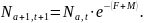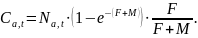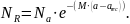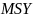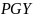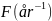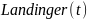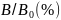Rapportserie:
Rapport fra havforskningen 2020-27ISSN: 1893-4536Publisert: 07.09.2020Oppdatert: 29.06.2024Prosjektnr: 15480Oppdragsgiver(e): HI Forskningsgruppe(r):
Bentisk økologiTema:
HaneskjellProgram:
Barentshavet og Polhavet Godkjent av:
Forskningsdirektør(er):
Geir Huse
Programleder(e):
Maria Fossheim og Elena Eriksen
Stock assessment of Iceland scallops (Chlamys islandica) in the Bear Island area
The Institute of Marine Research conducted an investigation of the standing stock and harvest potential of Iceland scallops (Chlamys islandica) around Bear Island and Spitsbergen Bank. Underwater video and a regular delta-dredge were used to collect data on shell density, size distribution and other biological data as input to an age-based population model for estimation of stock size and sustainable long term yield. The model outputs reveal a precautionary long term yield of approximately 15000 tons of scallops (round), which is presumed to be a conservative estimate.
Sammendrag
På bakgrunn av en bestilling fra Fiskeridirektoratet gjennomførte Havforskningsinstituttet en kartlegging av haneskjellressursene (Chlamys islandica) i Fiskevernsonen rundt Svalbard. Undersøkelsen ble konsentrert om områdene rundt Bjørnøya og Spitsbergenbanken.
Kartleggingen ble foretatt ved bruk av undervanns video og trekantskrape, og vi benyttet en aldersbasert populasjonsmodell for å beregne bestandsstørrelse og bærekraftig langtidsutbytte. Beregningene viser at bærekraftig langtidsutbytte fra de undersøkte feltene er på ca 15000 tonn rundskjell (skjell med skall og bløtdeler) årlig, og vi antar at dette er et konservativt estimat.
1 - Innledning
Med bakgrunn i deni nye interessen for fisket etter haneskjell fikk Havforskningsinstituttet en bestilling fra Fiskeridirektoratet om å foreta en ny kartlegging av statusen for bestanden av haneskjell i Fiskevernsonen rundt Svalbard.
De viktigste områdene for kommersiell utnyttelse av haneskjell (Chlamys islandica) har vært og er langs østkysten av Canada, Vest-Grønland og i Island (http://www.dfo-mpo.gc.ca/, Pedersen 1988, Shumway 1991). I Canada foregår det fortsatt et fiskeri etter dette skjellet, mens fisket på Island stanset i 2004 (Jonasson et al 2007). Her til lands ble haneskjell først og fremst fangstet på lokale felter i Troms og Finnmark (Venvik og Vahl 1979). Fangsten av haneskjell i Troms foregikk fra midten av 1970-tallet til ut på 1980-tallet og kvanta som ble landet var relativt små – mindre enn 200 tonn rundskjell hvert år (Norges råfisklag).
I 1985 tok fangsten av haneskjell i norske farvann nye dimensjoner ved at to havgående fartøyer ble utrustet for fiske etter haneskjell ved Jan Mayen. Samtidig ble det inngått kontrakter om både nybygg, ombygging av eksisterende fiskefartøy, samt innførsel av andre fartøytyper i Fartøyregisteret, for fiske etter haneskjell i norske farvann. Bakgrunnen for denne nye satsingen på haneskjell var at torskebestanden i Barentshavet var på et historisk lavmål samtidig med en kollaps i loddebestanden. I løpet av 1986 skjedde det en voldsom økning i antall fartøy og på slutten av året hadde i alt 26 større fartøy vært inne i dette fiskeriet.
Etter hvert flyttet fangstingen av skjell fra Jan Mayen til Fiskevernsonen rundt Svalbard, hvor felter ved Bjørnøya, Spitsbergenbanken og nord for Svalbard (Moffen) ble beskattet (Sundet & Rubach 1988, Opstad 1988). Kunnskapen om haneskjellressursene både ved Jan Mayen og i Fiskevernsonen rundt Svalbard var svært mangelfull på dette tidspunktet, og det ble derfor satt i gang en storstilt kartlegging av skjellressursene i norsk sone samtidig med at fisket foregikk. I denne kartleggingen ble det funnet store forekomster av haneskjell flere steder. I tillegg til feltet ved Jan Mayen (Straumflaket) ble det funnet betydelige skjellfelter ved Bjørnøya, Spitsbergenbanken og nord for Svalbard (Rubach og Sundet 1987, Sundet og Rubach 1988). Usikre anslag den gang indikerte at den stående bestanden av haneskjell i disse områdene kunne være flere hundre tusen tonn (Rubach og Sundet 1987).
Det var store havgående fartøy som deltok i dette fisket; ombygde og nybygde trålere, ringnotfartøy og andre typer fartøyer. Fangstredskapen var store skjellskraper (flere tonn) av jern med et nett av jernringer, og hvert fartøy opererte gjerne tre skraper samtidig. Alle skjell som ble fanget ble produsert ombord og hovedproduktet var frossen muskel av forskjellig størrelse. De totale landingene av frossen muskel fra dette fiskeriet nådde et maksimum på ca 4000 tonn i 1987, men minket raskt til noen hundre tonn i de påfølgende årene og 1992 var siste år med fangster fra norske skjellskrapere.
Det ble ikke gjennomført undersøkelser av hvilke effekter denne skrapeaktiviteten hadde på bunnfauna og sediment, men det var liten tvil om at påvirkningene var store. Størsteparten av det som kom opp med skrapene (stein, tomskall o.l.) ble kastet ut igjen og en av observasjonene vi gjorde var blant annet at felter som tidligere hadde en flat og slett bunn, i ettertid ble kjennetegnet av at store steiner lå oppå sedimentoverflaten. Videofilmer av bunnen viste også store furer i bunnen etter skrapingen siden bunnen i stor grad besto av en blanding av mudder, stein og tomskall.
Den store investeringen i haneskjellfisket på 1980-tallet førte til en kraftig overbeskatning av denne ressursen og alle skjellfelt ble nedfisket i løpet av kort tid. Fangstteknologien i seg selv bidro også til en lite effektiv utnyttelse av denne ressursen i og med at mye skjell ble knust og ødelagt i fangstprosessen. I tillegg førte den sterke begroingen av skjellene, spesielt ved Bjørnøya, til at de ikke lot seg produsere i fabrikkene ombord. Store mengder skjell ble derfor drept ned og kastet over bord uten å bli produsert.
De fleste fartøyene var inne i dette fiskeriet kun i kort tid og det var de spesialbygde skjellskraperne som holdt ut lengst. Betydelige overinvesteringer i ombygginger og fabrikkanlegg førte også til at en rekke fartøyer og rederier gikk konkurs.
I 2017 fikk Havforskningsinstituttet en henvendelse fra firmaet TauTech as om å delta i en utprøving av et nytt fangstredskap for haneskjell. Uttestingen av dette redskapet var et ledd i et prosjekt i flere faser hvor målet var å starte fangst av haneskjell i Fiskevernsonen rundt Svalbard. Første fase som besto i å undersøke effekten av dette nye fangstredskapet på bunnfauna og sediment på haneskjellfelt, er gjennomført og rapportert (Sundet et al 2019).
Før et eventuelt prøvefiske etter haneskjell kunne starte var en avhengig av å vite tilstanden til haneskjellressursene i Fiskevernsonen; om bestanden av skjell hadde tatt seg opp igjen etter den harde beskatningen på 1980-tallet. Det er også kjent at i perioden fram til i dag har det skjedd endringer i havmiljøet i dette området ved at det har foregått en oppvarming av hele Barentshavet (Fossheim et al 2015). Dette kan ha påvirket både rekruttering, vekst og utbredelse av haneskjellressursene i Fiskevernsonen.
2 - Haneskjellets biologi
Haneskjellet finnes utbredt langs hele kysten av Nord-Norge, med de største forekomstene i Troms og Finnmark (Wiborg 1963). I tillegg er det også store forekomster av haneskjell på østkysten av Canada, vestkysten av Grønland, vestkysten av Island, ved Jan Mayen og i Fiskevernsonen rundt Svalbard (http://www.dfo-mpo.gc.ca/, Pedersen 1988, Eriksson 1988, Rubach & Sundet 1987).
Haneskjellet ser ut til å trives best på noe varierende dyp; på kysten av Troms og Finnmark fra 20 - 60 m (Wiborg 1963), mens ved Bjørnøya fant en de høyeste konsentrasjonene mellom ca 60 – 100 m dyp (Rubach & Sundet 1987). Skjellet finnes vanligvis på bunnsubstrat av sand, skjell eller grov grus og oftest i bakkeskråninger, gjerne i tilknytning til områder med sterk strøm (Ekman 1953). På de enkelte feltene er skjellet svært flekkvis fordelt og som regel finner en områder med høy tetthet spredt rundt på det som defineres som et felt. Disse små områdene kan ha varierende utstrekning og mellom dem er det lavere tetthet eller ingen skjell. Selve skjellfeltet avgrenses gjerne av forhold som dybde, bunnsubstrat og strømforhold.
Sammenlignet med mer sørlige arter av kamskjell vokser haneskjellet langsomt og det tar om lag 6 - 8 år før skjellet når en størrelse på 60 mm skallhøyde som er minstemålet i fiskevernsonen. Veksten varierer fra område til område og ser ut til i større grad å være bestemt av næringstilgangen, enn av temperatur (Sundet & Vahl 1981). Ved kysten har haneskjellet en årlig skall-tilvekst på ca 10 mm fram til det blir 6 – 7 år gammel. Da avtar veksten etter hvert, og gamle skjell vokser omtrent ikke i skallstørrelse (Vahl 1981).
I motsetning til flere andre vanlige kamskjell er haneskjellet særkjønnet, og hanner og hunner har forskjellig farge på gonaden (Skreslet & Brun 1969). I kystpopulasjonene i Troms ser gytingen ut til å foregå epidemisk i månedsskiftet juni – juli og befruktningen skjer i de frie vannmassene (Skreslet 1973, Gruffydd 1976). Larvene er pelagiske i ca to måneder før de bunnslår, gjerne på grunne områder (Wallace 1982).
3 - Undersøkelsesområdet
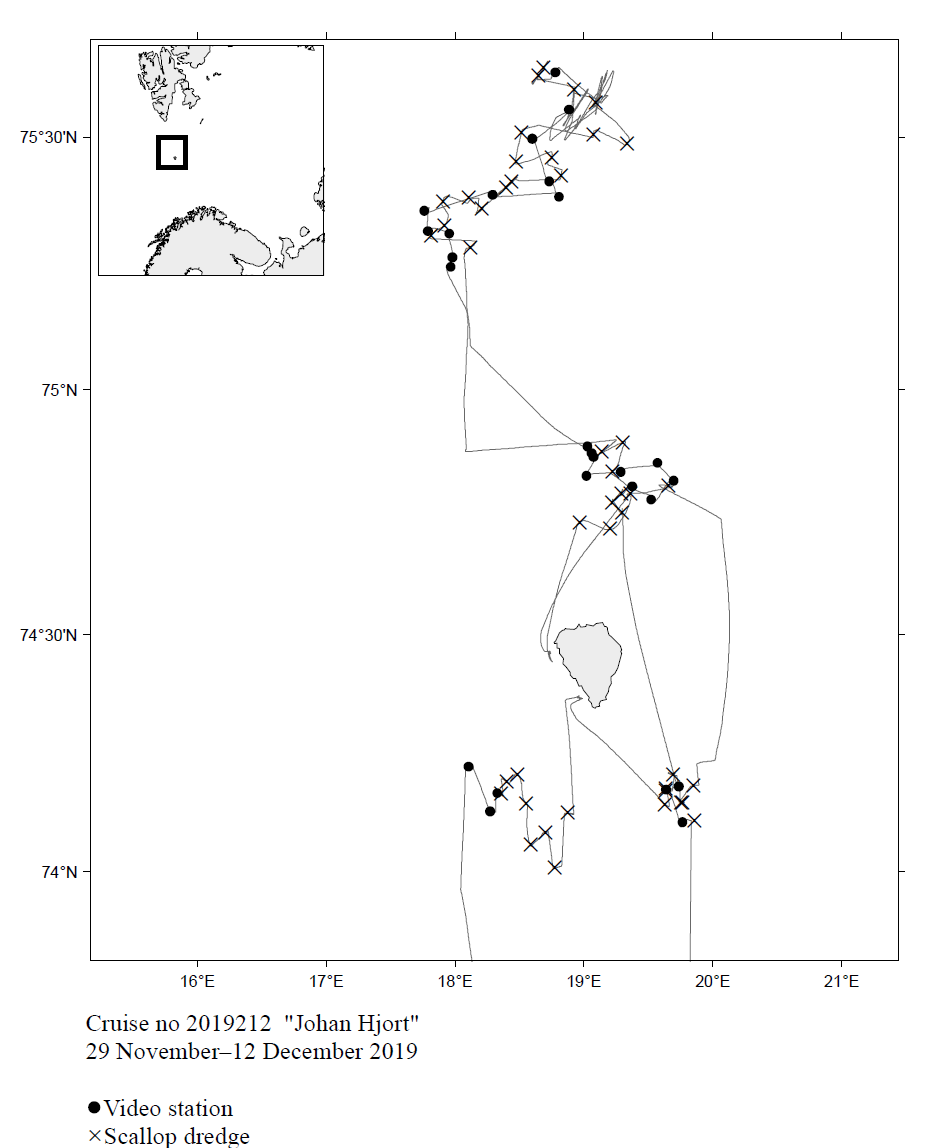
Feltene rundt Bjørnøya som ble undersøkt i desember 2019 er vist på kartet i Figur 1 sammen med posisjoner for trekant-skrapetrekk og videostasjoner.
Haneskjellfeltene ved Bjørnøya ble første gang beskrevet av Wiborg (1963) og senere av Venvik og Vahl (1979). Ved den omfattende kartleggingen i 1986 ble feltene undersøkt på nytt i tillegg til at feltenes utstrekning ble noe utvidet (Rubach og Sundet 1987). I ettertid er grensene for territorialfarvannet rundt Svalbard utvidet fra 4 til 12 nautiske mil, noe som innebærer at deler av skjellfeltene, spesielt på nordsiden av Svalbard, i dag ligger innenfor verneområder og er ikke tilgjengelig for fangst. Dette gjelder i liten grad for feltene ved Bjørnøya.
Skjellfeltene i dette området ligger i nær tilknytning til Polarfronten, overgangen mellom arktisk og atlantisk vann, og feltene nord for Bjørnøya blir enkelte år dekket av is. Dette er svært strømrike områder som i stor grad styres av tidevannet. Beliggenheten i Polarfrontområdet og ved iskanten, gjør at dette er et viktig område for primærproduksjon, noe som kan være grunnen til at det finnes mye haneskjell i dette området.
Figur 1. Viser kurslinjer og stasjoner med videoslede og trekantskrape fra toktet med FF Johan Hjort i desember 2019.
4 - Metoder
4.1 - Innsamling på tokt og bearbeidelse i laboratoriet
Innsamlingen av data foregikk på et tokt med FF Johan Hjort i tidsrommet 29.11 – 12.12 i 2019. Alle posisjoner for prøvestasjoner for både skrapetrekk og videoslede ble satt tilfeldig ut på forhånd, basert på data fra 1980-tallet når det gjelder forutsetninger om utbredelse av feltene.
I løpet av toktet ble det gjennomført i alt 44 trekk á 5 minutter med trekantskrape og 26 trekk á 60 minutter, med videoslede. Dårlige vær med mye kuling gjorde det ganske utfordrende å få et godt billedmateriale ved bruk av videosleden, som opprinnelig er konstruert for telling av krabbe og går med en viss avstand fra bunnen. Alle opptak fra video ble lagret på disker med sikkerhetskopier for videre bearbeidelse på land.
Innholdet i skrapetrekkene ble sortert på dekk og alle haneskjellene ble målt for skallhøyde. I tillegg ble det tatt prøver av skjell i forskjellige størrelse fra hvert av skjellfeltene for aldersbestemmelse i laboratoriet på land. Alderslesingen ble gjort som beskrevet av Johannessen (1973). Lukkemuskelen av skjell større enn 60 mm skallhøyde (minstemålet) fra to av feltene (Kveitehola og Concordia), ble dissekert ut og veid, og antall pr kg beregnet. I tillegg beregnet en forholdet mellom muskelvekt og skallhøyde.
En gjennomgang av videomaterialet i etterkant viste at det var bare deler av opptakene som kunne brukes i bestandsberegningene. Store deler av opptakene var svært uklare på grunn av marin snø og av bevegelsene til fartøyet. Dette gjorde det utfordrende å kunne se og telle enkeltskjell på bunnen, i tillegg til å skille mellom døde og levnede skjell. En må derfor anta at en del skjell ikke ble telt, noe som bidrar til et konservativt bestandsestimat.
4.2 - Bestandsberegninger
Skraping er en lite egnet metode for estimering av bestanden av skjell på grunn av at arealet som dekkes av hvert enkelt skrapetrekk kan variere mye selv om tiden for hvert trekk holdes konstant. Derfor ble registreninger av skjell fra videopptak benyttet til bestandsberegningene, mens data skrapeprøvene ble brukt til å beregne størrelsesfordeling, muskelvekt og utbytte fra de tre skjellfeltene. Den reduserte kvaliteten av videofilmene bidro til at tellingene av skjell ble begrenset til å benytte stillbilder fra filmopptakene. På hver stasjon ble det gjort filmopptak av ca. en times varighet, og fra hver slik film ble det plukket ut stillbilder med tilstrekkelig billedkvalitet til å telle enkeltskjell. Dette resulterte i at mellom 32 og 285 (gjennomsnittlig 122) enkeltbilder ble telt fra hver videostasjon. I den videre databehandlingen antok vi at hvert bilde utgjorde en tilfeldig, representativ delprøve av hele arealet som ble dekket på hver videostasjon. Hvert bilde representerer et areal på 0.65 × 0.65 m. To alternative modeller ble anvendt til å beregne bestandsstørrelsen: i modell 1 ble bestandsstørrelsen beregnet ut fra gjennomsnittlig skjelltetthet per bilde (0.65 × 0.65 m) på alle stasjonene for hvert felt, multiplisert med arealet av feltet. I modell 2 brukte vi skjelltettheten per bilde per felt til å modellere tetthet og totalantall skjell per felt. Modell 2 baserer seg på en antagelse om at hvert bilde er en uavhengig prøve fra feltet og at bildene fra en og samme stasjon dermed er uavhengige av hverandre. Hvert bilde representerer dermed en tilfeldig prøve fra hele feltet. Fordelen med denne modellen er at alle bildene kan analyseres som del av en fordeling, som gir et større antall uavhengige datapunkter enn om alle bildene fra hver stasjon er knyttet til hverandre.
De undersøkte skjellfeltene ble stratifisert i ettertid (post-stratifisering) for å inkludere innsamlet informasjon om forekomst og tetthet fra toktet sammen med dyp. Stratasystemet er derfor basert på alle skrape- og videostasjoner som er tatt innenfor 50 og 100 m dybdekonturene. Polygoner ble laget for hvert felt basert på disse stasjonene, mens de detaljerte grensene for polygonene følger dybdekotene. Dette sikrer at polygonene kun består av områder hvor vi har prøver fra. For å utforske mulige konsekvenser og bias av den anvendte post-stratifiseringen testet vi også et grid-system som bestod av identiske kvadrater (0.2 lengdegrader x 0.05 breddegrader) som grunnlag av bestandsberegningen. Her ble beregningene basert på de observerte gjennomsnittstetthetene per kvadrat, mens tettheten i kvadratene uten stasjoner ble ekstrapolert fra de omliggende kvadrater med observasjoner. Fordelen med grid-systemet er at her benyttes ingen a priori antagelser om utbredelsesområdet og kan derfor redusere mulig bias som følge av en bevisst stratadefinisjon. Ulempen er at antallet stasjoner og den tilfeldige fordelingen av stasjonene på 2019-toktet gjør at en ikke har stasjoner i alle grid-kvadratene selv om oppløsningen i grid-systemet er relativt lav med tanke på det komplekse og flekkvise fordelingsmønsteret av haneskjell.
4.3 - Modellestimater av langtidsutbytte
Vi benyttet en aldersbasert populasjonsmodell for å simulere mulige høstingsscenarier og til å beregne et bærekraftig langtidsutbytte. I modellen er endringen i antall skjell fra ett år til det neste et produkt av antall skjell per aldersklasse Na , og overlevelsesraten som er definert av fiskedødeligheten F og naturlig dødelighet M :
Fangst av antall skjell per aldersklasse og år Ca,t , er derfor beregnet som fiskeridødelighetens andel av alle skjell som dør i et år:
Alder ved rekruttering til fisket er satt til 7 år; den alderen haneskjell har ved skallhøyde 60 mm som er minstemålet, basert på alders-lengde-sammenhengene vist i pkt 5.3. Vi antar at alle aldersklasser som er 7 år og eldre er fullstendig rekruttert til fisket og at alle skjell er kjønnsmodne. Gytebestanden B er derfor summen av alle skjell eldre enn 7 år, og maksimumsalderen ble satt til 30 år.
Modellen inkluderer usikkerheter i utgangsbestanden (B0), naturlig dødelighet (M) og i rekrutteringen, og vi benytter bestandsstørrelsen og variasjonen i estimatene vist i pkt 4.2. som utgangsbestand. Data fra toktet ble brukt til å beregne alderssammensetningen i utgangsbestanden.
Naturlig dødelighet (M) i haneskjellbestandene i Fiskevernsonen er ikke kjent. Derfor benyttet vi et mål på naturlig dødelighet for haneskjellbestander fra østkysten av Canada (Naidu 1988). Denne undersøkelsen avslørte en relativ stor variasjon i M (0.02 til 0.2) for forskjellige områder og bestander, og det er uklart om denne variasjonen skyldes forskjell mellom lokale populasjoner, estimeringsusikkerhet eller variasjoner på grunn av forskjellig fiskepress. Uansett gir det et godt grunnlag for en nedre og øvre grense for M i modellen. For å ta høyde for den store usikkerheten i M, og for å teste mulige effekter av forskjellige fangstuttak, ble en tilfeldig verdi mellom 0.02 og 0.2 vektet med frekvensen i Naidu (1988) for M, benyttet ved hver simulering.
Den andre sentrale parameteren og store usikkerhetskilden er rekruttering. Siden vi ikke har noen tidsserie i overvåkingen av haneskjellbestandene, har vi heller ingen kunnskap om rekrutteringen og mulige sammenhenger med størrelsen på gytebiomassen eller med miljøvariasjoner. Vi benyttet derfor alderssammensetningen fra toktdata som utgangspunkt for å tilbakeberegne antall skjell per aldersklasse, ved hjelp av følgende modell:
Modellen beskriver en eksponentiell nedgang i antall skjell fra rekrutteringsalder arec til alder a som samsvarer med en dødelighetsrate M per år. Denne kan brukes til å tilbakeregne rekruttering NRfor alle aldersklasser fra 7 til 22 år (fra rekrutteringsalder til høyeste aldersklasse med nok skjellprøver), og det gir et estimat av hvor mange skjell som er rekruttert til bestanden i løpet av en 15 årsperiode. Dette indikerer gjennomsnittet og variasjonen i rekrutteringen. Fordelingen i rekruttering hvert år er et produkt av alderssammensetningen, bestandsestimatet og M(Figur 2). Den årlige rekrutteringen endrer seg i hver simulering, fordi et tilfeldig utvalg av bestandsestimater og Mdefinerer et gjennomsnitt og et standardavvik av en (log-transformert) rekrutteringsfordeling: Narec=η(μR, σR2). Dette er grunnlaget for å beskrive den stokastiske rekrutteringsdynamikken i bestanden.
Forholdet mellom rekruttering og gytebestand i de undersøkte haneskjellpopulasjonene er ikke kjent og kan derfor ikke parametriseres. Haneskjell har et høyt reproduksjonspotensiale i og med at de produserer mange egg pr individ, men en må likevel anta at det er en nedre og øvre terskel hvor rekrutteringen reduseres på grunn av gytebestandens størrelse. Lav gytebestand kan bidra til at mengden egg og larver blir for lav til å sikre rekruttering. En stor bestand av voksne haneskjell kan føre til tetthetsregulerende effekter ved at tilgjengelig substrat blir for lite (Vahl 1982). Ved setling har haneskjellarven spesifikke krav til substratet for å kunne feste seg og vokse opp (stein, tomskall etc.). Siden haneskjellet er en filtereter er det også tenkelig at en stor bestand kan predatere på egne larver og egg. Vi mangler tilstrekkelige data til å lage en gytebestand-rekrutteringsmodell og valgte derfor å benytte en «dobbelt hockeykølle»-sammenheng mellom gytebestand og rekruttering (Figur 3).
Dette innebærer at tilfeldig rekruttering er normalfordelt η (μR,σR )2dersom gytebestanden Bet år er lavere enn startbestanden B0 og høyere enn en andel av B0 som ligger imellom null og halv av startbestanden. I mange fiskebestander antas at den nedre grensen før bestandssstørrelsen påvirker rekrutteringen negativt, ligger på ca 30 % av en ufisket bestand. Denne nedre grensen defineres i fiskeriforvaltningen som den bestandsstørrelsen en ikke må falle under Blim. Siden vi mangler kunnskap om rekruttering i haneskjellbestandene i Fiskevernsonen, antar vi at bestanden kan ligge mellom 20 og 50 % av ufisket bestand, og i hver simulering ble en tilfeldig verdi mellom disse to verdier utvalgt. Dette følger de samme prinsippene som ved simulering av M, og bidrar til å kvantifisere usikkerheten og følsomheten i modellresultatene.
Vi gjennomførte i alt 10000 simuleringer over en periode på 50 år slik at resultatene inneholdt alle kombinasjoner av parameterverdier i et stort nok antall. Ut fra dette kan det etableres to referanseverdier til bruk i forvaltningen av bestanden: fiskedødeligheten FMSYsom maksimerer langsiktig fangst (MSY maximum sustainable yield») og FPGY, den laveste fiskedødeligheten som resulterer i et langtidsutbytte som er høyere enn 80% av MSY, det såkalte «pretty good yield» PGY («ganske bra utbytte») (Hilborn 2010, Rindorf et al 2016). FMSYvil maksimere utbyttet over tid og kan være problematisk i forhold til et føre-var-prinsipp, spesielt hvis usikkerhet er stor. Derfor er det viktig å sammenligne FMSY med en referanseverdi som er mer konservativ og som kombinerer forskjellige målsetninger; f. eks. et høyt utbytte som både ivaretar bærekraftighet og føre-var-prinsippet. Vi brukte her FPGYfordi det resulterer i et langtidsutbytte som er sammenlignbart med FMSY, men reduserer risikoen for å falle under Blim, og rekrutteringen reduseres. Beregningene inkluderer derfor også de bestandsstørrelsene som vil gi en MSY og PGY (BMSYog BPGY) og sannsynligheten for at bestanden vil ligge under bestandsstørrelsen Blim.
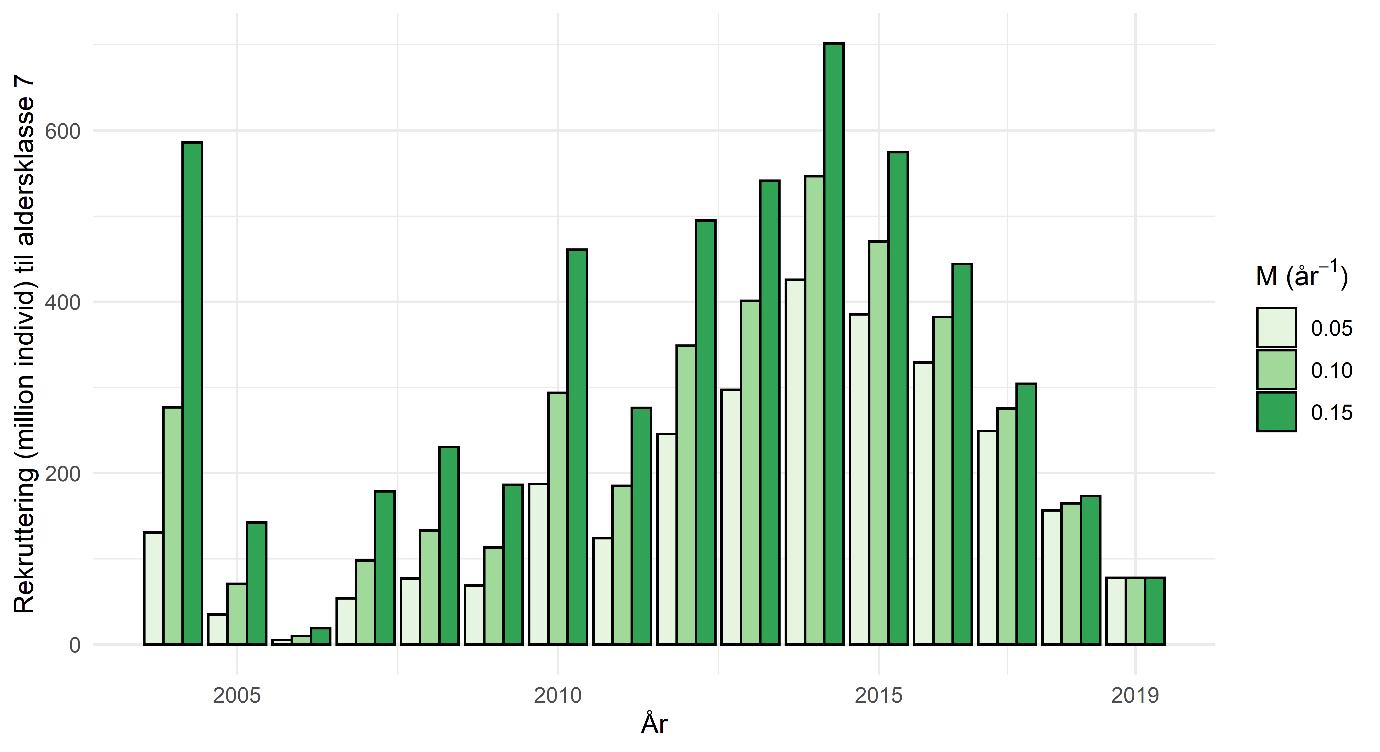
Figur 2. Tilbakeregnet rekruttering av 7 år gamle skjell, per år mellom 2004 og 2019, for tre forskjellige verdier av naturlig dødelighet (M). En høy M betyr at flere skjell dør hvert år og derfor øker de tilbakeregnete kohortstørrelsen jo høyere M er, og jo lengere tilbake i tid deres rekrutteringsalder ligger.
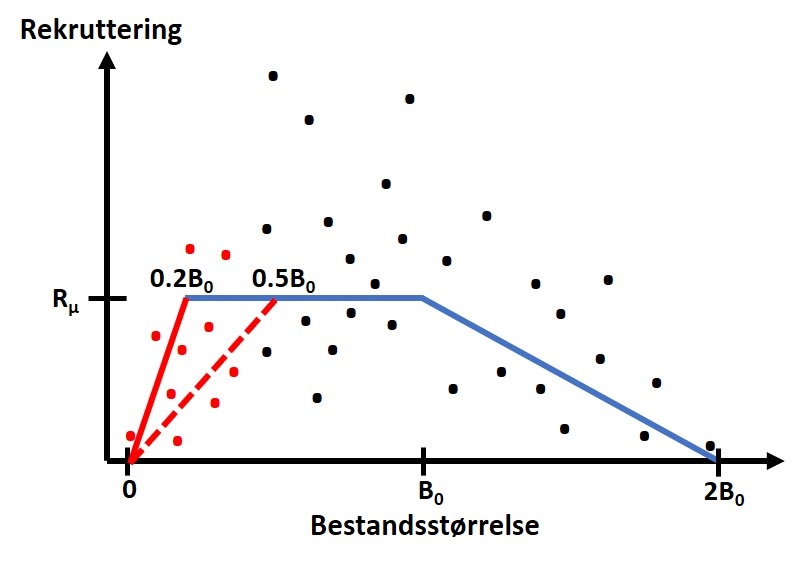
Figur 3. Illustrasjon av forholdet mellom rekruttering og bestandsstørrelse. Rekrutteringen blir redusert når størrelsen på bestanden minker eller går under en terskelverdi (50 % stiplet rød linje, og 20 % rød heltrukken linje). Rekrutteringen hvert år er en tilfeldig verdi utfra en fordeling som baseres på empiriske estimater av en gjennomsnittrekruttering Rμ og en variasjon eksemplifisert her som svarte og røde punkter (tilsvarende til rekruttering over og under terskelverdien).
5 - Resultater
5.1 - Utbredelse
På toktet i 2019 ble det tatt langt færre prøver enn i 1986 og 1987, men stasjonene i 2019 representerer en tilstrekkelig randomisert dekning av den antatte utstrekningen av de tre feltene (Figur 9). Forekomsten av skjell i 2019 viser at tetthet og mengde i stor grad ligner det som ble registrert i 1986/87, men det finnes også forskjeller. Spesielt gjelder dette området sørvest av Bjørnøya hvor det ble registrert betydelige mengder haneskjell i 1986, men kun få registreringer i 2019. På Kveitehola-feltet var registreringene av skjellmengde i 2019 på samme nivå som i 1986/87, mens de registrerte mengdene av skjell i skrapetrekkene var noe høyere i 2019 på Concordia-feltet og på feltet sørøst av Bjørnøya.
Observasjoner på toktet i 2019 bekrefter også at tettheten av haneskjell er veldig ujevnt og flekkvis fordelt på de enkelte feltene. Noe som innebærer at det er vanskelig å sammenligne stasjoner fra de to tidsperiodene som ikke ligger på nøyaktig samme plass. Forskjell mellom stasjoner og år kan derfor være tilfeldig selv om avstanden mellom stasjonene er liten.
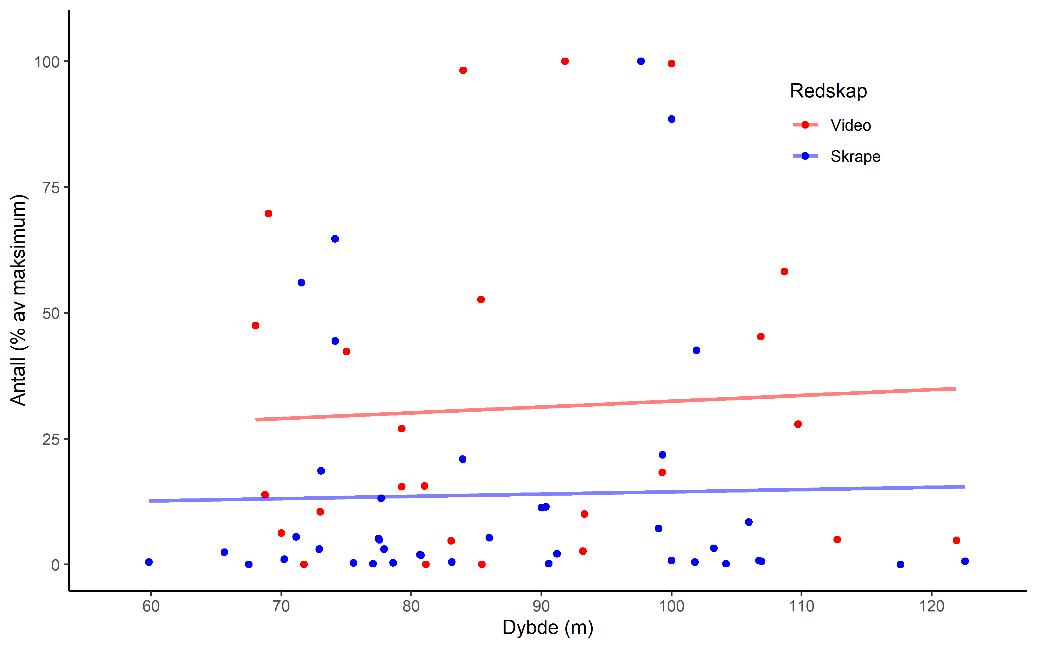
Skrape- og videostasjonene viser at selv om variasjonen i mengden registrerte skjell er veldig høy er det et likt mønster ved at høye antall i skrapetrekkene gjenspeilte seg i høye tettheter på videoopptakene fra samme område (Figur 5). Dataene fra toktet i 2019 viser ingen klar sammenheng mellom skjelltetthet og bunndybde i dybdeintervallet for undersøkelsen (60 – 125 m) (Figur 5), men stasjoner med store registeringer (>50% av maksimum) er lokalisert mellom 70 og 110m, med de høyeste antallene (>75%) mellom 80 og 100m. Dette bekrefter i stor grad registreringene som ble gjort i 1986 og 1987 hvor de høyeste tetthetene av skjell ble gjort i dybdeområdet 70 – 110 m dyp (Rubach og Sundet 1987). Resultatene fra undersøkelsene i 2019 og i 1986/87 viser at bunndybde indikerer en tilstrekkelig avgrensning av haneskjellfeltene, men en finner ingen gradienter i tetthet innenfor de gitte dybdegrensene.
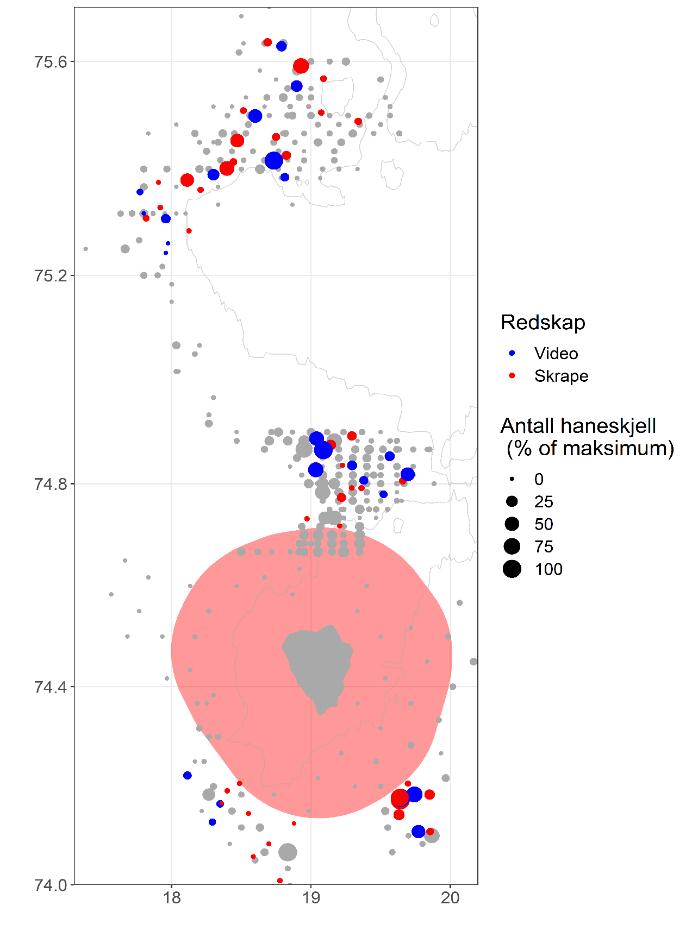
Figur 4. Oversiktskart over alle stasjoner på tidligere tokt i 1986 og 1988 (grå) og i 2019 (blå=videostasjon, rød=skrapestasjon). Punktene viser relativt antall registrerte skjell (prosent av maksimum registrerte fangster på alle skrapestasjoner eller videostasjoner). Grå linjer indikerer 50 og 100 meter dybdekoter og det rosa polygonet viser utstrekningen av naturvernområdet rundt Bjørnøya.
Figur 5. Relativt antall skjell (% av maksimumsfangst) i skrape-(blå) og på videostasjoner (rød) i forhold til median-bunndybde fra alle de undersøkte feltene. Punktene viser data fra hver stasjon, linjer er estimerte trender ved bruk av en glattingsmodell.
5.2 - Bestandsstruktur
a)
b)
c)
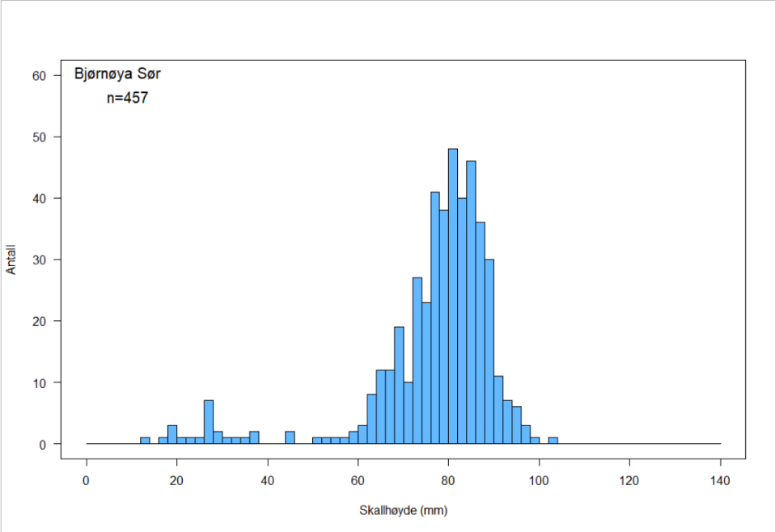
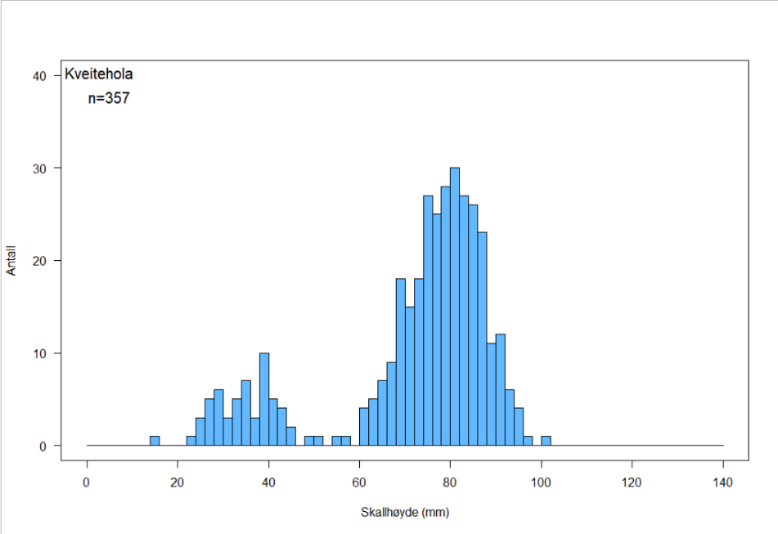
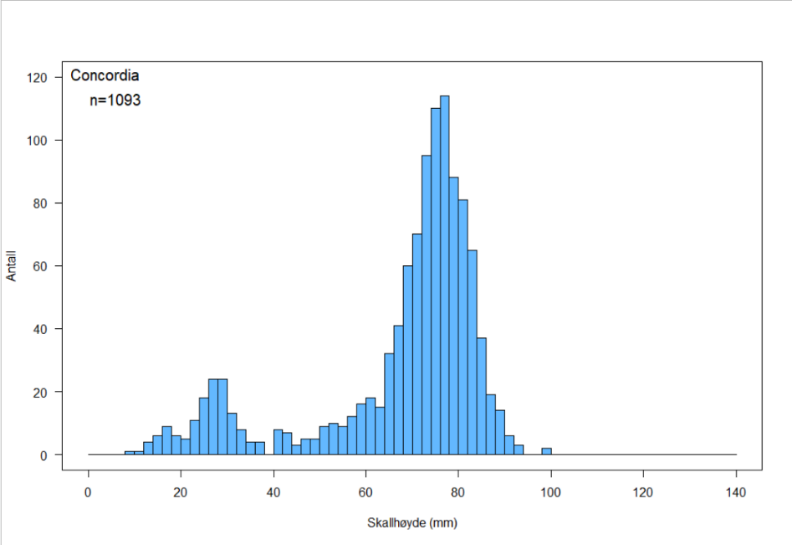
Figur 6. Skallhøydefordeling hos haneskjell fra Bjørnøya Sør (a), Kveitehola (b) og Concordia (c)
Skallhøydefordelingen på de tre feltene i Figur 6 a – c viser at alle tre skjellfeltene domineres av haneskjell med skallhøyde større enn ca 65 mm. Det kan se ut til at skjellene på Bjørnøya Sør og Kveitehola når en høyere maksimumsstørrelse enn de på Concordia, og at gjennomsnittsstørrelsen av fangstbare skjell (> 60 mm) er høyere på disse feltene.
Figurene viser også at det finnes en god del små haneskjell mellom 20 og 40 mm skallhøyde på disse feltene. Spesielt var forekomstene av småskjell høye på feltene Kveitehola og Concordia. Erfaringer fra tidligere undersøkelser ved bruk av skjellskraper viser at små skjell vanligvis er svært underrepresentert i prøvene. Dette kan skyldes at små haneskjell vanligvis er festet til tomskall av samme art og gjerne inne i såkalte cluckers. Dette gjør dem vanskeligere å finne under sorteringsarbeidet på dekk. I tillegg er ikke maskevidden i skrapenettet liten nok til å fange de aller minste skjellene.
Det ble registrert svært få cluckers på alle tre feltene. Totalt fant en 13 cluckers med fire som høyest antall på en skrapestasjon. Dette indikerer at den naturlige dødeligheten i haneskjellpopulasjonene på disse tre feltene er svært lav.
5.3 - Muskevekt og alder
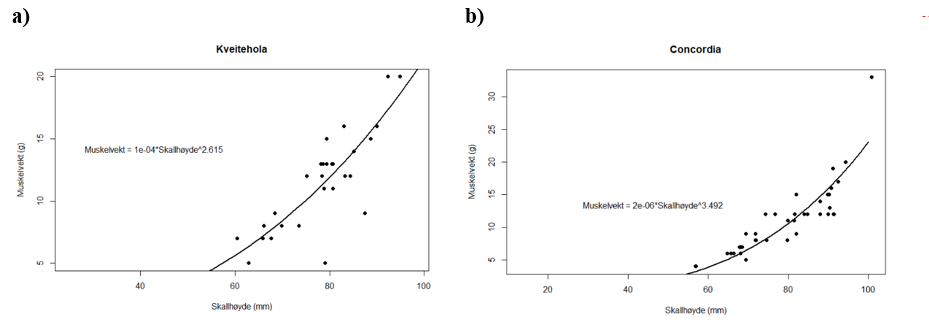
Forholdet mellom vekt av lukkemuskel og skallhøyde hos haneskjell fra Kveitehola og Concordiafeltet feltene er vist i Figur 7 a og b. I tillegg vises parametrene i funksjonen som beskriver forholdet mellom muskelvekt og skallhøyde beregnet fra modellen Muskelvekt = a*skallhøydeb .
Ut fra parametrene i modellen og figurene kan det se ut til at muskelvekten øker raskere med økt skallhøyde på Concordiafeltet enn i Kveitehola. Det ble ikke gjort registreringer av muskevekter fra Bjørnøya.
Figur 7. Forholdet mellom vekt av lukkemuskel og skallhøyde hos haneskjell fra Kveitehola (a) og Concordiafeltet.
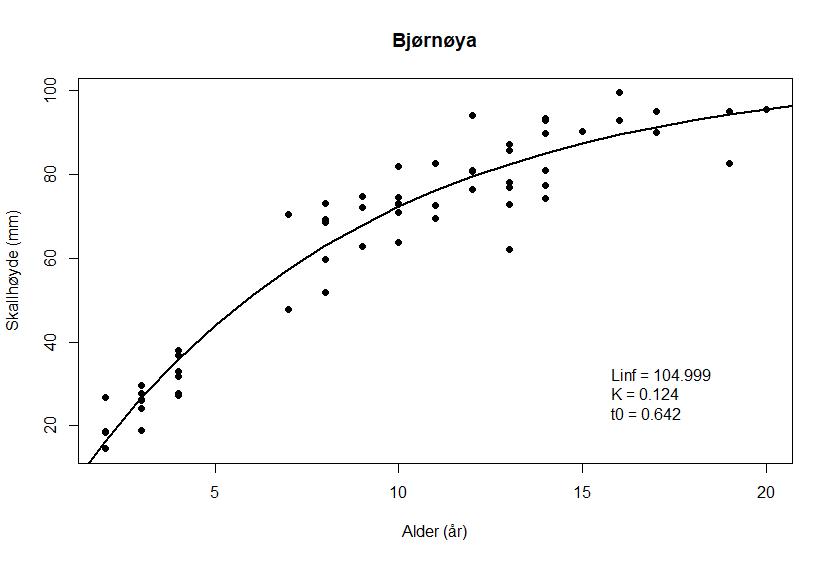
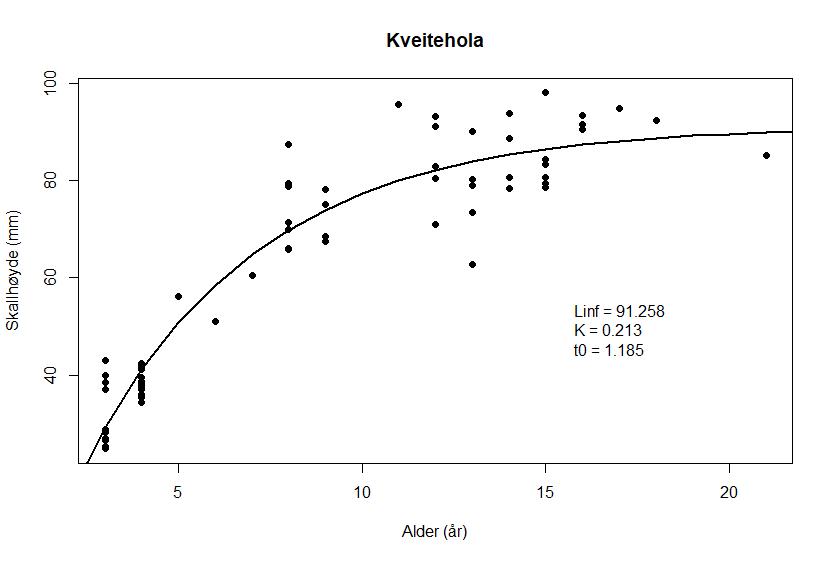
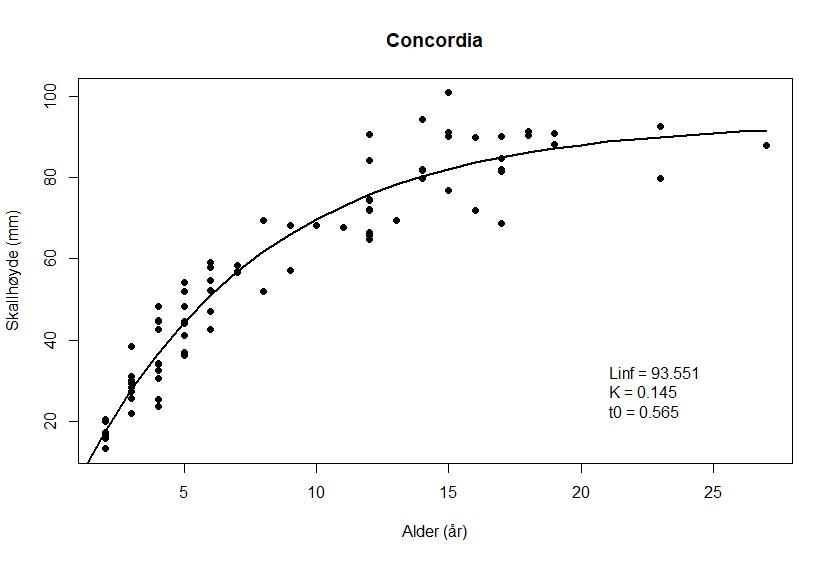
Forholdet mellom skallhøyde og alder hos haneskjell fra de tre feltene er vist i Figur 8 a – c
b)
c)
Figur 8. von Bertalanffy vekstfunksjon hos haneskjell fra feltene Bjørnøya sør (a), Kveitehola (b) og Concordia (c). Punktene angir observasjonene fra alderslesingen.
5.4 - Bestand- og biomasseestimater
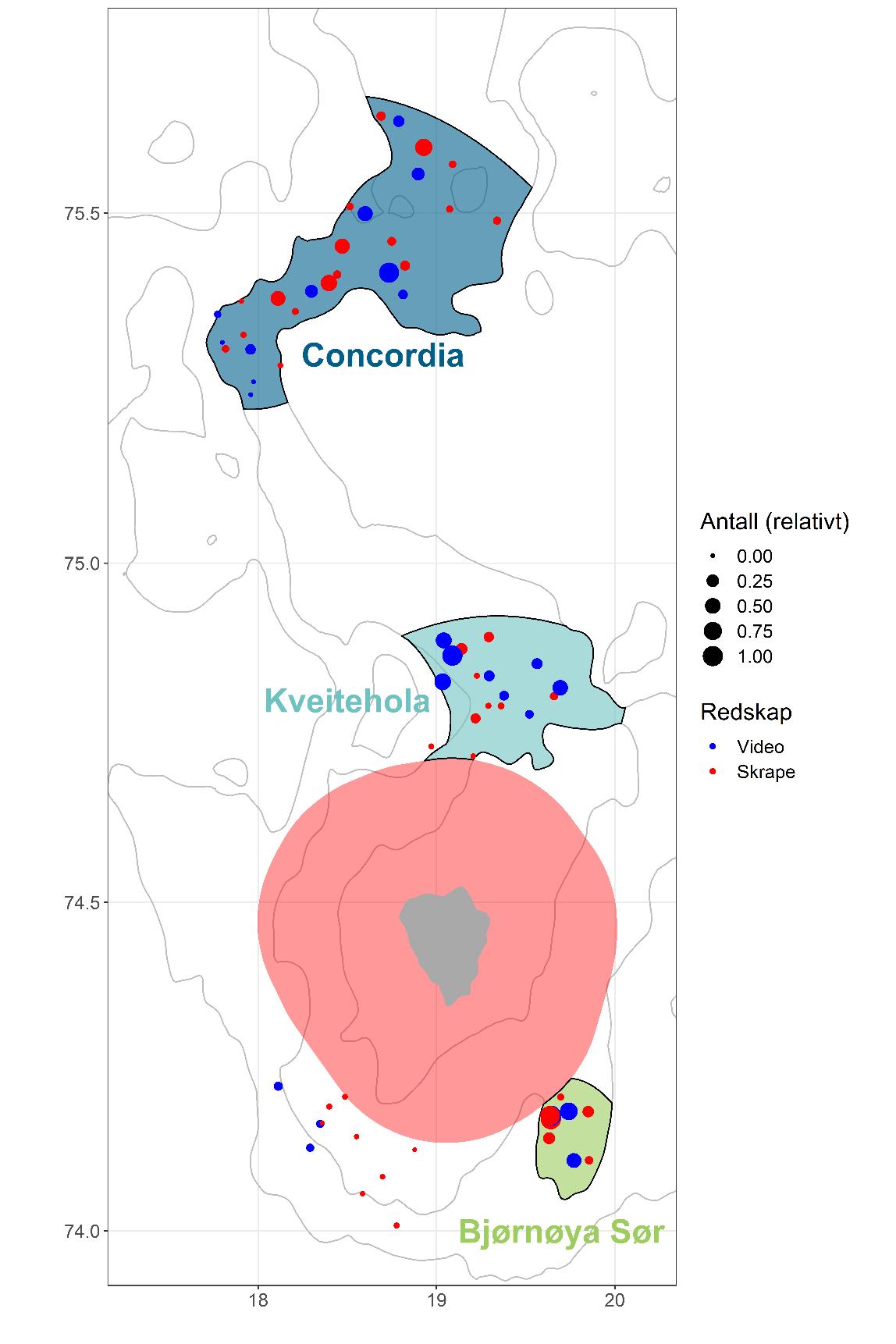
Basert på observasjonene på toktet i 2019, ble det besluttet at bestandsestimeringen skulle foretas for tre felter separat: sørøst av Bjørnøya (heretter kalt «Bjørnøya Sør»), nordøst av Bjørnøya («Kveitehola»), og et område på Spitsbergenbanken («Concordia»). Området sørvest av Bjørnøya ble ekskludert fra bestandsestimeringen fordi vi ikke fant forekomster vurdert som kommersielt interessante. Basert på metodene beskrevet under kap. 4.2. definerte vi et system med tre strata hvor hvert stratum avgrenser de enkelte feltene (Figur 9).
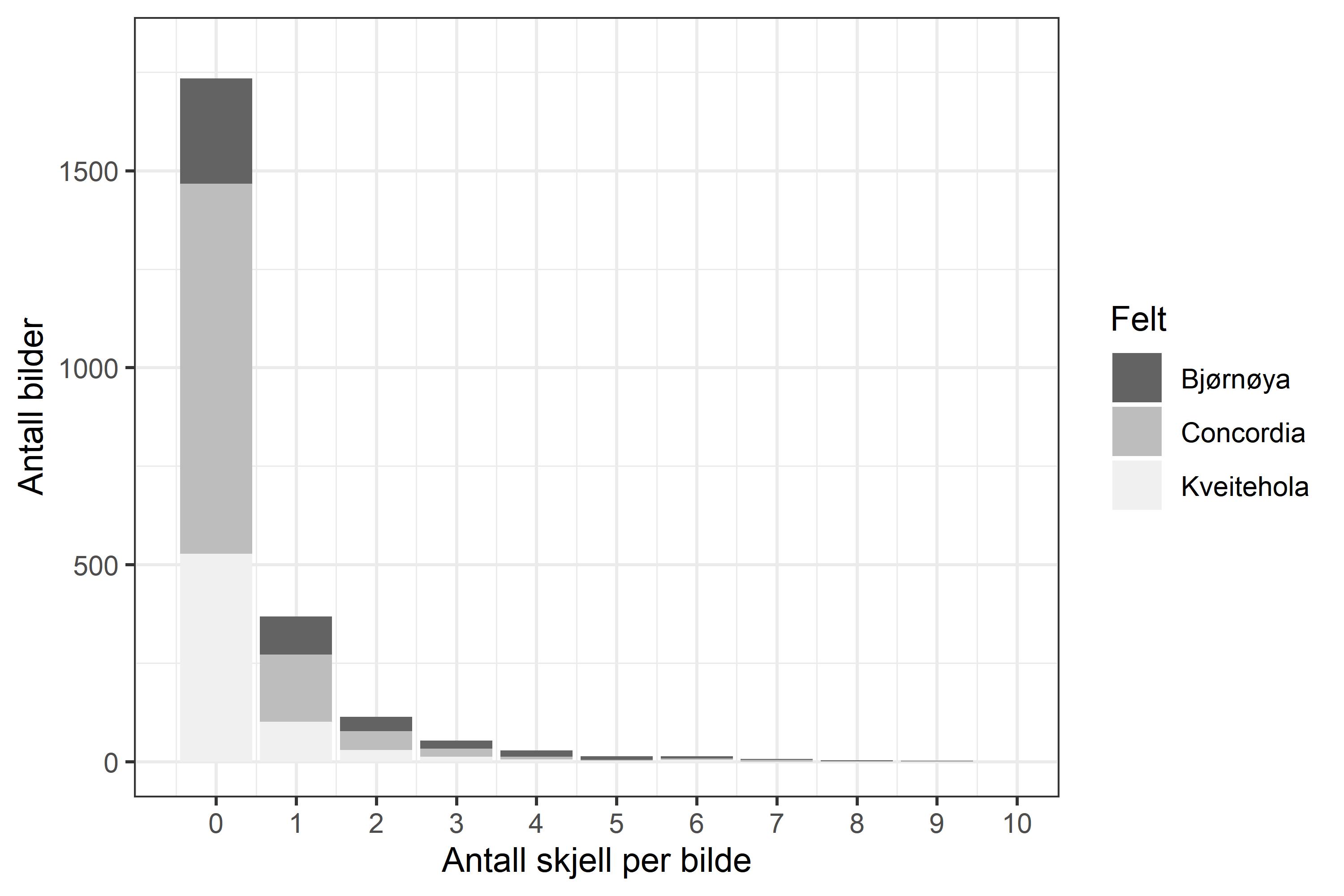
Tellingene fra videostasjonene ble brukt som grunnlag for bestandsestimeringene, og fordelingen av antall skjell per stasjon ble best forklart med en Poisson-fordeling (Figur 10). Resultatene viste høyest gjennomsnittlig antall skjell per bilde på Bjørnøya Sør (0.93), mens Concordia- og Kveitehola-feltene hadde lavere tetthet (0.32 og 0.46). Derimot er Bjørnøya Sør mindre i utstrekning enn de to andre med 191 km2 sammenlignet med Kveitehola som er 569 km2 og Concordia som er størst med 986 km2 (Tabell 1).
Concordia-feltet var feltet med høyest estimert antall skjell (742 millioner) og lavest på Bjørnøya Sør (422 millioner) (Tabell 1). På Kveitehola-feltet var estimatet lavere enn på Concordiafeltet, men høyere enn på Bjørnøya Sør (615 millioner). Våre undersøkelser viste at gjennomsnittlig rundvekt av haneskjell større enn 60 mm skallhøyde (minstemålet) er 60,3 og 70,6 g fra henholdsvis Concordia-feltet og Kveitehola-feltet. Det ble ikke gjort målinger av total individvekt av skjell fra Bjørnøya Sør. Ut fra størrelsessammensetningen er det rimelig å anta at individvekten hos skjell fra Bjørnøya sør er minst like stor som hos skjell fra Kveitehola. Basert på disse individvektene ble totalbiomassen beregnet til henholdsvis 29,5, 43 og 51,9 tusen tonn for feltene Bjørnøya Sør, Kveitehola og Concordia (Tabell 1). Dette betyr at den estimerte tettheten er høyest på feltet Bjørnøya Sør med 2,21 skjell/m2, mens tettheten på Kveitehola og Concordia var henholdsvis 1,08 og 0,75 skjell/m2. Arealet av feltene gir størst utslag på estimatene av den totale biomassen på det enkelte felt. Variasjonen i estimatene er størst for Kveitehola og Concordia. Noe som skyldes at det er flere stasjoner med store variasjoner mellom stasjonene her.
Felt
Antall (106 skjell)
Biomasse (1000 tonn)
Arealstørrelse (km2)
Tetthet (skjell/m2)
Estimat
95% KI
Estimat
95% KI
Bjørnøya Sør
421.88
±193.75
29.53
±13.56
191.18
2.21
Concordia
742.72
±571.86
51.99
±40.03
986.24
0.75
Kveitehola
614.67
±375.10
43.02
±26.26
568.54
1.08
Tabell 1 . Estimert antall skjell (i millioner), biomasse (i 1000 tonn) og areal (kvadratkilometer) av de tre feltene. Biomassen er beregnet med en gjennomsnittlig individvekt på henholdsvis X, Y og Z for de tre feltene. Konfidensintervaller (KI) indikerer usikkerheten i estimatene.
Figur 9. Kart som viser de tre feltene og alle stasjoner fra toktet i 2019 (blå=videostasjon, rød=skrapestasjon). Punktene er skalert i forhold til antall registrerte skjell (prosent av maksimum registrerte i alle skrape- og videostasjoner). Grå linjer indikerer 50,100 og 150 meter dybdekoter og det rosa skraverte området angir verneområdet rundt Bjørnøya.
Figur 10. Frekvensfordeling av antall skjell per bilde fra alle stasjoner og felter. Gråtoner indikerer de tre forskjellige feltene.
5.5 - Usikkerhet i estimatene
Konfidensintervallene i Tabell 1 understreker at det er betydelig usikkerhet rundt bestandsestimatene, men at variasjonen mellom stasjoner og derfor usikkerheten ligger i normalområdet for et forskningstokt og sammenlignbart med hva finnes i andre bestander. Konfidensintervallet viser imidlertid bare usikkerheten knyttet til variasjonen i tellingene, mellom stasjonene. I tillegg finnes det også usikkerheter som ikke er kvantifisert; definisjonen av strata, valg av beregningsmodell og usikkerhet knyttet til selve prøvetakingen.
De fleste metoder for bestandsestimering bygger på at observert antall eller biomasse i det undersøkte området er representativt for hele utbredelsesområdet. Dermed blir bestandens størrelse et produkt av tetthet/vekt per undersøkt arealenhet («swept area»), multiplisert med totale arealet av et felt. Dette betyr at det er en lineær sammenheng mellom bestandsestimater og arealstørrelse. Antagelser omkring utbredelsesområdet bidrar derfor betydelig til usikkerheten i de absolutte bestandsestimatene. Det gjelder spesielt i dette tilfellet fordi haneskjellet har spesifikke habitatkrav. De forskjellige habitatene er ujevnt fordelt og kan bidra til lave tettheter av skjell over store områder. Vi har forsøkt å redusere dette problemet ved å foreta en post-stratifisering av feltene som kun baserer seg på områder og bunndybder hvor skjell ble observert. Dette øker sannsynligheten for at observasjonene på de enkelte stasjonene vil være representative for hele skjellfeltet.
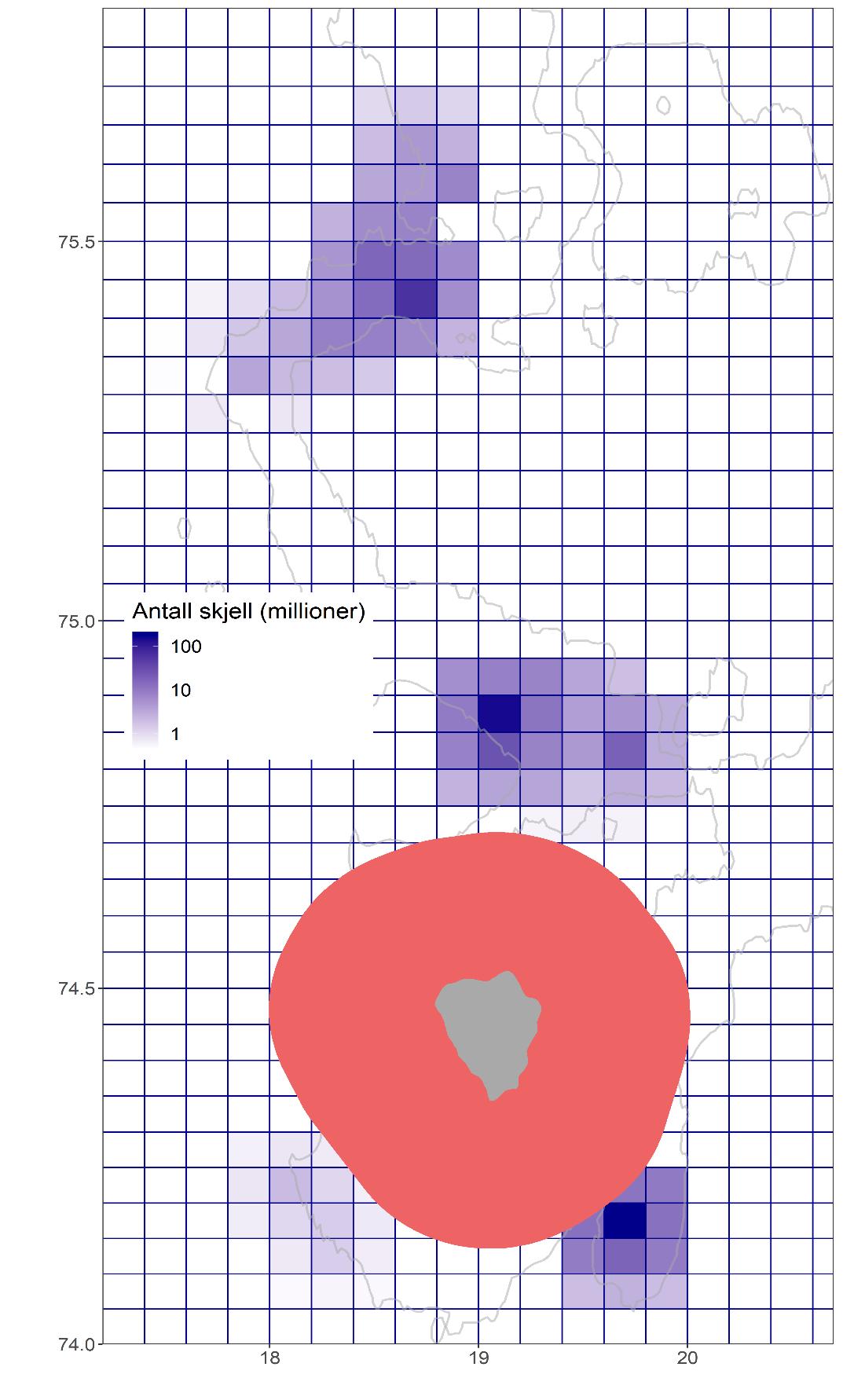
Det er likevel mulig at vi kan ha inkludert arealer med lave tettheter eller ingen skjell, eller utelukket områder med høye tettheter, slik at resultatene enten kan være over- eller underestimater. Gridsystemet som ble testet (Figur 11) viste at totalt antall skjell estimert i grid-systemet ikke skiller seg vesentlig fra estimatene i stratasystemet (Tabell 2). Størst avvik mellom de to tilnærmingene finner en på Bjørnøya Sør hvor estimatet basert på grid-systemet sannsynligvis overestimerer utbredelsesområdet, noe som bidrar til et for høyt totalantall. På Concordia-feltet og på Kveitehola-feltet er estimatene svært like ved de to tilnærmingene. Uansett er forskjellene små hvis en tar i betraktning den store usikkerheten knyttet til alle estimatene (Tabell 1). Selv om definisjonen av arealstørrelse fortsatt er en hovedfaktor i beregningen av de absolutte bestandsestimatene, bekrefter sammenligningen av de to tilnærmingene at stratasystemet sannsynligvis er den beste tilnærmingen for å estimere utstrekningen av feltene.
Valg av estimeringsmetode kan påvirke estimatet og dermed usikkerheten i stor grad. For å evaluere mulige effekter har vi testet en alternativ metode hvor en forutsetter at alle bildene fra hvert felt er representative for tetthetsfordelingen i hele feltet, uavhengig av stasjon. Hvis en ser på fordelingen av antall skjell per bilde på ett skjellfelt er sannsynligheten for å finne et gitt antall skjell per bilde, best forklart av en negativ binomialfordeling. Årsaken til dette er at en større variasjon i skjellfordelingen på hele feltet er større enn skjellfordelingen per stasjon, og at en negativ binomialfordeling er mindre følsom for store variasjoner i data enn Poisson-fordelingen. Den alternative metoden gir estimater som er noe lavere på Bjørnøya Sør og litt høyere på Concordia og Kveitehola, men er av samme størrelsesorden som ved bruk av modell 1 (Tabell 2). Dette indikerer at enkeltbilder er tilstrekkelig som tilfeldige og representative prøver fra feltet, og understreker at bestandsestimeringen er ufølsom i forhold til valg av metode. Uansett viser den statistiske analysen at antall skjell per bilde først og fremst henger sammen med stasjonene og i mindre grad feltene. Derfor valgte vi metoden med Poisson-gjennomsnitt per stasjon som hovedmetode.
Felt
Antall (106 skjell)
Metode
(1) Stratasystem
(2) Gridsystem
(3) Alternativmetodikk
Bjørnøya
422
525
388
Concordia
743
634
822
Kveitehola
615
627
664
Tabell 2 . Estimert antall skjell (middelverdi i millioner) ved tre forskjellige tilnærminger: 1. stratasystem og modell 1 (korresponderer til estimatene i tabell 1), 2. gridsystem og modell 1, 3. stratasystem og modell 2.
Figur 11. Estimert tetthet av haneskjell i et gridsystem. Gridsystemet består av identiske kvadrater av 0.2 lengdegrader og 0.05 breddegrader. Antallet i kvadrater uten stasjoner ble ekstrapolert fra de omliggende kvadrater med observasjoner. Grå linjer indikerer 50 og 100 meter dybdekoter og det rosa skraverte området viser verneområdet rundt Bjørnøya.
5.6 - Fiskerisimulering og referanseverdier
Gjennomsnittet av alle simuleringer indikerer en FMSY= 0.25 og en FPGY= 0.19 pr år (tabell 3). De potensielle landingene med en fiskeriintensitet basert på FPGYer omtrent 8% lavere enn ved FMSY, mens den tilsvarende stående bestanden i gjennomsnittet er 18% høyere. Begge strategier har en lav risiko for å komme under Blim .
Resultatene er delvis avhengig av de underliggende antagelsene i modellen, og noen parametre har en veldig stor innvirkning. Referanseverdiene i tabell 3 er derfor svært avhengig av de spesifikke antagelsene om rekruttering og naturlig dødelighet (figur 12). Kunnskapen om begge er begrenset. Variasjon i rekrutteringen baserer seg på alderssammensetningen, og forholdet mellom bestandsstørrelse og rekruttering er ukjent. Naturlig dødelighet (M) har et empirisk grunnlag fra andre haneskjellbestander, men variasjonen er stor. Alle antagelser om rekruttering og naturlig dødelighet er derfor svært usikre. Det gjør at resultatene fra modellsimuleringene også er usikre. Resultatene er også avhengig av antagelsen om at fiskeselektivitet ekskluderer alle skjell som er yngre enn 7 år og at umodne skjell i bestanden ikke påvirkes av fisket.
Referanseverdi
0.25
±0.01
0.19
±0.01
16157
±143
14928
±137
66
±1
78
±1
B<Blim (%)
2.0
±0.3
0.2
±0.1
Tabell 3: Gjennomsnittlige estimater med 95%-konfidensintervaller for alle feltene samlet, for fiskeridødelighet , landinger, bestandsstørrelse i forhold til opprinnelig bestand , samt risiko for at bestanden faller under Blim. Landingene representerer hele skjell (rundvekten), som er antatt å ligge på 70 g i gjennomsnitt for alle feltene (16157 t tilsvarer dermed 228 millioner skjell).
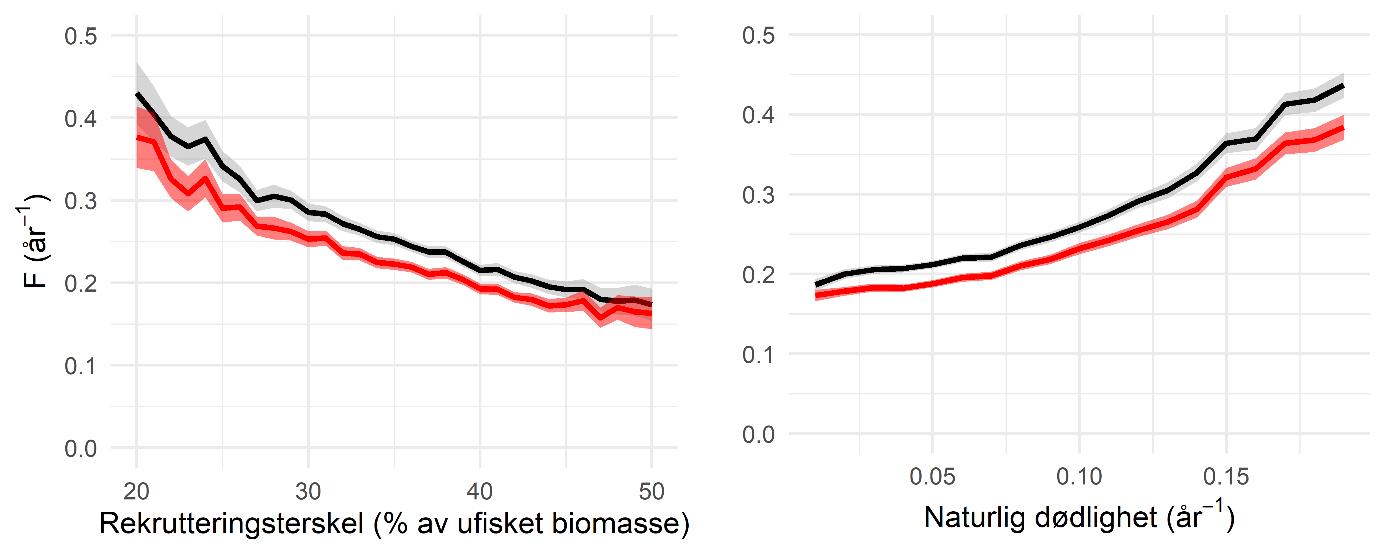
Figur 12. Forholdet mellom rekrutteringsterskel (venstre side) eller naturlig dødelighet (høyre side) med FMSY (svart linje) og FPGY (rød linje).
6 - Diskusjon og konklusjoner
Utbredelsen av haneskjell på de undersøkte feltene ser ut til å være ganske lik det som ble observert ved undersøkelsene på 1980-tallet, men mye tyder på at mengden skjell på feltet sørøst av Bjørnøya er større enn det var på den tiden. Posisjoneringsverktøyet (GPS) på 1980-tallet var imidlertid betydelig mer upresist enn det utstyret vi har tilgjengelig i dag, noe som gjør at sammenligningen mellom nå og da kan være vanskelig. Dessuten ble undersøkelsene på 1980-tallet foretatt gjennom en stratifisert prøvetaking hvor en gjorde skrapetrekk i et gridmønster med faste avstander mellom stasjonene. Dette bidro til at en lettere kunne avgrense feltene.
Innenfor de definerte feltene er haneskjellene veldig flekkvis fordelt i små områder med høye tettheter og mellomliggende deler hvor tettheten er betydelig lavere. Denne flekkvise foredlingen av skjell gjør det vanskelig å sammenligne med tidligere undersøkelser når en heller ikke har nøyaktig posisjon på de gamle stasjonene.
Områder med høye tettheter innenfor de definerte feltene ble registrert både i skrapetrekkene og på videoopptakene. Resultatene fra videosleden bekreftet registreringer med høye tettheter som ble gjort ved bruk av trekantskrapen og vise versa.
Resultatene avslørte ingen klare dybdegradienter i skjelltetthet innenfor det undersøkte dybdeintervallet (60 – 125 m), men stasjoner med høye tettheter (> 50 % av maksimum observert) lå mellom 70 og 110 m dyp. Denne dybdefordelingen samsvarer også med de observasjonene som ble gjort på 1980-tallet. Dette indikerer også at eventuelle endringer i temperatur eller isforhold i disse områdene ikke har påvirket haneskjellets preferansedyp i særlig grad.
Alle prøvene som ble målt viser at store skjell dominerer (skallhøyde > 65 mm) i skrapetrekkene og ut fra størrelsesfordelingen kan det se ut som at skjellene fra Bjørnøya sør og Kveitehola blir noe større enn skjellene fra Concordia-feltet. Dette gjenspeiles i vekstfunksjonene som beskriver sammenhengen mellom skallhøyde og alder, ved at alder ved en gitt størrelse er lavere på Bjørnøya sør og Kveitehola enn på Concordiafeltet. Forskjellene i veksthastighet og oppnådd størrelse kan skyldes at temperaturen gjennom året ved Bjørnøya sør og Kveitehola sannsynligvis er noe høyere enn på Concordiafeltet. Områdene i nordkanten av Spitsbergenbanken (Concordiafeltet) er i stor grad påvirket av polart vann som strømmer sørvestover, mens områdene rundt Bjørnøya og ved Kveitehols i større grad er påvirket av Atlantisk og varmere vann.
Den gjennomsnittlige tettheten av skjell var størst på Bjørnøya sør-feltet, men dette er et lite felt i utstrekning (191 km2) sammenlignet med de to andre feltene. Størst total biomasse var det på Concordiafeltet selv om den laveste tettheten ble observert her. Siden total biomasse er produktet av observert tetthet fra stasjonene, og arealet på feltet, er det derfor en lineær sammenheng mellom biomasse og areal.
Det er stor usikkerhet i bestandsestimatene for disse haneskjellfeltene. Dette har flere årsaker; blant annet er det stor variasjon mellom enkeltstasjonene når det gjelder observert tetthet. Billedkvaliteten på videoopptakene varierte mye og det var bare på deler av opptakene en kunne se skjellene tydelig. I tillegg kunne det av og til være vanskelig å avgjøre hvorvidt et skjell var levende eller ikke. Den største feilkilden ved bruk av videoopptakene var likevel at en ikke var i stand til å se alle skjellene, noe som bidrar til et underestimat av skjelltettheten.
I beregningene av et bærekraftig langtidsutbytte fra de tre undersøkte skjellfeltene har vi valgt to tilnærminger for fiskeridødelighet F: MSY («Maximum Sustainable Yield) som er mest brukte beregning av langtidsutbytte i fiskerier, og PGY («Pretty Good Yield») som også ivaretar en føre var tilnærming for høsting av ressurser. PGY-tilnærmingen gir ca 8 % lavere langtidsutbytte og reduserer også risikoen for redusert rekruttering i framtida. Vi har imidlertid ingen kunnskap om forholdet mellom bestandsstørrelse og rekruttering når gjelder disse haneskjellbestandene slik at det ikke er mulig å si hvorvidt valget av en PGY-tilnærming har noen betydning for de undersøkte skjellfeltene. Forskjellen i langtidsutbytte mellom de to tilnærmingene er imidlertid liten, spesielt hvis man tar størrelsen på usikkerheten (CI) for utbytteestimatene i betraktning.
Våre beregninger viser at et bærekraftig fiske av haneskjell fra de tre undersøkte feltene vil ligge på ca 15000 tonn rundskjell årlig. Det er som nevnt knyttet stor usikkerhet til våre estimater og en må kunne anta at estimatene av bestand og langtidsutbytte er konservative.
Bestand og fangstutbytte i våre beregninger og estimater gjøres på rundskjell, det vil si skjell med skall og alle bløtdeler, mens det i fiskeriet er utbytte i muskel som er viktig. Tidligere har en benyttet en tommelfingerregel om at lukkemuskelen utgjør ca 10 % av den totale vekta av skjellet, men det er indikasjoner på at denne prosentandelen er for lav. Dette forholdet ble ikke beregnet ved denne undersøkelsen og burde studeres nærmere. Bløtdelsvekta av lukkemuskelen varierer til dels mye gjennom året. Det er derfor rimelig å anta at også muskelutbyttet varierer mye (Sundet and Vahl 1981).
Haneskjellfeltet på Moffenflaket nord for Svalbard var det største skjellfeltet i Fiskevernsonen under fiskeriet på 1980-tallet, og en foreløpig sammenstilling av dataene på feltets utbredelse med de utvidete territorialgrensene på Svalbard (4 til 12 nm), indikerer at betydelige deler av dette skjellfeltet fortsatt ligger utenfor grensene for naturvernområdene, og er tilgjengelig for fiske. Dessverre ble en undersøkelse av dette feltet i juni i år forhindret av is, men dette feltet burde ha prioritet når det gjelder en fornyet bestandskartlegging. Forekomstene her vil kunne øke det årlige fangstpotensialet betydelig.
7 - Takk
Vi ønsker å takke skipper og mannskap på FF Johan Hjort for godt samarbeid og uvurderlig innsats under særdeles vanskelige arbeidsforhold. Vi er også Maria Jenssen stor takk skyldig for omfattende alderslesing av haneskjellmaterialet i vanskelig Corona-tider.
8 - Litteratur
Ekman, S. 1953. Zoogeography of the Sea. 417 s. London: Sidgwick and Jackson.
Gruffydd, Ll. D. 1976. The development of the larvae Chlamys islandica in the plankton and its salinity tolerance in the laboratory (Lamellibranchia, Pectinidae). Astarte, 8: 61 – 67.
Hilborn, R. (2010). "Pretty Good Yield and exploited fishes." Marine Policy 34(1): 193-196.
Johannessen, O.H. 1973. Age distribution in Chlamys islandica (Müller). Astarte,6: 15-20.Jonasson, J.P., Thorarinsdottir, G., Eiriksson, H. Solmundsson, J. and Marteinsdottir, G. 2007. Collapse of the fishery for Iceland scallop (Chlamys islandica) in Breidafjordur, West Iceland. ICES Journal of Marine Science, 64: 298-308.
Fossheim, M., Primicerio, R., Johannesen, E, Ingvaldsen, R.B., Aschan, M.M. and Dolgov, A.V. 2015. Nature Climate Change, vol. 5. July 2015
Naidu, K. (1988). "Estimating mortality rates in the Iceland scallop, Chlamys islandica." Journal of Shellfish Research 7(1): 61-71.
Opstad, D. E. Skjelltrålerbygginga – hva skjedde. Ottar nr. 170 2/1988. 8 s.
Pedersen, S.A. 1988. Kammuslinger, Chlamys islandica, ved Vestgrønland. [Iceland scallop, Chlamys islandica at West Greenland]. Rapport til Grønlands Hjemmestyre fra Grønlands Fiskeriundersøgelser, December 1988, 61s.
Rindorf, A., M. Cardinale, S. Shephard, J. A. A. De Oliveira, E. Hjorleifsson, A. Kempf, A. Luzenczyk, C. Millar, D. C. M. Miller, C. L. Needle, J. Simmonds and M. Vinther (2016). "Fishing for MSY: using “pretty good yield” ranges without impairing recruitment." ICES Journal of Marine Science 74(2): 525-534.
Rubach, S. og J.H. Sundet. 1987. Ressurskartlegging av haneskjell ( Chlamys islandica (O.F. Müller)) ved Jan Mayen og i Svalbardsonen i 1986. Rapport fra Institutt for fiskerifag, Universitetet i Tromsø, Ser. b: Ressursbiologi, 42 s.
Shumway, S. E. (ed.). 1991. Scallops: Biology, Ecology and Aquaculture. Developments in Aquaculture and Fisheries Science, 21. Elsevier, Amsterdam, Oxford, New York, Tokyo. 1095 p.
Skreslet, S. 1973. Spawning in Chlamys islandica (O.F. Müller) in relation to temperature variations caused by vernal meltwater discharge. Astarte, 2(1-2): 1-6.
Skreslet, S. and Brun, E. 1969. On the reproduction of Chlamys islandica (O.F. Müller) and itsrelation to depth and temperature. Astarte, 2 (1-2): 1-6.
Sundet, J. H. and O. Vahl. 1981. Seasonal changes in dry weight and Biochemical composition of the tissues of sexually mature and immature Iceland scallops, Chlamys islandica . J. Mar. Biol. Ass. U.K. vol. 61: 1001 - 1010.
Vahl, O. 1982. Long term variation in recruitment of the Iceland scallop, Chlamys islandica, from Northern Norway. Neth. J. Sea Res., 16: 80-87.
Venvik, T. and O. Vahl. 1979. Muligheter og begrensninger for fangst og produksjon av haneskjell. Institutt for fiskerifag, Universitetet i Tromsø, Ser. E, no. 2/79.
Wiborg, K. F. 1963. Some observations on the Iceland scallop Chlamys islandica (Müller) in Norwegian waters. Fiskdir. Skr. Ser. Havunders. vol. 13(6): 38 - 53.