Methods for mapping and monitoring of alien marine species in Norway
Suggestions for a national program
In Norway there is currently only monitoring programs for specific marine alien species and a general sporadic mapping of the occurrence of such species for some parts of the coast. A systematic national program for mapping and monitoring of alien species in Norwegian waters is vital.
When making such plans it is necessary to identify which are the most likely door knockers, how will they the get here, what is their preferred habitat and where will they most likely enter.
In this report we give an analysis of preferred habitat of established alien species and potential doorknockers which will create a basis for the choice of methods. Fouling on vessels is likely the most important vector for new introductions into the Norwegian coast. Here we give a detailed overview of ports with a high risk of introductions dependent on the frequency of port call directly from foreign ports. Based on this we suggest a rolling monitoring plan following international recommendation.
This report summarises existing international protocols for investigations of alien species in the marine environment and gives three suggestions for methodology for the Norwegian coast. The suggestions presented have different levels of intensity and thereby cost. Testing of the different methods is recommended to evaluate their efficiency and give a more exact cost estimate.
Forord
I 2021 fikk Havforskningsinstituttet (HI) i samarbeid med Norsk Institutt for naturforskning (NINA) i oppdrag fra Miljødirektoratet. Oppdraget skulle øke kunnskapen om fremmede marine arter i Norge og gi en samlet oversikt over nåværende kartlegging og overvåkning, samt vurdere vektorer for innførsel av nye arter til landet, presentere internasjonal overvåkningsmetodikk og komme med forslag til et nasjonalt overvåkningsprogram for fremmede marine arter for Norge. Dette er den andre rapporten som publiseres for å fullføre dette oppdraget.
Sammendrag
Norge har i dag overvåkning av enkelte fremmede marine arter og en sporadisk generell kartlegging av forekomsten av slike arter langs deler av kysten. Det er derfor behov for en systematisk plan for kartlegging og overvåkning av fremmede marine arter i Norge.
Når man skal lage en slik plan er det viktig å vite hvilke arter som kan forventes til landet, hvordan de kan komme seg hit, hvor de lever og hvor i landet de mest sannsynlig vil etablere seg først. Her gis en analyse av levested for etablerte arter og arter som forventes til landet, som kan danne basis for valg av metodikk.
Begroing av skrog er trolig den vektoren som i dag utgjør den største risikoen for introduksjon av fremmede marine arter. Her gis en detaljert oversikt over havner som har høy risiko for slike introduksjoner basert på frekvensen av anløp direkte fra utenlandsk havn. Denne vurderingen danner basis for ett rullerende overvåkningsprogram etter internasjonale anbefalinger.
Rapporten gir også en oppsummerende oversikt over eksisterende internasjonal metodikk og basert på dette gis tre alternative metodikkforslag for norskekysten. Forslagene som presenteres har ulik intensitet og vil ha store forskjeller i kostnader. Det anbefales at ulike metoder prøves ut for å vurdere effektiviteten og gi mer nøyaktige kostnadsoverslag.
1 - Innledning
Norge har verdens nest lengste kystlinje og det er utfordrende å kartlegge og overvåke fremmede marine arter langs hele kysten. De artene som allerede har etablert seg her er blitt oppdaget nærmest ved tilfeldigheter og gjerne i forbindelse med annen aktivitet slik som feltkurs ved universitetene og andre biodiversitetsprosjekt. Husa mfl. 2022 gir en samlet oversikt over overvåkning og kartlegging av fremmede marine arter i Norge. I perioden 2010-2014 foregikk det en generell kartlegging av fremmede marine arter i båthavner finansiert av Miljødirektoratet. Dette prosjektet har fortsatt til 2021 finansiert av Nærings- og fiskeridepartementet. Denne kartleggingen er likevel begrenset da kun deler av kysten i Sør-Norge er kartlagt og hvert område kun har vært undersøkt en gang. Det finnes også systematisk overvåkning av enkelte arter slik som kongekrabbe og stillehavsøsters.
Å ha god kunnskap om etablerte fremmede arters geografiske utbredelse og forekomst er viktig for å kunne vurdere omfanget av økologiske effekter. Tidlig kunnskap om nye arters etablering i landet er også viktig for å kunne sette inn tiltak mot problemarter. Det har til nå vært mindre fokus på fremmede arter i marine økosystemer enn i terrestrisk miljø. Det er behov for en mer systematisk overvåkning og kartlegging av fremmede marine arter, både etablerte arter og dørstokkarter (arter som forventes til landet).
Det er en utfordring å vite hvilke arter man skal leite etter, hvor man skal leite, hva slags metoder man bør bruke og hvor høy intensitet man bør ha for å oppdage nye arter.
Enhver art som kommer inn til norskekysten fra en annen biogeografisk region har potensiale til å etablere seg her dersom miljøforholdene er sammenlignbare med donorregionen. Her er mulighetene uendelige, det er derfor nyttig å ha fokus på arter som allerede er på farten og som har blitt introdusert til flere steder i verden eller i Europa. I regi av Artsdatabanken har det blitt gjennomført en evaluering av slike arter, en såkalt horisontskanning. Artene som er plukket ut vil bli risikovurdert sammen med etablerte fremmede arter ved revideringen av Fremmedartslista som skal publiseres i 2023 (https://www.artsdatabanken.no/fremmedartslista2018). Det vil likevel være viktig å ha øynene åpne for andre nye arter som kan komme inn til landet, som ikke omfattes av horisontskanningen.
Basert på vurderinger gjort i Husa mfl. (2022) må begroing på skip anses som en av de viktigste vektorene for introduksjon av fremmede marine arter til norskekysten. En analyse av skipstrafikk direkte fra utenlandsk havn til norske havner (Husa mfl. 2022) danner grunnlaget for å vurdere risikoen for hver enkelt havn langs norskekysten. Denne rapporten vil gi en sammenstilling av disse dataene og foreslå ett systematisk program for overvåkning av havner med høy risiko for introduksjoner av nye arter.
Når man skal velge metodikk for slike undersøkelser er det viktig å vite hvor potensielle nye arter kan finnes. Denne rapporten gir derfor en analyse av levested for etablerte artene og de fleste artene som forventes til landet. Det har de seinere årene blitt utviklet internasjonal metodikk for slik kartlegging og denne rapporten vil gi en gjennomgang av disse metodene for å se hva som er best egnet for et nasjonalt program for norskekysten.
2 - Habitatsanalyse av etablerte og forventede fremmede marine arter
Når en skal leite etter fremmede marine arter er det, i tillegg til å ha god kjennskap til stedegen flora og fauna, en fordel å ha et klart bilde av hvilke arter man skal se etter. Teoretisk kan enhver art som overføres fra en region med lignende temperaturforhold som langs norskekysten etablere seg her, men erfaringsmessig er det gjerne arter som allerede er introdusert til Europa eller andre steder i verden som sprer seg til nye områder. For å gi et bedre bilde av hvilke arter som mest sannsynlig vil dukke opp i Norge har Artsdatabanken satt i gang en evaluering av aktuelle dørstokkarter (arter som kan forventes til landet) for Norge, en såkalt horisontskanning inspirert av lignende prosesser internasjonalt. Artene på denne horisontskanningslisten er arter som er introdusert til andre land i Europa og/eller arter som er introdusert til en rekke andre land i de siste tiårene.
Når vi skal velge en metodikk som passer for kartlegging og overvåkning av fremmede arter i norske farvann er det viktig å vite hvor man skal undersøke for å finne de fremmede artene. Noen arter vil finnes overalt, mens det for andre arter kanskje må letes i spesielle habitat. Vi har derfor gjort en habitatanalyse for de etablerte artene og for dørstokkarter er basert på arter som allerede er vurdert for fremmedartslisten (Artsdatabanken, Fremmedartslista 2018 https://www.artsdatabanken.no/fremmedartslista2018) og dørstokkarter som står på horisont-skanningslisten til Artsdatabanken (Artsdatabanken, upubliserte data).
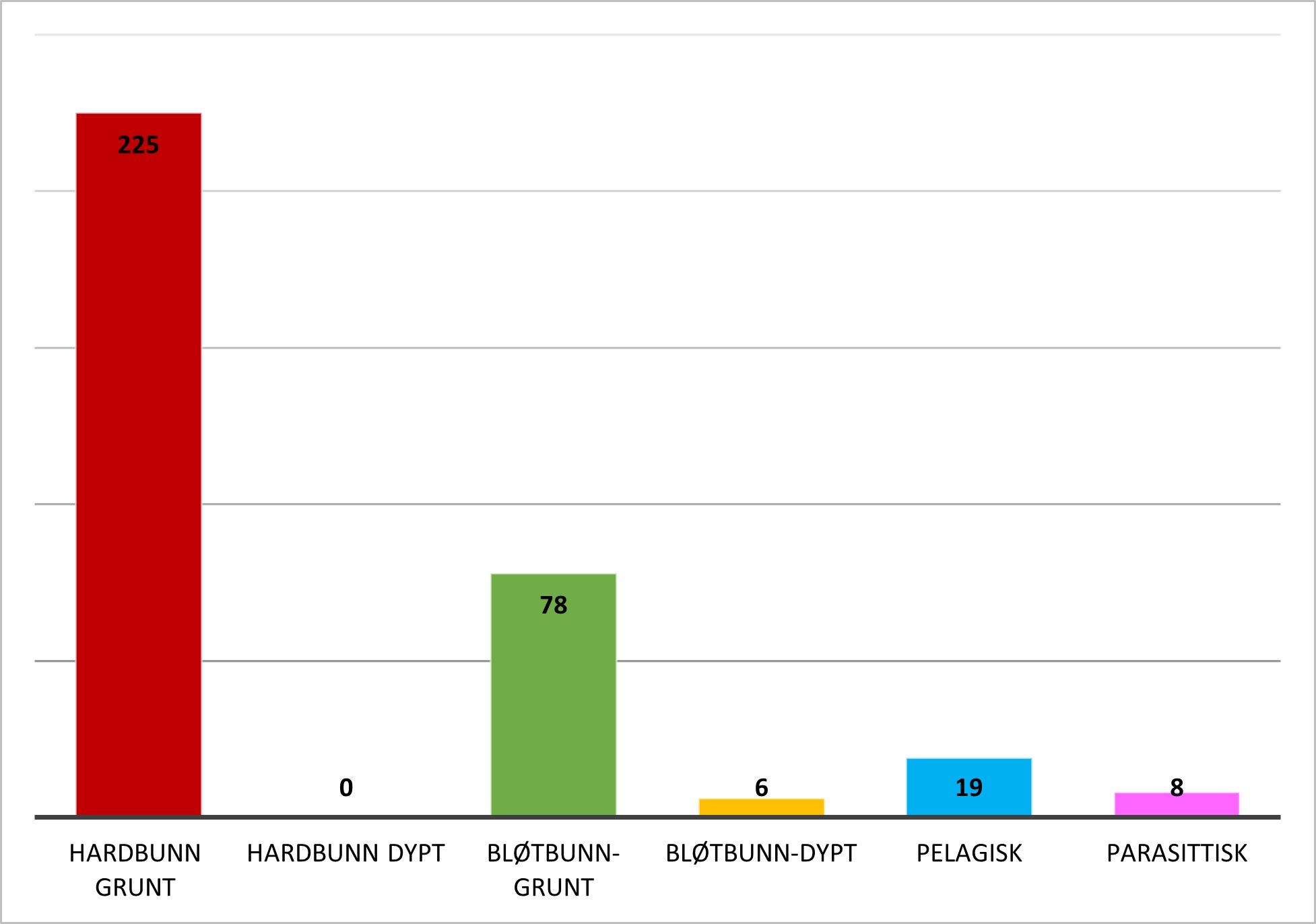
Mange fremmede marine arter trives i flere typer habitat og mange er særlig knyttet til kunstig substrat i marinaer og havner i begynnelsen av etableringen, mens de siden finner veien til naturlige habitat. Figur 1 gir en oversikt over hovedhabitat til 336 etablerte arter og dørstokkarter. Som vi ser er flest arter knyttet til grunn hardbunn og mange finnes på grunn bløtbunn, noen er pelagiske (19) (i vannmassene), noen er parasittiske (9) og noen få finnes på dyp bløtbunn (6). Dyp bunn defineres her som dypere enn 40 meter, mens grunn bunn er fra 0-40 meters dyp.
Figur 1. Marine evertebrater og makroalger: Hovedhabitat for 336 etablerte fremmede marine arter og dørstokkarter (vurderte i 2018 og Artsdatabankens horisontskanningslister 2021).
2.1 - Mikroalger
Mikroalger er ikke vurdert for fremmedartslisten i artsdatabanken, årsaken er manglende kunnskap om hvilke arter som er stedegne og hvilke som er fremmede. Vi har derfor ikke gjort noen analyser av de artene som står på europeiske horisontsskanningslister. Analyser av planteplankton inngår i HELCOM protokollen og prøver tas med planktonhåv.
2.2 - Makroalger
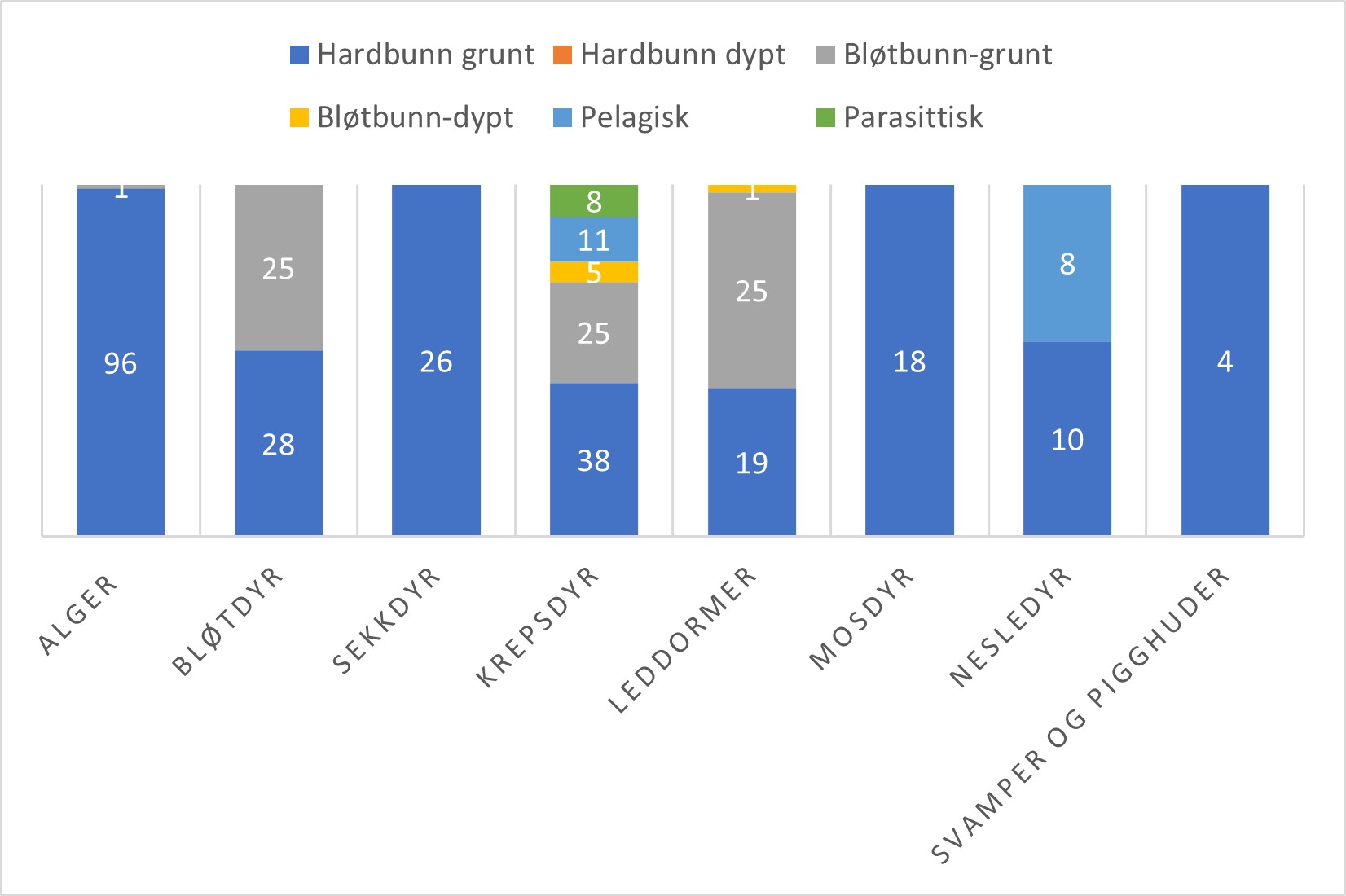
De fleste fremmede makroalger er knyttet til grunn hardbunn, med unntak av en art som er etablert i Norge og vokser som løsliggende på mudderbunn i beskyttede områder (Figur 2). Mange av de etablerte fremmede algeartene vokser på brygger, moloer og annen type kunstig substrat i småbåthavner og kan derfor fanges opp ved å undersøke slike levesteder. For noen arter vil de trolig vokse dypere ned slik at skrap på bunn med enten håndholdt trekantskrape eller kasterive kan fange opp flere arter. For rødalgen Agarophyton vermiculophyllum, som vokser på bløtbunn, må man ha en mer målrettet tilnærming og undersøke for arten i dens foretrukne habitat. For å finne den er visuell observasjon fra land kombinert med kasterive optimal. For å kunne kartlegge forekomstareal av arten bør det brukes en undervannsdrone, videorigg eller snorkling.
2.3 - Marine evertebrater
Marine bløtdyr omfatter blant annet muslinger, blekksprut og snegler. Om lag halvparten av de fremmede artene i denne gruppen er knyttet til hardbunn, mens resten finnes på grunn bløtbunn, enten over eller i sediment (Figur 2). For å finne disse må man undersøke ulike typer habitat på lokalitetene. Skraping av brygger og kai-pilarer, samt undersøkelser av steiner og svaberg vil kunne fange opp artene som er knyttet til hardbunn. De to fremmede østersboresneglene som er etablert i Danmark må man gjerne lete i østersforekomster for å finne. For å finne arter som er knyttet til bløtbunn er strandsøk å anbefale, gjerne kombinert med en skjellsanker for å finne arter i sedimentet. Man kan også bruke en håndholdt grabb eller en trekantskrape på grunn bløtbunn. Miljø-DNA prøver fra sediment kan gi en indikasjon på om målarter i denne grupper er å finne i området.
Marine sekkdyr eller sjøpunger er alle hovedsakelig knyttet til hardbunn og finnes ofte på brygger og tau i marinaer (Figur 2). Kolonidannende sjøpunger som for eksempel havnespy kan også vokse utover bløtbunn når de drypper ned fra bergvegger. Denne arten vokser også hovedsakelig dypere enn de andre sjøpungene (5-20 meter) og har i liten grad blitt funnet på småbåtbrygger. Det kan derfor være fornuftig å komplementere med undersøkelser med vannkikkert, dykking eller videotransekter.
Marine krepsdyr er den gruppen med størst variasjon i habitattype og dyp (Figur 2). De fleste artene lever på grunn hardbunn (38) eller grunn bløtbunn (25). Elleve arter lever hele livet i vannsøylen mens åtte er parasittiske. Noen arter (5) lever på dyp bløtbunn og dette er hovedsakelig store krabber. Bunnlevende krepsdyr kan fanges i teiner, ved skraping av bunnsubstrat og mange lever også inni klaser av skjell og lignende på tau og brygger. Pelagiske krepsdyr fanges med planktonhåv og parasittiske ved å undersøke vertsdyret.
Figur 2. Evertebrater og makroalger: Hovedhabitat for 336 etablerte fremmede marine arter og dørstokkarter (vurderte i 2018 og Artsdatabankens horisontskanningslister 2021) fordelt på artsgrupper.
Av fremmede leddormer lever 25 arter i grunne bløtbunnsområder, de fleste lever i sedimentet, mens 19 arter er knyttet til grunn hardbunn (Figur 2). Flere av disse er kalkrørsormer som vokser på skjell og annet hardt substrat eller bygger revlignende strukturer. Kun én dørstokkart finnes i dyp sedimentbunn. For å lete etter fremmede leddormer kreves det derfor en kombinasjon av undersøkelser som skraping og visuell observasjon av hardbunn samt en sedimentgrabb eller skraping av bløtbunn. Miljø-DNA prøver fra sediment kan gi en indikasjon på om målarter i denne grupper er å finne i området.
Fremmede mosdyr, svamper og pigghuder er alle knyttet til grunn hardbunn og kan finnes ved skraping av brygger, påler og visuelle undersøkelser av svaberg og annet hardt substrat (Figur 2).
Fremmede marine nesledyr omfatter både arter som er fastsittende på hardbunn og arter som lever fritt i vannmassene, hovedsakelig maneter (Figur 2). De fastsittende på hardbunn finnes ved skraping av brygger, påler og visuelle undersøkelser av hardbunn, mens de pelagiske finnes ved håvtrekk. Publikum er også flinke til å rapportere inn fremmede maneter gjennom Dugnad for havet (https://dugnadforhavet.no/).
2.4 - Fisk
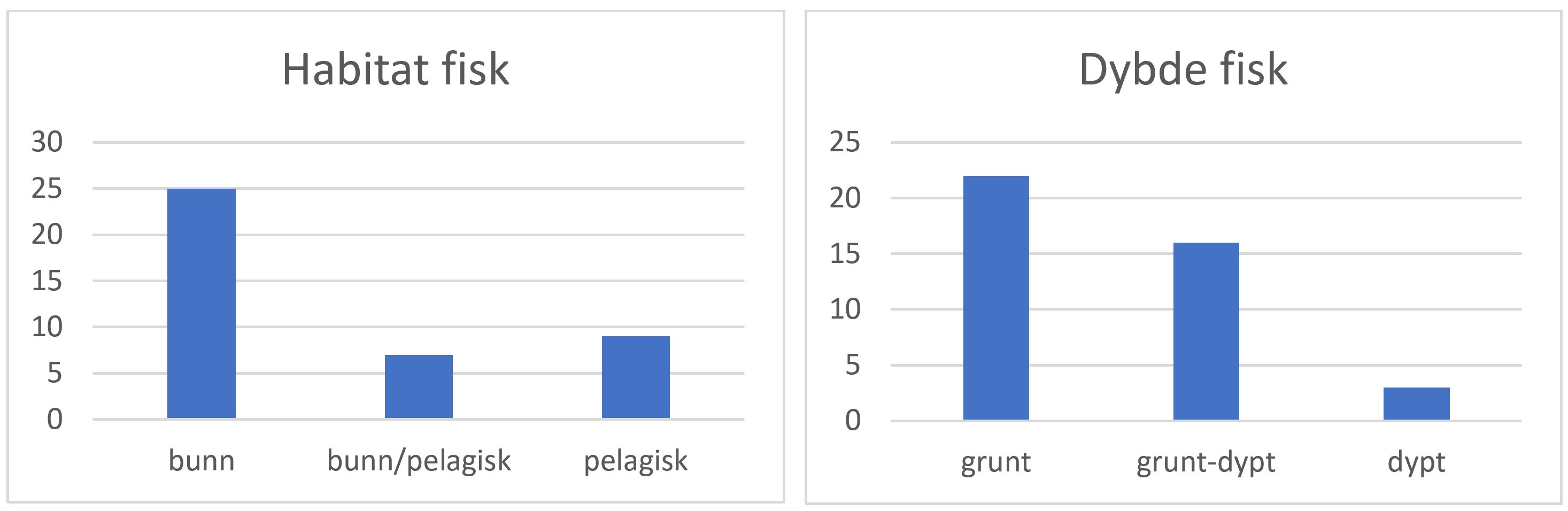
Hovedhabitatet for fremmed marin fisk, inkludert etablerte arter og dørstokkarter, er ved bunnen på grunt vann (Figur 3). Av de bunnlevende artene hvor informasjon om bunntype var lett tilgjengelig (f.eks. FishBase), var bløtbunn vanligere enn hardbunn.
Figur 3. Antall arter fremmed marin fisk for fastlands-Norge og Svalbard, som er bunnlevende og/eller pelagiske (til venstre), og om de lever grunt (<40 m) eller dypt (til høyre). Artene er fra fremmedartsdatabasen (Artsdatabanken 2018), van den Heuvel-Greve mfl. (2021), og fra horisontskanningen høsten 2021 (Artsdatabanken), se Tabell 1.
Det er to etablerte fremmede fiskearter med relevans for hav, pukkellaks og regnbueørret (Tabell 1). Begge er assosiert med høy risiko for påvirkning på norsk naturmangfold (Artsdatabanken 2018). I havet lever de pelagisk fra grunt vann ned imot 250 m. Pukkellaks er anadrom og gyter i ferskvann seint på sommer/høst. Den har en 2-årig livssyklus og alle hanner og hunner dør etter gyting. Den har kommet inn i Norge fra Russland, og har fått stor oppmerksomhet i senere år da den har hatt en enorm egenspredning (Berntsen mfl. 2020). Det har blitt gjort analyser og modellering basert på fangster av pukkellaks i 2017 og 2019 for å øke kunnskapen om hvilke miljøvariabler som kan forklare den romlige fordelingen i Norge i estuarier og elver. Man fant at elver nær Russland, elver med stor vannføring samt elver med mye villaks hadde store fangster av pukkellaks i både 2017 og 2019 (Berntsen mfl. 2021). Det forventes særlig mye pukkellaks i elver som munner ut i Varangerfjorden samt i store elver i Troms og Finnmark. Samtidig vil pukkellaksen trolig bli relativt tallrik i mange elver uavhengig av størrelse og lokalisering (Berntsen mfl. 2021). Regnbueørret har både en anadrom sjøvandrende form og en stasjonær form i ferskvann. Det er produksjon av regnbueørret i flere norske oppdrettsanlegg langs kysten og fra hvor det skjer rømninger (Anon. 2011). Oppdrettsanlegg finnes fra Rogaland til Nordland, først og fremst i Vestland, og den totale biomassen var i 2020 nesten 40.000 tonn (Fiskeridirektoratets akvakulturstatistikk, www.fdir.no). Selv om produksjonen av regnbueørret har vært økende i Norge siden midten av 1990-tallet har den i liten grad etablert naturlig reproduserende bestander (Skaala mfl. 2020).
Pukkellaks er dørstokkart for Svalbard. Den forekommer allerede i havområdene omkring Svalbard og på grunn av klimaendringene så forventes den å kunne reprodusere der i fremtiden, i ferskvann (Artsdatabanken 2018). Majoriteten av dørstokkarter og potensielle dørstokkarter for fastlands-Norge lever på forholdsvis grunt vann langs kysten (Tabell 1). Foretrukket habitat er for de aller fleste fiskearter ulike typer av bunn (leirbunn, sandbunn, steinbunn), mens noen er pelagiske. Mange er brakkvannsarter og det er relativt få som lever i havvann med høy, oseanisk salinitet. Det er også flere arter som veksler mellom ferskvann og havvann, for eksempel for å reprodusere (anadrome eller katadrome), eller av andre årsaker. Disse kan ofte oppholde seg i estuarier og elvemunninger.
Familien med flest dørstokkarter og potensielle dørstokkarter er Gobiidae, kutlingfisker (Tabell 1). Dette er stort sett kutlinger fra området omkring Svartehavet, det Kaspiske hav og Azovhavet («Ponto-kaspiske» området). Disse er generelt bentiske, gruntlevende brakkvannsarter. Svartmunnet kutling, Neogobius melanostomus, en dørstokkart med svært høy risiko (Artsdatabanken 2018), forekommer f.eks. i Østersjøen på grunt vann langs kysten under sommerhalvåret (<10 m dybde), men dypere vinterstid (Behrens mfl. 2021). Der forekommer den særlig i nærheten av store havner fordi den spres med skipstrafikk (Kotta mfl. 2016).
For å ha størst mulig sjanse å forhindre etablering av nye fremmede arter er det essensielt å oppdage introduksjoner tidlig. I Østersjøen, for eksempel, er det nå urealistisk å eliminere svartmunnet kutling fordi den er godt spredt og etablert (Ojaveer mfl. 2015). Tiltak for å bekjempe fremmede arter er mest kostnadseffektive jo tidligere i invasjonsprosessen man setter dem inn (Leung mfl. 2002). For fisk så synes miljø-DNA i kombinasjon med ulike fangstmetoder i kystsonen å være gode metoder for tidlig oppdagelse. Med tanke på viktige spredningsveier som båttrafikk og egenspredning, bør man være særlig observant på områder i nærheten av store havner, samt i Sørøst-Norge med nærhet til andre land i Norden og Nord-Europa. Det er også et stort potensial for å involvere for eksempel artsfiskere og sportsdykkermiljøer for å rapportere observasjoner av nye fremmede arter. Marmorert berguer, Sebastiscus marmoratus, ble f.eks. oppdaget i en fiskekonkurranse i Oslofjorden (Hansen & Karlsbakk 2018).
Tabell 1. Fremmed marin fisk for fastlands-Norge og deres foretrukne habitat sammenfattet på familienivå. Rene ferskvannsarter er ikke inkludert, men et utvalg av arter som forekommer i marint miljø er inkludert (anadrom, katadrom, brakkvann). Unge livsstadier forekommer ofte pelagisk (ikke notert i tabellen).
Familie
# Arter
Dybde(m)
Økosystem (og livsstil)
Habitat
1Etablerte fremmede arter:
Salmonidae
2
0-250
marin, brakkvann og ferskvann (anadrom)
pelagisk (marint)
1Dørstokkarter:
Anguillidae
1
0-460
marin, brakkvann og ferskvann (katadrom)
bunn
Gobiidae
1
0-30
brakkvann, ferskvann
sandbunn, steinbunn
Sciaenidae
1
<100
marin, brakkvann, kyst og estuarier
leirbunn, sand-leirbunn
Sebastidae
1
0-50
marin
steinbunn, kyst, nært land
2Potensielle dørstokkarter:
Anguillidae
1
1-400
marin, brakkvann og ferskvann (katadrom)
bunn
Argentinidae
1
>200
marin
bunn
Blennidae
2
0-15
marin
bunn, klippekyst
Cyprinidae
2
0-30
brakkvann og ferskvann
bunn/pelagisk, estuarier
Fundulidae
1
0-10
marin, brakkvann og ferskvann
bunn/pelagisk, kyst, fjæresone, elvemunning
Gobiidae
10
0-30
marin, brakkvann og ferskvann
bunn, kyst, elvemunning
Moronidae
1
0-10
marin, brakkvann og ferskvann
bunn, pelagisk, kyst
Mugilidae
1
5-10
marin, brakkvann og ferskvann (katadrom)
pelagisk, kyst
Salmonidae
7
0-250
marin, brakkvann, ferskvann (anadrom)
først og fremst pelagisk (marint)
Sebastidae
1
3-100
marin
steinbunn, kyst
Syngnathidae
1
0-5
marin, brakkvann
i vegetasjon over sand eller leirbunn
Tetraodontidae
1
50-250
marin
sand-, leir-, steinbunn
1Klassifisert som dørstokkart i Fremmedartsdatabasen 2018 (Artsdatabanken). 2Utvalg fra liste over arter til pågående horisontskanning i regi av Artsdatabanken 2021. Noen av disse var også med som potensielle dørstokkarter i Fremmedartsdatabasen (2018), men ble da vurdert til liten sannsynlighet for etablering.
3 - Geografisk vurdering av risiko knyttet til vektorer for nye introduksjoner
Basert på en gjennomgang av fremmede marine arter som er registrert i Norge (Husa mfl. 2022) ser vi at det er flest arter som er registrert eller har etablert seg langs kysten av sør og midt Norge til Trøndelagskysten (Figur 4). Dette skyldes høyst sannsynlig endring i sjøtemperaturen som blir lavere jo lengre nord man kommer, også for stedegne arter er det slik at artsmangfoldet synker mot nord langs kysten (Brattegard & Holte 1997). Mange arter er varmekjære og har sin nordlige grense et sted langs kysten vår, enten fordi vinteren er for kald for overlevelse eller fordi sommeren ikke er varm nok for reproduksjon. De fleste av våre fremmede marine arter har kommet til Norge som sekundær spredning fra andre land i Europa, der de først ble introdusert fra en annen biogeografisk region. Mange av de fremmede artene i Europa er også varmekjære og vil ha begrenset spredningspotensial i våre nordligste områder. I en omfattende undersøkelse av havnen i Narvik, som har mottatt ballastvann fra fremmede fartøy i mer enn 100 år, ble det kun registrert to fremmede arter, ingen av dem nye for Norge (Husa mfl. 2014). Arter som kommer inn fra nordlige deler av USA, Canada og Alaska og nordlige deler av Kina vil ha størst etableringspotensiale i nordområdene på fastlandet og på Svalbard.
Områdene nord for Trøndelag er i liten grad undersøkt, noe som reflekteres i at få arter er registrert. En generell kartlegging av fremmede arter i småbåthavner er kun gjennomført fra svenskegrensen til Sognefjorden, samt ett område på søre Sunnmøre (Husa mfl. 2022).
Figur 4. Geografisk utbredelse av fremmede marine arter i Norge, summert per fylke, pluss Svalbard og Jan Mayen. Figuren viser alle arter med etableringskategori C1, “overlever i naturen”, N = 39 (til venstre), og etableringskategori E “sprer seg, overlever og reproduserer flere steder”, N=27 (til høyre) (data fra Sandvik m. fl. 2019, figur fra Husa mfl. 2022).
En vurdering av vektorer for introduksjon av fremmede marine arter er gitt i Husa mfl. 2022. Vektorer som ballastvann, utsetting av arter for akvakultur eller økte matressurser, og import av levende sjømat er i stor grad regulert i Norge i dag og anses derfor som mindre relevante vektorer. Import av fremmede arter til private saltvannsakvarier er det vanskelig å få oversikt over, da mye foregår som netthandel. De fremmede artene som selges til slike akvarier fra butikk er hovedsakelig tropiske eller subtropiske og vil ha liten sannsynlighet for overlevelse i norsk natur. Da det også er liten sannsynlighet for at de havner i norsk natur har vi totalt sett vurdert import av fremmede arter til saltvannsakvarier til å ha liten risiko over hele landet. Arter som kommer med marint flytende søppel er ikke undersøkt i Norge, trolig vil sannsynligheten for at fremmede arter kommer seg til Norge som haikere på flytende søppel være størst i Skagerrak og på Vestlandet, da søppel som kommer inn her stammer fra Sverige, Danmark og Nordsjøen (Husa mfl. 2022). Vi vurderer derfor sannsynligheten for introduksjon av fremmede arter til å være høy i sørvestlige områder og moderat til liten i resten av landet.
Transport av rensefisk (leppefisk) fra Sverige kan føre med seg patogener, men også andre mikroorganismer, larver av fisk, evertebrater og sporer fra alger i transportvannet. Leppefisken fra Sverige havner i stor grad i Trøndelag da det er lite tilgang på lokalfanget leppefisk i disse områdene. Havforskningsinstituttet vurderer i sin risikovurdering av bruk av villfanget leppefisk i fiskeoppdrett at sannsynligheten for smittespredning ved transport er høy for områdene fra Lista til Nordland med dagens manglende krav til behandling av både transportmiddel og transportvann (Grefsrud mfl. 2021).
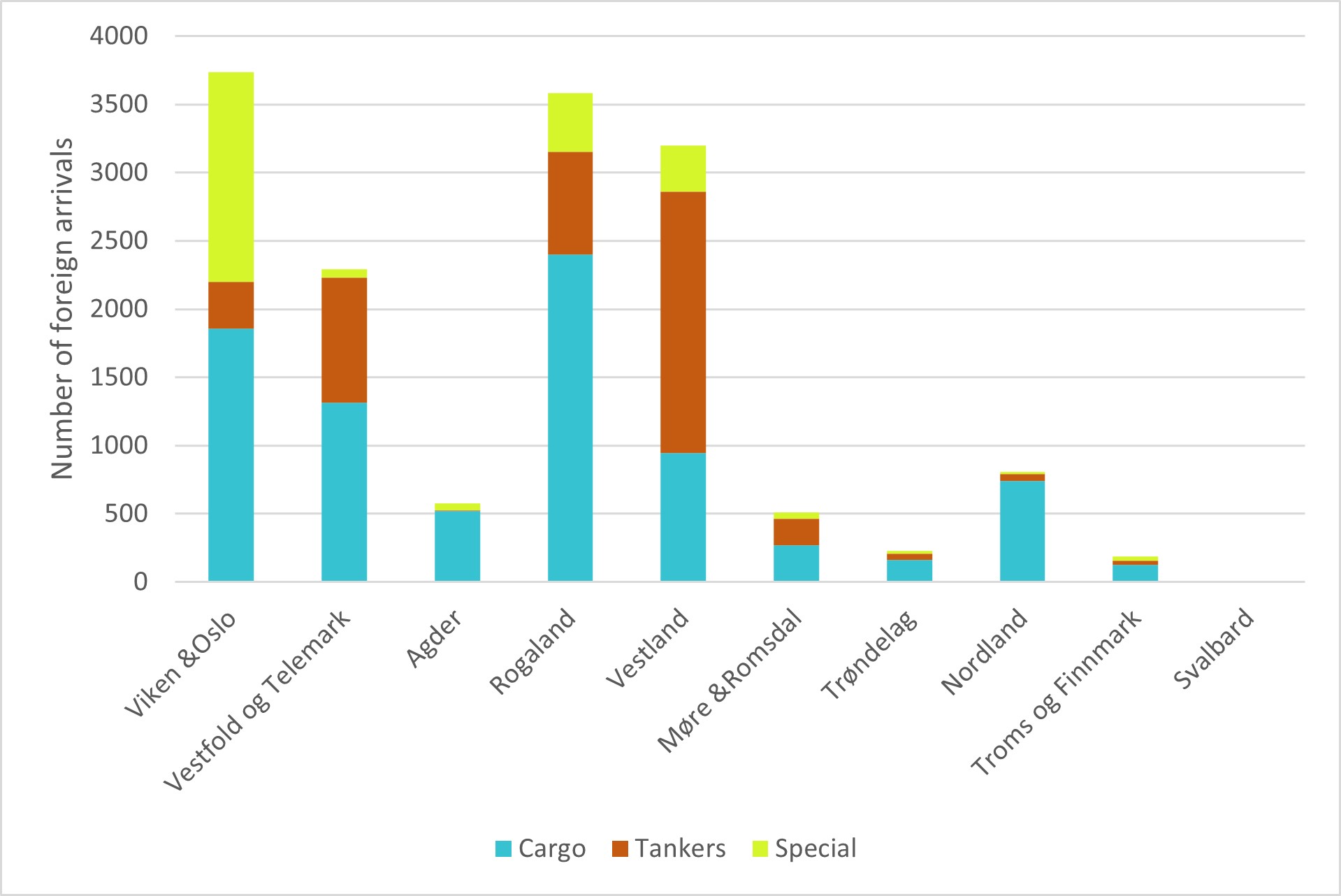
Begroing på skip er vurdert som den vektoren med høyest risiko for å medføre introduksjoner av fremmede marine arter (Husa mfl. 2022). Fylker med mange havner med skipsanløp fra fremmede havner har høyest risiko for at nye arter skal introduseres, men temperatur i donor og mottaker region spiller også en stor rolle for nye arters mulighet for å etablere seg.
Figur 5. Årlige ankomster av fartøy fra utenlandske havner (estimert for havner med >500 årlige ankomster i gruppen) I kategoriene lasteskip, tankskip og spesialfartøy i norske fylker (Data fra Marine Traffic.com, Figur fra Husa mfl. 2022).
Fylkene rundt Oslofjorden; Viken, Oslo, Vestfold & Telemark, samt Vestland og Rogaland fylke har mellom 3800 og 2300 årlige anløp fra utlandet og må defineres som områder med høy risiko for introduksjon av nye arter. De øvrige fylkene har mindre enn 1000 årlige anløp og defineres som områder med moderat til lav risiko (Figur 5).
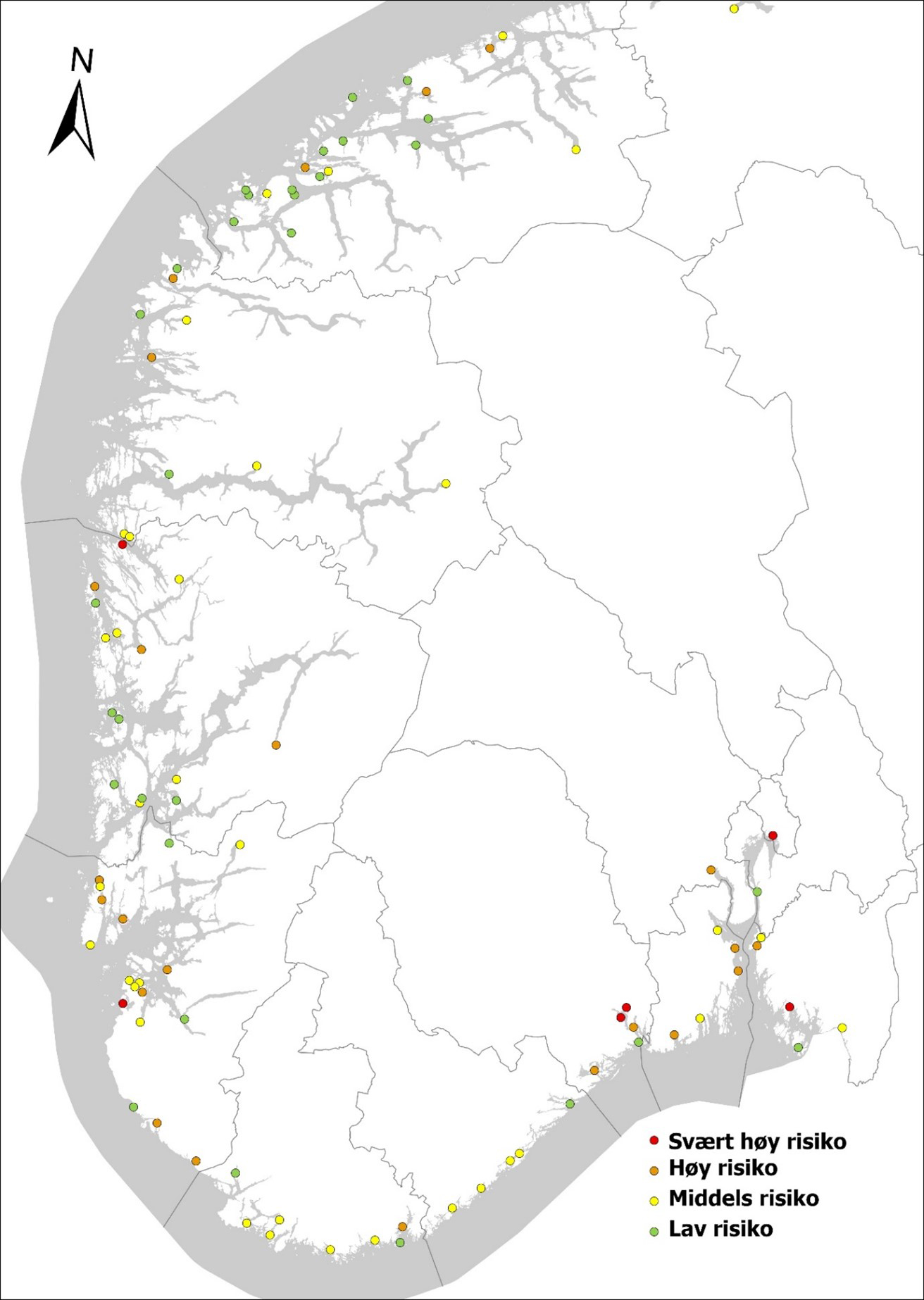
Tabell 2 gir en oversikt over de enkelte havnenes årlige anløp fra utlandet i de tre fartøyskategoriene spesialfartøy, tankskip og lasteskip. Vi har her angitt en risikokategori for havnene slik: Havner med ≤ 10 utenlandske anløp = lav risiko, havner som har > 10 og ≤ 100 utenlandske anløp = moderat risiko, havner som har > 100 og ≤ 500 utenlandske anløp = høy risiko, havner med > 500 utenlandske anløp har svært høy risiko.
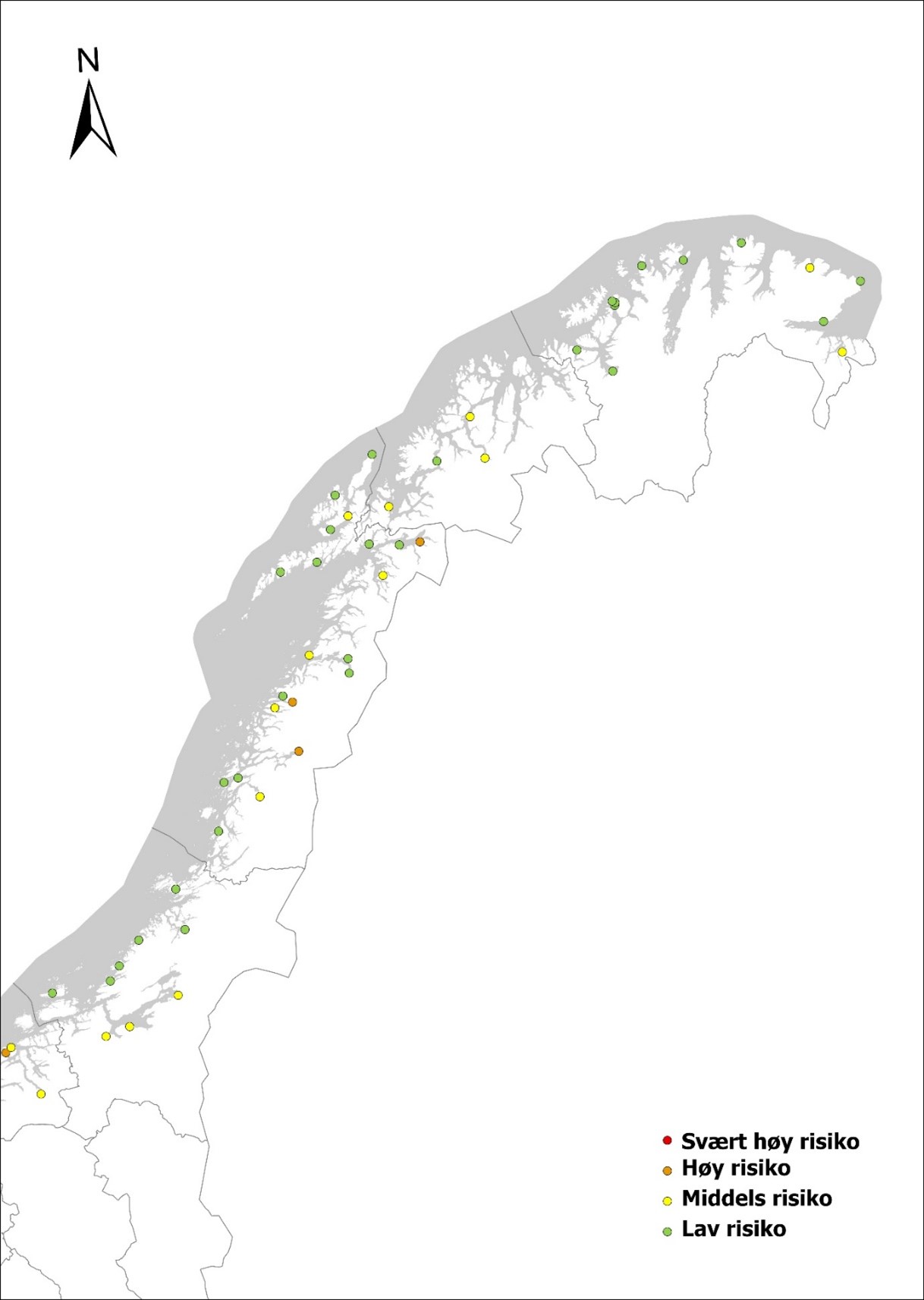
Noen havner har svært mange utenlandske anløp og har svært høy risiko slik som Mongstad som mottar mange utenlandske tankskip, industrihavnene Rafnes og Porsgrunn, Tananger som har høy aktivitet knyttet til oljeindustrien, samt Oslo og Fredrikstad. Fylkene rundt Oslofjorden har i tillegg syv havner i høy risiko. Agder har kun en havn som er i høy risiko, Kristiansand, men også Farsund bør anses som høy risiko da det er en opplagshavn for fartøy uten kontrakt. Rogaland har syv havner med høy risiko og Vestland har fem (Tabell 2, Figur 6). Fylkene nord for Vestland anses å ha lav til moderat risiko for området som helhet, men har noen havner med høy risiko som bør overvåkes. For Møre & Romsdal gjelder dette Ålesund, Elnesvågen og Averøya (Tabell 2, Figur 6), for Nordland Mo i Rana, Glomfjord og Narvik (Tabell 2, Figur 7). For Trøndelag, Nordland, Troms og Finnmark finnes også noen havner med moderat risiko slik som Trondheim, Orkanger og Verdal, Bergsneset, Harstad, Tromsø, Båtsfjord og Kirkenes som også bør overvåkes, om enn med litt lavere frekvens enn høyrisikohavner (Tabell 2, Figur 7). Mye av den utenlandske trafikken nord for Lofoten kommer fra havner i Nord-Russland. Marine arter fra Russland utgjør sannsynligvis en mindre trussel da flora, og fauna i stor grad vil være den samme. Dersom fartøyene kommer gjennom Nordøstpassasjen fra Stillehavet, anses risikoen for introduksjon av fremmede arter å være stor. Trafikken gjennom denne passasjen har vært økende og satt rekord med 62 passeringer i 2020, hovedsakelig av skip fra Kina.
Svalbard har bare et lite antall utenlandske fartøy, men flere anløp fra fastlandet. Statistikken for Svalbard (Husa mfl. 2022) bygger på data fra 2020-2021. Svalbard har hatt forsterkede restriksjoner for innreise på grunn av Covid-19 i denne perioden og man må anta at i et normalt år vil både passasjer og fritidstrafikken fra utenlandske havner være langt større. Havner på Svalbard bør likevel undersøkes med regelmessige mellomrom, da flora og fauna på Svalbard regnes å være ekstra sårbar.
Ferger som går til og fra Sverige, Danmark og Tyskland fra Tananger (Stavanger), Kristiansand, Larvik, Sandefjord og Oslo er ikke inkludert i tabell 2, men vil gi en ekstra risiko for disse havnene. Cruisetrafikken har også vært redusert i analyseperioden 2020-2021 på grunn av Covid-19 og derfor er data om cruisetrafikken hentet ut for 2019. Bergen er den travleste cruisehavnen i Norge, fulgt av Geiranger, Stavanger, Ålesund, Flåm, Tromsø, Nordkapp og Oslo. Detaljer om fergetrafikk og cruisetrafikk samt fiskebåter og fritidsfartøy er gitt i Husa mfl. 2022. Fergehavner og cruisehavner anses å ha høy risiko for introduserte arter og bør overvåkes.
Lasteskip og tankskip går gjerne fra havn til havn uten opphold og ligger ikke lenger i havn enn det tar å laste/losse fartøyet. Gruppen spesialskip kan være slepebåter, kranfartøy, løfteskip, lektere, ankerhåndteringsfartøy, supplybåter, boreskip og borerigger. Disse ligger ofte lenger i ro i en havn der det er arbeid som skal utføres og utgjør kanskje en større risiko for å ha med seg fremmede arter som de av begroing. Mange av dem er også for store til å ta i tørrdokk og kan på grunn av konstruksjonen være vanskelige å gjøre reine.
Risikoen for introduksjon av fremmede marine arter er ikke bare høy i havner med mange anløp fra utlandet, men kan også være høy i områder med relativt få ankomster, men der fartøyene som ankommer har oppholdt seg lenge et sted i havn eller på oljefelt i utlandet for deretter å bli liggende lenge på norskekysten, slik som opplagsplasser, skipsverft og opphuggingsverksted. Disse områdene bør også undersøkes. Slike områder kan identifiseres i hvert fylke fra opplagsregisteret og Marine Traffic.com.
Tabell 2. Årlige utenlandske anløp i kategoriene spesialfartøy, tankskip og lasteskip havner i hvert fylke (estimert for havner med >500 årlige ankomster I gruppen). Risikokategori er angitt slik: Havner med ≤ 10 utenlandske anløp= lav risiko, havner som har > 10 og ≤ 100 utenlandske anløp = moderat risiko, havner som har >100 og ≤500 utenlandske anløp= høy risiko, havner med > 500 utenlandske anløp har svært høy risiko (basert på data fra Marine Traffic, sammenstilt i Husa mfl. 2022).
Spesialfartøy
Tankskip
Lasteskip
Totalt anløp fra utlandet
Risiko
Oslo & Viken
Drammen
6
50
250
306
H
Drøbak
0
0
0
0
L
Kambo
3
1
46
50
M
Moss
0
2
100
102
H
Fredrikstad
6
206
520
732
SH
Hvaler
10
0
0
10
L
Halden
5
2
18
25
M
Oslo
1506
82
925
2513
SH
Vestfold & Telemark
Kragerø
11
0
153
164
H
Langesund
0
0
0
0
L
Brevik
112
1
252
365
H
Rafnes
0
434
100
534
SH
Porsgrunn
4
180
550
734
SH
Larvik
2
24
90
116
H
Sandefjord
1
79
0
80
M
Slagen
23
196
0
219
H
Horten
2
1
125
128
H
Holmestrand
0
0
30
30
M
Agder
Flekkefjord
3
0
3
6
L
Farsund
11
8
44
63
M
Hausvik
0
0
50
50
M
Lyngdal
3
0
15
18
M
Mandal
6
0
29
35
M
Høllen
0
0
10
10
M
Flekkerøy
0
0
0
0
L
Kristiansand
16
0
203
219
H
Lillesand
0
0
15
15
M
Grimstad
1
0
11
12
M
Arendal
4
0
56
60
M
Eydehavn
3
0
82
85
M
Risør
3
0
0
3
L
Rogaland
Ølen
2
0
5
7
L
Haugesund
94
73
547
714
H
Storasund
3
0
22
25
M
Husøya
22
42
285
349
H
Skudeneshavn
0
0
17
17
M
Kårstø
0
457
0
457
H
Sauda
0
0
94
94
M
Breiviken
0
0
269
269
H
Forsand
0
0
0
0
L
Sandnes
1
0
71
72
M
Stavanger
102
60
89
251
H
Dusavik
38
20
14
72
M
Mekjarvik
27
1
18
46
M
Tananger
109
73
530
712
SH
Sirevåg
0
0
8
8
L
Egersund
4
9
142
155
H
Jøssingfjord
1
0
105
106
H
Vestland
Hanøytangen
6
0
8
14
M
Ågotnes
16
1
45
62
M
Skipavik
6
0
49
55
M
Mongstad
47
1642
26
1715
SH
Sture
0
183
0
183
H
Kolsnes
0
0
3
3
L
Eikefet
0
2
64
66
M
Bergen
155
2
47
204
H
Storabø
2
0
5
7
L
Torangsvåg
1
0
3
4
L
Rubbestadneset
2
2
1
5
L
Stord havn
6
0
26
32
M
Leirvik
4
0
1
5
L
Odda
0
11
152
163
H
Husnes
0
0
15
15
M
Høylandsbygd
3
0
0
3
L
Sløvåg
5
6
87
98
M
Leirvik Hyllestad
0
0
2
2
L
Høyanger
2
0
26
28
M
Årdalstangen
0
0
42
42
M
Florø
56
52
195
303
H
Svelgen
0
6
42
48
M
Kalvåg
0
0
0
0
L
Måløy
25
6
89
120
H
Raudeberg
2
0
2
4
L
Møre & Romsdal
Larsnes
0
0
0
0
L
Mjølstadneset
2
0
1
3
L
Fosnavåg
1
0
0
1
L
Ulsteinvik
7
0
7
14
M
Hjørungavåg
0
0
3
3
L
Hareid
4
0
1
5
L
Ørsta
2
0
2
4
L
Fiskarstrand
2
0
1
3
L
Spjelkavik
5
15
6
26
M
Ålesund
15
0
90
105
H
Søvik
0
0
2
2
L
Steinshamn
0
0
0
0
L
Brattvåg
0
0
1
1
L
Vestnes
3
0
2
5
L
Molde
1
0
3
4
L
Elnesvågen
0
150
4
154
H
Harøysundet
0
0
3
3
L
Averøya
2
13
92
107
H
Sunnalsøra
0
0
37
37
M
Kristiansund
0
16
15
31
M
Trøndelag
Nordskaget
0
2
0
2
L
Orkanger
2
0
71
73
M
Trondheim
3
41
35
79
M
Verdal
12
1
56
69
M
Lysøysund
1
1
0
2
L
Kjerkeholmen
0
0
0
0
L
Sandviksberget
0
0
0
0
L
Namsos
0
0
0
0
L
Rørvik
3
0
0
3
L
Nordland
Brønnøysund
3
0
4
7
L
Herøy
0
0
0
0
L
Sandnessjøen
0
3
0
3
L
Mosjøen
0
22
71
93
M
Mo i Rana
2
0
246
248
H
Halsa Meløy
0
2
45
47
M
Glomfjord
0
12
104
116
H
Ørnes
0
0
0
0
L
Fauske
0
0
0
0
L
Rognan
1
0
4
5
L
Bodø
10
11
0
21
M
Kjøpsvik
0
0
28
28
M
Hekkelstrand
0
0
2
2
L
Narvik
0
0
196
196
H
Lødingen
0
0
2
2
L
Svolvær
0
0
0
0
L
Leknes
0
0
1
1
L
Stokkmarknes
0
1
10
11
L
Sortland
0
0
20
20
M
Myre
0
0
5
5
L
Andenes
0
1
0
1
L
Troms & Finnmark
Bergneset
0
0
11
11
M
Harstad
6
7
4
17
M
Finnsnes
0
0
0
0
L
Tromsø
0
12
22
34
M
Øksfjord
0
0
0
0
L
Alta
0
4
0
4
L
Rypefjord
0
4
2
6
L
Hammerfest
4
0
0
4
L
Melkeøya
0
2
0
2
L
Havøysund
0
0
0
0
L
Honningsvåg
0
0
0
0
L
Mehamn
0
0
0
0
L
Båtsfjord
0
0
45
45
M
Vardø
0
0
1
1
L
Vadsø
5
0
0
5
L
Kirkenes
17
1
41
59
M
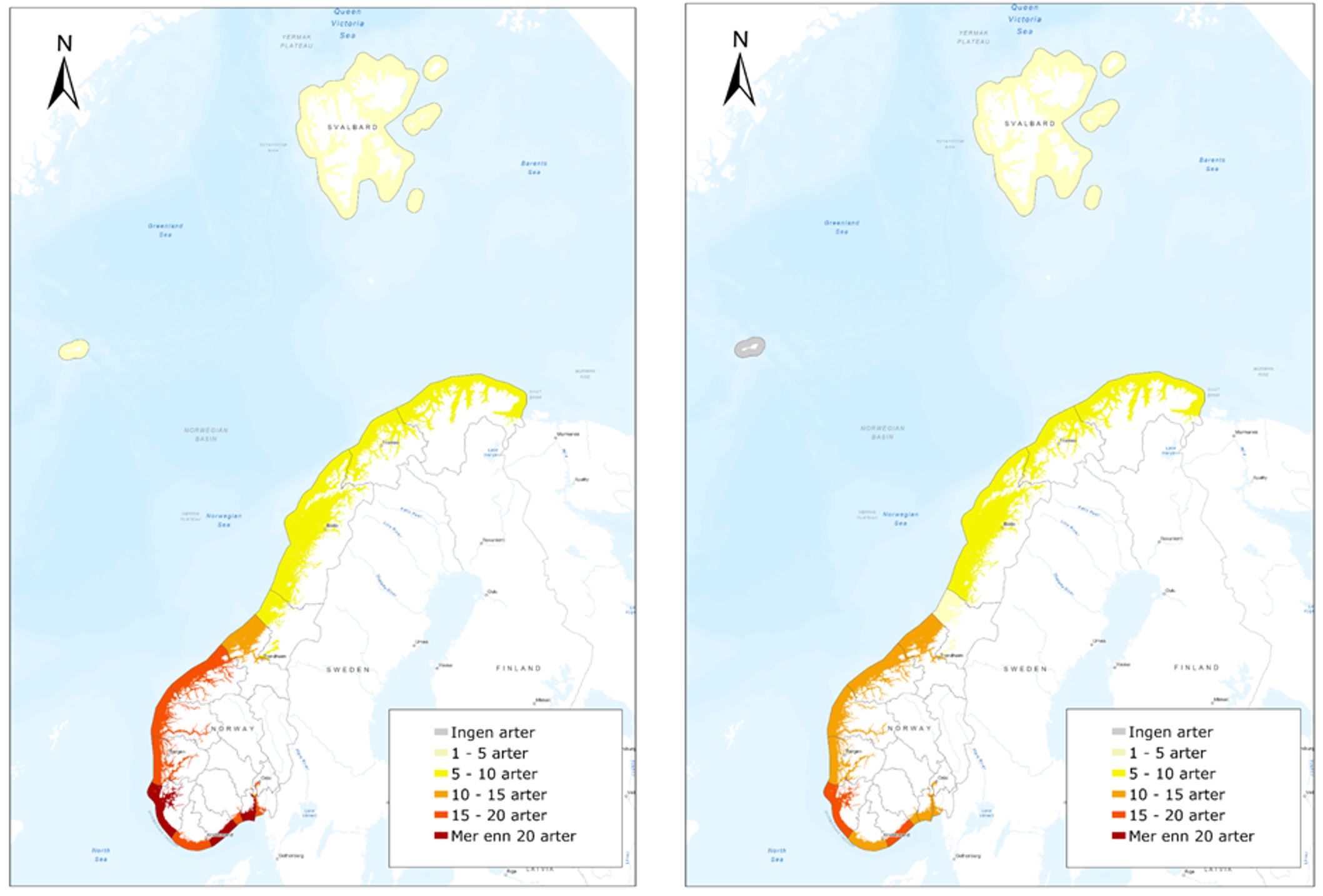
Figur 6. Oversiktskart over norske havner med en risikovurdering basert på årlige anløp av båter fra utenlandske havner i kategoriene lasteskip, tankskip og spesialfartøy (estimert for havner med > 500 årlige ankomster). Risikokategori er angitt slik: Havner med ≤ 10 utenlandske anløp= lav risiko, havner som har > 10 og ≤ 100 utenlandske anløp = moderat risiko, havner som har > 100 og ≤ 500 utenlandske anløp = høy risiko, havner med > 500 utenlandske anløp har svært høy risiko (basert på data fra Marine Traffic, sammenstilt i Husa mfl. 2022).Figur 7. Oversiktskart over norske havner med en risikovurdering basert på årlige anløp av båter fra utenlandske havner i kategoriene lasteskip, tankskip og spesialfartøy (estimert for havner med > 500 årlige ankomster). Risikokategori er angitt slik: Havner med ≤ 10 utenlandske anløp = lav risiko, havner som har > 10 og ≤ 100 utenlandske anløp = moderat risiko, havner som har > 100 og ≤ 500 utenlandske anløp = høy risiko, havner med > 500 utenlandske anløp har svært høy risiko (basert på data fra Marine Traffic, sammenstilt i Husa mfl. 2022).
4 - Internasjonal metodikk for kartlegging og overvåkning av fremmede marine arter
Siden 90-tallet har det blitt utarbeidet og utprøvd en rekke forskjellige måter å gjennomføre systematiske/standardiserte undersøkelser av forekomst av fremmede arter. Metodene varierer betydelig i omfang, hvilke organismegrupper de detekterer og hvilke de “overser”, men de fleste metodene har noen felles problemer som for eksempel: tilgjengelig taksonomisk ekspertise og ofte manglende bakgrunnsstudier.
Tidlig deteksjon av fremmede arter er utfordrende fordi tettheten av artene i begynnelsen ofte er lav. Dette har konsekvenser for den innsatsen, og derved kostnadene, en slik overvåkning bør ha.
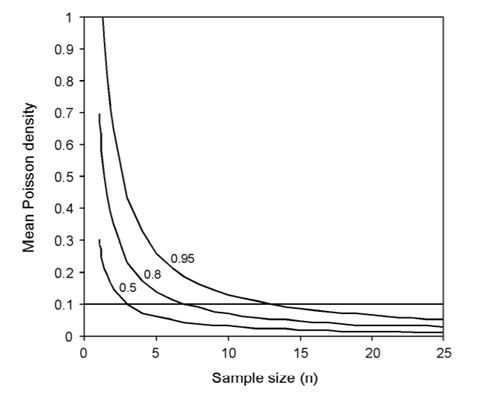
Hewitt & Martin (2001) skriver: “A power analysis to determine the appropriate sampling effort, using the method of Green and Young (1993) for rare species with a Poisson-distribution, suggests that a sample size of approximately 13 samples will be necessary to detect a species with a mean Poisson density of 0.1 individuals per sample unit at 95% probability” (Figur 8).
Figur 8. Styrkeanalyse av antall prøver (for eksempel kvadranter) som må tas for å detektere Poissonfordelte sjeldne arter med varierende midlere tetthet i kvadrantene ved tre forskjellige grader av styrke (1-β): 0,95, 0,8, 0,5. Den horisontale linjen representerer en tilfeldig valgt terskel for sjeldenhet. (Hewitt & Martin, 2001)
Analyse av styrke er viktig fordi det omhandler “type II feil”, - sannsynligheten for å ikke forkaste en feilaktig 0-hypotese. I kartlegging og overvåking av fremmede arter vil 0-hypoteser ha sin generelle form: “I lokasjonen er det ingen NIS-arter" / “I lokasjonen finnes ikke art xxx”. Som en ser av Figur 8: Hvis en har en fremmed art med midlere Poisson-fordelt tetthet på 0,1, vil en ved analyse av 5 prøver ha mellom 20 og 50 % sjanse (hhv 1-0,80 og 1-0,5) for ikke å forkaste 0-hypotesen når den er feil.
Skal en ha ambisjoner om å oppdage alle fremmede arter i f.eks. en havn eller marina, må en undersøke alle tilgjengelige habitater (Hewitt & Martin, 2001). Slike undersøkelser vil også være begrenset av vitenskapelige og logistiske faktorer, og ressursene som trengs for å gjennomføre dette vil neppe være/bli tilgjengelig verken for enkelthavner, andre typer lokaliteter eller i nasjonal skala (Hewitt and Martin, 2001). Med denne erkjennelsen er det rasjonelt å konsentrere undersøkelsene om de habitatene hvor det er størst sannsynlighet for å finne fremmede organismer.
4.1 - CRIMP (Centre for Research on Introduced Marine Pests): Revised Protocols for Baseline Port Surveys for introduced Marine Species (AUSTRALIA)
Australia har utviklet en metodikk for undersøkelser av fremmede marine arter i havner. Metodikken er prøvd ut i en rekke havner og en revidert protokoll for slike undersøkelser er publisert (Hewitt & Martin 2001). Denne metoden er omfattende, men gir også gode baselinjedata for havnene. Metodikken baserer seg i stor grad på bruk av dykkere, men gir muligheter for tilpasninger slik at man kan bruke grabb, slede/skrape og videotransekter i stedet for innsamling ved hjelp av dykkere.
4.1.1 - Oppsummering av protokoll
Det skal samles inn data på temperatur, saltholdighet og turbiditet på hver stasjon og andre metadata skal noteres. Det anbefales at hver havn undersøkes hvert 3-5 år. Mer detaljert protokoll og anbefaling om utstyr, fiksering og opparbeiding av prøver finnes i Hewitt & Martin (2001).
Planteplankton: Tre vertikale og ett horisontalt planktontrekk per stasjon anbefales. Prøvene tas tilbake til lab for kultivering og det skal sjekkes for utvikling av giftige alger med regelmessig frekvens. I tillegg skal det samles inn sedimentkjerner på bløtbunn, der hvilecyster av planteplankton skal dyrkes i lab og sjekkes for utvikling av giftige alger.
Zooplankton: Samles inn ved bruk av senkenett fra overflaten til omtrent en halv meter over bunn, en prøve per stasjon.
Mobile dyr på hardbunn/bløtbunn: Samles inn ved hjelp av tre krabbeteiner og tre reketeiner per stasjon. Teinene skal stå ute i omtrent 12 timer over natten.
Giftstasjoner: Små kutlinger og annen fisk samles inn ved hjelp av dykkere som har egnet gift (f. eks rotenon) i en plastpose som slippes ut mellom brygger og ved moloer. Bedøvet og død fisk samles opp av dykkere eller med håv fra land.
Visuelle søk: Dykkere foretar visuelle søk ved så svømme på tre ulike dyp langs kaier og moloer for å se etter og samle inn større arter.
Kaier og pilarer: Alle fortøyningsplasser i en havn undersøkes på med visuelle søk. Tre steder på hver kai merkes og det legges ut et vertikalt transekt som filmes og det tas stillbilder fra forankrede (eller holdte av dykker) kvadrater på 0,1 m2 på tre ulike dyp (0,5, 3 og 7 meters dyp). Dykkere skraper av og samler inn materiale i kvadratene som tas med tilbake til lab, sorteres og identifiseres.
Moloer, steinvegger og svaberg: Undersøkes på samme måte i vertikale transekt med 10-15 meters avstand
Bløtbunn: Undersøkes på samme måte med 50 meters transekter som legges perpendikulært ut fra bryggepilarer og/eller kaier. I tillegg til videofilming, stillbilder og innsamling av biota i kvadrater, skal det tas kjerneprøver til analyser av hvilecyster av planteplankton og dyr i sedimentet. Det skal også tas prøver til analyse av kornstørrelse og innhold av organisk karbon i sedimentet. På bløtbunn skal det også fiskes med strandnot på 25 meters lengde.
Fjæresone: Undersøkes visuelt samt innsamling i 0,1 m kvadrater, men har lav prioritet.
Småbåthavner: Undersøkes på samme måte, samt akvakulturanlegg og skipsvrak hvis det finnes i eller nær havnen.
4.2 - SERC-Protokoll for NIS (USA)
SERC (Smithsonian Environmental Research Center) har blant annet fokus på fremmede arter https://serc.si.edu/research/research-topics/biological-invasions, og har gjennom flere tiår utviklet metoder som kan benyttes til å detektere fremmede marine- og brakkvannsarter. De benytter ofte en enkel, standardisert metode som baserer seg på utplasserte begroingsplater (14 x 14 cm grå/mørk PVC), som monteres horisontalt 1 m over bunnen. Dette gir en lyseksponert side som favoriserer alger, og en skyggeside som favoriserer dyr. Platene er mattet med sandpapir. I en studie ble slike begroingsplaters evne til å fange opp fremmede arter sammenlignet med innsamlinger ved hjelp av dykker i like store kvadrater på brygger. Det viste seg at begge metoder var like gode både med hensyn til generell diversitet og antall fremmede arter. Denne metoden egner seg godt til å kartlegge påvekstorganismer som vanligvis slår seg ned på brygger/kunstig substrat, men fanger ikke opp arter som har spesielle krav til substrat/habitat slik som bløtbunnsorganismer og pelagiske arter (Marrafini mfl. 2017).
4.3 - HELCOM (land rundt Østersjøen)
HELCOM PROTOKOLLEN (det er utviklet en rekke protokoller for spesifikke habitattyper og analysemetoder). Metodikken bygger på CRIMP protokollen, men har byttet ut den utstrakte bruken av dykkere med håndholdt utstyr fra land. Metodene er tatt i bruk i en rekke land, men har ofte nasjonale og lokale tilpasninger. Danmark har utviklet et nasjonalt program basert på HELCOM-protokollen, med blant annet bruk av miljø-DNA (Andersen mfl. 2016, 2017, 2018), det samme har Finland (HELCOM 2013). Metodikken er også testet ut i Nederland. Figur 9 gir en samlet oversikt over metodikk og prøvetakingssteder.
4.3.1 - Oppsummering av protokoll (HELCOM 2013)
Da alle havner er ulike bør man bruke en stratifisert prøvetakningsdesign slik at alle ulike typer habitat/miljø blir inkludert. Som et minimum bør tre stasjoner i hver havn undersøkes. I store havner og i havner med mange ulike miljø bør antall stasjoner økes. Det er ikke publisert noe mål på hva som er nok prøvetaking i forhold til størrelsen på havnen, noe som også etterlyses av Kabuta (2014). Det er utarbeidet en liste med prioriterte områder i havner som bør undersøkes, basert på Hewitt og Martin (2001). Planlegging av undersøkelsen skal bygge på kjennskap til aktiviteter i havnen og områder med høy risiko skal prioriteres. Alle typer hardt substrat i havnen skal undersøkes med minst tre prøver fra hvert sted. Alle typer bløtbunn i havnen skal undersøkes med et minimum av tre prøver. Protokollen anbefaler gjenbesøk i havner basert på risiko med intervaller på 1-5 år. Videre anbefales det at man samler inn prøver av biota på hardbunn og bløtbunn i perioden sein juli til september. Planteplanktonprøver tas under vårblomstringen (februar-april) og nok en gang seint på sommeren. Begroingspanel kan settes ut når man tar de første prøvene og samles inn igjen seint på sommeren i forbindelse med resten av undersøkelsene.
Vannmiljø: For hver lokalitet skal som et minimum følgende miljødata registreres: temperatur, salinitet, turbiditet og klorofyll-a (se underkapittel om fytoplankton). En kan med fordel benytte en nedsenkbar logger (CTD) som logger både dyp, temperatur, turbiditet, klorofyll-a og salinitet.
Patogener: En vannprøve på 500 ml tas fra ca. 30 cm dyp på hver lokalitet. Prøvetakingen skal følge retningslinjene gitt i EUs Direktiv for badevann 2006/7/EC. Prøvedyp, vanndyp og annen relevant informasjon noteres på feltskjemaet. Dersom det finnes prøvetakingsprogram utført av lokale myndigheter i havnene er dette tilstrekkelig.
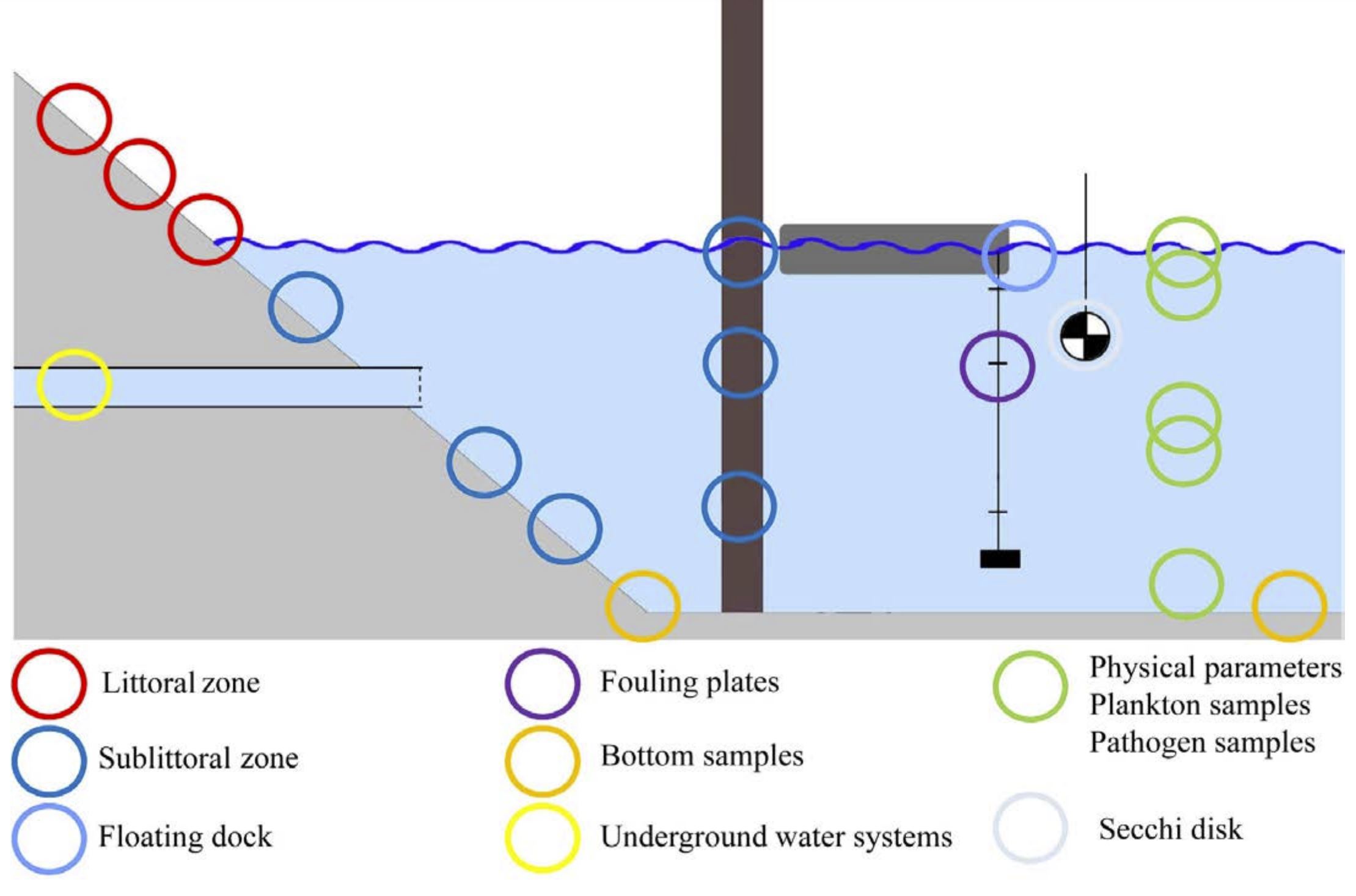
Figur 9. Skjematisk fremstilling av vanlige prøvetakingssteder ifølge HELCOM-protokoller. (Figur fra Kabuta mfl. 2014
Dyr i sediment: Bunnprøver kan tas med liten grabb som kan opereres fra kai/brygge. Temperatur, salinitet og oksygenmetning ved bunnen kan måles med nedsenkbar datalogger/CTD. Det bør tas minst 3 grabbprøver fra hver lokalitet med minst 15 m avstand. Sedimentkvalitet vurderes visuelt, og grabben bør ha nådd til minst 10 cm dyp for å få en tilfredsstillende prøve. Analysene gjøres i henhold til HELCOM COMBINE Manual Annex C-8: Soft bottom Macrozoobenthos.
Mobil epifauna, som for eksempel fisk og krepsdyr, kan fanges ved hjelp av ulike fangstmetoder. I HELCOM protokollen for havneundersøkelser (HELCOM 2013) anbefales det å bruke to typer feller (krabbeteiner og ørekyteteiner (minnow-traps) for å fange dyr av ulik størrelse. Som et forslag nevnes teiner med mål 63 cm x 42 cm x 20 cm, med 1,3 cm nettstørrelse, samt ørekyteteiner 42 cm lengde, 23 cm bredde, 6,4 mm nett og 2,5 cm åpning. Fellene bør sikres til fast struktur og merkes med blåse. Selv om feller er selektive fangstredskap så kan de gi informasjon om relativ forekomst. Man bør være oppmerksom på om det er variasjon i bunnsubstrat på lokaliteten slik at man dekker opp ulike typer bunn ved plassering av fellene. Fellene skal agnes med passende agn og ligge ute minimum 48 timer. Selv om det ikke er nevnt, så bør man også sjekke fellene tidligere, både med tanke på dyrevelferd og for å redusere sjansen for at rovdyr spiser opp byttedyr. Hvis den som håndterer fellene har god artskunnskap kan artsbestemmelse til stor del gjøres i felt, men med sikring av materialet i kjølebag og senere fryser for mulig verifisering / identifisering etterpå. Man bør også ta foto. Dessuten må man huske å notere fangst per tidsenhet per felle.
HELCOM-protokollen viser til at andre fangstmetoder enn feller ofte kan være umulige å bruke i havneområder. Andre fangstmetoder kan brukes i stedet for eller sammen med feller og kan for eksempel være en god måte for å fange fremmede fiskearter utenfor havner. På grunt vann kan fiske med stang, ruser, garn, kilenot og strandnot være effektivt. For fiske med stang finnes det stort potensiale for folkeforskning og involvering av artsfiskermiljøet i Norge. Som nevnt har enkelte funn av nye fremmede fisker blitt gjort på den måten (Hansen & Karlsbakk 2018). For dypvannsfisker og fisker lenger til havs er det mindre sjanse for å oppdage nye fremmede fiskearter. Det er mulig at fremmede arter kan fanges og dermed oppdages ved ulike typer tokt og tråling. Men for fisk så forekommer mesteparten av potensielle dørstokkarter på relativt grunt vann i kystsonen (Figur 4, Tabell 1). Visuell observasjon av epifauna ved hjelp av snorkling, dykking eller videoundersøkelser kan være et alternativ for enkelte arter i utvalgte områder. Nattdykk etter fisk er tatt med som metode i den danske protokollen (Andersen mfl. 2017). Akustiske overvåkingsmetoder kan også være en mulighet hvis arten man er ute etter er lager lyder (Rosten & Fossøy 2020).
Fastsittende organismer på hardbunn/kunstig substrat: Pilarer/stolper eller andre faste strukturer er regnet som høyprioritetsområder i CRIMP-protokollen (Hewitt & Martin, 2001). Minst tre prøver bør derfor tas fra slike strukturer på hver lokalitet. Lokalitetene bør være minst 10 m fra enden av en kai, for å unngå kant-effekter. En kan gjerne legge lokalitetene 10 –15 m fra hverandre. Hvis det ligger vrak i havnen er disse ofte “hot-spots” for fremmede arter, og kan med fordel undersøkes.
Hvis det er “hengende strukturer som kan løftes opp, kan prøver enkelt tas “tørt”. En søker ellers å skrape av begroingen på en ca. 10 x 10 cm rute for nærmere analyse. Hvis mulig tas det bilde av begroingen før en skraper av. Hvis det er mulig å dykke /snorkle, kan en ta prøver fra større dyp. Hvis ikke, vil prøver fra ned til 0,5 m (så langt en kan nå med armene fra overflaten) benyttes. (https://helcom.fi/wp-content/uploads/2021/06/HELCOM-Monitoring-guidelines-for-marinas.pdf).
Innsamling av fastsittende arter bygger her på CRIMP protokollen, men en ønsker å unngå dykking. Mange kaier er høye og bygd på høye pilarer noe som gjør at det er vanskelig å nå ned og få tatt prøver. HELCOM anbefaler derfor begroingsplater slik at man får prøver fra ulike dyp uten dykking. Begroingsplater som settes ut, og så hentes tilbake etter en angitt tid, har evne til å fange opp fastsittende organismer når de er i spredningsfasen. I HELCOM-protokollen benyttes 15 x 15 cm sandpapir-mattede PVC-plater. Disse settes ut på tre dyp for hver stasjon, minst 1,5 måned, fortrinnsvis i tidsrommet april-september (Figur 10, https://helcom.fi/wp-content/uploads/2021/06/HELCOM-Monitoring-guidelines-of-mobile-and-sessile-epifauna.pdf ). Kabuto (2014) anbefaler at man bruker 14 X 14 cm plater, analyserer hver side for seg og ikke henger alle platene fra samme tau. Effektiviteten av begroingsplater er undersøkt av blant annet (Marraffini mfl. 2017) og av (Tamburini mfl. 2021). Begge fant at slike plater er velegnet til å fange opp påvekstorganismer på hardbunn. Outinen mfl. (2021) sammenliknet skraping av begroing på faste strukturer med begroingsplater. De fant at begroingsplater var mer effektive enn skraping av andre strukturer både for å detektere fremmede arter og generell artsrikdom og det var lettere å gjøre tetthetsberegning av påveksten på standardiserte plater.
Figur 10. Rigging av begroingsplater, og eksempler på påvekst etter noen måneder. Figur fra HELCOM 2013.
I Europa finnes en rekke faste ruter mellom to havner (typisk fergeruter). Ballastvannkonvensjonen har åpning for å gi fritak for ballastvannbehandling for skip i slike ruter under bestemte forutsetninger (IMO Resolution MEPC. 162(56)). Fritak kan gjøres av havnestatenes myndigheter og er kun gyldig for disse havnestatenes jurisdiksjon. Havnestatene plikter også å varsle eventuelle tilgrensende havnestater om eventuell tillatelse. Mange land rundt Nordsjøen har høy konnektivitet og tilnærmet samme biota, men kan likevel ha betydelige forskjeller i forekomst av fremmede arter.
Det er derfor nedfelt krav til operatørene som skal søke om fritak. Fritak kan kun gis etter en risikovurdering som er nærmere beskrevet i: “Guidelines for risk assessment under regulation A-4 of the BWM Convention (G7).” Risikoanalysen fordrer ferske havneundersøkelser av de respektive havnene for å forsikre om at det er de samme fremmede artene i begge havnene. Unntak fra ballastvannreglene skal gjennomgås i perioder fra 12-36 måneder (IMO G7 MEPC.162(56)), og nødvendigheten av eventuell ny havneundersøkelse kan vurderes etter blant annet trafikkmengden i havnen.
Noen av disse rutene går mellom havner som ligger i henholdsvis OSPAR- og HELCOM-områder. OSPAR og HELCOM har derfor blitt enige om et felles regelverk (Joint harmonized procedure). Regelverket er i samsvar med IMO-kravene, og i den omforente prosedyren er det utviklet et sett med risikoanalyser i tillegg til de omtalte undersøkelsesprotokollene.
4.4 - RAS (Rapid Assessment Survey)
Denne metodikken er velutviklet i USA og er testet ut i ulike deler av verden. Tanken bak denne metoden er at fremmede marine arter ofte trives på kunstig substrat og at ved å samle inn prøver fra bøyer, tau, flytebrygger og lignende kan få en ganske god oversikt over hvilke arter som finnes i området. Hovedtrekkene ved metoden er at man undersøker småbåthavner, bruker kort tid (typisk 1‑2 timer) på hver havn og undersøker så mange habitat som mulig. Metodikken kan suppleres med for eksempel innsamling av bløtbunnsfauna med liten grabb, strandundersøkelser og miljø-DNA prøver. Typisk for denne metodikken er at ikke mye materiale tas med tilbake på lab, men at det meste sorteres i fremmede/stedegne arter og identifiseres i felt. Metodikken krever personell med god kunnskap om stedegne arter og fremmede marine arter. Metoden som har blitt brukt i Norge fra 2010 bygger på Rapid Coastal Survey (Minchin 2007), der man på forhånd utvikler ID kort for fremmede arter, både etablerte og dørstokkarter. Bare arter som anses som fremmede eller arter som er vanskelig å identifisere i felt tas med tilbake til lab for videre undersøkelser. Arter som er lett å identifisere registreres i felt med et estimat av forekomst. I Norge har man brukt to personer på hvert team og arbeidet 1 time i hver havn, man drar fra sted til sted i bil og det er ikke behov for båt, noe som gjør at man kan operere over større områder uten å være avhengig av tokt. Man har også utvidet metodikken til strandundersøkelser der det har vært mulig for å fange opp arter som lever på bløtbunn, samt dykking og snorkling på utvalgte lokaliteter (Husa mfl. 2012a, b, 2013). HELCOM veileder for RAS: (https://helcom.fi/wp-content/uploads/2019/08/Guidelines-for-monitoring-of-non-indigenous-species-by-eRAS.pdf)
4.5 - Visuell kartlegging med kamera
Visuell kartlegging av fremmede arter med undervannskamera/drone/ROV krever selvsagt at målarten er stor nok til å foreta en sikker identifikasjon basert på videoopptak. Denne metodikken kan være anvendbar dersom arten lever litt dypere ned enn i strandsonen eller på brygger. Bentisk slede eller trekantskrape fanger også opp arter som lever dypere nede, men gir ikke noe kvantifiserbart resultat. Videoundersøkelser ved hjelp av undervannsdrone er testet ut for Didemnum vexillum (havnespy) og fungerer bra for denne arten, dersom man har kvalifisert personell som kjenner arten godt. Videoundersøkelser med slede brukes også til kartlegging av snøkrabbe og kongekrabbe. Andre arter som dette kan egne seg for er f. eks store sekkdyr, djevletunge og andre store makroalger, skjell som lever oppå substratet og store krepsdyr. Utplassering av videorigg med timelapse-kamera kan også være nyttig for å få et estimat på forekomst av mobile arter som ofte blir tallrike, slik som penselkrabbe (Hemigrapsus takanoi) og svartmunnet kutling (Neogobiusmelanostomus). Videoundersøkelser med ROV kan kombineres med prøvetaking av organismer ved hjelp av manipulatorarm og gripeklo.
4.6 - Miljø-DNA
Analyser av miljø-DNA-prøver er en ny metode for overvåking av arter og økosystemer der innsamling av prøver ikke er avhengig av langvarig innsats eller taksonomisk ekspertise i felt (Thomsen & Willerslev 2015, Valentini mfl. 2016). Flere miljø-DNA studier har vist at metoden kan være mer sensitiv til å påvise sjeldne fremmede arter enn konvensjonelle metoder, men det er fortsatt et gap mellom resultatene og bruk av disse i praktisk forvaltning (Sepulveda mfl. 2020). I marine miljøer med store arealer og lav tetthet vil miljø-DNA kunne være et viktig første skritt for å påvise sjeldne invasive arter i en tidlig fase (Rey mfl. 2020, Sepulveda mf. 2020, Bowers mfl. 2021, Fernandez mfl. 2021).
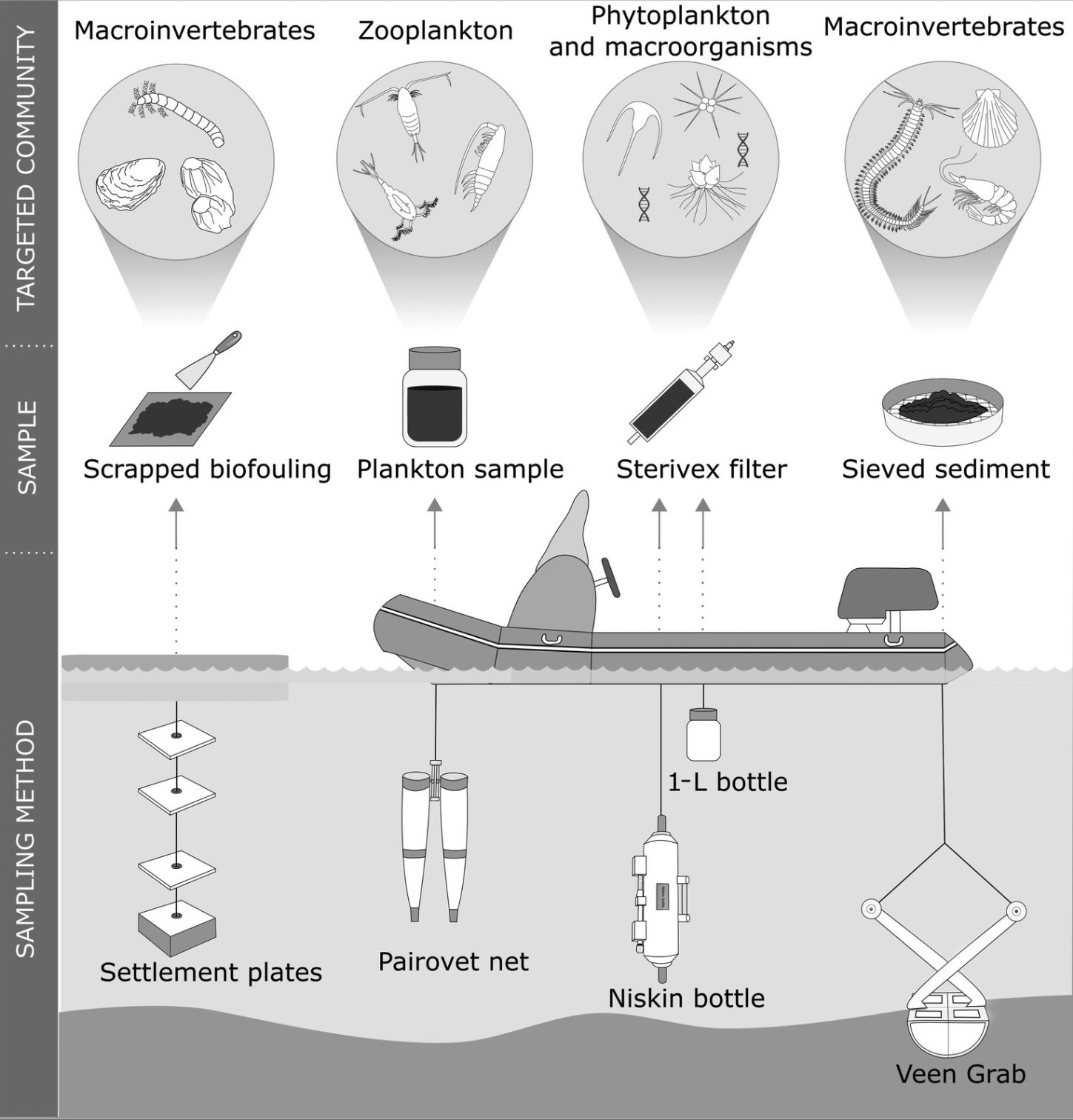
Miljø-DNA blir som oftest knyttet til filtrering av vann, men miljø-DNA inkluderer også mange andre prøvetyper (Taberlet mfl. 2012). Miljø-DNA inkluderer bl.a. sedimentprøver fra havbunnen, planktonprøver fra håvtrekk, begroingsprøver fra båter og brygger (Ardura & Zaiko 2018, Muñoz-Colmenero mfl. 2018, Fernandez mfl. 2021). Ulike prøvetyper vil fange opp ulike grupper av fremmede arter (Figur 11), og man bør derfor velge prøvetype egnet til arten man ønsker å påvise. Fisk er blant de best studerte gruppene så langt, og det finnes flere eksempler på at man kartlegger hele fiskesamfunn ved hjelp av vannprøver fra marine miljøer og artsgenerelle markører (Thomsen mfl. 2012, Thomsen mfl. 216, Fraija-Fernandez mfl. 2020). Evertebrater med harde eksoskjeletter lekker mindre DNA til vannet, og er dermed vanskeligere å påvise i vannprøver enn mykere organismer, som for eksempel fisk. Det er likevel eksempler på at fremmede krepsdyr og muslinger blir påvist med miljø-DNA i vannprøver (Ardura mfl. 2015, Forsström & Vasemägi 2016, Miralles mfl. 2016, Ardura & Zaiko 2018).
Vi skiller mellom to ulike metoder for å påvise arter gjennom miljø-DNA: artsspesifikke (qPCR/ddPCR) og artsgenerelle metoder (DNA-metastrekkoding/metagenomikk). For påvisning av spesifikke fremmede arter bruker man som oftest artsspesifikke markører som er designet for å kun amplifisere DNA fra en enkelt art. Utvikling av slike markører krever en hel del uttesting for å sikre at ikke andre arter gir et såkalt falskt positivt signal. Det har nylig blitt publisert en guide der slike artsspesifikke markører kan evalueres i forhold til hvor grundig de er testet (Thalinger mfl. 2021). Artsgenerelle markører blir som oftest brukt til DNA-metastrekkoding for å kunne lese millioner av DNA-strenger i en miljøprøve. Denne analysen krever en god del mer labarbeid, sekvensering på en «high-throughput» maskin og avanserte bioinformatiske analyser i etterkant. Avhengig av den genetiske markøren får man da en bred taksonomisk liste over arter i miljøprøven og kan potensielt påvise flere fremmede arter i den samme analysen (Djurhuus mfl. 2020, Duarte mfl. 2021). Men dersom man leter etter en sjelden fremmed art kan slike artsgenerelle markører være mindre sensitive enn en artsspesifikk markør (Zaiko mfl. 2018, Gargan mfl. 2021). Bruk av artsgenerelle markører krever gode referansebibliotek med kjente DNA-strenger fra artene man ønsker å påvise, men det er fortsatt store mangler for de fleste organismegrupper i marine miljøer. Offentlig databaser som Genbank (Benson mfl. 2006) og BOLD (Ratnasingham mfl. 2007) inneholder millioner av DNA-referanser fra hele verden, men også mange feil som kan gjøre indentifiseringen av arter vanskelig (Pentinsaari mfl. 2020).
Figur 11. Oversikt over prøvetype og innsamlingsmetode i forhold til DNA-bestemmelse av taksonomisk gruppe av fremmede arter. Figur gjengitt fra Rey mfl. 2020, alle innsamlingsmetoder var basert på HELCOM/OSPAR protokoller.
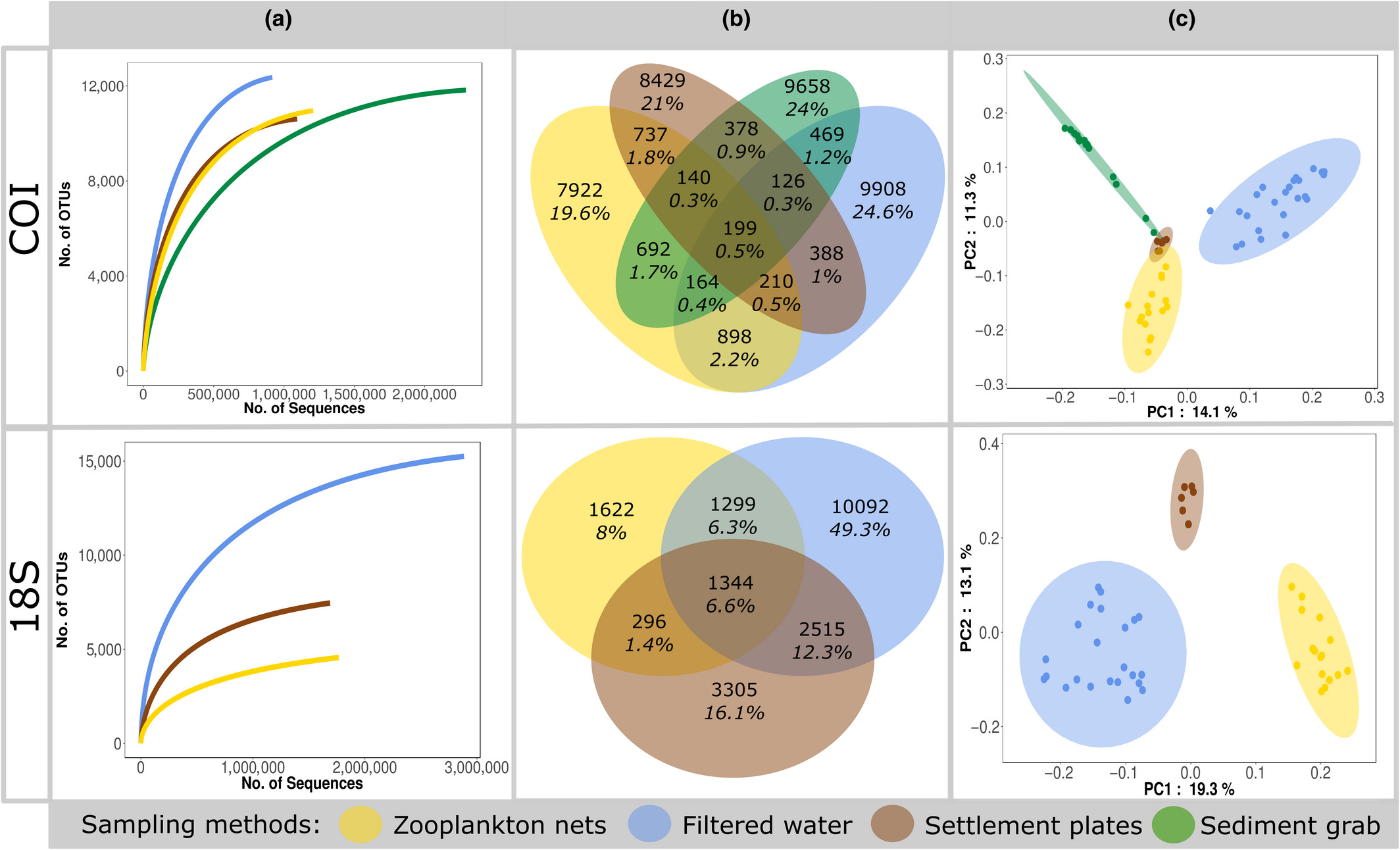
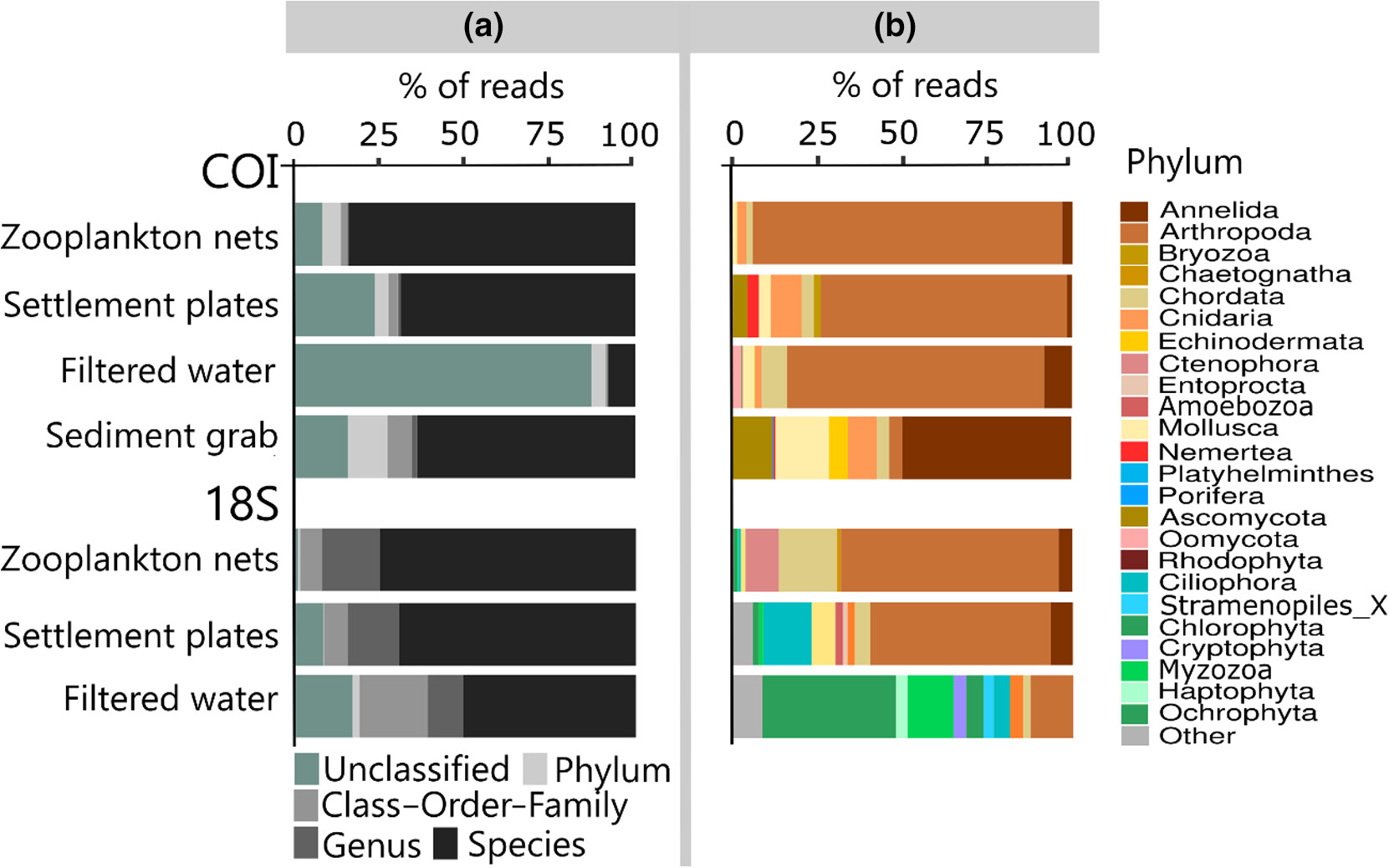
DNA-metastrekkoding av materialer fra ulike innsamlingsmetoder kan generere store komplekse artslister på tvers av mange organismegrupper og vil gi mye informasjon rundt kartlegging av både lokale og fremmede organismer. Fra Figur 12 ser vi at de ulike innsamlingsmetodene gir mange unike arter (b) og at artssamfunnene er svært ulike (c), mens figur 13 viser hvilke fylum som dominerer de ulike prøvematerialene (Rey mfl. 2020). Disse resultatene viser at de ulike prøvetypene og markørene komplementerer hverandre og at de til sammen gir et godt bilde av artsdiversiteten og dermed hvilke fremmede arter man finner i miljøet.
Figur 12. Resultater fra DNA-metastrekkoding av materiale fra ulike innsamlingsmetoder med to ulike markører (COI og 18S) der a) viser akkumulasjonskurver, b) viser overlapp i genetiske artsgrupperinger (OTUs) og c) viser en ordinasjon av artssamfunnene. Figur gjengitt fra Rey mfl. 2020, alle innsamlingsmetoder var basert på HELCOM/OSPAR protokoller.Figur 13. Resultater fra DNA-metastrekkoding av materiale fra ulike innsamlingsmetoder med to ulike markører (COI og 18S) der a) viser andel DNA-sekvenser bestemt til ulike taksonomiske nivåer og b) viser andel DNA-sekvenser fra ulike fylum. Figur gjengitt fra Rey mfl. 2020, alle innsamlingsmetoder var basert på HELCOM/OSPAR protokoller.
5 - Forslag til overvåkings- og kartleggingsplan av fremmede marine arter for Norge inkludert Svalbard
Basert på vurderinger gjort over og i Husa mfl. (2022) har vi her utarbeidet et forslag til metodikk for en nasjonal overvåknings og kartleggingsplan for norskekysten og Svalbard med søkelys på å både oppdage nye arter og overvåke forekomsten av etablerte fremmede arter.
5.1 - Pågående overvåking
Pågående, langsiktig overvåking av fremmede marine organismer er relativt beskjeden, selv om det foregår både overvåking og kartleggingsaktivitet (Husa mfl. 2022). Følgende arter dekkes av pågående overvåkningsprogram: pukkellaks, kongekrabbe, snøkrabbe, stillehavsøsters og utvalgte patogener. Vi forutsetter at disse programmene videreføres med den finansieringen som finnes i dag. Det bør etableres et eget overvåkningsprogram for de nye invasive artene Didemnum vexillum,Corella eymota, Hemigrapsus takanoi, Grateloupia turuturu.
5.2 - Hvilke områder bør overvåkes?
Overvåking av høyrisikoområder vil øke sjansen for å oppdage fremmede arter tidlig og bør prioriteres. For marine arter vil dette først og fremst være havner og skipsleier (Lehtiniemi 2015). Statistikk på havneanløp og skipstrafikk (Husa mfl. 2022) tyder på at risikoen er meget stor flere områder i Norge, nær de største havnene, særlig i Sør- og Vest-Norge. Den romlige fordelingen av allerede etablerte fremmede arter styrker også dette (Figur 4).
Selv om nye arter kan dukke opp i områder med lav risiko, er det hensiktsmessig å konsentrere overvåkningen i havner med svært høy og høy risiko. Da begroing av fremmed biota på skip må anses som den største risikovektoren for introduksjon av fremmede arter til norskekysten i dag, vil havnene med størst risiko være de som mottar flest anløp fra utlandet i løpet av et år. Basert på vurderinger gjort av skipsanløp fra utlandet (Husa mfl. 2022) er 44 havner valgt ut for regelmessig overvåkning (Tabell 3). Det bør vurderes om noen flere av havnene i moderat risiko skal overvåkes i tillegg (Tabell 2). To havner er valgt ut på Svalbard, selv om vurderingen basert på data fra 2020-2021 viser få fartøyanløp fra utenlandske havner. I et normalår kan situasjonen være noe annerledes, da Svalbard har hatt strenge Covid-19 restriksjoner i denne perioden. Disse to havnene bør overvåkes med tanke på at de marine økosystemene på Svalbard regnes som ekstra sårbare for introduksjoner av fremmede arter. Tabell 2 viser også forslag til antall stasjoner basert på havnenes størrelse og anbefalinger gitt i HELCOM protokollen. I tillegg vises forslag til hvordan feltarbeidet kan rulleres mellom år slik at hver havn blir undersøkt minst hvert 5. år som anbefalt både i HELCOM og CRIMP protokollen. Det bør vurderes om havner med svært høy risiko skal undersøkes med kortere intervall, f. eks hvert 3. år.
Tabell 3. Utvalg av havner foreslått for regelmessig overvåkning i norske fylker. Tabellen viser også forslag til antall stasjoner per havn basert på størrelsen til havnen og hvordan undersøkelsene kan rulleres i en femårsperiode. i.e. = forklaring. * tre havner i Trøndelag foreslås som opsjon
Risikokategori cargo/spesial/tankskip
Tilleggsrisiko
Antall stasjoner
År
Oslo & Viken
Drammen
Høy risiko
3
1
Moss
Høy risiko
3
1
Fredrikstad
Svært høy risiko
3
1
Oslo
Svært høy risiko
Cruisehavn, utenlandske ferger
7
1
Vestfold & Telemark
Kragerø
Høy risiko
3
1
Brevik
Høy risiko
3
1
Rafnes
Svært høy risiko
5
1
Porsgrunn
Svært høy risiko
5
1
Larvik
Høy risiko
Utenlandske ferger
3
1
Sandefjord
Moderat risiko
Utenlandske ferger
3
1
Slagen
Høy risiko
3
1
Horten
Høy risiko
3
1
Agder
Farsund
Moderat risiko
Offshore opplag
3
2
Kristiansand
Høy risiko
Utenlandske ferger
5
2
Rogaland
Ølen
Lav risiko
Plattformopplag
3
2
Haugesund
Høy risiko
Offshore opplag
5
2
Storasund
Moderat risiko
Offshore opplag
3
2
Husøya
Høy risiko
Offshore opplag
3
2
Kårstø
Høy risiko
5
2
Breiviken
Høy risiko
3
2
Stavanger
Høy risiko
Cruisehavn
5
2
Tananger
Svært høy risiko
Utenlandske ferger
5
2
Egersund
Høy risiko
3
2
Jøssingfjord
Høy risiko
3
2
Vestland
Hanøytangen
Moderat risiko
Plattformopplag
3
3
Ågotnes
Moderat risiko
Plattformopplag
3
3
Skipavik
Moderat risiko
Plattformopplag
3
3
Mongstad
Svært høy risiko
5
3
Sture
Høy risiko
3
3
Bergen
Høy risiko
Cruisehavn, utenlandske ferger
5
3
Odda
Høy risiko
3
3
Florø
Høy risiko
3
3
Måløy
Høy risiko
3
3
Flåm
Cruisehavn
3
3
Møre & Romsdal
Ålesund
Høy risiko
Cruisehavn
3
4
Elnesvågen
Høy risiko
3
4
Averøya
Høy risiko
3
4
Geiranger
Cruisehavn
3
4
Trøndelag
Trondheim
Moderat
3
*
Orkanger
Moderat
3
*
Verdal
Moderat
3
*
Nordland
Mo i Rana
Høy risiko
3
4
Glomfjord
Høy risiko
3
4
Narvik
Høy risiko
3
4
Troms & Finnmark
Tromsø
Moderat risiko
Cruisehavn
3
4
Svalbard
Longyearbyen
Lav risiko
Ekstra sårbarhet
3
4
New Ålesund
Lav risiko
Ekstra sårbarhet
3
4
5.3 - Når skal det overvåkes?
Alle protokoller anbefaler at innsamling av arter i felt gjøres i perioden juli til september da påvekstsamfunnene normalt er velutviklet.
5.4 - Forarbeid
Identifisering av særlige risikoområder i hver havn bør utføres basert på data fra Marine Traffic eller havnemyndighetene. Områder med høy frekvens av utenlandske anløp bør prioriteres.
Analyser av de ulike områdene i havnen basert på dyp, substrat, kaier, flytebrygger og andre installasjoner slik at man sikrer at alle typer habitat blir undersøkt
Utarbeiding av en detaljert protokoll for innsamlingen og preservering av prøver basert på protokoller for metoden som er valgt.
Man må søke om tillatelse for adkomst til ISPS havner.
Sette opp en tidsplan for arbeidet og en plan for datalagring og publisering
5.5 - Målarter
For at ikke arbeidet med å identifisere arter skal bli for omfattende og kostbart er det en fordel å på forhånd utarbeide en liste med aktuelle målarter. Dette er gjort både for Australia (CRIMP) og for landene rundt Østersjøen (HELCOM). For begge metoder er det utarbeidet lister basert på omfattende kriterier og risikovurderinger. Sverige har anvendt den Belgiske ISEIA protokollen (ISEIA 2009), men har inkludert noen egne kriterier slik som risiko for effekter på naturlige ressurser og human helse. Danmark har valgt ut en liste med 50 arter, hvorav kun fire er dørstokkarter for Danmark, resten er etablerte (Andersen mfl. 2016). For norske målartlister er det naturlig å bruke lister fra Artsdatabankens vurdering av fremmede marine arter og de nye dørstokkartene som går videre fra horisontskannings-listene til vurdering i 2022/23. Dette vil gi et relativt høyt antall arter som skal inngå i kartleggingen, men når man legger så mye arbeid ned i innsamling av arter bør også potensielle nye arter for Norge inkluderes. Fremmede marine arter er så lite kartlagt i Norge at mange av dørstokkartene sannsynligvis allerede er etablert her. Arter som er vurdert til svært høy, høy og potensielt høy risiko bør ha størst fokus, mens arter som er vurdert til lav risiko og ingen kjent risiko bør inkluderes, men ha lavere prioritet. Det er en fordel å organisere målartlistene etter habitat/metode slik at det gjør arbeidet med identifisering lettere. Gode identitetskort med de karakteristiske kjennetegn og gode bilder av artene er en stor fordel. I kartleggingsarbeidet av fremmede arter med Rapid Coastal Survey metoden ble det utarbeidet slike ID kort for omtrent 35 arter, både etablerte og dørstokkarter (Figur 14).
Figur 14. Eksempel på ID kort for fremmede marine arter til bruk for identifisering i felt og lab.
5.6 - Målarter med eksisterende genetiske markører og DNA-referanser
Artsspesifikke genetiske markører for qPCR/ddPCR analyser eksisterer allerede for mange av de marine artene på den norske fremmedartslista. Artsspesifikke analyser kan relativ raskt teste for forekomst av enkeltarter på tvers av mange prøver. Eksempelvis har NINA og HI i 2021 undersøkt forekomst av svartmunnet kutling (Negogobius melanostomus) i en rekke miljø-DNA prøver innsamlet i Oslofjordområdet og Trondheim, samt gjennomført en Rapid Coastal Survey av havnespy (Didemnum vexillum) i Rogaland og Vestland ved hjelp av artsspesifikke miljø-DNA analyser. Danmark har gjennom sitt MONIS prosjekt (Monitoring of Non-Indigenous Species in Danish Marine Water) og utviklet artsspesifikke markører for en rekke fremmede marine arter (Andersen mfl. 2016, 2018). Nylig ble 18 av disse markørene testet i 16 danske havner og resultatet viste at enkelte arter som for eksempel Amerikansk lobemanet (Mnemiopsis leidyi) ble påvist i samtlige havner (Knudsen mfl. 2022).
Krepsdyr med kraftige skall viser seg å avgi lite DNA til miljøet, og store hummere og krabber kan derfor være vanskelig å påvise ved hjelp av vannprøver og miljø-DNA analyser, men det er utviklet og testet artsspesifikke markører for blant annet amerikansk hummer (Homarus americanus), kongekrabbe (Paralithodes camtschaticus) og kinesisk ullhåndskrabbe (Eriocheir sinensis) (Andersen mfl. 2018). Hvor sensitive vannprøver er for å påvise disse artene er fortsatt uvisst, men markørene kan også brukes til å identifisere larvestadier av disse artene i planktonprøver.
Artsgenerelle genetiske markører for DNA-metastrekkoding kan påvise svært mange arter i en og samme analyse, og kan dermed også påvise flere fremmede arter på en effektiv måte. Men siden de generelle markørene vil feste seg på mange ulike arter samtidig, vil man ikke nødvendigvis finne den samme arten hver gang. Gode protokoller med stor nok sekvenseringsdybde er derfor viktig for denne metoden. DNA-metastrekkoding forutsetter også at arten man ønsker å påvise finnes i DNA-referanse bibliotekene som for eksempel Genbank (Benson mfl. 2006) og BOLD (Ratnasingham & Hebert 2007), og at den ikke har nært beslektede arter som har nesten identisk strekkode. For komplekse prøver som for eksempel håvtrekk, begroingsplater og bunnprøver med potensielt mange arter vil DNA-metastrekkoding være til stor hjelp for å identifisere arter som ofte kan være vanskelig å skille fra hverandre eller som har livsstadier som ikke lar seg identifisere ved hjelp av morfologiske analyser. Rent kostnadsmessig vil det ofte heller ikke være mulig å bestemme alle artsgrupper ved hjelp av morfologi da dette vil ta svært lang tid og kreve mange taksonomiske eksperter for de ulike organismegruppene. Det har også blitt færre taksonomiske eksperter over tid og det er i dag vanskelig å finne kvalifiserte eksperter for mange organismegrupper.
5.7 - Alternativ I. Tradisjonell HELCOM-metodikk
Alle havner i Tabell 3 undersøkes hvert 5. år med tradisjonell HELCOM metodikk (HELCOM 2013) som fremgår av Tabell 4. Antall stasjoner i hver havn avgjøres av størrelse på havnen, men det er ikke gitt noen konkrete anbefalinger bortsett fra at man bør ha minst tre stasjoner i hver havn. I Tabell 4 er det gitt forslag til antall stasjoner per havn, men dette kan justeres ved en nærmere vurdering av havnen. Genetiske metoder kan inngå i undersøkelsen, slik som beskrevet i alternativ II.
Tabell 4. Parametere, metodikk og antall prøver per stasjon etter anbefalinger i HELCOM-metodikk
Parameter
Metodikk/utstyr
Antall prøver per stasjon
Temperatur, salinitet, klorofyll-a og turbidet
CTD/secciskive
1
Patogener
Vannprøve
1
Planteplankton
Håvtrekk
3 x 2 (vår + sommer)
Dyreplankton
Håvtrekk med to ulike maskevidder
6
Dyr i sediment
Håndholdt grabb 0.025 m2
3
Fisk og krepsdyr
Krabbeteiner/små ruser
6 (48 timer)
Fastsittende organismer
Skrapeprøver
Minst 3
Fastsittende organismer
Begroingspanel
9 (1,5-3 mnd)
5.8 - Alternativ II. Tilpasset HELCOM/CRIMP metodikk med genetiske metoder
Alle havner i Tabell 3 undersøkes hvert 5. år med tilpasset HELCOM/CRIMP metodikk som fremgår av Tabell 5. Antall stasjoner i hver havn avgjøres av størrelse på havnen, men det er ikke gitt noen konkrete anbefalinger bortsett fra at man bør ha minst tre stasjoner i hver havn. I Tabell 3 er det gitt forslag til antall stasjoner per havn, men dette kan justeres ved en nærmere vurdering av havnen.
Tabell 5. Parametere, metodikk og antall prøver per stasjon ved tilpasset metodikk
Parameter
Metodikk/utstyr
Antall prøver per stasjon
Temperatur, salinitet, klorofyll-a og turbidet
CTD/secciskive
1
Patogener
Prøver av påvekstorganismer
3
Planteplankton
Vannprøve/DNA
1
Dyreplankton
Håvtrekk med to ulike maskevidder (3 til DNA)
6
Dyr i sediment
Håndholdt grabb 0.025 m2/DNA
3
Fisk og krepsdyr
Krabbeteiner/ørekytteiner/DNA
6 (12 timer)
Fisk
Vannprøve (DNA)
1
Fastsittende organismer
Skrapeprøver grunt
Minst 3, flere hvis ikke dykkere brukes
Alternativ 1: Fastsittende organismer
Skrapeprøver/dykkere
9
Alternativ 2: Fastsittende organismer/epifauna
Videotransekt/skrapetrekk
50-100 m X 6
Patogener: HELCOM foreslår prøver av vann i havnene som skal analyseres i henhold til EU’s badevannsdirektiv. Disse analysene har hovedfokus på humane patogener. I Norge har man regelmessig overvåkning av badevannskvalitet som følger dette direktivet. Denne overvåkningen vil sannsynligvis være tilstrekkelig til å avdekke fremmede humane patogener, det er derfor ikke hensiktsmessig å gjennomføre disse analysene i alle havner. I dag foregår det overvåkning av patogener som påvirker oppdrettsorganismer og en begrenset overvåkning av patogener i europeisk østers. Tidligere når vi hadde store utslipp av ballastvann var kanskje dette med overføring av patogener i vann et større problem enn i dag. Overføring av vann fra utlandet skjer hovedsakelig med transportvann av leppefisk fra Sverige. Det bør tas prøver av utslippsvannet for å få oversikt over hvor stort problemet er og hvilken risiko organismene i dette vannet utgjør. Vi har i dag liten kunnskap om hvilke mikroorganismer som kan overføres med påvekst på båter fra utlandet. Vi foreslår derfor at man utfører prøvetaking av skjell, østers og andre påvekstorganismer på et utvalg av fartøy for å identifisere målarter for patogener. Når man har identifisert en gruppe aktuelle risikoorganismer kan man ta prøver av skjell og andre påvekstorganismer på brygger i havneområder og undersøke disse med histologi og genetiske metoder.
Planteplankton: Ettersom artssammensetningen i planteplanktonsamfunn overvåkes som en del av ØKOKYST programmet (Miljødirektoratet) og fremmede mikroalger rapporteres inn, ser vi det som lite hensiktsmessig å ta så mange planktonprøver i hver havn som HELCOM protokollen anbefaler. I stedet kan det tas en vannprøve for DNA i havner som ligger lenger unna en ØKOKYST stasjon enn 50 km. Vannprøver og miljø-DNA har vist seg å være effektive for påvisning av planteplankton (Jacobs-Palmer mfl. 2021), og ballastvann fra båter har vært brukt til å påvise transportveien til fremmede arter (Zaiko mfl. 2015). Genetiske metoder bør testes ut sammenlignet med konvensjonelle metoder i et utvalg av havner.
Dyreplankton: HELCOM foreslår seks håvtrekk for dyreplankton i havnene. Trekkene med stor maskevidde skal fange opp geleplankton, som hovedsakelig skal artsbestemmes i felt så langt som mulig, er ikke så tidskrevende, mens trekkene med mindre plankton er tidskrevende å artsbestemme. Vi foreslår derfor at man gjennomfører de seks håvtrekkene og identifiserer geleplankton i felt, mens en samleprøve av mindre plankton analyseres ved DNA-metastrekkoding. Dyreplankton er antakelig den best studerte gruppen når det gjelder DNA-analyser for påvisning av fremmede organismer (Duarte mfl. 2021). DNA-metastrekkoding er brukt på vannprøver fra sjøen (Rey mfl. 2020, Westfall mfl. 2020), ballastvann (Zaiko mfl. 2015, Darling mfl. 2018) og lensevann fra båter (Fletcher mfl. 2017) samt bulkprøver av plankton (Stefanni mfl. 2018). Som for planteplankton vil håvtrekk egne seg svært bra for DNA-metastrekkoding og gi en effektiv artsbestemmelse av mange fremmede arter samtidig (Couton mfl. 2019). Genetiske metoder bør testes ut sammenlignet med konvensjonelle metoder i et utvalg av havner.
Dyr i sediment: Analyser av infauna i sediment er ganske kostnadskrevende. Vi foreslår derfor tre sedimentprøver som tas med håndholdt grabb som analyseres med genetisk metodikk. Det er viktig at noen av grabbhuggene tas også i helt grunne områder. Sedimentprøver fra grabbprøver kan brukes direkte inn i DNA-metastrekkoding analyser og har blant annet vært brukt til å påvise flere marine fremmede arter på Svalbard (Heuvel-Greve 2021). Små delprøver av sedimentene kan lagres på etanol for videre genetiske analyser og vil kunne oppbevares i lang tid. Den samme grabbprøven kan derfor brukes både til konvensjonelle analyser og DNA-metastrekkoding. Sammenligninger av DNA-metastrekkoding fra vannprøver og sedimentprøver viser at artssamfunnet man fanger opp er svært forskjellig (Holman mfl. 2019, Rey mfl. 2020), og at prøvetypene derfor vil påvise ulike fremmede arter. Genetiske metoder bør testes ut sammenlignet med konvensjonelle metoder i et utvalg av havner.
Fastsittende organismer: I et land som Norge, med sin lange kystlinje, kan det være logistisk vanskelig å bruke begroingspanel i og med at de skal stå ute 1,5- 3 måneder i sommerhalvåret og man må reise langt for å sette de ut og ta de inn igjen. Rinde mfl. (2017) testet ut bruk av begroingsplater i Oslofjorden og erfarte at det er lett å miste platene i utsettingsperioden. Når tau henges ut fra brygger og kaier vil det uunngåelig gnages på tauene og platene vil lett falle av. Andersen mfl. (2017) anbefaler at panelene festes under flytebrygger for mindre slitasje. Mange av høyrisikohavnene er rene industrihavner med kontinuerlig båttrafikk der det kan være vanskelig å få festet panelene på en slik måte at de ikke skades av båttrafikk. I disse havnene kan det også være en prosess i forkant for å få tilgang til havnen så det kan være en fordel om man kan gjøre ferdig alt arbeidet på en gang. Vi foreslår derfor at man tester ut begroingspaneler i noen nærliggende havner først og sammenligner resultatene med innsamling ved hjelp av dykkere ifølge CRIMP protokollen, samt innsamling på tilgjengelig hardt substrat. Identifisering av tidlige stadier av fastsittende arter kan være svært tidkrevende, og genetiske metoder kan testes ut for slike plater. Platene trenger da heller ikke stå ute så lenge som anbefalt i HELCOM-protokollene da det vil være mulig å identifisere arter på et tidlig stadium i utviklingen. Flere studier har også brukt DNA-metastrekkoding for å artsbestemme materiale fra begroingsplater (Zaiko mfl. 2016, Rey mfl. 2020).
Alternativ til dykking kan være innsamling på alle mulige harde substrat i havnen, samt bruk av liten håndholdt trekantskrape for dypere bunnprøver.
CRIMP protokollen anbefaler at dykkere undersøker horisontale transekter på tre ulike dyp langs kaier og moloer for å undersøke bentisk og epibentisk biota. De åpner opp for alternativ bruk av video for disse undersøkelsene dersom det er vanskelig å dykke i havnen. Vi anbefaler derfor lignende videotransekter kjørt med undervannsdrone eller liten ROV med manipulatorarm for å samle inn prøver. Denne metodikken vil kunne fange opp større arter/kolonier slik som for eksempel djevletunge og havnespy. Videotransekter bør kjøres minst i tre hardbunnstransekt og tre bløtbunnstransekt (tre dyp; 0,5, 3 og 5-10 meter). Videotransektene bør være 50-100 meter lange. Dersom man ikke har manipulatorarm kan en håndholdt trekantskrape samle inn prøver i transektene, enten fra land eller gummibåt.
Innsamling av hardbunnsorganismer bør tilpasses den enkelte havn. Hvis det er mange småbåthavner i havnen, kan man bruke RAS metodikk og sikre prøver fra dypere vann ved å dra opp tau etc. Dersom det er en industrihavn og/eller ISPS havn uten slike strukturer i nærheten bør man planlegge for å bruke dykkere til skrapeprøver på tre dyp.
Fisk og krepsdyr: For å fange fisk og større krepsdyr som lever på bunn anbefaler vi samme oppsett som i alternativ I, men reduserer fangsttiden med 12 timer over natten. Dette burde være tilstrekkelig til at man klarer å fange et representativt utvalg, og er også anbefalt i CRIMP-protokollen. På denne måten kan man gjennomføre undersøkelsene i et område på kortere tid og kan redusere behovet for innkjøp av teiner. Vi anbefaler også at metodikken suppleres med miljø-DNA prøver for et utvalg av fiske-målarter. Miljø-DNA for påvisning av fisk i marine vannprøver kan i mange tilfeller være en sensitiv og kostnadseffektiv metode (Thomsen mfl. 2012, 2016). Prøvene bør som oftest tas under sommerhalvåret da temperaturen er høyere og fisken er mer aktiv. Det er heller ikke uvanlig at fisker går ned på dypere vann om vinteren og dermed blir vanskeligere å oppdage (f.eks. Behrens mfl. 2021). Enkeltarts analyser av pukkellaks har vært utført ved hjelp av folkeforskning (citizen science: www.1000rivers.net) mens NINA og HI har brukt slike analyser for å undersøke forekomst av svartmunnet kutling i Oslofjordområdet og Trondheim. DNA-metastrekkoding av vannprøver påviser som oftest langt flere fiskearter enn konvensjonelle fangstmetoder (Thomsen mfl. 2012). Vannprøver i høy-risikoområder som store havner med mye internasjonal trafikk synes å være en god strategi for påvisning av fremmede fiskearter.
DNA-prøver versus innsamlede prøver: Det er viktig å ta vare på de fysiske innsamlede prøvene av planteplankton, dyreplankton, dyr i sediment slik at man kan verifisere eventuelle DNA signal med prøver i ettertid dersom det er nødvendig. Oppbevaring av fysiske referansesamlinger bør skje i samarbeid med de Naturhistoriske museene.
5.9 - Alternativ III. Tilpasset Rapid Coastal Survey (RAS) med genetiske metoder
Et mindre kostnadskrevende alternativ vil være å bruke RAS metodikk, med litt tilpasninger for å fange opp andre grupper enn fastsittende arter. Undersøkelser av småbåthavner med RAS metoden har vært gjennomført på om lag 450 stasjoner fra svenskegrensen til Sognefjorden og i et område fra Stadt til Ålesund i perioden 2010-2021 (Husa mfl. 2022). Metodikken har blitt utvidet til også å omfatte målrettete strandsøk for å se etter krepsdyr og bløtdyr.
Vi foreslår at alle havner med høy risiko og svært høy risiko (Tabell 3), samt alle havner med moderat risiko (Tabell 2) undersøkes. Det vil gis to kostnadsoverslag med og uten undersøkelse av havner med moderat risiko. Det vil gi et totalantall på 77 havner. Flytebrygger i havnen og i andre småbåthavner innenfor 5 km radius fra havnen skal undersøkes (Tabell 6). Det bør være minst tre brygger/småbåthavner som undersøkes (opptil syv i større havner), og man bør forsøke å legge stasjonene slik at flere typer habitat dekkes (høy/lav salinitet, bølgeeksponert/beskyttet etc.). Tre flytebrygger i hver småbåthavn undersøkes og det tas minst tre skrapeprøver av et standardisert areal 0,1 m2 på hver brygge. Tau dras opp og det forsøkes å ta standardiserte skrapeprøver fra ulike dyp på disse. Det foretas en visuell inspeksjon langs bryggene og det tas prøver av mistenkelig biota. Alt materiell sorteres og skannes for målarter i felt hvis mulig og det tas prøver og foto av målarter for dokumentasjon. Det tas prøver av arter som er vanskelig å bestemme i felt for seinere identifisering i lab. I tillegg til standard undersøkelser på brygger bør det kjøres minst 3 x 50 meters transekt på 3-10 meters dyp undervannsdrone med god billedkvalitet langs kaier og moloer. Det bør gjennomføres strandundersøkelser langs et minst 100 meter langt transekt der man også løfter på stein for å se etter mobile krepsdyr. Strandundersøkelsene bør dekke både hardt og bløtt substrat, og en kasterive kan benyttes for å samle inn løsliggende alger (f.eks. Agarophyton vermiculophyllum). I tillegg settes det minst tre krabbeteiner og tre ørekyteteiner i hver hovedhavn som står ute 12 timer over natten. Det bør tas prøver til DNA analyser som beskrevet i Alternativ II, men kun en prøve per stasjon. Det bør også samles inn prøver av påvekstorganismer for analyse av patogener.
Noen høyrisiko-industrihavner har ikke småbåthavner eller andre lave brygger i nærheten. En bør derfor vurdere å samle inn fastsittende arter med dykkere som beskrevet og prisestimert i Alternativ II.
DNA-prøver versus innsamlede prøver: Det er viktig å ta vare på de fysiske innsamlede prøvene av planteplankton, dyreplankton, dyr i sediment slik at man kan verifisere eventuelle DNA signal med prøver i ettertid dersom det er nødvendig.
Tabell 6. Parametere, metodikk og antall prøver per stasjon ved RAS metodikk og genetiske metoder. En stasjon er her lik 3 flytebrygger i en havn.
Parameter
Metodikk/utstyr
Antall prøver per stasjon
Temperatur, salinitet, klorofyll-a og turbidet
CTD/secciskive
1
Patogener
Prøver av påvekstorganismer
3
Planteplankton
Vannprøve/DNA
1
Dyreplankton
Ett håvtrekk/DNA
1
Dyr i sediment
Håndholdt grabb 0.025 m2/DNA
1
Fisk og krepsdyr
Krabbeteiner/små ruser/DNA
6 (12 timer) (Minst ett sted i hovedhavn)
Fisk
Vannprøve/DNA
1
Fastsittende organismer
Skrapeprøver/brygger/tau
9
Alternativ 2: Fastsittende organismer/epifauna
Videotransekt
3
Epifauna
Strandsøk
1
6 - Referanser
Andersen, JH, Kallenbach E, Hesselsøe M, Knudsen, SW, Møller, PR, Bekkevold, D, Hansen, BK & Thaulow, J. 2016. Steps toward nation-wide monitoring of non-indigenous species in Danish marine waters under the Marine Strategy Framework Directive. Rapport 7022-2016 DK3. NIVA Denmark.
Andersen JH, Kallenbach E, Thaulow J, Hesselsøe M, Bekkevold D, Hansen BK, Jacobsen LMW, Olesen CA, Møller PR, Knudsen SW. 2018. Development of species-specific eDNA-based test systems for monitoring of non-indigenous species in Danish marine waters. RAPPORT L.NR. 7204-2017. Norwegian Institute for Water Research.
Andersen JH, Brink M, Kallenbach E, Hesselsøe M, Knudsen SW, Støttrup JG, Rask Møller P, Ekrem W, Fagerli C, Oug E. 2017. Sampling protocol for monitoring of non-indigenous species in selected Danish harbours. NIVA report 7175-2017.
Andersen, JH, Kallenbach, E, Kjeldgaard, MB, Knudsen, SW, Dale, T, Eikrem, W, Fagerli, C, Green, G, Hobæk, A, Oug, E, Thaulow, J, Hesselsøe, M, Bekkevold, D, Jacobsen, LMW, Kuhn, J, Møller, PR, Olesen, CA, Car, H & Stuer-Lauridsen, F. i trykk. A baseline study of the occurrence of non-indigenous species in Danish harbours. NIVA Denmark Report.
Anon. 2011. Prognoser for lakseinnsig, regnbueørret og klimaendringer: utfordringer for forvaltningen. Temarapport fra Vitenskapelig råd for lakseforvaltning Nr 2, 45 s.
Ardura, A, Zaiko, A, Martinez, JL, Samulioviene, A, Semenova, A & Garcia-Vazquez, E. 2015. eDNA and specific primers for early detection of invasive species – A case study on the bivalve Rangia cuneata, currently spreading in Europe. Marine Environmental Research 112, Part B: 48-55.
Ardura A, Zaiko A. 2018. PCR-based assay for Mya arenaria detection from marine environmental samples and tracking its invasion in coastal ecosystems. Journal for Nature Conservation 43: 1-7.
Behrens JW, Ryberg MP, Einberg H, Eschbaum R, Florin A-B et al. 2021. Seasonal depth distribution and thermal experience of the non-indigenous round goby Neogobius melanostomus in the Baltic Sea: implications to key trophic relations. Biological Invasions 24, Pages 527–541. https://doi.org/10.1007/s10530-021-02662-w
Benson DA, Karsch-Mizrachi I, Lipman DJ, Ostell J, Wheeler DL. 2006. GenBank. Nucleic Acids Research 34. Issue suppl_1, 1 January 2006, Pages D16-D20.
Berntsen HH, Sandlund OT, Thorstad EB & Fiske P. 2020. Pukkellaks i Norge, 2019. NINA Rapport 1821. Norsk institutt for naturforskning. https://hdl.handle.net/11250/2651741
Berntsen HH, Diserud OH, Hanssen F, Sandlund OT. 2021. Pukkellaks i Norge: kan vi forutse hvor den etablerer seg i fremtiden? Nåværende og mulig fremtidig utbredelse. NINA Rapport 2004. Norsk institutt for naturforskning. https://hdl.handle.net/11250/2755582
Bowers HA, Pochon X, von Ammon U, Gemmell N, Stanton J-AL, Jeunen G-J, Sherman CDH, Zaiko A. 2021. Towards the Optimization of eDNA/eRNA Sampling Technologies for Marine Biosecurity Surveillance. Water 13(8).
Brattegard T, Holthe T. (editors), 1997. Distribution of marine, benthic macro-organisms in Norway. Research Report for DN Nr. 1997-1. Directorate for Nature Management. 409 s.
Couton, M, Comtet, T, Le Cam, S, Corre, E & Viard, F. 2019. Metabarcoding on planktonic larval stages: an efficient approach for detecting and investigating life cycle dynamics of benthic aliens 10: 657-689.
Darling, JA, Martinson, J, Gong, Y, Okum, S, Pilgrim, E, Lohan, KMP, Carney, KJ & Ruiz, GM. 2018. Ballast Water Exchange and Invasion Risk Posed by Intracoastal Vessel Traffic: An Evaluation Using High Throughput Sequencing. Environmental Science & Technology 52(17): 9926-9936.
Djurhuus, A, Closek, CJ, Kelly, RP, Pitz, KJ, Michisaki, RP, Starks, HA, Walz, KR, Andruszkiewicz, EA, Olesin, E, Hubbard, K, Montes, E, Otis, D, Muller-Karger, FE, Chavez, FP, Boehm, AB & Breitbart, M. 2020. Environmental DNA reveals seasonal shifts and potential interactions in a marine community. Nature Communications 11(1): 254.
Duarte, S, Vieira, PE, Lavrador, AS & Costa, FO. 2021. Status and prospects of marine NIS detection and monitoring through (e)DNA metabarcoding. Science of The Total Environment 751: 141729.
Fernandez, S, Miller, DL, Holman, LE, Gittenberger, A, Ardura, A, Rius, M & Mirimin, L. 2021. Environmental DNA sampling protocols for the surveillance of marine non-indigenous species in Irish coastal waters. Marine Pollution Bulletin 172: 112893.
Fletcher, LM, Zaiko, A, Atalah, J, Richter, I, Dufour, CM, Pochon, X, Wood, SA & Hopkins, GA. 2017. Bilge water as a vector for the spread of marine pests: a morphological, metabarcoding and experimental assessment. Biological Invasions 19(10): 2851-2867.
Forsström, T & Vasemägi, A. 2016. Can environmental DNA (eDNA) be used for detection and monitoring of introduced crab species in the Baltic Sea? Marine Pollution Bulletin 109(1): 350-355.
Fraija-Fernandez, N, Bouquieaux, MC, Rey, A, Mendibil, I, Cotano, U, Irigoien, X, Santos, M & Rodriguez-Ezpeleta, N. 2020. Marine water environmental DNA metabarcoding provides a comprehensive fish diversity assessment and reveals spatial patterns in a large oceanic area. Ecol Evol 10(14): 7560-7584.
Gargan, LM, Mo, TA, Carlsson, JEL, Ball, B, Fossøy, F & Carlsson, J. 2021. Development of an environmental DNA assay and field validation for the detection of invasive pink salmon (Oncorhynchus gorbuscha). Environmental DNA, in press.
Grefsrud ES, Karlsen Ø, Kvamme BO, Glover K, Husa V, Hansen PK, Grøsvik BE, Samuelsen O, Sandlund N, Stien LH, Svåsand T. 2021. Risikorapport norsk fiskeoppdrett 2021-kunnskapsstatus. Rapport fra Havforskningen 2021-7.
Hansen H, Karlsbakk E. 2018. Pacific false kelpfish, Sebastiscus marmoratus (Cuvier, 1829) (Scorpaeniformes, Sebastidae) found in Norwegian waters. BioInvasions Records 7:73-78.
HELCOM 2013. HELCOM ALIENS 2- Non-native species port survey protocols, target species selection and risk assessment tools for the Baltic Sea. 34 pp.
Heyvel-Greve MJ, Brink AM, Glorius ST, De Groot A, Laros I, Renaud PE, Pettersen R, Weslawski JM, Kuklinski P, Murk AJ. 2021. Early detection of marine non-indigenous species on Svalbard by DNA metabarcoding of sediment. Polar Biology 44: 653-665.
Holman, LE, de Bruyn, M, Creer, S, Carvalho, G, Robidart, J & Rius, M. 2019. Detection of introduced and resident marine species using environmental DNA metabarcoding of sediment and water. Scientific Reports 9(1): 11559.
Husa V, Heggøy E, Sjøtun K, Agnalt AL, Johansen PO, Glenner H & Hatlen K. 2012 a. Kartlegging av fremmede marine arter i Hordaland. Utredning for DN 2-2012. Direktoratet for naturforvaltning.
Husa V, Heggøy E, Agnalt A-L, Sjøtun K, Svensen R, Rokkan-Iversen K & Alvestad T. 2012 b. Kartlegging av fremmede marine arter i Rogaland. Utredning for DN 3-2012. Direktoratet for naturforvaltning.
Husa V, Agnalt AL, Svensen R, Rokkan Iversen K, Steen H, Jelmert A, Farestvedt E & Petersen H. 2013. Kartlegging av fremmede marine arter i indre og ytre Oslofjord. Utredning for DN 4-2013. Direktoratet for naturforvaltning
Husa V, Agnalt AL, Svensen R, Floerl O, Brattegard T, Rokkan-Iversen K, Glenner H, Bruntveit L. 2014. På jakt etter fremmede marine arter i Narvik havn. Rapport fra Miljødirektoratet M222-2014
Husa V, Agnalt A-L, Falkenhaug T, Fossøy F, Forsgren E, Grefsrud ES, Hansen F, Jelmert A, Mortensen S. 2022. Alien marine species in Norway. Mapping, monitoring and assessment of vectors for introductions. Rapport fra Havforskningen.
ISEIA protocol. 2009. Guidelines for environmental impact assessment and list classification of non-native organisms in Belgium. Ver 2.6 (07/12/2009). ISEIA_protocol (biodiversity.be)
Jacobs-Palmer, E, Gallego, R, Cribari, K, Keller, AG & Kelly, RP. 2021. Environmental DNA Metabarcoding for Simultaneous Monitoring and Ecological Assessment of Many Harmful Algae. Frontiers in Ecology and Evolution 9: 262.
Kabuta S, Gittenberg A, Brus D, Smolders S. 2014. Joint HELCOM/OSPAR Task Group on Ballast Water Management Convention Exemptions Fifth Meeting, Madrid, Spain, 1-2 December Rotterdam Port Sampling Project Report.
Knudsen, SW, Hesselsøe, M, Thaulow, J, Agersnap, S, Hansen, BK, Jacobsen, MW, Bekkevold, D, Jensen, SKS, Møller, PR & Andersen, JH. 2022. Monitoring of environmental DNA from introduced species of algae, dinoflagellates and animals in the North Eastern Atlantic. Science of The Total Environment: 153093.
Kotta J, Nurkse K, Puntila R, Ojaveer H. 2016. Shipping and natural environmental conditions determine the distribution of the invasive non-indigenous round goby Neogobius melanostomus in a regional sea. Estaurine, Coastal Shelf Science 169:15-24.
Lehtiniemi M, Ojaveer H, David M, Galil B, Gollasch S, McKenzie C, Minchin D, Occhipinti-Ambrogi A, Olenin S, Pederson J. 2015. Dose of truth – monitoring marine non-indigenous species to serve legislative requirements. Marine Policy 54:26-35.
Leung B, Lodge DM, Finnoff D, Shogren JF, Lewis MA, Lamberti G. 2002. An ounce of prevention or a pound of cure: bioeconomic risk analysis of invasive species. Proceedings of the Royal Society B 269:2407-2413.
Marraffini M, Ashton G, Brown C, Chang A, Ruiz G. 2017. Settlement plates as monitoring devices for non-indigenous species in marine fouling communities. MBI. 8:559–566. doi:https://doi.org/10.3391/mbi.2017.8.4.11
Minchin D. 2007. Rapid coastal survey for targeted alien species associated with floating pontoons in Ireland. Aquatic Invasions 2:63-70.
Miralles, L, Dopico, E, Devlo-Delva, F & Garcia-Vazquez, E. 2016. Controlling populations of invasive pygmy mussel (Xenostrobus securis) through citizen science and environmental DNA. Marine Pollution Bulletin 110(1): 127-132.
Muñoz-Colmenero, M, Ardura, A, Clusa, L, Miralles, L, Gower, F, Zaiko, A & Garcia-Vazquez, E. 2018. New specific molecular marker detects Ficopomatus enigmaticus from water eDNA before positive results of conventional sampling. Journal for Nature Conservation 43: 173-178.
Ojaveer H, Galil BS, Lehtiniemi M, Christoffersen M, Clink S, Florin A-B, Gruszka P, Puntila R, Behrens J. 2015. Twentyfive years of invasion: management of the round goby Neogobius melanostomus in the Baltic Sea. Management of Biological Invasions 6:329-339.
Outinen,O, Puntila-Dodd,R, Barda,I, Brzana,R, Hegele-Drywa,J, Monta Kalnina,M, Kostanda,M, Lindqvist,A, Normant-Saremba,M, Ścibik,M, Strake,S, Vuolamo J, and Lehtiniemi, M. (2021) .The role of marinas in the establishment and spread of non-indigenous species in Baltic Sea fouling communities, Biofouling, DOI: 10.1080/08927014.2021.1996564
Pentinsaari, M, Ratnasingham, S, Miller, SE & Hebert, PDN. 2020. BOLD and GenBank revisited – Do identification errors arise in the lab or in the sequence libraries? PLOS ONE 15(4): e0231814.
Ratnasingham, S & Hebert, PDN. 2007. BOLD: The Barcode of Life Data System. Molecular Ecology Notes 7(3):355-364.
Rey, A, Basurko, OC & Rodriguez-Ezpeleta, N. 2020. Considerations for metabarcoding-based port biological baseline surveys aimed at marine nonindigenous species monitoring and risk assessments. Ecology and Evolution 10(5): 2452-2465.
Rinde E, Gitmark JK, Hjermann DØ, Fagerli CW, Røst Kile M, Christie H. 2017. Utvikling av metodikk for overvåkning av fremmede marine arter. NIVA-rapport 7131-2017.
Rosten, C. M. & Fossøy, F. 2020. Bruk av lyd til overvåking av norsk natur. En mulighetsstudie. NINA Rapport 1925. Norsk institutt for naturforskning.
Sandvik H, Dolmen D, Elven R, Falkenhaug T, Forsgren E, Hansen H et al. 2019. Alien plants, animals, fungi and algae in Norway: an inventory of neobiota. Biological Invasions 21: 2997–3012. http://dx.doi.org/10.1007/s10530-019-02058-x
Sepulveda, AJ, Nelson, NM, Jerde, CL & Luikart, G. 2020. Are environmental DNA methods ready for aquatic invasive species management? Trends in Ecology & Evolution 35(8): 668-678.
Skaala, Ø, Borgstrøm, R, Solberg, MF, Fiske, P, Hindar, K, Jensen, JA, Mo, TA, Sægrov, H, Barlaup BT. 2020. Regnbueørret (Onchorhynchus mykiss); produksjon og interaksjoner med stedegne arter. Rapport fra Havforskningen 2020-21. 22 s.
Stefanni, S, Stanković, D, Borme, D, de Olazabal, A, Juretić, T, Pallavicini, A & Tirelli, V. 2018. Multi-marker metabarcoding approach to study mesozooplankton at basin scale. Scientific Reports 8(1): 12085.
Taberlet, P, Coissac, E, Hajibabaei, M & Rieseberg, LH. 2012. Environmental DNA. Molecular Ecology 21(8): 1789-1793.
Tamburini M, Keppel E, Marchini, A, Repetto MF, Ruiz GM, Ferrario J and Occhipinti-Ambrogi A (2021). Monitoring Non-indigenous Species in Port Habitats: First Application of a Standardized North American Protocol in the Mediterranean Sea. Frontiers in Marine Science 8:700730. doi: 10.3389/fmars.2021.700730
Thalinger, B, Deiner, K, Harper, LR, Rees, HC, Blackman, RC, Sint, D, Traugott, M, Goldberg, CS & Bruce, K. 2021. A validation scale to determine the readiness of environmental DNA assays for routine species monitoring. Environmental DNA 3(4): 823-836.
Thomsen, PF, Kielgast, J, Iversen, LL, Møller, PR, Rasmussen, M & Willerslev, E. 2012. Detection of a diverse marine fish fauna using environmental DNA from seawater samples. PLoS ONE 7(8): e41732.
Thomsen, PF, Møller, PR, Sigsgaard, EE, Knudsen, SW, Jørgensen, OA & Willerslev, E. 2016. Environmental DNA from seawater samples correlate with trawl catches of subarctic, deepwater fishes. PLOS ONE 11(11): e0165252.
Thomsen, PF & Willerslev, E. 2015. Environmental DNA - An emerging tool in conservation for monitoring past and present biodiversity. Biological Conservation 183(0): 4-18.
Valentini, A, Taberlet, P, Miaud, C, Civade, R, Herder, J, Thomsen, PF, Bellemain, E, Besnard, A, Coissac, E, Boyer, F, Gaboriaud, C, Jean, P, Poulet, N, Roset, N, Copp, GH, Geniez, P, Pont, D, Argillier, C, Baudoin, J-M, Peroux, T, Crivelli, AJ, Olivier, A, Acqueberge, M, Le Brun, M, Møller, PR, Willerslev, E & Dejean, T. 2016. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Molecular Ecology 25(4): 929-942.
van den Heuvel-Greve, MJ, van den Brink, AM, Glorius, ST, de Groot, GA, Laros, I, Renaud, PE, Pettersen, R, Węsławski, JM, Kuklinski, P & Murk, AJ. 2021. Early detection of marine non-indigenous species on Svalbard by DNA metabarcoding of sediment. Polar Biology 44(4): 653-665.
Westfall, KM, Therriault, TW & Abbott, CL. 2020. A new approach to molecular biosurveillance of invasive species using DNA metabarcoding. Global Change Biology 26(2): 1012-1022.
Zaiko, A, Martinez, JL, Schmidt-Petersen, J, Ribicic, D, Samuiloviene, A & Garcia-Vazquez, E. 2015. Metabarcoding approach for the ballast water surveillance – An advantageous solution or an awkward challenge? Marine Pollution Bulletin 92(1): 25-34.
Zaiko, A, Schimanski, K, Pochon, X, Hopkins, GA, Goldstien, S, Floerl, O & Wood, SA. 2016. Metabarcoding improves detection of eukaryotes from early biofouling communities: implications for pest monitoring and pathway management. Biofouling 32(6): 671-684.
Zaiko, A, Pochon, X, Garcia-Vazquez, E, Olenin, S & Wood, SA. 2018. Advantages and Limitations of Environmental DNA/RNA Tools for Marine Biosecurity: Management and Surveillance of Non-indigenous Species. Frontiers in Marine Science 5(322).