Havforskningsinstituttet anbefaler at hovedtrekkene i 2023- reguleringen av fisket etter leppefisk videreføres for 2024 – men tilrår at det presiseres at bestemmelsen i §37, 4. ledd i Forskrift om drift av akvakulturanlegg også gjelder utsatt rensefisk. Det betyr at oppdrettere som mottar leppefisk må ha en maskestørrelse på nota som brukes som er liten nok til at den utsatte leppefisken ikke kan rømme. Å minimere sannsynligheten for rømning er viktig av flere grunner. Rømt fisk kan øke risikoen for sykdomsspredning og endringer i genetisk struktur i lokale bestander, spesielt dersom fisken ikke er lokalfanget. I tillegg blir fisken ikke brukt til det formålet den er ment til, og det kan føre til høyere fiskeintensitet for å erstatte rømt fisk. I praksis vil dette tilsi at minstemålet for levert fisk må tilpasses maskestørrelsen i kjøpers not. Det er gjennomført forsøk for grønngylt og bergnebb med representative lengdeutvalg. Metode, resultater og konklusjoner er vedlagt i eget notat [kapittel 2]. Havforskningsinstituttet har følgende anbefalingene om justerte minstestørrelser for bergnebb og grønngylt:
Bergnebb som leveres til oppdrettsanlegg med nøter på 26 mm nominell maskeåpning eller mindre må minst være 12.5 cm for å redusere sannsynligheten for rømning. For nøter med større nominell maskeåpning enn 26 mm anbefales et minstemål på 13.5 cm. Brukes det større maskeåpning enn 30 mm, anbefales det å ikke tillate utsett av bergnebb.
Grønngylt over gjeldende minstemål (12 cm) kan settes ut i nøter med de to minste maskestørrelsene (26 og 30 mm nominell maskeåpning) uten fare for rømming. Informasjon innhentet fra notprodusenter viser at 92 % av nøter som selges til smoltutsett har 26-30 mm maskeåpning. Brukes det større maskeåpning enn 30 mm, anbefales det å ikke tillate utsett av grønngylt.
Grasgylt brukes i liten grad som rensefisk, men vi anbefaler at man for denne arten følger samme regler som for bergnebb. Berggylt har et minstemål på 22 cm, og er ikke vurdert her.
For øvrig bør oppdrettere merke seg at for nøter av polyamid (PA) er maskeåpning på vått notlin oftest noe større enn nominell maskeåpning som er målt på tørt notlin. Beregninger viser at minste fiskestørrelse for å unngå rømming er sensitiv for selv små endringer i maskestørrelsen. Som eksempel vil en økning av maskeåpningen fra 26 til 27 mm endre minste anbefalte fiskelengde for utsett av bergnebb fra 11,3 til 11,7 cm.
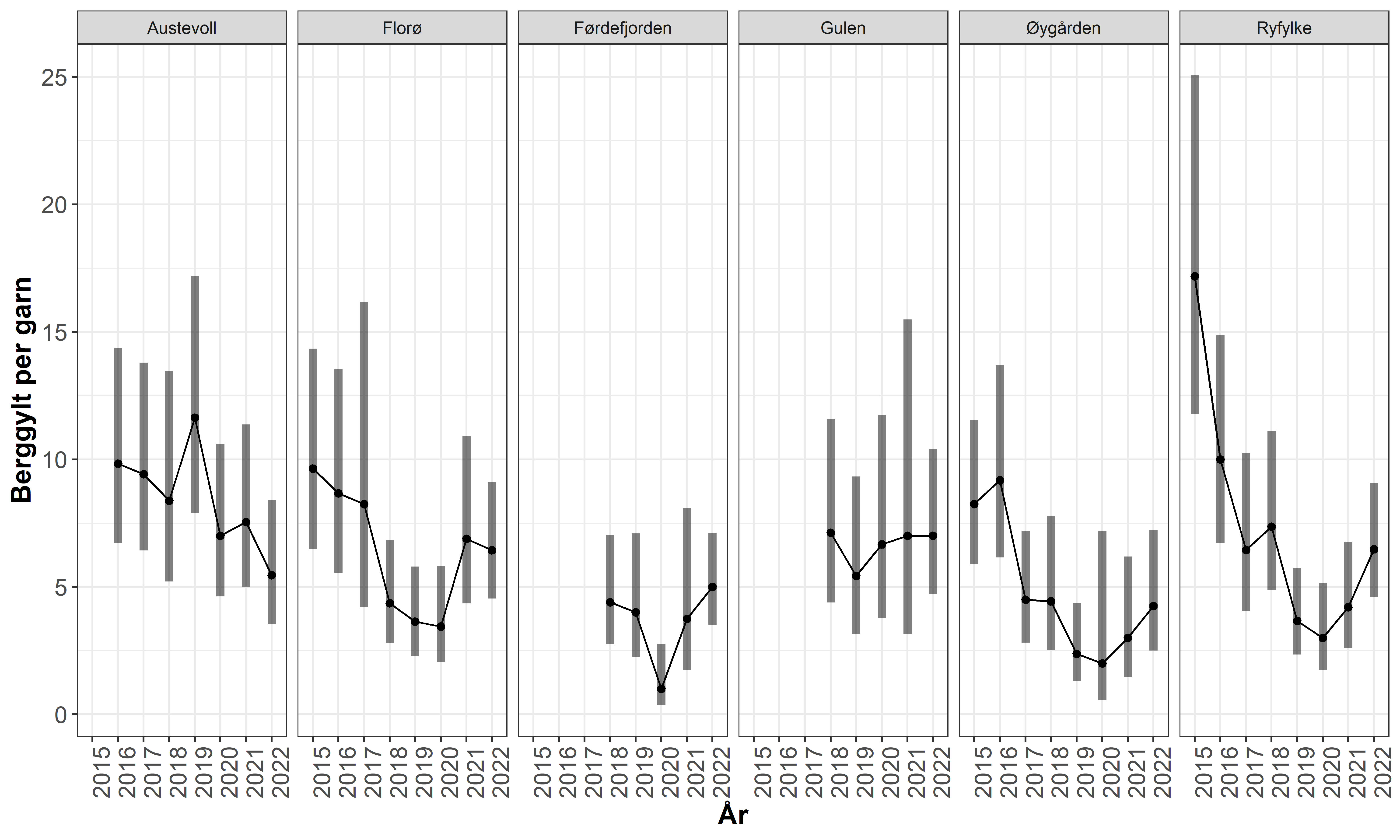
Når det gjelder bestandssituasjonen og fiskeripåvirkning er det ikke kommet frem ny kunnskap som tilsier at det behov for endringer i reguleringene. En oppdatert kunnskapsstøtte som beskriver og diskuterer utviklingen i relevante tidsserier er vedlagt [kapittel 3]. Det er gjennom tidligere og pågående studier dokumentert at fiskeriet kan ha en bestandsregulerende effekt på både bergnebb, grønngylt og berggylt, men det er kun for berggylt man har tydelige indikasjoner på overfiske, og da i områder på Vestlandet. Det er viktig å merke seg at det i forkant av 2022 sesongen ble innført omfattende reguleringer for beskatningen av berggylt i form av økt minstemål og maksimalmål. De siste to årene har det vært en positiv utvikling i fangstrater av Berggylt på Havforskningsinstituttets rusetokt på flere av stasjonene på Vestlandet, men nivåene er fortsatt lavere enn i 2015-2016 (Figur 1.1). Dette kan tyde på at økt minstemål og maksimalmål har hatt ønsket effekt, og det er behov for å opprettholde disse reguleringene for å fortsette gjenoppbyggingen av bestandene.
Figur 1.1. Utvikling i fangst-rate for berggylt på ulike stasjoner i garn tokt på Vestlandet 2015-2022.
Det er viktig å understreke at det stedvis kan forventes lokalt overfiske på en eller flere av de tre målartene i områder med høy fiskeintensitet over tid, noe som i utgangspunktet vil kunne påvirke rekrutteringspotensialet til leppefisk, men også kunne medføre indirekte økologiske konsekvenser. Det viktigste tiltaket for å motvirke dette er minstemål og maksmål tilpasset artenes biologi (størrelse ved kjønnsmodning/kjønnsskifte). Havforskningsinstituttet vurderer at de nåværende reguleringene hensyntar dette, og anbefalingene om å sette minstemål for bergnebb tilpasset notens størrelse vil i praksis bety en økning av minstemålet.
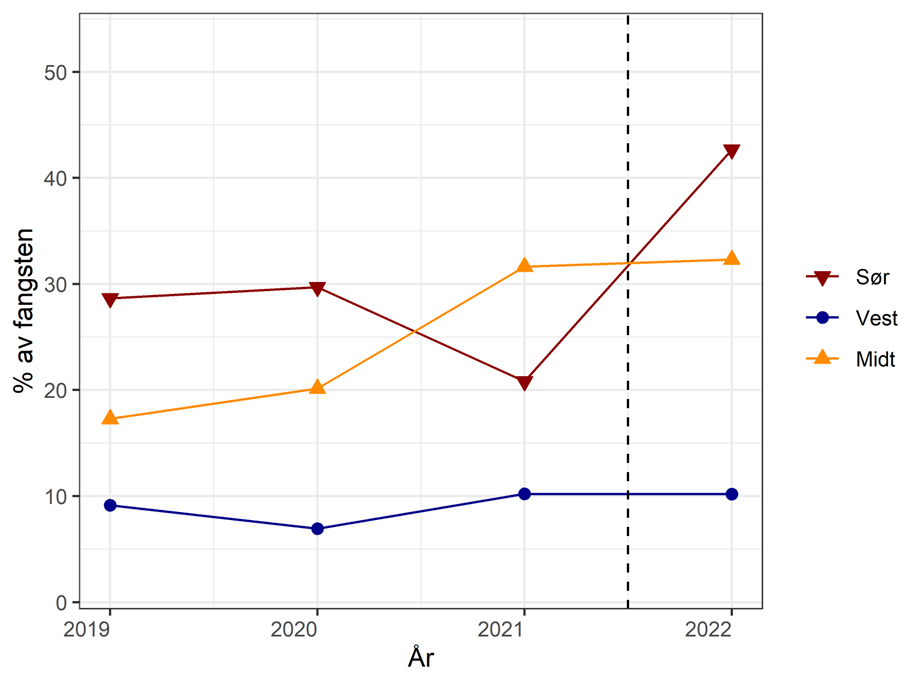
Når det gjelder fangstområde Sør så er fangstene gått betydelig tilbake de siste årene. I 2022 ble under 25 % av kvoten tatt, og det ligger an til å bli fangster på tilsvarende nivå i år. Det kan dermed stilles spørsmål ved om det er behov for like strenge minstemål og maksimalmål for berggylte i dette fangstområdet. Havforskningsinstituttet velger å opprettholde anbefalingen om å ha like reguleringer her, da vi på generelt grunnlag anbefaler bruk av lokalfanget leppefisk for å unngå økt sannsynlighet for smitte eller genetisk utveksling mellom naturlig adskilte bestander. For berggylt og grønngylt er det store genetiske mellom populasjonene på Sør- og Vestlandet, og det geografiske skillet sammenfaller med grensen mellom Region Sør og Vest. Hoveddelen av leppefisk som fanges på Sørlandet transporteres til Vestlandet og Midt-Norge. I tillegg er minstemål og maksimalmål satt for å hindre lokalt overfiske, og der det er mange fiskere på et mindre område kan dette fortsatt være en utfordring. I Region Midt ble litt over 50 % av kvoten tatt i 2022. Havforskningsinstituttet anbefaler at gjeldende reguleringer videreføres også her, da det er mindre kunnskap om bestandene i dette området enn i de to andre fangstområdene.
Det vil i de neste årene komme ny kunnskap om sammenhengen mellom fiskeintensitet og fiskeripåvirkning på målartene og økosystemet for øvrig. I år ble sporingsdata (AIS/VMS) for 2022 tilgjengeliggjort, og disse har blitt benyttet til å utvikle en modell for å estimere fiskeintensitet i og rundt Austevoll kommune. For grønngylt finner her en negativ sammenheng mellom et områdes fiskeintensitet i 2022 og fangstrater i Havforskningsinstituttets rusetokt i 2023. Det er ikke signifikante effekter for de andre artene, men det er viktig å understreke at dette er en modell som er under utvikling og det er for tidlig å trekke endelige konklusjoner før fiskepresset i dette områder har blitt sammenliknet med andre områder og foregående år. Havforskningsinstituttet vil derfor anmode om at også sporingsdata for 2020, 2021 og 2023 blir tilgjengeliggjort så fort som mulig slik at man kan utvikle og validere modellen og undersøke endringer i fiskeintensitet over tid på regionalt og nasjonalt nivå. Det er vedlagt et notat som beskriver modellen, analyse av variasjon i fangstrater som funksjon av fiskeintensitet og betraktninger for veien videre for dette arbeidet [kapittel 4].
Vedlegg:
Kapittel 2. Betraktninger om rømming av leppefisk fra oppdrettsnøter
Ved utsett av leppefisk i oppdrettsnøter, må fiskestørrelsen tilpasses maskeåpningen på nota for å unngå rømming. Havforskningsinstituttet har estimert minste fiskelengde for bergnebb og grønngylte for rømmingssikkert utsett for vanlig brukte maskestørrelser i oppdrettsnøter. Beregningene for bergnebb er gjort ved bruk av to forskjellige metodikker; hhv trengningsforsøk i kar og modellberegninger ved bruk av programpakken FishSelect. For grønngylte er det kun gjort trengningsforsøk.
2.2 - Materiale og metoder
I trengningsforsøkene ble det brukt PA notlin med nominelle maskeåpninger på hhv 26, 30, 40 og 50 mm. Maskeåpningene ble kontrollmålt ved forsøksstart med et skyvelære. Målingene ble gjort med vått notlin. Etter avsluttet forsøk ble det gjort en ny serie målinger av maskeåpning der en sammenlignet målinger gjort med et skyvelære og en Omega-måler. Disse målingene ble gjort for tørt notlin. Alle målinger er gjort i samsvar med ISO 16663-1.
For hvert trengningsforsøk (maskeåpning og art) ble retensjon modellert som en funksjon av fiskelengde (logit-modell). Av beregningsmessige hensyn er L 99 (lengden der 99% av fisk med denne lengden holdes tilbake) benyttet som mål på minste fiskelengde som vil at hindre rømming for den gitte maskeåpningen. Fisken som ble benyttet i trengningsforsøkene ble fanget i nærområdet til Forskningsstasjonen Austevoll
FishSelect (Herrmann et al. 2009, 2012) er en sammensetting av metoder (morfometri- og kompressibilitetsmålinger), verktøy og programvare som er utviklet for å bestemme om en fisk kan trenge gjennom en gitt maske eller annen åpning av definert form. Metoden er benyttet til å studere ulike fiskeredskapers selektivitet.
Bergnebbene som ble benyttet til morfometri- og kompressibilitetsmålingene ble samlet inn av en lokal fisker på Hitra i september 2017. Det ble benyttet 100 fisk som dekket lengdeintervallet fra 78 til 160 mm. FishSelect gjør bereninger for stive, halvslakke og slakke masker. I våre analyser har vi benyttet estimatene for slakke masker, som vi anser som det mest realistiske alternativet for ei oppdrettsnot. Dette alternativet gir også det mest konservative estimatet med hensyn til rømmingsrisiko.
2.3 - Resultater
2.3.1 - Trengningsforsøkene
Maskemålinger: Kontrollmålingene viste at maskeåpningene for vått notlin var 5-10% større enn nominell maskeåpning (Tabell 2.1). Det var godt samsvar mellom målingene gjort med skyvelære og Omega (Tabell 2.2).
Maskeåpning (mm)
Nominell
Målt
26
28,6 ± 0,63
30
31,3 ± 0,54
40
43,1 ± 0,97
Tabell 2.1 . Nominell maskeåpning og målt maskeåpning (med standardavvik for vått notlin.
Maskeåpning
Nominell
Skyvelære
Omega
26
24,6 ± 0,52
24,6 ± 0,52
30
27,5 ± 0,85
27,0 ± 0,82
40
37,9 ± 0,32
37,3 ± 0,82
Tabell 2.2. Maskeåpning (mm) for de tre nominelle maskestørrelsene målt med skyvelære og Omega-måleren. Målingene ble gjort på tørt notlin.
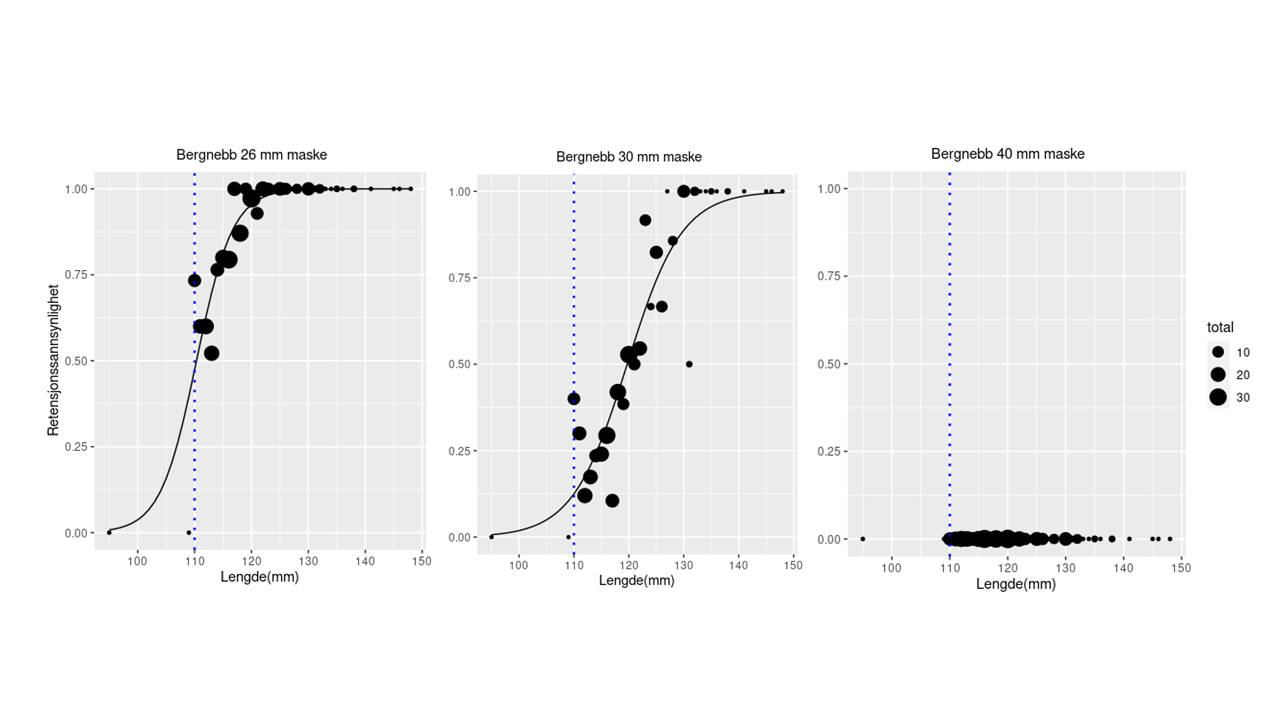
Bergnebb: Til trengningsforsøkene med bergnebb ble det benyttet 296 fisk i lengdeintervallet 95-148 mm. De beregnede retensjonskurvene for de tre maskeåpningene er vist på Fig. 2.1. Den beregnede L 99 -verdien for notlinet med nominell maskeåpning på 26 mm var 12,4 cm, mens den var 13,5 cm for notlinet med nominell maskeåpning på 30 mm. Ved bruk av notlin med nominell maskeåpning på 40 mm og 50 mm var det ingen retensjon (alle fisk rømte).
Figur 2.1. Retensjon for bergnebb for nominell maskepning på 26 (øvre panel, til venstre), 30 (øvre panel, til høyre) og 40 mm (nederste panel). Kurven angir modellert retensjon (logit-modell) basert på de observerte data (fylte sirkler). Størrelsen på symbolene angir antall observasjoner for gitt lengdeintervall.
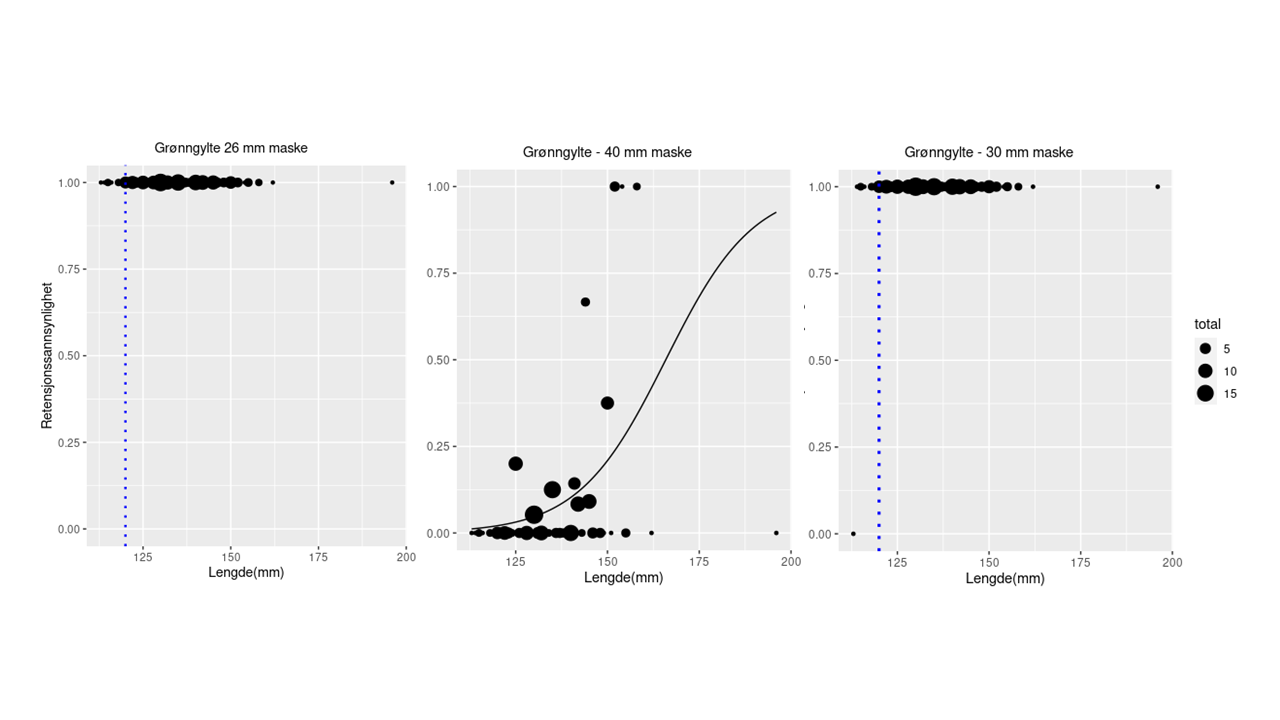
Grønngylte: Til trengningsforsøkene med grønngylte ble det benyttet 392 fisk i lengdeintervallet 11,3-19,6 cm. De beregnede retensjonskurvene for de tre maskeåpningene er vist på Figur 2.2. For 26 mm nominell maskeåpning ble alle fisk holdt tilbake, mens det for 30 mm nominell maskeåpning rømte kun 1 fisk på 11.3 cm. L99 kan ikke beregnes, men er mindre enn minstemålet for grønngylte (12 cm) for begge disse maskeåpningene. For 40 mm nominell maskeåpning ble L 99 beregnet til 22.1 cm. Dette estimatet er imidlertid beheftet med stor usikkerhet fordi det så få data i seleksjonskurvens øvre del. Enkeltobservasjonen med rømming av en fisk på 19,6 cm får derfor stor innvirkning på den beregnede modellen. For 50 mm var det ingen retensjon.
Figur 2.2. Retensjon for grønngylte for nominell maskevidde på 26 (øvre panel, til venstre), 30 (øvre panel, til høyre) og 40 mm (nederste panel). Kurven viser modellert retensjon (logit-modell) basert på de observerte data (fylte sirkler). Størrelsen på symbolene angir antall observasjoner for gitt lengdeintervall.
2.3.2 - FishSelect
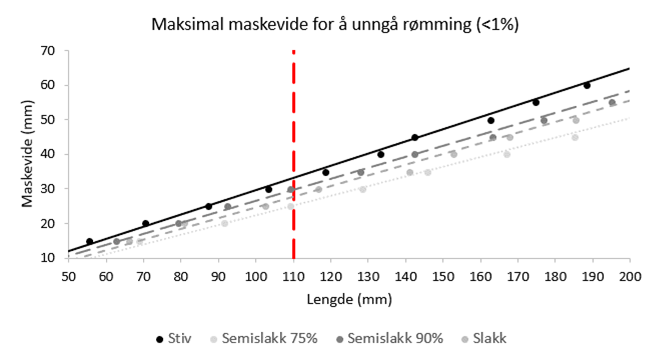
Beregningene fra FishSelect-modelleringen av L99 vs fiskelengde for bergnebb er vist på Figur 2.3. L 99 verdier for de samme tre maskeåpningene som ble benyttet i trengningsforsøkene var hhv 12.7, 13.5 og 17.8 cm.
Figur 2.3. FishSelect beregninger av største maskevidde som kan benyttes ved utsett av bergnebb av gitt minstelengde for å sikre en rømmingssannsynlighet på <= 1%. Beregningene er gjort for stive, halv-slakke og slakke masker. Den røde, stiplede linjen angir minstemålet for bergnebb.
2.4 - Konklusjon
Faktisk maskeåpning er bestemmende for hvilken minstestørrelse det må være på leppefisk for å unngå rømming gjennom maskene i ei oppdrettsnot. Nominell maskeåpning som er gjort på tørt notlin vil ofte være mindre enn maskeåpningen på vått notlin. Våre målinger antyder en forskjell på 5-10%. Oppgitt maskevidde i notsertifikat bør derfor gis som faktiske målinger på vått notlin.
Beregningene våre viser at det ved utsett av bergnebb i oppdrettsnøter med vanlig brukte maskevidder må forventes rømming. Data fra Akvagroup viser at av de solgte nøtene for første smoltutsett har 16% en maskeåpning på 26 mm, 18% har 28 mm og 58% har 30 mm maskeåpning, mens de resterende 8% har andre maskestørrelser (Karen Kvalheim, pers. medd.). Selv med den minste maskevidden (26 mm nominell maskeåpning), vil kun fisk over 12.2-12,4 cm kunne benyttes ved utsett dersom en skal unngå rømming. Benyttes 30 mm maskeåpning økes minstestørrelsen til 13.2-13,5 cm. Dette betyr at en stor andel av de kommersielle bergnebbfangstene over minstemål ikke vil kunne benyttes til utsett. FishSelect simuleringene tilsier en største maskeåpning på 25 mm for å sikre at all bergnebb over minstemål holdes tilbake i nota.
For grønngylte viser imidlertid beregningene at all grønngylte over minstemål kan benyttes i nøter med de to minste maskestørrelsene (26 og 30 mm nominell maskeåpning) uten fare for rømming.
Fisket etter leppefisk er et blandingsfiske hvor fangstene består av ulike arter salgbar leppefisk, samt bifangst av undermåls leppefisk og andre arter. Fisket etter leppefisk er regulert ved redskapsbegrensning, seleksjonsinnretninger i redskap som brukes, minste- og maksmål båtkvoter og fangstperiode og håndtering av bifangst. Det fiskes i hovedsak med spesialbygde teiner, mens et lite mindretall av fiskerne benytter også ruser. Selv om det forekommer variasjoner, gir teiner generelt sett lavere bifangst av andre arter sammenliknet med ruser (Halvorsen mfl., 2017b, 2017a). Ruser er mindre selektive og har et økt innslag av arter som torsk, krabber og ål i forhold til teiner. Fra 2021 vil det være forbudt å bruke ruser i fiske etter leppefisk, men det blir gitt dispensasjon fram til og med 2024 for yrkesfiskere som benytter merkeregistrerte fartøy og har fisket og landet mer enn 3000 stykk leppefisk med ruser i to av de siste tre årene. Fra 2015 måtte både teiner og ruser ha fluktåpninger for å selektere ut den minste fisken. Dessuten ble det påbud om inngangssperre for å hindre oter, stor fisk og hummer å gå inn i fiskeredskapene.
I de fleste områder foregår fisket etter leppefisk i nærheten av oppdrettsanleggene de leveres til. Fisken holdes i en eller flere oppbevaringstanker om bord, og leveres direkte til fiskeoppdrettsanleggene, eller har en kort mellomlagring før levering. Er det større avstander mellom fiskeområder og oppdrett (som er tilfelle fra Sørlandet og Sverige), samles leppefisken opp i tanker eller samlemerder og fraktes videre med tankbiler (VKM 2019).
For oppdretterne er det er viktig at fangstmetodene er skånsomme og at fisken er lytefri ved levering. Hvis fisken står lenge i redskapen kan den få skader, og det er derfor innført regler for ståtid (maks ett døgn utenom helligdager). Det er ikke funnet forskjeller i kvalitet og overlevelse av fisk som er fisket med henholdsvis ruser eller teiner (Skiftesvik mfl., 2014b). All bifangst og leppefisk under minstemål skal etter gjeldene regelverk straks slippes tilbake i sjøen på en slik måte at denne finner tilbake til sitt naturlige miljø og unngår å bli skadet. Hvis fisken sorteres mens fiskerne forflytter seg fra en lokalitet til en annen og blir sluppet ut i åpen sjø, vil den trolig ha redusert overlevelse (Halvorsen mfl., 2021, Cresci mfl., 2022). Mange fiskere har montert rør for å slippe ut fisken under vannflaten. Når den slippes ut igjen på fangststedet (nært land) øker dette muligheten for at den kommer trygt til bunns og i skjul.
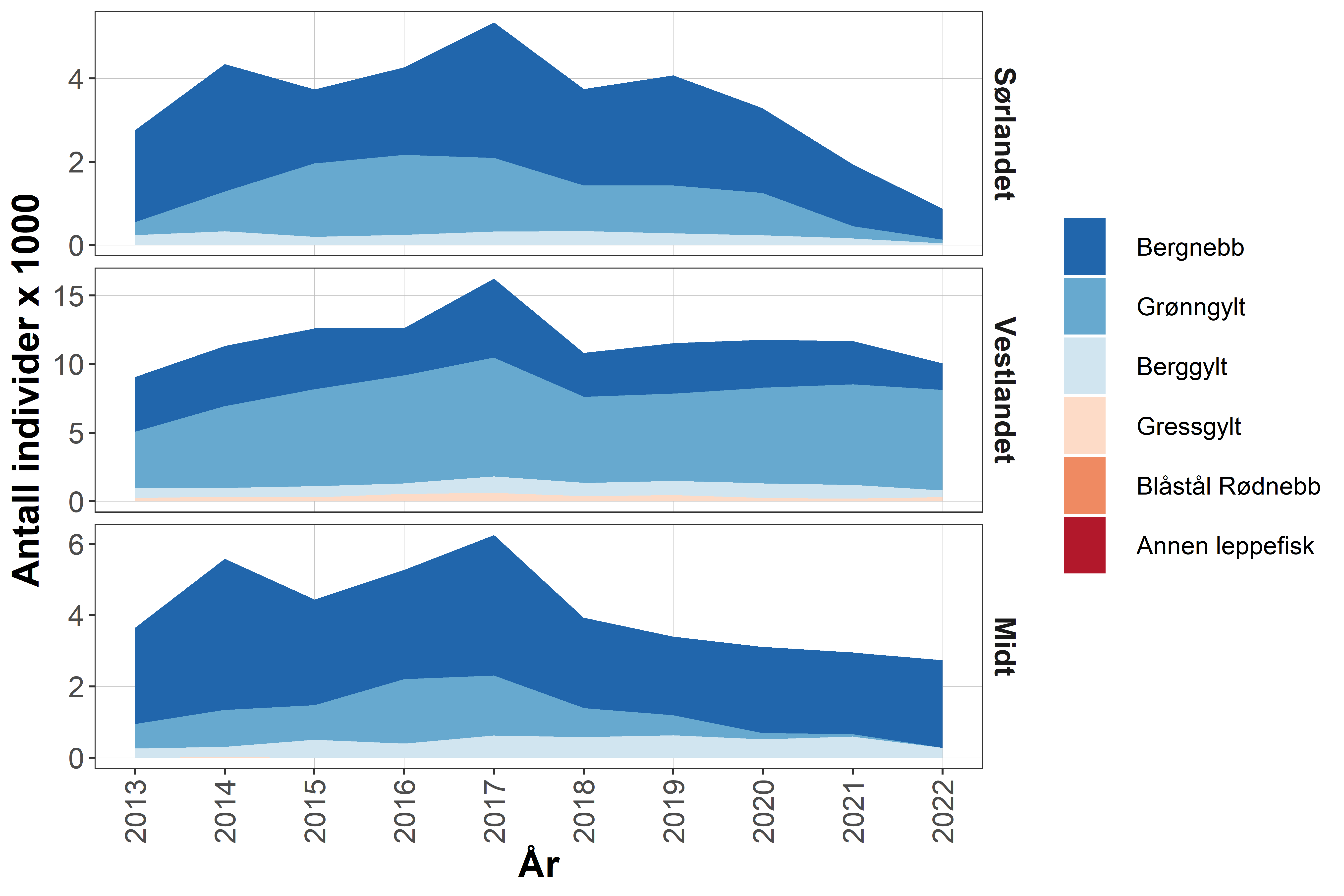
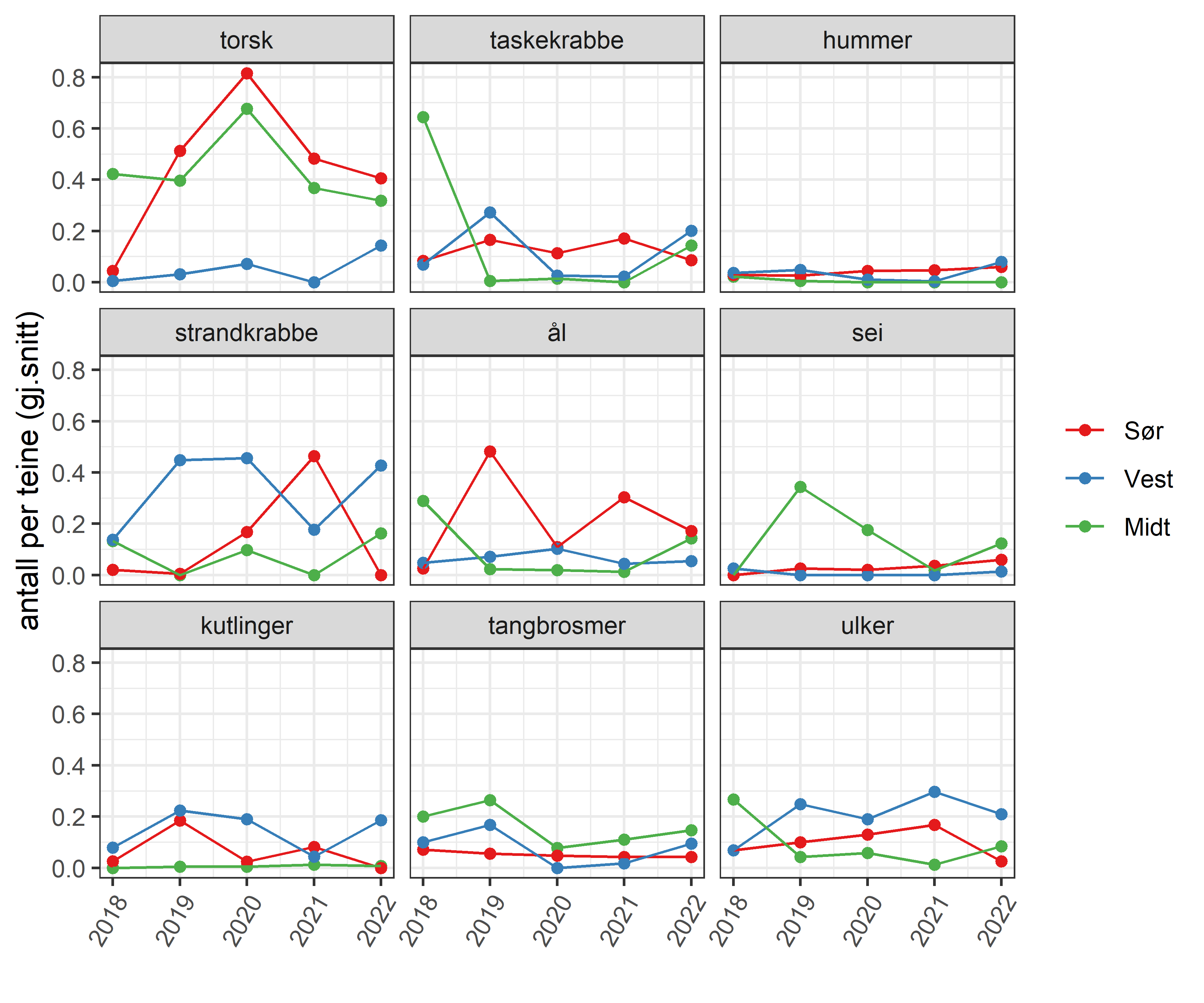
Fangstene av leppefisk har variert med etterspørsel og reguleringsendringer. I 2012 var det registrerte uttaket rett over 11 millioner vill leppefisk på landsbasis, mens 2017 hadde det økt til i underkant av 28 millioner. Dette var nesten 10 millioner mer enn anbefalt uttak. Fisket ble kvoteregulert fra 2018, og uttaket av leppefisk har på landsbasis stabilisert seg og er nå under anbefalt uttak (Tabell 3.1). Totalkvoten ble fordelt på tre fangstområder, 4 millioner til Sør (fra grensa mot Sverige til Varnes fyr på Lista), 10 millioner til Vest (fra og med Varnes fyr på Lista til 62°N) og 4 millioner til Midt (nord for 62°N). Det har inntil i 2022 vært fisket mellom 1-1,7 millioner flere leppefisk enn anbefalt kvote i fangstområde Vest. Kvoten i Vest er 2,5 ganger høyere enn i de andre to regionene, men denne fordelingen reflekter ikke størrelsen på fiskbart areal i regionene (areal i sjø 0-10 m dyp, Tabell 3.1). Dette tilsier at fisketrykket på leppefiskbestandene er betydelig høyere i fangstområde «Vest» i begge de to andre fangstområdene, men det er viktig å understreke at en slik arealberegning ikke har tatt hensyn til eventuelle forskjeller i produktivitet og habitatkvalitet i de ulike områdene. I årene 2020-2021 har det vært levert omtrent like mye av bergnebb og grønngylt. I 2022 var det derimot en overvekt av grønngylt som ble fisket med ca 7,4 millioner individer mot ca 5,1 millioner bergnebb, men det er tydelige forskjeller mellom de ulike fangstområdene (Figur 3.1).
2021
2022
2023
Fangst-område
Kvote (x1000)
Areal 0-10 m (km2)
Kvote/areal (km2)
Uttak (x1000)
Utnyttelse
Uttak (x1000)
Utnyttelse
Uttak (x1000)
Utnyttelse
Sør
4000
515.8
7.8
1937
48 %
871
22 %
633
16 %
Vest
10000
686.0
14.6
11676
117 %
10053
101 %
10140
101 %
Midt
4000
1738.8
2.3
2953
74 %
2735
68 %
2055
51 %
Total
18000
2940.5
6.1
16567
92 %
13659
76 %
12827
71 %
Tabell 3.1 Oversikt over totalt uttak, kvoter og beregnet areal på leppefiskhabitat i de tre fangstområdene. Nord-grensen for fangstområde Midt (Nord for 62 °N ) er satt ved fylkesgrensen mellom Trøndelag og Nordland
Figur 3.1. Utviklingen i rapporterte fangster i fisket etter leppefisk fordelt på art – i perioden 2013 til 2022. Merk en nedadgående andel grønngylt på Sørlandet som skyldes lavere etterspørsel etter grønngylt fra dette området. Data fra Fiskeridirektoratet. https://www.fiskeridir.no/Yrkesfiske/Tall-og-analyse/Fangst-og-kvoter/Fangst/Fangst-av-leppefisk
3.2 - Fiskeripåvirkning på leppefiskbestandene
Fisket etter leppefisk vil påvirke de ulike artene forskjellig. Fangbarheten forventes å være artsspesifikk, noe som i stor grad avhenger av artenes habitat-overlapp med fiskeriet, som for eksempel dybde. Grønngylt og berggylt har høyest forekomst på 0-6 meter, som også er måldybde for dette fiskeriet (Halvorsen mfl., 2020b). Bergnebb, grasgylt og rødnebb/blåstål finnes også like stor eller økende tetthet ned mot 20 meters dyp. Sannsynligheten for at disse artene blir påvirket negativt av fiskeriet derfor lavere . Merkeforsøk viser at grønngylt og bergnebb har høyest fangbarhet i teiner, mens den er tilsynelatende lavere for berggylt (Halvorsen mfl., 2020c, Ruud 2020). De underliggende årsakene for disse forskjellene er ukjente, men antas at de skyldes ulik grad av tiltrekking til agn, samt motivasjon og evne til å ta seg inn og ut av teinene (Ruud 2020).
Det selektive fiskeriet har potensiale til å endre arts-, størrelses - og kjønnsfordeling i leppefisksamfunnene langs kysten (Darwall mfl., 1992, Skiftesvik mfl., 2014a, Halvorsen 2017b), spesielt i fangstområde Vest der fiskeintensiteten er høyest. Leppefiskartene med kortest generasjonstid vil kunne restitueres raskere etter et overfiske enn arter med lengre generasjonstid. I tillegg er de artsspesifikke minstemålene i ulik grad er tilpasset størrelse ved kjønnsmodning. Berggylt stått i en særklasse tidligere, med et minstemål som har vært på 14 cm, mens den kjønnsmodnes som hunn rundt 22 cm, og skifter kjønn til hann ved 34-40 cm (Muncaster mfl., 2013). Med andre ord, et høyt uttak av bestanden over 14 cm kunne ha en sterk påvirkning på kjønnsfordeling og produktiviteten hos berggylt. I reguleringene for 2022 er det tatt hensyn til dette og minstemålet for berggylt er økt til 22 cm, og det er innført et maksimalmål på 28 cm. Generasjonstiden er lengst for berggylt og bergnebb, noe som tilsier at disse artene er mer sårbare for overfiske enn den mer kortlivede grønngylta. Bergnebb kjønnsmodnes når den er 6-7 cm (de er da 2-3 år gamle), godt før oppnådd fangststørrelse, men på denne størrelsen har hunnene lav fekunditet. Grønngylt har kort generasjonstid på Sørlandet (maks alder 4 år), og vil derfor ha potensial til å hente seg inn raskere enn bergnebb og berggylt. På Vestlandet og nordover kjønnsmodnes grønngylt senere og lever lengre (inntil 8 år), noe som gir økt sårbarhet for overfiske. Artenes reproduktive strategi påvirker også sårbarhet for fiske. Hos grønngylt, berggylt og rødnebb/blåstål utøver hannene yngelpleie og er større enn hunnene, og et målrettet, størrelsesselektivt fiske kan derfor antas å redusere andelen egg og larver som overlever (Kindsvater mfl., 2020). Det ble for øvrig ikke registrert fangst av rødnebb/blåstål hverken i 2020 eller i 2021 og registrert fangst i 2022 var minimal.
Det påvist store lokale og regionale variasjoner i den naturlige tettheten av de ulike leppefiskartene (Skiftesvik mfl., 2015, Halvorsen mfl., 2016). Dette skyldes at leppefisk er svært stedbundne og har spesifikke habitatpreferanser, slik at tettheten påvirkes i stor grad av romlig variasjon i miljøbetingelser (Halvorsen mfl., 2020b). En gitt fiskeintensitet vil gi ulik påvirkning i ulike områder, siden det er påvist stor geografisk forskjell i vekst og kjønnsmodning og også i grad av genetisk isolasjon (Halvorsen mfl., 2016, Olsen mfl., 2018, Jansson mfl., 2023a). Dette påvirker igjen bestandenes produktivitet, og dermed deres tåleevne til fiskeri.
Tidligere forskning på effektene av fisket har fokusert på å sammenlikne bestander i fredningsområder med nærliggende områder åpent for fiske. På Skagerrakkysten er det påvist høyere tetthet av bergnebb og grønngylt i fire hummerverneområder (Halvorsen mfl., 2017b), mens det i en annen undersøkelse ikke ble funnet noen tydelig effekt av et null-fiskeområde i Tvedestrand på hverken berggylt, grønngylt og bergnebb i perioden 2010-2019 (Reamon 2020). Bourlat mfl. (2021) observerte heller ingen forskjeller i leppefisksamfunnet mellom fiskede og fredede områder på den Svenske vestkysten. Derimot har en merkestudie gjennomført i Austevoll på Vestlandet vist at fiskeriet tar ut minst 40% av grønngylt over minstemål i løpet av to måneders fiske (Halvorsen mfl., 2017c). Her samlet man inn informasjon om fisketrykk i studieområdet ved daglig overvåking med båt, men det ble ikke sammenliknet med fiskeintensiteten i nærliggende områder. Disse fire studiene kommer altså til ulike konklusjoner om hvorvidt fiskeriet har påvirket de lokale leppefiskbestandene. Resultatene kan heller ikke uten videre generaliseres til større områder når det mangler informasjon om hvordan fiskeintensiteten i og rundt studieområdene - informasjon som det inntil i år ikke har vært mulig å fremskaffe med godt nok presisjonsnivå. Det er rimelig å anta at tetthet av oppdrettsanlegg og/eller nærhet til transportruter og annen infrastruktur har påvirket hvordan leppefisk-fiskeriet fordeler seg langs kysten. I noen områder har det over tid vært en høy konsentrasjon av fiskere, som for eksempel i Austevoll og kommunene rundt. Videre vil sannsynligvis mer variable faktorer som vær og tilstedeværelse av konkurrerende fiskere ha en sterk innvirkning på hvordan fiskerne velger sine fiskeplasser. På Vestlandet rapporterer fiskere at de stadig skifter lokalitet når fangstratene for salgbar fisk faller under et visst nivå (Bollinger 2020). En mobil flåte med uforutsigbart bevegelsesmønster understøttes av posisjonsdata fra teinene til referansefiskere i fangstområde Vest, hvor man har sett at noen fartøy opererer på fiskeplasser som ligger flere 10-talls km unna hverandre i samme sesong. På Sørlandet, hvor det er færre fiskere og det som regel brukes mindre fartøy har ofte fiskerne inndelt ulike områder seg imellom, noe som fører til mindre forflytninger.
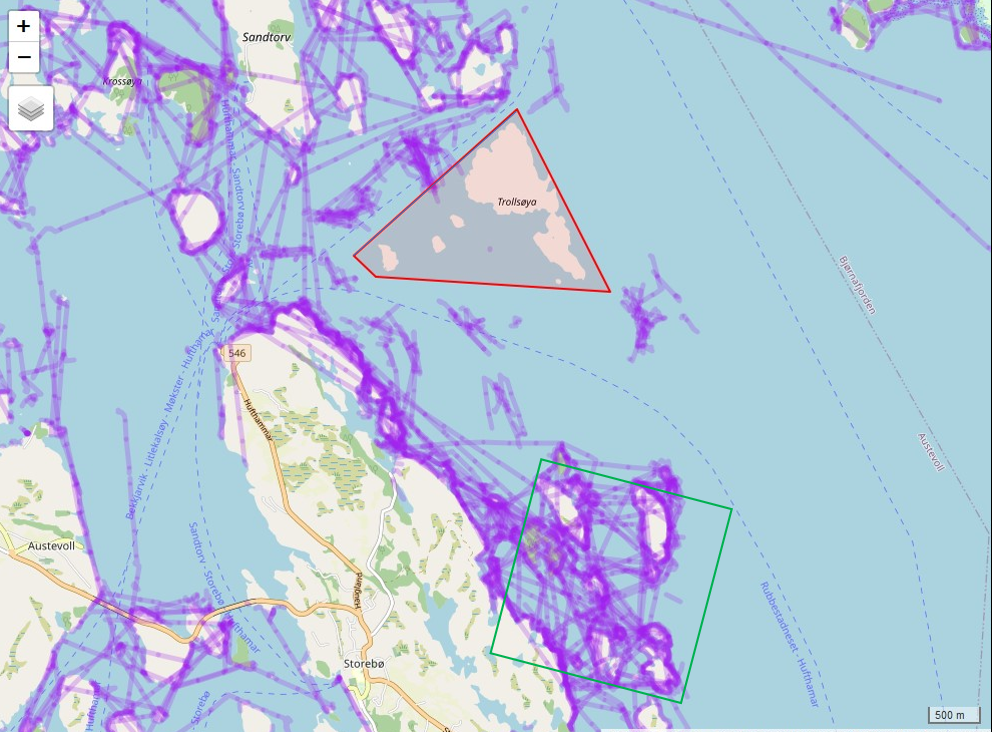
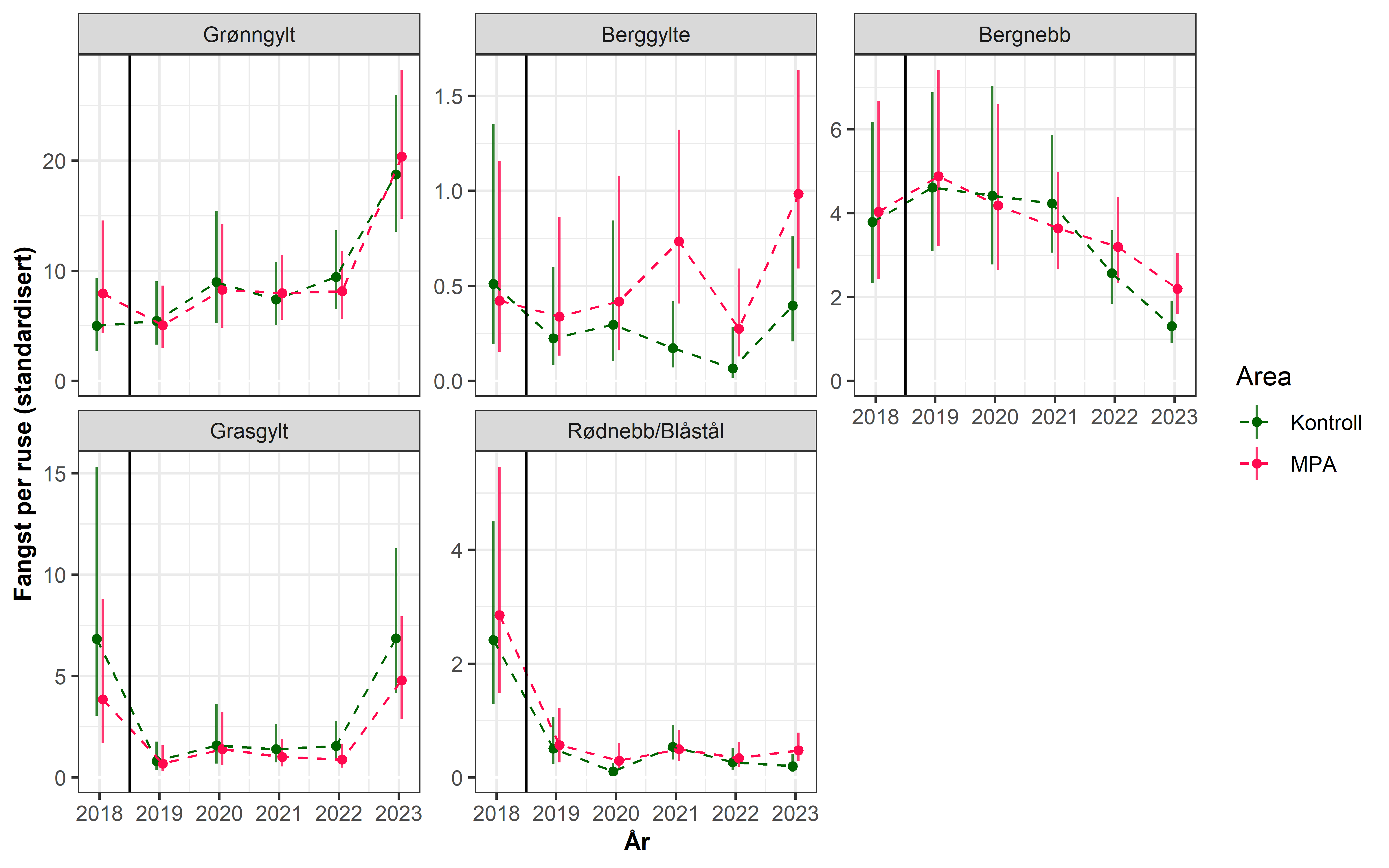
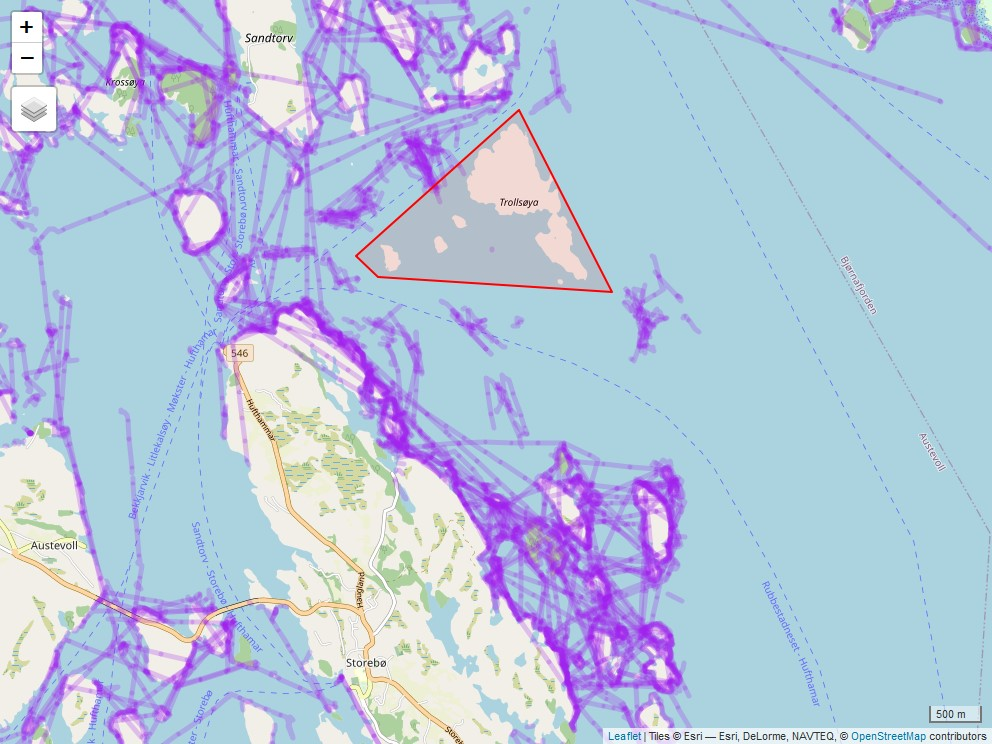
Fra 2020 innførte Fiskeridirektoratet påbud om obligatorisk elektronisk sporing (AIS/VMS) av alle deltagende fartøy (med unntak av ungdomsfiskere). Fiskeridirektoratet/Kystverket har tilgjengeliggjort data fra 2022, og Norsk institutt for Naturforskning (NINA) og HI har samarbeidet med å utvikle en modell som estimerer fiskeintensitet ved bruk av sporingsdataene til fartøyene (se eget notat). Dette er data som er essensielle for å kunne undersøke hvilke effekter fiskeriet har på målartene og økosystemet for øvrig. I første omgang har det blitt laget en modell for Austevoll og områdene rundt. HI har siden 2018 gjennomført et årlig rusetokt for å følge utviklingen i Trollsøya hummerfredningsområde og et nærliggende kontroll-område åpent for fiske siden 2018. Sporingsdataene viser at det ikke skjedde noe fiskeri i selve verneområdet i 2022, mens kontrollområdet var et relativt hardt fisket område (Figur 3.2). Resultatene etter fem år med vern viser at fangstratene for berggylt har doblet seg i verneområdet i forhold til kontrollområdet, mens det er ingen signifikante forskjeller for grønngylt - hvor vi merker oss at fangstene har steget kraftig i begge områder, spesielt mellom 2022 og 2023 (Figur 3.3). For bergnebb er det en generell nedgang de siste 4 årene, og i 2022 var det signifikant mindre bergnebb i det fiskede området. Det er ingen forskjeller mellom områdene for de ikke-kommersielle leppefisk artene grasgylt og rødnebb/blåstål. Fraværet av tydelige effekter på de andre artene, kombinert med store år-til-år svingninger for de mer kortlivede artene, grønngylt, grasgylt og rødnebb/blåstål som sammenfaller i begge områder, tyder dette på at naturlig variasjon i rekruttering og/eller dødelighet har en sterkere populasjonsregulerende effekt enn fiskeriet har på disse artene. Det er likevel for tidlig å konkludere, da det er behov for å bruke sporingsdata på større geografiske skalaer for å sammenlikne det relative fiskeintensiteten mellom ulike områder. Videre så vil tilgjengeliggjøring av sporingsdata fra 2020 og 2021 vil være viktig å inkludere i modellen – da det kan være at den romlige fordelingen av fiskeriet i Austevoll var annerledes i de årene. Det vil også arbeides med å estimere fiskeintensitet der referansefiskerne opererer slik at man kan undersøke fiskerpåvirkning i andre regioner og områder.
Figur 3.2. Fiskespor rundt i hummerfredningsområdet i Austevoll (rød polygon) og i kontrollområdet (grønn firkant). Det har blitt gjennomført standardisert rusetokt i disse områdene 2018-2023 (Figur 3.3).
Figur 3.3. Utvikling i standardisert CPUE (Fangst per ruse av overmåls leppefisk; 95 % konfidensintervall) i Trollsøya hummerfredningsområde (MPA) og nærliggende kontrollområde i Austevoll.
Det er videre gjennomført merkestudier over flere år i et nærliggende verneområde lengre sør på Austevoll. Her har man i et delområde gjennomført et kontrollert fiskeri i regi av HI over tre år. De foreløpige resultatene der tyder på at både grønngylt og bergnebb kompenserer for økt fiskedødelighet med redusert naturlig dødelighet. For grønngylt observerte man i tillegg raskere vekst etter man reduserte tettheten i bestanden. Dette gjenspeiles også i fiskeriuavhengige data fra andre områder hvor man ser at gjennomsnittstørrelsen på grønngylt er lavere der tettheten (fangst-ratene) er høyere (Halvorsen mfl. 2016). Slike tetthetsavhengige responser kan være viktige for at bestandene tilpasser seg beskatningstrykket fra fiskeriet og opprettholder produktiviteten. Resultatene fra merkeforsøket på Austevoll skal publiseres i en fagfelle-vurdert artikkel i løpet av 2024.
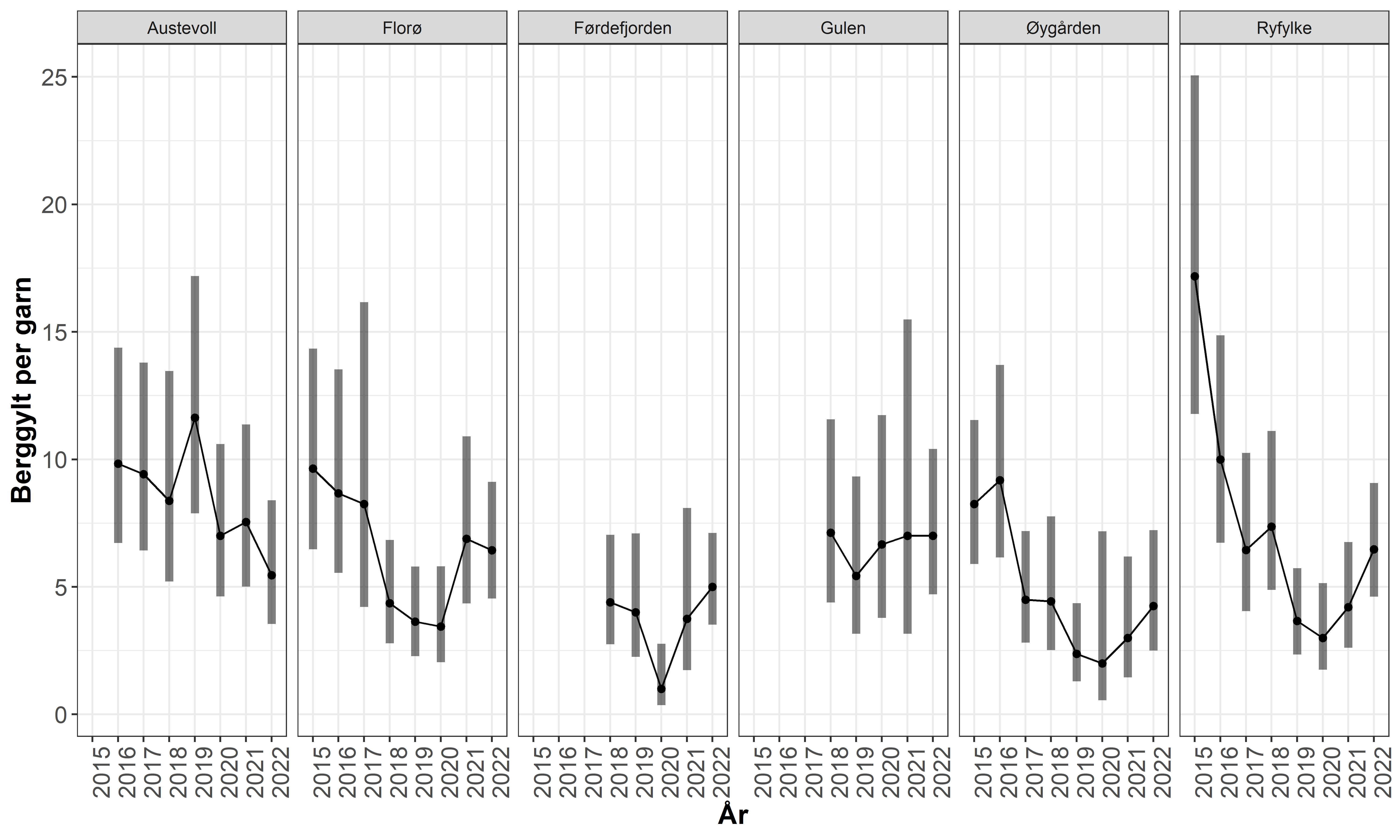
At fiskeriet har redusert berggyltbestandene på Vestlandet understøttes også av data fra garn-tokt og referansefiskere. Siden 2015 har det blitt gjennomført et standardisert ruse-garn tokt på faste lokaliteter på Vestlandet (Ryfylke-Florø). Fisket gjennomføres i perioden september-oktober med småmaskede trollgarn og åleruser. Flere av lokalitetene viser her en tydelig negativ utvikling for stor berggylt. Dette underbygger et helhetsbilde om at berggyltbestandene på Vestlandet stedvis har blitt påvirket av et høyt fiske press . De siste to årene har det likevel vært positiv trend i fem av områdene, mens i Austevoll var fangsten i 2022 den dårligste i tidsserien så langt (Figur 3.3). Denne dataserien vil være viktig for å vurdere effekten av økt minstemål og innføring av maksimalmål for berggylte som innført før 2022 sesongen.
Figur 3.4. Utvikling i fangst-rate for berggylt på ulike stasjoner i garn tokt på Vestlandet 2015-2022. Punktene viser estimert fangstrate ved en GLM modell med 95 % konfidensintervall rundt estimatene.
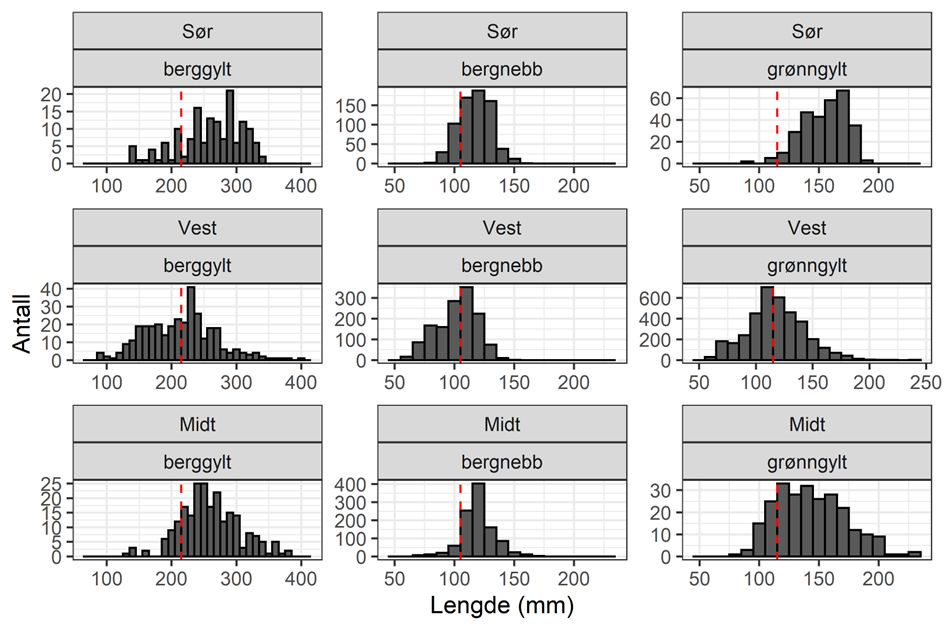
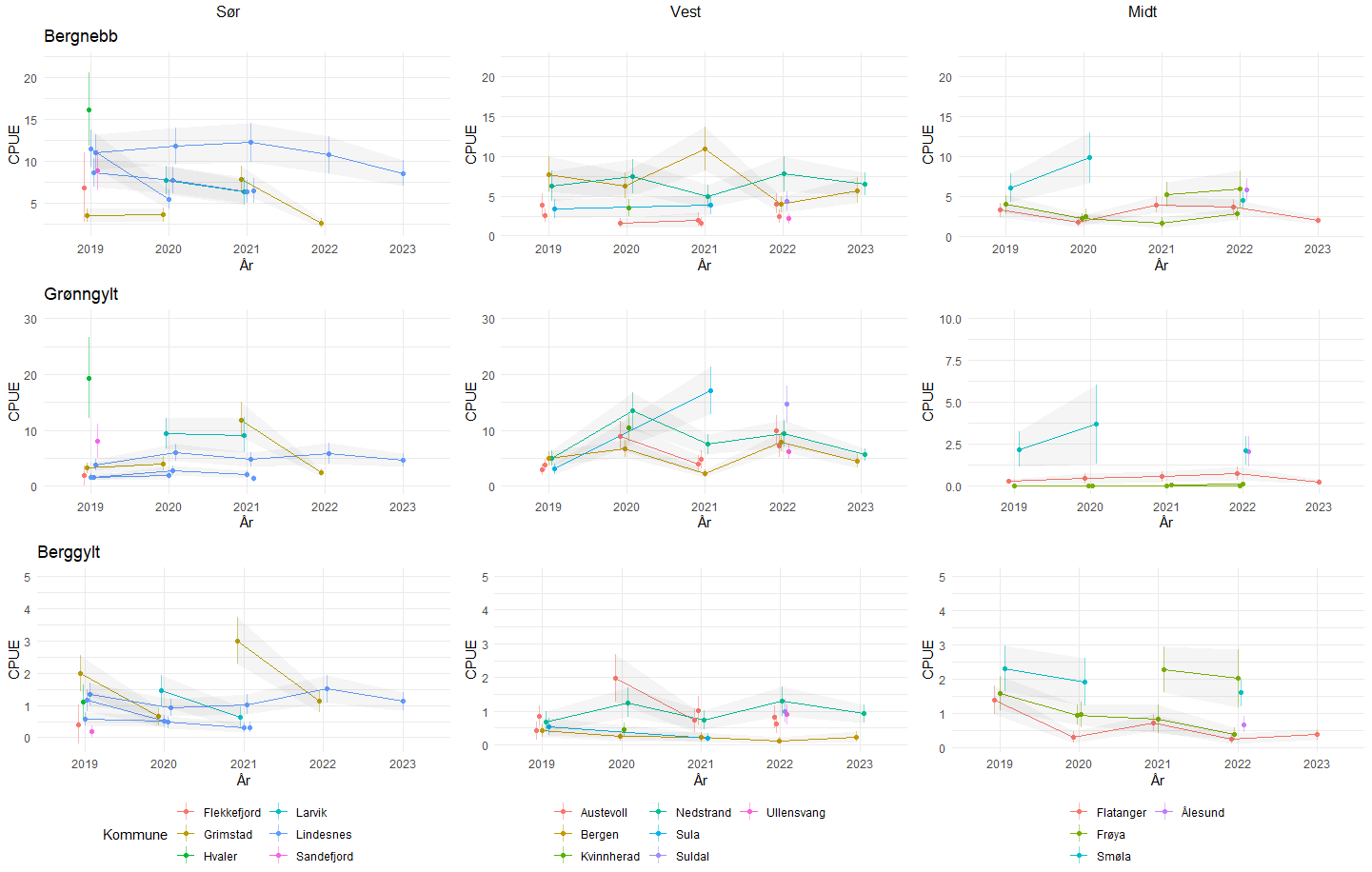
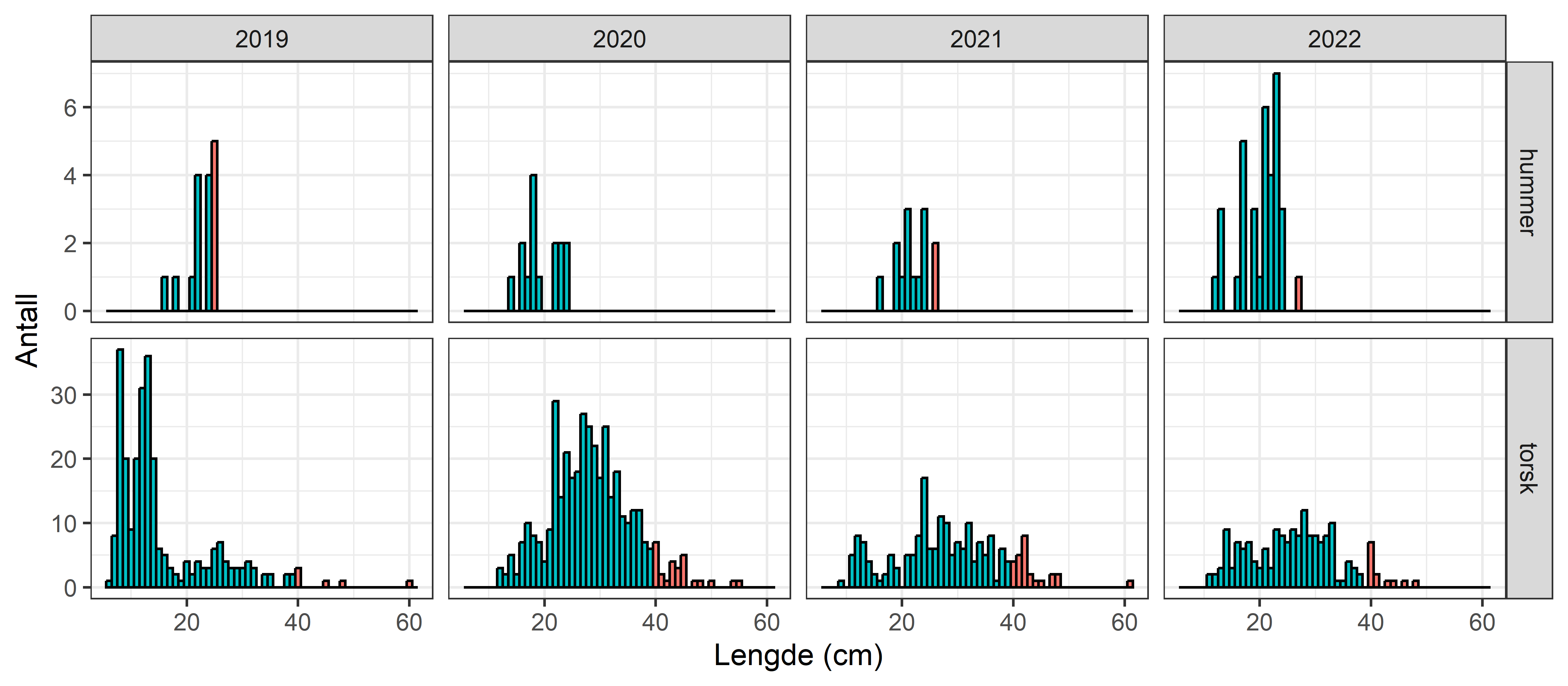
HI har hatt et nettverk av referansefiskere som årlig har levert fangstdata fra 700-800 teinetrekk, hvor all leppefisk blir lengdemålt. Lengdefordelingene fra 2022 viser at det er en tydelig forskyvning mot mindre individer for berggylt i Vest i forhold til de to andre fangstområdene (Figur 3.5). Dette gjenspeiles også i at Vest har en betydelig lavere andel av berggylt som er større eller like store som maksimalmålet (28 cm, Figur 3.6 ). Andelen berggylt som er større eller lik det nylig innførte maksimalmålet (28 cm) er betydelig lavere i Vest enn i de andre fangstområdene, og det er ikke observert noen økning i år etter at maksimalmålet ble innført (Figur 3.6). Fraværet av større individer er indikasjon på høy fiskedødelighet hos berggylt. Også for bergnebb og spesielt grønngylt så er lengdefordelingen mer venstre-forskjøvet (mot minstemålene) i Vest enn i de andre regionene. Det har vist seg å være utfordringer med å rekruttere fiskere og holde på de samme over flere sesonger. Det er stor variasjon i fangstratene mellom fiskere, i noen tilfeller også innen samme kommune (Figur 3.7). Dette reflekterer etter alt å dømme stor lokal variasjon i leppefiskbestandene. Spesielt for fiskere i fangstområde Vest er det observert at fiskerne kan flytte seg store avstander innad og mellom sesong. Fra 2023 ble det besluttet å skalere ned på denne datainnsamlingen, hvor kun fiskerne som hadde levert konsistente data over minst 3 år blir med videre (2 fiskere i fangstområde Vest, 1 i Sør og 1 i midt). Det vil derfor jobbes med å utvikle en fangst-innsats indeks basert på sporingsdata og daglig fangststatistikk, hvor man da unngår sårbarheten som kommer med et begrenset utvalg av fiskere med høy utskiftning av fiskere. Tidsserien fra referansefiskerne har likevel gitt svært viktig kunnskap om hvordan leppefiskbestandene varierer i tid og rom, samt om bifangst.
Figur 3.5. Lengdefordeling for grønngylt, berggylt og bergnebb i fangstene til referansefiskerne 2022. NB – ulike skalaer på x- og y-akser. Rød linje er minstemål.
Figur 3.6. Andel berggylt som er større (eller like store) som maksimalmålet på 28 cm i de tre fangstområdene. Data fra referansefiskere 2019-2022.
Figur 3.7. Utvikling i fangst-rate for referansefiskere i perioden 2019-2023. Punktene viser estimert fangstrate ved en GLMM modell med 95 % konfidensintervall rundt estimatene for hver fisker. Estimatene er standardisert for ståtid, temperatur, fangstdyp og eksponering (estimert middels bølgehøyde), slik at de er sammenliknbare mellom områder. I 2023 deltok 4 fiskere. Fangstkommune for fiskerne er oppgitt og angitt med fargekoder.
3.3 - Genetisk bestandsstruktur og relevans for forvaltning av leppefisk
Leppefisk lever i geografisk atskilte bestander med begrenset grad av utveksling. Leppefisk som flyttes over lange avstander og deretter rømmer, kan blande seg med lokale bestander og på denne måten påvirke den bestandsgenetiske strukturen på mottaksstedet. Leppefiskene er små, og selv små hull i nøtene representerer en rømningsvei. Dette gjelder særlig bergnebb, som er slankere enn de andre artene (Sistiaga mfl., 2021). Grønngylt er rapportert å rømme i mindre grad (Woll mfl., 2013). Når oppdretterne skifter til nøter med større maskevidde, kan dette resultere i at små leppefisk forsvinner ut av noten. All håndtering av laks medfører en fare for rømming eller dødelighet for rensefisken. Det er også rapportert at noen oppdrettere tidligere hadde en praksis hvor leppefisk ble satt fri etter bruk. Studier av bergnebb og grønngylt tyder på at en innblanding av importert fisk i lokale bestander kan ha funnet sted i et importområde i Trøndelag (Jansson mfl., 2017, Faust mfl., 2018). Et studium gjennomført på grønngylt, viser indikasjoner på at det hittil spesielt er de nordligste områdene som er mest påvirket. En teori er at det er enklere for rømt fisk å få rotfeste i områder med nylig etablerte populasjoner, har lav populasjonstetthet og /eller høy fiskeintensitet på lokale populasjoner (Faust mfl., 2021). Nylige publiserte funn viser at bergnebb kan være sårbar for genetisk påvirkning mellom ulike geografisk spredte populasjoner. Voksne individer flytter seg i liten grad mellom områder noe som kan gi både regional og lokal populasjonsstruktur (Jansson mfl., 2023a).
Transport representerer derfor en fare for en genetisk påvirkning av lokale leppefiskbestander. Faktorer som spiller inn, vil variere mellom arter av leppefisk. Arter med lav overlevelse etter utsett, gjør faren for innblanding i ville bestander mindre. Berggylt vil antakeligvis være følsom for genetisk endring som følge av å ha lav populasjons tetthet og høy skjevfordeling mellom kjønn. Det er også fare knyttet til bruk av oppdrettet berggylt, som kommer fra få stamfisk, men blir spredd ut til mange oppdrettslokaliteter. Villfanget berggylt har en høyere lovlig minstestørrelse ved fangst enn de andre leppefiskartene, så sannsynligheten for at denne rømmer er lav. Imidlertid kan oppdrettet berggylt settes ut ved mindre størrelser og disse kan ha større sannsynlighet for å rømme gjennom notmaskene enn villfanget fisk. Oppdrettet berggylt er ofte ikke stedegen. Selv om det er større sannsynlighet at liten berggylt rømmer, vil den lange tiden det tar før de rømte små berggyltene kjønnsmodner (ved 22-23 cm) minske sannsynligheten for innblanding med lokal populasjon. Nylige gjennomførte studier gir indikasjoner på at inndelingen av populasjonsstrukturen til berggylt ligner grønngylt med en nordvestlig og sørøstlig inndeling. Skillet går ved Jæren hvor sandstrendene utgjør en naturlig barriere for genetisk utveksling (Seljestad mfl., 2020). Studiet konkluderer med at transport av berggylt fra Sverige og Sørlandet til Vestlandet og Midt-Norge, vil kunne føre til en miksing av og endring av genetisk forskjellige populasjoner. Et ennå upublisert studium av Jansson et al. støtter dette ytterligere. Sammenligningen av hele genomene av berggylter samlet inn fra Smøla og Flødevigen, viste betydelige genetiske forskjeller og tegn på divergerende lokala tilpasninger.
Arten som sannsynligvis påvirkes minst er bergnebb. Med sine pelagiske egg vil frekvensen av genutveksling naturlig sett være høyere sammenlignet med andre leppefisk. I tillegg har denne arten høyest populasjonstetthet og størst utbredelse. Men bergnebb er også den vanligste arten i transportene, har best overlevelse etter transport og rømmer mest (Woll mfl., 2013). Bergnebb er småvokst og kan lett unnslippe gjennom maskene. Den er sannsynligvis også mer robust. Ved en undersøkelse av bergnebb samlet inn langs en syd-nord-gradient ble det vist en genetisk variasjon som fulgte gradienten relatert til avstand (Jansson mfl., 2017). Dette viser at det meste av genflyten skjer mellom nærliggende områder, selv om havstrømmer kan transportere fritt svevende egg og larver over store avstander. En nylig publisert studie av Jansson mfl. (2023a) viste at mange bergnebbpopulasjoner var genetisk sett noe unike, spesielt i nord. Dette støtter forvaltning på et mer lokalt nivå.
Endringer i den bestandsgenetiske strukturen vil også kunne skje i lokale populasjoner ved overfiske, som følge av tap av genetisk variasjon. Dette er en ofte ignorert, men like vel viktig faktor. Leppefiskene lever i mer eller mindre isolerte, lokale populasjoner. Bruk av leppefisk som er fraktet nordover (eksempelvis fra Sverige til Nordland) kan representere akvakultur med arter som ikke forekommer naturlig i området (grønngylt i nordlige områder) og/eller bruk av ikke-stedegen fisk.
3.4 - Sortering og gjenutsetting av leppefisk og bifangst
Det er en varierende grad av bifangst i fisket, som også varierer med sesong og område. Regelverket er tydelig på at all bifangst (undermåls leppefisk og andre arter) skal gjenutsettes på grunt vann. Siden formålet med fiskeriet er å levere levedyktig leppefisk, er det vurdert til å være et skånsomt fiske på grunn av kort ståtid og fordi fisket foregår på grunt vann (0-8 m). Mange fiskere har montert rør for å slippe ut fisken under vannflaten. Når den slippes ut igjen på fangststedet (nært land) øker dette muligheten for at den kommer trygt til bunns og i skjul. Det er planer om ny forskningsaktivitet for å undersøke i hvilken grad dette gjennomføres i praksis.
En stor del av bifangsten er undermåls leppefisk, som i hovedsak slippes fri (Figur 3.9; Halvorsen mfl., 2016b). Det er også påbud om seleksjonsinnretning (12 mm spalter) for å redusere fangsten av undermåls fisk. Dette har vist seg effektivt i å redusere innslaget av undermåls bergnebb og delvis undermåls grønngylt (Jørgensen mfl., 2017). Størrelsene på åpningen er tilpasset med en målsetning om at undermåls bergnebb skal sorteres ut på fiskedypet. Høyere minstemål og tykkere kroppsfasong hos grønngylt og berggylt betyr at en større andel av undermåls fisk av disse artene ikke kan rømme ut av fluktåpningene. Fluktåpningene er mer effektive i teiner enn i ruser (Halvorsen mfl., 2017a, Jørgensen mfl., 2017).
Leppefisk er svært stedegne arter med små hjemmeområder, og merkeforsøk har vist at de raskt finner tilbake dit om de flyttes og slippes ut opptil 400 m fra fangststedet, gitt at det skjer langs samme kystlinje (Neghabat 2022). Om de derimot må krysse dypere vann (15-20 m) med bløtbunn er det en mye lavere andel som vender tilbake (samme avstand), men skjebnen til disse er ikke kjent. Det kan enten være de da slår seg ned i et nytt område, eller blir spist i forsøket for å ta seg tilbake. Det finnes per i dag heller ikke informasjon om i hvor stor grad sortering og gjenutsetting skjer andre steder en fangstområdet. For å vite om dette medfører økt dødelighet eller har andre negative effekter (sykdomsoverføring, økt konkurranse, forstyrrelse av lokale dominanshierarkier) trengs det mer forskning.
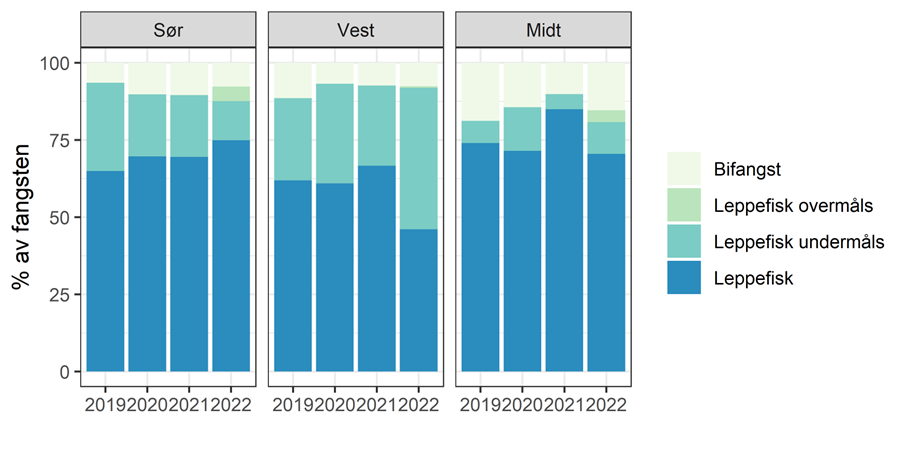
Havforskningsinstituttet har undersøkt artsfordelingen i fangstene som tas i leppefiskteiner med 9x6 cm ovale åpninger. Datagrunnlaget er totalt 2049 teinetrekk fra referansefiskere gjort i 2019-2021. Leppefisk over minstemålet utgjør 60-75% av fangsten (i antall i individer, Figur 3.9). Andelen av leppefisk som settes ut varierer betydelig mellom artene og har vært spesielt lav for berggylt som skyldes at minstemålet beskyttet kun en liten andel av den naturlige størrelsesfordelingen. Fra før 2022-sesongen ble minstemålet for berggylt endret fra 14 cm til 22 cm, og det ble innført et maksmål på 28 cm. Andelen fisk over minstemål som settes ut igjen varierer, og er avhengig av etterspørsel. Fra 2021 har det vært lav etterspørsel etter grønngylt fra Sørlandet, og mesteparten av grønngylten som blir fanget blir nå satt ut igjen. Det er relativt lite bifangst av andre arter (9% i gjennomsnitt i 2021), og andelen er høyest i Midt (nord for 62 grader) og torsk er den mest vanlige bifangstarten i Sør (ca av totalfangsten; Figur 3.10) og Region Midt (~2% av totaltfangsten). Tangbrosme er mest vanlig i Vest (vel 0.5% av totalfangsten).
Fangst av større arter, som torsk, lyr, taskekrabbe og ål kan spise eller skade leppefisk og andre mindre fiskearter i redskapen. Dette er et noe større problem i ruser enn i teiner, da teiner har lavere andel bifangst av større arter. For eksempel ser man at i teiner med bifangst av torsk har færre bergnebb enn de uten torsk, men en medvirkende faktor kan være at torskens tilstedeværelse gjør det mindre attraktivt for mindre leppefisk å gå inn i teinen, eller at de rømmer ut av fluktåpningene. Når det fanges torsk i HI’s forsøksfiske har de ofte en eller flere merkede leppefisk i magen, noe som tyder på at det skjer predasjon i redskapen. Andelen redskap med større bifangstarter i teiner er likevel såpass lav til at vi antar at dødelighet som følge av skade og predasjon er relativt lav i forhold til uttaket av leppefisk til oppdrett, men det er ønskelig med mer forskning på dette. For eksempel kan kameraobservasjoner og data fra merkeforsøk brukes til å kvantifisere predasjon i redskapen. En effektiv måte å redusere denne utfordringen på er å bruke mindre innganger i teiner som reduserer bifangst av torsk og andre større predatorer. Fiskere i områder med mye torsk bruker ofte teiner med mindre innganger av nettopp denne årsaken.
Ser man bort i fra predasjon og skade påført av større arter er vår vurdering at de fleste bifangstarter håndterer fangst og utsett bra dersom de settes ut igjen umiddelbart der de er fisket, slik som krabbe, hummer, ål, torsk og undermåls leppefisk. Vi antar at skadeomfanget på eventuelle sårbare arter vil øke i forhold til hvor mange ganger de fiskes og settes ut igjen.
Figur 3.9. Prosentvis andel av leppefisk, undermåls leppefisk og bifangst i referansefiske 2019-2022.
Figur 3.10. Oversikt over bifangst i referansefiske fordelt på art, oppgitt som gjennomsnitt antall individer per teinetrekk. Data fra 3573 teinetrekk fra referansefiskere i perioden 2018 - 2022.
HI har mottatt henvendelser fra publikum som har uttrykt bekymring for ulovlig uttak av hummer som tas som bifangst. Tallene fra referansefiskerne viser at det fanges relativt lite hummer, totalt 52 hummer på 2049 teinetrekk (Figur 3.10) og at de som tas i all hovedsak er under minstemål (Figur 3.11). Bare 7 var 25 cm eller over av de 52 som ble fanget. Fiskerne rapporterer at disse settes ut igjen. I Havforskningsinstituttets omfattende forskningsfiske i Austevoll og Flødevigen med ruser og teiner fanges det av og til hummer, og per 2021 er ikke registrert skadet eller død hummer i disse studiene. Havforskningsinstituttet har også omfattende fangst-merke-gjenfangst tokt på hummer i Skagerrak, og samlet tilsier disse erfaringene at hummer ikke tar skade av å bli fanget og sluppet ut igjen (Fernandes-Chacon mfl., 2020). Ved fangst av flere hummere i samme hummerteine er det rapportert om klo-tap og død hummer, mest sannsynlig som følge av aggressive interaksjoner i redskapen (Sørdalen mfl., 2020). Det er betydelig lavere fangstrater av hummer i leppefiskredskap sammenliknet med hummerteiner, og det er svært sjelden blitt fanget mer en hummer per teine/ruse i Havforskningsinstituttets prøvefiske etter leppefisk.
Figur 3.11. Lengdefordeling av bifangstartene hummer og torsk i referansefiske. Rød farge: over minstemål.
Zimmermann mfl. (2020) beregnet den totale bifangsten av taskekrabbe i leppefisk-fisket til gjennomsnittlig 235 krabber per fartøy per år for fartøy som fisker med ruser, mens for fartøy som fisker med teiner er gjennomsnittet 2911 krabber per år. Utkastraten av taskekrabbe er høy og ligger i gjennomsnitt på 72% over alle år og redskapstyper (varierer mellom 44 og 100%), men vi kan anta at de fleste krabber som gjennutsettes overlever. Vi antar derfor at gjennomsnittlig mer enn en fjerdedel av krabbene som tas som bifangst, landes og brukes til agn. Ekstrapolert til hele leppefiskkvoten kan dette bety at omtrent 150 000 krabber (rundt 60 tonn) brukes til agn per år, men en stor usikkerhet er knyttet til dette estimatet.
I 2015 ble det påbudt med kryss eller not i ytterste kalv på ruser og maksstørrelse på inngangskalv i teiner brukt til fiske etter leppefisk. Dette har redusert bifangsten av oter, hummer, krabbe og stor fisk. Under hele referanseperioden 2015-2018, dvs. etter at inngangssperre ble innført, er det kun rapportert 5 skarver og 1 oter som fanget av referansefiskerne. Ingen i teiner, bare i ruser.
Overholdes regelverket vil overlevelsen til både fangst og bifangstarter antageligvis være høy grunnet kort ståtid og ved at fisket kun foregår på grunt vann. Havforskningsinstituttet har ikke kjennskap til at kontrollmyndighetene har avdekket brudd på regelverket rundt gjenutsetting eller rapporter om ulovlig beholdt bifangst. Det er likevel en del kunnskapshull, spesielt knyttet til omfang og konsekvenser av gjenutsetting av bifangst i ulike avstander fra fangststedet, samt predasjon i redskapen. Hvis det forekommer avvik fra regelverket ved at bifangst beholdes eller settes ut på dypere vann, så er det sannsynlig at fisket i en eller annen grad påvirker bestandene av bifangstarter.
3.5 - Mulige indirekte økologiske effekter av fisket etter leppefisk
Arter kan påvirkes indirekte (negativt eller positivt) ved en endring i artssammensetning som en følge av uttak fra bestandene. Leppefisk har en sentral rolle i kystøkosystemet. De er opportunistiske beitere på en rekke bunnlevende og fastsittende dyr som en rekke arter virvelløse dyr, blant annet krepsdyr, snegler, muslinger, børstemark (Alvsvåg, 1993, Sayer mfl., 1995; Deady and Fives, 1995 , Bourlat mfl., 2021). Videre er de byttedyr for større fiskearter og sjøfugl (Steven 1933, Nedreaas mfl., 2008, Olsen mfl., 2018). Leppefisk er også føde for større fisk, som kysttorsk, lyr, lange og ål, og i noen områder sjøfugl (skarv). Hvis lokale bestander av leppefisk fiskes ned, vil dette kunne resultere i et redusert fødetilbud for andre arter (Bergström mfl., 2016).
En nedfisking av lokale leppefiskbestander vil kunne ha en effekt på mengde og tetthet av disse organismene og således på bunnfauna og flora (begroing). En studie fra Sverige har påpekt at et fiske av leppefisk kan i så måte ha en positiv effekt på tareskogen, ved at en for høy tetthet av leppefisk beiter ned snegler og krepsdyr som igjen beiter på trådalger (Östman mfl., 2016). I en annen nylig publisert eksperimentell studie ble det rapportert at høy tetthet av bergnebb kan ha en positiv effekt på flerårige makroalger og forfatterne advarer mot at en reduserte bestander av leppefisk kan påvirke bunnsamfunn i negativ retning dominert av trådformede grønnalger (Kraufvelin mfl., 2020). Samme studiet viser at blåskjell er en viktig del av dietten til leppefisk - og det er blitt fremsatt en hypotese om at redusert forekomst av blåskjell kan ha en sammenheng med økt tetthet av leppefisk. De økologiske interaksjonene er likevel for lite undersøkt til at man kan si noe håndfast om økologiske effekter av overuttak av leppefisk, spesielt siden fødevalg og habitatvalg er ulikt både mellom arter og størrelsesklasser. Det er ingen kjente studier av effekter av fiskeriet på økosystemet som helhet, som en følge av en eventuell endret artssammensetning, størrelsesfordeling og tetthet av leppefisk. Bifangst bestående av undermåls leppefisk og andre arter skal etter regelverket settes ut igjen på en slik måte at de overlever. Det er et klart behov for målrettet forskning for å bedre forstå de økologiske sammenhengene mellom leppefisk og andre arter, og hvordan disse påvirkes av endringer i arts og størrelsessammensetning av leppefisk som en følge av fiskeriet.
3.6 - Kunnskapsstyrke og forskningsprioriteringer
Det har blitt gjennomført omfattende forskning på Austevoll på fiskerieffekter de siste 8 årene. Resultatene tyder på at fiskeriet har en begrenset påvirkning på grønngylt og bergnebb på tross av høy fiskeintensitet i dette området (Kunnskapstøtte til Fiskeridirektoratet 2019, Halvorsen mfl., 2020c, Halvorsen mfl., 2021). Berggylt er lite tallrik i dette område, men etablering av bevaringsområde har doblet fangstratene på 5 år. Data fra ruse-garn toktet gir et inntrykk at det har pågått et overfiske på berggylt, men at trenden har snudd noe de siste par årene. Med tilgjengeliggjøring av sporingsdata vil det bli mulig å få detaljert oversikt over hvordan fiskertrykk fordeler seg i tid og rom, noe som vil gjøre det mulig å analysere hvordan fiskeintensitet påvirker fangstrater. Dette er et prioritert arbeid de neste årene og det er allerede laget en god modell for å estimere fiskeintensitet i og rundt Austevoll som skal oppskaleres til nasjonalt nivå i 2024. Det er behov for å undersøke konsekvensene av selektivt uttak av artene med kjønnsdimorfisme, hvor hannene står for yngelpleie og større hanner bygger større og flere reir (Kindsvater mfl., 2020). HI har delfinansiert en PhD-stilling som skal jobbe med dette i perioden 2023-2026. Undersøkelser av dette vil være viktig for å skaffe mer kunnskap om hvilke konsekvenser fisket etter leppefisk vil ha både på kort og lengre sikt. Videre så mangler en del kunnskap om de ulike leppefiskartenes økologiske funksjon som predator og byttedyr, og hvordan endringer i leppefiskbestandene kan ha negative/positive konsekvenser forandre deler av kystøkosystemet. Disse problemstillingen er fokus for prosjektet «Forståelse av menneskeskapte trofiske kaskader i kystøkosystemer – CASCADES”, som er et samarbeid mellom Norsk institutt for Naturforskning (NINA), HI og Universitetet i Bergen (UiB) og internasjonale partnere.
Det er begrenset forståelse av hvilke mekanismer som regulerer rekruttering og variasjoner i bestandene (f.eks sommer- og vintertemperatur, tetthetseffekter), og i hvilken grad fiskeriet har en regulerende innvirkning på bestandsstørrelsene hos de ulike artene. Pågående forskning ved Havforskningsinstituttets forskningsstasjon på Austevoll undersøker dette. Datagrunnlaget er for bestandsutviklingen er god på Sørlandet gjennom strandnotundersøkelsen og årlige rusetokt. Det er tilsynelatende ikke noen klar nedgang i bestandene etter fisket ble intensivert fra 2010. For resten av landet er det samlet inn data på lengde og fangst-per-enhet-innsats fra referansefiskere siden 2011. Det har vært utfordrende å konkludere med bestandsendringer basert på disse dataene, grunnet stor variasjon i fangstrater innad og mellom år, og mellom fiskere, men hovedinntrykket er at det er relativt stabile fangstrater de siste fem årene. Referansefiskerdataene har gitt omfattende kunnskap om bifangst, og om regelverket følges og bifangst slippes ut raskt nært land kan man anta at det er liten grad av påvirkning. Det er likevel ønskelig med mer konkret kunnskap om skjebnen til utsatt bifangst, både leppefisk og andre arter. For dette er det gjort forsøk som undersøker hvorvidt leppefisk evner å ta seg tilbake når de blir sluppet ut i ulike avstander fra fangststedet. Det er også samlet inn data på fiskeripraksis for utsetting av bifangst i ulike deler av landet i 2023. Resultatene fra disse studiene er planlagt publisert i løpet 202 4 .
4 - Modellering av fiskeintensitet i leppefisk-fiskeriet ved bruk av elektronisk fartøysporing
Fiskeriet etter leppefisk foregår langs store deler av Norskekysten, hvor flere hundre mindre fartøy deltar. Fangstredskapen er i hovedsak agnede teiner på lenker. Lengden på lenken (og antall teiner), kan variere betydelig. Fiskeriet foregår på grunt vann (<10 meter) og i hovedsak på hardbunn. Fiskeriet har tre målarter; grønngylt, bergnebb og berggylt. Grasgylt er også en vanlig art i dette fisket, men blir kun sporadisk brukt som rensefisk (< 2 % av totalfangsten på Vestlandet). Det fanges også en del rødnebb/blåstål, men disse blir ikke brukt som rensefisk. All bifangst må slippes tilbake i sjø nært land i henhold til gjeldene reguleringer. Et fellestrekk for de fem leppefiskartene er en høy grad av stedbundenhet, noe som tilsier at høy fiskeintensitet kan ha en betydelig effekt på lokale, demografisk isolerte bestander (begrenset utveksling av juveniler og voksne individer med nærliggende bestander). Det har vært begrenset kunnskap om den romlige fordelingen av fiskeintensitet da fiskerne ikke er pliktige om å rapportere hvor de fisker. Fra 2020 ble det innført obligatorisk elektronisk sporing i dette fiskeriet (med unntak av ungdomsfiskere). Data samles inn ved bruk av AIS (Automatic Identification System). Dette er et sporingssystem for å identifisere og lokalisere fartøy ved elektronisk utveksling av data med andre nærliggende fartøy, AIS-basestasjoner og satellitter. Alternativt kan fartøyene bruke VMS (Vessel Monitoring System), som er et satellittbasert overvåkingssystem.
AIS /VMS data kan brukes til å få verdifull informasjon om fordeling av fisketrykk i tid og rom, og gi mulighet for å bedre studier av generelle effekter av fiskeriet på bestandene i de kommende årene. Dette vil skje i prosjektet « Forståelse av menneskeskapte trofiske kaskader i kystøkosystemer – CASCADES”, som er et samarbeid mellom Norsk institutt for Naturforskning (NINA), Havforskningsinstituttet (HI) og Universitetet i Bergen (UiB) og internasjonale partnere, finansiert av Norges forskningsråd. Sentralt i prosjektet er at vi vil bruke sporingsdata til å undersøke sammenhengen mellom fisketrykk i leppefisk-fiskeriet og direkte og indirekte konsekvenser for kystøkosystemet. I denne rapporten oppsummerer vi status, utfordringer og muligheter for prosjektet. Dette er et pågående arbeid, og det kan derfor komme endringer i metodikk og resultater, men det vil presenteres foreløpige data og bruk for å estimere fiskeintensitet for Austevoll-området.
4.2 - Data og metodebeskrivelse
Sporingsdata blir innsamlet og lagret av Kystverket. Data fra fiskesesongen 2022 ble gjort tilgjengelig våren 2023. Vi har frem til nå hatt fokus på å lage en modell for Austevoll og omkringliggende kystområder. Austevoll ble valgt da dette er en av kommunene med høyest antall registrerte fartøy og flere oppdrettsanlegg som mottar leppefisk. I tillegg har dette vært et fokusområde for forskning på leppefiskbestandene gjennom flere år, og det er flere marine verneområder. All analyse av fiskeintensitet er gjort i statistikkprogrammet R. Alle R-skript er skrevet slik at de kan skaleres opp til det nasjonale fiskeriet når modelljusteringen er fullført.
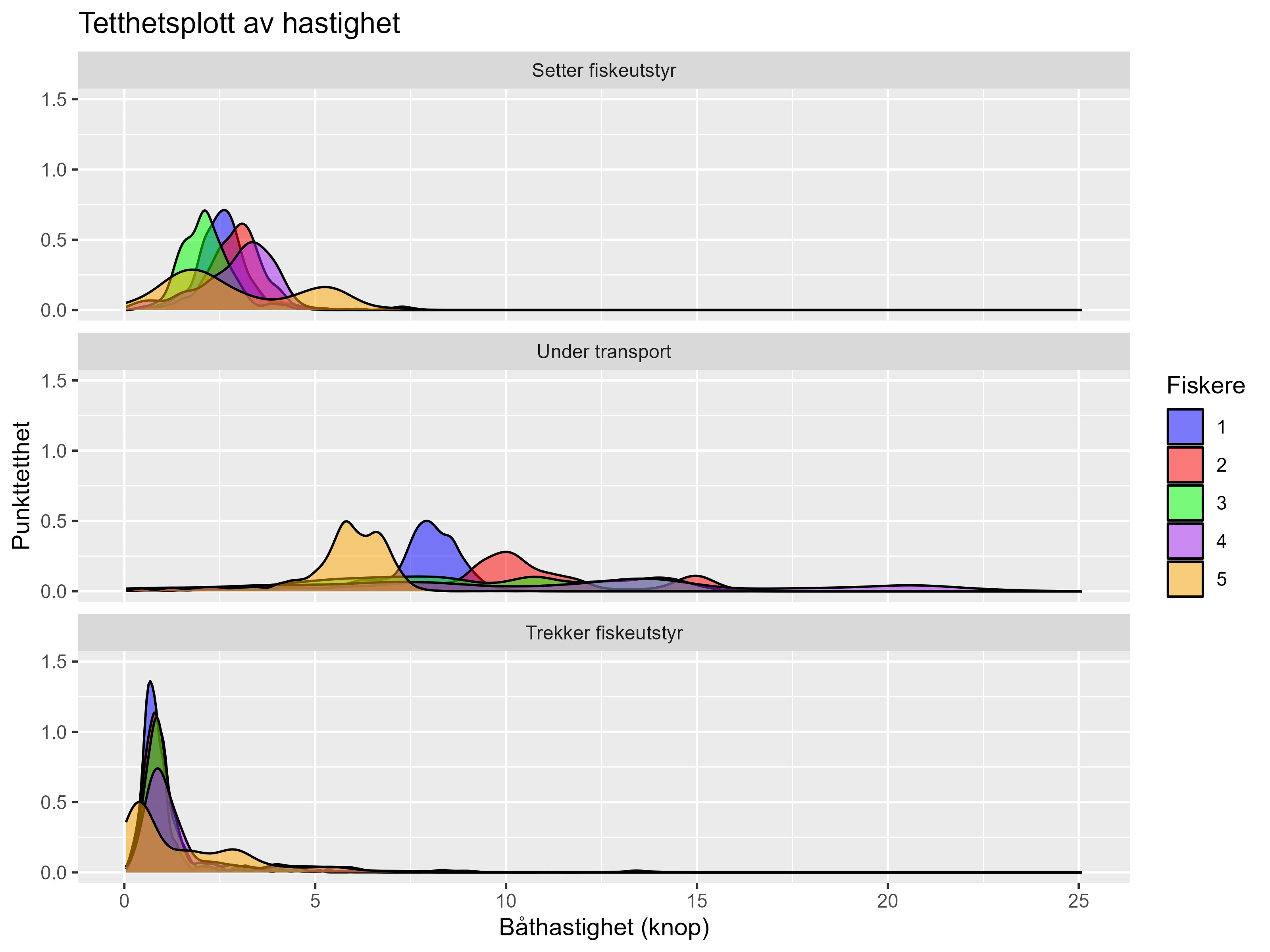
AIS og VMS data gir informasjon om posisjon og hastighet til fartøyet. Ved fangst og trekk av redskap (aktivt fiske ) vil fartøyene gå i lav hastighet nært land – slik at fiskeriaktivitet gir definerte mønstre i dataene. Første steg er derfor å definere hastighetsspennet for aktivt fiske og deretter gruppere datapunktene romlig innenfor disse hastighetsverdiene. For å få validerte observasjoner av aktivitetsmønsteret og hastigheter så gjennomførte vi en datainnsamling fra et utvalg av fiskefartøy i juli 2023, hvor vi manuelt registrerte fiskernes aktivitet i tid og rom. Data på posisjon og hastighet ble samlet inn ved hjelp av håndholdt GPS. Disse ble så brukt for å definere hvilke hastigheter som korresponderte til ulike aktiviteter (Figur 4.1). For å modellere fiskeriaktivitet må sporingsdataene først filtreres for å ekskludere spor der fiskeren legger til kai eller leverer fisk til oppdrett siden hastighetene for disse aktivitetene overlapper den hastigheten fartøyene har når de fisker. I tillegg er det betydelige forskjeller i AIS- og VMS-data som gir utfordringer. Sendefrekvens for AIS er variabel og blant annet avhengig av fartøyets hastighet. Oppløsningen kan derfor variere betydelig i løpet av en fisketur (fra nesten en gang i sekundet til flere minutter). VMS har en standard sendefrekvens på et signal per hvert tiende minutt. Mangelen på standardiserte sendefrekvenser av AIS-data nødvendiggjør en gruppering av fiskepunkter etter fisker, hvor spor som representerer aktivt fiske (fiskespor) kan tegnes mellom sendepunkter. Disse fiskesporene ble deretter lagt over et polygon-nett (Figur 4.2), og romlig fiskeintensitet kan da estimeres som antall fiskespor per polygon (Figur 4.3). Størrelsen på polygonene er justerbar, noe som gjør det mulig for oss å modellere fiskeintensitet på ulike geografiske skalaer.
I utgangspunktet var det ønskelig å bruke samme metodikk for VMS-dataene som for AIS for å kunne gjøre en 1:1 sammenlikning av fiskeintensitet for fartøy med ulike type sendeutstyr, men den tidsmessige oppløsningen i VMS-data er for lav for å muliggjøre en gruppering av punkter mellom separate hendelser med sett og trekk av utstyr. Ifølge ombord-observasjonene bruker et fartøy i gjennomsnitt 7 minutter på sett/trekk av utstyr. De fleste hendelsene med aktivt fiske blir derfor representert av et enkelt punkt. En sendefrekvens på 10 minutter er vanligvis ikke nok til å gruppere flere punkter med aktivt fiske til spor. En slik filtrering ville føre til at mye av det faktiske fiskeintensiteten ikke blir detektert. Det ble derfor valgt å modellere VMS data basert på tetthet av punkter istedenfor spor. Dette medfører antageligvis at VMS data gir en noe redusert presisjon og romlig oppløsning når det brukes til å estimere fiskeintensitet i dette spesifikke fiskeriet. 2022 datasettet fra Austevoll inkluderer 50 fartøy med AIS og 4 fartøy med VMS.
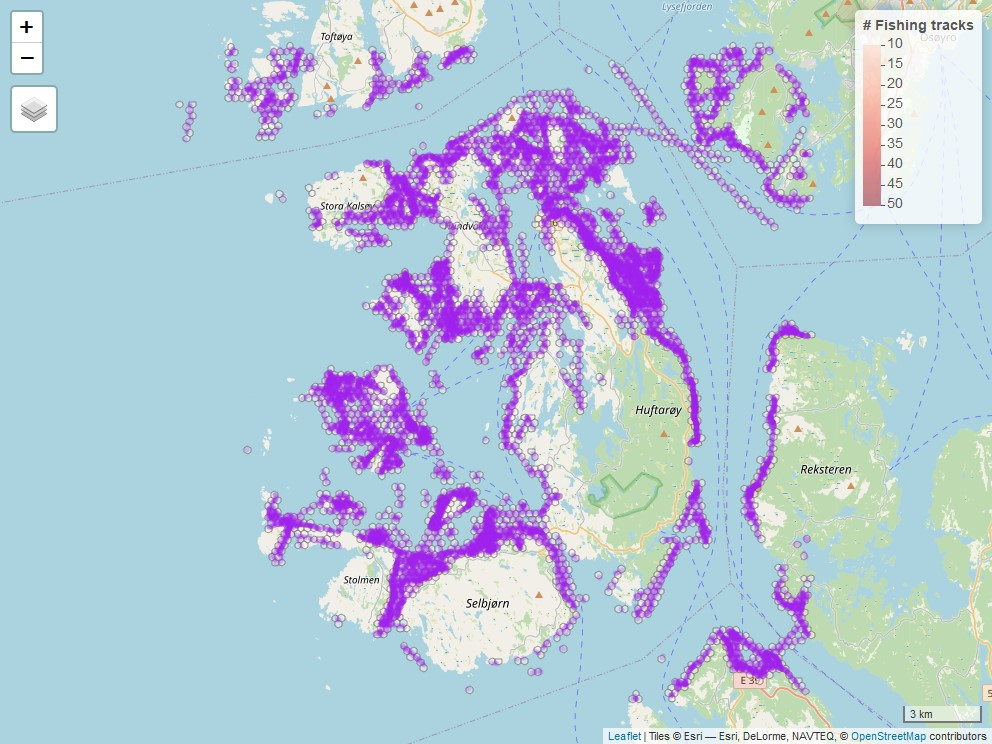
Den nåværende modellen av fiskeintensitet er kan visualiseres i et interaktivt kart, hvor fartøyenes hjemmehavner, oppdrettslokasjoner, fiskespor og polygoner kan slås av og på, sammen med flere bakgrunnskart. Man kan zoome inn på spesifikke områder av kystlinjen for å undersøke fiskespor rundt interesseområder. For eksempel kan man undersøke overholdelse av fiskereguleringer i fredningsområder (Figur 4.4). Det interaktive kartet kan utforskes i en nettleser.
Fiskepressmodellen gir per nå et godt bilde av fiskeriaktivitet i og rundt Austevoll, men det gjenstår litt justeringer og ytterligere filtreringssteg før den kan oppskaleres på nasjonalt nivå. Den største mulige feilkilden i den nåværende modellen er relatert til punkter (og relaterte spor) som har registrerte hastighetsverdier som faller innenfor vår definisjon av fiskeintensitet, selv om de skjer på dyp som ikke kan tilskrives aktivt fiske etter leppefisk. Slike spor skjer eksempelvis når et fartøy tar pauser i fisket et stykke fra land. Filtrering av disse punktene kan oppnås ved å beregne avstand til nærmeste kystlinje, og filtrere bort punkter som overstiger en definert avstand fra land. Dybdedata kan også brukes, selv om det sannsynligvis ville være mye mer krevende å beregne, og antageligvis kun gi en begrenset forbedring sammenliknet med den romlige filtreringen beskrevet ovenfor.
I tillegg er det behov for å validere modellen for ulike deler av kysten der fiskeripraksis kan være annerledes. For eksempel er det vanlig å bruke mindre åpne fartøy på Sørlandet og færre teiner på lenke. Derfor kan bevegelsesmønstret være annerledes. Det er samlet inn ombord data fra to fiskere på Sørlandet som vil benyttes til å definere hastigheter for mindre fartøy.
Figur 4.1. Tetthetsplott av hastighetsverdier for når fiskerne setter og trekker fiskeutstyr og under transport. Dataene er samlet inn på kommersielle leppefisk-fartøy av forskningspersonell med håndholdt GPS. Antall GPS-punkter er henholdsvis 3731, 6263 og 50,811 for de tre aktivitetskategoriene.
Figur 4.2. Plassering av AIS fiskespor i og rundt Austevoll Kommune i løpet av fiskesesongen 2022.
Figur 4.3. Antall fiskespor, gruppert etter sted. Polygonstørrelse er satt til 250 meter i tverrsnitt.
Figur 4.4. Mangel på fiskespor i hummerfredningsområdet i Austevoll (rød polygon).
4.3 - Anvendelse: Sammenheng mellom fiskeintensitet og fangstrater for leppefisk
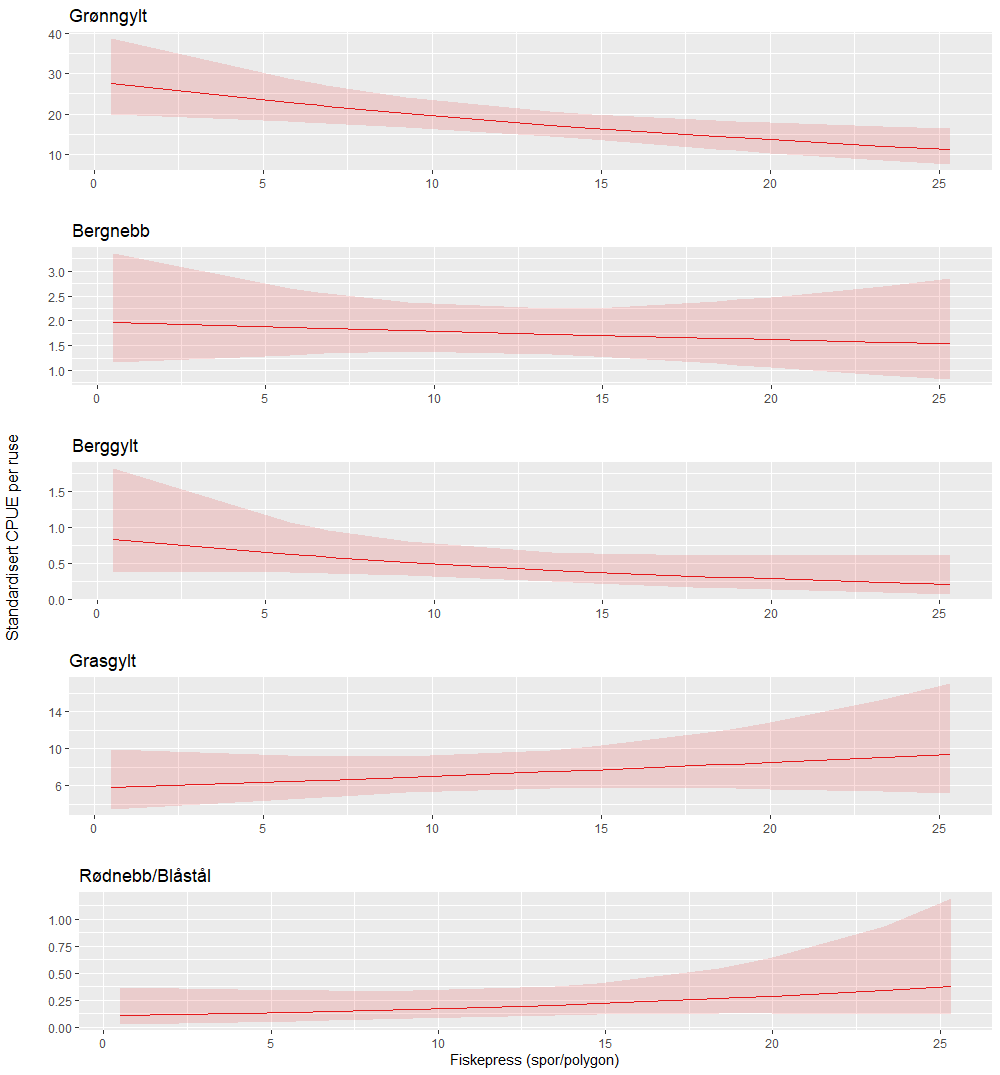
Modellen vil bli et sentralt verktøy for å forstå i hvilken grad fiskeintensitet påvirker leppefiskbestandene og har eventuelt andre økologiske konsekvenser. For å demonstrere dette har vi gjennomført noen foreløpige analyser om fiskeintensitet forklarer forskjeller i fangstrater for de ulike leppefisk artene. Det ble gjennomført et forsøksfiske med ruser i ulike områder i Austevoll i juni 2023 (Figur 4.5). All fangst i rusene ble lengdemålt og satt ut igjen på fangststedet. Med modellen for fiskeintensitet var det mulig å teste effekten av fiskeintensitet på fangst-per-ruse (Catch-per-unit-effort – CPUE). Dette ble gjort ved å gruppere stasjonene i mindre områder (stasjonene lå innen 500m fra hverandre og/eller ikke adskilt av dyp dypere enn 20m) og deretter regne gjennomsnittlig verdi av fiskeintensitet (antall spor i polygonene der stasjonene ligger) for hvert område. Stasjoner i verneområder er ekskludert fra analysen. Gjennomsnittsverdi for fiskeintensitet ble så inkludert i en GLM (Generalized linear model) sammen med data på temperatur, dybde og ståtid for å estimere CPUE av leppefisk (over minstemål). De undersøkte områdene hadde gjennomsnittlige verdier for fiskeintensitet som varierte mellom 0.5 og 25 spor per polygon.
Grønngylt var den eneste av artene hvor man finner en signifikant effekt av fiskeintensitet – her er CPUE lavere i områder som hadde høyt antall fiskespor i 2022 (Figur 4.6). Det var også nær signifikante effekter på CPUE for berggylt. De ikke-kommersielle artene, grasgylt og rødnebb/blåstål hadde ingen signifikant effekt av fiskeintensitet, men trenden for disse artene er at CPUE øker med fiskeintensitet, altså motsatt av det vi observerte for de tre kommersielle artene. Dette er viktig å understreke at dette pågående analyser og før man konkluderer så er det behov for å modellere fiskeintensitet i for 2020 og 2021 før man konkluderer, da vi per nå ikke kjenner til om det er de samme områdene som fiskes hardest hvert år eller om det flytter seg etter som fiskerne responderer på endrede fangstrater. Dette er spesielt viktig for berggylt og bergnebb, artene som har en lengre generasjonstid. For disse er det rimelig å forvente at det kumulative fiskeintensitet et område har blitt utsatt for over flere år vil ha større forklaringsverdi enn fiskeintensiteten det foregående år. Likevel demonstrerer dette en nyttig bruk av fiskeintensitet modellen som gjør det mulig å kvantifisere effekten av fiskeriet på bestandene på en helt ny måte. Den samme metodikken kan brukes på å undersøke om fiskeintensitet forklarer fangstrater hos referansefiskerne eller i det årlige garntoktet som brukes for å følge utviklingen i berggylt-bestanden over et større område på Vestlandskysten.
Figur 4.5. Kart over ruse stasjoner i Austevoll kommune. Nærliggende stasjoner ble gruppert i områder som fikk beregnet en gjennomsnittlig fiskeintensitet (antall fiskespor per polygon).
Figur 4.6. Estimert CPUE (leppefisk over minstemål) per art i forhold til fiskeintensitet. Det er brukt en GLMM-modell med standardisering av temperatur, dybde og ståtid. Merk ulike skalaer på y-aksene.
4.4 - Veien videre og forventninger om tilgjengeliggjøring av data:
Det mulig å knytte spesifikke fisketurer/fartøy til fangststatistikk fra Fiskeridirektoratet. Dette kan brukes til å utvikle en fangst-per-enhet-innsats indeks, samt detaljerte data på mengde leppefisk fisket i ulike geografiske områder, for eksempel produksjonsområder. Per i dag kan fangststatistikken knyttes til fartøy, men det er ikke info om hvor fartøyet har fisket – slik at man kun har statistikk på de ulike forvaltningsregionene. Slike data vil gjøre det mulig å modellere fiskeintensitet ved å estimere antall fisk tatt fra forskjellige områder, snarere enn ved å kvantifisere fiskepunkter. Fiskeintensitet-modellen kan bli bedre ved å inkorporere informasjon om fangst per enhetsinnsats og endringer i fangst over løpet av fiskesesongen – noe som forbedrer modellnøyaktigheten betydelig.
For at sporingsdata skal kunne brukes til å studere effekten av leppefiskeriet på regionalt og nasjonalt nivå er det behov for en forutsigbar tilgjengeliggjøring av data. Data fra 2020 og 2021 er ikke tilgjengelige per dags dato. I prosjektet er det planlagt å bruke data fra 2020-2024 til å estimere endringer i fiskeintensitet i tid og rom i ulike regioner, slik at forvaltningen og andre interessenter kan få tilgang til oppdatert informasjon om hvordan dette er fordelt. Dette er eksempelvis etterspurt av HIs årlige risikovurdering av norsk oppdrett, hvor man vurderer konsekvenser og risiko i de ni produksjonsområdene langs kysten. Effekter av fangst og bruk av rensefisk har et eget kapittel. Videre, ved å sammenligne områder med varierende fiskeintensitet, er det et mål å undersøke sammenhengene mellom fiskeaktivitet og endringer i tetthet, art- og størrelsessammensetning i lokale leppefiskbestander. Dette vil videre brukes til å undersøke indirekte økologiske effekter av fisket etter leppefisk på andre arter. Det vil være viktige leveranser i CASCADES prosjektet samt HIs overvåkning. Det er planlagt flere publikasjoner, og selve metodikken for å estimere fiskeintensitet ved sporingsdata er planlagt publisert i 2024.
5 - Referanser
Alvsvåg J (1993). Fødeval, vekst og energiallokering hos gress gylt ( Centrolabrus exoletus L.) og grønngylt ( Symphodus melops L.) (Pisces: Labridae) på vestkysten av Noreg, og diettoverlapping med O og l-gruppe torsk ( Gadus morhua L.). University of Bergen.
Balasch, J.C.; Tort, L. Netting the stress responses in fish. Frontiers in Endocrinology 2019, 10, doi:10.3389/fendo.2019.00062.
Bergström L, Karlsson M, Bergström U, Pihl L, Kraufvelin P (2016) Distribution of mesopredatory fish determined by habitat variables in a predator-depleted coastal system. Marine Biology 163 (10): p. 201.
Bollinger PJ (2020). Biological control of salmon lice: A critical analysis of knowledge production and development in the Norwegian cleaner fish industry. Master thesis, University of Life Sciences.
Bourlat SJ, Faust E, Wennhage H, Wikström A, Rigby K, Vigo M, Kraly P, Selander E, André C (2021) Wrasse fishery on the Swedish West Coast: towards ecosystem-based management. ICES J. Mar. Sci., 78, 1386-1397, doi:10.1093/icesjms/fsaa249
Darwall WRT, Costello MJ, Donnelly R, Lysaght S (1992). Implications of life-history strategies for a new wrasse fishery. Journal of Fish Biology 41 (sb): 111–123.
Deady S, Fives JM (1995). Diet of ballan wrasse, Labrus bergylta, and some comparisons with the diet of corkwing wrasse, Crenilabrus melops. Journal of the Marine Biological Association of the United Kingdom 75 (03): 651–665. Cambridge University Press.
Faust E, Jansson E, André C, Halvorsen KT, Dahle G, Knutsen H, Quintela M, Glover KA (2021) Not that clean: Aquaculture-mediated translocation of cleaner fish has led to hybridization on the northern edge of the species' range. Evolutionary Applications, 14, 1572-1587, doi:https://doi.org/10.1111/eva.13220
Faust E, Halvorsen KT, Andersen P, Knutsen H, André C (2018). Cleaner fish escape salmon farms and hybridize with local wrasse populations. Royal Society Open Science 5(3): 171752.
Fernández‐Chacón AD, Villegas‐Ríos E, Moland ML, Baskett EM, Carlson SM (2020). Protected areas buffer against harvest selection and rebuild phenotypic complexity. Ecological Applications:eap. 2108. Ecological Society of America.
Halvorsen KT, Sørdalen TK, Durif C, Knutsen H, Olsen EM, Skiftesvik AB, Rustand TE, Bjelland RM, Vøllestad LA (2016). Male-biased sexual size dimorphism in the nest building corkwing wrasse (Symphodus melops: implications for a size regulated fishery. ICES Journal of Marine Science 73 (10): 2586–2594.
Halvorsen KT, Bjelland R, Jørgensen T, Skiftesvik A B (2017a). Forsøksfiske for selektiv fangst av berggylt. Fisken og Havet (8).
Halvorsen KT, Larsen T, Sørdalen TK, Vøllestad LA, Knutsen H, Olsen EM (2017b). Impact of harvesting cleaner fish for salmonid aquaculture assessed from replicated coastal marine protected areas. Marine Biology Research 13(4): 359–369.
Halvorsen KT, Sørdalen TK, Vøllestad LA, Skiftesvik AB, Espeland SH, Olsen EM (2017c). Sex- and size-selective harvesting of corkwing wrasse (Symphodus melops)–a cleaner fish used in salmonid aquaculture. ICES Journal of Marine Science 74(3): 660–669.
Halvorsen K, Skiftesvik AB, Jørgensen T (2019). Kunnskapsstøtte – anbefaling om redusert inngangsstørrelse i teiner i fisket etter leppefisk.
Halvorsen KT, Sørdalen TK, Larsen T, Browman HI, Rafoss T, Albretsen J, Skiftesvik AB (2020b). Mind the depth: The vertical dimension of a small-scale fishery shapes selection on species, size and sex in wrasses. Marine and Coastal Fisheries 12 (6), 404–422. https://doi.org/10.1002/mcf2.10131
Halvorsen K, Sørdalen TK, Larsen T, Rafoss T, Skiftesvik AB (2020c). Kunnskapsbasert innovasjon for optimal ressursutnyttelse i leppefiskeriet. Rapport fra Havforskningen.
Halvorsen K., Skiftesvk, AB., Larsen T., Otterå H., og AF Chacón. 2021. Kunnskapsstøtte og råd for regulering av fisket etter leppefisk i 2022. Rapport fra Havforskningen 2021-54 ISSN: 1893-4536
Jansson E, Quintela M, Dahle G, Albretsen J, Knutsen H, André C, Strand Å, Mortensen S, Taggart JB, Karlsbakk E, Kvamme BO, Glover KA (2017). Genetic analysis of goldsinny wrasse reveals evolutionary insights into population connectivity and potential evidence of inadvertent translocations via aquaculture. ICES Journal of Marine Science. doi:10.1093/icesjms/fsx046
Jansson E, André C, Quintela M, Halvorsen KT, Besnier F, Ayllon F, Faust E, Knutsen H, Strand Å, Glover KA (2023a) Genetic study reveals local differentiation persisting in the face of high connectivity and a genomic inversion likely linked with sexual antagonism in a common marine fish. ICES J. Mar. Sci. 2023, doi:10.1093/icesjms/fsad042.
Jansson E, Faust E, Bekkevold D, Quintela M, Durif C, Halvorsen KT, Dahle G, Pampoulie C, Kennedy J, Whittaker B, et al. (2023b) Global, regional, and cryptic population structure in a high gene-flow transatlantic fish. PLOS ONE 2023, 18, e0283351, doi:10.1371/journal.pone.0283351.
Jørgensen T, Bjelland R, Halvorsen K, Durif C, Shema S, Skiftesvik AB (2017). Seleksjon i leppefiskredskap. Fisken og Havet (2).
Kindsvater HK, Halvorsen, KT, Sørdalen, TK, Alonzo SH (2020). The consequences of size‐selective fishing mortality for larval production and sustainable yield in species with obligate male care. Fish and Fisheries. Early view online https://doi.org/10.1111/faf.12491
Kraufvelin P, Christie PH, Gitmark JK (2020) Top-down release of mesopredatory fish is a weaker structuring driver of temperate rocky shore communities than bottom-up nutrient enrichment. Marine Biology 167(4): 1–20. Springer Berlin Heidelberg.
Muncaster, S., B. Norberg, and E. Andersson. 2013. Natural sex change in the temperate protogynous Ballan wrasse Labrus bergylta. Journal of Fish Biology 82(6): 1858–1870. Blackwell Publishing Ltd.
Nedreaas K, Aglen A, Gjøsæter J, Jørstad K, Knutsen H, Smedstad O, Svåsand T, Ågotnes P (2008). Kysttorskforvaltning på Vestlandet og langs Skagerrak-kysten. Vurdering av status for kysttorsk på strekningen svenskegrensen–Stad med forslag om forvaltningstiltak. Fisken og Havet, 5–2008. 106 pp.
Neghabath N (2022). Homing ability of goldsinny wrasse (Ctenolabrus rupestris) and corkwing wrasse (Symphodus melops). Masteroppgave, Universitetet I Agder.
Olsen EM, Halvorsen KT, Larsen T, Kuparinen A (2018). Potential for managing life history diversity in a commercially exploited intermediate predator, the goldsinny wrasse ( Ctenolabrus rupestris ). ICES Journal of Marine Science 76(1): 357–357.
Reamon, M (2020). Assessing the impacts of marine protected areas on wrasse populations in Norway. Master thesis. University of Agder.
Ruud, M (2020). Catchability and selectivity in a multispecies wrasse fishery in Western Norway. Master thesis. University of Agder.
Sayer MDJ, Gibson RN, Atkinson RJA (1995). Growth, diet and condition of goldsinny on the west coast of Scotland. Journal of Fish Biology 46 (June 1993): 317–340.
Seljestad GW, Quintela M, Faust E, Halvorsen K, Besnier F, Jansson E, Dahle G, Knutsen H, André C, Folkvord A, Glover KA (2020) “A cleaner-break”: Genetic divergence between geographic groups and sympatric phenotypes revealed in ballan wrasse ( Labrus bergylta ). Ecology and Evolution. Crossref DOI link: https://doi.org/10.1002/ECE3.6404
Sistiaga, M, Herrmann, B, Jørgensen T (2021) Prediction of goldsinny wrasse ( Ctenolabrus rupestris ) minimum size required to avoid escape through salmon ( Salmo salar ) farm nets. Aquaculture, 543, 737024, doi:https://doi.org/10.1016/j.aquaculture.2021.737024.
Skiftesvik AB , Bjelland RM, Durif CMFI, Johansen IS, Browman HI (2013). Delousing of Atlantic salmon ( Salmo salar ) by cultured vs. wild ballan wrasse ( Labrus bergylta ). Aquaculture 402-403, 113–118.
Skiftesvik AB, Blom G, Agnalt A-L, Durif CMF, Browman HI, Bjelland RM, Harkestad LH, Farestveit E, Paulsen OI, Fauske M, Havelin T, Johnsen K, Mortensen S (2014a). Wrasse (Labridae) as cleaner fish in salmonid aquaculture – The Hardangerfjord as a case study. Mar Biol Res 10, 289–300.
Skiftesvik AB, Durif D, Bjelland R, Browman H, Holm E, Jørgensen T, Løkkeborg S, Utne Palm AC, Skar Tysseland AB, Hoddevik Ulvestad B, Aasen A, Axelsen BE, Mortensen S, Sælemyr L, Skår CK, Karlsbakk E, Einen ACB, Nordbø J, Omdal LM, Isachsen CH, Espeland SH, Semb Johannesen I, Olausson S og Elen E ( 2014b). Bestander og fangstkvaliteter av leppefisk. Rapport fra Havforskningen nr. 3–2014. Sluttrapport FHF prosjekt 900609.
Skiftesvik AB, Durif CMF, Bjelland RM, Browman HI (2015). Distribution and habitat preferences of five species of wrasse (Family Labridae) in a Norwegian fjord. ICES Journal of Marine Science 72(3): 890–899.
Skiftesvik AB, Bjelland R, Durif C, Moltumyr L, Hjellum RB, Halvorsen KT (2018) Adferd og artssamspill I laksemerder. Rapport fra Havforskningen nr. 33-2018, Sluttrapport FHF prosjekt 900978.
Steven GA (1933). The food consumed by shags and cormorants around the shores of Cornwall (England). Journal of the Marine Biological Association of the United Kingdom 19 (01): 277. Cambridge University Press.
Sørdalen TK, Halvorsen KT, Vøllestad LA, Moland E, Olsen EM (2020). Marine protected areas rescue a sexually selected trait in European lobster. Evolutionary Applications: 1–12.
Treasurer JW (2012). Diseases of north European wrasse (Labridae) and possible interactions with cohabited farmed salmon, Salmo salar L. Review article. J Fish Dis 35, 555–562.
VKM, Eli Rueness, Paul R. Berg, Snorre Gulla, Kim Halvorsen, Johanna Järnegren, Martin Malmstrøm, Tor Atle Mo, Espen Rimstad, Hugo de Boer, Katrine Eldegard, Kjetil Hindar, Lars Robert Hole, Kyrre Kausrud, Lawrence Kirkendall, Inger Måren, Erlend B. Nilsen, Eva B. Thorstad, Anders Nielsen and Gaute Velle (2019) Assessment of the risk to Norwegian biodiversity from import of wrasses and other cleaner fish for use in aquaculture. Opinion of the Panel on Alien Organisms and Trade in Endangered Species (CITES) of the Norwegian Scientific Committee for Food and Environment. VKM report 2019: 15, ISBN: 978-82-8259-330-4, ISSN: 2535–4019. Norwegian Scientific Committee for Food and Environment (VKM), Oslo, Norway
Wienerroither R, Johannesen E, Dolgov A, Byrkjedal I, Bjelland O, Drevetnyak K, Eriksen K, Høines Å, Langhelle G, Langøy H, Prokhorova T, Prozorkevich D, Wenneck T (2011). Atlas of the Barents Sea Fishes. IMR/PINRO Joint Report Series (ISSN 1502-8828). 1, 272 s.
Östman Ö, Eklöf J, Eriksson BK, Olsson J, Moksnes PO, Bergström U (2016). Top-down control as important as nutrient enrichment for eutrophication effects in North Atlantic coastal ecosystems. Journal of Applied Ecology 53(4): 1138–1147.