre, i kyststrømmen, og på Vesterålbankene over tålegrense for laks. I tillegg er det å forvente at strømhastighet nær overflaten vil overstige det vi måler på 50 m på grunn av sterkere vinddrevet strøm. Dette gjelder både for vinter- og sommerhalvåret. Imidlertid er målt strømhastighet på bankene og de flatere områdene mellom omtrent 64-67 °N lavere enn for laksens tålegrense. Basert på målt strøm nær overflaten (20 og 2 m) er de stadig under tålegrense, men disse målingene må altså regnes som relativt usikre.
2.4 - Sammenligning av observert og modellert strøm
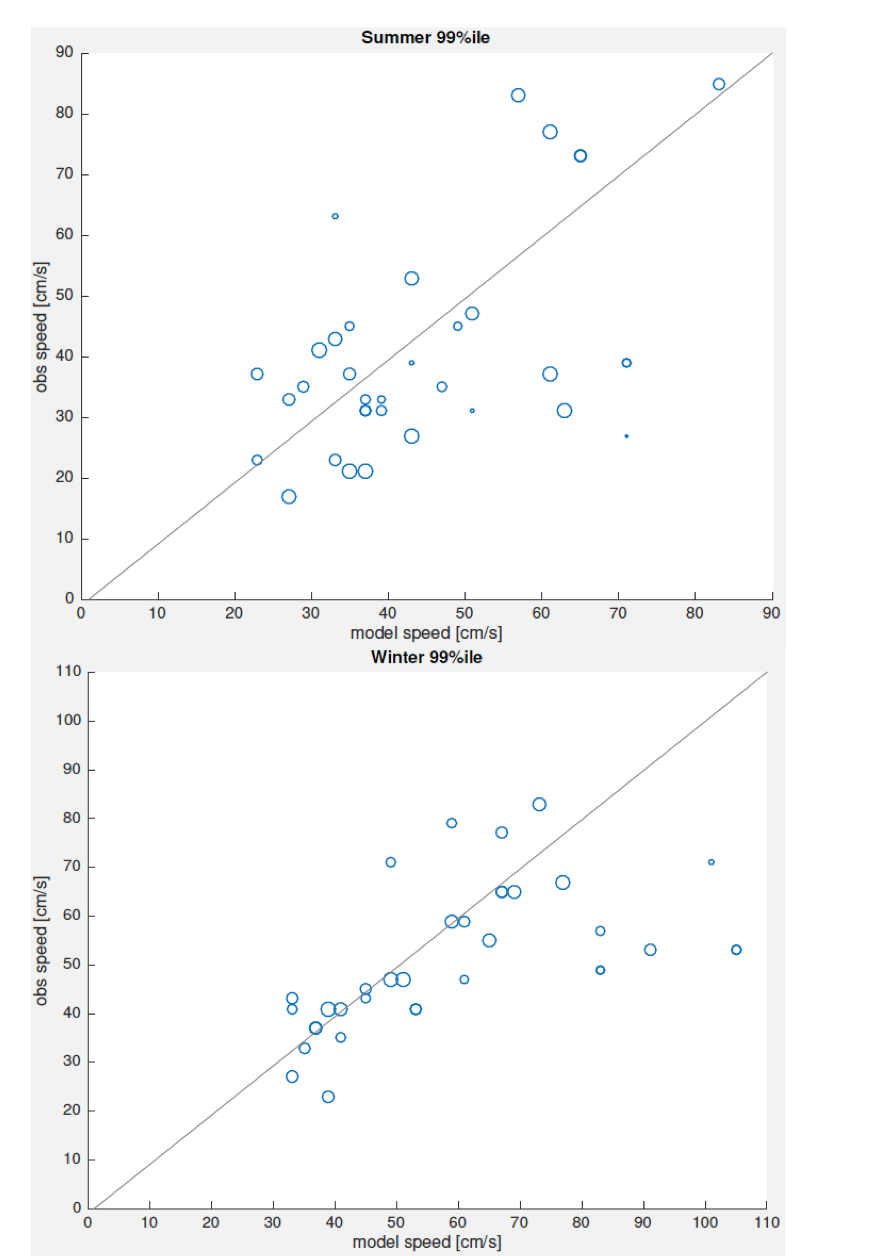
Sammenligning mellom observert og modellert strømhastighet er viktig for å vurdere om det er systematiske forskjeller, enten under- eller overestimat, i modellert strøm. Siden observert og modellert strøm ikke er fra samme periode, og målingene typisk er av kortere varighet, har vi sammenlignet persentiler av strømhastighet for modell mot observasjon i samme posisjon og dyp (Figur 2.8).
Sammenligningen av modellert og observert strøm samsvarer godt. Generelt kan man si at for sommerhalvåret er det ikke systematiske forskjeller, mens for vinterhalvåret det det en tendens til at modellert strøm ligger noe høyere for de sterkeste strømmene. Det er liten tendens til at modellen underestimerer strømhastighet. Det betyr at vi relativt sikkert kan anta at modellert strømfart er realistisk på 50 m dyp. Det betyr at «grunnstrømmen» som den modellerte overflatestrømmen rir på er rimelig, og det er en nødvendig betingelse for at strømmen i øvre lag skal være riktig. Siden prosessene for hvordan strømmen varierer mot overflaten er representert i havmodellen basert på beste kunnskap om overflatestrøm kan vi anta at strømhastighet nærmere overflaten også er realistisk.
2.5 - Referanser
Albretsen J, Sperrevik AK, Sandvik AD, Asplin L (2011) NorKyst-800 Report No. 1 User Manual and technical descriptions.
Beldring S, Engeland K, Roald LA, Sælthun NR,Voksø A (2003) Estimation of parameters in a distributed precipitation-runoff model for Norway, Hydrology and earth System Sciences, 7 (3): 304-316.
Egbert GD, Erofeeva SY (2002) Efficient Inverse Modeling of Barotropic Ocean Tides. J. Atmos. Oceanic Technol., 19,183204.
Hurrel, James W. (2003). The North Atlantic Oscillation: Climatic Significance and Environmental Impact. American Geophysical Union. ISBN 9780875909943.
Hvas M, Folkedal O, Oppedal F (2019) Havbasert oppdrett – hvor mye vannstrøm tåler laks og rensefisk? Fiskevelferd og grenseverdier. Rapport fra Havforskningen nr x.
Ingvaldsen R, Asplin L and Loeng H (2006) Volumefluxes into the Barents Sea from current measurements and numerical modelling. ICES JMS.
IPCC, 2019: Summary for Policymakers. In: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate [H.- O. Pörtner, D.C. Roberts, V. Masson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, M. Nicolai, A. Okem, J. Petzold, B. Rama, N. Weyer (eds.)].
Lien VS, Gusdal Y, Albretsen J, Melsom A, Vikebø FB (2013) Evaluation of a Nordic Seas 4 km numerical ocean model hindcast archive (SVIM), 1960-2011. Fisken og Havet 7/2013.https://brage.bibsys.no/xmlui/bitstream/handle/11250/113861/FoH_7-2013.pdf?sequence=1
Lien VS, Gusdal Y, Vikebø, FB (2014) Along-shelf hydrographic anomalies in the Nordic Seas (1960–2011): locally generated or advective signals? Ocean Dynamics 64: 1047.https://doi.org/10.1007/s10236-014-0736-3
Orvik KA and Skagseth Ø (2005) Heat flux variations in the eastern Norwegian Atlantic Current toward the Arctic from moored instruments, 1995-2005. Geophysical Research Letters 32.
3 - Havbølger
Havbølger kan påvirke installasjoner til havs og medføre rømning eller redusert fiskevelferd. Som beskrevet i Hvas et al. (2019) er effektene av bølger på havbruk lite studert. Men det er åpenbart at for havbruk til havs blir dette en mer aktuell problemstilling. Da er det nødvendig å kjenne til bølgemiljøet for ulike potensielle lokaliteter og effekter på installasjoner og fisk ved ulike bølgehøyder og -perioder. I det følgende oppsummeres bølgemiljø for kysten og tilgrensende hav basert på bølgemodellen som brukes operasjonelt på Meteorologisk Institutt.
3.1 - Bakgrunn
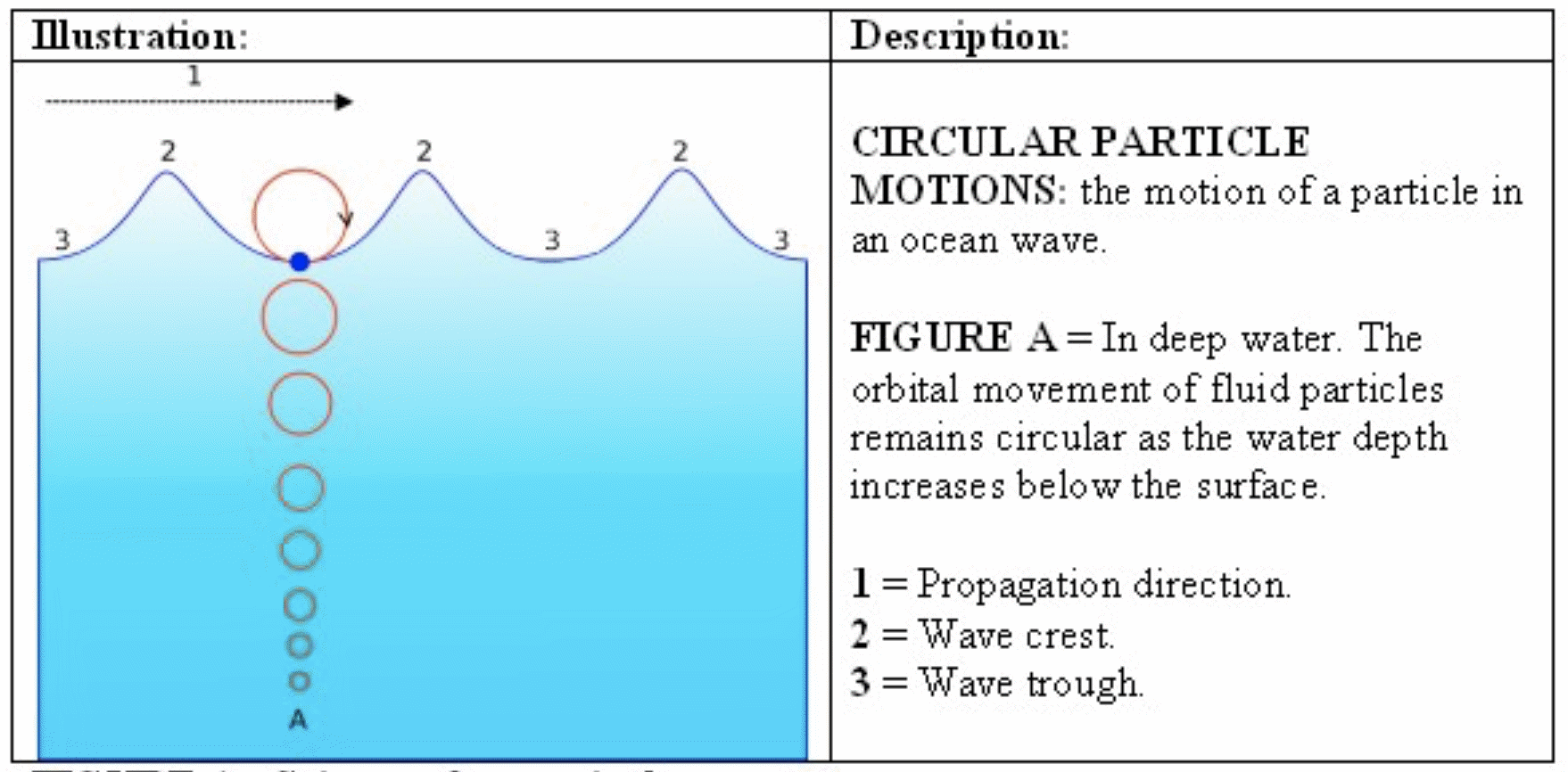
En havbølge er en overflatebevegelse som opptrer i det øverste laget i havet og brer seg som en bølge. Havbølger skyldes vanligvis vinder og kan reise store avstander før de slår mot land. De kan variere i størrelse fra små krusninger til flere titalls meter høye. Det er lite bevegelser av individuelle vannpartikler i bølgeretningen til tross for den store energimengden og kraften den dytter forover med. I stedet beveger de seg i hovedsak i ellipser. Når havdypet er større enn bølgelengden, vil de enkelte vannpartiklene i bølgen tilnærmet bevege seg i sirkler. Disse sirklenes radius avtar raskt nedover i dypet.
I bølgetoppen og i dypet under den vil vannet bevege seg horisontalt og i samme retning som bølgen. I bølgedalen og i dypet under den vil vannet bevege seg horisontalt og i motsatt retning. Mellom bølgetopp og bølgedal vil vannet bevege seg vertikalt, opp foran toppen og ned bak toppen, sett i bølgeretningen (se Figur 3.1). Denne sirkelbevegelsen er ikke helt uten fremdrift, fordi vannpartiklene har litt større fart fremover i bølgetoppen enn de har bakover i bølgedalen. Denne fremdriften kalles Stokesdrift.
Vindsjøen er sammensatt av en mengde bølgetog med forskjellige periodee og bølgehøydee og må derfor beskrives statistisk. Man utarbeider diagrammer som angir bølgeenergiens fordeling (frekvens) på de forskjellige bølgeperioder og -høyder (bølgespektre).
Man oppgir gjerne den signifikante bølgehøyden, som er middelverdien av den tredjedel av bølgene som er høyest. Den maksimale bølgehøyde kan bli mye større enn dette, og forekommer ved samvirkning (interferens) mellom to eller flere bølger. Man har observert bølger på 20–30 meter i Nordsjøen/Norskehavet. Særlig ved konstruksjoner som står på havbunnen, f.eks. petroleumsplattformer, er man interessert i å vite den største bølgehøyde som kan inntreffe f.eks. i de neste hundre år (hundreårsbølgen). Denne beregnes ut fra observerte og/eller modellerte bølgedata sammen med visse statistiske antagelser. Ved Utsira er f.eks. hundreårsbølgen beregnet til 23 m, men den kan bli enda større i åpent hav. Kilde: https://snl.no/havbølger.
Bølgeformen forplanter seg normalt med en hastighet som avhenger av bølgelengden (horisontal avstand mellom to nabobølgetopper).
Sammenhengen mellom bølgelengde og periode kan settes opp i følgende formel når havdypet er større enn bølgelengden, L=gT2/2π. Her står L for bølgelengden, g står for tyngdens akselerasjon, T står for bølgenes periode og tegnet π er den matematiske størrelsen pi. Ifølge formelen vil f.eks. en bølge med periode på 5 sekunder være 39 m lang, en bølge med periode på 10 sekunder være 156 m lang og en bølge med periode på 15s være 351 m lang.
3.2 - Dannelse av bølger
De fleste bølger er resultat av vind, og flere faktorer har innvirkning på dannelsen av vindbølger:
-
Vindhastigheten
-
Hvor lenge vinden har blåst over et gitt område
-
Avstanden vinden har blåst over hav (strøklengden)
-
Topografien
Disse faktorene avgjør i stor grad størrelsene og formene på bølgene. Dess større en av de tre første variablene er, dess større blir bølgen.
Bølgeenergien er styrt av en balanse mellom lastene som vil øke bølgene og forhold som demper dem ut. Det er en rekke forhold som påvirker bølgene. Bølgeenergien er fordelt på flere bølgefrekvenser og vil bl.a. ha følgende påvirkninger:
-
Bølgebryting vil redusere energien i bølgene.
-
At vanndybden varierer. For eksempel vil en strand virke som bølgedemper. Generelt er bølgene størst på dypt vann der påvirkningen av havbunnen er liten.
-
At man kan ha kunstige bølgedempere som forstyrrer bølgene og dermed tar ut energi. Topografien og kunstige endringer av topografien, som ved moloer, vil dempe bølger.
-
Bølger med ulike retninger og størrelser kan påvirke hverandre og en kan få resonanseffekter. De kan ha opphav i dønninger eller andre lavtrykk i området.
-
Høye vindhastigheter påvirker bølgeprofilet.
-
Når bølgehastigheten er større enn vindhastigheten eller går mot bølgene, vil vinden dempe bølgene, og bølgene kan gi energi til vinden.
Strøm mot bølgene vil kunne medføre bølgebryting og reduksjon i bølgeenergien. Strøm med bølgene kan endre formen på bølgene og energifordelingen mellom ulike bølgefrekvenser
3.3 - Beregning av bølger
Det er utviklet flere metoder for varsling av vindsjø og dønning. Slik varsling, som inngår i varsling av sjøgang, har betydning for kystområder og havneanlegg, og selvsagt også for skipsfarten. Virkningen på kystområder vil også avhenge av havbunnens form. Meteorologisk institutt har utviklet og tilpasset den numeriske bølgemodellen WAM («The Wave Model», Hasselmann et al., 1988), og lager varsler for alle de nærliggende havområdene med denne flere ganger om dagen. WAM har vært i drift hos Meteorologisk institutt siden 1988 og er et viktig varslingsverktøy. Den horisontale oppløsningen for de nordiske hav er 4 x 4 km. WAM beregner todimensjonale bølgespektra, med 25 frekvenser og 24 retninger, og i denne analysen har vi hentet ut signifikant bølgehøyde, bølgetopp-periode (kan omregnes til bølgelengde) og Stokesdrift i to retninger. Vi har tatt ut felter fra modellen hver time i ett år (2018) og antar at dette er en representativ periode som fanger opp de fleste bølgesituasjonene. Ut fra timesverdiene har vi beregnet median verdi og 95-persentil for hvert gridpunkt.
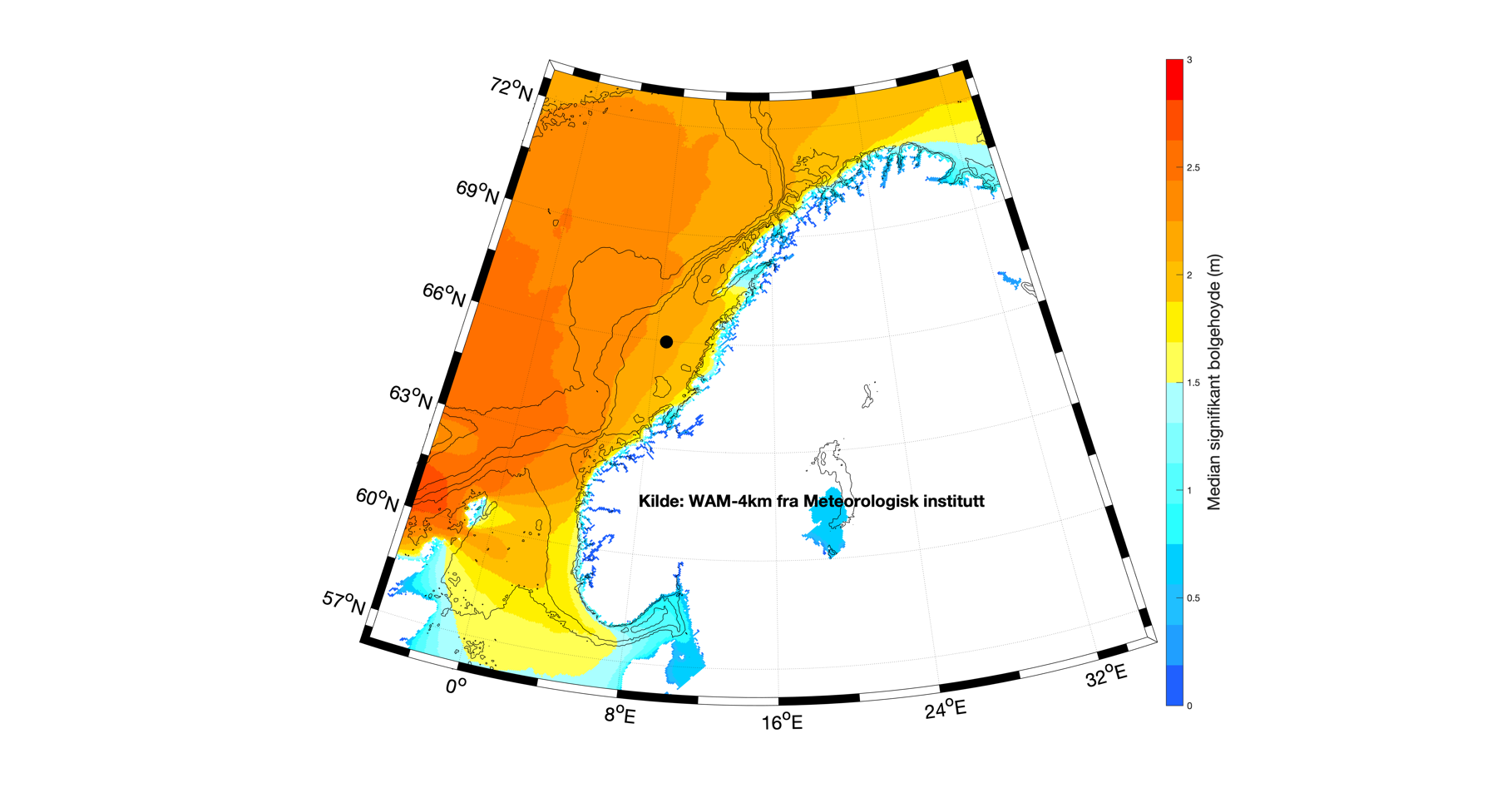
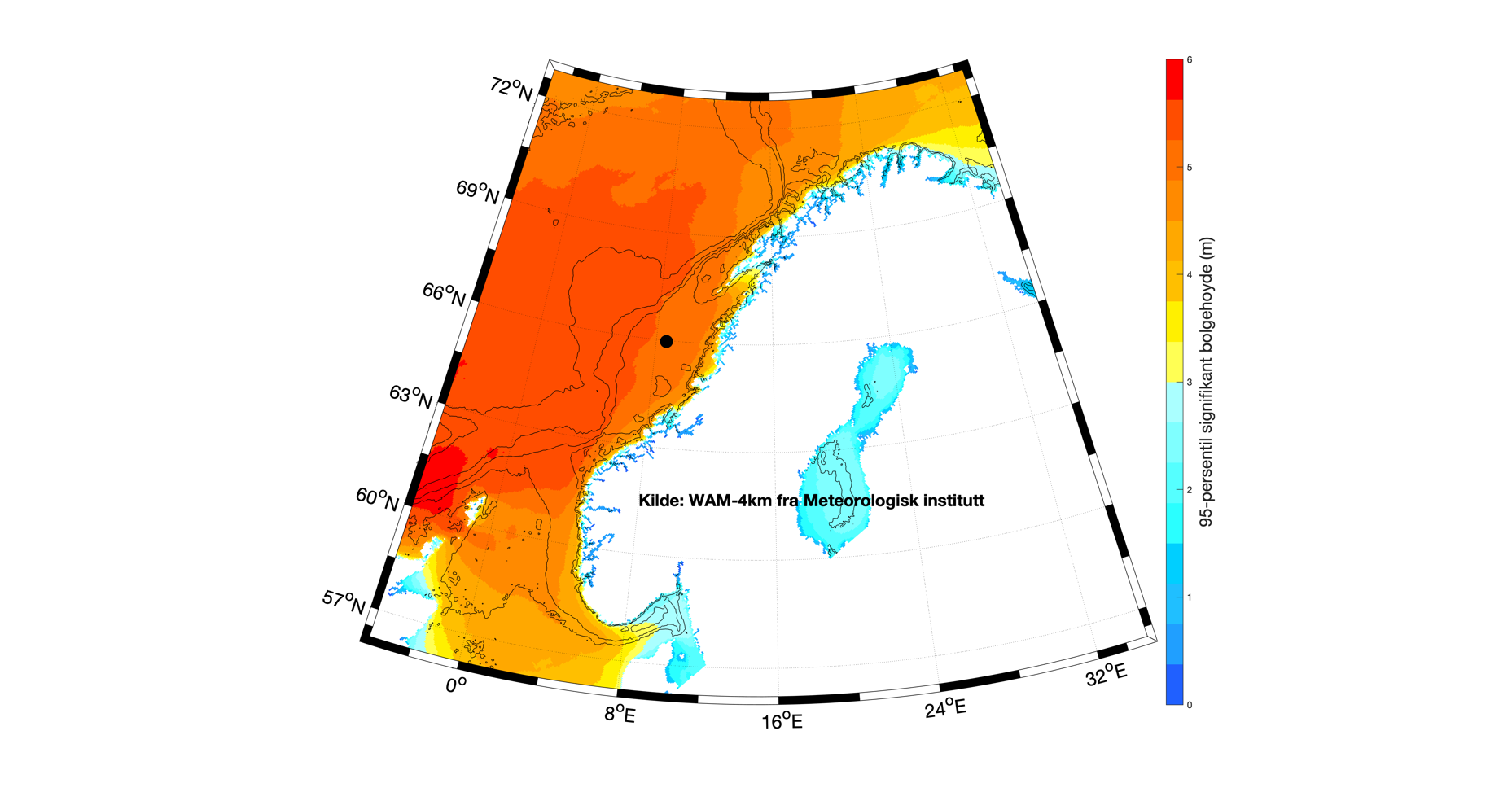
Når man ser på gjennomsnittlig (median) bølgehøyde for Nordsjøen, Norskehavet og områdene langs kontinentalsokkelen, så finner man de største verdiene nær norskekysten fra Sogn og Fjordane og nordover (Figur 3.2). Vi finner en liten gradient av mindre bølger inn mot land, men det vil først og fremst være de grunnere dybdeforholdene som reduserer bølgehøyden. Kartet som viser 95-persentilen av sign. bølgehøyde (Figur 3.3) gir stort sett det samme mønsteret som middelverdien, men verdiene er rundt det dobbelte. Merk at maksimal bølgehøyde vil overgå dette, og i tillegg at enkeltbølger kan være 2-3 ganger høyere enn maksimal signifikant bølgehøyde.
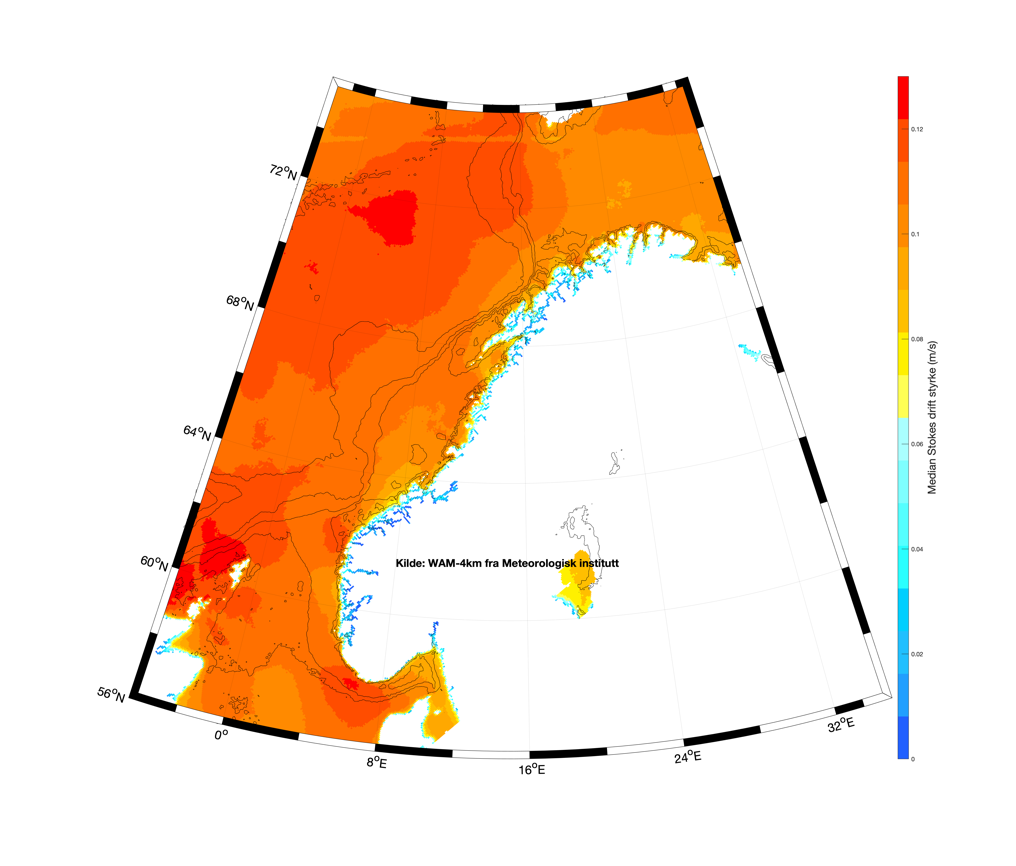
Stokesdriften, selve fremdriften til bølgetogene, kan man også hente ut av de samme bølgemodellene, og median verdi er angitt i Figur 3.4. Verdiene over Vøringplatået og langs norskekysten er relativt jevne med drift rundt 0,1 m/s. De høyeste styrkene av Stokesdriften vil da ligge typisk rundt 0,2 m/s.
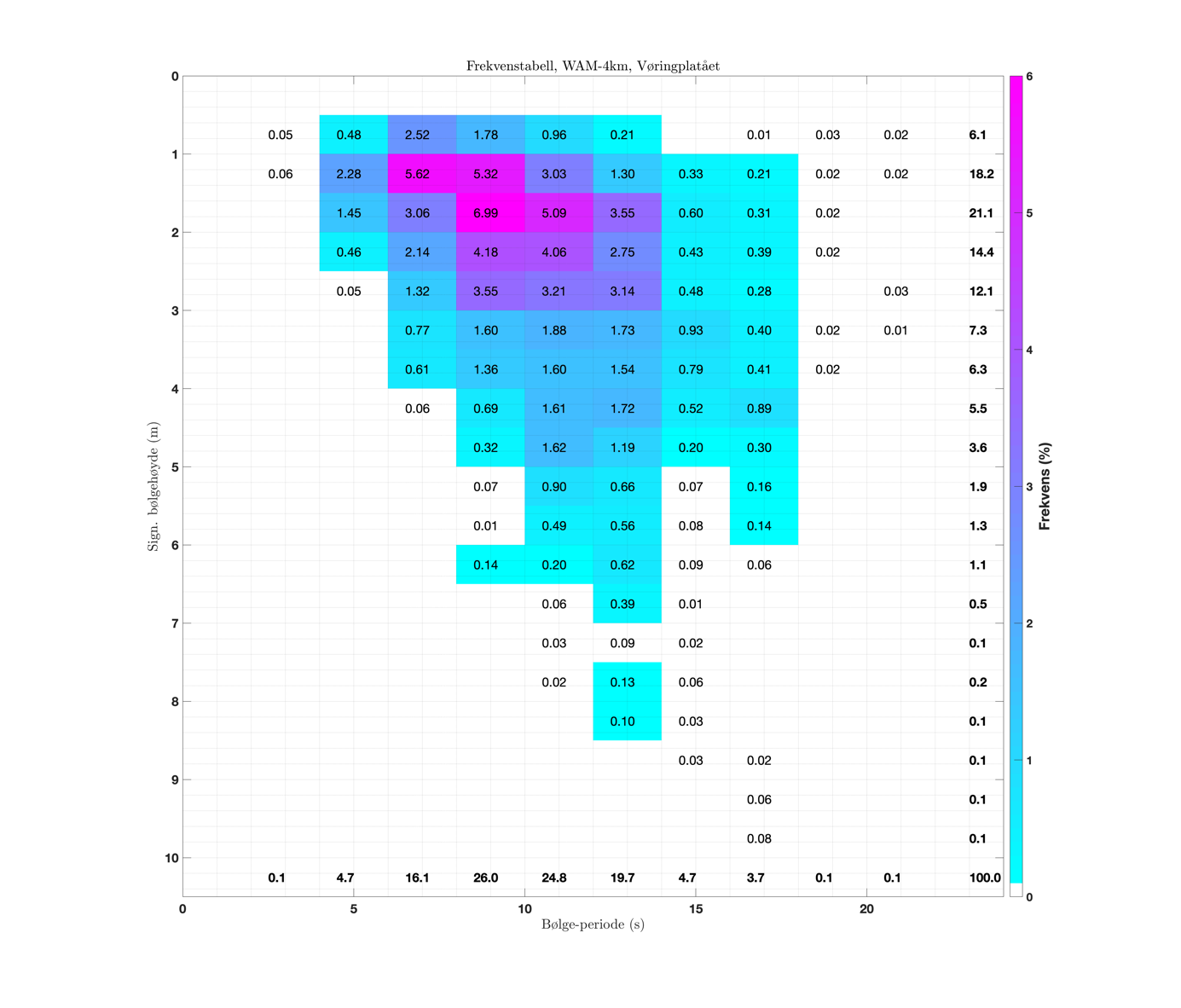
Fordelingen av bølgehøyder og bølgelengder er relativt jevn utenfor norskekysten med noe større bølger lenger vekk fra kysten og noe større bølger utenfor Stadt enn utenfor kysten av Nordland. En frekvenstabell er lagd ut fra signifikant bølgehøyde og bølgeperiode (Figur 3.5) for et punkt midt på Vøringplatået (angitt med sort prikk i Figur 3.2) hvor dybden er rundt 350 m. Basert på timesverdier for hele 2018 fra den operasjonelle bølgemodellen WAM-4km, er hver tilstand av signifikant bølgehøyde og bølgeperiode kategorisert i den todimensjonale frekvenstabellen hvor verdiene er angitt i %. Typisk for bølgetilstand er at man har vindsjøen som relativt lave bølger med kortere periode (kortere bølgelengde) og dønningene som de høyeste bølgene med lengre periode. Ut fra modellerte bølger midt på Vøringplatået, så ser man at de fleste bølgene har signifikant bølgehøyde mellom 1 og 3 m og med bølgeperiode på 6-14 s, sistnevnte tilsvarer en bølgelengde på rundt 50-300 m.
3.4 - Referanser
Hasselmann S, Hasselmann K, Bauer E, Janssen PAEM, Komen G, Bertotti L, Lionello P, Guillaume A, Cardone V, Greenwood J, Reistad M, Zambresky L & Ewing J (1988). The WAM model - a third generation ocean wave prediction model. Journal of Physical Oceanography, 18, 1775-1810.
Hvas M, Folkedal O, Oppedal F (2019) Havbasert oppdrett – hvor mye vannstrøm tåler laks og rensefisk? Fiskevelferd og grenseverdier. Rapport fra Havforskningen nr x.
4 - Kunnskapsgrunnlag om vandringsruter for postsmolt og laks
Kunnskapen om vandringsrutene for postsmolt fra norske lakseelver og ut til beiteområdene i havet er begrenset. Det samme er kunnskapen om hvordan større laks eventuelt benytter kystnære områder for beiting, og vandringsrutene til kjønnsmoden laks som returnerer til elvene for å gyte. Det er derfor også vanskelig å gjøre en vurdering av eventuell påvirkning fra arealer avsatt til offshore oppdrett på ville laksebestander. Eventuell spredning av sykdom og parasitter fra offshore anlegg vil kunne modelleres, men det er mer komplisert å fastslå om dette vil kunne påvirke ville laksebestander så lenge kunnskapen om hvor de oppholder seg og vandringsruter de følger er så begrenset. I det følgende presenterer vi kunnskapsgrunnlaget, og peker på hvordan mer detaljert og relevant kunnskap kan framskaffes.
I en samlet sårbarhetsvurdering av hvordan områder avsatt til offshore oppdrett vil kunne påvirke ville laksebestander er det også viktig å ta i betraktning den belastning postsmolten utsettes for på kysten i intensive oppdrettsområder. Postsmolt som vandrer gjennom slike områder vil kunne være svekket og ytterligere påvirkning fra områder med offshore oppdrett vil derfor gi relativt større utslag enn om påvirkning fra kyst var lav. Områder med høy samlet belastning kan blant annet identifiseres gjennom det pågående programmet for overvåkning av lakselus, og vurderingene som gjøres inn mot «trafikklyssystemet».
Dersom effekt fra havbruk til havs på vandrende laks skal kunne vurderes uten stor usikkerhet må kunnskapsgrunnlaget bedres. En risikoanalyse som kartlegger mulige negative konsekvenser for ville laksebestander bør gjennomføres.
4.1 - Utvandrende postsmolt
Kunnskapen om hvordan lakseunger vandrer ut av elvene som smolt og videre ut gjennom fjordene som postsmolt er relativt god. Men når de forlater kysten og vandrer videre ut i havet mot beiteområdene er kunnskapen mer begrenset. Den kunnskapen vi har i dag om vandring og fordeling av postsmolt i Norskehavet stammer i hovedsak fra pelagiske trålhal på forskningstokt. Enkelte av disse trålstasjonene med fangster av postsmolt i Norskehavet og Barentshavet er gjennomført i forbindelse med målrettede studier av laks i havet, men postsmolt tas også av og til som bifangst i pelagiske trålhal etter andre arter som sild og makrell.
I forbindelse med arbeidet med en vitenskapelig publikasjon som forventes innsendt i løpet av høsten 2019 har vi i samarbeid med kolleger ved andre institusjoner satt sammen en database med alle stedfestede registeringer av postsmolt i Norskehavet og nærliggende områder. Totalt inneholder denne databasen over 7000 individer fanget siden 1995. For de aller fleste av disse individene finnes det ingen informasjon om hvilket vassdrag de kommer fra, men det er fanget enkelte merkede individer hvor det har vært mulig å fastslå opprinnelse. For ca 4000 individer er det gjennomført genetiske analyser og individene er sammenliknet med en Europeisk database med genetiske profiler av mer enn 300 lakseelver. For disse individene er det mulig å bestemme regional tilhørighet, men, med noen få unntak, gir ikke de genetiske metodene som ble benyttet tilstrekkelig presisjon til å si hvilket vassdrag de kommer fra, eller kartlegge vandringsruter i detalj. Disse analysene ble gjennomført i løpet av EU-prosjektet SALSEA-Merge, og resultatene er nå under publisering i regi av NFR-prosjektet SeaSalar. Det arbeides også med utvidede genetiske analyser som vil kunne øke presisjonen i tilordningen av disse individene, og resultatene av dette arbeidet forventes å foreligge høsten 2020.
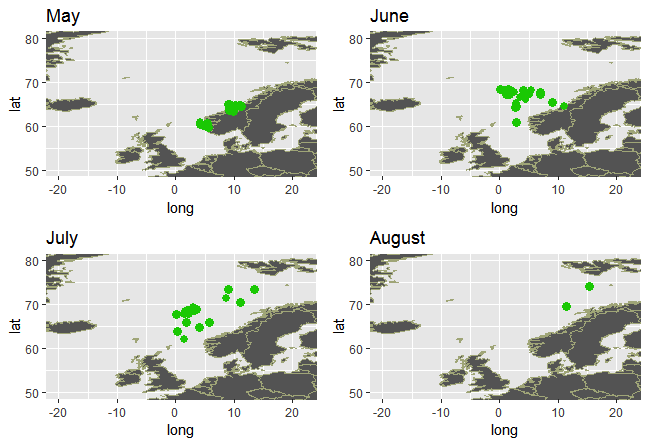
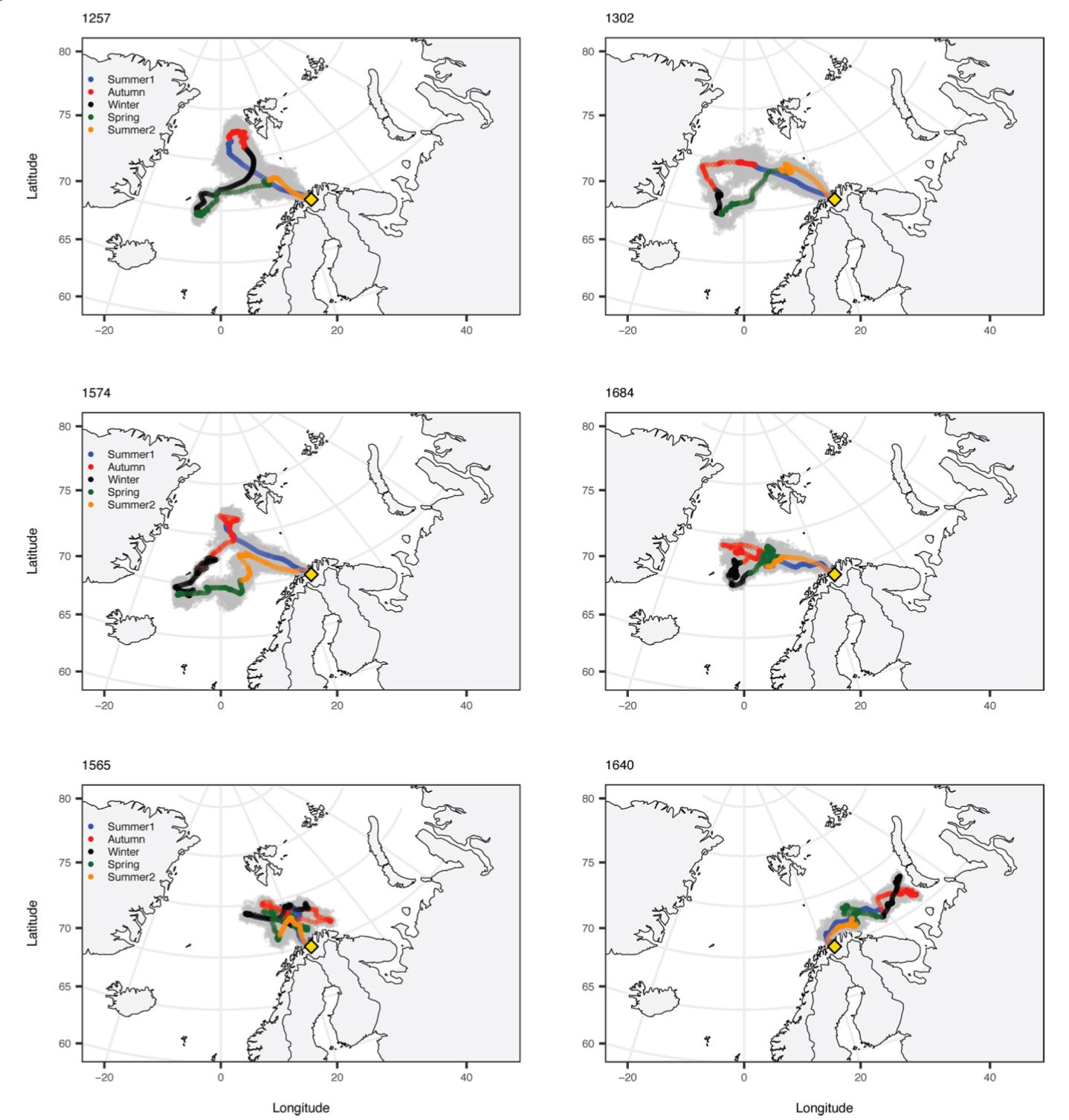
Disse fangstene, som stort sett er gjort langt til havs, gir liten informasjon om vandringsrutene fra kysten og ut i havområdene. Det er usikkert om postsmolt svømmer direkte ut til beiteområdene i havet, eller om den følger kyststrømmen nordover før den går ut i havet. Merkegjenfangster gir heller ikke et entydig svar på dette, og vandringsstrategier kan nok variere mellom regioner og vassdrag, og kanskje mellom år. Genetiske analyser, hvor postsmolten er tilordnet region, kan tyde på at en del av postsmolten krysser kyststrømmen og går rett ut i beiteområdene i Norskehavet. I Figur 4.1 er det vist et eksempel med tilordnet postsmolt fra områdene mellom Møre og Lofoten.
I en artikkel basert på data fra SALSEA-Merge prosjektet modellerte Kjell Arne Mork et al. (2012) hvordan postsmolt vandrer i Norskehavet. Vandringen kan påvirkes av både biologiske og fysiske faktorer, blant annet strøm og temperatur. I dette studiet viste de også hvordan vandringsmønster varierer mellom år som følge av variasjon i ulike parametere i modellen. I Figur 4.2 er modellert vandring til postsmolt fra en bestand i Irland og en bestand i Sør-Norge, i ulike år, vist.
4.2 - Tilbakevandrende laks
Kunnskapen om større fisk, i hovedsak fra merkeforsøk med ulike merketyper, er noe bedre. Det er gjennomført merkestudier med såkalte vinterstøinger, altså utgytt laks. Disse laksene ble merket når de forlot elven om våren for å returnere til beiteområdene i havet. Vinterstøinger fra tre elver i Norge ble merket; Neiden, Alta og Orkla. I tillegg pågår det nå merkeforsøk med vinterstøinger i Etneelva og Nidelva i Arendal. Noen av disse merkestudiene er gjennomført med DST-merker (Data Storage Tags) som registrerer hydrografiske data som temperatur og dyp, og i noen tilfeller lys. Etter gjenfangst av fisk med slike merker ved neste oppgang i elva, kan man ut fra lagrede data i merket rekonstruere vandringsruten i havet. Figur 4.3. er et eksempel på bruk av slike data fra fisk merket og gjenfanget i Altaelva (Strøm et al., 2018).
Laks fanget i sjølaksefisket i Nordland, Troms og Finnmark ble analysert genetisk og tilordnet til Norske og Russiske vassdrag i EU-prosjektet Kolarctic Salmon. Fangststed og tidspunkt for fangst ga informasjon om vandringsmønsteret på kysten for tilbakevandrende laks, og deler av disse resultatene er nylig publisert (Svenning et al., 2019). Resultatene fra dette studiet viste at laks fra ulike elver kom inn til kysten på ulik måte. Mens laks fra noen elver kom inn til kysten nær elva de var på vei til, kom laks fra andre elver inn til kysten over en lengre strekning.
4.3 - Hvordan fremskaffe mer relevant kunnskap om villaks og effekter av havbruk til havs?
Som forklart over har man generelt sett lite kunnskap om hvilken rute postsmolt tar fra elven og ut i havet. Hvordan denne vandringen eventuelt varierer med geografisk posisjon, utvandringstidspunkt, fysiske miljøforholdene eller byttedyrstilgang i havet har man ingen kunnskap om. For å forbedre kunnskapsgrunnlaget er det i første rekke behov for innsamling av mer laks fra havet. Dette kan gjøres med pelagisk tråling da dette er en godt egnet fangstmetode i åpne farvann for postsmolt. Det er spesielt behov for data fra perioden slutten av mai til begynnelsen av juli. Dedikerte tråltokt i juni over flere år vil kunne gi bedre innsikt i hvordan postsmolt vandrer ut fra elvene og hvor stor mellomårlig variasjon det er i vandringsmønsteret. En slik innsamling vil kreve systematisk tråling i overflaten og genetisk identifikasjon for postsmoltens opprinnelse. En slik biologisk innsamling vil også kunne gi innsikt i den individuelle tilstanden til postsmolten i områder der det kan være aktuelt med havbruk til havs. Dette inkluderer størrelse og kondisjon, samt påvirkning fra parasitter og infeksjoner.
Et alternativ til innsamling av postsmolt med pelagisk tråling er å merke postsmolt med små akustiske merker som kan registreres med lyttebøyer. Man merker da smolten før den forlater elven og en eventuell deteksjon vil gi presis informasjon om postsmolten opprinnelse. Rekkevidden til lyttebøyene er ca 200 m og merket fisk som passerer på større avstand vil ikke bli dedikert. Lyttebøyene vil ikke registrere umerket postsmolt eller si noe om fiskens tilstand, og vil således gi mindre informasjon enn det man oppnår ved pelagisk tråling. Fordelen med akustiske merker og lyttebøyer er lavere kostnadene enn for tråltokt, samt at datainnsamlingen kan foregå kontinuerlig over lang tid i viktige områder.
Utvikling av individbaserte vandringsmodeller for laks kan også gi ny kunnskap om laksens vandring i havet. En slik modell har allerede blitt benyttet for laks som vandrer ut fra Norskekysten (Mork et al., 2012, se Figur 4.2). Videre modellutvikling vil dra stor nytte av laks fanget i havet som beskrevet over. Uten bedre data for kalibrering og validering av modeller vil man ikke kunne gi presise prediksjoner med en vandringsmodell for laks.
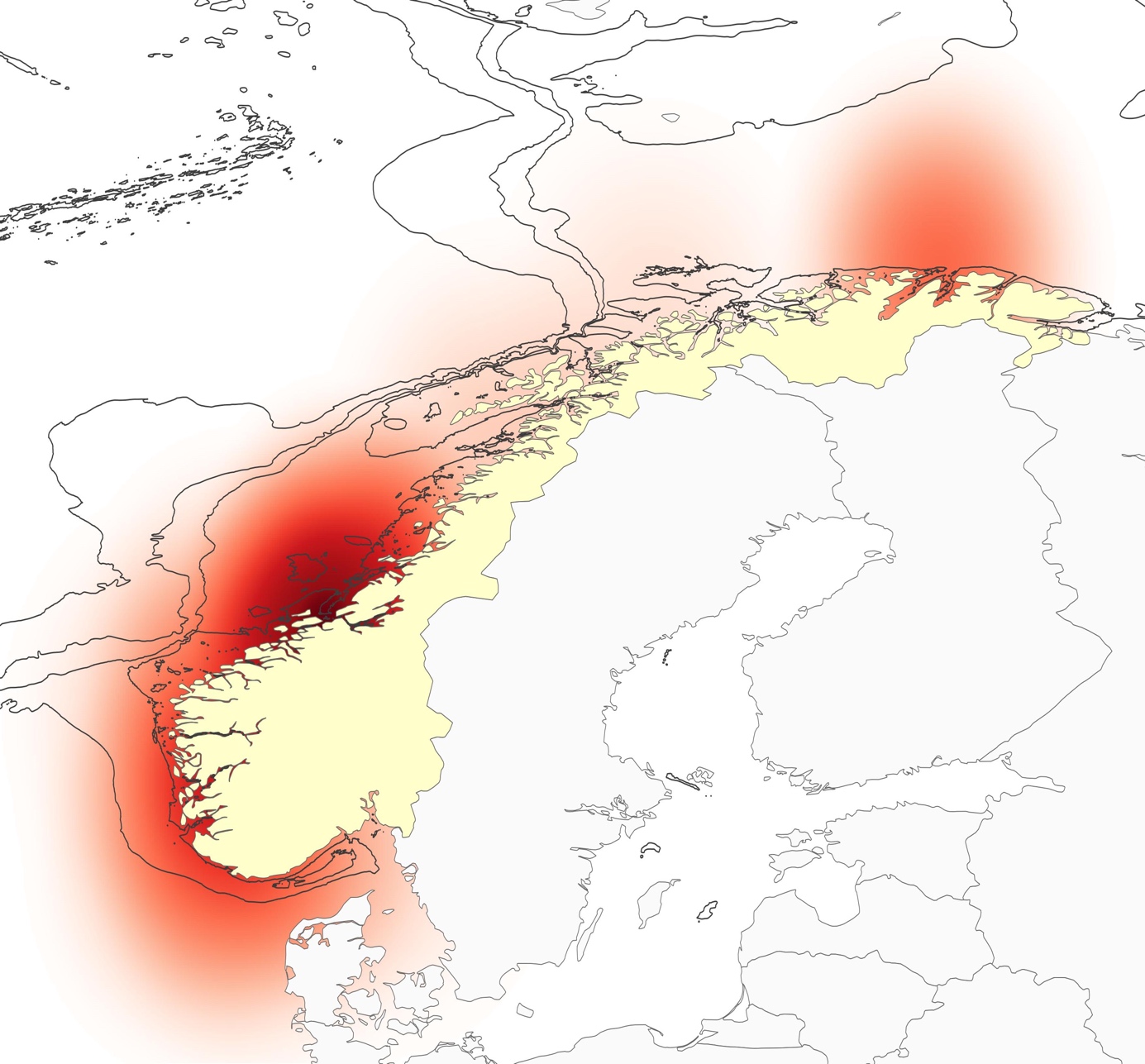
I mangel på konkret kunnskap om vandringsruter har vi forsøkt å illustrere hvordan smoltproduksjonen varierer langs kysten. I enkelte områder er det store vassdrag som sender mye smolt ut i havet, mens andre regioner har i hovedsak mindre vassdrag med begrenset produksjon. I kartet i Figur 4.4. har vi illustrert dette ved å aggregere teoretisk smoltproduksjon for produksjonsområdene langs kysten og plottet dette som et tetthetsplott. Teoretisk smoltproduksjon som er brukt som grunnlag for kartet er hentet fra en rapport utarbeidet av ekspertgruppen for vurdering av lakseluspåvirkning (www.regjeringen.no/contentassets/f7e4ce6a71c44fd5965f64e0913dae35/hovedrapport.pdf). I kartet kan man se hvordan de store elvene i Trondheimsfjorden sender mye smolt ut i Frohavet, og også hvordan området utenfor Tanafjorden vil ha relativt høy smolttetthet.
4.4 - Referanser
Mork K, Gilbey J, Hansen L, Jensen A, Jacobsen J, Holm M, Holst J, O ´ Maiole´ı´digh N, Vikebo F, McGinnity P, Melle W, Thomas K, Verspoor E, Wennevik V (2012) Modelling the migration of post-smolt Atlantic salmon (Salmo salar) in the Northeast Atlantic. ICES J Mar Sci. doi:10.1093/icesjms/fss108
Strøm, J. F., Thorstad, E. B., Hedger, R. D. & Rikardsen, A. H. (2018) Revealing the full ocean migration of individual Atlantic salmon. Anim. Biotelemetry6, 2, https://doi.org/10.1186/s40317-018-0146-2
Svenning, M.-A., M. Falkegård, E. Niemelä, J.-P. Vähä, V. Wennevik, M. Ozerov, S. Prusov, J. B. Dempson, M. Power, Fauchald, and D. Gomez-Uchida (2019) Coastal migration patterns of the four largest Barents Sea Atlantic salmon stocks inferred using genetic stock identification methods. ICES Journal of Marine Science
5 - Gyteområder og -vandring for fisk
Identifisering av områder egnet for havbruk til havs må ta hensyn til kjente gytefelt for kommersielt høstede bestander og nøkkelarter på kontinentalsokkelen langs kysten. Pågående forskning som NFR-prosjektet Salcod (294631/E40) ledet av Pål Arne Bjørn ved Havforskningsinstituttet undersøker hvordan kystnær havbruk påvirker habitatsbruk for lokal fisk. Lignende problemstillinger bør også undersøkes for større bestander på gytevandring. Tilsvarende peker seismikkrådgivning nettopp på forstyrrelser på gyteplasser og i gytevandringsruter som viktig grunner til å fraråde seismikkaktivitet i noen tidsrom og for enkelte områder. Det er avgjørende å skaffe til veie kunnskap om slik adferdsendring som kan finne sted ved havbruk til havs med anlegg og biomasse med oppdrettsfisk i langt større skala enn sammenlignet med det som er tilfelle kystnært og i fjordene.
5.1 - Gyteområder og -perioder for ulike fiskeslag
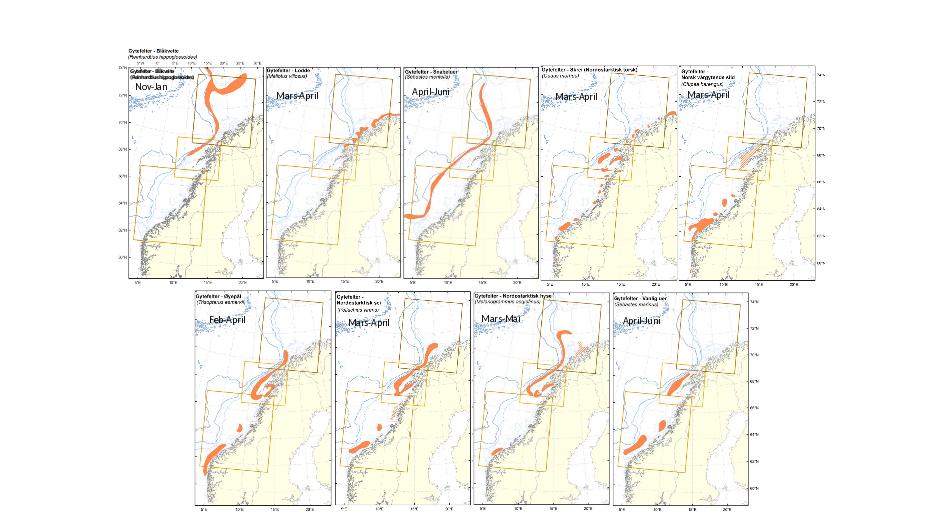
Figur 5.1 og 5.2 viser oversikt over gyteområder og -perioder for fiskebestander som gyter på Norskekysten som oppsummert i rapportene KILO (Sundby et al., 2013) og KINO (Sundby et al. 2017) samt tilgjengelig på http://www.imr.no/geodata/geodataHI.html. Gyting i det aktuelle området foregår som regel tett på kyst eller på sokkelkanten. For arter på sokkelen fra omtrent ved Møre og nordover foregår gyting typisk i mars – april for at de nyklekkede eggene skal treffe produksjonen av Calanus Finmarchicus på sokkelen som trigges av våroppblomstringen for planteplankton (Melle og Skjoldal, 1998; Vikebø et al., 2019). Dette sikrer fiskelarvene tilstrekkelig og egnede byttedyr i riktig størrelse til rett tid. Det er også arter som gyter tidligere (Blåkveite) eller lengre utover våren (uer) med tilhørende lengre eller kortere eggstadievarighet for å tilpasse seg lokale miljøforhold som temperatur. I Nordsjøen foregår gytingen over en lengre periode ettersom planktonoppblomstringen ikke er like konsentrert i tid.
Generelt er det liten overlapp mellom kjente gyteområder og de foreslåtte lokalitetene 1-13 fra Fiskeridirektoratet, selv om område 1 er relativt tett på kyststrømmen og gyteplasser for noen av artene som vist i Figur 5.1. Samtidig er område 10 umiddelbart nedstrøms for det største sildegyteområdet hvor det typisk finnes nyklekkede sildelarver i april og mai.
5.2 - Referanser
Melle, W., and Skjoldal, H.R. (1998). Reproduction and development of Calanus finmarchicus, C. glacialis and C. hyperboreus in the Barents Sea. Marine Ecology Progress Series, 169: 211–228.
Sundby et al. (2017) Dynamic Mapping of the North Sea Spawning – Report of the KINO Project. Fisken og Havet nr. 2.
Sundby et al. (2013) KunnskapsInnhenting Barentshavet-Lofoten-Vesterålen (KILO). Fisken og Havet nr. 3.
Vikebø FB, Strand KO and Sundby S (2019) Wind Intensity Is Key to Phytoplankton Spring Bloom Under Climate Change. Front. Mar. Sci. 6:518. doi: 10.3389/fmars.2019.00518
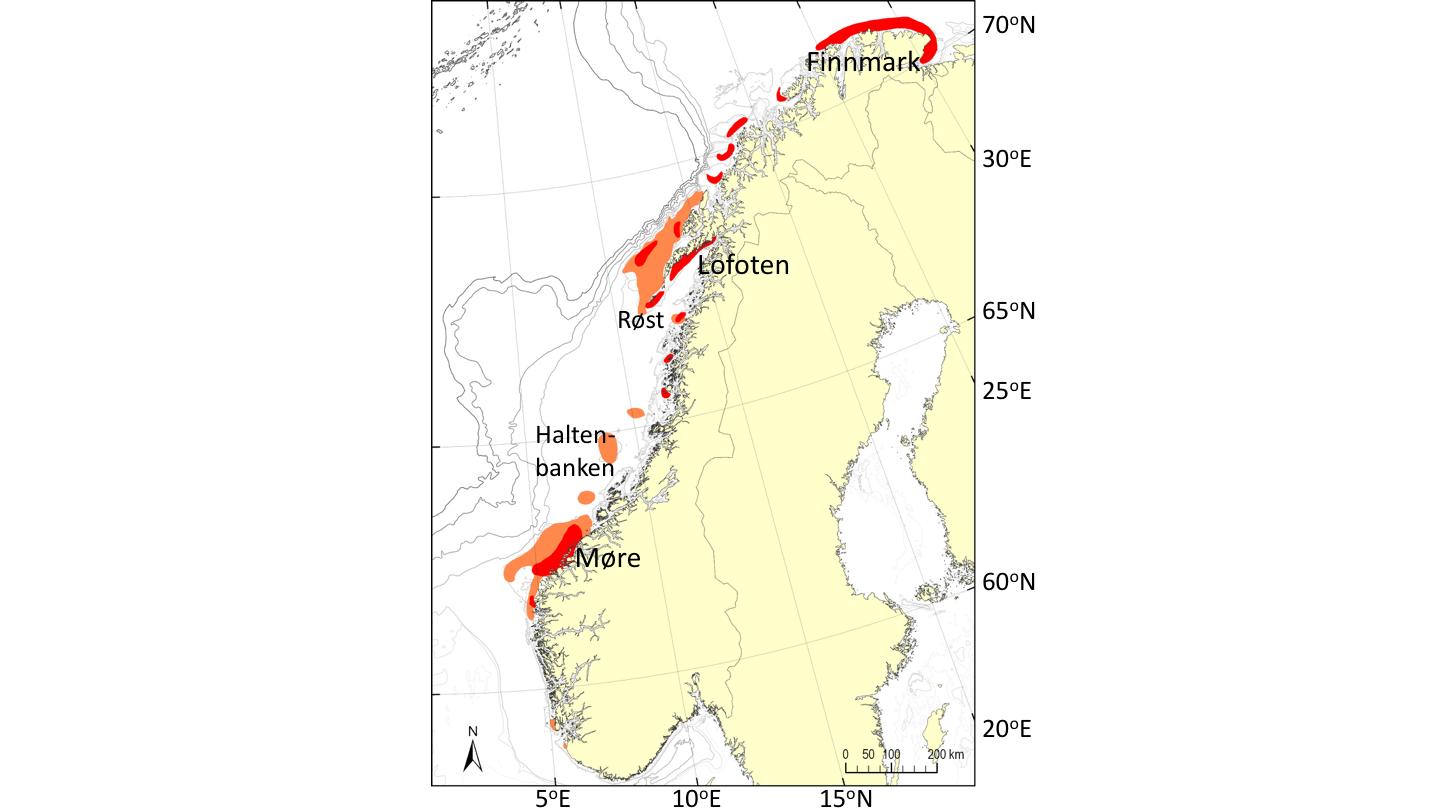
6 - Sårbare marine økosystem
Ved utpeking av lokaliteter egnet for havbruk til havs må man ta hensyn til eksisterende anbefalinger om områder som er definert som sårbare og/eller særlig verdifulle. Videre må man ta hensyn til kandidatområder for marint vern og områder som skal fungere som referanseområder for å vurdere endringer som følge av naturlig eller antropogen påvirkning. Områder tiltenkt akvakulturproduksjon på norsk sokkel bør kartlegges spesifikt for lokal artssammensetning og artsmangfold tidlig i prosessen, slik at man kan unngå å legge produksjonsanlegg over områder med sårbar og/eller funksjonelt viktige økosystem.
6.1 - Sentrale begreper
Sårbare marine økosystem består av arter og/eller samfunn av arter som er særlig sensitive for ytre, menneskeskapt påvirkning. Sårbarhet i seg selv er et kombinasjonsbegrep som beskriver hvor sensitiv en art er for en gitt påvirkning, moderert av sannsynligheten for at arten vil bli eksponert for den gitte påvirkningsfaktoren.
Hvor sensitiv en gitt art er, er en funksjon av to komponenter: 1) en arts motstandsdyktighet (i.e. resistens) mot endring i miljøet og 2) dens evne til å vende tilbake til dens opprinnelige naturtilstand etter endt påvirkning (i.e. resiliens, Holling, 1973). Resistens beskriver således en arts evne til å tåle eller tilpasse seg til forstyrrelser. Altså det å utsettes for en stressfaktor men forbli innenfor den opprinnelige naturtilstand. Resiliens beskriver en arts evne til å innhente seg og returnere til dens opprinnelige tilstand etter endt forstyrrelse. Det betyr at en art vil defineres som sensitiv når den er lett påvirkelig av ytre faktorer (har lav resistens) og/eller når det tar lang tid for arten å restituere seg (lav resiliens). For eksempel er fastsittende arter mer sensitive mot lokal forhøyet sedimentasjon enn mobile arter fordi de ikke kan flytte på seg. Arter med uregelmessig rekruttering og langsom vekst er sensitive for påvirkning fordi det kan ta lang tid for dem å etablere nye populasjoner eller økosystem hvis de forsvinner fra nåværende lokalitet.
En rekke ulike begreper blir brukt i forvaltningen i omtalen av sårbare marine økosystem. En oversikt over dem som blir brukt mest er presentert nedenfor. En rekke organer arbeider med forvaltning av sårbare marine økosystem. Relevante rødlister fra de forskjellige organene blir også beskrevet her.
6.2 - Relevante kilder for informasjon
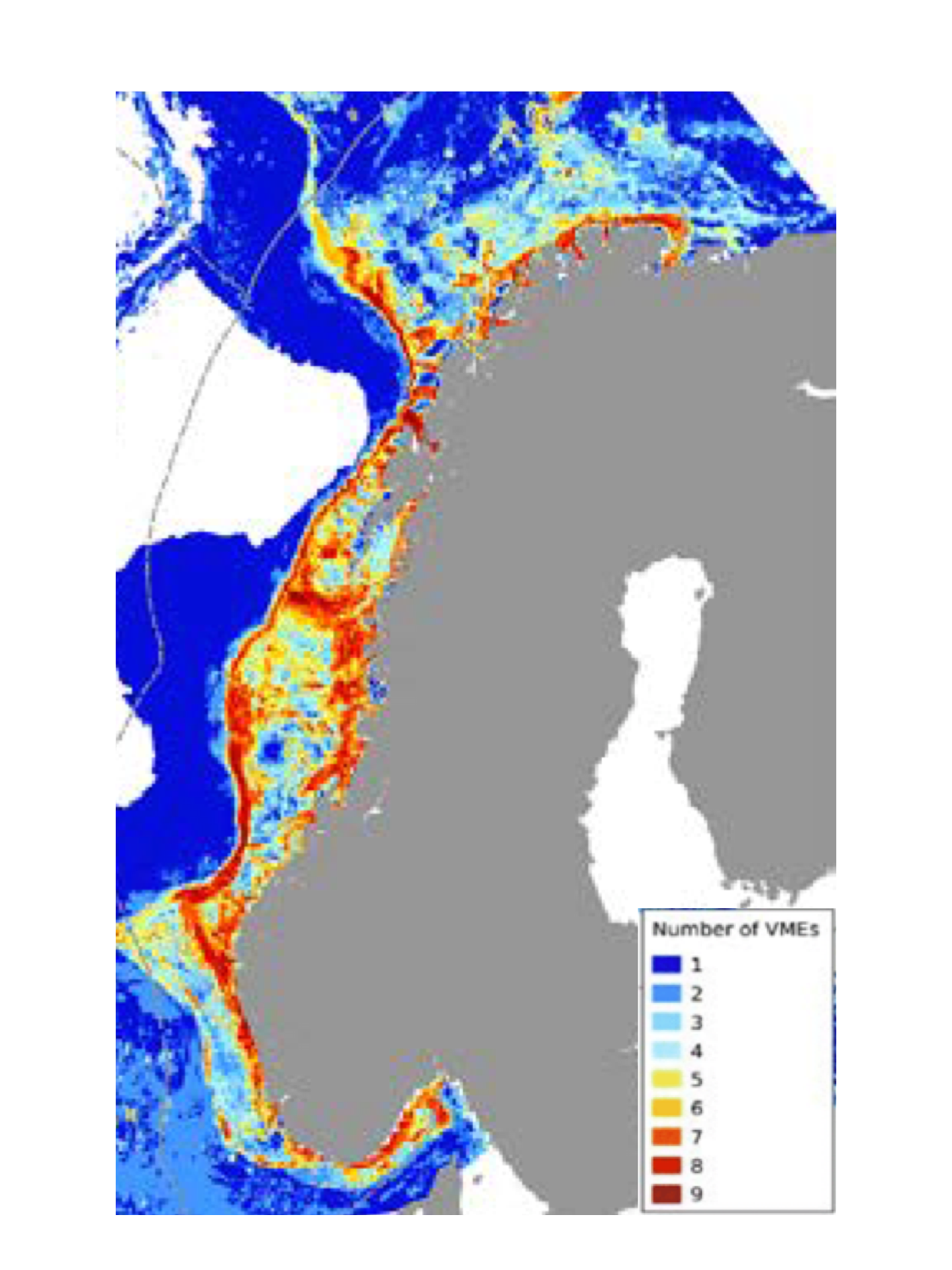
Vulnerable Marine Ecosystems (VMEs)
Begrepet Vulnerable Marine Ecosystems (VME) brukes hyppig i internasjonalt forvaltningsarbeid. Det stammer fra arbeid i FNs generalforsamling (UNGA) med å begrense bunnfiske med særlig negativ virkning på bunndyrssamfunn og/eller fiskepopulasjoner. VMEs defineres som en art eller dyregruppe som tilfredsstiller et eller flere av følgende kriterier: 1) unikhet, 2) økologisk funksjon/funksjonell signifikans, 3) ømfintlighet, 4) livshistorietrekk som bidrar til langsom restitusjon og 5) strukturell kompleksitet (FAO 2009, Artikkel 42). Flere undergrupper av koraller (deriblant steinkoraller, bløtkoraller, hornkoraller og hydrokoraller) inneholder mange arter som er følsomme og potensielt sårbare og som hyppig bygger opp sårbare marine økosystemer (VMEs). Samme gjelder de store svampene (Demospongia) som bygger «ostur» områder i Barentshavet og den norske kontinentalsokkel. Ostur er benevnelsen på ett begrenset område der store svamp utgjør mer enn 90 % av biomassen. FAOs liste inneholder også flere geologiske strukturer som kan være leveområder for VME arter. Eksempel på dette er varme havkilder, kalde gassoppkomster, undersjøiske raviner og renner.
Økologisk og biologisk signifikante områder
I konvensjonen for biologisk mangfold arbeider man med å identifisere økologisk og biologisk signifikante områder (EBSA). EBSA er spesielle områder i havet som fyller en viktig funksjon og som dermed har et stort bidrag til å opprettholde et friskt og livskraftig hav. Utvalg av EBSA baserer seg på flere kriterier som delvis overlapper med de i VMEs; 1) hvor unik eller sjelden ett område er, 2) om det har spesiell betydning for spesifikke livshistoriestadier, 3) om området er viktig for noen truede eller avtagende arter eller habitat, 4) hvor sensitive artene som bygger økosystemet er for ytre påvirkning og 5) om området støtter høy biologisk produksjon. Det arbeides for tiden med identifikasjon av EBSAs i nordøstlige deler av Atlanterhavet.
OSPAR
På OSPARs liste over truede og/eller avtagende arter og habitater i Region I (Nordøstlige Atlanteren) står flere arter og habitat som er relativt vanlig i norske farvann, dvs artene Arctica islandica, purpursnegl Nucella lapillus og flatøsters Osterea edulis samt habitatene dypvannskorallrev, korallskog, svampsamfunn, ruglbunn/mergelbunn/løsliggende kalkalger, oskjellfelt, kalkrørsormrev (Sabellaria spinulosa), samfunn av sjøfjær og gravende megabentos, ålegrasenger, undervannsfjell, varme havkilder, karbonatskorper og tidevannsflater (Agreement 2008-6). Gjennom ratifisering av avtalen har Norge forpliktet seg til å stanse videre reduksjon av populasjonene av disse arter/habitat i norske farvann (Bergen statement §24) (North-East Atlantic Environment Strategy (§1.2c).
Norsk rødliste for naturtyper
Norsk rødliste for naturtyper (Artsdatabanken) vurderer 5 naturtyper som forekommer i marine dypvannsområder som truede (Buhl-Mortensen et al., 2018). Korallrev, hardbunnskorallskog og svampspikelbunn i Barentshavet blir vurdert som nær truet. Bambuskorallskog i Nordsjøen og grisehalekorallskog blir vurdert som sterkt truet. I motsetning til OSPAR vurderer den norske rødlisten kaldt gassoppkomme (opphav til karbonatskorper) og varme havkilder som livskraftige naturtyper. Naturtypen samfunn av sjøfjær og gravende megabentos er ikke blitt vurdert.
På grunt vann blir 4 naturtyper vurdert som truet. Sukkertareskog blir vurdert som sterkt truet, stortareskog nær truet og fingertareskog og blåskjellbunn blir vurdert som sårbare naturtyper. I tillegg står ruglbunn på listen men er ikke vurdert på grunn av datamangel (Gundersen et al., 2018). I motsetning til OSPAR blir ålegrasenger i den norske rødlisten for naturtyper vurdert som en livskraftig naturtype, mens oskjellfelt og kalkrørsormrev ikke er blitt vurdert.
Utvalgte naturtyper
Miljødirektoratets Håndbok 19 definerer følgende utvalgte naturtyper i marint miljø: større tareskogforekomster, sterke tidevannsstrømmer, fjorder med naturlig lavt oksygeninnhold i bunnvannet, spesielt dype fjordområder, poller, litoralbassenger, israndavsetninger, bløtbunnsområder i strandsonen, korallforekomster, løstliggende kalkalger, ålegrasenger og andre undervannsenger og skjellsandforekomster. Det blir sagt at det skal tas særskilt hensyn til en utvalgt naturtype slik at man unngår en forringelse av naturtypens utbredelse og forekomstenes økologiske tilstand.
Forvaltningsplanene
Forvaltningsplanene for de norske havområdene har identifisert enkelte områder som særlig verdifulle og sårbare i miljø- og ressurssammenheng, såkalt særlig verdifulle områder (SVO). SVOer er hotspots for biodiversitet og produksjon, og her forventer man at mulige menneskeskapte skadevirkninger kan få langvarige eller irreversible konsekvenser. Områdene er selektert basert på informasjon om områdets økologiske funksjon og kriterier som beskriver hvor sårbart området er (som unikhet, uberørthet, representativitet og vitenskapelig verdi).
Utbredelse av sårbare bunnøkosystem på norsk kontinentalsokkel
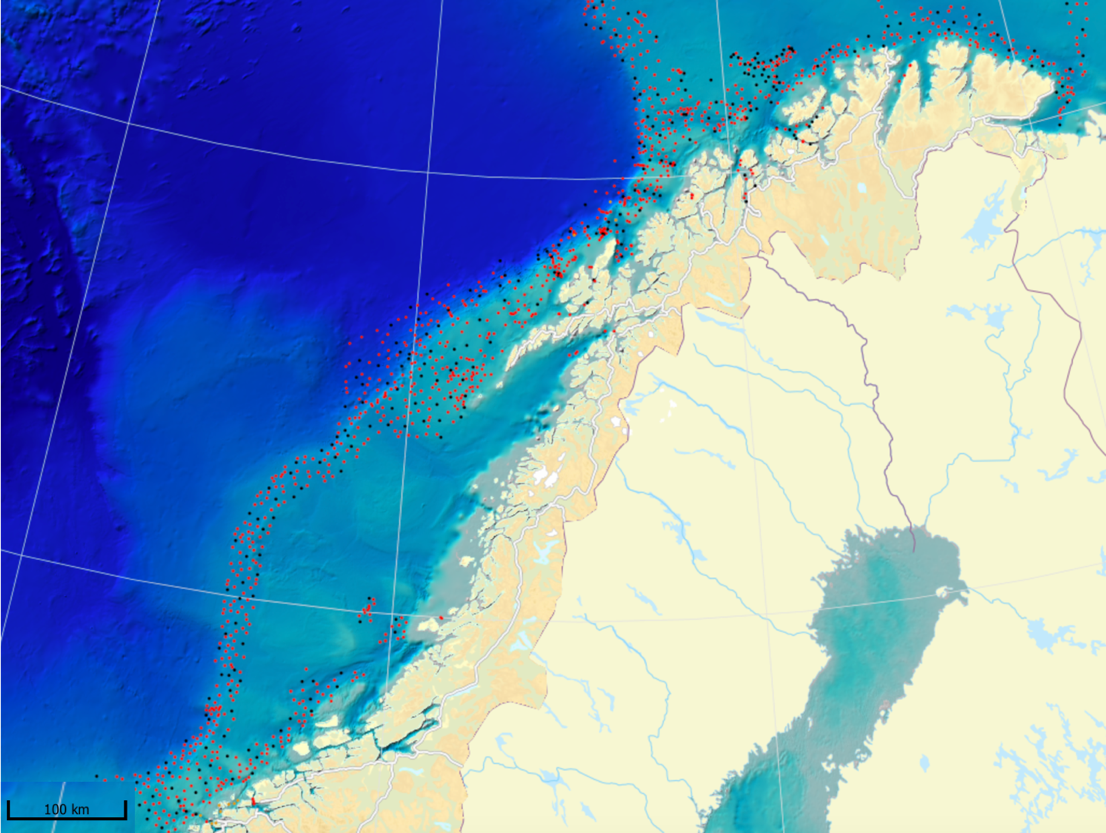
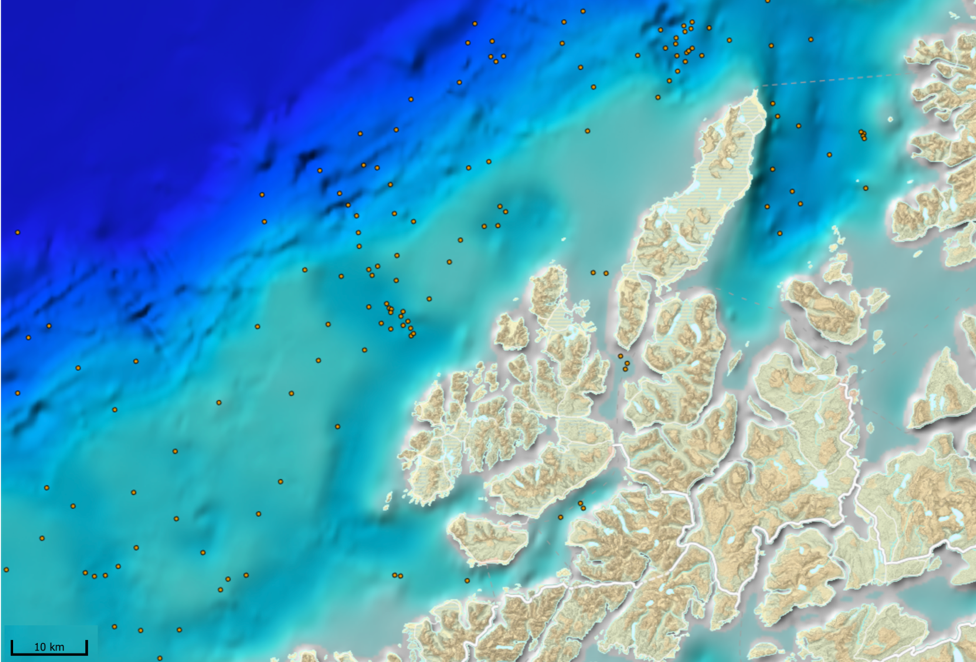
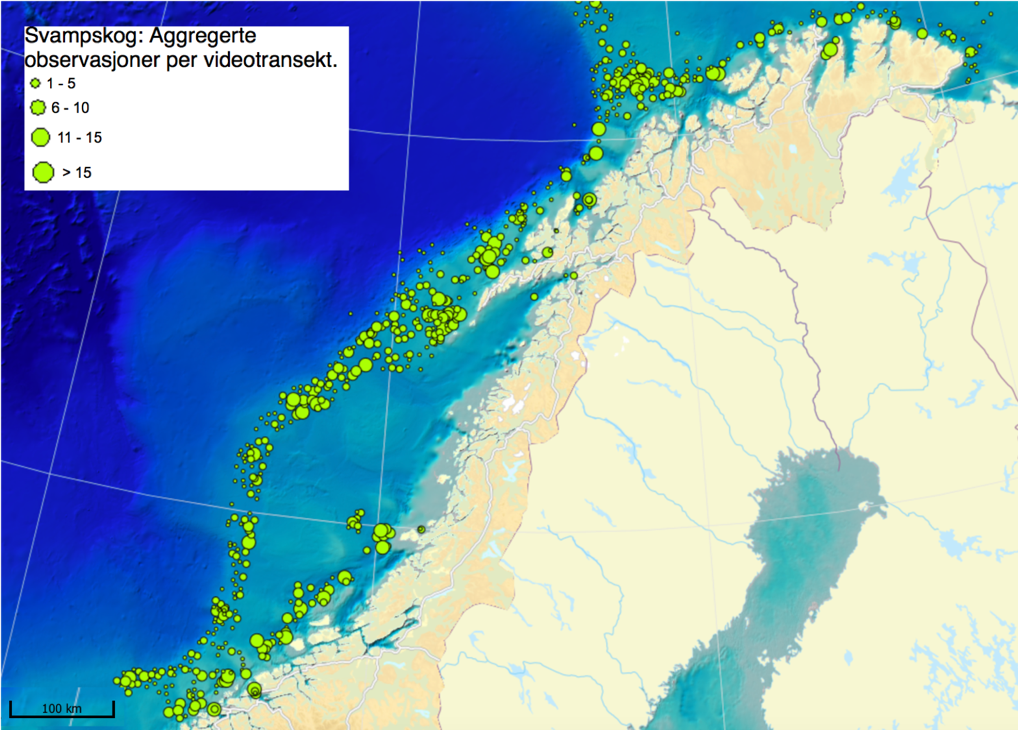
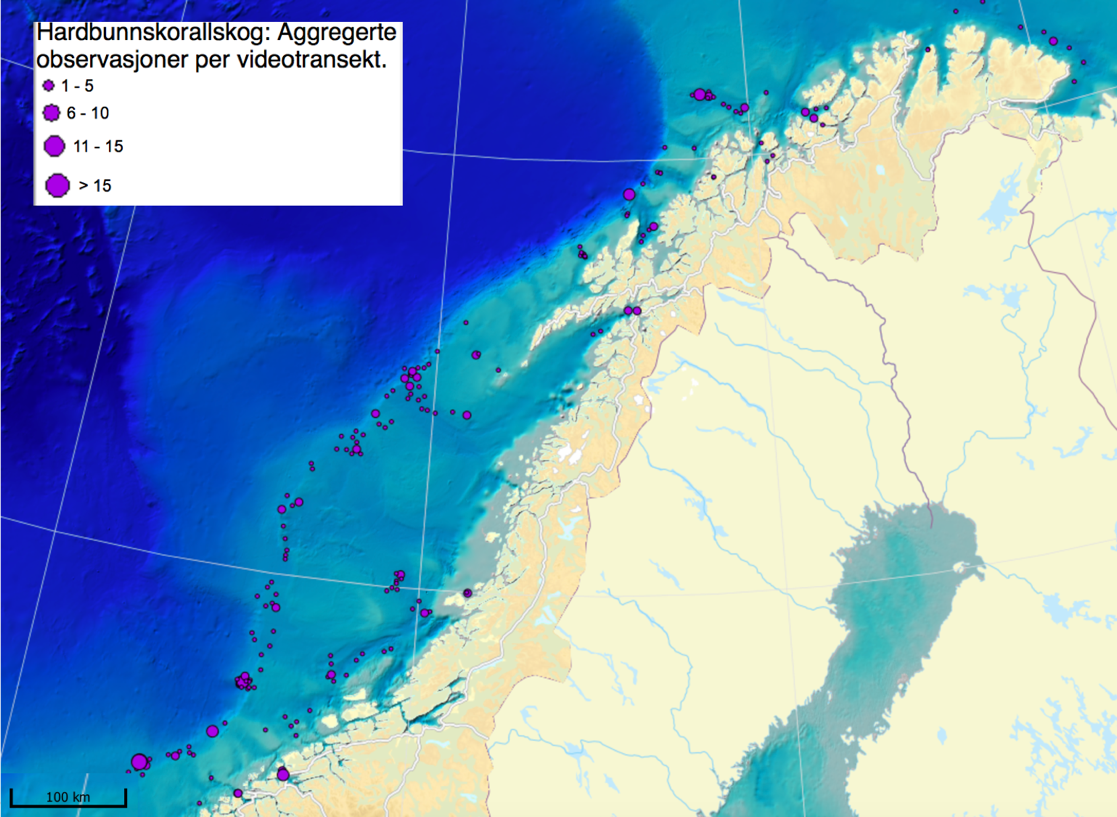
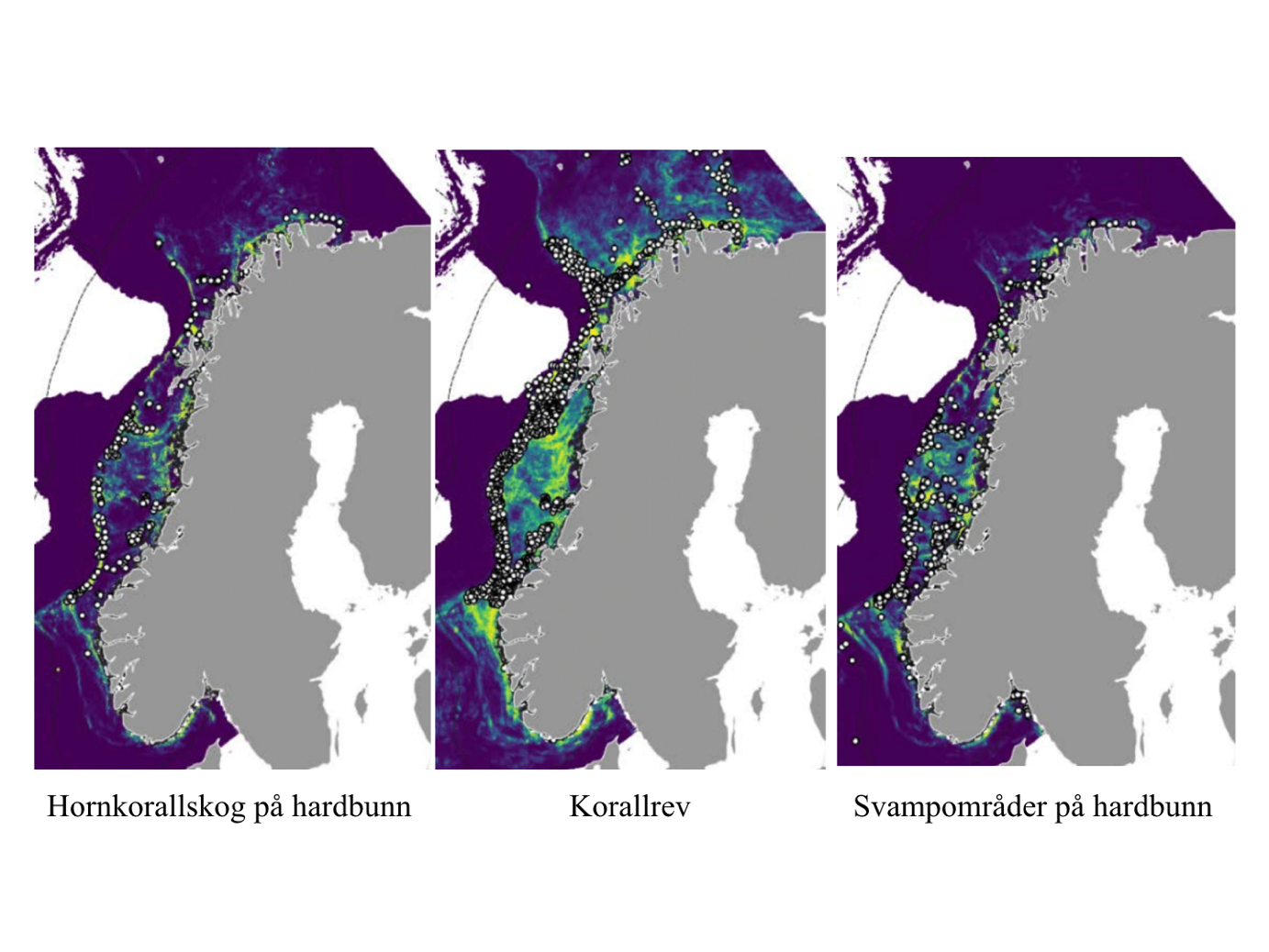
MAREANO har siden 2006 kartlagt dybde, bunnforhold, arter og naturtyper i norske havområder. Figur 6.1 viser en oversikt av stasjoner hvor informasjon om bunndyrssamfunnets struktur finnes, enten fra undervannsvideo, bomtrål, slede og/eller prøvetaking med grabb. Det er et meget omfattende arbeid som har gitt oss ny og viktig kunnskap om utbredelsen av marine bunndyr på Norsk sokkel. Til trass for et meget intensivt kartleggingsarbeid de siste 15 år er det imidlertid fortsatt store kunnskapshull og det er store områder som fremdeles er ukjente med hensyn til artssammensetning og artsmangfold (Figur 6.2). Noen områder vil bli dekket av fremtidig kartlegging i regi av Mareano, men det er ikke sannsynlig at den kartleggingen vil bli foretatt med et tettere stasjonsnett. Observerte forekomster av svamper (Figur 6.3.) og hornkoraller (Figur 6.4.) gitt begrenset arealdekning på sokkelen gir svært nyttig informasjon om forekomst og tetthet, men kan samtidig gi et skjevt bilde av utbredelse dersom man ikke er observant på hvilke områder som er undersøkt. Imidlertid gir observasjonene nyttig informasjon om egnede miljøforhold som gir grunnlag for å modellere utbredelse i de områder som enda ikke er undersøkt.
Mange nye områder med sårbar bunnfauna er blitt lokalisert gjennom Mareano. Men det er altså kun et lite fragment av det totale areal av norsk sokkel som er undersøkt. Parallelt pågår derfor allerede et utstrakt arbeid med å modellere og predikere hvor på Norsk sokkel det med høy sannsynlighet finnes sårbare habitat (Figur 6.5). Slike prediksjoner (Buhl-Mortensen et al., 2019) viser at store deler av den Norske sokkelen er gode leveområder for sårbare naturtyper, dvs hardbunnskorallskog (hornkoraller, blomkålkorall og hydrokoraller), bløtbunnskorallskog (hornkoraller og begerkorall), sjøfjærbunn, kaldtvannskorallrev, svampområder på hardbunn, svampområder på bløtbunn og dyphavssvampområder. Kart over områder hvor vi predikerer forekomst av sårbar bunnfauna kan bli brukt for å velge ut områder før nøyere granskning og visuell kartlegging.
6.3 - Økologisk funksjon
Den Norske kontinentalsokkelen er på mange måter karakterisert av sine tette forekomster av korallrev, korallskoger og svampsamfunn. Disse økosystemene er oaser av høy biodiversitet og produktivitet. I Norske korallrev alene er det blitt dokumentert over 1300 arter av evertebrater og fisk (Freiwald et al., 2002). Sammen spiller disse økosystemene en svært viktig rolle for karbonsyklusen på sokkelen. Nyere forskning antyder at korallrevene er ansvarlig for 30 % av all omsetning av organisk materiale ved bunnen til tross for at de dekker kun 1 % av arealet på Norsk sokkel (Cathalot et al., 2015). Korallrev fyller flere av kriteriene for å karakteriseres som sårbart økosystem, dvs økologisk funksjon/funksjonell signifikans, ømfintlighet, og livshistorietrekk som bidrar til langsom restitusjon og strukturell kompleksitet.
6.4 - Effekter av akvakultur på sårbare økosystem på dypt vann
Det er kjent fra en rekke laboratorieforsøk at steinkorallen Lophelia pertusa, som er den arten som bygger de store korallrevene i Norge, tåler sedimentering av uorganiske partikler relativt bra (Brooke et al., 2009; Larsson et al., 2013; Allers et al., 2013). De levende korallene kvitter seg med partikler som er sedimentert på polyppene ved å produsere et slimlag som den skaller av sammen med sedimentet. Kostnaden av denne forsvarsmekanismen er ikke målbar i korttidseksperimenter (1–3 måneder). Hvordan korallen reagerer på økt sedimentering av organiske partikler er mindre kjent, men man vet at koraller naturlig lever i områder med relativt lav tilgang på mat og derfor er tilpasset dette. Pilotstudier som Havforskningsinstituttet har gjennomført i felt rundt et anlegg i Langenuen (Kutti et al., 2015) viste høy overlevelse, men en gradvis reduksjon i vekst hos L. pertusa jo nærmere anleggene korallene hadde vokst. Nær anleggene var veksten og produksjon av nye polypper etter fem måneder halvparten av det som anses som normalt og som er blitt målt ved korallrevet Nakken ved Huglo i Langenuen. Erosjonen på det døde kalkskjelettet av bakterier, alger, foraminifere og svamp var imidlertid fordoblet. Det er tidligere vist at det er sonen innenfor 250 m fra et anlegg som har størst sannsynlighet for å bli kraftig påvirket av nedfall av organisk materiale. Større anlegg til havs kan ha langt større påvirkningsradius og kan estimeres i numeriske spredningsmodeller. Nye transplantasjonsstudier på Møre er blitt gjennomført og data er under opparbeiding. Vi forventer at ha resultatene klare på slutten av dette året (2019).
I korttidseksponeringsforsøk har man sett ved hjelp av målinger av svampers energiomsetning og biomarkører at rester fra fôret til oppdrettsfisk i de konsentrasjonene vi måler i nærheten av anleggene kan påvirke svampene negativt, men uten å føre til dødelighet. Nye transplantasjonsstudier er pågående for å bekrefte eventuelle langtidseffekter av sedimentasjon av organisk avfall fra anlegg på svamp. De vil bli plukket opp sommeren 2020, etter 2 år i sjøen.
Studier hvor vi testet 96 timers eksponering av dypvannskoraller, skjell og svamp for løste kobber-ioner i konsentrasjoner fra 0 til 1000 µg/L viser tydelig at de forskjellige artene har ulik toleranse for kobber. Mest sensitiv er øyekorall Lophelia pertusa med en LC50 på 30 µg/L. Til sammenligning har sjøtre Paragorgea arborea en LC50 på 80 µg/L. Dypvannskorallen Dentomuricea meteor, en vanlig art på Azorene, har en LC50 på 137 µg/L (Martins et al., 2018). Korallen har ingen mulighet for å beskytte de ytre lagene med celler mot forurensing løst i vann. Dypvannsskjellet Acesta excavata, som kan velge å stenge skallet når den eksponeres for uønskede omgivelser, hadde i våre eksperimenter en LC50 på 344 µg/L. LC50 er definert som den konsentrasjonen som er dødelig for 50 % av testorganismene. Det pågår et arbeid for å undersøke konsentrasjoner av løst og bundet kobber i området rundt anlegg.
Lophelia pertusa og Acesta excavata er tolerante mot korttidseksponering (1h) for hydrogenperoxid i de konsentrasjoner som så langt er blitt målt på 100 m dyp etter en avlusningsepisode. Skjellet lukker seg under eksponering og korallen drar inn polyppene sine. Ingen cellulær stressreaksjon ble observert etter eksponeringen (Øystein Gjelsvik pers. com.). Resultat fra eksperimentet vil bli publisert i løpet av 2020. Det finnes ingen annen kunnskap om hvor tolerante dypvannskoraller og -skjell er for kjemikalieutslipp. Det at man hyppig finner korallkolonier på oljeplattformer førte tidlig til den konklusjon at koraller ikke er veldig sensitive til kjemikalieutslipp som skjer i forbindelse med oljeproduksjon (Bell & Smith, 1999). Denne påstanden er blitt kritisert (Roberts et al., 2000) og til dels motbevist med dokumentasjon av permanente skader som følge av sedimentering av oljeholdig koagulanter fra Deepwater Horizon-utslippet (Fischer et al., 2014).
I Norsk rødliste for naturtyper 2018 (Artsdatabanken) klassifiseres korallrev og korallskog som nær truet. De er spesielt truet i områder med mye bunntråling og der det ikke er verneområder for korall. I Vestnorske fjorder, der det fiskes mest med line og garn, er oppdrettsaktivitet den største trusselen for korallrevene i dag. Hvis man flytter store oppdrettsanlegg ut på sokkelen vil akvakultur utgjøre en potensiell trussel også her. I tillegg vil sannsynligvis havforsuring, høyere sjøtemperaturer og lavere oksygeninnhold i bunnvann bli et problem for korallene. En kombinasjon av ulike påvirkningsfaktorer kan gjøre korallene ekstra sårbare i fremtiden. Grunnet lang levetid, langsom vekst og sensitivitet overfor fysiske forstyrrelser er korallrev klassifisert som sårbart habitat. Å reetablere et korallrev vil ta fra flere hundre til over tusen år.
6.5 - Effekter av akvakultur på sårbare økosystem på grunt vann – tareskog og mergelbunn
De offshore-anleggene som er planlagt i dag ligger alle i åpne fjorder og kan derfor påvirke tareskogforekomster og mergelbunn, som begge er klassifisert som sårbare økosystem. Det samme vil gjelde for eventuelt mobile anlegg som blir fraktet inn mot kysten til roligere omgivelser under høst- og vinter-månedene.
Tareplanter kan påvirkes på flere ulike måter av utslipp fra oppdrett. Teoretisk vil pulser av ekstra næringssalter i form av ammonium kunne øke tareplantenes vekst fordi de får tilgang på mer nitrogen. Økt nitrogen i form av ammonium kan også stimulerer vekst av hurtigvoksende opportunistiske påvekstarter (Worm & Sommer, 2000). Et dekke av slike arter på bladet, særlig i sommerhalvåret, kan redusere lys og næringstilgang for tareplantene og føre til lavere vekstrater i vinterhalvåret. Forsøk gjennomført av Havforskningsinstituttet viste ingen effekt på hverken vekst eller påvekst på sukkertare satt ut nær anlegg i forhold til tare på kontrollstasjoner (Haugland, 2019). Derimot er sukkertare veldig sensitiv til eksponering for hydrogenperoxid. En behandlingsdose tilsvarende den som blir tilsatt i fiskemerden er dødelig for unge sukkertare-planter. Ved eksponering for 5 % av behandlingsdosen er dødeligheten på 50 % (Haugland et al., 2018). Studier av tareskogssamfunn nær anlegg på eksponert kyst viste at disse samfunnene i liten grad ble påvirket av utslipp av næringssalt og organiske partikler fra anlegg på eksponert kyst (Taraldset Haugland, 2019).
Løstliggende kalkalger finnes naturlig i strømrike områder der det er mindre sannsynlighet for å bli begravet i sedimenter. Kalkalger er sensitive for sedimentering, og dersom de blir dekket av et lag med finkornet sediment hindrer dette gassutvekslingen i cellene. Sediment som inneholder hydrogensulfid (H2S), som gjerne dannes ved oppdrettsanlegg, har vist seg å være fatalt for kalkalger, og 14 dagers dekke av slikt sediment førte til 100 % dødelighet (Wilson et al., 2004). Utslipp av organisk materiale påvirker også dyresamfunnene i kalkalgehabitatet ved at de fleste krepsdyr forsvinner og blir erstattet med opportunistiske børstemark (Sanz-Lazaro et al., 2011; Hall-Spencer, 2006). Eksperiment på havforskningsinstituttet våren 2019 viste at økt sedimentasjon av organiske partikler leder til stopp i veksten hos kalkalger. Uorganiske partikler har ikke den samme effekten. Hvite pigmentfrie prikker ble observert på kalkalger som ble eksponert for organiske partikler i en måned (Legrand pers. com).
Både sukkertare og stortare etablerer seg raskt igjen hvis de blir utsatt for negative miljøpåvirkninger. Mens det tar 4–8 år før stortareskog er fullstendig rehabilitert, vokser sukkertare (som er en hurtigvoksende treårig plante) raskt tilbake. Løstliggende kalkalger/mergel er meget saktevoksende (0.5–1.5 mm årlig, Blake & Maggs, 2003) og det vil ta lang tid å regenerere en mergelbunn hvis den utsettes for skader.
6.6 - Identifisere/beskrive kunnskapshull
For mobile anlegg som veksler mellom en væreksponert sommerlokalitet og en beskyttet lokalitet om vinteren bør biomassen i anlegget tilpasses den lokalitet med lavest kapasitet til å assimilere organisk materiale.
Mareano-programmet har så langt dekket utvalgte deler av disse havområdene, men det kan være nødvendig å rette slik kartlegging mer spesifikt inn mot områder som av andre grunner er vurdert som godt egnet for havgående havbruksanlegg. Det kan også være aktuelt å samarbeide med Mareano-programmet for å videreutvikle gode kartløsninger eller andre dataprodukter som er spesielt godt egnet for bruk til vurdering av lokalisering av havgående havbruksanlegg.
Utover dette gir de oppdaterte forvaltningsplanene for de ulike norske havområdene viktig sammenstilling av tilgjengelige data og vurderinger (http://www.miljodirektoratet.no/no/Havforum/Forside/).
Det er behov for å vurdere samlet påvirkning på kyst ettersom næringer utvikles og intensiveres. Spesielt fordi kystøkosystemer også er i endring som følge av klimaendringer. IPCCs spesialrapport om hav og is (IPCC, 2019) peker blant annet på økende temperaturer og forsuring, samt redusert planteplanktonproduksjon og oksygennivå. Disse endringene kommer samtidig med økt menneskelig påvirkning som fører til habitatsforringelse gjennom eksempelvis organisk belastning og fysiske inngrep. Det er derfor avgjørende at fremtidige utredninger og tilhørende forskning vurderer effekter av multiple stressorer og ikke kun belastning fra enkeltstressorer.
6.7 - Referanser
Allers, E., Abed, R.M.M., Wehrmann, L.M., Wang, T., Larsson, A.I., Purser, A., de Beer, D. (2013) Resistance of Lophelia pertusa to coverage by sediment and petroleum drill cuttings. Marine Pollution Bulletin.
Bell N, Smith J (1999) Coral growing on North Sea oil rigs. Nature 402:601
Blake, C. , and Maggs, C. A. (2003). Comparative growth rates and internal banding periodicity of maerl species (Corallinales, Rhodophyta) from northern Europe. Phycologia 42, 606–612.
Brooke, S.D., Holmes, M.W., Young, C.M. 2009. Sediment tolerance of two different morphotypes of the deep-sea coral Lophelia pertusa from the Gulf of Mexico. Marine Ecology Progress Series. 390:137-144.
Buhl-Mortensen, P., Bakken, T., Oug, E. og Rapp H. T. (2018). Marint dypvann. Norsk rødliste for naturtyper 2018. Artsdatabanken. Hentet (22.08.2018) fra www.artsdatabanken.no/Pages/260223
Buhl-Mortensen et al. (2019) Vulnerable marine ecosystems (VME) Coral and sponge VMEs in Arctic and sub-Arctic waters - Distribution and threats
Cathalot, C. & Oevelen, D. & Cox, Tom & Kutti, Tina & Lavaleye, Marc & Duineveld, Gerard & Meysman, Filip. (2015). Cold-water coral reefs and adjacent sponge grounds: Hotspots of benthic respiration and organic carbon cycling in the deep sea. Frontiers in Marine Science. 2. 10.3389/fmars.2015.00037.
Fischer, B.B., Kwiatkowski, M., Ackermann, M., Krismer, J., Roffler, S., Suter, M.J.F. et al. (2014). Phenotypic plasticity influences the ecoevolutionary dynamics of a predator-prey system. Ecology, 95, 3080– 3092.
Freiwald, A., Hühnerbach, V., Lindberg, B., Wilson, J.B., Campbell, J. (2002) The Sula reef complex, Norwegian shelf. Facies 47:179-200.
Gundersen, H., Bekkby, T., Oug, E., Norderhaug, K. M., Fredriksen, S. og Rinde, E. (2018). Marint gruntvann. Norsk rødliste for naturtyper 2018. Artsdatabanken. Hentet (dato) https://www.artsdatabanken.no/Pages/259183/Marint_gruntvann
Hall-Spencer, J. , White, N. , Gillespie, E. , Gillham, K. , and Foggo, A. (2006). Impact of fish farms on maerl beds in strongly tidal areas. Marine Ecology Progress Series 326, 1–9.
Haugland, Barbro (2019) Effecs of fish-farm effluents on kelp-forest ecosystems - kelp performance, associated species, and habitats. PhD Thesis. University of Oslo. 2019.
Haugland, Barbro & Rastrick, Samuel & Agnalt, Ann-Lisbeth & Husa, Vivian & Kutti, Tina & Samuelsen, Ole. (2018). Mortality and reduced photosynthetic performance in sugar kelp Saccharina latissima caused by the salmon-lice therapeutant hydrogen peroxide. Aquaculture Environment Interactions. 11. 10.3354/aei00292.
Holling, C. S. (1973) Resilience and stability of ecological systems. Annual Review of Ecological Systems, 4, 1-23.
IPCC, 2019: Summary for Policymakers. In: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate [H.- O. Pörtner, D.C. Roberts, V. Masson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, M. Nicolai, A. Okem, J. Petzold, B. Rama, N. Weyer (eds.)].
Kutti T, Bannister RJ, Fosså JH, Krogness CM, Tjensvoll I, Søvik G (2015) Metabolic responses of the deep-water sponge Geodia barretti to suspended bottom sediment, simulated mine tailings and drill cuttings. Journal of Experimental Marine Biology and Ecology 473: 64-72
Larsson, A.I., van Oevelen, D., Purser, A., Thomsen, L. 2013. Tolerance to long-term exposure to suspended benthic sediments and drill cuttings in the cold-water coral Lophelia pertusa. Marine Pollution Bulletin 70: 176-188.
Martins I, Godinho A, Goulart J, Carreiro-Silva M (2018) Assessment of Cu sub-lethal toxicity (LC50) in the cold-water gorgonian Dentomuricea meteor under a deep-sea mining activity scenario. Environmental Pollution 240: 903-907.
Sanz-Lázaro, Carlos & Marín, Arnaldo. (2011). Diversity Patterns of Benthic Macrofauna Caused by Marine Fish Farming. Diversity. 3. 176-199. 10.3390/d3020176.
Worm B, Sommer U (2000) Rapid direct and indirect effects of a single nutrient pulse in a seaweed-epiphyte-grazer system. Marine Ecology Progress Series 202:282-288
7 - Sjøpattedyrvandring
7.1 - Datatilgjengelighet
Rapporten Seamounts in the OSPAR Maritime Area (Kutti et al., 2019) som er utarbeidet av Havforskningsinstituttet på oppdrag fra Miljødirektoratet, og som er en del av Norges bidrag til OSPAR, oppsummerer observasjoner av sjøpattedyr i Nordøstlige deler av Nord-Atlanteren (Figur 7.1). Basert på disse observasjonene er ansamlinger langs Norskekysten konsentrert nord og nordvest for Lofoten og videre nordøstover til Barentshavsåpningen. I tillegg er det større ansamlinger mellom Stadt og Haltenbanken og videre vestover ut i åpent hav. Imidlertid fremheves det i rapporten at det er en betydelig underestimering av forekomster ut fra kyst som følge av manglende kartlegging. I tillegg stammer mesteparten av data fra Norske farvann i denne figuren fra ‘WorldWhaling Database: Individual Whale Catches, North Atlantic’, for perioden 1901-1971. Det vil være nødvendig å oppdatere denne oversikten med nyere data for å få representativt bilde på utbredelsen for dagens situasjon.
Ettersom vandringsruter og adferd kan tenkes å påvirkes av store havbruksinstallasjoner til havs – større enn de tettheter vi ser ved kyst – bør kartleggingen av habitatbruk for kjente sjøpattedyr langs Norskekysten oppskaleres. Dette kan gjøres med satellittsendere som HI har gjort for Knølhval (Figur 7.2), delvis i samarbeid med Universitet i Tromsø. Disse dataene må oppfattes som eksempler på vandringer ved senvinter og tidlig vår da et flertall av individene ser ut til å følge gytevandringene til Norsk Vårgytende (NVG) sild.
Databasen (http://www.imr.no/geodata/geodataHI.html) gir tilgang til utbredelseskart for blant annet sjøpattedyr. Under (https://www.imr.no/forskning/forskningsdata/karttjenester/nb-no) kan man laste ned underlagsdataene for bruk i GIS-verktøy. Figur 7.3 sammenstiller disse dataene i et rutenett, der farge-intensiteten indikerer antall sjøpattedyrarter som kan påtreffes innenfor hver rute. De arter som er tatt med i denne sammenstilling er Finnhval, Grindhval, Grønlandsel, Havert, Hvalross, Hvithval, Klappmyss, Knølhval, Narhval, Nebbhval, Nise, Ringsel, Spekkhogger, Spermhval, Kvitnos, Kvitskjeving, Steinkobbe, Storkobbe og Vågehval.
7.2 - 7.2 - Kystsel
Kystsel
To arter av sel påtreffes året rundt langs Norskekysten. Antall steinkobbe er estimert til ~10,000 dyr (2011-2016), mens havertpopulasjonen er estimert til 8,700 dyr (2006-2011). Nye tellinger av havert i området Sør-Trøndelag til Lofoten i september–oktober 2014 og 2015 viste imidlertid en betydelig nedgang i ungeproduksjonen. Antall unger som ble født i dette området i 2014–2015 var kun ca. 40 % sammenlignet med forrige telling i 2007–2008. Grunnen til den kraftige nedgangen antas å være økt bifangst, mest sannsynlig i garn som brukes i fiske etter breiflabb. Figur 7.4 viser liggeplasser til de to artene av sel som oppholder seg langs norskekysten; steinkobbe og havert. Steinkobbe er forholdsvis jevnt spredt langs kysten, med særlig store kolonier i Sør-Trøndelag (Frøya-området), Nordland (Myken), Vesterålen (Stø og Nordmela) samt i Troms (Måsvær). Langs Mørekysten finnes også mange mindre, men tett beliggende, lokaliteter. De største forekomstene av havert påtreffes langs kysten fra Sør-Trøndelag til Lofoten. Flere av de store forekomstene i Trøndelag og Nordland ligger forholdsvis nær delområdene 1, 3, 5, 10 og 11 som Fiskeridirektoratet har identifisert som særlig aktuelle for havbasert akvakultur (jfr. Fig 2.4).
På tross av forholdsvis gode data på hårfellings- og kastelokaliteter på land, er det store usikkerheter knyttet til utbredelse og vandringer til havs. Dette begrenser sterkt vår evne til å si noe om i hvor stor utstrekning sel vil vandre ut til de havområder som Fiskeridirektoratet har identifisert som særlig godtegnet til havbasert oppdrettsvirksomhet. Disse usikkerhetene er fordi det er foretatt svært få og til dels småskala merkeforsøk i Norge. På steinkobbe har slike forsøk i hovedsak blitt utført i Finnmark (Porsangerfjord), Vesterålen (Stø) samt et mindre forsøk langs Telemarkskysten. Disse forsøkene viser imidlertid at steinkobbene holder seg forholdsvis nært kysten, og ikke foretar langere vandringer fra sine landbaserte kjerneområder. Det er derfor lite sannsynlig at steinkobbe vil komme i kontakt med havbaserte oppdrettsanlegg i de angitte havområdene i noen særlig grad. Når det gjelder havert er merkeforsøk i norske farvann begrenset til fem haverter merket på Froan i 1991-1992, og fem merket ved Myken i 2012. Disse begrensede forsøk tyder på at haverten foretar lengre vandringer langs kysten, og også bruker havområder utenfor kystsonen. Det er derfor mer sannsynlig at havert kan komme i kontakt med anlegg innenfor de delområdene som er identifisert av Fiskeridirektoratet.
Disse antakelser understøttes til viss grad av resultater fra et betydelig mer omfattende merkeforsøk langs kysten av Storbritannia. Figur 7.5 viser utbredelseskart til både steinkobbe og havert. Disse er basert på habitatmodellering fra vandringer til 280 steinkobber og 112 havert med GPS-merker. Resultatene fra habitatmodelleringen er deretter satt sammen med data fra tellinger på landbaserte lokaliteter, for å gi et sannsynlig bilde av utbredelsen til havs på populasjonsnivå. Tilsvarende habitatmodellering basert på GPS-merket sel fra flere kolonier langs Norskekysten ville innebære en betydelig forbedring av kunnskapsgrunnlaget for å gi råd, ikke bare når det gjelder denne utredningen, men også mer generelt.
Andre selarter
Arktiske selarter forekommer i mindre utstrekning i havområdene langs Norskekysten. En art som trolig kan komme til å overlappe med noen av de foreslåtte havområdene er klappmyss, som ofte foretar regelmessige beitevandringer fra pakk-isen langs Grønlandskysten til beiteområder på sokkelen rundt Norskehavet. Satellittmerking fra perioden ~1995-2008 viser at flere av disse områdene ligger langsnorskekysten (Fig 7.6). I den utstrekning klappmyssen bruker Norsk sokkel som beiteområder vil de kunne vandre gjennom område nr 12, og kan også forekomme innenfor de mindre områdene langs Trøndelag- og Nordlandskysten.
Grønlandsselen er mer knyttet til Arktiske farvann, men kan komme til å overlappe med foreslåtte havområder i Barentshavet (område 6-9).
7.3 - 7.3 - Referanser
Kutti et al. (2019) Seamounts in the OSPAR MaritimeArea. Rapport fra Havforskningen nr xx.
Russell, D. J. F., Jones, E.L., and Morris, C. D. (2017) Updated Seal Usage Maps: The Estimated at-sea Distribution of Grey and Harbour Seals. Scottish Marine and Freshwater Science Report Vol 8 No 25, 25pp.
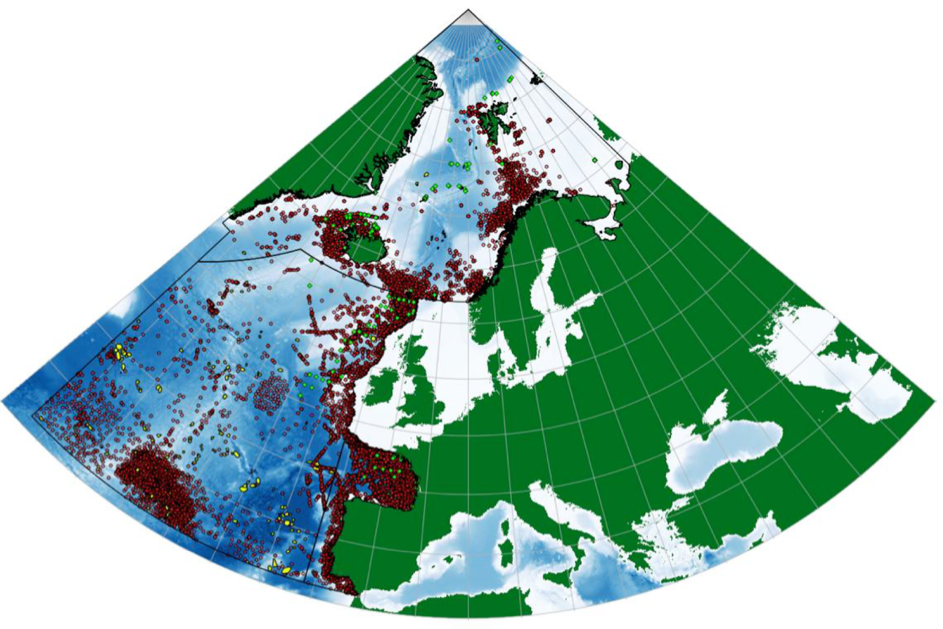
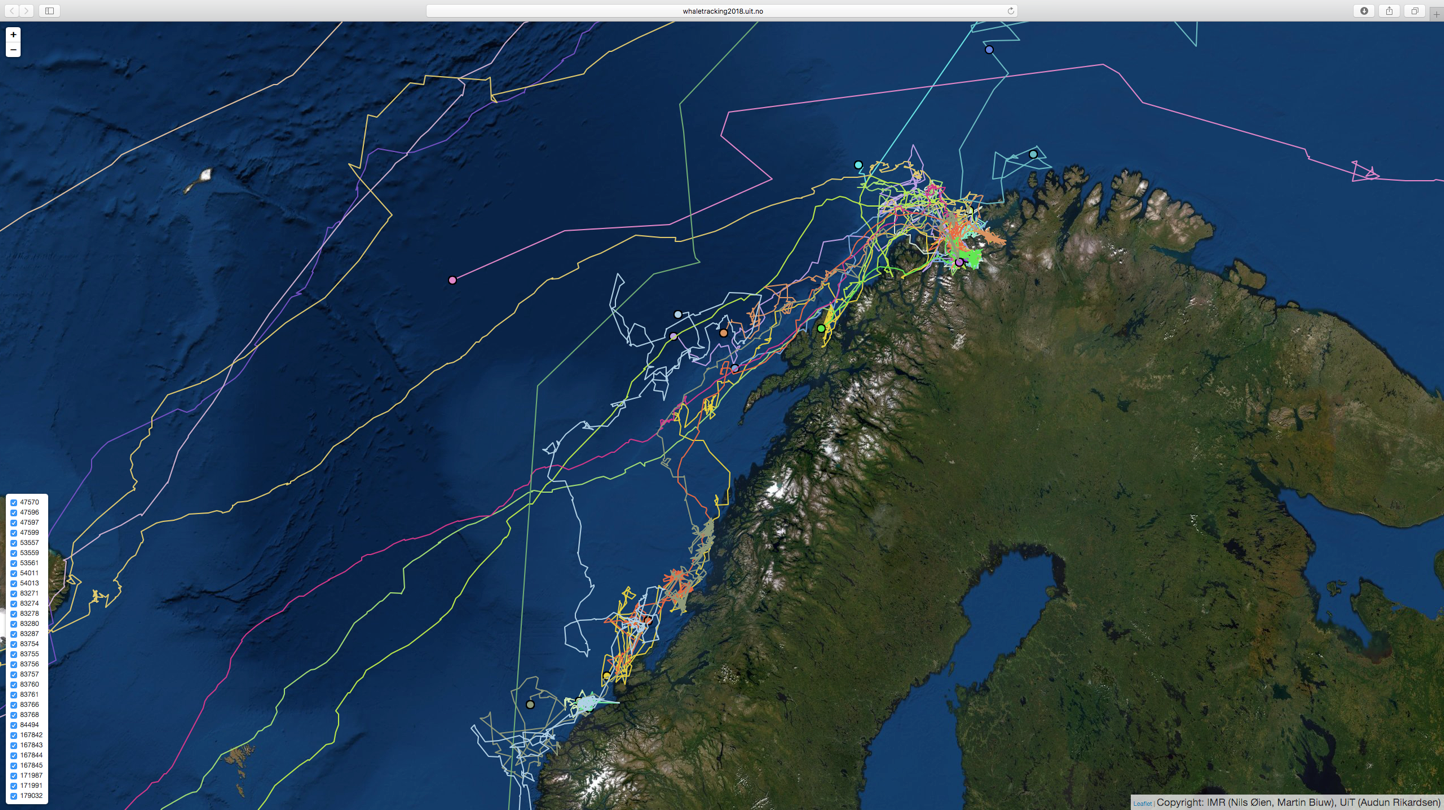
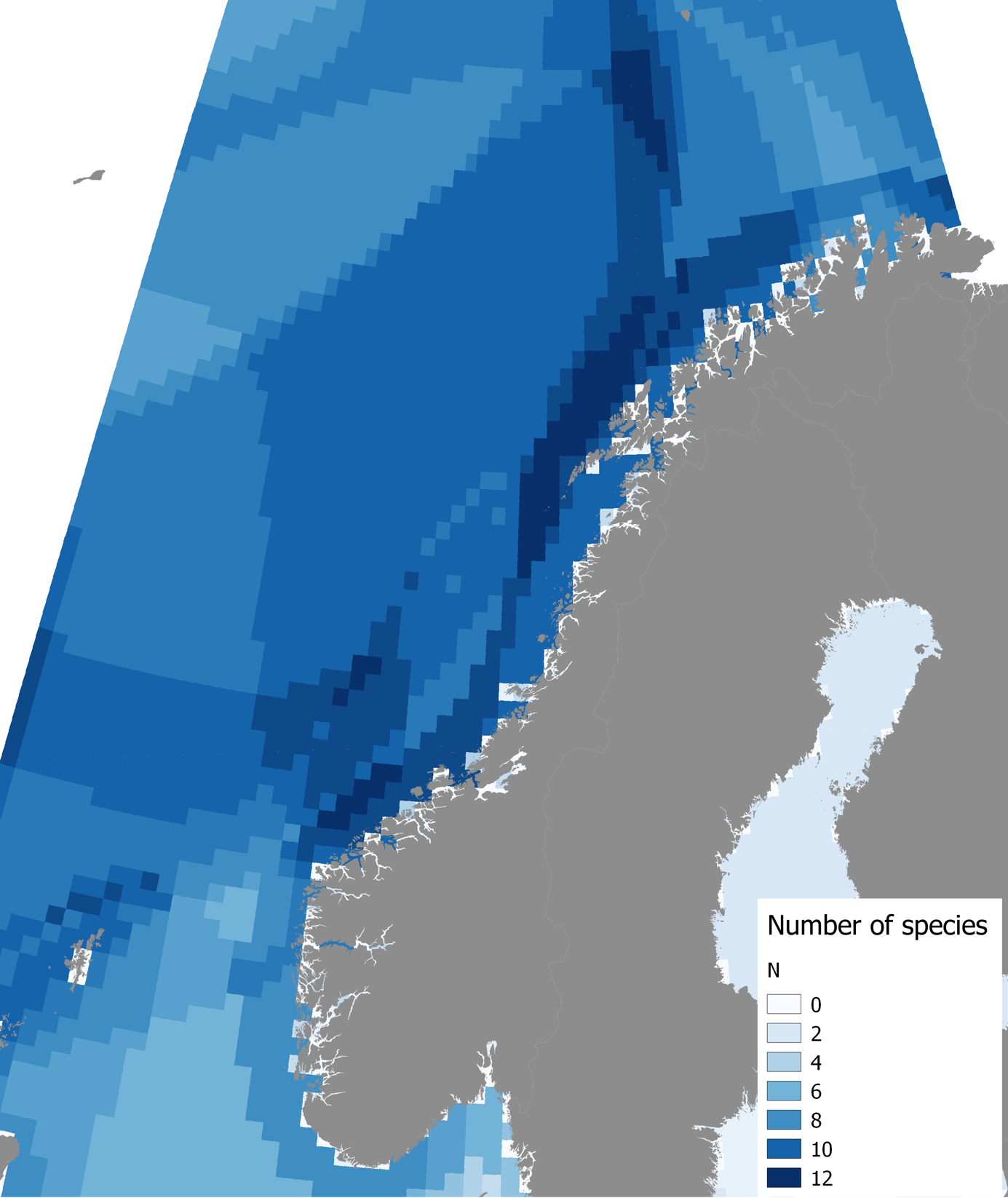
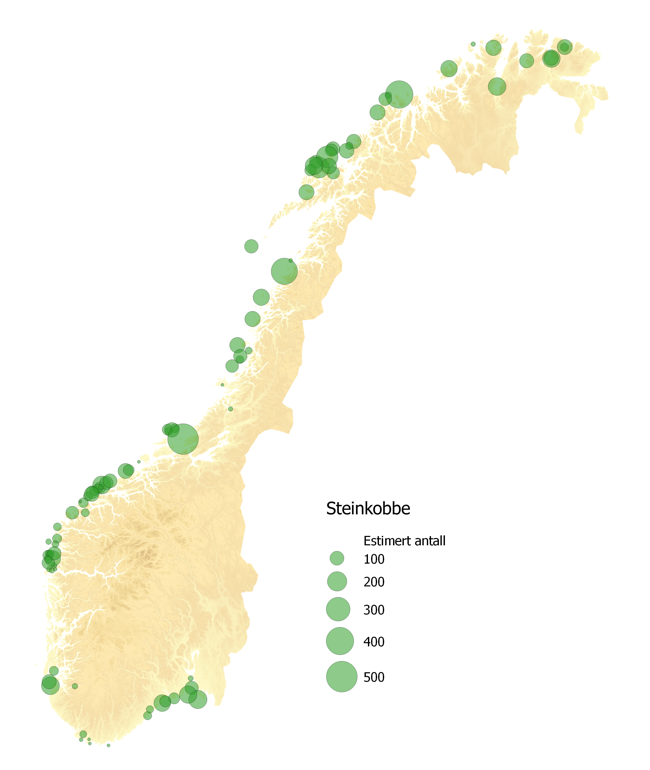
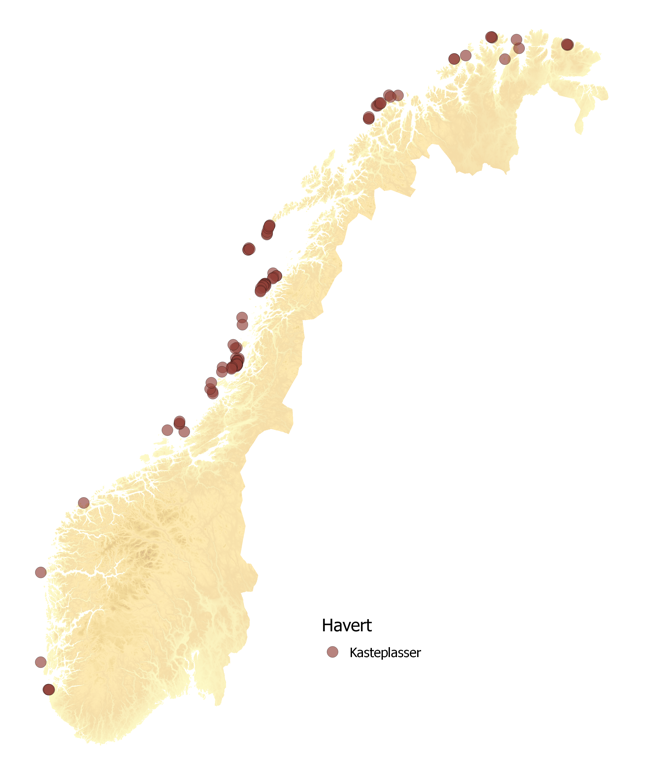
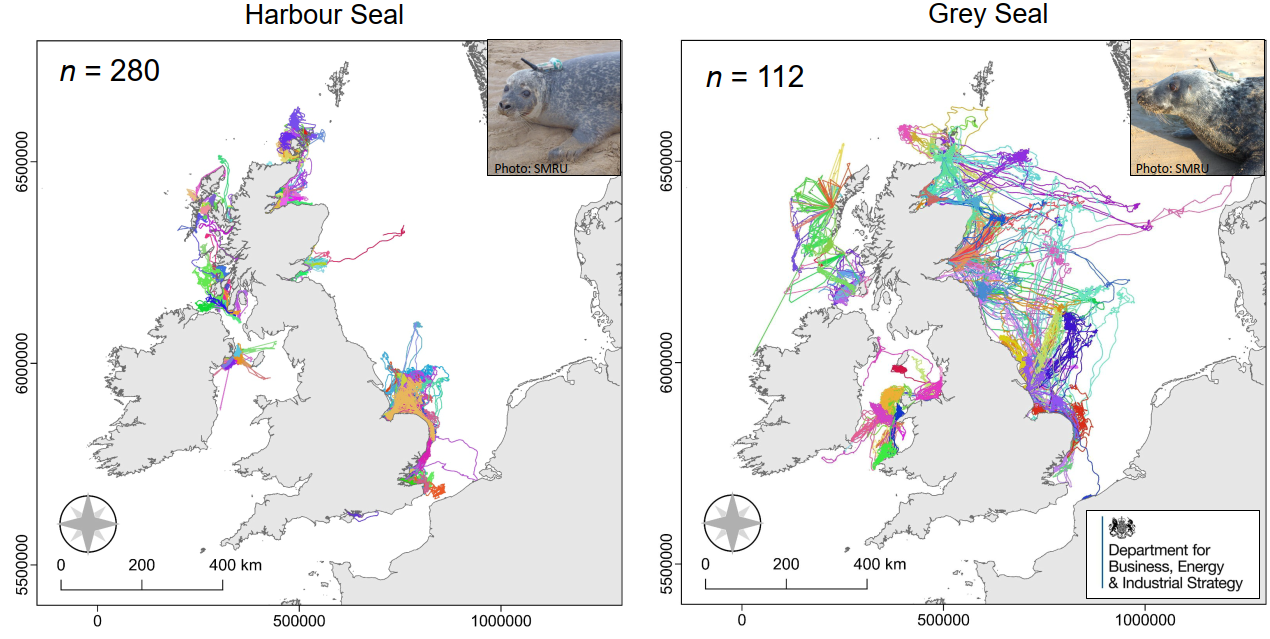
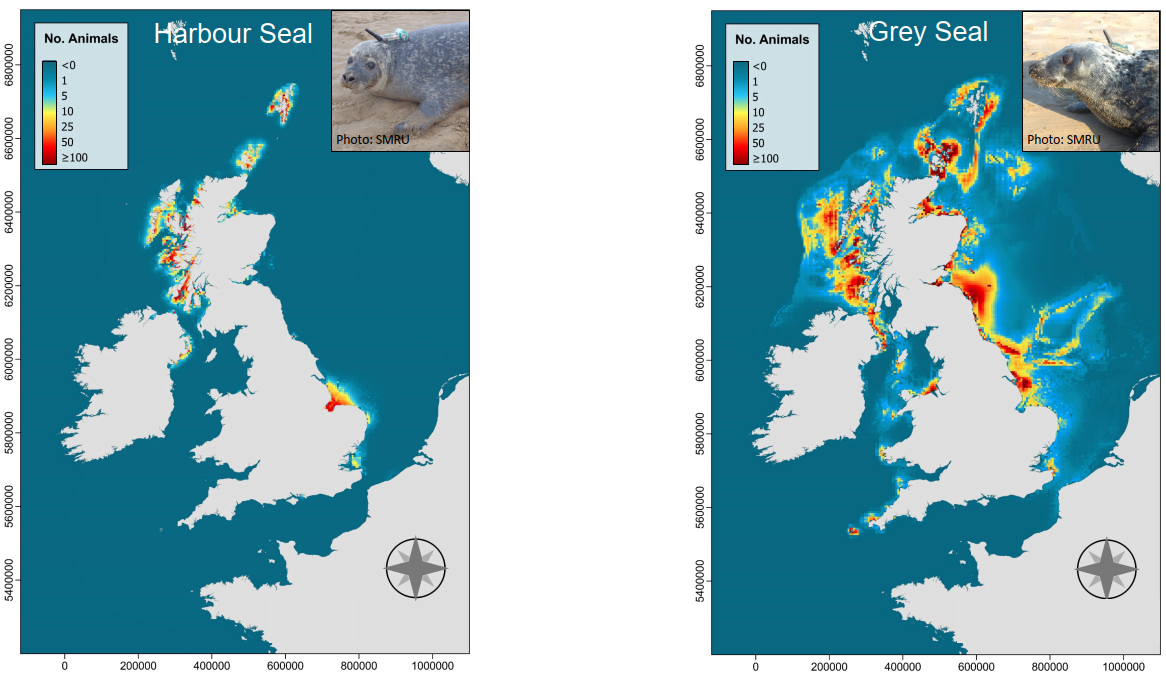
8 - Havbruk til havs – sykdom
8.1 - Smittespredning
I tillegg til lakselus gir andre parasitter (her: virus, bakterier, parasitter) opphav til en rekke smittefarlige sykdommer i oppdrett av fisk i Norge. Dette utgjør et alvorlig problem både for oppdrett av fisk, men også for mulige miljøpåvirkninger. De fleste av de kjente sykdommene i Norge i dag påvirker laks, eller laksefisk, men noen har et bredere vertsspekter. De fleste sykdomsproblemer i oppdrett i Norge har sitt opphav fra bestander av Norsk villfisk, og viser at smitte vil kunne skje fra villfisk til oppdrettsfisk. Åpne produksjonssystemer gjør at det er tett kontakt mellom fisk og parasitter inne i og utenfor merdene. Derfor vil infisert og syk oppdrettsfisk kunne spre parasitter til miljøet, og villfisk vil kunne gi opphav til smitte hos oppdrettsfisk.
Det er sannsynlig at det spres betydelige mengder patogener til miljøet fra smittet og syk fisk i oppdrett, og at villfisk i nærheten av anlegget kommer i kontakt med patogenene (eksponering). Utskilling av parasitter kan være fritt til vannet, gjennom slim, avføring, kjønnsprodukter eller når verten går i oppløsning etter at den er død. Dette gjelder både for oppdrettsfisk og villfisk.
Konsekvensene av kontakt med et patogen vil variere, og er oftest en kompleks interaksjon mellom ulike verter, patogener, predatorer og andre miljøfaktorer. Noen patogener vil smitte mange ulike verter, mens andre kun smitter noen få. Miljøet har stor innvirkning på interaksjonen mellom patogen og vert. Det fysisk-kjemiske miljøet vil i tillegg ha betydning for spredningspotensialet til en parasitt ved å påvirke hvor lenge parasitten er smittsom.
Det er svært sannsynlig at en får sykdom også på lokaliteter til havs, og disse vil på samme måte som lokaliteter i kyst og fjordstrøk utgjøre en smittespredningsfare. Store lokaliteter med mye biomasse og høyt antall fisk vil kunne slippe ut betydelige mengder parasitter. På den andre siden vil god strøm og store vannmasser fortynne slik at det i stor grad blir lave konsentrasjoner av agens, og sannsynligvis spredte verter. Siden vandringsmønstre til villaks er lite til moderat kjent er det per i dag ikke mulig å si noe konkret om risiko for at smitte vil påvirke disse fiskene. Men muligheten for smittespredning fra slike anlegg til vandrende villaks er helt klart tilstede.
Som ved kysten vil oppdrettsanlegg tiltrekke seg fisk som vil beite på fôrspill og faeces. Anleggene vil i tillegg kunne ha en «rev»-effekt som samler fisk. Dette vil kunne øke tettheten av potensielle verter ved anleggene. Dersom anleggene tiltrekker seg laksefisk på vandring vil disse kunne bli utsatt for et svært høyt smittepress fra laksepatogener når de passerer. Tett kontakt med mange ulike arter fisk øker også muligheten for at parasitter fra oppdrett kan etablere seg i «nye» verter, og at patogener fra villfisk greier å etablere seg hos laks. Hvor sannsynlig dette er, er svært vanskelig å si noe om, men selv om massedød av fisk i naturen er uvanlig, har det forekommet i Norge. De alvorligste tilfellene av slike episoder påvist hos villfisk i Norge er knyttet til introduksjoner av eksotiske patogener. Lokaliteter til havs vil i noe grad forventes å møte andre verter, eller andre bestander av verter, enn lokaliteter ved kysten og i fjordene. Disse vertene kan ha andre parasitter eller varianter av disse enn det som er kjent på kyst. Det kan altså introduseres nye parasitter, eller nye varianter av parasitter, som kan bli et fremtidig problem for oppdrett, og over tid derfor også bli et problem ift smittespredning tilbake til havet.
9 - Appendix
9.1 - Modellert strøm og temperatur
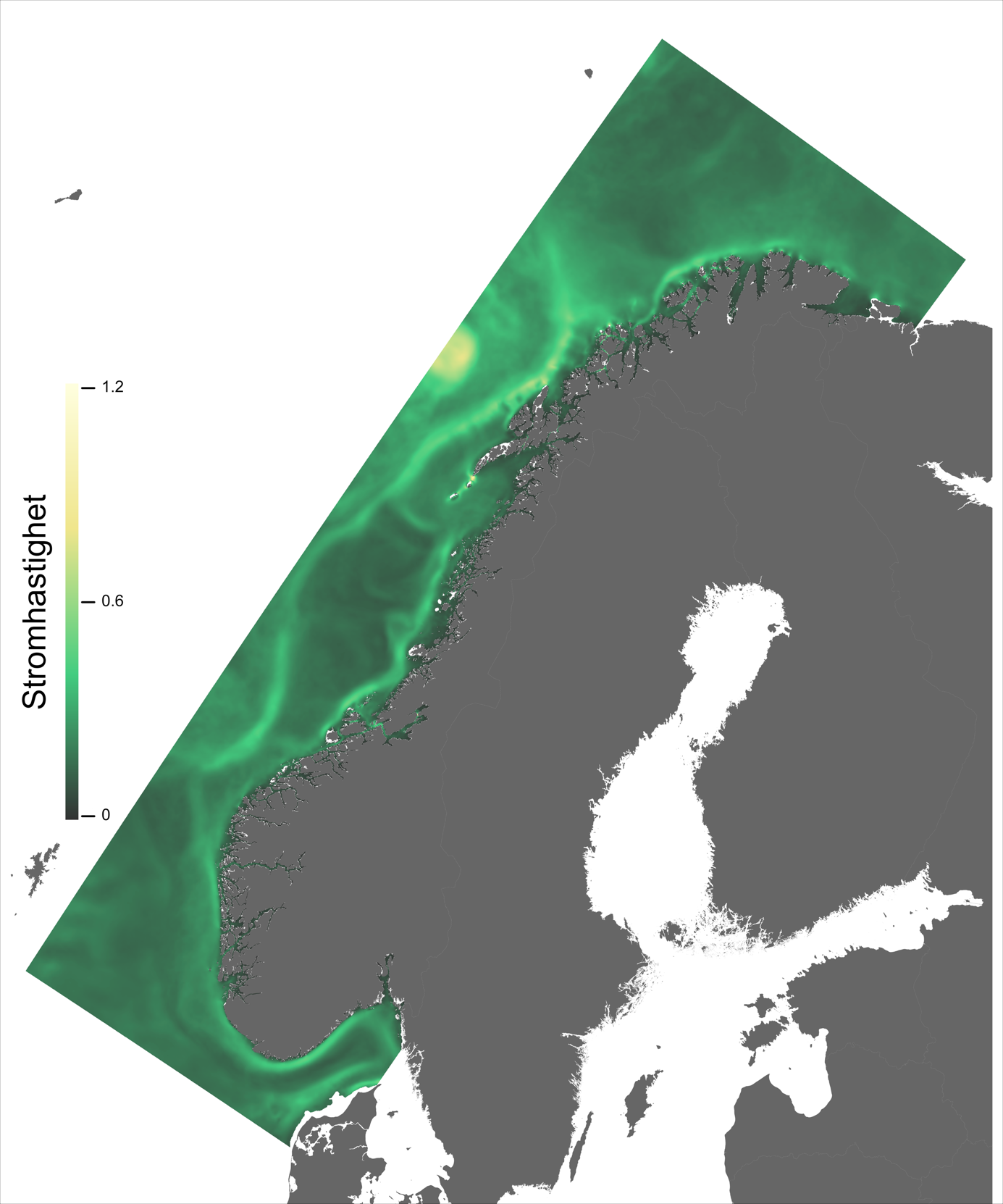
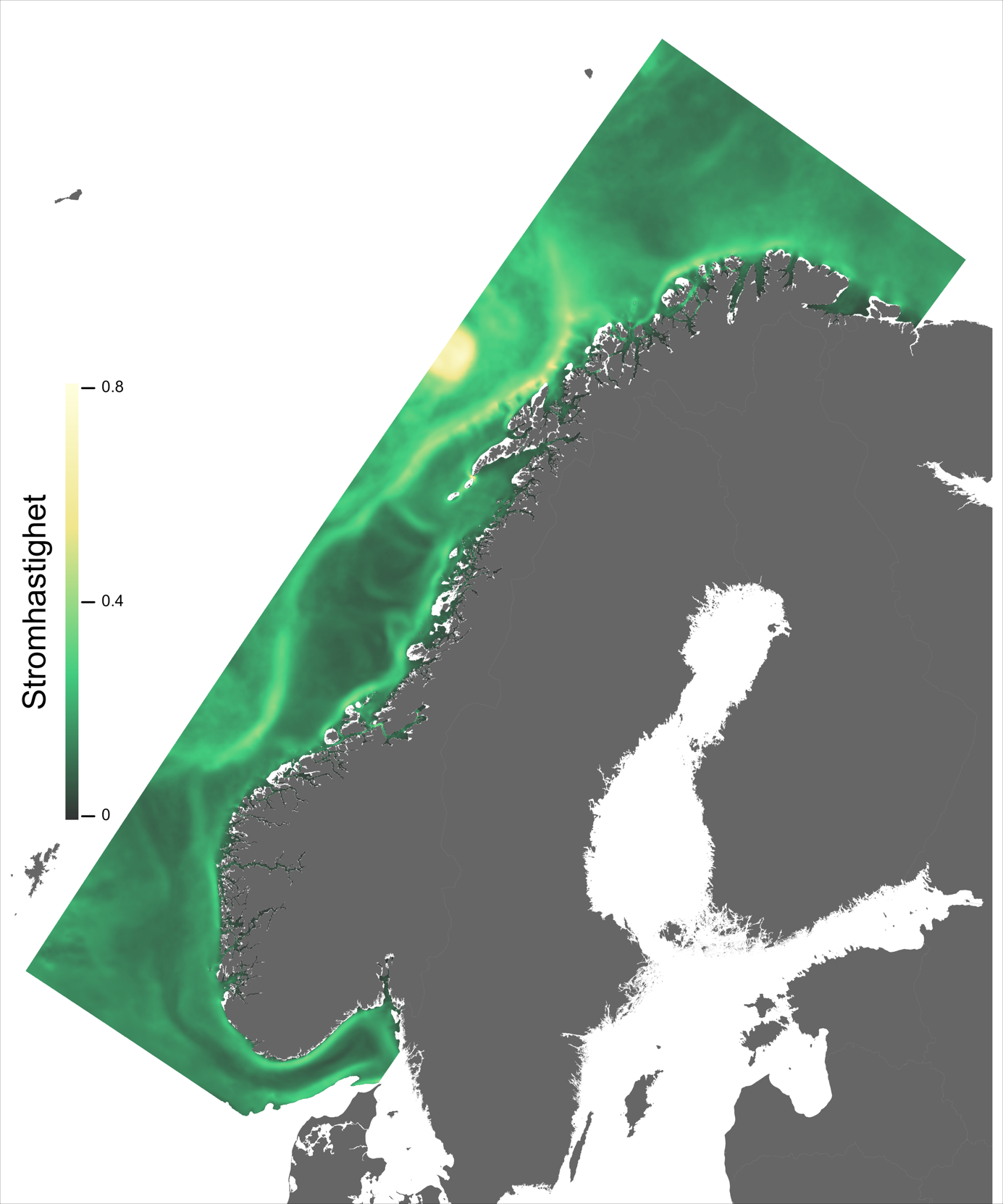
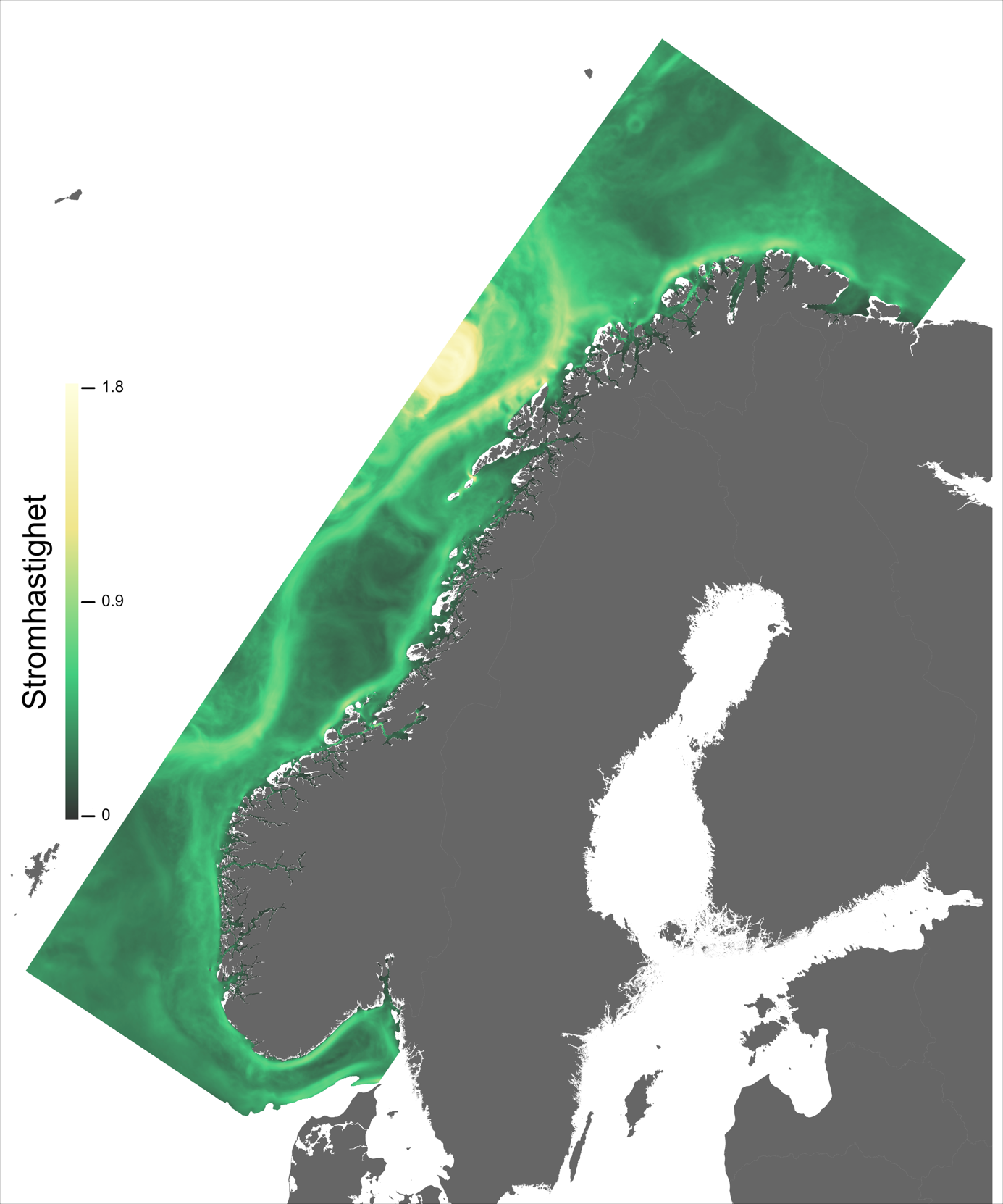
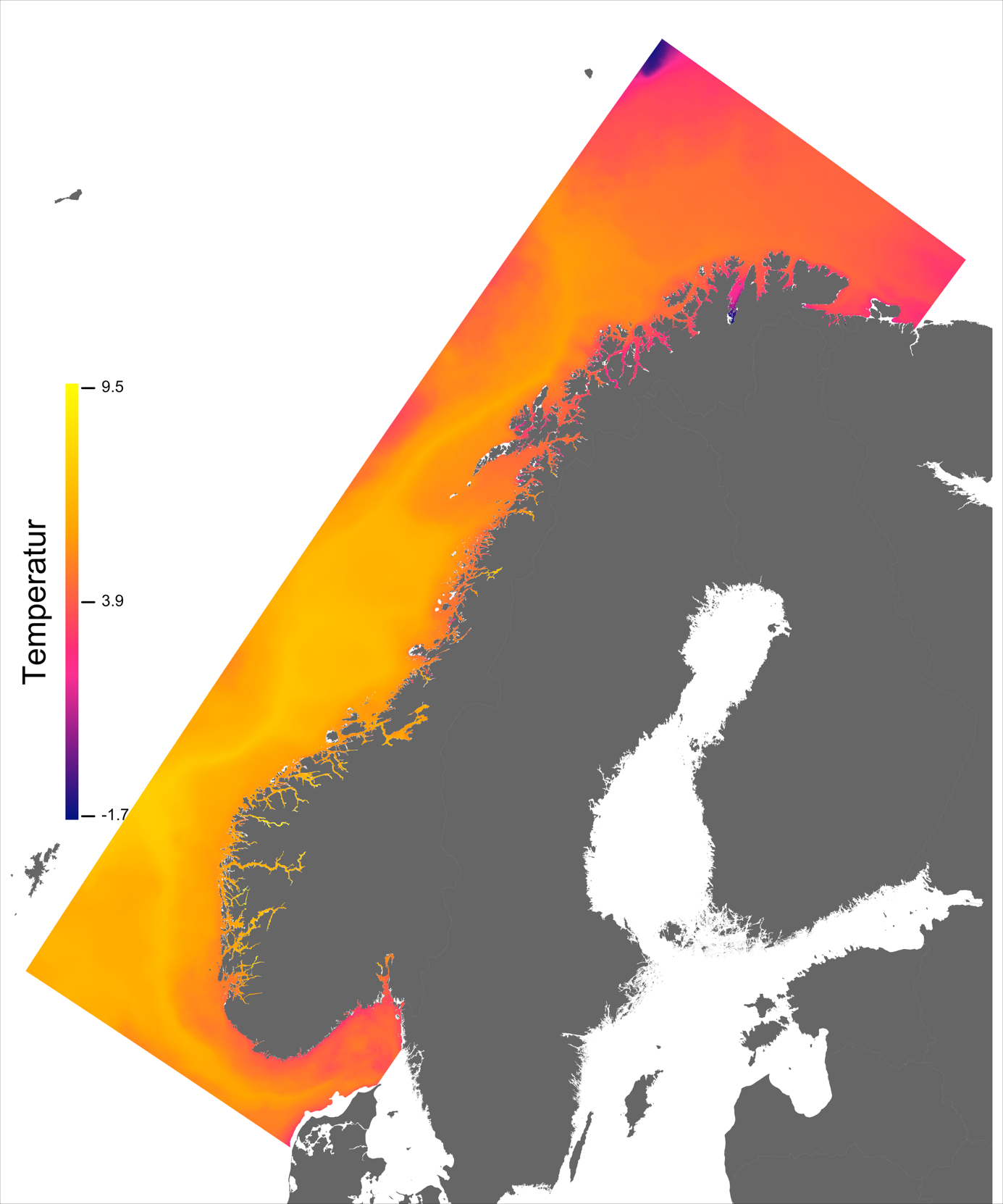
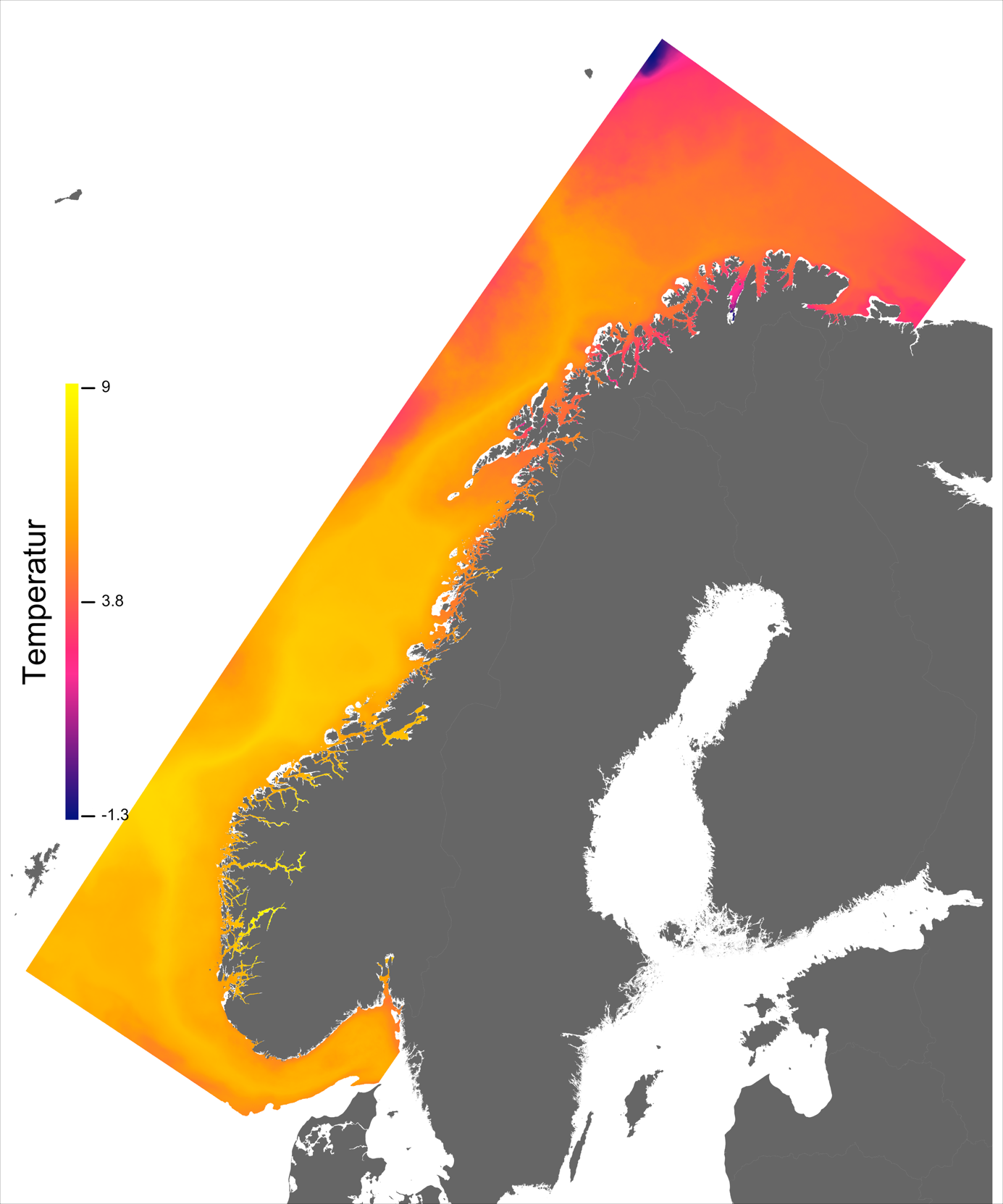
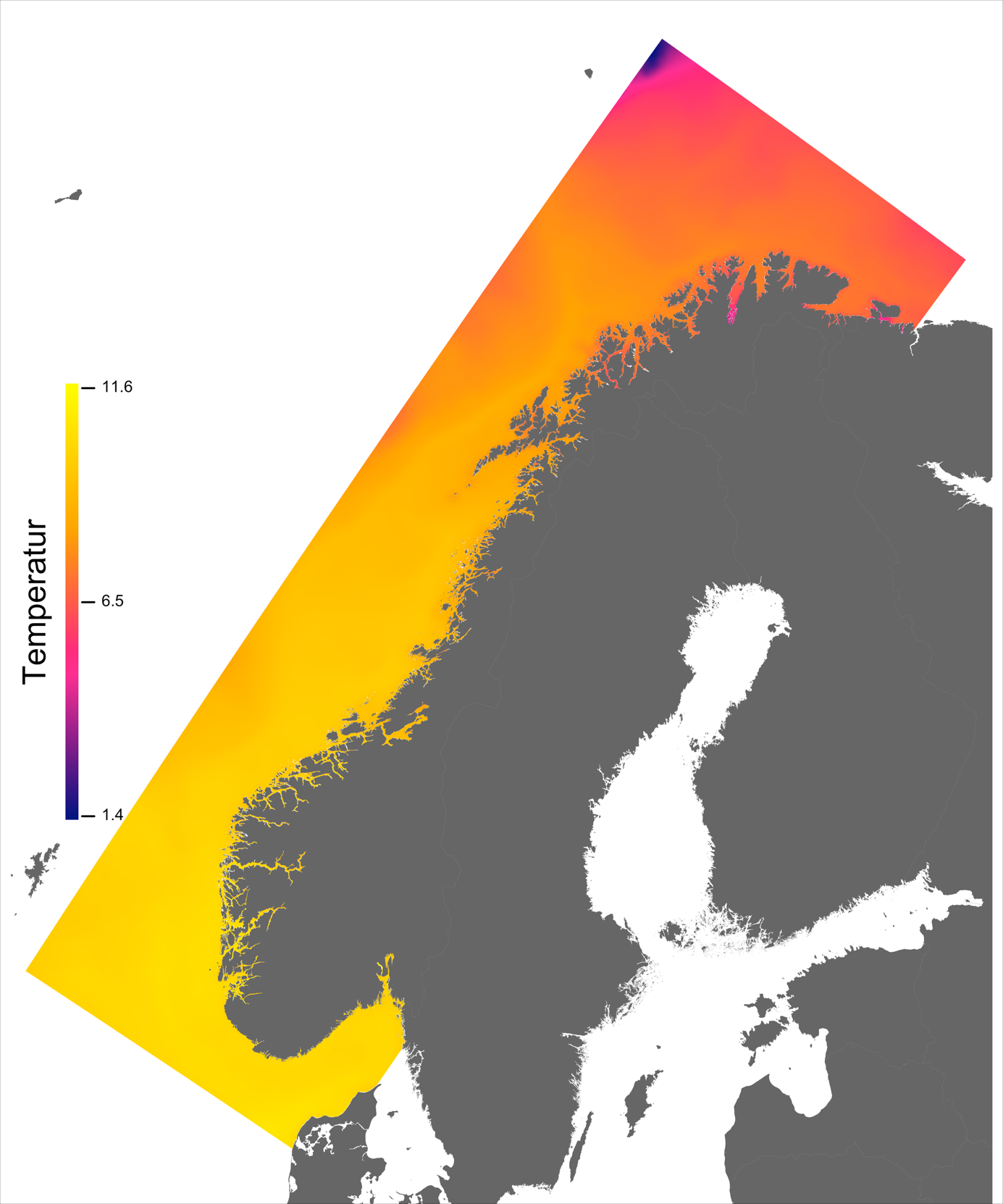
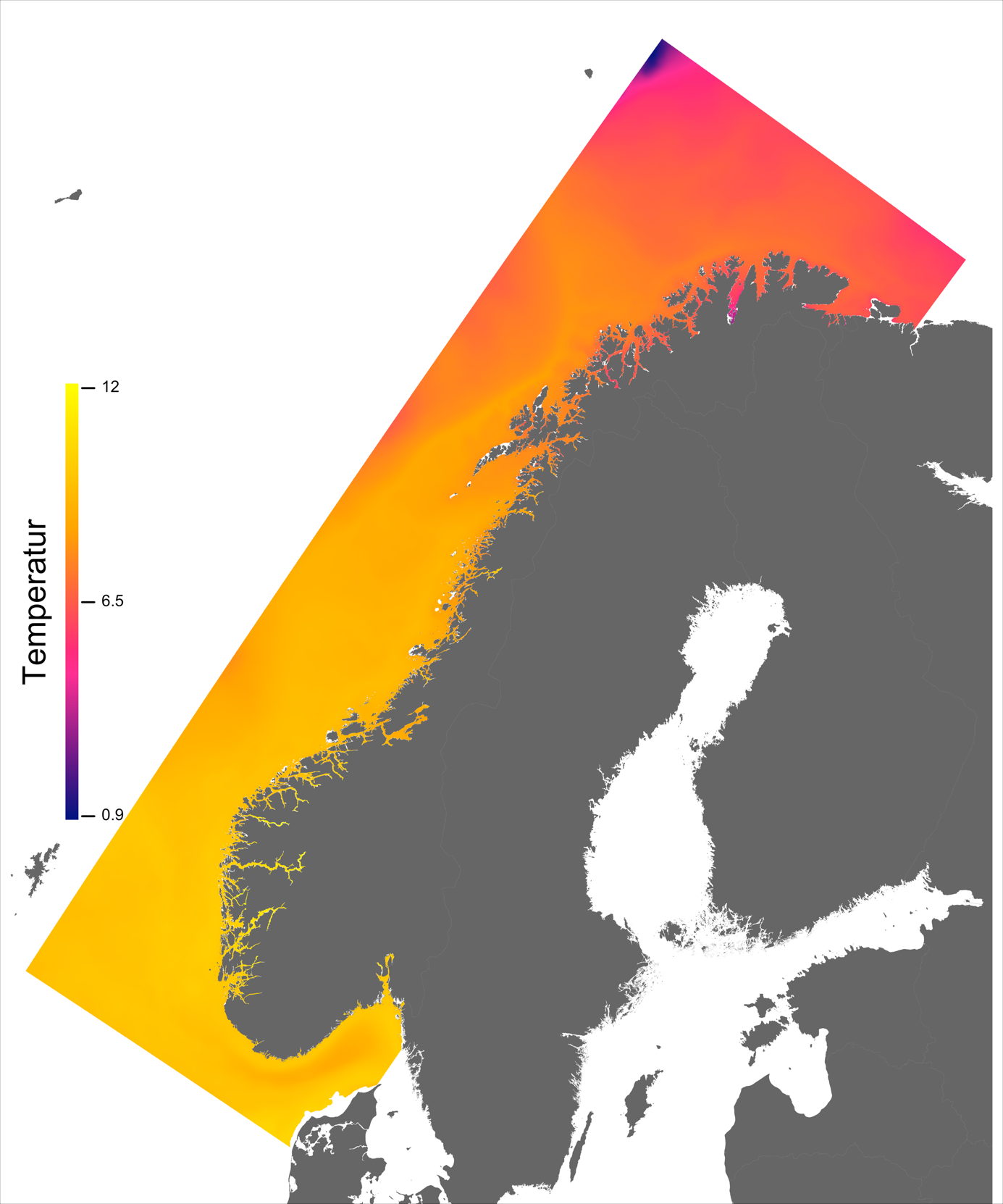
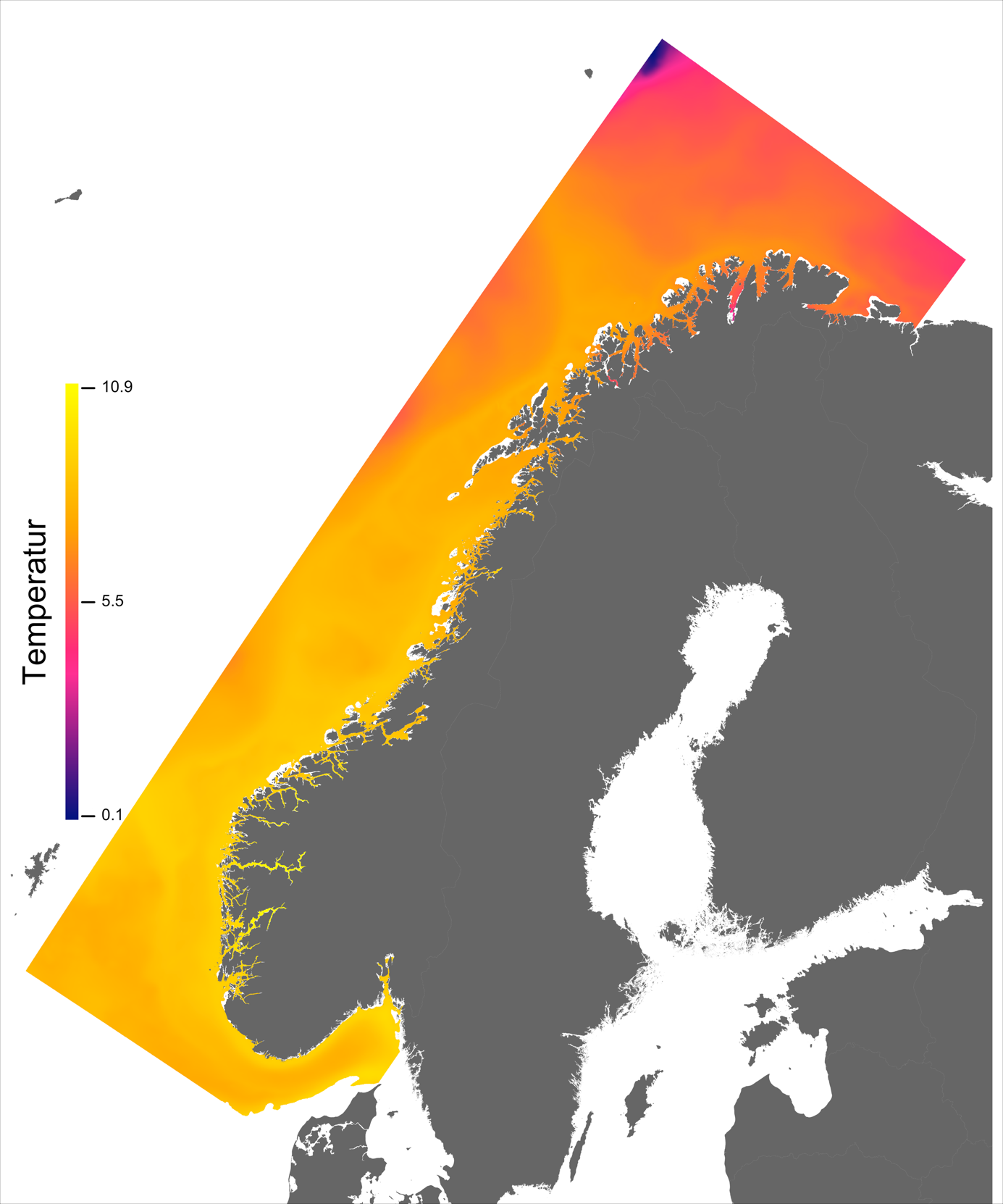
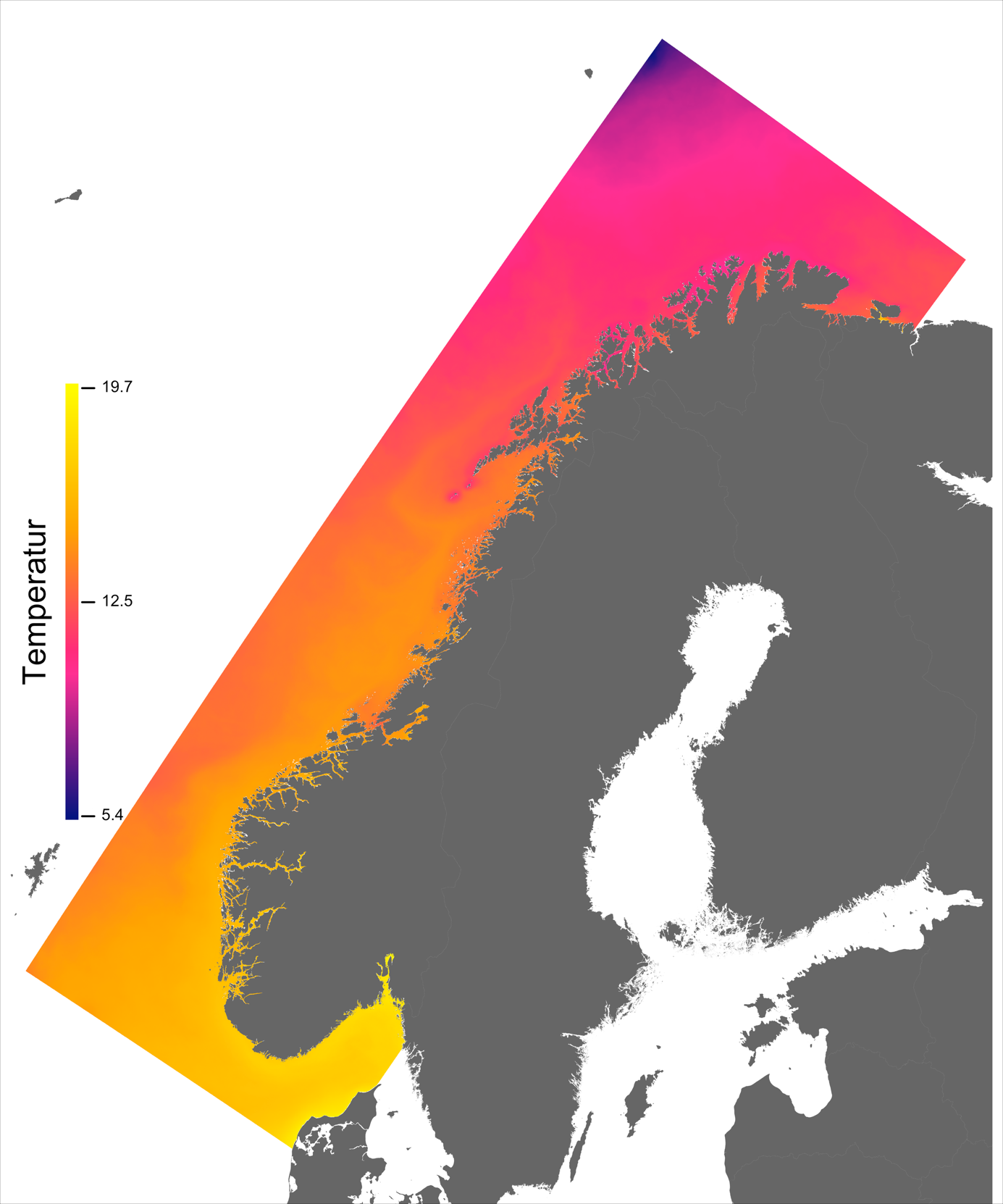
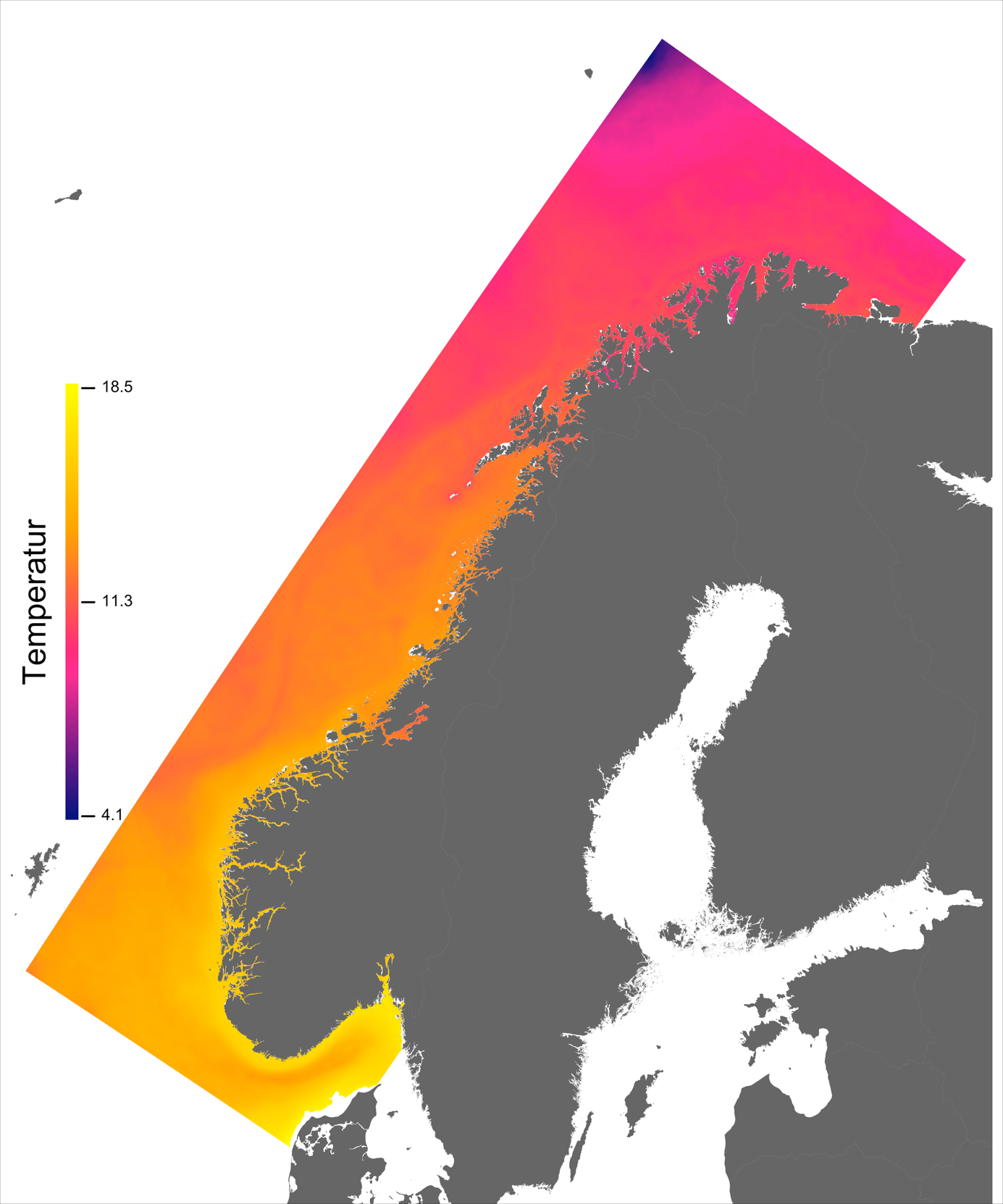
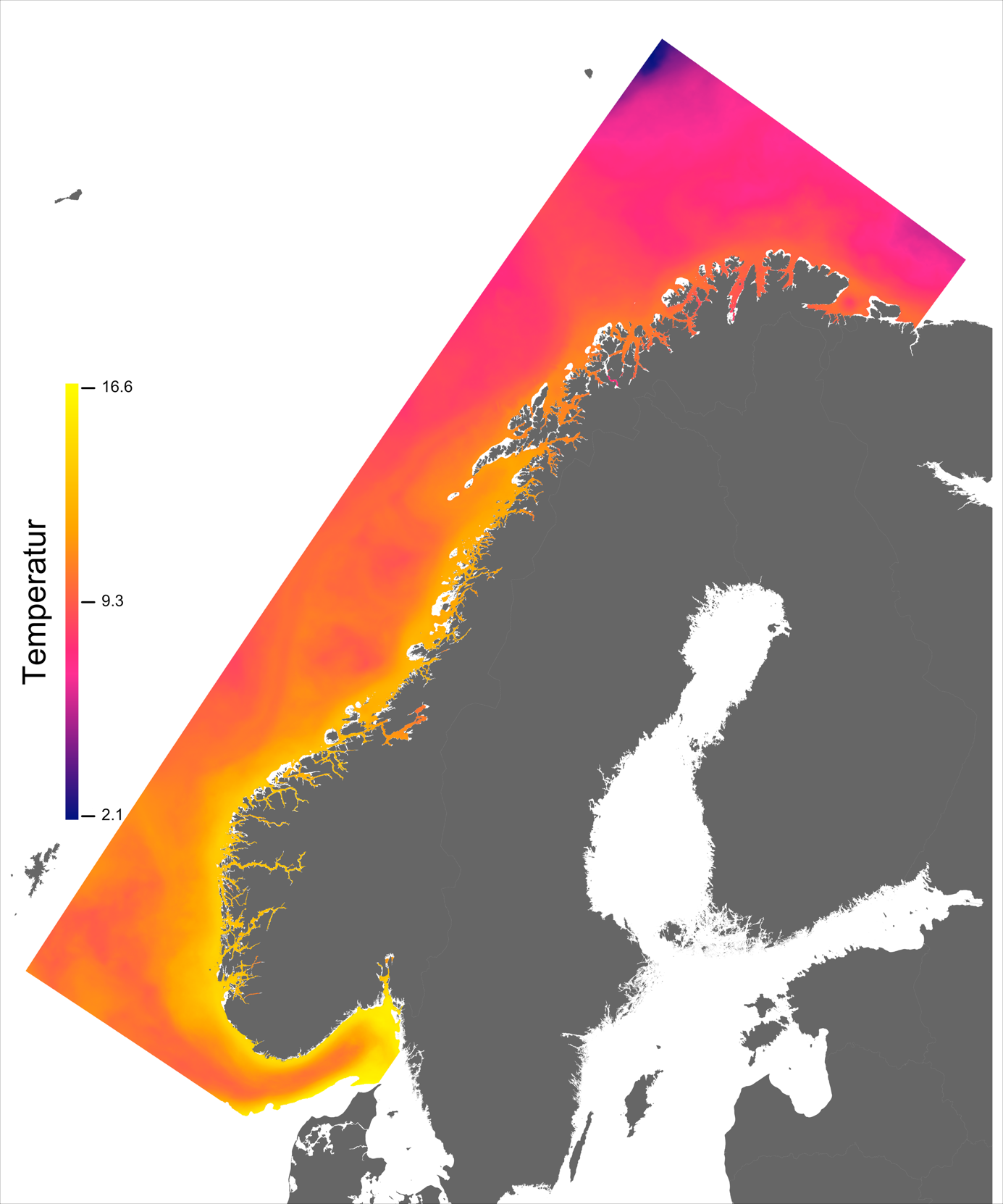
9.2 - Observert strøm
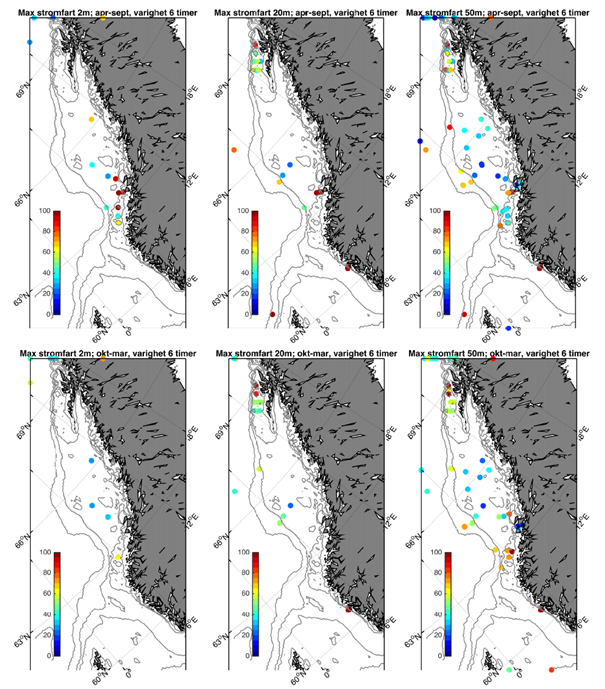
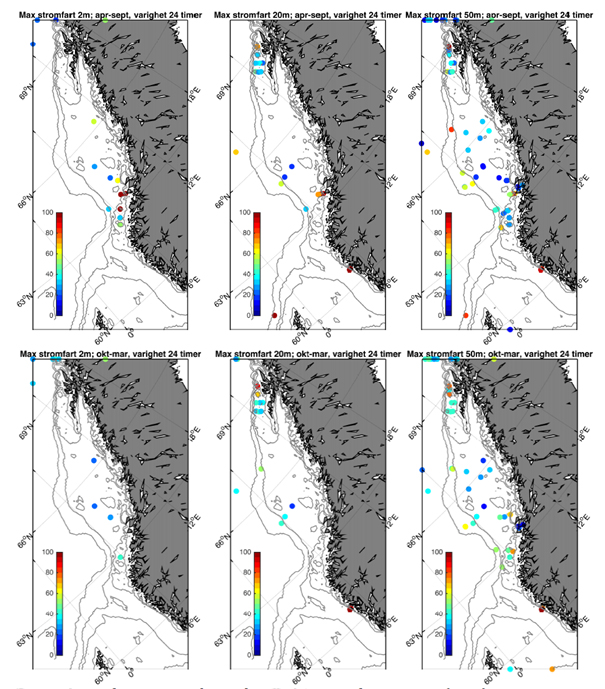
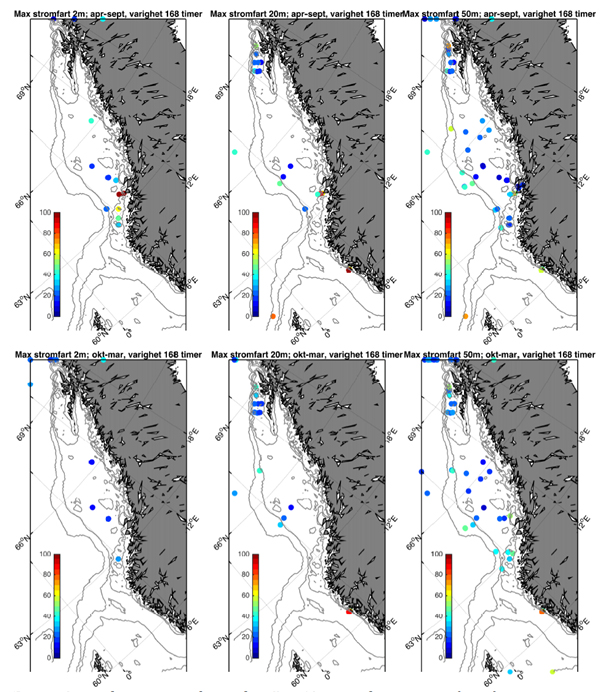
9.3 - Relativ egnethet i de foreløpige og første utpekte områdene fra Fiskeridirektoratet
Sett i sammenheng med de 12 områdene som Fiskeridirektoratet foreløpig har utpekt som aktuelle basert på liten overlapp med annen bruk (Figur 9.16), fremstår følgende områder som egnet (fra sør til nord, navngitt basert på nærmeste kjente topografisk struktur og områdenummer); Norskerenna sør (2), Norskerenna nord (4), Frøyabanken sør/nord (10 og 11), Haltenbanken (3), Sklinnabanken (1) og Trænabanken (5). Mindre egnede områder finner vi i Barentshavet, basert på de generelt lave temperaturer (Figur 9.16b), og inkluderer Tromsøflaket (7) og Bjørnøyrenna (6). Områdene 8, 9 og 12 dekkes ikke i sin helhet av strøm- og temperaturarkivene vi har benyttet, men de delene som er representert her kommer ikke fordelaktig ut med hensyn til hverken strøm (Figur 9.16a) eller temperatur mht svømmeevne (Figur 9.16b).
Figur 2.17 viser frekvensfordeling av relativ egnethet for alle 800 x 800 m ruter i kartet i Figur 2.16a. Det er en overvekt av ruter som anses som relativt mindre egnet. Her er imidlertid relativ egnethet mht strøm fremstilt som en lineær funksjon av modellert strømhastighet mellom 0 og henholdsvis 0,48 (vedvarende strøm) og 0,6 m/s (kritisk strøm). Dvs at for modellert 95 persentil av strøm som overstiger 0,3 m/s får relativ egnethet under 0,5. Fra Hvas et al. (2019) vet vi imidlertid at undersøkelser ikke tilsier en lineær avhengighet for strøm, men mer sannsynlig velegnede strømforhold til en nærmer seg tersklene for enten vedvarende eller kritiske strømhastigheter (altså avhengig av varigheten på strømhendelsene) og så en mer brå avtagende relativ egnethet. Mao gir forenklingen med en lineær avhengig for strømhastigheter en bias mot mindre relativ egnethet i Figur 2.17.
Figur 2.18 viser relativ egnethet for de 12 områdene utpekt av Fiskeridirektoratet som funksjon av modellert strømhastighet, men her med variabilitet innad i hvert delområde basert på de modellerte tidsseriene. De fleste områder er nokså homogene med hensyn til miljøforhold (liten variabilitet i relativ egnethet), men vi ser betydelig variabilitet i delområder 9. Dette område er ikke godt dekket av modell men har åpenbart betydelig variabilitet i strømhastigheter knyttet til innstrømning av Atlantisk vann i Barentshavet og generelt vekslende værforhold. Det er en tendens til mindre variasjon i relativ egnethet nærmere kyst og større variabilitet i mer åpne farvann. Delområde 4 og 7 er ved forgreningspunkter for kantstrømmen (inngangen til henholdsvis Norskerenna og Barentshavet) og variabiliteten her illustrerer hvor dynamiske disse område er med hensyn til strøm og fronter.
Relativt egnethet basert på miljøforhold som tilsier god fiskevelferd rapportert i Hvas et al. (2019) innenfor hvert av de 12 foreløpig utpekte delområdene styres av ulike parametere. Figur 2.15 viser relativt egnethet basert på medianverdier for hvert delområde 1-12 for midlere og ekstreme (5 og 95 persentil) temperaturer og strømhastigheter for de ulike dypene 2, 20 og 50 m. De enkelte kart som viser alle de faktiske verdiene i hele områder er vedlagt i 9 – Appendix, Figur 9.1 – 9.14. Vi ser at strømhastigheter er spesielt høye i delområde 12. Høye strømhastigheter og lave temperaturer trekker også relativ egnethet ned for 8 og 9. Områdene 6 og 7 har relativt høye strømhastigheter, men til forskjell fra lengre sør spesielt lave temperaturer som reduserer ytelse og dermed tåleevne for høye strømhastigheter. Områdene 2, 10 og 11 ligger tett inntil sentrale strømgrener for kyststrømmen og har relativt høye strømhastigheter. Dette er til dels også tilfelle for områdene 1 og 4. Men i motsetning til områdene i Barentshavet er temperaturene mer fordelaktig i forhold til påvirkning på svømmeevne. De resterende delområder 3 og 5 fremstår som relativt gunstige både med hensyn til strøm og temperatur og avstand til kant- og kyststrøm, selv om område 5 delvis overlapper med Trænatråget som skjærer innover sokkelen og gir opphav til topografisk forsterkning av strøm.
Fra strømobservasjonene (Figur 2.6 og 2.7) ser vi at målt strømhastighet under laksens vedvarende svømmekapasitet sammenfaller med område 3 og 5 som foreløpig utpekt fra Fiskeridirektoratet. Område 1, 10 og 11 er mer usikker fordi de er relativt nær kjernen av kyststrømmen eller kantstrømmen. Det er ikke tilgjengelig strømmålinger som vi kjenner til som kan fortelle om strømhastigheter i de andre foreslåtte områdene annet enn i deler av område 6 og en sørlig del av område 12 hvor det er strømrigger i forbindelse med henholdsvis Fugløy- og Svinøysnittet.