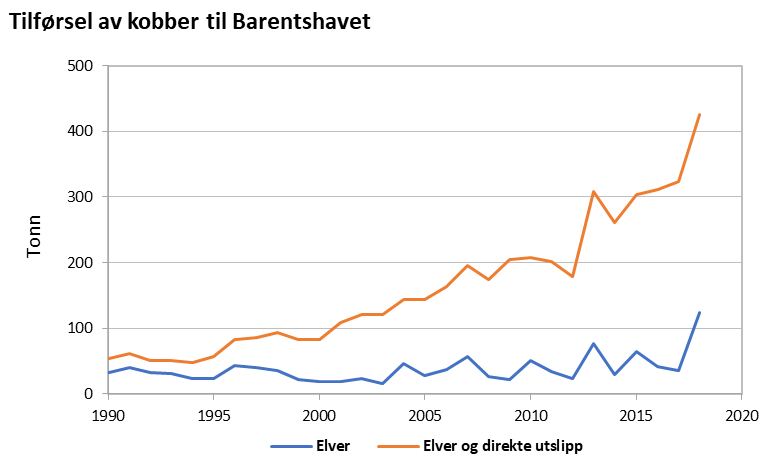This report addresses three major themes: (1) Dominant features in the status and trends of the Barents Sea since 2016, with an emphasis on describing how central processes affect ecosystem condition; (2) a more detailed description of the status and trends in various components of the ecosystem; and (3) a description of related fisheries management measures implemented in the Barents Sea.
Dominant trends and central processes
Natural variation in the Barents Sea includes considerable increase or decrease in temperature within a few years. After nearly four decades of a generally increasing trend, water temperatures dropped significantly after 2016, and remained at the same level in 2019 as observed during the early 2000s. The extent of sea ice in the Barents Sea declined significantly during the decades of warming. However, over the period 2016-2019, the extent of sea ice has increased and the ice edge has moved soutwards. Just as warming up through 2016 had significant effects on the Barents Sea ecosystem, in the form of increased distribution of southern species and declines in Arctic populations, increasing numbers in certain Arctic zooplankton and fish species have been observed after 2016. This strengthens our understanding of how climate variation affects this ecosystem. Modeling suggests that during 2026-2035 sea temperatures will reach about the same level as during the 2010-2019 period, and that there will be intense warming during the 2060-2069 period.
Natural up-and-down swings in the capelin population are central to the dynamics of the Barents Sea. The stock has undergone several significant fluctuations in recent decades, with the mature stock varying from less than 200,000 tonnes to up to three million tonnes. While the first of these declines had significant consequences throughout the ecosystem, the consequences have been less significant following subsequent declines. After being at a low level in 2016, there was a moderate upturn during 2017-2018, before again falling to a low level in 2019. Consequences of this recent stock decline are now under investigation. The snow crab population has spread westward since 2016 and is now well-established east of Svalbard. Most monitored seabird populations continue to decline, as observed for populations of black-legged kittiwake, common murre (common guillemot), and Atlantic puffin along the mainland coast, and for thick-billed murre (Brünnich's guillemot) throughout the management plan area.
Status and trends for various components
Water temperature in the Barents Sea is largely influenced by the amount of Atlantic water flowing into areas to the west; this inflow slowed during the 2016-2019 period, although temperatures remained considerably higher than during earlier cool periods. Levels of ocean acidification in the Barents Sea vary considerably; this is largely due to variation in water masses, biological production, and sea ice formation. Trends for pH and calcium carbonate saturation are weak, but generally declining; thus, indicating increasing ocean acidification.
Phytoplankton (primary) production has increased significantly during the 1998-2019 period. This is a result of climate change, where increasing temperatures and decreases in the extent of sea ice have opened new areas for high production and extended the growing season. The increase has been particularly pronounced in the northeastern part of the Barents Sea.
Medium-sized zooplankton provide direct or indirect nutrition to large parts of the ecosystem. Biomass of this species group has been relatively stable since 1995. As temperatures in the Barents Sea have warmed, several species of Arctic zooplankton have declined, while the abundance of more warmwater species has increased. Concurrent with decreased temperatures since 2016, an increased occurrence of pelagic amphipods, a distinctly Arctic species group, was observed in 2019.
Total biomass of large benthic species has increased in the Barents Sea over the past 14 years. The king crab population is considered to be stable east of the North Cape. With westward spread of the snow crab since 2016, it is believed that this species has colonized most of its potential habitat; but its occurrences in the west and north remains too small to be of commercial interest. The northern shrimp stock has remained stable at a relatively high level since 2005.
There are still large stocks of cod and haddock in the Barents Sea. Cod now occur in most areas, and in areas further north and northeast than previously observed. It is not clear the extent to which large stock size causes the population to expand northward, or the extent to which this is linked to the period of rising water temperatures.
Declining seabird populations have been observed over the past decade and throughout the entire monitoring period. Food availability, both during and outside the breeding season, is critically important, and likely the cause of negative outcomes for most seabird species. It is difficult to determine what factors contribute to decreased food availability. However, secondary effects of climate-related changes, lower production of prey and fishery removals, have been proposed.
Stocks of marine mammals in the Barents Sea are now affected by both increases related to conservation measures and the effects of climate change. Walrus is an example of a species that has increased significantly in abundance after decades of conservation. The ice-dependent ringed seal population appears to have been negatively affected by warming in the Barents Sea. The polar bear population is likely to be affected both positively through conservation measures and negatively by the effects of climate change. Over the past five years, there has been increased occurrence of several whale species along the coast of Svalbard; this has contributed to significantly increased knowledge of several ice-associated whale species. The summer occurrence of minke whales in the Barents Sea has increased as a result of changing environmental conditions. Stock conditions are good for grey seals and harbour seals along the mainland coast of the Barents Sea, unlike conditions farther south along the Norwegian coast.
Non-indigenous species in the Barents Sea include king crab and humpback salmon (pink salmon). The king crab population is so large that it can have significant effects on the ecosystem near the mainland coast. Several other non-indigenous species are poised to expand distribution into the Barents Sea and to the areas around Svalbard over the next 50 years.
Many of the most endangered marine species, for which Norway has special management responsibility, occur in the Lofoten-Barents Sea region. Of species considered threatened on the Norwegian Red List (published in 2015), 26 inhabit this region. On the 2018 Red List of Habitats, four habitat types in this marine area are considered to be severely or critically endangered.
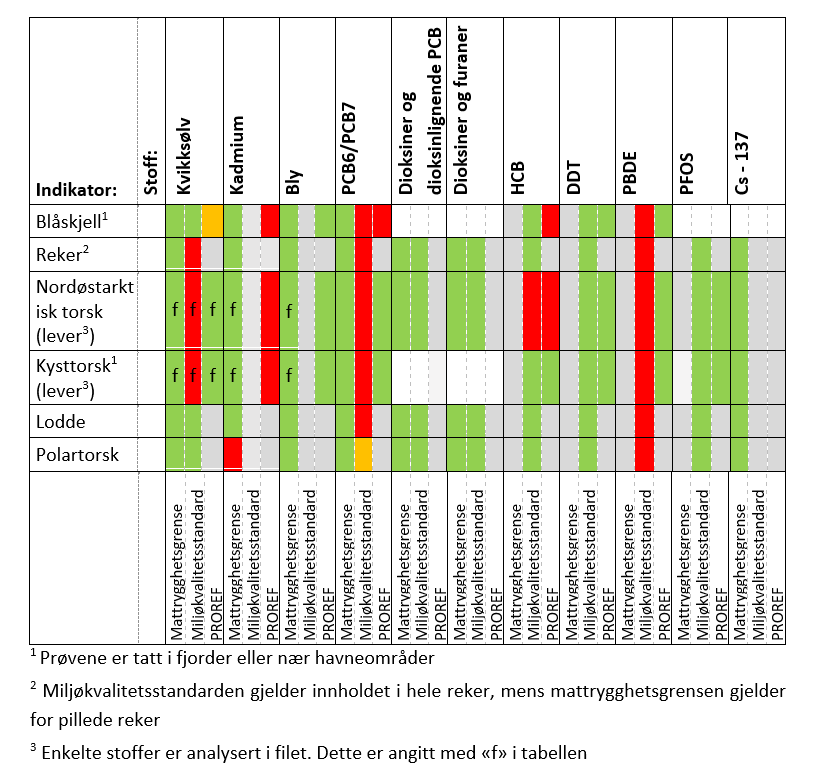
Contaminants and levels of pollutants are largely stable or decreasing. Measurements in sediments near Svalbard have shown relatively high levels of polyaromatic hydrocarbons, probably due to natural coal deposits. Levels of pollutants in fish, shrimp, and mussels are generally low, and well within the requirements for safe seafood. Most environmental toxins are also below levels believed to affect vulnerable parts of the ecosystem. Polar bears have high levels of fat-soluble pollutants which become concentrated at higher levels of the food chain. Levels of many pollutants in polar bears are declining in concert with declines in other components of the ecosystem, while levels of some novel contaminants are increasing. Environmental pollutants can have harmful effects on polar bears, especially in combination with decreased food availability.
In the Barents Sea, fisheries for stocks of cod, haddock, and deep-water redfish have been managed sustainably for many years. This coincides with an approximate 75% reduction in hours trawled by Norwegian fishermen from the end of the 1980s through 2009; hours trawled remains at about the same level currently.
Sammendrag
I rapporten dekkes tre hovedtemaer: (1) Dominerende trekk i status og utvikling i økosystemet i Barentshavet siden 2016 med vekt på å beskrive hvordan sentrale prosesser påvirker tilstanden; (2) en mer detaljert beskrivelse av status og utvikling for de ulike komponentene i økosystemet; og (3) en beskrivelse av fiskeriforvaltningen i Barentshavet.
Dominerende trekk i utvikling og sentrale prosesser
Det er betydelig naturlig variasjon i temperatur i Barentshavet som gjør at den kan øke eller avta mye i løpet av få år. Etter at vanntemperaturen generelt økte gjennom nærmere fire tiår, har den falt betydelig etter 2016 og var i 2019 på samme nivå som tidlig på 2000-tallet. Mens utbredelsen av havis i Barentshavet avtok betydelig i tiårene som vanntemperaturen økte, har utbredelsen av havis økt og iskanten trukket seg sørover fra 2016 til 2019. På samme måte som oppvarmingen fram mot 2016 hadde betydelige effekter på økosystemet i Barentshavet, i form av økt utbredelse av sørlige arter og nedgang i bestander av arktiske arter, er det observert økende mengder av visse arktiske dyreplankton- og fiskearter etter 2016. Dette styrker kunnskapen om hvordan variasjon i klima påvirker dette økosystemet. Modellering tyder på at vanntemperaturene i 2026-2035 vil komme ut på omtrent samme nivå som perioden 2010-2019 og at det vil komme en kraftig oppvarming i perioden 2060-2069.
Loddas naturlige opp- og nedganger er sentral for dynamikken i Barentshavet. Bestanden har gjennomgått flere betydelige svingninger de siste tiårene, der den modne bestanden har variert fra under 200 000 tonn til opp mot tre millioner tonn. Mens den første av nedgangene fikk betydelige konsekvenser for økosystemet, har følgene vært mindre for senere nedganger. Etter å ha vært på et lavt nivå i 2016 hadde den en moderat oppgang i 2017 og 2018 før den igjen falt til et lavt nivå i 2019. Det forskes på hvilke konsekvenser den siste bestandsnedgangen vil kunne få. Snøkrabbe har spredd seg vestover siden 2016 og er nå vel etablert på østsiden av Svalbard. Nedgangene i de fleste overvåkede sjøfuglbestandene fortsetter, og observeres for krykkje, lomvi og lunde langs fastlandskysten og for polarlomvi i hele forvaltningsplanområdet.
Status og utvikling for de ulike komponentene
Vanntemperaturen i Barentshavet påvirkes i stor grad av hvor mye atlanterhavsvann som strømmer inn i området i vest, og denne innstrømmingen avtok fra 2016 til 2019. Temperaturen er fortsatt betydelig høyere enn under tidligere kjølige perioder. Det er store variasjoner i havforsuring i Barentshavet, som i stor grad skyldes variasjoner i vannmasser, biologisk produksjon og isdannelse. Trendene for pH og kalkmetning i Barentshavet er svake, men viser generelt en nedgang, i retning av økt havforsuring.
Primærproduksjon i planteplanktonet har økt betydelig i perioden 1998-2019. Årsaken er endringer i klima, der økende temperatur og minkende utbredelse av havis har åpnet nye områder for høy produksjon og forlenget vekstsesongen. Økningen har vært særlig utpreget i den nordøstlige delen av Barentshavet.
Det mellomstore dyreplanktonet gir direkte eller indirekte næring til store deler av økosystemet. Biomassen av dette planktonet har vært relativt stabil siden 1995. Under oppvarmingen av Barentshavet har flere arter av arktisk dyreplankton gått tilbake mens mengden av mer varmekjære arter har økt. Etter at temperaturen hadde gått ned siden 2016, ble det i 2019 observert økte mengder av pelagiske amfipoder, en utpreget arktisk gruppe.
Den samlede biomassen av store bunndyrarter har økt i Barentshavet de siste 14 årene. Bestanden av kongekrabbe er vurdert å være stabil øst for Nordkapp. Med spredningen vestover, siden 2016, antas det at snøkrabben har kolonisert det meste av sitt potensielle leveområde, men forekomstene i vest og nord er fortsatt for små til å være kommersielt interessante. Bestanden av reke har vært stabil på et relativt høyt nivå siden 2005.
Det er fremdeles store bestander av torsk og hyse i Barentshavet. Torsken dekker nå store deler av Barentshavet, også lenger mot nord og nordøst enn tidligere. Det er ikke klart om det er den store bestanden som gjør at den har trukket nordover, eller om dette også er koblet til perioden med økende temperatur i vannet.
Nedgangen i de fleste overvåkede sjøfuglbestandene ses både for de siste ti årene og når en ser samlet på hele tidsperioden de har vært overvåket. Mattilgang i og utenfor hekketiden er viktig, og det er sannsynlig at de negative endringene for de fleste sjøfuglartene skyldes redusert næringstilgang. Det er imidlertid vanskelig å konkludere med hva endringene i næringstilgangen skyldes, men sekundæreffekter av klimarelaterte endringer, lavere produksjon av byttedyr eller uttak av fiskeressurser, har vært foreslått.
Bestander av sjøpattedyr i Barentshavet påvirkes nå både av vekst etter fredning og klimaendringer. Hvalross er eksempel på en art som har økt betydelig i antall etter flere tiår med fredning. Den istilknyttede arten ringsel ser ut til å ha vært negativt påvirket av oppvarmingen i Barentshavet, mens bestanden av isbjørn sannsynligvis er påvirket av både fredning og klimaendringer. Det er økte forekomster av flere hvalarter langs kysten av Svalbard og en betydelig økning i kunnskap om flere istilknyttede hvalarter i løpet av de siste fem årene. Sommerforekomsten av vågehval i Barentshavet har økt som følge av endrede miljøforhold. Bestandssituasjonen er god for havert og steinkobbe langs fastlandskysten av Barentshavet, til forskjell fra lenger sør langs norskekysten.
Fremmede arter i Barentshavet inkluderer kongekrabbe og pukkellaks. Bestanden av kongekrabbe er så stor at den kan ha betydelige effekter på økosystemet nær fastlandskysten. Flere fremmede arter er vurdert å kunne spre seg inn i Barentshavet og til områdene rundt Svalbard de neste 50 årene.
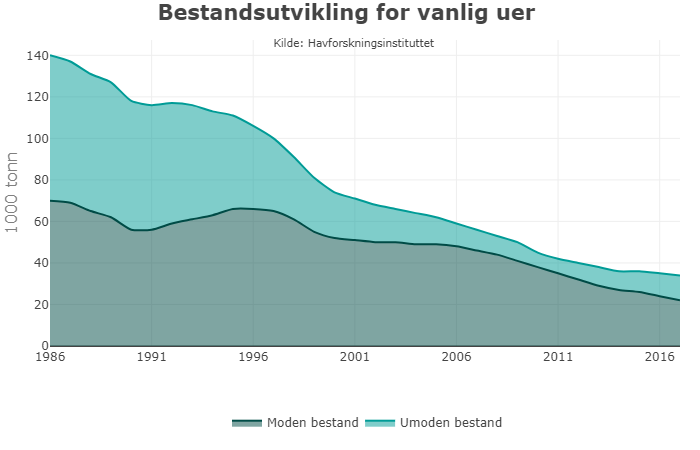
Mange av de mest truede marine artene som Norge har et særskilt forvaltningsansvar for finnes i havområdet Lofoten-Barentshavet. Av artene som er vurdert som truet i den sist publiserte norske rødlista fra 2015, har 26 sitt leveområde her. Halvparten av artene er i kategoriene kritisk og sterkt truet, som for eksempel lomvi langs fastlandskysten og vanlig uer. Fire av naturtypene i havområdet er vurdert som sterkt eller kritisk truet på rødlista over naturtyper fra 2018.
Tilførsler og nivåer av forurensende stoffer er stort sett stabile eller nedadgående. Målinger i sedimenter har vist forholdsvis høye nivåer av polyaromatiske hydrokarboner nær Svalbard som trolig skyldes naturlige kullforekomster. Nivåene av forurensede stoffer i fisk, reker og blåskjell er stort sett lave og godt innenfor krav til trygg sjømat. For de fleste miljøgiftene er de også under nivåene som antas å kunne føre til effekter på de mest sårbare delene av økosystemet. Isbjørn har høye nivåer av fettløselige miljøgifter som oppkonsentreres i næringskjeden. Nivåene av mange miljøgifter i isbjørn avtar imidlertid i takt med nedgang i økosystemet ellers, mens nivåene av noen «nye» stoffer øker. Miljøgiftnivåene kan ha skadelige effekter på isbjørn, særlig i kombinasjon med sult.
Bestandene av torsk, hyse og snabeluer i Barentshavet fiskes bærekraftig og har vært det i mange år. Etter at antall tråltimer med norske trålere gikk ned med rundt 75 % fra slutten av 1980-tallet til 2009, har det etter det ligget på omtrent samme nivå.
Rapporten kan siteres slik:
Arneberg, P, van der Meeren, G.I., Frantzen, S. og Vee, I. (red.) (2020). Status for miljøet i Barentshavet – rapport fra Overvåkingsgruppen 2020. Fisken og Havet nr. 2020-13, Havforskningsinstituttet.
1 - Innledning
Som en del av oppfølgingen av de tre helhetlige forvaltningsplanene for norske havområder, skal Overvåkingsgruppen, hvert tredje år, rapportere om status i miljøet i havområdene Barentshavet, Norskehavet og Nordsjøen/Skagerrak. Rapportene alternerer mellom de ulike områdene, slik at det i 2018 ble rapportert for Nordsjøen og Skagerrak, i 2019 for Norskehavet, mens turen i år er kommet til Barentshavet.
Formålet med rapporten er å beskrive hovedtrekk i status og vesentlige utviklingstrekk for miljøet i Barentshavet de senere årene. Forrige rapport fra Overvåkingsgruppen for Barentshavet var i stor grad basert på data frem til og med 2016. I rapporten er det derfor vurdert hvordan tilstanden i Barentshavet har endret seg fra 2016.
En helhetlig vurdering av tilstanden for miljøet i Barentshavet er gitt i kapittel 2. Her er de viktigste trekkene i status for miljøet og de viktigste endringene siden 2016 beskrevet. Det er også drøftet hva som er årsaken til status og endringer. Dette er basert på Overvåkingsgruppens indikatorer for området, publisert forskningslitteratur og ulike rapporter, blant annet flere rapporter fra det Internasjonale havforskningsrådet (ICES). En mer detaljert beskrivelse av status og utvikling for de ulike komponentene i økosystemet, samt for en del av den menneskelige påvirkningen er gitt i kapittel 4.
I Overvåkingsgruppens miljøstatusrapporter bruker det å være et kapittel med en utdypende beskrivelse av ett eller flere utvalgte tema. I år handler dette om fiskeriforvaltning i Barentshavet. Dette er gitt i kapittel 3.
Kapittel 5 dreier seg om særlig verdifulle og sårbare områder. Her er det gitt en kort oppsummering for elementer som ligger til grunn for identifisering av slike områder og som Overvåkingsgruppen har indikatorer for.
Også i årets rapport er det vurdert om referanse- og tiltaksgrenser er overskredet for de indikatorene Overvåkingsgruppen har for området og hvor slike verdier er fastsatt. Dette er beskrevet i kapittel 6. Overvåkingsgruppens indikatorer publiseres elektronisk og oppdateres løpende på Miljøstatus i Norge sine nettsider (www.miljostatus.no). I kapittel 6 er det gitt en oversikt over alle indikatorer som inngår i grunnlaget for denne rapporten. Her er det også gitt lenker til de fullstendige rapporteringene på nettsidene til Miljøstatus i Norge.
En rekke institusjoner har bidratt til denne rapporten, enten ved å levere data til indikatorene eller på andre måter til utforming av teksten. På omslagssiden er det angitt hvilke institusjoner dette dreier seg om.
2 - Dominerende trekk i utviklingen av økosystemet
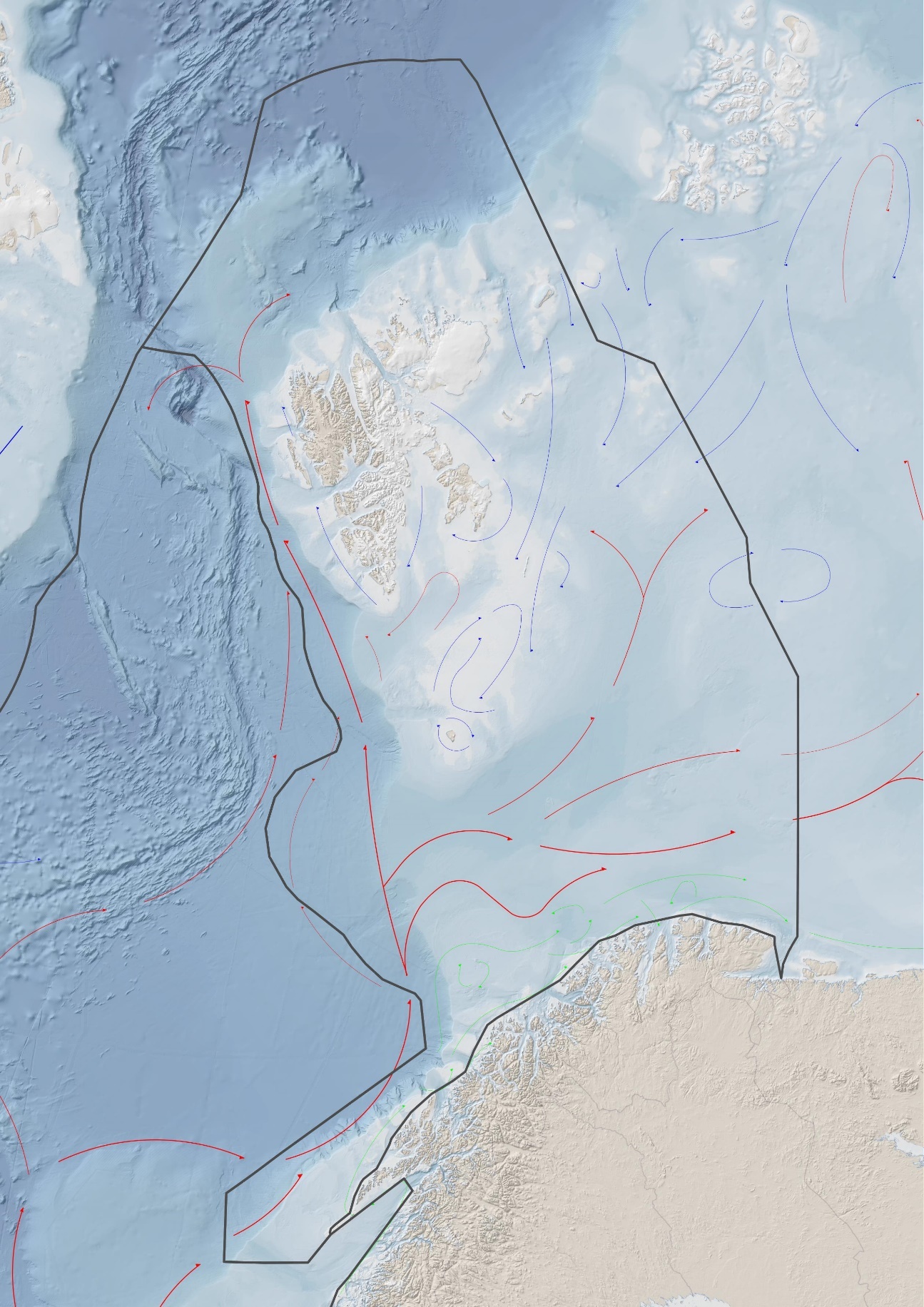
Helhetlig tilstandsvurdering Dette kapittelet beskriver dominerende trekk i miljøtilstand i Barentshavet og utviklingen siden 2016, som datagrunnlaget var hentet fra da Overvåkingsgruppen sist rapporterte om havområdet (Arneberg og Jelmert 2017). Det er lagt vekt på å beskrive hvordan sentrale prosesser påvirker tilstanden. Viktig ny kunnskap om årsaker til status og endringer er beskrevet. Det er gitt en kort omtale av forventet utvikling de neste to årene for noen elementer samt en kort omtale av det nye fagsystemet for vurdering av økologisk tilstand, som er blitt testet for den arktiske delen av Barentshavet. Fordi status og utvikling best kan beskrives og forstås ved å se helhetlig på havområdet, omfatter vurderingen i dette kapittelet hele Barentshavet. Barentshavet avgrenses geografisk mot Norskehavet i vest, dvs. øst for en linje trukket fra Nordkapp over Bjørnøya til Sørkapp på Spitsbergen, og øst mot Frans Josefs Land i nordøst og Novaja Semlja i øst. I forvaltningsplanarbeidet er kystområdene utenfor Lofoten, Vesterålen, Troms og Vest-Finnmark også inkludert i Barentshavet, så begrepet «Barentshavet» i denne sammenheng omfatter også disse kystområdene (figur 2.1).
Figur 2.1. Forvaltningsplanområdet for Barentshavet og havområdene utenfor Lofoten. I kartet er det angitt hovedstrømmer av atlantisk vann (rødt) og arktisk vann (blått). Kilde: Barentswatch/Arealverktøyet.
En dominerende årsak til forandringer i økosystemet i Barentshavet er endringer i klima, som påvirker en rekke komponenter i systemet. Området er også preget av naturlig dynamikk. Det har gitt seg utslag i at loddebestanden, en sentral komponent i økosystemet, nok en gang har kollapset i løpet av de siste årene. Andre viktige utviklingstrekk er fortsatt spredning av snøkrabbe og fortsatt nedgang i sjøfuglbestandene.
2.1 - Endringer i klima og effekter på økosystemet
Det er betydelig naturlig variasjon i vanntemperatur i Barentshavet. Denne variasjonen kan spille seg ut over korte tidsrom og gjøre at temperatur kan gå mye opp eller ned i løpet av få år. Den menneskeskapte endringen i klima kan registreres først når man ser over lange tidsrom, typisk flere tiår. Vanntemperaturen har vært økende de siste 40 årene med markante variasjoner (figur 2.2, samme som figur 4.1). Mens slutten av 1970-tallet var en kald periode, var det relativt varmere i 1930- og 1950-årene, men ikke så varmt som etter 2006. Etter betydelig temperaturøkning fra 1990 var tiåret mellom 2006 og 2016 spesielt varmt med 2016 som det varmeste året. Etter 2016 har temperaturen gått ned og var i 2019 på samme nivå som tidlig på 2000-tallet, men fremdeles over langtidsmiddelet. Vanntemperaturen i Barentshavet er i stor grad påvirket av atlanterhavsvannet som strømmer inn fra Norskehavet, og den siste temperaturnedgangen har sammenheng med lavere temperaturer i atlanterhavsvannet. Det er mindre variasjon i temperaturen i det arktiske vannlaget, som vanligvis ligger mellom 0 og -1,9 grader, men etter 2005 har denne temperaturen økt og i noen tilfeller vært over null grader på slutten av sommeren (Lind et al. 2018). Utbredelsen av havis varierer gjennom året og har minket betydelig om vinteren de siste tiårene. Utbredelsen av havis har økt noe etter 2016, trolig på grunn av noe kjøligere vann og økende tilførsel av sjøis fra Polhavet.
Figur 2.2. Temperatur i kjernen av atlanterhavsvannet som strømmer inn i Barentshavet fra sørvest i snittene Fugløya-Bjørnøya og Vardø-Nord fra 1950 og frem til og med 2019. Svarte linjer viser årlige verdier (på sensommeren), tykke linjer viser tre års løpende gjennomsnitt og stiplede svarte linjer viser langtidsmiddelet (1981-2010).
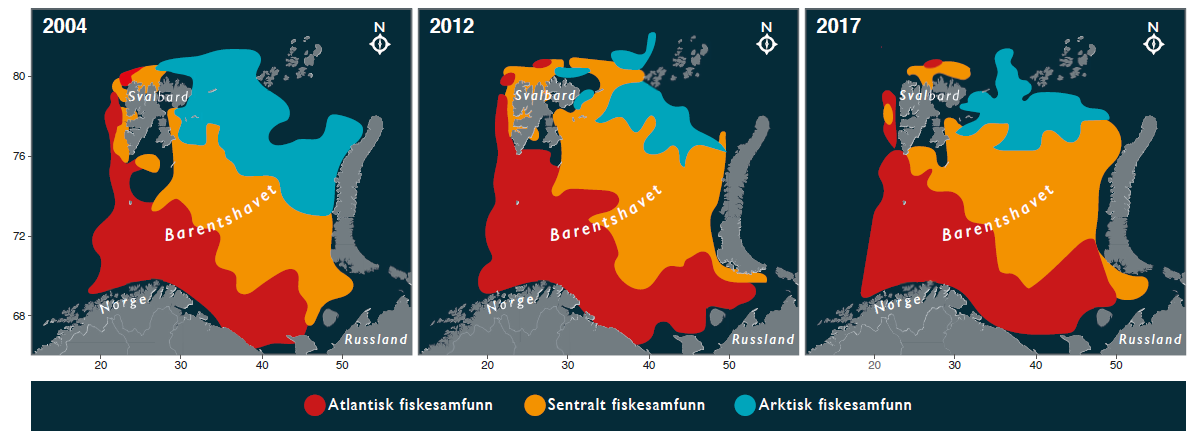
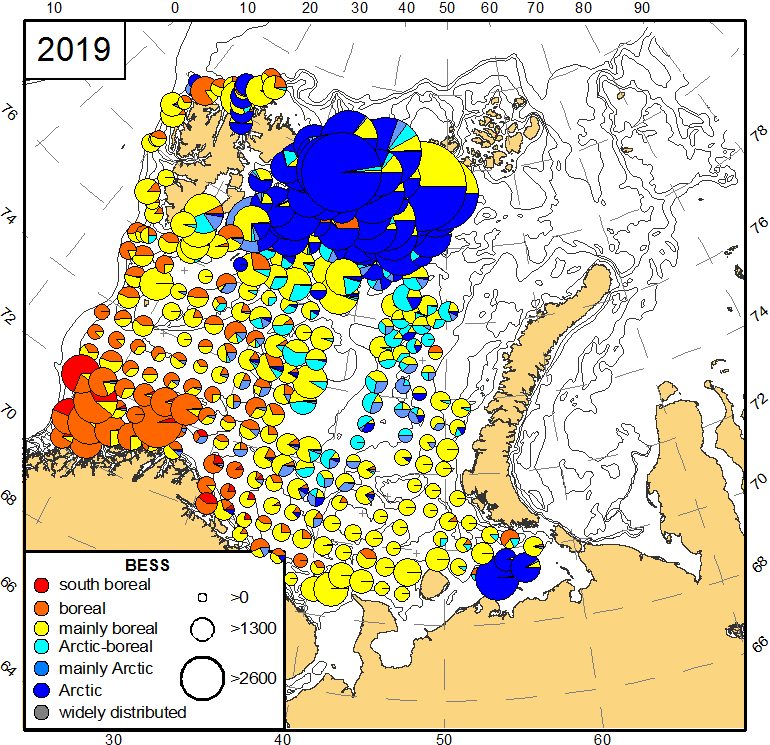
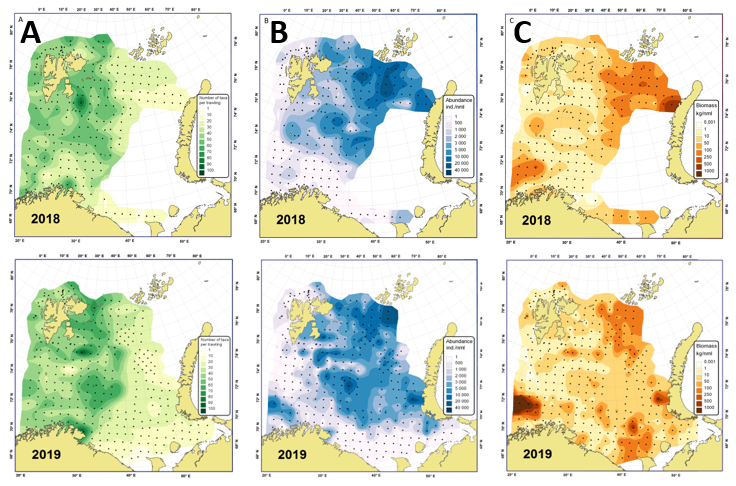
ICES-gruppen WGIBAR (Working Group on the Integrated Assessments of the Barents Sea) har under sitt årlige møte i februar 2020 vurdert om de siste årenes nedkjøling har reversert noen av de endringene oppvarmingen forårsaket inntil 2016. I Overvåkingsgruppens forrige statusrapport for Barentshavet ble det beskrevet hvordan oppvarmingen fra begynnelsen av 2000-tallet har ført til at en rekke arter har flyttet sin utbredelse mot nord og øst. Dette har vært særlig godt dokumentert for fisk, og som har betydd at arktiske fiskearter har fått utbredelsen begrenset til de nordligste delene av Barentshavet (Fossheim et al. 2015) (figur 2.3). For 2019 rapporterer WGIBAR tegn på reversering av dette. Fangstene av ikke-kommersielle fiskearter som er klassifisert som arktiske eller for det meste arktiske var i 2019 de høyeste siden 2016 (figur 2.4) (ICES 2020).
Figur 2.3. Endring fra 2004 til 2017 i utbredelse av atlantiske, sentrale og arktiske fiskesamfunn i Barentshavet. Basert på oppdatert analyse etter Fossheim et al. (2015).
Figur 2.4. Utbredelse av ikke-kommersielle fiskearter fra økotokt i august-september i 2017 og 2019. Størrelsen på sirkler viser størrelsen på fangster (antall individer per nautisk mil). Med blå farger markert arktiske fiskearter (ICES 2020) .
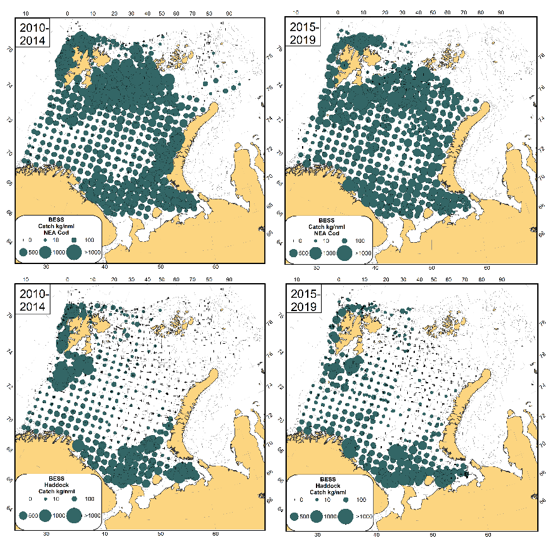
Torsk og hyse spredte seg i flere år nordover parallelt med at bestandsstørrelser og temperatur har økt. Bestandene var på sine høyeste nivå i 2013 for torsk og 2010 for hyse. Begge bestandene spredte seg i årene før dette inn i områder med nye matressurser som var blitt isfrie nord i Barentshavet på grunn av oppvarmingen. Parallelt med at Barentshavet har blitt kjøligere og bestandene mindre, har de begge artene trukket seg sørover de senere årene (figur 2.5) (ICES 2020). Dette forventes å ha betydning også for andre deler av økosystemet. For eksempel har et nytt studium vist at fiskesamfunnene i ulike områder blir likere når torskebestanden øker, antagelig som en konsekvens av at torsken beiter omfattende og jevnt på mange matressurser over større områder (Ellingsen et al. 2020).
Figur 2.5. Utbredelse av torsk (oppe) og hyse (nede) fra økotokt i august-september for to perioder 2010-2014 og 2015-2019. Størrelsen på sirkler indikerer størrelsen på fangster (kg per nautisk mil). Figuren er tatt fra ICES (2020).
Mengden amfipoder i frie vannmasser har tidligere minket som en følge av oppvarmingen, og også for denne gruppen har det vært en økning de siste årene, med dobbelt så høye mengder estimert i 2019 som under de varmeste årene (2015-2016). Det er fortsatt høye nivåer av krill (ICES 2020).
Mengden havis er knyttet til både luft og havtemperatur, og parallelt med nedgangen i temperatur har utbredelsen av havis økt i Barentshavet (se kapittel 4.1.3). Polartorsk er en utpreget arktisk art og et viktig byttedyr for en rekke arter i den arktiske delen av Barentshavet. Polartorsk gyter under isen, og isen er viktig for overlevelse av eggene. Eggene har en tynn hinne og er avhengige av gode isforhold som verner mot slitasje fra bølger og virvler i sjøen. Temperaturen er også mer stabil under isen, som også bidrar til økt overlevelse (Eriksen et al. 2019; Eriksen et al. 2015; Huserbråten et al. 2019). Mens mengden under ett år gammel polartorsk i mange år har ligget på et svært lavt nivå, var det i 2019 for første gang på flere år en betydelig økning opp fra dette nivået. De lave forekomstene i varme år faller sammen med at isdekket over hovedgytefeltene sørøst i Barentshavet var redusert og variabelt disse årene. Gytefeltene ved Svalbard har vært mer stabilt i de varme årene, men er mindre viktige for rekrutteringen.
Økningen i utbredelse av havis har som beskrevet i kapittel 4.1.3 også ført til at iskanten i 2019 har hatt en sørligere posisjon enn i det varmeste året (2016).
Det at de siste årenes avkjøling har reversert noen av effektene av oppvarmingen, gjør at vi nå får enda bedre kunnskap om hvordan naturlig variasjon i klima påvirker økosystemet. Selv om mye forskningsarbeid gjenstår, tegner det seg et bilde av sterke effekter av endringer i klima på geografisk utbredelse og mengdeforhold av arter, der arktiske arter trekker seg nordover og minker under oppvarminger og det motsatte under nedkjølinger. For atlantiske (sørlige) arter er bildet motsatt. Hvordan økosystemet vil utvikle seg i fremtiden vil derfor påvirkes betydelig av utvikling i klimaet i Barentshavet. Resultater fra modellkjøringer presentert i kapittel 4.1.6 tyder på at det vil bli en avkjøling i perioden 2026-2035, som vil føre til at havtemperaturene kommer til omtrent samme nivå som i 2010-2019, det vil si betydelig høyere enn de kalde periodene i forrige århundre. Deretter kommer det en kraftig oppvarming i perioden 2060-2069.
De antatte utslagene vil være størst i østlige og nordlige deler av Barentshavet. Om klimaet utvikler seg slik, vil økosystemet variere innenfor kjente klimarammer de nærmeste årene. Etter dette må vi forvente at økosystemet beveger seg ut i ukjent klimaterreng, der pilotvurderingen fra fagsystem for økologisk tilstand for arktisk del av Barentshavet (Jepsen et al. 2019) gir en antydning av retning og omfang av forventede økologiske endringer.
2.2 - Naturlig dynamikk
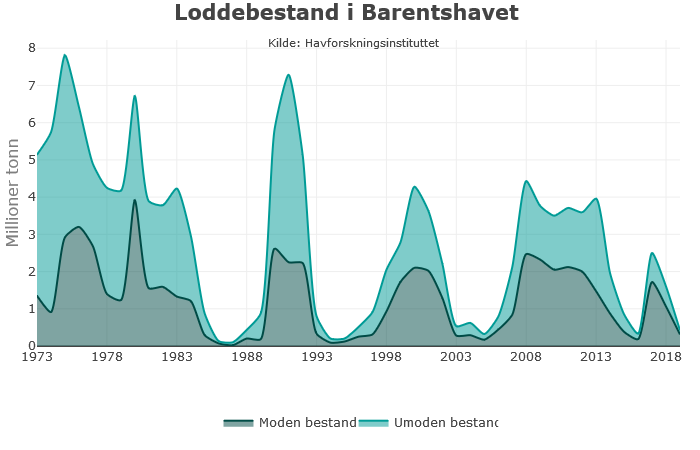
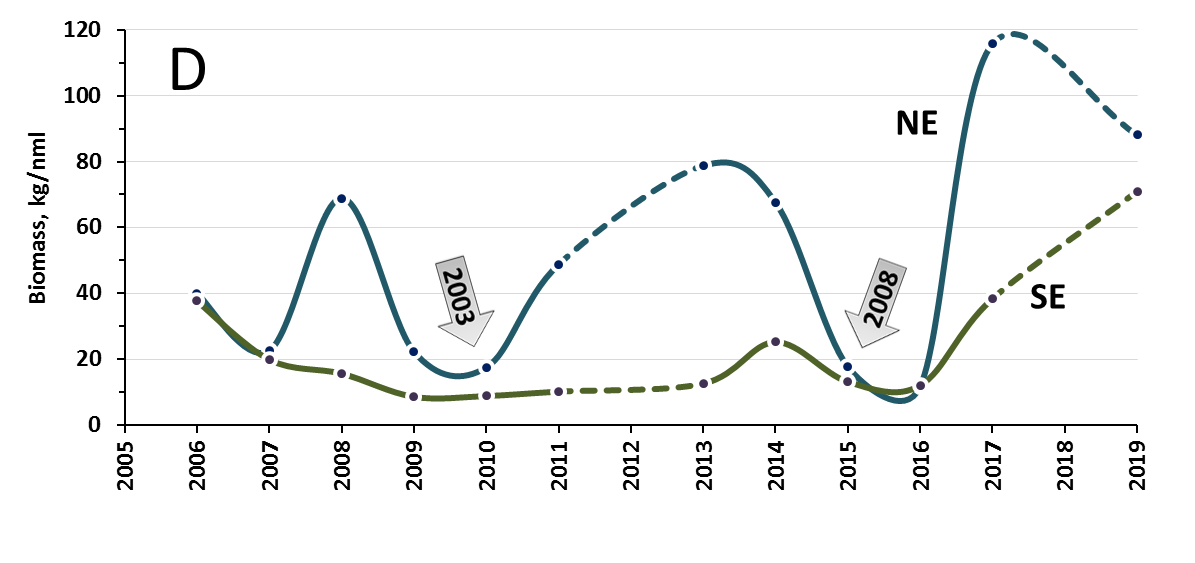
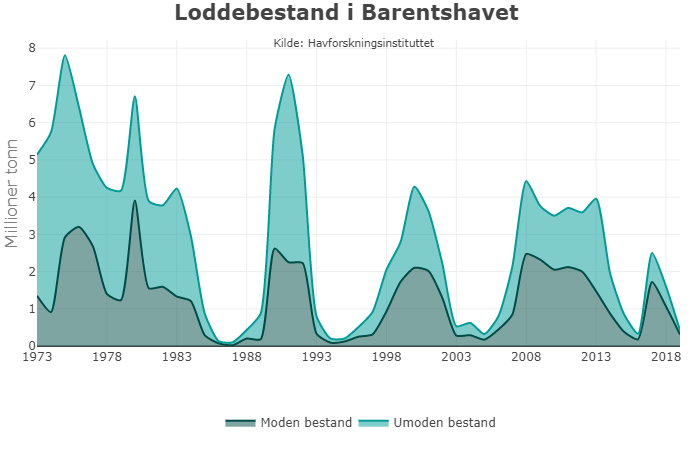
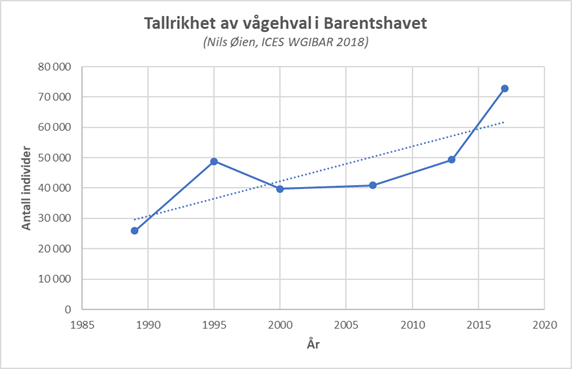
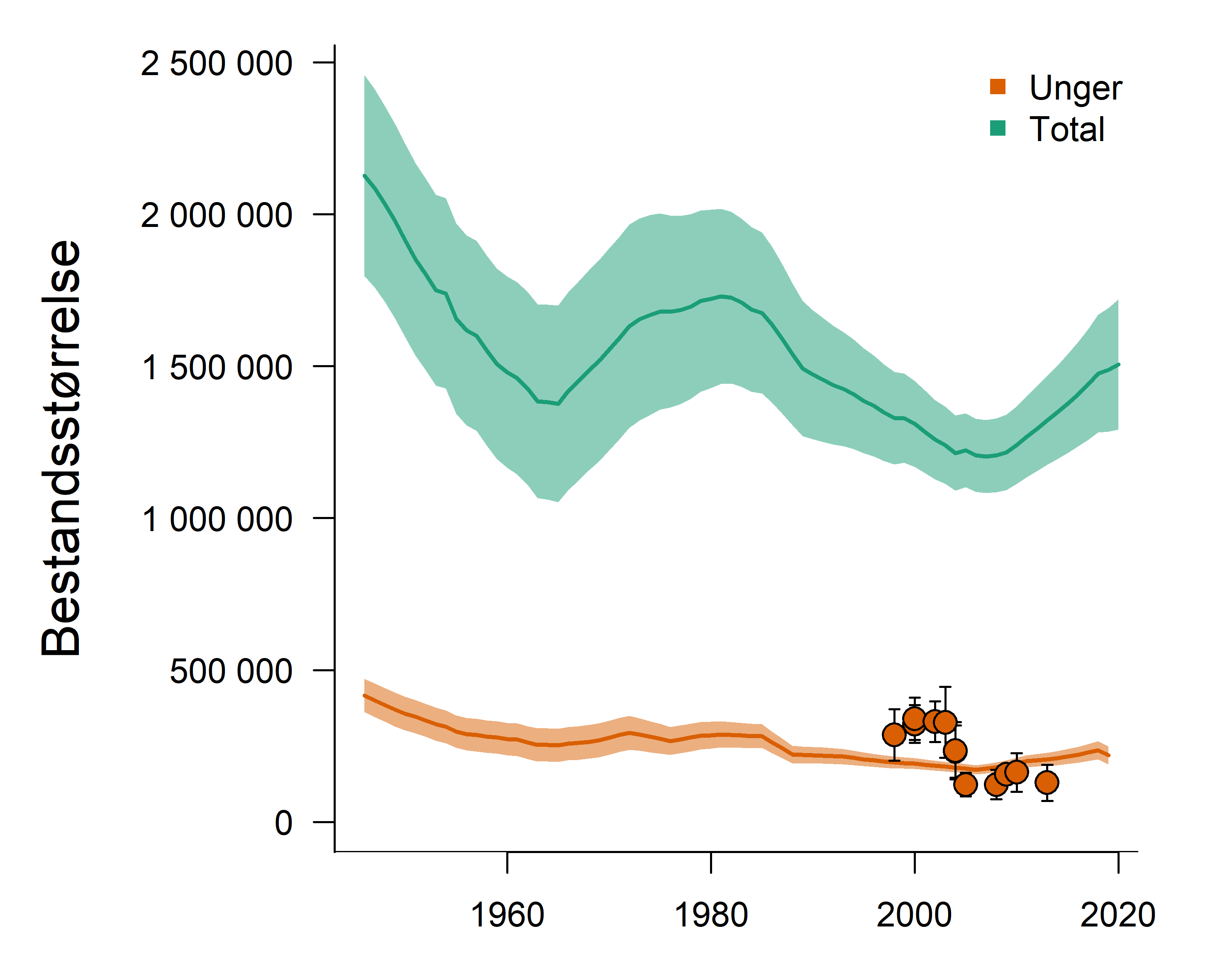
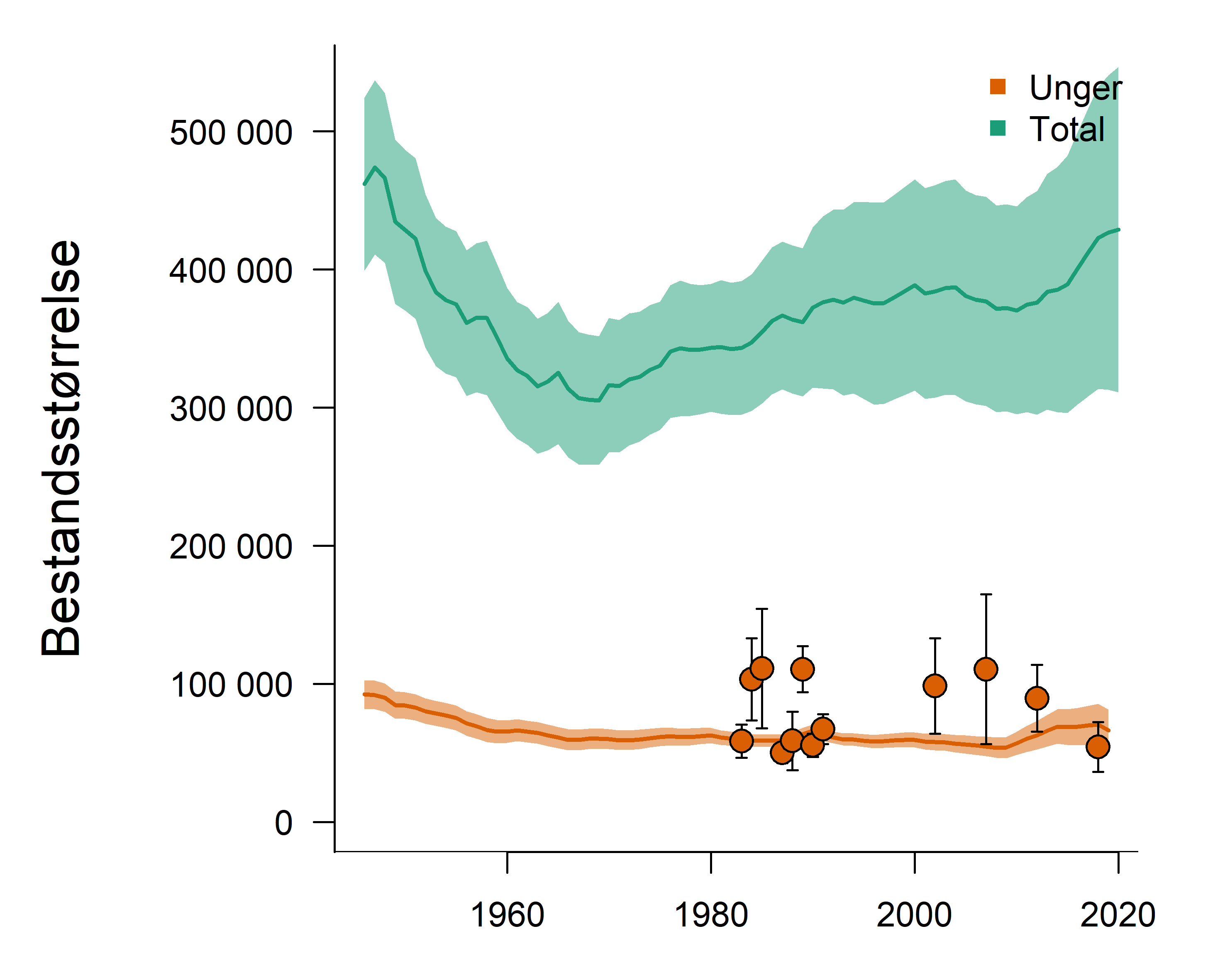
Marine økosystemer har stor naturlig variabilitet over tid. Variasjon i rekruttering og overlevelse hos sentrale arter vil bidra til det. Lodde er en viktig art i økosystemet i Barentshavet og er derfor en sentral indikatorart for Barentshavet. Den er viktig næring for blant annet nordøstarktisk torsk og flere arter av sjøfugl og sjøpattedyr. Størrelsen på loddebestanden er samtidig svært variabel. Siden systematiske målinger av bestanden startet i 1973, har bestandsstørrelsen falt til lave nivåer fem ganger, i 1987, 1995, 2005, 2016 og igjen i 2019. Årsaken til de betydelige bestandsnedgangene er sammensatte, men svikt i rekruttering på grunn av økt predasjon på loddelarver spiller en avgjørende rolle i de tre første tilfellene. I første rekke ser ungsild ut til å være den viktigste predatoren i dette bildet, men 0-gruppe hyse og torsk kan også ha spilt en rolle (Gjøsæter et al. 2015; Gjøsæter et al. 2016). I de to siste bestandsnedgangene har overlevelsen av loddelarver vært relativt god mens dødeligheten for ett- og toåringer har vært forhøyet. En mulig årsak til dette er økt predasjon fra fisk, sjøfugl og sjøpattedyr. Når det gjelder sjøpattedyr, har det i de senere årene vært observert flere arter samt økende antall av vågehval i Barentshavet om sommeren (ICES 2020).
Mens den første betydelige bestandsnedgangen (i 1987) fikk omfattende konsekvenser i resten av økosystemet, inkludert økt dødelighet blant sel, sjøfugl og torsk (Hjermann et al. 2004), hadde de to neste nedgangene (1995 og 2005) langt mindre effekter på økosystemet (Johannesen et al. 2012). Årsaken til dette er antagelig at det var økt tilgang til alternative byttedyr under den andre og tredje kollapsen (Gjøsæter et al. 2009). Det gjenstår å se hvilke konsekvenser den siste bestandsnedgangen vil få, noe som vil følges opp gjennom tokt, forskning og rapportering fra WGIBAR og Overvåkingsgruppen.
Figur 2.6. Utvikling i estimert biomasse av lodde i Barentshavet.
2.3 - Snøkrabbe som sprer seg og sjøfuglbestander som minker
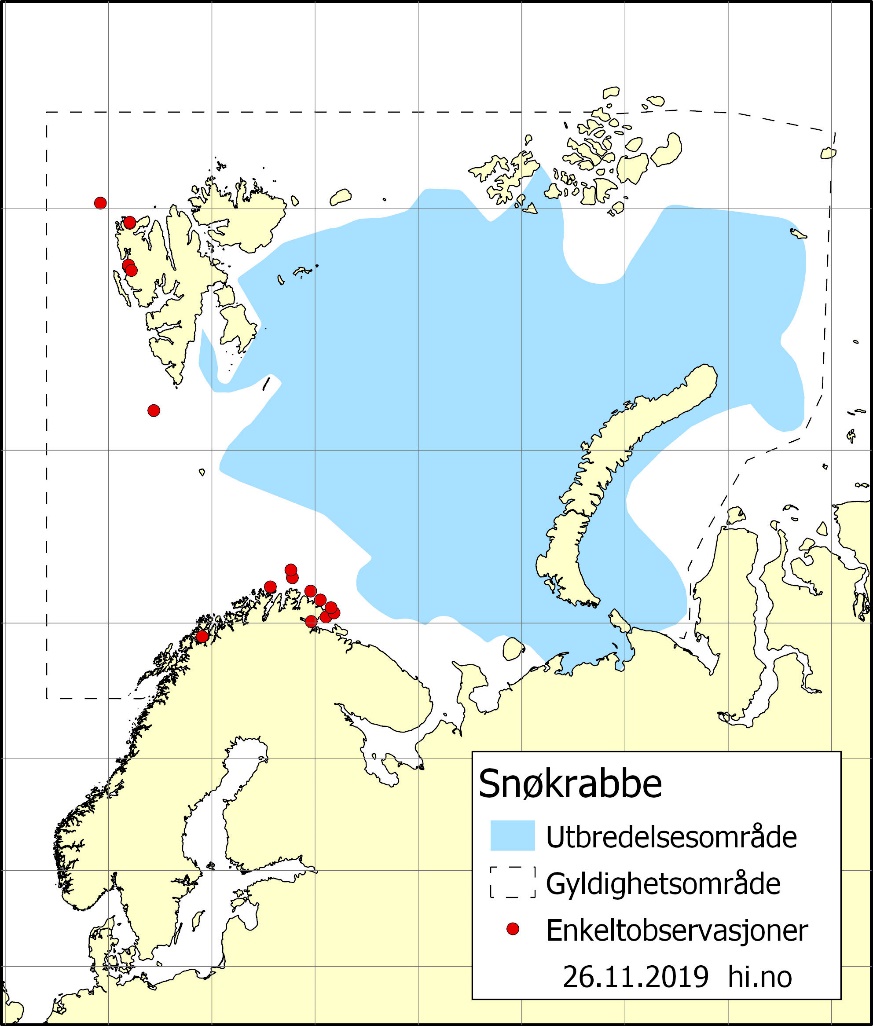
Siden forrige rapport fra Overvåkingsgruppen, har snøkrabben spredd seg vestover til farvannene rundt Svalbard. Arten er vel etablert på østsiden av øygruppen og det er gjort observasjoner av enkeltindivider også på vestkysten av Spitsbergen (figur 2.7). Det er uklart hvordan bestanden vil kunne vokse på vestsiden av Spitsbergen, fordi snøkrabbe her kan påvirkes negativt av store mengder torsk (et viktig rovdyr) og høyere vanntemperaturer.
Figur 2.7. Estimert utbredelsesområde av snøkrabbe i Barentshavet og vestlige deler av Karahavet. Den stiplede linjen viser det undersøkte området. Enkeltobservasjoner utenfor utbredelsesområdet er vist med røde prikker.
De fleste sjøfuglindikatorene viser en nedgang i hekkebestandene, både i de siste ti årene og samlet over tidsperioden de har vært overvåket. Dette gjelder både krykkje, lomvi og lunde langs fastlandskysten samt polarlomvi i hele området. Mattilgang i og utenfor hekketida er viktig, og det er sannsynlig at de negative endringene for de fleste sjøfuglartene skyldes redusert næringstilgang. Det er imidlertid vanskelig å konkludere med hva endringene i næringstilgangen skyldes, men sekundæreffekter av klimarelaterte endringer, lavere produksjon av byttedyr eller uttak av fiskeressurser har vært foreslått.
2.4 - Forventet utvikling i 2020 og 2021
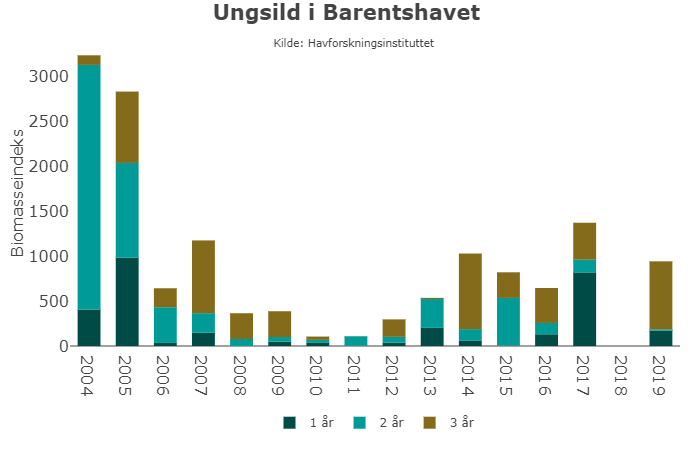
Det forventes at bestanden av torsk vil minke noe fordi eldre fisk går ut av bestanden mens kun en ny sterk årsklasse er observert de siste årene (2017). Hysebestanden er forventet å øke på grunn av en sterk 2016-årsklasse og en litt over middels 2017-årsklasse. Mengden ungsild (1-4 år gammel) var relativt stor i 2019, men vil minke i 2020 fordi den sterke 2016-årsklassen vil vandre ut av Barentshavet til Norskehavet. Bestanden av polartorsk forventes å fortsette å minke fordi det ikke har vært observert noen sterke årsklasser de siste årene. Loddebestanden kan vokse dersom overlevelsen av den sterke 2019-årsklassen blir god.
2.5 - Fagsystem for vurdering av økologisk tilstand
Siden forrige statusrapportering har fagsystemet for vurdering av økologisk tilstand blitt utviklet (Nybø og Evju 2017) og prøvd ut for den arktiske delen av Barentshavet (Jepsen et al. 2019). I systemet blir økologisk tilstand vurdert for syv følgende økosystemegenskaper:
Primærproduksjon
Fordeling av biomasse mellom ulike trofiske nivåer
Funksjonell sammensetning innen trofiske nivåer
Funksjonelt viktige arter, habitatbyggende arter og biofysiske strukturer
Landskapsøkologiske mønstre
Biologisk mangfold representert ved økosystemenes genetiske mangfold, artssammensetning og artsutskifting
Abiotiske forhold
Avvik fra god økologisk tilstand er definert som å inntreffe når økosystemet er blitt betydelig påvirket av menneskelige aktiviteter (inkludert klimaendringer). Systemet er basert på at det først etableres indikatorer for alle de syv økosystemegenskapene. For alle indikatorene vurderes det så i hvilken grad de viser endringer som kan knyttes til menneskeskapt påvirkning. Deretter gjøres det en samlet vurdering for alle indikatorene innen en gitt økosystemegenskap før det gjøres en kvalitativ vurdering for økosystemet som helhet.
Fra utprøvingen for den arktiske delen av Barentshavet er det konkludert med at det ikke er god økologisk tilstand fordi klimaet gjennom flere tiår har blitt varmere, og at flere av økosystemegenskapene viser endringer som kan knyttes til oppvarmingen (Jepsen et al. 2019). Resultatene fra denne pilotvurderingen må anses som foreløpige. En endelig vurdering vil foreligge i løpet av 2020.
3 - Spesialtema: Norsk fiskeriforvaltning i Barentshavet
3.1 - Innledning
Barentshavet, sammen med kyst og havområdene fra Lofoten og nordover, er i biologisk forstand et av verdens mest produktive havområder, med gyte- og oppvekstområdene for en rekke viktige fiskeslag i Nordøst Atlanteren. Denne produktiviteten gir grunnlag for store bestander av fisk, sjøpattedyr og sjøfugl. Sommerstid har minst 20 millioner sjøfugl, fordelt på 40 ulike arter og 1600 hekkekolonier tilhold i havområdet. Barentshavet er det viktigste området for norsk fiskerinæring når det gjelder høsting, og siden 2015 har samlet fangst ligget i overkant av 900 000 tonn, om lag en tredjedel av de totale norske landinger av fisk, mens fangstverdien fra området i de samme årene utgjorde vel 50 % av den samlede verdien fra de norske fiskeriene.
Den geografiske avgrensningen av Barentshavet er beskrevet i kapittel 2. Store deler av havbunnen i området er kontinentalsokkel, og havområdet regnes for å være et subarktisk hav da det ligger omtrent på grensen mellom Arktis og temperert sone. Nordgående havstrømmer transporterer atlantisk vann øst- og nordover, mens sørgående havstrømmer transporterer arktisk vann sørover. Tilførselen av atlantisk vann gjør deler av området spesielt sammenlignet med andre arktiske områder på tilsvarende breddegrad. Denne tilførselen bidrar også til at området er relativt lett tilgjengelig for fiske i store deler av året.
3.2 - Forvaltning av fiskeressursene
Barentshavet er delt inn i følgende fire soner:
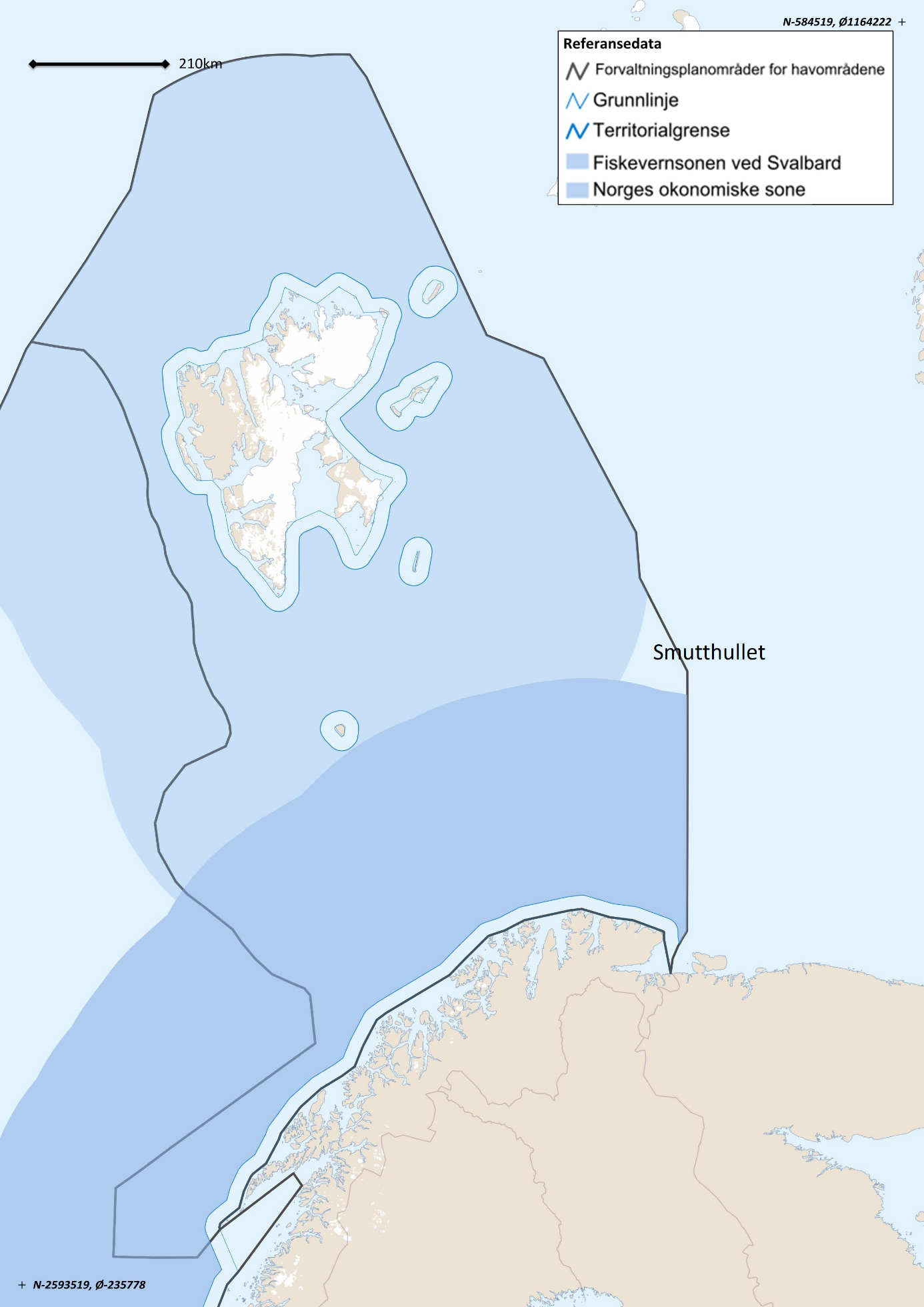
Norsk økonomisk sone (NØS) Fiskevernesonen ved Svalbard Russisk økonomisk sone (RØS) Internasjonalt farvann (Smutthullet)
Figur 3.1. Kart over Norges maritime grenser. Kilde: Arealverktøyet/Barentswatch.
Den norske eksklusive sonen på 200 nautiske mil (NØS) og fiskevernesonen ved Svalbard ble opprettet med virkning fra 1977, med basis i FNs havrettskonvensjon. En viktig del av det institusjonelle grunnlaget for å innføre en forvaltning for å sikre en forsvarlig utnyttelse av fiskeressursene var med dette på plass. NØS omfatter i tillegg til Barentshavet også Norskehavet og Nordsjøen. Norsk fiskerilovgivning gjelder innenfor alle disse tre områdene. Den 20. september 2010 undertegnet Norge og Russland en avtale om en ny sonedeling mellom de to landene i Barentshavet og Polhavet. Avtalen ble godkjent av Stortinget og den russiske dumaen trådte i kraft 7. juli 2011.
Fiskeriene i NØS og i Fiskevernesonen ved Svalbard er regulert i henhold til norsk lov, mens fisket i RØS er underlagt russisk forvaltning, og North East Arctic Fishery Commission (NEAFC) når det gjelder fisket i internasjonalt farvann (Smutthullet), et område der det hovedsakelig fiskes reke.
De fleste og viktigste fiskebestandene vandrer mellom NØS og andre soner, og for å sikre en god forvaltning av fiskeressursene, er et samarbeid med andre land en nødvendig forutsetning. Flere av de viktigste forvaltningsbeslutningene blir således tatt i internasjonale forhandlinger, der resultatene av disse forhandlinger danner grunnlaget for den nasjonale beslutningsprosessen.
Forvaltningen av livet i havet er basert på de best tilgjengelige vitenskapelige råd når det gjelder kunnskap om bestandsstørrelser og økosystemene som disse bestandene er en del av. De fleste bestandene av interesse for Norge i Barentshavet, blir vurdert i samarbeid mellom forskere fra flere land gjennom Det internasjonale rådet for havforskning (ICES). Råd og anbefalinger fra ICES utgjør grunnlaget for fastsettelsen av fiskekvoter.
3.2.1 - Den blandede norsk-russiske fiskerikommisjon
Etter opprettelsen av de økonomiske sonene, inngikk Norge og Sovjetunionen i 1975 en samarbeidsavtale for å sikre en rasjonell forvaltning av de felles marine ressursene i Barentshavet. Som en videreføring av denne avtalen, ble det året etter enighet om å opprette en kommisjon, Den norsk-sovjetiske fiskerikommisjon (fra 1991, Den blandede norsk-russiske fiskerikommisjon), for å behandle praktiske spørsmål i forbindelse med regulering av fiskeriene i området.
Det nære samarbeidet med Russland om forvaltningen av fiskeressursene i Barentshavet, har bidratt til å sikre stabile og bærekraftige bestander. Samarbeidet er formalisert i Den blandede norsk-russiske fiskerikommisjon som løpet av mer enn 45 år har bidratt til å utvikle et tett samarbeid, som fastsetter totalkvoter, forvaltningsstrategier, tekniske reguleringstiltak og rammer for et kontrollsamarbeid.
Forhandlingene mellom Norge og Russland starter etter at ICES har lagt fram sine kvotetilrådninger for det kommende år. Den tosidige kvoteavtalen mellom Norge og Russland avtaler totalkvoter på fellesbestander i Barentshavet og kvotefordelingen mellom landene, samt en kvoteavsetning til tredjeland. Denne avsetningen vil være et viktig element i fiskeriavtalene med EU, Færøyane, Grønland og Island, forhandlinger som vanligvis begynner etter at forhandlingene med Russland er ferdige.
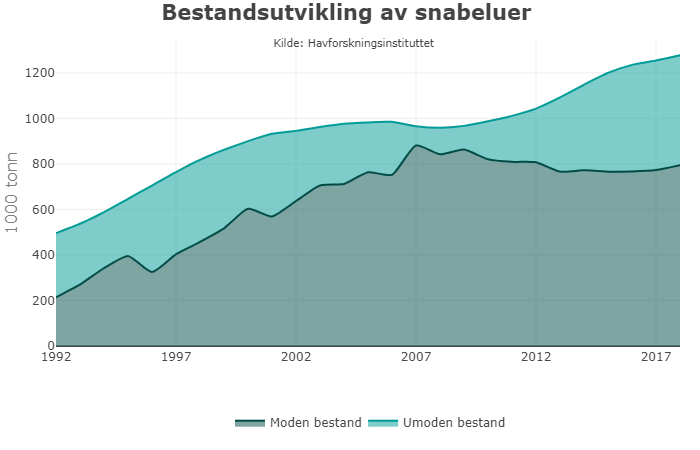
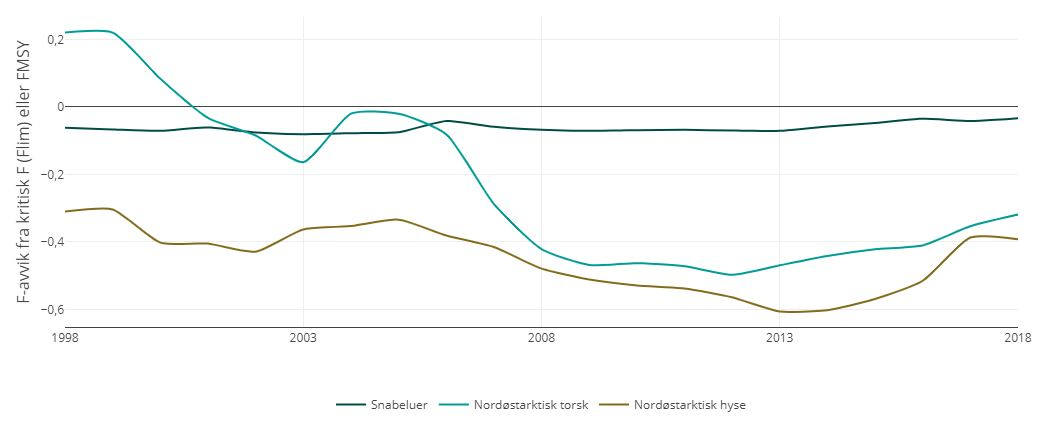
Til fellesbestandene Barentshavet regnes nord-østarktisk torsk, nord-østarktisk hyse, blåkveite, snabeluer og lodde. Adgangen til å delta i fisket i hverandres soner blir avtalt, og avtalene vil også omfatte spørsmål knyttet til teknisk regelverk, fiskerivitenskapelig samarbeid med mer.
Norge og Russland har utviklet høstingsregler for torsk, hyse og lodde som angir bestandsnivå og tiltaksgrenser for kvotefastsettelsen av disse artene. Høstingsreglene følger en beskatningsstrategi som skal tilrettelegge for høy langsiktig avkastning av disse bestandene, noe som krever relativt stabile totalkvoter fra år til år, og forutsetter full utnyttelse av tilgjengelig informasjon om bestandsutviklingen til disse artene.
Fra begynnelsen av 1990-tallet ble fiskerisamarbeidet mellom Norge og Russland utvidet til også å omfatte kontrollspørsmål, og i 1993 ble det etablert et utvalg, Det permanente utvalg for forvaltnings- og kontrollspørsmål. Kontrollsamarbeidet inkluderer utveksling av informasjon om omlasting til havs og landinger i tredjelands havner av fisk som tilhører fellesbestandene i Barentshavet og Norskehavet, felles retningslinjer for hvordan inspeksjoner til havs skal foregå, samt arbeide for å harmonisere det tekniske regelverket i tillegg til spørsmål knyttet til utkast av fisk med mer.
3.2.2 - Nasjonale reguleringer
Når forhandlingene med andre land er fullført, starter den nasjonale behandlingsprosessen for regulering av fiskebestandene. Med utgangspunkt i den inngåtte avtalen utarbeider Fiskeridirektoratet forslag til nasjonale reguleringer - innspill som legges fram for Reguleringsmøtet, et forum der fiskeriorganisasjoner, fagforeninger, Sametinget, lokale myndigheter og andre berørte parter deltar. Reguleringsmøtet er et åpent høringsmøte, og er et rådgivende organ for Nærings- og fiskeridepartementet når det gjelder reguleringen av de fleste kommersielle fiskeslag som norske fiskere utnytter.
Innspillene fra Reguleringsmøtene danner grunnlaget for de endelige reguleringsforskriftene som fastsettes av Nærings- og fiskeridepartementet, forskrifter som angir kvotefordelingen mellom ulike fartøygrupper, samt retningslinjer for gjennomføringen av fisket i det kommende året.
Reguleringsmøtene har bidratt og bidrar til å involvere næringsaktørene i forvaltningen av fiskeressursene, møter som gir fiskerne mulighet til å påvirke utformingen av prinsippene for forvaltningen, og har bidratt til god kontakt og dialog med myndigheter, og havforskere. Reguleringsmøtene er og har vært et viktig forum for å skape en forståelse for nødvendigheten av å regulere fiskeriaktivitet, en forståelse som har bidratt til å gi reguleringene høy grad av legitimitet i næringen.
3.3 - Lovverket som regulerer utnyttelsen av de marine ressurser
Forvaltningen av marine ressurser er forankret i folkeretten og er hjemlet i en rekke norske lover, hvorav de viktigste er:
Lov om retten til å delta i fiske og fangst (deltakerloven) Lov om Norges økonomiske sone (soneloven) Lov om Norges territorialfarvann (territorialfarvannsloven) Lov om førstehandsomsetning førstehåndsomsetning av viltlevande marine ressursar (fiskeslagslova) Lov om Kystvakten (kystvaktloven) Lov om forvaltning av viltlevande marine ressursar (havressurslova)
Av disse lovene er havressursloven den mest sentrale loven som etablerer et helhetlig forvaltningsregime som regulerer utnyttelsen av de viltlevende marine ressursene. Formålet er å sikre en bærekraftig og lønnsom forvaltning av de viltlevende marine ressursene, samt å medvirke til å sikre sysselsetting og bosetting i kystdistriktene. Loven er således både en miljølov og en næringslov, og binder sammen de ulike lovene listet ovenfor.
Havressursloven innfører forvaltningsprinsippet som en grunnleggende regel i forvaltningen av de viltlevende marine ressursene, et prinsipp som setter krav til at forskning og forvaltning skal overvåke de enkelte bestandene, og med jevne mellomrom vurdere tilstanden. På bakgrunn av denne vurderingen skal eventuelle tiltak som anses som nødvendige iverksettes. Disse tiltakene skal sikre en ansvarlig forvaltning av den enkelte bestand, tiltak som i form av en forskrift blir fastsatt med hjemmel i denne havressursloven.
3.3.1 - Regulering av enkelte arter
Når det gjelder reguleringene av enkeltarter, gir havressursloven grunnlag for detaljerte forskrifter når det gjelder kvoter på fartøynivå. Den mest detaljerte artsreguleringen etter nordøstarktisk torsk er å finne i «Forskrift om regulering av fisket etter torsk, hyse og sei nord for 62 °N i 2020» (J-8-2020), en forskrift som angir detaljerte bestemmelser om totalkvoter, gruppekvoter og fartøykvoter innenfor de enkelte fartøygruppene, tillegg til bestemmelser om bifangst og kvoteberegning ved utskiftning/ombygging av fartøy m.m. Denne forskriften gjelder for både Norskehavet og Barentshavet, dvs. områdene i NØS nord for 62 °N. Reguleringsforskriftene for andre arter har tilsvarende oppbygning, men der reguleringsbestemmelsene er mindre detaljerte.
Nordøstarktisk torsk og hyse er de desidert viktigste artene i Barentshavet. I 2019 utgjorde førstehåndsverdien på torsk og hyse henholdsvis vel 7,7 og 1,5 milliarder kroner, nesten 70 % av samlet førstehåndsverdi i området. Andre økonomisk viktige arter er sei og blåkveite, reke og lodde.
Omfanget og detaljgraden i reguleringsforskriftene for de viktigste artene i Barentshavet gjenspeiler den økonomiske betydningen av disse artene. Forvaltningen av disse artene er basert på et omfattende kunnskapsgrunnlag med basis i lange tidsserier, der uttak er basert på analytiske bestandsvurderinger og vedtatte forvaltnings/høstingsregler.
Et annet viktig fiskeri i Barentshavet er trålfisket etter reke, som til nå ikke har vært kvoteregulert fordi de samlede rekefangstene har ligget langt under kvotetilrådningene fra ICES. Utøvelsen av dette trålfisket er i NØS regulert av en egen forskrift om tekniske reguleringer som stiller detaljerte krav utformingen av til fangstredskapet.
3.3.2 - Tekniske reguleringer
Utøvelsen av alt fiske er regulert ved en egen forskrift, «Forskrift om utøvelse av fiske i sjøen» (J-255-2019), som blant annet definerer minstemål for en rekke arter, størrelsen på maskevidden i ulike redskapstyper og bifangstbestemmelser som angir tillatt mengde av fisk under minstemål. Et viktig formål med forskriften er å begrense uttak av fiske under minstemål. Utøvelsesforskriften inneholder i tillegg en rekke krav til tekniske detaljer med hensyn til utforming og bruk av redskap, og dette gjelder særlig den tekniske utformingen av trålredskap. I tillegg til utøvelsesforskriften er trålfisket også regulert med egne forskrifter som påbyr bruk av sorteringsrist i flere områder, for ytterligere å redusere bifangst av fisk under minstemål (J-181-2017 og J-26-2020).
Bifangst av andre arter, dvs. mindre bestander det som ikke er utsatt for et målrettet fiske, kan representere en stor reguleringsmessig utfordring, da de kun forekommer som tilfeldig og uunngåelig bifangst i fisket etter andre arter. I Barentshavet gjelder dette arter som for eksempel skater og rokker, flyndre m.m. Storskate og vanlig uer er arter som er plassert på Rødlisten og regnes som kritisk truet. Det foregår ikke et målrettet fiske etter disse artene, men de forkommer som tilfeldig og uunngåelig bifangst.
3.4 - Områdereguleringer
Fiskeridirektoratets sjøtjeneste (tidligere Overvåkningstjenesten for fiskefelt) ble opprettet i 1984. Inspektørene fra Fiskeridirektoratets sjøtjeneste følger fiskefartøyene i flere uker. Kontrollene er basert på risikovurdering, det vil si at inspektørene oppsøker de fiskeriene der de vet det for eksempel kan være mye liten fisk/reker og/eller bifangst.
Fiskeridirektoratets sjøtjeneste arbeider i hele vernesonen og i norsk økonomisk sone. På grunnlag av representative prøver fra det kommersielle fisket, kan fisket nærmere angitte områder bli stoppet dersom prøvene viser for høy innblanding av fisk/reke under minstemål, eller for høy innblanding av andre arter. Det er områder der det fiskes med torsketrål, snurrevad, reketrål, seinot eller etter arter som sild og lodde, som oftest blir berørt av midlertidig stenging. I 2019 ble inntil seks områder stengt for fiske i lengre og kortere perioder.
3.5 - Sjøpattedyr
Fangsten av vågehval er regulert med kvoter fastsatt som gjelder for alle havområdene i NØS. De årlige kvotene er fastsatt med utgangspunkt i beregningsmodeller utarbeidet av vitenskapskomiteen i Den internasjonale kvalfangstkommisjonen (IWC), som gir god sikkerhet for bærekraftig fangst på vågehvalbestanden. Barentshavet er det viktigste fangstområdet, og de siste årene har så å si all fangst blitt tatt i området, og i 2019 ble 426 av totalt 429 dyr skutt i Barentshavet.
I Barentshavet har norske fartøy drevet fangst av grønlandssel og klappmyss i Østisen, det vil si farvann rundt Kvitsjøen på kvoter tildelt fra Russland. Selkvotene er basert på rådgivning fra en arbeidsgruppe i Den blandete norsk-russiske fiskerikommisjon. I de to siste årene har det har ikke vært drevet norsk fangst i disse områdene. Norske fartøy som deltar i fangst av sel må ha egen tillatelse, og fangst og fangstutøvelse er regulert med egne forskrifter.
3.6 - Nye arter: Kongekrabbe og snøkrabbe
Kongekrabbe er en introdusert art i Barentshavet, satt ut av russiske forskere i Murmanskfjorden på 1960-tallet for å etablere en ny fiskeressurs. Det opprinnelige utbredelsesområde er Kamtsjatka-halvøyen i det nordlige Stillehavet. Kongekrabben har siden spredd seg østover og vestover langs kystlinjen og ut i åpne havområder i det sørlige Barentshavet, og har etter hvert blitt en verdifull ressurs for teinefiske i fjordene og i kystnære farvann i Øst-Finnmark .
Kongekrabbe forvaltes etter to målsettinger, der den ene er å opprettholde et langsiktig fiskeri innenfor et avgrenset område i Øst-Finnmark, og den andre er å begrense spredningen av kongekrabbe til områder vest for dette området.
Snøkrabbe ble første gang registrert i Barentshavet av russiske forskere i 1996, og åtte år senere ble det registrert fangster i norsk del av Barentshavet. Snøkrabben er nå en etablert bestand i Barentshavet, der hovedtyngden av bestanden fortsatt befinner seg i russisk farvann. Den sprer seg også nord og vestover til farvann rundt Svalbard.
Norge og Russland ble i 2015 enige om at snøkrabben skal betraktes som en sedentær art, dvs. en stedegen bunnart, og kan derfor forvaltes etter sokkelgivning på samme måte som olje, og sokkelstatene Norge og Russland kan med dette alene forvalte bestanden av snøkrabbe i Barentshavet. I 2017 ble derfor tredjeland utestengt fra å fiske snøkrabbe i Barentshavet, som medførte at EU-land som hadde fisket snøkrabbe de foregående årene ble utestengt fra fiskeriet fra og med 2017.
I 2017 fastsatte Nærings- og fiskeridepartementet en kvote på 4000 tonn snøkrabbe for norsk sone i Barentshavet, mens kvoten for 2019 var på 4 500 tonn. I 2018 og 2019 fisket norske fartøy henholdsvis vel 2800 tonn og vel 4000 tonn snøkrabbe.
3.7 - Vern av bunnhabitat
I området rundt Bjørnøya er det et forbud mot å drive fiske innenfor 20 nautiske mil av grunnlinjen, med unntak av et fiske med reketrål og skjellskraping, og et absolutt forbud innenfor verneområde 4 nautiske mil av grunnlinjen som rundt Bjørnøya (J-23-2018 og J-24-2018).
I 2011 ble alt fiske med bunnredskap på større dyp enn 1000 m forbudt i norske farvann, og i 2019 ble det innført et forbud mot fiske med bunnredskap i åtte områder i fiskevernsonen ved Svalbard. Dessuten ble det innført restriksjoner på fiske i «nye» områder i fiskevernsonen, dvs. områder som tidligere ikke har blitt utnyttet.
Ved sammenstøt med sårbare habitater (koraller, svamper), og dersom det er mer enn 60kg korall eller 800kg svamper i fangsten, skal fartøy rapportere hendelsen til Fiskeridirektoratet og flytte minst to nautiske mil før fisket kan gjenopptas (Jfr. J-40-2016).
4 - Tilstanden i økosystemet for de ulike komponentene
4.1 - Klima
Et karakteristisk trekk ved Barentshavet er at klimatiske faktorer som vanntemperaturer og havisforhold naturlig varierer betydelig fra år til år. Vanntemperaturen i Barentshavet har økt i de siste 40 årene, men med markante variasjoner. Parallelt med temperaturøkningen har utbredelsen av havis avtatt. Reduksjonen i havisens utbredelse i Arktis, og særlig i Barentshavet, siden satellittmålinger startet i 1979 er godt dokumentert. De tre siste årene har imidlertid både temperaturen og mengden innstrømmende atlanterhavsvann til Barentshavet avtatt, og dette har ført til en økning i området dekket av havis om vinteren. I 2019 var temperaturene på samme nivå som tidlig på 2000-tallet, men fortsatt betydelig høyere enn under tidligere kjølige perioder.
4.1.1 - Vanntemperatur og vannmasser
Vanntemperaturen i Barentshavet har økt de siste 40 årene, men med markante variasjoner (figur 4.1). Atlanterhavsvannet som strømmer inn i Barentshavet i sørvest er avgjørende for havklimaet i dette området, og tidsserier fra snittene Fugløya-Bjørnøya og Vardø-Nord viser at vanntemperaturen på sensommeren har økt med omkring 1,5 °C siden slutten av 1970-tallet. På slutten av 1970-tallet var Barentshavet inne i en relativt kald periode av den naturlige klimasyklusen til Atlantic Multidecadal Oscillation (AMO) (Boitsov et al. 2012; Smedsrud et al. 2013), så deler av økningen i vanntemperatur etter dette kan knyttes til naturlig klimavariabilitet. Også i 1930- og 1950-årene var det varmt i Barentshavet, men ikke fullt så varmt som i årene etter 2006. Årene med høyest temperatur i det innstrømmende atlanterhavsvannet var 2006 og 2015. Etter 2015 har vanntemperaturene gått ned, og i 2019 var de nede på samme nivå som de var tidlig på 2000-tallet. Dette har sammenheng med lavere temperaturer i atlanterhavsvannet i Norskehavet.
Figur 4.1. Temperatur i kjernen av atlanterhavsvannet som strømmer inn i Barentshavet fra sørvest i snittene Fugløya-Bjørnøya og Vardø-Nord fra 1950 og frem til og med 2019. Svarte linjer viser årlige verdier (på sensommeren), tykke linjer viser tre års løpende gjennomsnitt og stiplede svarte linjer viser langtidsmiddelet (1981-2010).
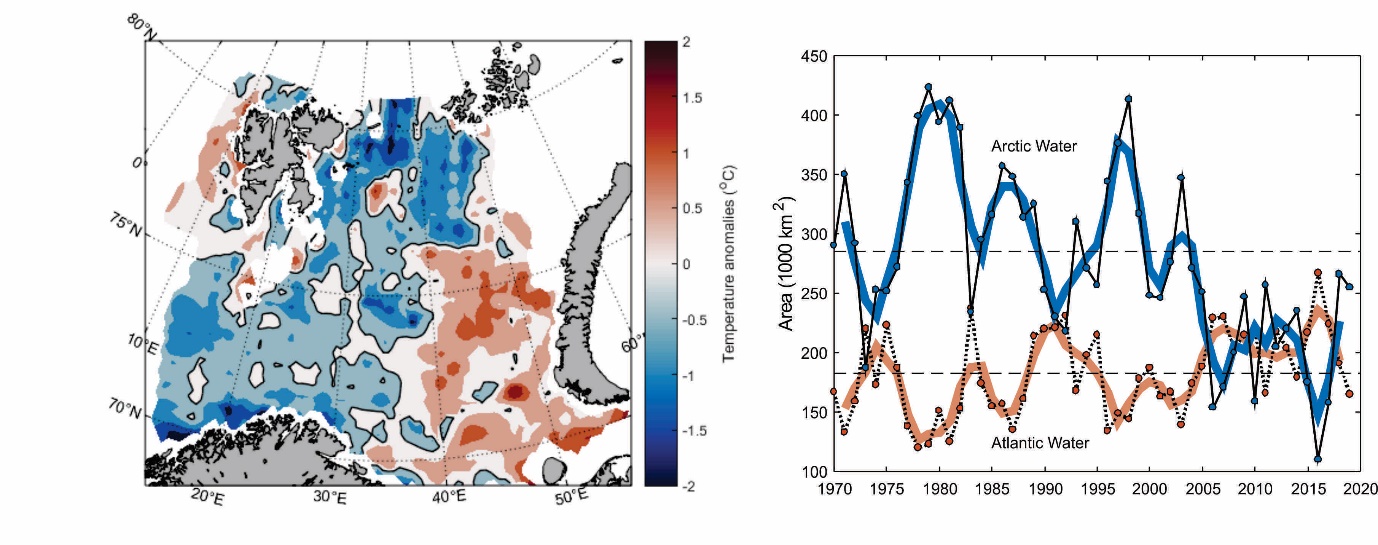
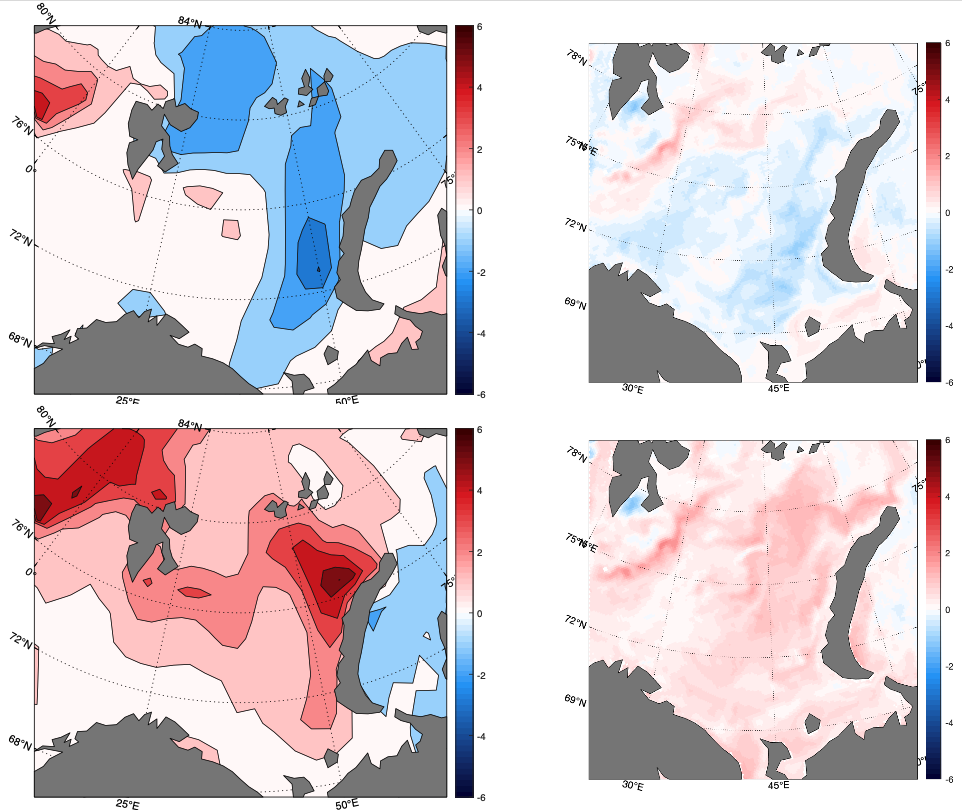
Målinger fra hele Barentshavet på sensommeren 2019 (figur 4.2) viser at mesteparten av det vestlige Barentshavet på det tidspunktet hadde temperaturer som lå mellom 0 og 0,5 °C under langtidsmiddelet. De laveste temperaturene ble funnet helt i nord og sørvest. Øst av omkring 40 °Ø var temperaturene mellom 0 og 1 °C over langtidsmiddelet.
Mens det sørlige Barentshavet er dekket av varmt atlanterhavsvann, er den nordlige delen dekket av kaldt arktisk vann (Loeng 1991). Skillet mellom de to områdene kalles polarfronten. Tidsserier av areal dekket av arktisk vann viser et tydelig skille før og etter rekordåret 2006, med betydelig større utbredelse av kaldt arktisk vann før enn etter (figur 4.2). Dette sammenfaller med perioden med sterk oppvarming av arktisk vann (Lind et al. 2018). Arealet dekket av varmt atlanterhavsvann viser en mer gradvis økning over hele perioden, og varierer i stor grad i takt med temperaturen på atlanterhavsvannet som strømmer inn fra sørvest. Størst utbredelse av atlanterhavsvann, og minst utbredelse av arktisk vann, ble observert i 2016, altså året etter det var rekordhøye temperaturer i det innstrømmende atlanterhavsvannet. Etter 2016 har utbredelsen av atlanterhavsvann gått ned samtidig som utbredelsen av kaldt arktisk vann har økt. Utbredelsen av de to vannmassene var i 2019 tilbake til samme nivå som det var rundt 2004-2005.
Figur 4.2. Figuren til venstre viser temperaturavvik i 50-200 m dyp på sensommeren 2019 i forhold til langtidsmiddelet for perioden 1981-2010. Figuren til høyre viser arealet i Barentshavet som er dekket av varmt atlanterhavsvann (varmere enn 3 °C) og kaldt arktisk vann (temperaturer under 0 °C) på sensommeren fra 1970 og frem til og med 2019. Svarte linjer viser årlige verdier, tykke linjer viser tre års løpende gjennomsnitt og stiplede svarte linjer viser langtidsmiddelet (1981-2010).
4.1.2 - Innstrømming av atlanterhavsvann
I tillegg til variasjoner i temperaturen på atlanterhavsvannet, er det også variasjoner i mengden atlanterhavsvann som strømmer inn i Barentshavet (Ingvaldsen et al. 2004). Høy innstrømming gir økning i temperatur, fordi det innstrømmende vannet jevnt over er varmere enn vannet i Barentshavet. Dessuten bringer vannet med seg dyreplankton og fiskelarver inn i området (se for eksempel Dalpadado et al. (2012)). Dynamikken i primærproduksjonen og energistrømmen gjennom næringskjeden påvirkes av innstrømningen. Den samlede hastigheten på innstrømningen til Barentshavet er på vel 3 Sverdrup (1 Sverdrup (Sv) = 1 million m3/s). Mesteparten av dette, omkring 2 Sv, er atlanterhavsvann. Innstrømningen av atlanterhavsvann til Barentshavet varierer mye, som vist i figur 4.3. Årsaken til de observerte variasjonene er knyttet til variasjoner i vindforholdene vest i Barentshavet (Ingvaldsen et al. 2004).
Innstrømmingen varierer i perioder på flere år. Årene 2006 og 2015-2016 var ekstreme år med svært høy innstrømming, og i disse årene var også temperaturen i det innstrømmende vannet høy. Disse årene strømmet det altså inn både mer og varmere atlanterhavsvann enn de andre årene. I 2017 var det ganske moderat innstrømming, og i 2018 steg den noe igjen. Måleserien har foreløpig bare data tilgjengelig frem til våren 2019, så det er ikke kjent hvordan innstrømmingen har vært resten av 2019.
Figur 4.3. Avvik i innstrømming (lilla linjer) og temperatur (grønne linjer) av atlanterhavsvann inn i Barentshavet. Målingene er gjort i området mellom norskekysten og Bjørnøya (Fugløya–Bjørnøya-snittet). Avviket er målt i forhold til middelet over perioden 1997-2019. Innstrømningen er oppgitt i Sverdrup (1 Sv = 1 million m3/s) mens temperaturen er gitt i °C. Tre måneders (tynne linjer) og ett års (tykke linjer) glidende middel er vist.
Vanligvis er innstrømmingen størst om vinteren og minst om våren (figur 4.4), og de ekstreme årene 2006 og 2015-2016 hadde svært høy innstrømming om vinteren. Etter 2015-2016 har innstrømmingen om vinteren og våren gått ned med nesten 2 Sv. Over samme periode har innstrømmingen om sommeren og høsten økt, men ikke like mye. Det har dermed vært en endring i sesongvariasjonen i innstrømmingen de siste 5-6 årene. Nedgangen i innstrømming av atlanterhavsvann om vinteren, og den samtidige nedgangen i temperaturen i atlanterhavsvannet, har ført til endringer i havis (se kapittel 4.1.3).
Figur 4.4. Innstrømming av atlanterhavsvann i fire sesonger. Innstrømningen er oppgitt i Sverdrup (1 Sv = 1 million m3/s) og middel over perioden 1997-2019 er vist med stiplet linje.
4.1.3 - Havisen i Barentshavet
Utbredelsen av havis i Barentshavet er et sesongmessig fenomen. Isdannelsen begynner vanligvis om høsten i de nordlige og østlige delene av regionen, og iskantsonen strekker seg sør- og vestover gjennom vinteren. Maksimal isutbredelse er vanligvis i april. Når solinnstrålingen øker utover våren starter vårsmeltingen. Da minker iskonsentrasjonen og iskantsonen trekker seg tilbake, nord- og østover. Denne prosessen foregår frem til slutten av august og første halvdel av september, og havområdet har oftest et sesongminimum i isutbredelse i september. Trenden over tidsrommet 1979-2017 viser at isen har blitt dannet senere på høsten/vinteren og smeltet tidligere på våren (Stroeve og Notz 2018 ), og i senere år har mesteparten av Barentshavet har vært isfritt i sommermånedene (f.eks. Onarheim et al. (2018)).
Havområdet er karakterisert av store mellomårlige variasjoner og langtidstrender i isforhold (se figur 4.5). Mellomårlig variasjon i isutbredelse er nært knyttet til variasjon i temperatur i sjøen og lufta samt i mengde innstrømmende atlantisk vann (Årthun et al. 2012), som er påvirket av nordlige vinder og lavtrykksaktivitet i de nordiske hav (Smedsrud et al. 2013; Sorteberg og Kvingedal 2006). I tillegg er lokale vindforhold (Onarheim og Årthun 2017) og istransport fra nord og øst inn til i Barentshavet viktig for mengden sjøis (Lind et al. 2018). Det har vært en markant nedgang i tilførsel av havis fra områder nord for Barentshavet etter 2005 (Lind et al. 2018). Dette kan skyldes mindre havis i tilgrensende havområder og endrede værmønstre.
For Barentshavet er midlere isutbredelse beregnet for månedene april og september i perioden 1979–2019, innenfor et nærmere angitt areal (72 °N - 82 °N; 10 °Ø - 60 °Ø). Isutbredelsen er målt som summen av areal med iskonsentrasjon over 15%. Tidsserien er basert på passive mikrobølgedata fra instrumentene SMMR (Nimbus-7) og SSM/I-SSMIS (DMSP) (Cavalieri et al. 1996, oppdateres årlig), med en romlig oppløsning på 25 km. Data fra det siste året i tidsserien har en foreløpig status. Etter ett år blir datakvaliteten for disse forbedret med reprosessering. For å ha med de mest aktuelle data i denne rapporten, er foreløpige data for 2019 tatt med.
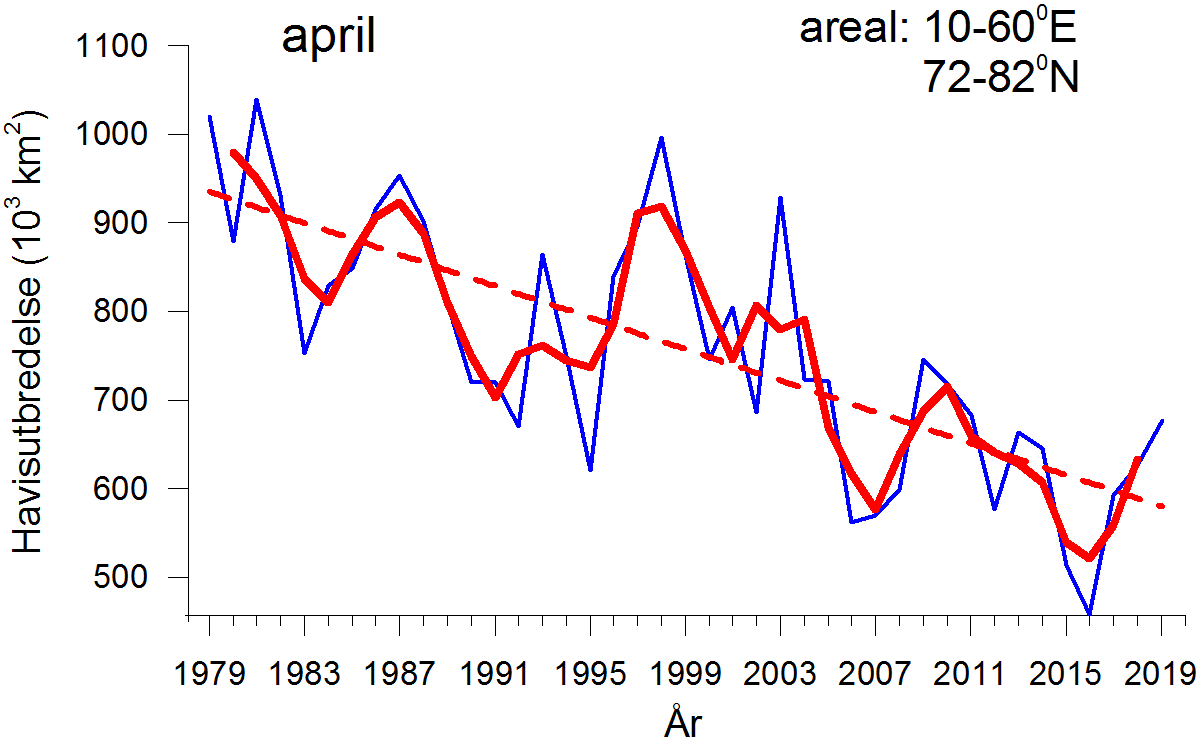
For april, måneden med størst isutbredelse i Barentshavet, viser tidsserien en negativ trend i perioden 1979–2019 i tillegg til betydelig variasjon mellom år (figur 4.5, øvre del). Tidsserien for havisutbredelse i april 1979–2019 viser fem år som skilte seg ut med stor havisutbredelse (1979, 1981, 1987, 1998 og 2003) og seks år med spesielt liten utbredelse (2006, 2007, 2008, 2012, 2015 og 2016). Året 2016 var et absolutt minimum for isutbredelse i april i hele perioden.
For september, måneden med minst isutbredelse, viser tidsserien også en negativ trend for hele perioden 1979-2019 (figur 4.5, nedre del). I september har det vært fem år med spesielt stor havisutbredelse (1982, 1989, 1993, 2003 og 2014) og ni år med spesielt liten isutbredelse (1979, 1984, 1996, 2001, 2004, 2011, 2012, 2013 og 2018) i Barentshavet. I flere år etter 2010 har Barentshavet vært helt isfritt om sommeren.
Et viktig kjennetegn til havisen i Barentshavet er den store variasjonen med årstidene og mellom år. Dette regnes som naturlige variasjoner og fører til at store deler av Barentshavet er isfritt deler av året, i større grad enn i andre arktiske randhav. Den sørvestlige delen av Barentshavet har åpent vann hele året fordi varmt atlanterhavsvann fyller hele vannsøyla her. Det nordlige Barentshavet er lagdelt med en kald, ferskere arktisk vannmasse over atlantisk vann. Her beskytter arktisk vann isdekket mot varmen i det atlantiske vannlaget under, på samme måte som i andre isdekte havområder i Arktis (Lind et al. 2016). Tetthetsforskjellen mellom det arktiske og det atlantiske vannlaget angir styrken på lagdelingen, og tetthetsforskjellen er styrt av saltholdigheten i det arktiske vannet (Lind et al. 2016). Jo ferskere det arktiske vannlaget er, jo vanskeligere er det å blande opp atlantisk vann. Etter 2005 har dette arktiske vannlaget blitt varmere og saltere, og beskytter dermed isdekket dårligere (Lind et al. 2018).
Figur 4.5. Isutbredelse for Barentshavet 1979–2019 (terskel: > 15 % iskonsentrasjon; data er basert på observasjoner fra passive mikrobølgesatellitter) vist som maksimum utbredelse (april) og minimumsutbredelse (september). Det er vist månedsmiddelverdier (tynne kurver), løpende middelverdier over tre år (april og september henholdsvis; tykk linje), og den lineære trenden. Den mellomårlige variasjonen er stor, men også trender er tydelig både ved maksimums- og minimumsutbredelse. Data fra 2019 er inkludert i diagrammene, men datakvaliteten her er lavere inntil de er prosessert (gjøres først ett år etter observasjonen).
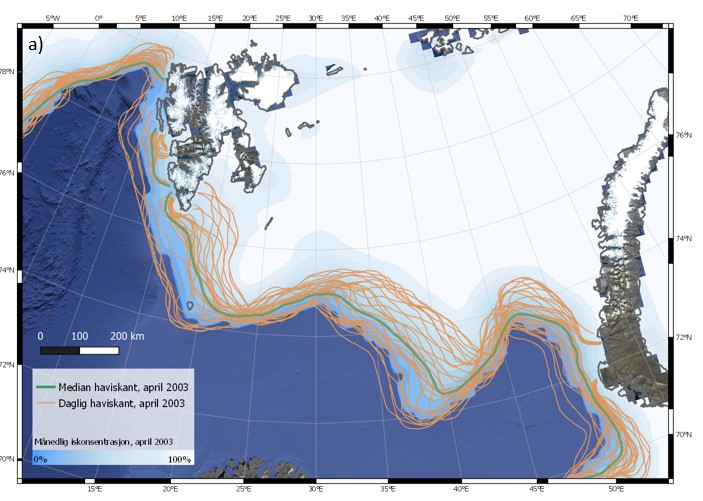
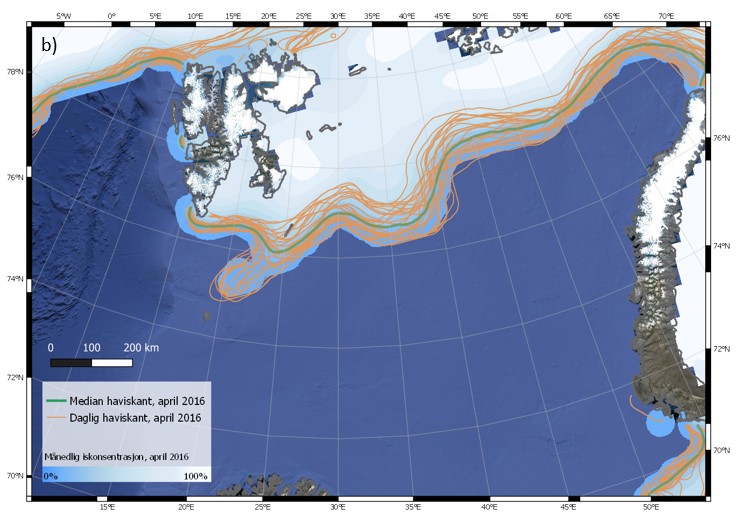
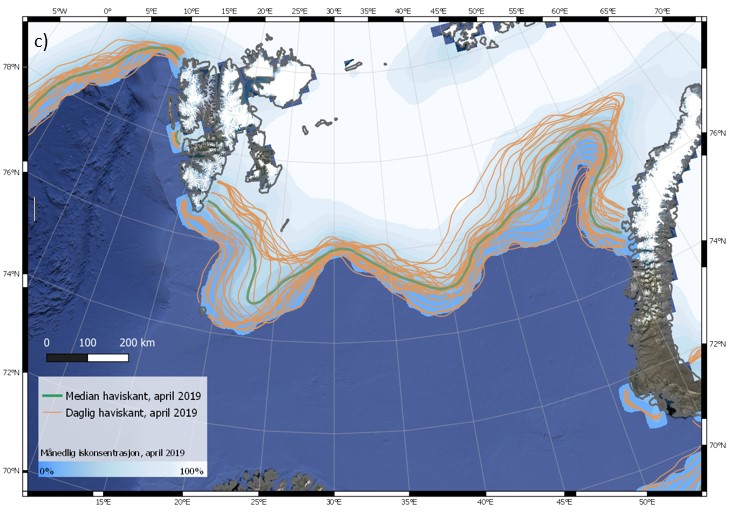
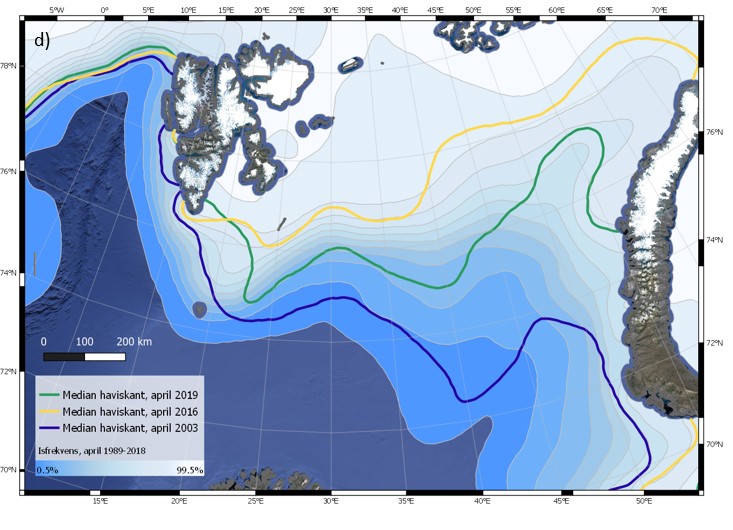
Iskanten kan også variere mye fra dag til dag. Eksempler på dette er vist i Figur 4.6 (a-c) for tre ulike år, 2003, 2016 og 2019. Spesielt for april 2003 blir det synlig at iskantens forløp endres på kort tid i et nokså stort område. Faktorer som bidrar til raske endringer av iskantposisjonen er vind, havstrømmer, samt temperaturforhold som kan påvirke frysing og smelting. Sammenligning av median for april måned for de tre angitte årene (figur 4.6 d) viser tydelig hvor variabel isutbredelsen i april kan være i Barentshavet. Den varierer svært mye mellom dager og år sammenlignet med iskantposisjonen i Framstredet vest for Svalbard, der iskantmedian for april i de tre angitte årene ligger nært hverandre (figur 4.6 d). Også iskantposisjoner for enkeltdagene i april for de tre årene viser et tilsvarende mønster, med større variasjoner i Barentshavet.
Figur 4.6 (a-d). Haviskanten i Barentshavet vist for hver dag i april måned (15 % iskonsentrasjon; oransje kurver) vist sammen med månedsmedian (grønn kurve) og midlere iskonsentrasjon for april (blåhvit bakgrunn) i 2003 (a), 2016 (b) og 2019 (c). Tilsvarende er median haviskant (15 % iskonsentrasjon) for april for de samme tre årene (blå, grønn og oransje kurver) vist sammen med midlere isfrekvens for april i perioden 1989–2018 (blåhvit bakgrunn; d; for isfrekvens se Itkin et al. (2014)). Havisdataene er basert på observasjonen fra passive mikrobølgesatellitter og bearbeidet av National Snow and Ice Data Center (Meier et al. 2017a; Meier et al. 2017b).
Det er nokså begrenset med informasjon om havistykkelse i Barentshavet, men flere studier peker på at det er en trend mot tynnere is over de siste tiår (for det nordvestlige Barentshavet se King et al. (2017) og for fastis ved Hopen se Gerland et al. (2008)). Dette er i samsvar med sammenligning av observasjoner siden 2010, og med tidligere observasjoner nord for Svalbard, i grenseområdet mellom Barentshavet og Polbassenget (Renner et al. 2013; Rosel et al. 2018).
Det er relevant å sammenligne utviklingen i isdekke i Barentshavet med det øvrige Arktis. Reduksjon av havisdekke i Arktis på større romlig skala siden 1980-tallet er kjent og beskrevet (f.eks. Barber et al. (2017); Perovich et al. (2019)), og flere studier har vist at de største endringene ble observert i den Eurasiske sektoren av Arktis, spesielt i Barentshavet (f.eks. Lind et al. (2018); Onarheim et al. (2018); Onarheim og Årthun (2017); Pavlova et al. (2014); Stroeve og Notz (2018)). Mellomårlige variasjoner (minimumsisutbredelse) for hele Arktis korrelerer ikke med utviklingen i Barentshavet, men noen av ekstremårene med svært lite is sammenfaller, som 2012 og 2016 (Wang et al. 2019). Trender peker nedover, i samme retning, for både Arktis generelt og Barentshavet, men nedgangen i vintersjøis er eksepsjonelt stor i Barentshavet (Onarheim og Årthun 2017; Screen og Simmonds 2010). Det nordlige Barentshavet er del av et «hotspot» som har det største tapet av havis og den raskeste oppvarmingen av nedre atmosfære på vinterstid i hele Arktis (f.eks. Screen og Simmonds (2010)). Dette henger sammen med den markante nedgangen i tilførsel av sjøis til Barentshavet etter 2005. Sjøisen, som smelter om sommeren, er den primære ferskvannstilførselen til det arktiske vannet og viktig for å opprettholde lagdelingen. Nedgangen i tilførsel av sjøis har derfor ført til et stort tap av ferskvann, med 40% reduksjon i de øvre 100 m av havet i 2010-2016 sammenlignet med klimatologisk middel for 1970-1999 (Lind et al. 2018). Dette har gitt svakere lagdeling og økt oppblanding av atlantisk vann til øvre vannmasser, og denne prosessen bidrar til at det nordlige Barentshavet har sterkere oppvarming og istap enn resten av Arktis (Lind et al. 2018).
Ved siden av langtidsovervåking bidrar prosesstudier til en bedre forståelse av havisprosesser i Barentshavet, noe som også kan forbedre tolkningen av langtidsendringene og konsekvensene av disse. To nyere viktige slike prosesstudier/prosjekter er N-ICE2015, en studie der Norsk Polarinstitutts tidligere fartøy RV «Lance» drev med isen nord for Svalbard, gjennomført i 2015 (Granskog et al. 2018); og prosjektet «Arven etter Nansen», et stort norsk prosjekt påbegynt i 2018 som skal vare til 2023 (se Reigstad et al. (2019) og arvenetternansen.com). Arven etter Nansen skal gi integrert kunnskap om havklima og økosystem i rask endring, med fokus på det nordlige Barentshavet.
4.1.4 - Havforsuring i Barentshavet
Det er store variasjoner i havforsuring i Barentshavet. Trendene for pH og aragonittmetning (kalkmetning) i Barentshavet er svake, men viser generelt en nedgang, i retning av økt havforsuring. Klimaet i polarområdene endres raskt i takt med økt atmosfærisk CO2 -konsentrasjon, samtidig som havets biogeokjemi forandres. Havet tar opp CO2, hvilket bidrar til at pH og karbonationer minker i havet, såkalt havforsuring. Undermetning av aragonitt (dvs. ΩAr <1) betyr at kalkskall løses kjemisk og kalkskalldannende marine organismer kan få problemer med danning av kalkskall (Fransson et al. 2016). Redusert havisdekke og oppvarming bidrar til økt mengde ferskvann ved overflaten, økt lagdeling av vannmassene og endringer i primærproduksjonen. Disse endringene medfører endringer i karbonsyklusen og utveksling av CO2 mellom havet og atmosfæren (Chierici og Fransson 2018). På grunn av sesong- og mellomårlig variasjon av disse driverne er det behov for lange tidsserier for å forstå trender i havforsuring.
Det er store variasjoner i Barentshavet på grunn av variasjoner i vannmasser, biologisk produksjon og isdannelse i de nordlige delene. Opptak av atmosfærisk CO2 i Barentshavet er til stor del styrt av biologiske prosesser. I den nordlige delen bidrar isdannelse og is-smeltevann til lav pH og alkalinitet (havets buffer mot forsuring) og videre CO2 opptak. På samme tid er det en økning i alkalinitet og temperatur i sør, noe som kan skyldes at det er økt tilførsel av atlanterhavsvann til området (Chierici et al. 2017). Atlanterhavsvannet fører også med seg CO2 som er tatt opp fra atmosfæren, som videre bidrar til å øke havforsuring i Barentshavet. De to ulike vannmassene arktisk vann og atlanterhavsvann møter hverandre og danner en polarfront på grunn av store forskjeller hovedsakelig i salt og temperatur, men også i kjemiske konsentrasjoner. De ulike betingelsene i polarfronten og vannmasser i sør og nord bidrar til variasjoner i biogeokjemiske prosesser, havets karbonopptak og havforsuring (Chierici og Fransson 2018).
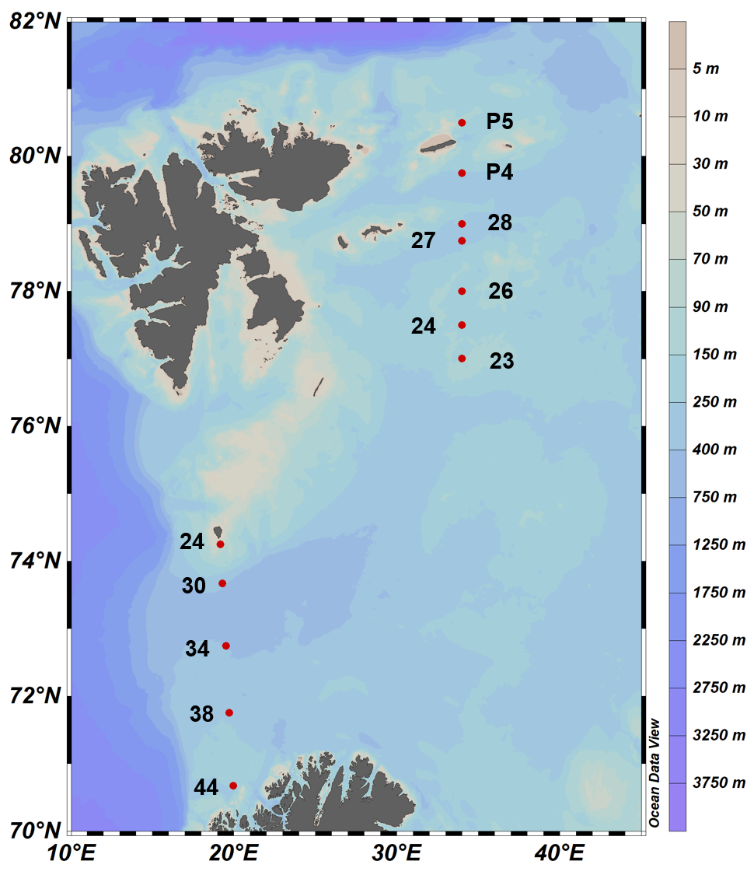
Figur 4.7. Kart og stasjonsnummer fra Fugløya-Bjørnøya-seksjonen i januar 2018 og i den nordlige delen av Barentshavet i august og september 2018. Havforsuring er studert i Fugløya-Bjørnøya-seksjonen siden 2011 mens tidsserier i nordlige Barentshavet ble initiert i 2013.
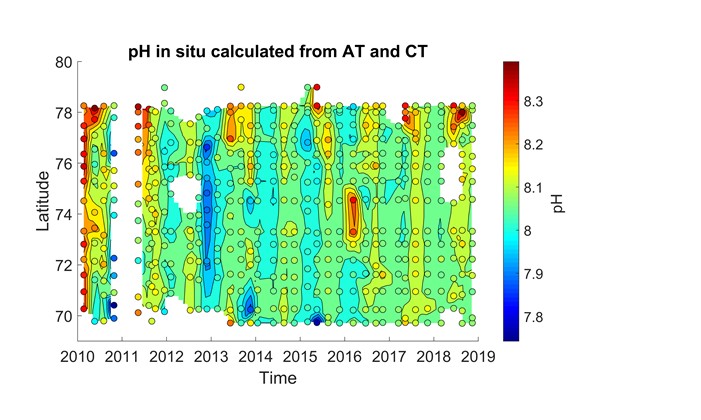
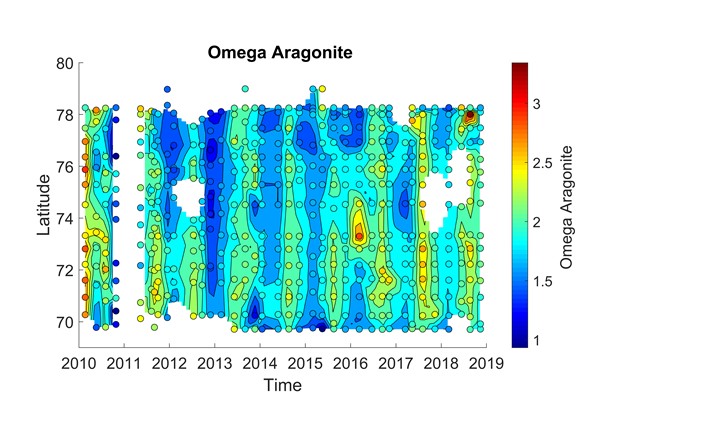
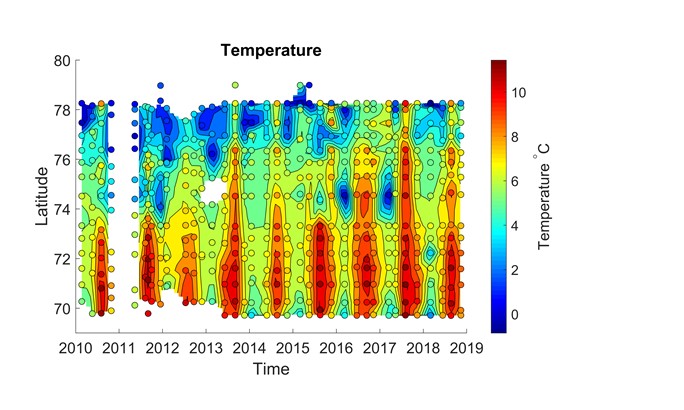
Det er store regionale forskjeller i havforsuring i sør og i nord i Barentshavet (figur 4.7), og dette vises tydelig i overflatevann mellom Svalbard og fastlands-Norge (figur 4.8 og figur 4.9). I nordlige deler påvirkes karbonsystemet av det kalde og ferske vannet fra Arktis og av smeltevann, mens lenger sør spiller det varme atlanterhavsvannet en større rolle (figur 4.10). Det er generelt lavere pH i overflaten i sør enn i nord, og det er tydelig høyere pH om sommeren enn om vinteren (figur 4.8). Når det gjelder kalkmetning av aragonitt (ΩAr), er det høyere ΩAr i den sørlige delen enn i den nordlige, og det var en tydelig undermetning av ΩAr (ΩAr <1) i 2013 både om sommeren og vinteren (figur 4.9). En økning av ΩAr i overflaten fra 2015 til 2018 om sommeren kan delvis bero på en tregraders økning i overflatetemperatur i de nordlige delene fra 2015 til 2018 (figur 4.9 og figur 4.10)
Figur 4.8. pH in situ i overflatevann (i farger) mellom Svalbard og fastlands-Norge (y-akse) i løpet av årene 2010 til 2018 (x-akse). Sirkler viser måledata og fargede områder rundt er beregnede verdier.Figur 4.9. Kalkmetning (ΩAr, omega aragonitt) i overflatevann (i farger) mellom Svalbard og fastlands-Norge (y-akse) i løpet av årene 2010 til 2018 (x-akse). Sirkler viser måledata og fargede områder rundt er beregnede verdier.Figur 4.10. Temperatur (°C) i overflatevann (i farger) mellom Svalbard og fastlands-Norge (y-akse) i løpet av årene 2010 til 2018 (x-akse). Sirkler viser måledata og fargede områder rundt er beregnete verdier.
I de dypeste stasjonene (300-400 m) ved Fugløya-Bjørnøya-seksjonen (72,8 °N, figur 4.7) varierte vanntemperaturen med tre grader og konsentrasjonen av uorganisk løst karbon (CT) økte signifikant i perioden 2012 til 2018 (Jones et al. 2019). Disse mellomårlige variasjonene er assosiert med variasjon i tilførsel av atlanterhavsvann i sør og det arktiske vannet i nord (figur 4.11). Generelt var det kalde arktiske vannet assosiert med høyere CT, spesielt synlig i 2018. Saltholdighet ved 300-400 m dyp viste en endring fra 2012 til 2018 til ferskere og mer arktisk vann. Fra 2016 til 2018 minket aragonittmetningen (ΩAr) fra 1,8 til 1,4 (figur 4.11).
Figur 4.11. Trendanalyse av vanntemperatur, saltholdighet, CT og aragonittmetning (ΩAr) ved 300-400 m dyp, stasjon 72,8 °N i Fugløya-Bjørnøya-seksjonen.
I den nordlige seksjonen i Barentshavet ved 250-300 m dyp (79.5 °N, figur 4.7) vistes endringer i havforsuringsparametene fra 2012 til 2018 (figur 4.11 og figur 4.12). I denne perioden ble havvannet ferskere, kaldere og pH økte generelt med 0,006 per år, men uten en tydelig trend. Aragonittmetning varierte i løpet av perioden med middelverdi på 1,3 (Jones et al. 2019). Dette området er i økende grad påvirket av kaldt og ferskt arktisk vann, hvilket også bidrar til stor variasjon i karbonkjemien.
Figur 4.12. Trendanalyse av vanntemperatur, saltholdighet, pH og aragonittmetning (ΩAr) ved 250-300 m dyp, stasjon 79,5 °N i nordlige Barentshavet.
4.1.5 - Årsaker til status og endringer i klima
Overflatetemperaturen i Arktis har økt dobbelt så mye som det globale gjennomsnittet (Meredith et al. 2019 - in press). Den globale temperaturøkningen skyldes menneskeskapte utslipp (IPCC 2014). At temperaturen stiger dobbelt så raskt i Arktis, skyldes hovedsakelig at havisdekket har blitt mindre som følge av global oppvarming, og at det mørke havvannet absorberer mer energi fra solen nå enn det den lyse isen gjorde før (Meredith et al. 2019 - in press). På toppen av denne trenden fra global oppvarming kommer naturlig variabilitet i temperatur og mengde av det innstrømmende atlanterhavsvannet og i tilførsel av sjøis til Barentshavet. Årsaken til de observerte mellomårlige variasjonene i innstrømming av atlanterhavsvann er knyttet til variasjoner i vindforholdene vest i Barentshavet (Ingvaldsen et al. 2004). Disse kan igjen henge sammen med storskala atmosfæriske fenomener som den nordatlantiske oscillasjonen (NAO) og den arktiske oscillasjonen (AO) (Hurrell et al. 2003; Årthun et al. 2019). NAO er det sterkeste klimamønsteret vinterstid i det nordatlantiske området og representerer variasjon i forskjellen i lufttrykk ved havoverflaten mellom lavtrykket i nord (Island) og høytrykket lengre sør (Azorene). NAO er kjent for å påvirke vind og nedbørsmønstre, og varierer på mellomårlig skala (Hurrell et al. 2003). Høy NAO fører vanligvis til høyere vanntemperaturer i Barentshavet, men den sammenhengen har blitt svakere etter 1990-tallet (Johannesen et al. 2012).
Innstrømningen av atlanterhavsvann til Barentshavet er også assosiert med storskala fenomener i havet som den atlantiske multidekadiske oscillasjon (AMO) og oppstrøms sirkulasjonsendringer (AMOC - Atlantic meridional overturning circulation) (Årthun et al. 2019). AMO representerer storskala temperaturvariasjoner i Atlanterhavet som svinger med en periode på 60-70 år (Schlesinger og Ramankutty 1994), mens AMOC karakteriseres av varmt og salt vann som transporteres nordover nær overflaten og avkjøles, for deretter å synke ned og strømme sørover på større dyp. Endringer i AMOC kan også påvirke Barentshavet i fremtiden (jfr. 4.1.6).
Økte temperaturer i hav og luft bidrar til redusert havisdekke, noe som altså gir en selvforsterkende effekt. Mengden atlanterhavsvann som strømmer inn fra Norskehavet er en annen faktor som varierer mellom år og som er viktig for utviklingen i isdekke i Barentshavet (Årthun et al. 2012). Vind, lokale havstrømmer og nedbør påvirker også havisen, men innen så store områder som havisindikatoren (figur 4.12) dekker vil enkelthendelser av lokal art påvirke minimalt over tid, mens regionale værmønstre, naturlig variabilitet og klimaendring kan gi mer fremtredende endringer.
4.1.6 - Fremtidsbilder
Forventet temperaturutvikling i atmosfære og hav Det som skjer i Barentshavet påvirkes av prosesser på større skala. Innstrømningen av relativt varmt og salt atlanterhavsvann til Barentshavet påvirkes som nevnt i 4.1.5 av prosesser på større skala, og Barentshavets beliggenhet er videre helt sentral for varmeutvekslingen med atmosfæren og dypvannsdannelse (Lien og Ådlandsvik 2011) og storstilt thermohalin sirkulasjon. AMOC har blitt svekket siden perioden 1850-1900 (middels sannsynlighet) og forventes med stor sannsynlighet («very likely») å bli svekket ytterligere i løpet av det 21. århundret (IPCC 2019 - in press). Likevel er det knyttet stor usikkerhet til hvordan en fremtidig økning i overflatetemperaturen vil påvirke dypvannsdannelsen i Barentshavet ettersom utviklingen av saltholdigheten og ferskvannsmengden og lagdelingen i nord er svært usikker (Lind et al. 2018; Sandø et al. 2014b).
Modellresultatene presentert her for lufttemperatur er hentet fra den norske jordsystemmodellen NorESM (Bentsen et al. 2013), mens overflatetemperatur i havet er nedskalert fra NorESM med den regionale havmodellen ROMS (Shchepetkin og McWilliams 2005). Scenariet som er brukt for fremtidig utslipp av klimagasser er RCP4.5, som er et av utslippsscenariene FNs klimapanel (IPCC) har arbeidet med. Dette scenariet innebærer en ekstra varmetilførsel på 4,5 W/m 2 til jord-atmosfæresystemet i år 2100 i forhold til førindustriell tid (1765). I RCP4.5 vil klimagasskonsentrasjonene i atmosfæren øke mest frem mot rundt 2060, men stabiliseres ved slutten av århundret. Her har vi sett på fremtidsendringer i to perioder, 2026-2035 og 2060-2069. Det er sammenlignet med klima i perioden 2010-2019.
Basert på modellkjøringen med NorESM forventer en at lufttemperaturen om vinteren vil gå ned med 1-2 grader i det meste av det nordlige og østlige Barentshavet frem mot midten av 2030-årene, noen steder litt mer (figur 4.13). Dette er sammenfallende med områder der vanntemperaturen går ned og iskonsentrasjonen og -tykkelsen går opp. Dette skyldes som nevnt mest sannsynlig naturlig variabilitet, og da spesielt AMO, som altså representerer storskala temperaturvariasjoner i Atlanterhavet som svinger med en periode på 60-70 år. I 2060-årene forventes situasjonen å være motsatt, med en lufttemperaturøkning om vinteren på opp mot 6 grader i nordøst. Om sommeren forventes det en økning på opp til 2 grader, noe mer i den siste perioden enn i den første (ikke vist).
For temperatur i havet forventes endringene om vinteren i den første perioden generelt å være preget av en oppvarming på rundt 0,5 grader i nordvest og en tilsvarende nedkjøling i sentrale deler og vest for Novaya Semlya. I den andre perioden forventes en oppvarming i hele Barentshavet, med unntak av Storfjorden på Svalbard. Den største økningen på 1-1,5 grader forventes i den nordligste halvparten, mens den i sør forventes å ligge på rundt 0,5 grader. Om sommeren forventes temperaturøkningene å være noe mindre (ikke vist).
Figur 4.13. Forventede endringer i atmosfæretemperatur fra NorESM (°C, venstre) og overflatetemperatur i havet fra ROMS (°C, høyre) for mars fra årene 2010-2019 til henholdsvis 2026-2035 (øverst) og 2060-2069 (nederst). Blått indikerer nedkjøling og rødt indikerer oppvarming.
Forventet utvikling for havis FN sitt klimapanel (IPCC) vurderer i sin spesialrapport på hav og kryosfære (is), som ble lansert i september 2019 (Meredith et al. 2019 - in press), historisk utvikling og framskrivinger til år 2100 av utbredelse og tykkelse av havis i Arktis. Vurderingsmaterialet består i hovedsak av er i stor grad en rekke kjøringer av ulike modeller (multi-modell ensemble av simuleringer), samlet under begrepet CMIP5 (Coupled Model Intercomparison Project Phase 5). Rapporten bekrefter tidligere resultater om at utbredelsen av havis i Arktis vil avta videre utover i dette århundret. Mens det er stor sikkerhet (high confidence) bak dette resultatet er det uenighet mellom modellene om når Arktis blir isfritt om sommeren og om hvor lenge den isfrie sesongen vil vare. CMIP5 modellene har ikke god nok oppløsning til å kunne gi holdbare framskrivinger av issituasjonen spesifikt for Barentshavet. Det er håp om at neste generasjons ensemblekjøringer, CMIP6, med bedre oppløsning, som en nå er godt i gang med og som vil bidra inn til IPCC sine neste hovedrapporter, vil takle dette.
En analyse som ble publisert nylig viser at disse ensemblekjøringene gir et mer realistisk estimat av sjøisdekket i september sammenlignet med tidligere CMIP-eksperimenter, gitt et visst utslipp av klimagasser med tilhørende global oppvarming (Notz et al. 2020).
Regionale modeller for Barentshavet og nedskaleringer fra globale modeller gir et bilde av forventet utvikling. Modellkjøringene beskrevet i forrige delkapittel viser forventede endringer i mengde og tykkelse på havis i Barentshavet om man følger utslippsscenariet i RCP4.5 (figur 4.14). Endringene i isutbredelse og -tykkelse fra samme modellsimulering i figur 4.14 gjenspeiler i stor grad endringer i overflatetemperatur som vist i figur 4.13 i de områdene hvor man har sjøis om vinteren. Det innebærer at noe mindre is er forventet i den nordvestre delen og mer i den østre delen vest av Novaya Semlya i den første perioden, og generelt mindre is i den siste perioden. Endringene er hovedsakelig forårsaket av endringer i vanntemperatur og i mindre grad endringer i atmosfæretemperatur, selv om sistnevntes betydning er økende med global oppvarming (Sandø et al. 2014a).
Figur 4.14. Forventede endringer i iskonsentrasjon fra ROMS (deler av hver gridcelle som er dekket av is på en skala fra 0 til 1, venstre) og istykkelse (m, høyre) for mars fra årene 2010-2019 til henholdsvis 2026-2035 (øverst) og 2060-2069 (nederst). Blått indikerer reduksjon i iskonsentrasjon og rødt indikerer økning.
4.2 - Plankton
4.2.1 - Planteplankton
Satellittbaserte estimater av planteplanktonets årlige netto primærproduksjon for Barentshavet i sin helhet viser en oppadgående trend gjennom perioden 1998-2019. Endringene i klima de siste 40 årene, med økende temperatur og minkende utbredelse av havis, har redusert arealene hvor planteplanktonets primærproduksjon begrenses ved lysskygging fra is samt gitt en lenger produktiv sesong. Økende årlig primærproduksjon har vært særlig utpreget i den nordøstlige delen av Barentshavet.
Planteplankton står for det meste av primærproduksjonen (dvs. planteveksten) i Barentshavet og utgjør derfor hovedgrunnlaget for det øvrige økosystemet. For hele Barentshavet bidrar isalger med mindre enn tre prosent (Wassmann et al. 2006), men i isfylte farvann kan primærproduksjonen hos isalger være i størrelsesorden 20 % (Jin et al. 2012, Fernandez-Mendez et al. 2015). Siden isalger ikke overvåkes i Barentshavet, fokuserer denne rapporten på planteplanktonets produksjon.
Biomasse av planteplankton måles fra satellitt som mengde klorofyll a over vekstsesongen, og på dette grunnlaget modelleres primærproduksjonen. Disse estimatene av primærproduksjon må tolkes med forsiktighet, fordi en rekke forhold kan gjøre det vanskelig å måle klorofyll a fra satellitt. Dette inkluderer skydekke, oppblomstringer i dypere lag av overflatevann og under is, oppløste sedimenter, farget organisk materiale samt mulige svakheter i modellene som brukes til å estimere produksjonen (Ardyna et al. 2013; Arrigo et al. 2012; Arrigo og van Dijken 2015; Assmy et al. 2017; Engelsen et al. 2002; Engelsen et al. 2004; Joint og Groom 2000). Slik estimering fanger heller ikke opp variasjon i artssammensetning av planteplanktonet (Assmy et al. 2017), som kan ha stor betydning for beitende organismer som dyreplankton og derved påvirke også høyere nivåer i økosystemet. Satellittbaserte estimater av årlig primærproduksjon hos planteplanktonet i Barentshavet har indikert en stigende trend for de siste tiårene med betydelig mellomårsvariasjon (Arrigo og van Dijken 2015; Dalpadado et al. 2014). I løpet av perioden 1998-2017 har estimert årlig netto primærproduksjon for hele Barentshavet mer enn fordoblet seg, fra ca. 40 til over 100 Tg C, som tilsvarer en årlig økning på 3-6% (Dalpadado et al. in press). En oppadgående trend ses også når tidsserien forlenges til 2019 (Knut Yngve Børhsiem, pers. komm.). Forklaringen på den økte årlige primærproduksjonen i Barentshavet er høyere vanntemperatur og dermed redusert isdekke og lenger sesongperiode med åpent vann. Større arealer med åpent vann gjør at mer sollys når øvre vannlag, og lengre isfrie perioder gir utvidet produksjonssesong (Arrigo og van Dijken 2015; Dalpadado et al. 2014; Renaut et al. 2018). Dette henger sammen med «atlantifisering» av Barentshavet (Polyakov et al. 2017). Det er ikke klart om økning i primærproduksjonen er basert på ny eller regenerert produksjon; økning i sistnevnte vil ikke bidra til økning i høstbare ressurser. Satellittmålinger viser også at økningen av antall dager med åpent vann om høsten (pga. senere isdannelse om høsten) har bidratt til en høstoppblomstring, særlig i nordlige deler av Barentshavet (Ardyna et al. 2014). Prediksjoner fra modellering av et fremtidig klimascenario for Barentshavet har vist en gjennomsnittlig økning i primærproduksjonen på 36 % for årene 2046-2064 (Skaret et al. 2014). I tillegg viste disse resultatene at den største økningen forventes å skje i nordlige og østlige deler av Barentshavet, der isen er betydelig redusert.
Flere studier har i de siste årene vist at oppblomstringer av planteplankton under isen har blitt mer vanlige i hele det arktiske kontinentalsokkelområdet (Arrigo et al. 2012; Arrigo et al. 2014; Mundy et al. 2009; Mundy et al. 2014; Oziel et al. 2019), og dette gjelder også for Barentshavet (Assmy et al. 2017; Johnsen et al. 2018; Olsen et al. 2019). Modelleringsstudier viser at oppblomstringer under isen vil øke grunnet bedre lysforhold som skyldes tynnere is med mer smeltedammer (Horvat et al. 2017). Siden oppblomstringer under isen ikke oppdages av satellittmålinger, er det sannsynlig at økningen av primærproduksjon de siste årene er enda høyere enn det som er anslått på grunnlag av de isfrie arealene (Renaut et al. 2018). I tillegg har planteplanktonvekst i lag mellom isen og snøen, såkalte «snø-infiltrasjonssamfunn», sannsynligvis øket de siste årene pga. av endrede forhold mellom istykkelse og snøtykkelse (Fernández-Méndez et al. 2018).
Konsentrasjon av næringssaltet silikat i det innstrømmende atlanterhavsvannet har vist en nedgang på ca 30 % gjennom de siste 25 årene (Hatun et al. 2017; Rey 2012). Silikat er et viktig byggestoff for kiselalger (diatomeer), og nedgangen i dette næringssaltet kan motvirke økningen i primærproduksjon og endringer i artssammensetning av planteplankton. Mangel på silikat kan føre til at oppblomstringen av kiselalger forkortes og til at bidraget til våroppblomstringen av Phaeocystis, en mindre spiselig algeart, økes (Krause et al. 2019). Dette kan påvirke næringsopptaket til kopepoder (Calanus spp.) og dermed det pelagiske næringsnettet.
I løpet av de siste 20 årene har det vært flere observasjoner av sørlige arter av planteplankton i Barentshavet. I forbindelse med atlantifiseringen har forekomst og oppblomstringer av den atlantiske kalkalgen Emiliania huxleyi og picocyanobakterien Synechococcus spredd seg nordøstover i Barentshavet (Hegseth og Sundfjord 2008; Oziel et al. 2019; Paulsen et al. 2016) .
4.2.2 - Dyreplankton
Det mellomstore dyreplanktonet sin biomasse om høsten i den norske delen av Barentshavet har vært relativt stabil siden 1995. Parallelt med oppvarmingen i Barentshavet og reduksjonen av arealet dekket av arktiske vannmasser og av havis, pågår det en «atlantifisering» av det biologiske samfunnet. Mer varmekjære, atlantiske arter av dyreplankton har blitt mer vanlige, mens arktiske arter går usikre tider i møte. For eksempel er den arktiske sokkelarten Calanus glacialis under press, mest sannsynlig grunnet reduserte arealer dekket av kalde, arktiske vannmasser og havis. Den atlantiske og mer varmekjære arten C. finmarchicus har stort sett vært tallrik ved inngangen til Barentshavet de siste ti årene. Indekser for krill i Barentshavet har vært stigende de siste tiårene.
Hoppekreps er en gruppe av mellomstore dyreplankton hvor det er observert endringer i utbredelse og mengdeforhold av ulike arter. Hoppekreps utgjør en stor del av dyreplanktonet i Barentshavet, og biomassen domineres av Calanus -artene (Aarflot et al. 2017; Dalpadado et al. 2012). Tre arter forekommer i store deler av Barentshavet: Calanus finmarchicus (raudåte), Calanusglacialis (ishavsåte), og Calanus hyperboreus (feitåte). Mens C. finmarchicus dominerer i de atlantiske vannmassene, er C. glacialis og C. hyperboreus i første rekke knyttet til kaldere arktiske vannmasser (Falk-Petersen et al. 2009). Det er også forskjeller mellom de sistnevnte, siden C. glacialis er en sokkel-art, mens C. hyperboreus er en oseanisk art med hovedproduksjonsområde i det dype Grønlandlandshavet (Hirche 1997). Generelt observeres en økning av den atlantiske arten C. finmarchicus fra atlantisk innstrømming til Polhavet (Basedow et al. 2018; Hop et al. 2019).
Sammensetningen av Calanus-artene langs Fugløya–Bjørnøya (FB) snittet ved den sørvestlige inngangen til Barentshavet viser at forekomsten av C. finmarchicus var relativt stabil i perioden 1995-2009. I 2010 ble det registrert en meget høy forekomst (årsgjennomsnitt), og med unntak av en uvanlig lav mengde i 2013, har arten vært tallrik fra 2010 og frem til og med 2019 (ICES 2019c; ICES 2020). Samtidig har den årlige forekomsten av C. glacialis vært relativt lav siden slutten av nittitallet, og det synes å ha vært en nedadgående trend i bestanden av denne arten, med noe variasjon mellom år (Dalpadado et al. 2012; ICES 2019c). I Kongsfjorden, på vestkysten av Spitsbergen, varierer relativ sammensetning av atlantisk C. finmarchicus og arktisk C. glacialis stort sett med mengden av innstrømning av atlanterhavsvann. I årene 1996-2016 har mengden C. finmarchicus og total biomasse av dyrepankton økt, mens mengden av C. glacialis totalt ikke har minket, men kun avtatt relativt til mengden av atlantiske arter (Hop et al. 2019). Det har gjennomgående vært registrert relativt lave årlige antall av C. hyperboreus langs FB-snittet. Den mer varmekjære, sørlige arten Calanus helgolandicus observeres ofte langs FB-snittet (Dalpadado et al. 2012). Denne arten har økt i antall i Nordsjøen de siste tiårene, og forekommer også i Norskehavet utenfor kysten av Norge (Dupont et al. 2017), og mest om høsten (upubliserte data fra Continuous Plankton Recorder Survey). Med høyere vanntemperatur er det forventet at arten vil spre seg med havstrømmene inn i mer nordlige havområder. De største mengdene av C. helgolandicus langs FB-snittet forekommer i perioden desember-februar, men overstiger da sjelden noen få hundre individer per kvadratmeter av de eldste stadiene. Om våren og sommeren er den praktisk talt fraværende. Havforskningsinstituttets tidsserie for FB-snittet indikerer ingen økning i mengden av denne arten ved inngangen til Barentshavet i senere år.
Parallelt med at områdene med kalde arktiske vannmasser har minket siden rundt år 2000, har mengden av typiske kaldtvannsarter i Barentshavet minket. Et eksempel er forekomst av pelagiske amfipoder rundt Svalbard. Tidligere studier har vist at mengdene av den arktiske arten Themisto libellula (svartåte), som er et viktig byttedyr for polartorsk og sjøfugl og er avhengig av kalde (arktiske) vannmasser (ICES 2019c; Stige et al. 2019), har avtatt i det nordlige Barentshavet (Dalpadado et al. 2012). Denne nedgangen har foregått i perioden med oppvarming av havområdet. Parallelt med at temperaturen siden 2016 i varierende grad har gått ned igjen, har en sett en økning av pelagiske amfipoder igjen, spesielt i 2019 og nord og øst av Svalbard (ICES 2020). Mens mengden av svartåte avtok frem til 2016, har flere studier vist at mengden av den mer varmekjære amfipoden Themisto abyssorum har økt, både i Barentshavet, Framstredet og i Kongsfjorden (Dalpadado et al. 2016; Hop et al. 2019; Soltwedel et al. 2016). I tillegg er det observert økende antall av sørlige krillarter som Meganyctiphanes norvegica og Nematoscelis megalops i Barentshavet og i fjorder på Svalbard (Eriksen et al. 2017; Hop et al. 2019). Bifangster fra norsk-russiske høstundersøkelser av 0-gruppe fisk ved tråling i de øvre 60 meterne over det meste av Barentshavet har vist en økende trend i mengden av krill gjennom de siste tiårene (Eriksen et al. 2017). Tilsvarende har russisk langtidsovervåkning vist en økende trend i en krill-indeks basert på planktonnett festet til bunntrål. Den stigende trenden i de russiske dataene har blitt forklart ved økt adveksjon og mengde av Thysanoessa inermis og Meganyctiphanes norvegica med oppvarmingen siden 1980-tallet (Dolgov et al. 2013; Orlova et al. 2015; Zhukova et al. 2009). En økning av boreale krillarter har også vært oppdaget i nettprøver og krykkjediett i Kongsfjorden (Hop et al. 2019; Vihtakari et al. 2018).
Årsaken til at arktiske arter er i tilbakegang i Barentshavet mens tettheten av mer varmekjære arter øker, er sannsynligvis knyttet til økt vanntemperatur. En reduksjon av arealene dekket av kalde, arktiske vannmasser, og mindre havis, er ugunstig for arktiske kaldtvannsarter av dyreplankton. Dette medfører at mer varmekjære, boreale arter delvis eller helt vil kunne erstatte de arktiske artene. Mengden av C. finmarchicus i Barentshavet påvirkes også ved tilførsel med innstrømmende vannmasser fra sørligere breddegrader (Torgersen og Huse 2005). Generelt er arktiske dyreplanktonarter er ofte mer fettrike enn de mer varmekjære artene (Falk-Petersen et al. 2009). Et skifte i retning av mer varmekjære og mindre arter kan derfor få konsekvenser for artene som beiter på dyreplanktonet, og dermed også for økosystemet i det nordlige Barentshavet i sin helhet. Det ser særlig ut til at arktiske planktonspisere er avhengige av en fettrik diett og byttedyr, f.eks. C. glacialis , som er av en viss størrelse (Balazy et al. 2018). Disse artene kan derfor være utsatt for den typen endringer vi ser i dyreplanktonsamfunnet i Barentshavet (Hovinen et al. 2014). Likevel betyr ikke et skifte til mindre fettrike men mer tallrike dyreplanktonarter nødvendigvis at mattilførselen til dyr lenger oppe i næringskjeden blir mindre effektiv, men det kan få konsekvenser for visuelle predatorer som beiter på visse størrelsesfraksjoner av byttedyr. Renaud et al. (2018) påpeker at de aktuelle artene overlapper sterkt mht. størrelsesfordeling, og at fettmengden er størrelses- fremfor artsavhengig. Basert på en matematisk modell vurderer forfatterne at økt temperatur og kortere generasjonstid hos de atlantiske artene kan mer enn kompensere for at individene er mindre og har lavere fettreserver, slik at energitransporten i økosystemet opprettholdes (Renaud et al. 2018).
4.3 - Benthos
Bunndyrsamfunn er dynamiske og viser raske endringer ved endring i utbredelse av nye arter som snøkrabbe og kongekrabbe, samt endringer i vandring av bentiske arter. Endringer i samfunnsstruktur og -sammensetning gjenspeiler naturlige og antropogene faktorer. Biomasse av megabenthos i Barentshavet har økt de siste fjorten årene, og svingninger i biomasse gjenspeiler temperatursvingninger med en tidsforsinkelse på syv år. Snøkrabbe har de siste årene spredd seg til hele Barentshavet, og er nå observert også på vestsiden av Svalbard.
Det finnes mer enn 3000 arter av bentiske virvelløse dyr som er registrert i Barentshavet (Sirenko 2001). Her vil vi kun fokusere på megabenthos (>2 cm) som er samlet inn med trål og registrert (arter, antall og biomasse) under økosystemtoktet. Dette inkluderer hovedsakelig større bunndyr med lang levetid. Økosystemtoktets bunndyrprøvetaking begynte i 2005 på russisk side og 2009 på norsk side, og har nå en relativt kort tidsserie sammenlignet med plankton og fisk. Derfor må tolkningen av langsiktige trender for megabenthos-data gjøres med aktsomhet, og artsbestemmelse blir stadig bedre og derfor mer detaljert over tid.
Beskrivelse av megafauna Biomassefordelingen av hoved-bunndyrsgruppene i Barentshavet viser at i vest domineres biomassen av svamper (hovedsakelig arter innenfor familien Geodiidae), mens i den sentrale og nordlige delen av havområdet dominerer pigghuder (hovedsakelig slangestjerner) (figur 4.15). I nordøst dominerer nesledyrene, hvor en stor andel av biomassen består av arten dyphavssjøfjær (Umbellula encrinus), sammen med pigghuder. I sørøst dominerer krepsdyr sammen med pigghuder.
Figur 4.15. Biomassefordelingen av hovedgruppene av bunndyr per areal med integrert gjennomsnitt for perioden 2012-2017. Grå Annelida/Leddormer, rød Arthropoda/Leddyr blå Bryozoa/Mosdyr, mørkegrønn Chordata/Ryggstrengdyr, grønn Cnidaria/Nesledyr, gul Echinodermata/Pigghuder, lilla Mollusca/Bløtdyr og rosa Porifera/Svamp.
Status for megabenthos i 2018 og 2019 Resultatene fra 2019 viser høy biodiversitet med stort antall arter (figur 4.16 A) i den vestlige delen av Barentshavet (norsk sone) og noe lavere biodiversitet i øst (russisk sone). Den generelle fordelingen av forekomst og biomasse for 2019 ligner fordelingen i 2018. Forekomst (figur 4.16 B) viser et stort antall individer i den nordøstlige og midtre delen av havområdet, inkludert kystvannområdet til Novaya Zemlya. Men i sør og langs skråningen i vest er forekomsten lav. Biomassen (figur 4.16 C) er høy i nordøst, i sørøst (nær Novaya Zemlya-kysten, Kanin-halvøya og på Nord-Kanin-banken), og spesielt i sørvest.
Figur 4.16. Megabenthos-utbredelse fra økosystemtoktene i 2018 og 2019. A) antall arter totalt B) antall individer per nautisk mil C) biomasse per nautisk mil.
Årlige svingninger av den gjennomsnittlige biomassen for megabenthos For å estimere den langsiktige dynamikken i megabenthos ble de årlige endringene av den gjennomsnittlige biomassen beregnet for hele Barentshavet (figur 4.17 B). Havområdet ble delt opp i fire soner; nordøst-, nordvest-, sørøst- og sørvestdelen av Barentshavet (figur 4.17 C, D) (ICES 2018; ICES 2019c).
I 2018 var den østlige delen av Barentshavet bare delvis dekket av Økosystemtoktet (figur 4.16). Dette gjorde det umulig å estimere den årlige biomasse-trenden i den østlige delen av Barentshavet, samt estimeringen for “hele Barentshavet” (figur 4.17 D, B). Data fra den vestlige delen av Barentshavet viser moderat økning av biomasse for megabenthos i 2019 sammenlignet med 2018 (figur 4.17 C).
De siste fjorten årene med overvåkning viser en moderat trend med økende biomasse av megabenthos (figur 4.17 B). Samtidig viser svingninger i biomasse i løpet av året positiv korrelasjon med vanntemperaturen på Kola-sektoren, med en tidsforsinkelse på syv år, unntatt i nordvestlig sone (figur 4.17).
Tilsvarende respons på endring av miljøforhold har også blitt dokumentert for makrobenthos (>1 mm) i Barentshavet, med en tidsforsinkelse på omtrent fire år (Denisenko 2013; Lyubina et al. 2012; Lyubina et al. 2016). Forskjellen i tidsforsinkelsen (dvs. fra endring i temperatur til en respons fra benthos) mellom mer opportunistisk makrobenthos (grabbprøver) og mer langtidslevende megabenthos (trålbifangst) er et resultat av ulike størrelser og livslengder til gruppene av bentiske organismer.
Figur 4. 17. Variasjon over tid av årlig gjennomsnittstemperatur i dybdelaget 0-200 m for 3-7 stasjoner i Kola-området (A) (http://www.pinro.ru) og den årlige gjennomsnittlige svingningen i biomasse av megabenthos i hele Barentshavet (B), vestlig del (C) og østlig del (D). Hele Barentshavet er innenfor 68-80 °N, 15-62 °Ø, "NØ" - nord-østlig sone (74-80 °N, 40-62 °Ø), "NV" - nordvestlig sone (74-80 °N, 15-40 °Ø), “SØ” - sørøstlig sone (68-74 °N, 40-62 °Ø), “SV” - sørvestlig sone (68-74 °N, 15-40 °Ø). Biomasse av Pandalus borealis og alle fangster over ett tonn er ekskludert. Røde sirkler i graf A viser kalde år som kan ha forårsaket nedgang i biomasse i årene 2010, 2015 og sannsynligvis 2018 (er vist med grå piler). Den stiplede linjen i graf B viser en lineær trend.
Kongekrabbe De årlige toktene på kongekrabbe i og utenfor kvoteregulert område gir oss kunnskap om den relative bestandsutviklingen i kvoteområdet og status på spredningen vest for kvoteområdet (vest for Nordkapp). I de siste årene er det kun små endringer i krabbebestanden i kvoteområdet. Vest for Nordkapp dukker det stadig opp enkeltkrabber flere steder i Troms, og Balsfjorden ser ut til å bli et nytt «hot spot» for kongekrabbe. Her ble det landet mer enn syv tonn krabbe fra det frie fisket i 2019. I og med at krabben ser ut til å formere seg her forventer en at bestanden lokalt i Balsfjorden vil øke i tiden som kommer selv om den ble noe nedfisket i 2019. Utbredelsen nord for det kvoteregulerte området ser fortsatt ut til å være minimal.
Dypvannsreke Den totale mengden av reke har vært stabil på et relativt høyt nivå siden 2005.
Snøkrabbe Bestanden av snøkrabbe på norsk sokkel har økt betydelig siden 2010, men de største forekomstene er fortsatt i russisk sone. I tillegg til den kontinuerlige spredningen vest- og nordover i sentrale deler av Barentshavet, har det også blitt gjort enkeltfunn nordvest og sørvest av Spitsbergen. Det antas derfor at snøkrabben har kolonisert store deler av potensielt leveområde, men tettheten av krabbe i disse områdene er for lav til at de er av kommersiell interesse. Fiskeriet startet opp i 2012, og innsatsen og landingene har økt siden den gangen. Fra og med 2017 ble det innført en total kvote per år på hannkrabber større enn 10 cm skallbredde. Etter at fiskeriet ble utestengt fra Smutthullet, foregår fisket nå hovedsakelig i områdene rundt Sentralbanken i Fiskevernsonen ved Svalbard. Det er forventet at spredning og økning av tettheten vil fortsette og at den vil begrenses av bunntemperaturer og dybde.
Biotoper i Barentshavet I arbeidet med forvaltningsplanene er det nyttig å kartlegge forekomst og utbredelse av ulike biotoper i havområdene. Det kan for eksempel danne grunnlag for å identifisere områder som er sårbare for ulike typer påvirkning samt truede naturtyper, og kan gi bidrag til økt forståelse av dynamikk og sammenhenger i økosystemene. Innen MAREANO har det i løpet av høsten 2019 blitt identifisert og kartlagt 27 biotoper i Barentshavet ved hjelp av statistisk analyse av resultater fra videodata fra 757 havbunnslokaliteter (Buhl-Mortensen et al. 2020). Disse dataene dekker en mindre del av Barentshavet til sammen. For å lage kart som også dekker områder som ikke er undersøkt, ble det først identifisert en rekke miljøvariable (for eksempel dybde, terrengtyper, temperatur, saltholdighet og strømningsforhold) som er knyttet relativt godt til (dvs. korrelert med) de kartlagte biotoptypene. Deretter ble dette brukt til å modellere forekomsten av biotoper utenfor de kartlagte områdene (Buhl-Mortensen et al. 2020).
Figuren under viser et kart med fordelingen av biotoptyper i Barentshavet basert på slike modeller. Hvilke typer vannmasser som er i et område er viktig for forekomst av biotoptypene. Av de 27 biotopene er 12 biotoper preget av varmt eller raskt strømmende vann (atlantisk vann og arktisk intermediært vann), mens 15 biotoper forekommer i kaldt eller langsomt strømmende vann (arktisk vann og norskehavs-dypvann). I fremtiden er det aktuelt å bruke samme type modellering til å genere biotopkart for hele området som omfattes av kartleggingen i MAREANO, det vil si Norskehavet og Barentshavet.
Figur 4.18. Kart som viser modellert fordeling av de 27 generelle biotopene over hele kartleggingsområdet. Fargene er gruppert i henhold til vannmasse (grønne = Norskehavs-dypvann, lilla = Arktisk intermediært vann, gule / brune = Atlantisk vann, blå = Arktisk vann). Den generelle plasseringen av polarfronten er indikert med en blå linje.
4.4 - Fisk
Lodden er på et lavmål mens mengden ungsild er relativ stabil. Det er fremdeles store bestander av torsk og hyse i Barentshavet. Utbredelsen av torsken dekker nå store deler av Barentshavet, også lenger mot nord og nordøst enn tidligere. Det er ikke klart om det er den store bestanden som gjør at den har trukket nordover, eller om dette også er koblet til perioden med økende temperatur i vannet. Ellers er det ingen større endringer i fiskesamfunnet.
Teksten om fiskebestander er basert på rapporter fra arbeidsgruppene i ICES for henholdsvis for Barentshavet (WGIBAR) (ICES 2020) og for arktiske fiskebestander (AFWG) (ICES 2019a) som kildereferanser.
Ungsild De siste årene har mengden sild i Barentshavet vært i nedgang, men ser nå ut til å ha stabilisert seg.
Figur 4.19. Grafen viser fordeling mellom årsklassene av ungsild i Barentshavet, vist som biomasseindeks. I 2018 ble det ikke beregnet sildeindeks på grunn av manglende toktdekning.
Den siste store ungsild-årsklassen i Barentshavet var i 2004. Denne er nå utvandret, og det har vært lite ungsild igjen i Barentshavet de siste årene. I 2013 det ble det imidlertid observert relativt mange nullåringer av sild i Barentshavet. Denne årsklassen er nå stort sett ute av Barentshavet.
I 2017 ble det observert mange ettåringer. Biomasseindeksen for årsklassen fra 2016 var da den høyeste siden 2004-årsklassen ble målt som ettåringer i 2005. Høy biomasseindeks for 2016-årsklassen ble også bekreftet i 2019, - nå som et relativt høyt estimat av treåringer. På grunn av manglende toktdekning ble det ikke beregnet sildeindeks i 2018.
Lodde Bestanden av lodde gjennomgår store naturlige svingninger, og den modne bestanden har fluktuert mellom under 200 000 tonn og over tre millioner tonn siden 1983. Den er for tiden på et lavt nivå.
Figur 4.20. Beregnede størrelser av umoden bestand og moden/gytebestand av lodde i Barentshavet i millioner tonn. Til sammen angir dette totalbestand.
Bestanden ble i 2015 regnet for å være kollapset, det fjerde bestandssammenbruddet siden 1983. I 2017 viste målingene at bestanden igjen var stor nok til at en kunne åpne for et fiskeri, og vinteren 2018 ble det fisket 195 000 tonn lodde. Resultatene fra 2018 viste en liten bestandsnedgang, mens resultatene fra 2019 indikerte at bestanden var under nivået fra 2015 og fisket ble stoppet. Rekrutteringa har vært svak de siste seks årene. Den Norsk-Russiske Fiskerikommisjon, som bestemmer endelig fiskekvote på loddebestanden, vedtok anbefaling om fortsatt null fiske i 2020.
Kolmule I 2008 ble det registrert en markant reduksjon i kolmulebestanden i Barentshavet. Etter dette har den vært variabel på et lavt nivå, med en topp i 2016.
Kolmulen økte trolig i utbredelse og mengde i Barentshavet fra 1996 til 2004. Etter dette har det vært en klar nedgang. Fra 2008 har mengde kolmule variert på et lavt nivå, men med en topp i 2016 (figur 4.21 og figur 4.22).
Figur 4.21. Akustisk biomasseindeks for kolmule i Barentshavet. Viser beregnet fordeling mellom ulike aldersklasser i 1000 tonn.
Mesteparten av kolmulen i Barentshavet kommer inn fra Norskehavet når det strømmer varmt vann inn i Barentshavet fra sørvest. Derfor er det sannsynlig at det er en sterk sammenheng mellom mengden kolmule i Barentshavet, mengden kolmule i Norskehavet og innstrømming av varmt vann til Barentshavet (Belikov et al. 2011).
Figur 4.22. Grafen viser beregnet totalbestand av kolmule, i 1000 tonn. I beregningene av kolmulebestandens størrelse, brukes metoden SAM (State-space Assessent Model), som er en ny standardmetode etter hvert brukt på flere bestander i ICES.
Nordøstarktisk torsk Gytebestanden av nordøstarktisk torsk er i dag på et høyt nivå. Den nordøstarktiske torsken er den største torskebestanden i verden.
Figur 4.23. Beregnet totalbestand av nordøstarktisk torsk, i 1000 tonn.
Både totalbestanden og gytebestanden har vokst siden 2006 og nådde toppen i 2013. Etter dette har det vært en nedgang, men både totalbestand og gytebestand er fortsatt godt over langtidsgjennomsnittet for 1946-2018 (figur 4.23). Gytebestanden i 2019 er beregnet til 1,5 millioner tonn. Dette er langt over den tiltaksgrensen som er satt av fiskeriforvaltningen.
Gytebestandens størrelse har vært over føre-var-nivået siden 2002. Fiskedødeligheten ble redusert betydelig fra godt over kritisk nivå i 1999 til under føre-var-nivået fra og med 2008. I de siste årene har den igjen økt noe og var i 2017 på føre-var-nivået.
Lavt fiskepress har bidratt til å holde bestanden på et høyt nivå, og i tillegg har god tilgang på mat og relativt høye temperaturer bidratt til at vi i mange år nå har hatt svært gode bestander av både torsk og hyse i Barentshavet. Temperaturøkning har gitt fisken et større leveområde og bedre tilgang til mat.
Blåkveite Bekymringer for bestandssituasjonen gjorde at fisket etter blåkveite ble strengt regulert i 1992. Etter det steg bestanden frem til 2014. De siste årene har vi sett en svak nedgang i bestanden.
Den fangstbare mengden blåkveite, definert som fisk på 45 cm eller lengre, har i hovedsak steget jevnt fra 1992 og frem til 2014. Dette tyder på at fangstraten, som har vært høvelig stabil siden 1992, ikke har vært for høy i denne perioden. De siste årene har vi sett en svakt nedadgående trend i biomasse. Fangsttrykket har økt litt de siste årene og modellkjøringer viser at dagens fangsttrykk vil redusere bestanden noe, men ikke til under beregnet føre-var nivå på 500 000 tonn de nærmeste årene.
Blåkveitas videre bestandsutvikling avhenger også av rekrutteringen som er variabel. En god årsklasse vil ha stor effekt på blåkveitas bestandsutvikling. Dagens bestandsvurdering gjøres ved hjelp av en lengdebasert modell. Uten bruk av alder er det vanskelig å angi et nøyaktig tidspunkt for sterke årsklasser og nye data kan endre tidligere rekrutteringsestimater. Siste rekruteringsestimater indikerer relativt gode årsklasser rundt 2010 og 2014.
Vanlig uer Vanlig uer er klassifisert som en truet art. Bestanden er lav og fortsetter å synke. De senere årene er det satt i gang tiltak for å bedre situasjonen.
Data fra Havforskningsinstituttets tokt og trålfiskeriene viser en klar reduksjon i bestanden av vanlig uer, og bestanden er nå på det laveste nivået som noen gang har vært målt (figur 4.24). Bestanden har hatt liten rekruttering siden sent på 1990-tallet. Det internasjonale havforskningsrådet (ICES) vurderer bestanden til å ha redusert reproduksjonsevne og at den nå er på et historisk lavmål. Bedre rekruttering i enkelte årsklasser etter 2003 førte til en økning i umoden bestand, mens totalbestanden målt i tonn minket fordi totalvekt av moden uer i fangst ble større enn bidrag av rekrutter til biomassen. Det er også en viss risiko for at noe av den registrerte yngelen tilhører den større snabeluerbestanden, siden artene er like av utseende på yngelstadiet.
Gytebestanden er svært svak og minker fortsatt. Hvis fangstene opprettholdes på siste års nivå (5000 tonn årlig) og rekrutteringen er på nivå med snittet for årene 2001-2011, indikerer modellen som brukes at bestanden vil kollapse i nær framtid. Derfor er det, med unntak av et svært begrenset fiske med juksa, innført forbud mot alt direkte fiske etter vanlig uer. I tillegg er det innført reguleringer for å hindre uønsket bifangst av uer i andre fiskerier.
Figur 4.24. Beregnede størrelser av moden og umoden bestand av vanlig uer i Barentshavet og Norskehavet. Til sammen angir dette totalbestand.
Snabeluer Bestanden av snabeluer har en positiv utvikling. Rekrutteringen til bestanden var svak fra 1996 til 2004, men har blitt klart styrket etter dette (figur 4.25).
Ifølge Det internasjonale havforskningsrådet (ICES) har snabeluerbestanden blitt restituert til et bærekraftig reproduksjonsnivå. Rekrutteringen var svak fra 1996 til 2004, men har etter dette blitt klart styrket. Gytebestanden forventes likevel fortsatt å minke i noen år. Dette skyldes både rekrutteringssvikten fra 1996 til 2004 og at snabelueren ofte først blir kjønnsmoden ved 12 års-alderen.
I gjenoppbyggingsfasen av snabeluerbestanden, som varte fram til 2014, var snabeluer bare tillatt å fiske som uunngåelig bifangst i norske havområder. Fra og med 2014 har man igjen kunnet fiske direkte etter denne arten innenfor definerte områder. ICES vedtok i 2014 at det kommersielle fiskeriet kan ta ut opp til 30 000 tonn i 2015, 2016 og 2017 og 32 658 tonn i 2018 (inkludert bifangst og utkast).
Figur 4.25. Beregnede størrelser av moden og umoden bestand av snabeluer i Barentshavet og Norskehavet. Til sammen angir dette totalbestand.
4.5 - Sjøfugl og sjøpattedyr
4.5.1 - Sjøfugl
De fleste sjøfuglindikatorene viser en nedgang i hekkebestandene, både i de siste ti årene og samlet over tidsperioden de har vært overvåket. Dette gjelder både krykkje, lomvi og lunde langs fastlandskysten samt polarlomvi i hele området. Mattilgang i og utenfor hekketida er viktig, og det er sannsynlig at de negative endringene for de fleste sjøfuglartene skyldes redusert næringstilgang. Det er imidlertid vanskelig å konkludere med hva endringene i næringstilgangen skyldes, men sekundæreffekter av klimarelaterte endringer, lavere produksjon av byttedyr eller uttak av fiskeressurser har vært foreslått.
Lomvi Tilstanden for den nordnorske bestanden av lomvi er svært alvorlig, og det kan være et tidsspørsmål før arten forsvinner som hekkefugl i mange fuglefjell langs norskekysten (Erikstad et al. 2007). Dette gjelder spesielt for koloniene på Vedøy og Hjelmsøy der hekkebestandene i 2019 var 99 prosent lavere enn hva de var ved begynnelsen av 1980-tallet da overvåkingen startet. Lomvibestandene på Hornøya og Bjørnøya er i økning, og det samme er bestanden av lomvi på Hjelmsøya som hekker skjult i steinur. Både lomvi og krykkje som hekker langs fastlandskysten er sårbar for forstyrrelser fra en økende havørnbestand (Hipfner et al. 2012). I noen lomvikolonier observerer man at fuglene i økende grad hekker inne i steinurer der de er mer beskyttet for denne forstyrrelsen.
Polarlomvi Polarlomvibestanden på Hjelmsøya er i praksis forsvunnet, mens bestandene på Bjørnøya og vestkysten av Spitsbergen er halvert siden slutten av 1990-tallet. Nedgangen på Svalbard ser ut til å henge sammen med en langsiktig oppvarming av vannmassene i polarlomviens vinterområder rundt Island (den «subatlantiske gyre»). Denne oppvarmingen fører sannsynligvis til endret/redusert forekomst av byttedyrene polarlomvien er avhengig av, med påfølgende redusert overlevelse (Descamps et al. 2013; Fluhr et al. 2017).
Krykkje Alle krykkjekoloniene langs fastlandet, med unntak av Anda i Vesterålen, er redusert med 84-98 prosent siden tidlig på 1980-tallet, mens bestandene på Bjørnøya og Spitsbergen har holdt seg stabile eller kun vist en svak negativ endring de siste 10 årene. Krykkja henter næringen sin i havoverflaten, og er følgelig avhengig av at byttedyr er tilgjengelig i vannmassenes øvre lag. Studier tyder på at den observerte tilbakegangen i hekkebestanden av krykkje er relatert til næringsforholdene. For eksempel viste Reiertsen et al. (2014) at voksenoverlevelsen hos krykkjer på Hornøya var relatert til tilgangen på lodde i hekketiden og en nakensnegl (Thecosomata spp.) i overvintringsområdene ved Grand Banks utenfor Newfoundland.
Lunde Lunde kan dykke dypt, og kan derfor fange byttedyr (sild, lodde og tobis) som står langt nede i vannmassene. Lundebestanden på Røst (Hernyken) har vist seg å være avhengig av en god rekruttering av sildelarver som driver forbi på rett tidspunkt for å ha en vellykket hekkesesong (Anker-Nilssen 1992; Cury et al. 2011). Selv om bestanden av voksen sild nå er stor, er det ikke gitt at gytesesongene treffer til rett tid eller at tilgang på sildelarver er tilsvarende. De ti siste årene, 2009–2019, har således vært svært dårlige, med mer eller mindre fullstendig hekkesvikt for lundene på Røst som resultat. Lundene på Anda har en god tilgang på tobis fra en lokal bestand og kan supplere med dette i år med liten tilgang på sild. Hekkebestanden her er likevel redusert siden begynnelsen av 1980-tallet. Hekkebestanden på Gjesvær har holdt seg stabil, mens den har vært svakt negativ på Hornøya.
Romlig fordeling av sjøfugl Fra økosystemtoktet i september finner vi at utredelsen til flere sjøfuglarter har blitt forskjøvet nordover de siste 10 årene. Forskyvningen har vært spesiell stor for polarmåke (500 km), krykkje (400 km) og havhest (300 km). Svartbak, alkekonge og polarlomvi viser også en nordlig forskyvning på rundt 150 km. Kun lunde, lomvi og gråmåke viser ingen signifikante forskyvninger i nord-sør retning (figur 4.26).
Figur 4.26. Tyngdepunkt i fordelingen av sjøfugl i den vestlige delen av Barentshavet i september (nordlig retning) fra 2009 til 2019. Øverst er overflatebeitende fugl, nederst er alkefuglene. Stjerner angir signifikante positive trender (* 0.05, ** 0.01). Plassering av Ny Ålesund, Bjørnøya og Hammerfest i nordlig retning er vist med stiplede linjer. Kilde: Norsk institutt for naturforskning (NINA).
Det er stor variasjon i artenes tallrikhet fra år til år, noe som primært reflekterer vandringer av fugl inn og ut av Barentshavet. Forekomsten av havhest, krykkje og gråmåke har minket i det vestlige Barentshavet de siste ti årene. For de andre artene finner vi ingen signifikante trender. Havsule har spredt seg nord til Bjørnøya og har hekket der årlig siden 2011 (Barrett et al. 2017).
Generelt for sjøfugl Mattilgang i hekketida er viktig, og det er sannsynlig at de negative endringene for de fleste sjøfuglartene skyldes redusert næringstilgang. I Barentshavet er det spesielt null- og ettårig torskefisk, sild, tobis og lodde som er viktige næringsemner for sjøfugl. Særlig bestandene av ungsild, lodde og polartorsk viser store naturlige bestandssvingninger. Ungsilda i Barentshavet beiter på bl.a. lodde, og man ser derfor sjelden store bestander av disse artene samtidig (Fauchald et al. 2015). Unge årsklasser av torskefisk som transporteres inn i Barentshavet fra gytefeltene lengre sør er viktige næringsemner for sjøfugl i Barentshavet, og er sannsynligvis bestemmende for plasseringen av de største sjøfuglkoloniene (Sandvik et al. 2016). Sjøfuglbestandene i Barentshavet kan nyttiggjøre seg de pelagiske fiskebestandene av lodde og sild, i tillegg til tobis og ung torskefisk og har derved et relativt allsidig næringsgrunnlag. Krykkjekoloniene sør i Barentshavet er i sterk tilbakegang, i likhet med de fleste kolonier i Nord-Atlanteren (Descamps et al. 2017). Krykkjene spiser det samme som de pelagiske alkefuglene, og det er tilsynelatende ingen åpenbare endringer i næringsgrunnlaget i Barentshavet som kan forklare den store nedgangen. Det er derfor grunn til å anta at bestandsnedgangen delvis kan skyldes endringer i næringstilgang i overvintringsområdene utenfor Newfoundland (Frederiksen et al. 2012; Reiertsen et al. 2014).
Endringer i næringstilgangen for sjøfugl kan skyldes mange faktorer. Lavere produksjon av byttedyr, naturlig predasjon på yngre årsklasser av fisk (f.eks. torsk som spiser ungsild som spiser lodde), uttak av fiskeressurser eller sekundæreffekter av klimarelaterte endringer har vært foreslått. (Cury et al. 2011; Grémillet et al. 2018; Reiertsen et al. 2012; Sandvik et al. 2005; Sandvik et al. 2012; Sandvik et al. 2014). Det er for eksempel vist at overlevelse hos alkekonge i de vestlige delene av Barentshavet har gått ned under oppvarmingen av dette området (Hovinen et al. 2014). Alkekonge ernærer seg i stor grad av hoppekreps i Calanus-slekten i hekkeperioden, der de fettrike, større arktiske artene er de foretrukne. En mulig forklaring på nedgangen i overlevelse er at det har blitt lavere forekomster av disse artene på grunn av oppvarmingen (Hovinen et al. 2014 kapittel 4.2.2). For krykkje på fastlandskysten er det også vist at antall og størrelse av egg som legges, samt størrelsen på eggene, går ned når innstrømming av varmt vann til Barentshavet øker. Årsakene kan være at varme forhold gjør krykkjas foretrukne byttedyr, som lodde, mindre tilgjengelig i hekkeperioden eller at de varme forholdene på andre måter endrer næringstilbudet (Barrett et al. 2017). I de siste årene er det observert en voksende makrellbestand som har spredt seg langt nord i Barentshavet. Denne spiser store mengder dyreplankton og små fisk (f.eks. Bachiller et al. (2016); Havforskningsinstituttet (2020b); Jansen et al. (2019)), og er en viktig konkurrent i sjøfuglenes matfat. Den eneste sjøfuglarten som kan utnytte makrellen er havsule, og den har nå spredt seg nordover i Barentshavet, og hekker på Bjørnøya.
Klimaeffektene kan også påvirke sjøfuglbestandene direkte og høyere frekvens av ekstremvær kan påvirke både overlevelse og hekkesuksess (Guéry et al. in press). Klimaendringer påvirker også oseanografiske forhold som bestemmer tilgjengeligheten av næring for sjøfugl. Den kraftige nedgangen i krykkjebestandene har vært sammenfallende med en rask og plutselig økning i overflatetemperatur i 1990-årene (Descamps et al. 2017). Denne brå oppvarmingen kan ha igangsatt storskala, sirkumpolare endringer i de marine økosystemene (Beaugrand et al. 2015) som har ført til at næringsgrunnlaget for krykkja mange steder har blitt kraftig forringet. Det kan altså se ut til at selve oppvarmingen av havet har mindre betydning for krykkjebestandene, men at takten oppvarmingen skjer i er helt avgjørende.
Endringer i oseanografiske forhold bestemmer også tilgjengeligheten av fiskelarver for hekkende sjøfugl. En studie der man har modellert driften av fiskelarver langs kysten viste at de viktigste fuglekoloniene er lokalisert ved områder med naturlig opphopning av larver (Sandvik et al. 2016). Hvis klimaendringer endrer oseanografiske forhold i så stor grad at det påvirker driften og retensjonsområdene for torskelarver, vil dette kunne få store konsekvenser for sjøfuglbestandene ikke bare i Barentshavet, men langs hele kysten.
Det er målt høye nivåer av miljøgifter i fugleegg (bl.a. i de arktiske artene polarmåke og ismåke, f.eks. Erikstad et al. (2011)), og resultatene gir grunn til bekymring for miljøgifter som påvirkningsfaktor for sjøfugl i norske havområder. Miljøgifter kan påvirke sjøfugl på forskjellig vis, for eksempel gjennom lavere reproduksjon, skjev kjønnsfordeling hos avkom og økt dødelighet (se Bustnes et al. (2015) og referanser i denne).
Selv om hovedårsaken til at mange sjøfuglbestander går ned sannsynligvis er knyttet til endringer i næringstilgangen, ser man også at en økende havørnbestand påvirker klippehekkende fugl (f.eks. lomvi og krykkje) negativt (Hipfner et al. 2012). For eksempel er lomviene som tidligere hekket på åpne fjellhyller på Hjelmsøya nå borte, og det er en økende bestand som hekker skjult i steinur, og man opplever fullstendig hekkesvikt både hos krykkje og havørn enkelte år i sentrale sjøfuglkolonier (f.eks. hos lomvi på Hornøya i 2019). Det drøftes om hvorvidt effektene av redusert næringstilgang gjør sjøfugl mer sårbar i forhold til predasjon (Hipfner et al. 2012) (se også Christensen-Dalsgaard et al. (2018)).
4.5.2 - Sjøpattedyr
Både vekst etter fredning (for noen arter) og klimaendringer påvirker bestander av sjøpattedyr i Barentshavet nå. Hvalross er eksempel på en art som har økt betydelig i antall etter flere tiår med fredning. Ringsel er en av flere arter som er avhengig av havis, og studier tyder på at ringsel nå bruker mer tid og energi på å få tak i mat enn tidligere på grunn av nedgang i mengde havis. I tillegg er ringselene helt avhengige av stabil is og nok snø på denne til at de kan lage kastehuler oppe på isen som beskytter de nyfødte mot vær, vind og predasjon. Slike forhold har vært mangelvare de siste 10-15 årene, spesielt på vestsiden av Svalbard, slik at man antar at rekrutteringen for denne nøkkelarten har vært ekstremt lav. Isbjørn i Barentshavet er et annet eksempel på en bestand som kan være påvirket av både fredning og klimaendringer. Det er økte forekomster av flere hvalarter langs kysten av Svalbard og en betydelig økning i kunnskap om flere istilknyttede hvalarter i løpet av de siste fem årene. Sommerforekomsten av vågehval i Barentshavet har økt som følge av endret miljøforhold. Bestandsstatus for grønlandssel i Barentshavet er usikker pga manglende tellinger, men antas å være stabil eller svakt økende. Situasjonen er god for sel langs fastlandskysten av Barentshavet, til forskjell fra kystsel lenger sør langs norskekysten.
Romlig fordeling av hval Indikatoren «Romlig fordeling av hval i Barentshavet» er basert på økosystemtokter om høsten og har vært en indikator i forvaltningsplanen for Barentshavet siden 2004. Siden starten av økosystemtoktet i Barentshavet har havområdet i nord vært preget av økende innstrømming av varmt atlanterhavsvann, og varierende loddemengde. Innledende analyser av romlig fordeling av de vanligste artene av hval og byttedyr viser at de ulike hvalartene fordeler seg i forhold til spesifikke habitat; knølhval, vågehval og finnhval beiter gjerne på nordlige banker, i områder med lodde, polartorsk, og krill, mens kvitnos beiter både i det sørlige Barentshavet og i polarfronten - sannsynligvis blant annet på lodde.
I perioden 2003–2009 var det lite endring i sjøpattedyrenes fordeling og derfor lite romlig respons på gjenoppbyggingen av loddebestanden fra 2006-2007 (Skern-Mauritzen et al. 2011). Deretter (2010-2013) så man en nordlig forflytning av kvitnos, til områder nord for polarfronten samtidig som det ble observert færre kvitnos i det sørlige Barentshavet. Bardehvalene syntes også å være mer konsentrert i nord nå enn i tidligere år (se også Storrie et al. (2018)). Analyser av data fra økosystemtoktene fra 2010-12 viser at fordelingen av vågehval i august-september var knyttet mest til forekomsten av lodde, mens fordelingen av finnhval og knølhval er knyttet til høy forekomst av både lodde og krill (Ressler et al. 2015). Ingen sterke korrelasjoner ble funnet i romlig fordeling mellom kvitnos og forskjellige byttedyr, noe som kan tyde på at de har en variabel diett (Fall og Skern-Mauritzen 2014).
Data fra vågehvalfangst har vist en nedgang i tykkelsen på spekklaget over perioden 1993-2013 (Solvang et al. 2017). Prøver fra fangst, tatt tidligere i sesongen enn økosystemtoktene, har vist et bredt spekter av byttedyr som inkluderer krill, sild, lodde og torskefisk. I løpet av studieperioden har den største spekktykkelse vært observert i perioder med mye ungsild i Barentshavet. Dårlig rekruttering hos norsk vårgytende sild de senere år er derfor en mulig årsak til nedgangen i kroppskondisjon hos vågehval. Økende konkurranse med den store torskebestanden om både sild, krill og lodde har sannsynligvis også spilt en rolle (Bogstad et al. 2015). Siden 2013 har det de fleste årene vært sjøpattedyrobservatører på de ordinære økosystemtoktene i Barentshavet. Disse dataene er fortsatt under analyse, men det generelle bilde er at fordelingen ligner det som hittil er beskrevet (ICES 2019c). På telletokter for vågehval er det blitt observert en del finnhval ved vestkysten av Svalbard og blåhval nord for Svalbard. Knølhval ses oftest øst for Svalbard i områder med mye lodde. Sommerforekomsten av knølhval i Barentshavet har ligget stabilt på omkring 4000 dyr siden år 2000, mens de var veldig sjeldne på 1980 og 1990 tallet. Det observeres en del nise i den sørlige delen av Barentshavet under vågehvaltoktene, og disse dataene inngår nå i evalueringen av bestandssituasjonen for nise i Barentshavsområdet (Bjørge 2020).
Bifangst av nise Bifangst av nise er valgt ut som indikator for menneskelig påvirkning i Barentshavet og overvåkingen skjer ved hjelp av kystreferanseflåten. Rapporteringen for denne indikatoren har imidlertid vært innstilt noen år på grunn av problemer med å tolke data i fravær av bestandsanslag for nisebestanden. I tillegg ble det oppdaget feil i datagrunnlaget for oppskalering av observerte bifangster til totale uttak.
Tellinger i 2015 ga et bestandsestimat på om lag 83 700 (59 430-107 980) niser i norske farvann nord for 62 °N opp til Vestfjorden. Niser i fjorder utenom Vestfjorden er ikke inkludert og antas å kunne øke estimatet med rundt 15 % (NAMMCO og Havforskningsinstituttet 2019). Senere tellinger i Barentshavet, Balsfjorden og Varangerfjorden bekrefter at det kan være betydelige antall her, men nøyaktige estimater er enda ikke klare (Bjørge 2020).
Etter korreksjon har man fått et estimat for bifangst på rundt 3000 niser per år for hele norskekysten, de fleste i Norskehavsområdet (Bjørge 2017). Det antas imidlertid også at de tilgjengelige bifangstestimater fra garnfiskerier etter torsk og breiflabb kun utgjør rundt 80 % av de totale bifangster i garnfiskeriene, så estimatet for andel av bestanden som tas som bifangst er usikkert (NAMMCO og Havforskningsinstituttet 2019). Det er derfor gjort studier på tiltak for å redusere bifangsten.
Foreløpige resultater fra norske eksperimenter med alarmer på garn («pingere») viser at disse sannsynligvis kan redusere bifangsten av nise med 70%-100%. Det jobbes med å innføre bruk av «pingere» i regulære fiskerier i områder med høy bifangstrate langs norskekysten, primært i Vestfjorden. For å få oversikt over bifangsten er det krav om at alle fiskerier skal rapportere bifangst av sjøpattedyr i sine elektroniske fangstdagbøker. Det rapporteres likevel lite om dette. I tillegg jobbes det med å få satt opp kameraer, som skal overvåke fangst og bifangst på 30 båter i tillegg til de eksisterende 30 båtene i referanseflåten (Bjørge 2020). Ved riktig plassering vil kameraer også kunne gi informasjon om hvor mange niser som faller ut av garnene under haling, noe som antas å være en kilde til underestimering av bifangsten. Manglende evne til å kunne dokumentere at bifangst av sjøpattedyr er på bærekraftige nivåer vil kunne lede til amerikanske importrestriksjoner for norsk sjømat fra 2022 (Bjørge 2018). Nedgang i breiflabbfiskeriet har gitt noe minkende bifangst av nise over perioden 2006–2015. Når effekt av bifangst skal vurderes, er det av verdi å også trekke inn relevant bakgrunnsinformasjon om nise og kunnskap om annen påvirkning, fordi dette kan ha betydning for effekten bifangsten har. I løpet av 2016 og 2017 ble det innsamlet biologiske prøver fra bifangede niser for å få viktig bakgrunnsinformasjon om bestandsstruktur, reproduksjonsrater og miljøgiftbelastning. Genetiske analyser viste at niser langs norskekysten ser ut til å tilhøre samme bestand, men det er fortsatt usikkerheter omkring utveksling med bestander i tilgrensende områder (Quintela et al. 2020). Genetiske studier utenfor Norge tyder på at nisehunner kan være mere stedfaste enn hanner (NAMMCO og Havforskningsinstituttet 2019). Analyser av kjønnsorganer fra de samme bifangede nisene viser reproduksjonsrater i det øvre sjikt av tilsvarende verdier for andre europeiske nisebestander. Relativt høye kvikksølvnivåer ble funnet i niser fra Lofoten, men ikke fra Finnmark (NAMMCO og Havforskningsinstituttet 2019). Det er foreløpig ikke gjort analyser for organiske miljøgifter. Tannhvaler som nise er spesielt utsatt for opphopning av miljøgifter fordi de befinner seg høyt i næringskjeden og har dårligere evne til å bryte ned miljøgifter enn f.eks. seler.
På grunn av liten kroppsstørrelse må niser ha en høy beiteintensitet for å motvirke varmetap (Wisniewska et al. 2016; Wisniewska et al. 2018). Dette kan gjøre arten spesielt sårbar overfor forstyrrelser i beiteområdene, som skipstrafikk, seismikk, påledriving mm. Nyere undersøkelser har dokumentert forbigående reduksjoner i beiteintensitet som respons på både påledriving (innen 20 km radius; (Dahne et al. 2013)) og 3D seismikk (innen 8-12 km radius; (Sarnocińska et al. 2020)). Betydningen av dette på bestandsnivå vil avhenge av det totale omfanget av forstyrrelser og den tilgjengelige mengden av byttedyr per nise i et gitt område (NAMMCO og Havforskningsinstituttet 2019). Visse deler av Barentshavet har tidvis en høy aktivitet av seismiske undersøkelser. Det eksisterer foreløpig ingen studier av hvordan dette påvirker beiteatferden hos niser og andre sjøpattedyr i dette området .
Vågehval Sommerforekomsten av vågehval i Barentshavet har siden 1980 tallet blitt regelmessig estimert som en del overvåkingen av hele den nordøstatlantiske vågehvalbestanden. Frem til 2013 har delestimatene for Barentshavet stort sett ligget stabilt på 40 000-50 000 vågehval, men senere tellinger har vist en økning til rundt 70 000 vågehval (se figur 4.27). Dette skyldes antakelig en forflytning av vågehval fra Norskehavet som respons på miljøendringer, men mer detaljerte årsakssammenhenger er foreløpig uklare. Totalestimatene for den nordøstatlantiske vågehvalbestanden baseres på seksårige tellesykluser i alle relevante sommerbeiteområder. Bestandsstørrelsen har ligget på et stabilt nivå fra 1995 til og med til det siste estimat fra 2008-2013 på 89 600 dyr. Telledata for perioden 2014-2019 er under analyse. Bestandsestimatene danner grunnlag for beregning av kvotene til norsk hvalfangst ut fra en metode utviklet av den internasjonale hvalfangst-kommisjon (IWC). Kvotene skal sikre langsiktig overlevelse av bestanden og et langsiktig, bærekraftig utbytte av fangsten. Kvotene for den norske vågehvalfangsten baseres på den såkalte reviderte forvaltningsprosedyren (Revised Management Procedure), som er utviklet av IWC. Langtidsmålet norske myndigheter har satt for bestanden er at den skal styres mot et nivå på 60 % av den opprinnelige bestanden. Dette skal sikre både langsiktig overlevelse av bestanden og et stabilt fangstutbytte.
Figur 4.27. Sommerforekomst av vågehval i Barentshavet. Kilde: (ICES, 2019a).
Grønlandssel Grønlandssel har en anslått forekomst på rundt 1,9 millioner dyr i Barentshavet i sommer- og høstmånedene. Av disse utgjør rundt 1,5 millioner sel Østisbestanden som har sitt yngleområde på is inne i Kvitsjøen, mens ca. 0,4 millioner er fra Vestisbestanden (ICES 2019b). Ungetellinger i Kvitsjøen viste en halvering over perioden 2003-2005 og har siden holdt seg på et stabilt lavt nivå frem til siste telling i 2013 (se figur 4.28). Modellen antyder en svak stigning i bestanden, men det finnes ikke oppdaterte ungeproduksjonsestimater som kan underbygge dette. Det er også betydelig usikkerhet om formeringsratene for denne bestanden (ICES 2019b). Drektighetsraten for 2018 lå imidlertid på et historisk høyt nivå. I 2015 og 2016 var det svært dårlige isforhold i kasteområdene i Kvitsjøen og det antas at ungedødeligheten i disse årene har vært noe forhøyet.
Figur 4.28. Modellert utvikling for Barentshavsbestanden av grønlandssel. Grønn kurve viser utvikling for totalbestanden, oransje kurve viser utviklingen i ungeproduksjonen. 95 % konfidensintervaller er vist med stiplete linjer. Resultater fra ungetellinger er med fylte sirkler og 95 % konfidensintervaller.
En ny telling i Grønlandshavet i 2018 viste en nær halvering av ungeproduksjonen sammenlignet med siste telling i 2012 (figur 4.29). Årsaken til denne reduksjonen er ikke kjent, men kan være knyttet til endring i reproduksjonsrater. Nye reproduksjonsdata fra 2019 er under opparbeiding. Det er kjent fra Nordvestatlanteren at drektighetsratene for grønlandssel kan variere mye fra år til år, særlig når bestanden er stor i forhold til næringsgrunnlaget. Dette kan ha stor betydning for ungeproduksjonen i et gitt år, men med den nåværende hyppighet av tellinger og innsamling av reproduksjonsmateriale i Nordøstatlanten (ca. hvert 5. år) er det vanskelig å knytte disse svingninger til økologiske endringer. Det gjøres imidlertid i øyeblikket forsøk på å studere dette for både Barentshavsbestanden og Grønlandshavsbestanden av grønlandssel.
ICES sitt forvaltningsråd på grønlandssel innebærer vanligvis en årlig fangst som med stor sannsynlighet vil stabilisere bestanden over en femtenårsperiode. På bakgrunn av usikkerhet knyttet til nedgang i ungeproduksjon har ICES anbefalt at fangstnivået baseres på mest mulig konservativ beregningsmåte i både Øst- og Vestisen. Konklusjonen fra Den blandete norsk-russiske fiskerikommisjonen for sesongen 2020 følger rådene fra ICES. I Vestisen ligger anbefalt fangstuttak på 11 548 dyr. I Østisen ligger anbefalt fangstnivå på 21 172 dyr.
Figur 4.29. Modellert utvikling for Grønlandshavsbestanden av grønlandssel. Grønn kurve viser utvikling for totalbestanden, oransje kurve viser utviklingen i ungeproduksjonen. 95 % konfidensintervaller er vist med stiplete linjer. Resultater fra ungetellinger er med fylte sirkler og 95 % konfidensintervaller.
Havert og steinkobbe Ved de siste tellingene utført frem til 2017 var bestandene av havert og steinkobbe i forvaltningsplanområdet for Barentshavet på eller over målnivåene i de respektive forvaltningsplanene for disse artene. Målnivåene er en rettesnor for fastsettelse av jaktkvoter. Innenfor forvaltningsplanområdet for Barentshavet er forvaltningsmålet for havert en bestand som produserer rundt 190 unger per år. For steinkobbe er forvaltningsmålet en totalbestand på rundt 3 420 dyr. Disse målnivåene er tilnærmet lik estimerte bestandsnivåer for perioden 1996-99, før en periode med høyt jaktuttak i perioden 2003-2010. Utviklingen i området fra Vesterålen og nordover står i sterk kontrast til utviklingen i Trøndelag og deler av Nordland, hvor begge arter har vist en til dels kraftig tilbakegang (Bjørge 2020). Nye tellinger av steinkobbe i Troms og Finnmark er planlagt i 2021.
Ringsel Estimater for bestandsstørrelse og trender finnes ikke for ringselene på Svalbard da det ikke har vært foretatt tellinger siden 2001 (Krafft et al. 2006). Det finnes likevel en del nyere økologiske studier som gir innsikt om utfordringer for denne selarten som følge av endringer i isforholdene. Ringselene på Svalbard har to forskjellige strategier etter at yngling og hårfelling er over. Den ene er å vandre nordover opp til iskanten og være der frem til høsten (Hamilton et al. 2015). Den andre er å oppholde seg kystnært og da hovedsakelig nær brefronter (Hamilton et al. 2016). Begge steder har tradisjonelt rikelig tilgang på næring samt hvileplattformer i form av havis eller breis. Det er i hovedsak yngre dyr som vandrer nordover mens voksne dyr forblir inne ved kysten. I de senere år har iskanten i nord befunnet seg nord for kontinentalsokkelen og over det dype Nordishavet. Satellittsporing av ringsel før og etter at iskantene flyttet seg så langt nord viser at selene fortsatt oppsøker dette området og oppholder seg i områder med samme iskonsentrasjon ((Hamilton et al. 2015), se figur 4.30). Etter at de har kommet hit svømmer de lengre, dykker lengre, er mindre tid i overflaten og ligger mindre oppe på isen og hviler enn før ((Hamilton et al. 2015), se figur 4.31).
Figur 4.30. Vandringsmønstre til ringsel fra Svalbard som ble utstyrt med satellittsendere i perioden 2002-2003 (gule linjer) og 2010-2012 (røde linjer). Hvite linjer viser gjennomsnittlig (heltrukne) og minimums (stiplede) isutbredelse i september for de ulike periodene. Data Center, Boulder, CO). Den svarte prikkete linjer angir grensen for kontinentalsokkelen.
Figur 4.31. Endringer i atferd hos ringsel i iskantsonen mellom periodene 2002-2003 (hvor iskanten var over sokkelen i Barentshavet) og i 2010-12 (hvor iskanten var over det dype Nordishavet). Røde tall indikerer signifikante forskjeller mellom de to periodene. Det stiplede rektangelet forklarer hvilke parametere som er sammenliknet. Kilde: Hamilton et al. (2015).
De viser også mindre såkalt "area-restricted search" - noe som tyder på at de ikke finner konsentrasjoner med mat. Alle disse endringene tyder på at de må jobbe mer for å finne mat, noe som igjen vil kunne ha innvirkning på dyrenes kondisjon med mulige følger for reproduksjon og overlevelse (Hamilton et al. 2015) For de voksne ringselene som oppholder seg langs kysten av Svalbard viser satellittsporing at disse holder seg svært tilknyttet brefronter - mer i dagens situasjon enn før isforholdene endret seg i fjordene på vestsiden av Svalbard (Hamilton et al. 2016; Hamilton et al. 2019). Nå er det enkeltindivider som oppholder seg foran en og samme brefront hele sporingsperioden. Også for disse voksne selene er det påvist endring i dykkeatferd som tyder på at de må jobbe mer for å finne mat (Hamilton et al. 2016). I de siste årene er det også gjort flere registreringer av ringsel som legger seg opp og hviler på land, noe som tidligere var svært uvanlig for denne svært isavhengige selarten. Det er til og med gjort registreringer av at ringsel legger seg opp på land i grupper sammen med steinkobber, noe som er en utvikling ingen hadde forutsett i forbindelse med klimaendringer og mangel på havis for denne selarten (Lydersen et al. 2017).
Hvalross Etter flere tiårs fredning av hvalross på Svalbard ser en nå tydelig vekst i bestanden. Et nytt estimat på hvalrossantallet på Svalbard basert på en flytelling av alle liggeplassene i august 2018, viste at det da var 5 503 (CI: 5031-6036) hvalross på øygruppen i det tidsrommet tellingen foregikk. Dette er en klar økning fra forrige telling i 2012 hvor antallet ble estimert til 3 886 (CI: 3553-4262) hvalross (Kovacs et al. 2014). Sammenliknet med den første systematiske tellingen av hvalross på Svalbard i 2006 (Lydersen et al. 2008) hvor det bare ble sett en årskalv, sees det stadig flere slike ved de nyere tellingene.
Grønlandshval I de senere år rapporteres grønlandshval fra turistbåter i Svalbardområdet stort sett årlig men i lavt antall (Storrie et al. 2018). Et unntak er turistbåten MV Plancius som hvert år følger iskanten fra Jan Mayen og nordover på vei opp til Svalbard og som rapporterer om større ansamlinger i Framstredet hvert å i juni (De Boer et al. 2019). Over 100 individer er observert ved enkelte anledninger innenfor små områder langs iskanten mellom 75 °N og 77 °N. I 2017 ble 16 hval utstyrt med satellittsendere i dette området, og sporingsdata viste at disse dyrene spredte seg over hele det antatte utbredelsesområdet til denne bestanden, fra kysten av Grønland via Svalbard og inn i det russiske området (Kovacs et al. innsendt). Samtidig har data fra passiv akustisk overvåkning (lyttebøyer som står ute hele året og registrerer all lyd) vist at i enkelte områder hører man grønlandshval døgnet rundt de fleste månedene i året; særlig i vintermånedene som er dyrenes paringstid (Ahonen et al. 2017; Stafford et al. 2012). En telling av hval i iskanten nord for Svalbard og vestover i 2015 førte til et estimat på 343 (95 % CI 136−862) grønlandshval i dette området (Vacquié-Garcia et al. 2017a). Tellingen ble foretatt med skip og helikopter. Alle observasjonene av denne hvalarten ble gjort fra helikopter - ingen fra skip. Med andre ord oppholder grønlandshvalene seg oftest lenger inne i isen enn man tidligere har antatt, og denne tellingen, samt data fra de akustiske lyttebøyene og fra turistbåter, tyder på det står noe bedre til med denne bestanden enn tidligere antatt.
Narhval En telling av hval i iskanten nord for Svalbard og vestover i 2015 førte til et estimat på 837 (95 % CI 314−2 233) narhval i dette området (Vacquié-Garcia et al. 2017a). Narhval rapporteres sjelden fra Svalbardområdet, men ser altså ut til å være mindre sjelden dypt inne i isen. Disse hvalene ble observert langt fra iskanten og gjerne på slutten av transektlinjene, som generelt gikk inn til 185 km nord for iskanten. Dette kan tyde på at man bare så vidt har undersøkt sørlige deler av denne artens utbredelsesområde. I tillegg hører man narhval vokalisering og ekkolokalisering året rundt fra en lyttebøye i Framstredet på ca. 79˚N (Ahonen et al. 2019). Dette er et område hvor man stort sett har isdekke hele året. Narhval fra andre steder tilbringer sommermånedene i isfrie farvann, men det er tydelig at vi her, basert både på de akustiske undersøkelsene og flytellingene, har å gjøre med en til nå ubeskrevet bestand som oppholder seg inne i isen hele tiden.
Steinkobbe Steinkobbene på Svalbard utgjør en egen genetisk bestand (Andersen et al. 2011) som er verdens nordligste og som ved forrige (og eneste) telling ble estimert til å bestå av rundt 2000 individer (Merkel et al. 2013). Hovedutbredelsesområdet er på vestsiden av Prins Karls Forland, og satellittsporing av dyr herfra viste at de holder seg i kystnære områder hele året hovedsakelig på vestsiden av Spitsbergen (Blanchet et al. 2014). De unngår området med mye is om vinteren, men legger seg opp og hviler på isflak nær kontinentalskråningen. Atlanterhavsvannet i Spitsbergenstrømmen ble vist å være et viktig beitehabitat. Rapporter om observasjoner av steinkobber i områder utenom det tidligere kjerneområdet rundt Prins Karls Forland blir mer og mer vanlig (Norsk Polarinstitutt marine pattedyrdatabase, https://data.npolar.no/sighting/). Steinkobbene på Svalbard synes å være "klimavinnere" ettersom det blir mindre og mindre sjøis i området, mens de mer endemisk arktiske selartene risikerer å være tapere med den pågående reduksjonen i is-utbredelse og stigende vanntemperaturer (Blanchet et al. 2015; Blanchet et al. 2016; Blanchet et al. 2014).
Storkobbe Vi vet svært lite om bestandsforhold til storkobbene i dette området. Det er en kystnær sel som finnes spredt utover hele Svalbardområdet hvor de hovedsakelig ernærer seg av bentiske byttedyr eller dyr som oppholder nær bunnen (Hjelset et al. 1999). Sporingsstudier av voksne dyr viser en stor individuell variasjon (Hamilton et al. 2018), samt at de utnytter ulike deler av fjordsystemene de deler med ringsel og dermed reduserer potensiell konkurranse (Hamilton et al. 2019). De er avhengige av isplattformer for fødsel og diing av ungene sine og det er nylig vist at veksten hos ungene i perioden etter at sjøisen ble mangelvare på vestkysten av Svalbard i mai (som er yngletiden), ikke ble redusert. Dette fordi mødrene benyttet seg av isbrekalvinger som die- og fødeplattformer (Kovacs et al. 2020). Dette er imidlertid en kortvarig løsning siden breene i dette området gradvis trekker seg tilbake og etter hvert ender opp på land.
Hvithval Hvithval er den oftest observerte hvalarten på Svalbard. Den holder seg svært nær kysten og foran breer i den isfrie delen av året (Lydersen et al. 2001; Vacquié-Garcia et al. 2018). Gjennom vinteren presses den ut fra kysten, men oppholder seg inne i isen nær Svalbard. I de senere år med mer innstrømming av atlanterhavsvann i fjordene på vestsiden av Svalbard viser det seg at hvithvalene her tilbringer mindre tid foran breer og mer tid ute i disse «nye» vannmassene og da antakelig spiser på mer boreale byttedyr ((Hamilton et al. 2019), se figur 4.32). I 2018 ble for første gang en flytelling av hvithval utført som dekket hele Svalbard (over 7000 km ble fløyet) og som resulterte i et estimat på 549 (95 % CI: 436-723; (Vacquié-Garcia et al. 2020)). Dette tallet var overraskende lite og tyder på at bestanden har problemer med å ta seg opp etter at den ble «kommersielt utryddet» på 1960-tallet. Nye data om dykking hos disse hvalene (Vacquié-Garcia et al. 2019) ble brukt til å lage en korreksjonsfaktor for tid i overflaten i forhold til tid i neddykket tilstand som ble anvendt i dette estimatet.
Figur 4.32. Tid tilbrakt foran brefronter (5 km) for ringsel og hvithval før og etter endringer i miljøforholdene (a og b) og gjennom sesongen (c og d) på vestkysten av Svalbard.
Generelt om hval Forekomsten av større hvalarter som blåhval, finnhval og knølhval i kystnære farvann på Svalbard har økt de senere årene (Storrie et al. 2018). Dette gjelder i første rekke fjordene på vestsiden av Spitsbergen, men også i Storfjorden, Hinlopen og nordkysten av Spitsbergen (Vacquié-Garcia et al. 2017b). Det er også en trend at man finner disse store hvalene lengre og lengre mot nord ettersom iskanten trekker seg denne veien (Storrie et al. 2018).
Isbjørn Barentshavområdet har en av i alt nitten antatte arktiske delbestander av isbjørn, men med høy genetisk utveksling mot nabobestandene i vest og øst (Peacock et al. 2015). Bestanden var estimert til omtrent 2 650 (95 % konfidensintervall ca. 1 900-3 600) dyr i august 2004 (Aars et al. 2009), og var antatt å ha økt betraktelig etter fredningen i 1973 (Aars et al. 2009; Derocher 2005). Tilgjengeligheten av sjøishabitat har i senere år blitt redusert mye raskere i Barentshavområdet enn for andre isbjørnbestander (Stern og Laidre 2016), og reproduserende binner har i økende grad vært forhindret fra å nå tradisjonelt viktige hiområder øst på Svalbard (Aars 2013; Derocher et al. 2011). Det antas at en økende andel av bestanden går i hi i russisk Arktis (Franz Josef Land). Leveområdene er også forskjøvet mye lengre nordover, siden iskanten er det området der de fleste isbjørnene jakter mye av året (Lone et al. 2017). Rundt Svalbard bruker isbjørn nå mer tid på land, og tar mer egg og fugl i sommerhalvåret (Prop et al. 2015). De jakter også mindre foran brefronter på sommeren, siden disse områdene har mindre fastis enn før (Hamilton et al. 2017). Økende avstand med åpent vann mellom iskanten og øyene på Svalbard fører til lengre svømmeturer som krever mer energi (Lone et al. 2017). Isbjørn i Barentshavområdet utsettes for høye nivåer av miljøgifter, med sannsynlige negative effekter (Routti et al. 2019). Til tross for både dette, og det raske tapet av sjøishabitat, ser det ut som den lokale bestanden på Svalbard har holdt seg på rundt 300 bjørn fra 2004 til 2015, og at antallet bjørn totalt på norsk side er like høyt som før, eller har økt (Aars et al. 2017). Det er ingen tegn til at kondisjon (dvs. fettlager) hos isbjørnene har gått ned over tid, eller at ungeproduksjon har gått mye ned (www.mosj.no). Det antas at bestanden opp til nå har vært stabil eller økende fordi den fortsatt ikke har nådd en nedadgående bærekapasitet etter at den var langt under dette ved fredningen i 1973 (Aars et al. 2017).
4.6 - Fremmede arter
Fremmede arter i Barentshavet inkluderer kongekrabbe og pukkellaks. Bestanden av kongekrabbe er så stor at den kan ha betydelige effekter på økosystemet nær fastlandskysten. Flere fremmede arter er vurdert å kunne spre seg inn i Barentshavet og til områdene rundt Svalbard de neste 50 årene
Pukkellaks Pukkellaks (Oncorhynchus gorbuscha) finnes naturlig i nordlige Stillehavet, men ble innført til Nordvest-Russland i flere omganger i andre halvpart av 1900-tallet. Fra og med 1960 har det blitt tatt pukkellaks også i norske elver. Pukkellaks er en engangsgyter (den dør etter gyting), og populasjonen følger vanligvis en distinkt toårssyklus. I Norge er det gytepopulasjonene i oddetallsår som dominerer. Tidligere års statistikk har vært basert på fangst i elv, og det var betydelig øking i fangstene i 2017 (hele landet) og 2019 (særlig konsentrert i Finnmark). I 2019 ble også sjøfisket pukkellaks rapportert (SSB tabell 09243: Sjøfiske. Fangst, etter fiskeslag og vektklasse (K) (1993 – 2019)). I kommunene fra Nordkapp til Sør-Varanger ble det fisket 4 037 pukkellaks i sjø. Fremtidig rapportering av pukkellaks i sjø vil kunne klargjøre hvordan utviklingen for arten er også i sjøfasen.
Vitenskapskomiteen for Mat og Miljø har nylig gjennomført en risikovurdering for pukkellaks. Hovedkonklusjonen er at invasjon av pukkellaks i norsk kystfarvann og norske vassdrag vil ha negative konsekvenser på biologisk mangfold, produktiviteten til lokal laksefisk og akvakultur. Regionalt og internasjonalt samarbeid må til for å redusere omfanget av pukkellaks (VKM Rapport 2020: 01).
Andre fremmede arter Den nye fremmedartslisten (med risikovurdering også for 50 år inn i fremtiden) kom i 2018. En beskrivelse av metodene for risikovurdering er gitt hos Artsdatabanken.
For Barentshavet har kongekrabbe (Paralithodes camtschaticus) en så stor biomasse at den kan ha betydelig effekter på økosystemet. Kongekrabbe er vurdert til SE (=svært høy risiko). Kongekrabbe er allerede etablert i store deler av forvaltningsplanområdet, og er forventet å spre seg sørvest til Helgelandskysten i løpet av de kommende 50 år. Snøkrabbe (Cionoecetes opilio) stod tidligere på listen over fremmede arter, men er tatt ut fordi nye genetiske undersøkelser tyder på at arten ikke stammer fra Canada eller Grønland og derfor sannsynligvis ikke har blitt innført med ballastvann. Hovedteorien er nå at arten har vandret til Barentshavet fra Beringstredet, langs nordkysten av Russland (Havforskningsinstituttet 2020a).
Spøkelseskrepsen (Caprella mutica) er vurdert til SE, og er hittil funnet nord til Troms, men er fortrinnsvis knyttet til naturlig eller menneskeskapt kystnær hardbunn, og har antatt lav forekomst i forvaltningsområdet. Det er likevel sannsynlig at den vil kolonisere Finnmarkskysten i løpet av få år.
Grønnalgen pollpryd (Codium fragile) er vurdert til SE og er forventet å forekomme langs hele Finnmarkskysten i løpet av 50 år.
Det er vurdert 24 marine invertebrater for Svalbard. Av disse er 23 «dørstokkarter» (De finnes der ikke for øyeblikket, men finnes enten i nærheten, eller har miljøkrav som passer, og potensiell transportvei til Svalbard). I tillegg til høyrisikoarten kongekrabbe, er krepsdyret Ischyrocerus commensalis vurdert til PH (=potensiell høy risiko). Brunalgen østerstyv (Colpomenia peregrina) er også vurdert til PH og forventes å finnes langs hele norskekysten i løpet av få år, og på Svalbard i løpet av de kommende 50 år.
4.7 - Truede arter og naturtyper
Det er generelt få arter og naturtyper som er vurdert som truede i Barentshavet–Lofoten, men dette er delvis på grunn av kunnskapsmangel. For de aller fleste marine arter uten kommersielle interesser har vi svært begrenset kunnskap om både forekomst og bestandsendringer. 26 arter er vurdert til å være truede i Barentshavet–Lofoten. Sammenliknet med 2010 er fem arter vurdert som mindre truet mens situasjonen er mer alvorlig for seks arter, de fleste sjøfugler. Fire av naturtypene i havområdet ble vurdert til å være sterkt eller kritisk truet i Rødlistea for naturtyper fra 2018.
Norsk rødliste for arter er en oversikt over arter som er vurdert å ha en risiko for å dø ut fra Norge. Artene på rødlista er plassert i ulike kategorier, vurdert ut fra et kriteriesett som er utviklet av Den internasjonale naturvernunionen (IUCN). Listen ble sist oppdatert i 2015. Norsk rødliste for naturtyper tar utgangspunkt i Artsdatabankens klassifiseringssystem for naturtyper fra 2009. I 2011 ble marine naturtyper vurdert og klassifisert for første gang, og siste revisjon er fra 2018 og med vurdering av marine naturtyper etter inndeling i NiN-systemet.
Status for rødlistede arter i Barentshavet er oppsummert i tabell 4.1. Sammenliknet med 2010 er fire av de 26 artene i Barentshavet- Lofoten som er vurdert som truet på den norske rødlista fra 2015, vurdert som mindre truet. For seks arter er situasjonen vurdert som mer alvorlig. Dette gjelder fire sjøfugler (havhest, alke, makrellterne og polarlomvi på Svalbard) og blåhval og ringsel (Svalbard). Situasjonen for grønlandshval, storskate og lomvi på fastlandet er fortsatt svært kritisk. Sabinemåke på Svalbard er den eneste av sjøfuglene som har endret kategori til en mindre truet. Resten av artene er plassert i samme kategori.
Blant fiskeartene er det nå bare storskate som regnes som kritisk truet (CR). For snabeluer har tilstanden forbedret seg så mye at den har gått ut av lista for truede arter. Det samme har ikke skjedd for vanlig uer, som fortsatt regnes som sterkt truet (EN). Også for ål og pigghå har det skjedd en endring til en bedre kategori selv om begge fortsatt regnes som truede.
Også for steinkobbe har det skjedd en endring slik at denne nå ikke regnes som truet. Steinkobbe som befinner seg nær Svalbard har ikke hatt samme oppgangen og regnes fortsatt som truet.
Av arter som pr i dag ikke er listet som truet, men som det likevel er verdt å merke seg er at polartorsk (Boreogadus saida) har endret status fra livskraftig (LC) til nær truet (NT). Det samme har blåsteinbit (Anarhichas denticulatus). For polartorsk tyder toktdata på en nordlig og østlig forflytning de senere årene og en sannsynlig reduksjon i tallrikheten ved Svalbard. Basert på forskningsdata for perioden 1980-2008 viser polartorsken en preferanse for temperaturer mellom 2- 5.5 °C. Områder med denne temperaturen er blitt kraftig redusert de senere årene.
Rødlista for naturtyper bygger på Artsdatabanken sitt inndelingssystem "Naturtyper i Norge". I de marine dypvannsområdene i Barentshavet finner vi to naturtyper: - korallrev og grisehalekorallbunn. For korallrev har nye funn nasjonalt bidratt til at andel av forringet totalareal er blitt redusert og ble vurdert til å være mindre truet, fra sårbar til nær truet i rødliste 2018. Utviklingen for grisehalekorallbunn er forverret på grunn av pågående påvirkning siden forrige rødlistevurdering og ble i 2018 vurdert som sterkt truet.
Utover disse er flere arter og naturtyper vurdert, men det er generelt store kunnskapsmangler og er derfor vanskelig å håndtere. For eksempel er det mangel på kunnskap knyttet til svampsamfunn og andre habitater som gjør det vanskelig å vurdere status i henhold til rødlista.
Tabellene under viser et utvalg truede arter og naturtyper som vurderes å være relevante for forvaltning i planområdet. Utvalget av arter i saltvannssystemer tar med artsgruppene pattedyr med marin tilknytning, sjøfugler og fisker, men ikke bløtdyr, krepsdyr og leddormer. For sjøfugl har vi holdt oss til de som har marin tilknytning hele året. Arter som er merket med * gjelder Svalbard. Kun marine naturtyper som er dannet av koralldyr, på dypt vann, er tatt med.
Tabell 4.1. Tabellen viser truede arter og hvilken risikokategori artene ble plassert i rødlistene fra 2006, 2010 og 2015. Den viser også hva slags menneskelig og naturlig påvirkning som kan ha betydning for artene. De som er markert gul og grønn hadde en forbedring i utviklingen fra 2010 til 2015. De som er farget med rødt hadde en forverring i utviklingen. Grønn markering betyr at den er vurdert å ikke være truet. De truede artene er plassert i følgende kategorier:
CR = kritisk truet EN = sterkt truet VU = sårbar NT = nær truet LC = sikre bestander (levedyktige) DD = datamangel NA = ikke egnet, skal ikke vurderes
*Gjelder artsforekomster på Svalbard. Kilde: Artsdatabanken (2018). Norsk rødliste for naturtyper 2018. Hentet (dato) fra https://www.artsdatabanken.no/rodlistefornaturtyper. Kategoriene LC og NT står for levedyktig hhv nær truet, DD innebærer at det er mangel på data. Rødt=forverring, gult=bedring, grønt=ikke truet
Vitenskapelig navn
Norsk navn
Status i rødliste fra
Påvirkningsfaktorer
Pattedyr
2006
2010
2015
Phoca vitulina
Steinkobbe
VU VU*
VU VU*
LC VU*
Fastlandsbestanden av steinkobbe er vurdert som levedyktig *Svalbardbestanden –klimaendringer, risiko for oljeutslipp
Pusa hispida
Ringsel*
LC
VU
Ursus maritimus
Isbjørn *
VU
VU
VU
Klimaendring, menneskelig forstyrelse, forurensing, påvirkning på habitat (framtidig petroleumsaktivitet),
Odobenus rosmarus
Hvalross*
VU
VU
VU
Menneskelig forstyrelse, påvirkning på habitat, klimaendringer, risiko for framtida forurensing
Cystophora cristata
Klappmyss*
VU
EN
EN
Høsting (opphørt), klimaendringer
Balaena mysticetus
Grønlandshval
CR
CR
CR
Påvirkning på habitat, klimaendringer og høsting (historisk)
Lutra lutra
Oter
VU
VU
VU
Høsting (flora og faunakriminalitet), påvirkning på habitat, forurensing
Høsting (påvirkning på næringsdyr), påvirkning fra stedegne arter
Cepphus grylle
Teist
NT
VU
VU
Fremmede arter (predatorer), høsting (påvirkning på næringsdyr), menneskelig forstyrelse
Fratercula artica
Lunde
VU
VU
VU
Høsting, påvirkning fra stedegne arter (næringsdyr, predatorer)
Alca torda
Alke
NT*
VU EN*
EN
Rissa tridactyla
Krykkje
VU
EN
EN
Høsting (påvirkning på næringsdyr), fremmede arter (predatorer), klimatiske endringer
Fulmarus glacialis
Havhest
-
NT
EN
Påvirkning fra stedegne arter (rovdyr, næringsdyr)
Sterna hirundo
Makrellterne
VU
VU
EN
Påvirkning fra stedegne arter (næringsdyr),
Uria lomvia
Polarlomvi*
NT
VU
EN
Pagophila eburnea
Ismåke*
EN
VU
VU
Forurensing, klimatiske endringer
Xema sabini
Sabinemåke*
EN
EN
VU
Koralldyr
Radicipes gracilis
Grisehalekorall
-
-
VU
Påvirkning på habitat
Vitenskapelig navn (til den dominerende art som danner naturtypen)
Norske navn
Status i rødliste fra
Påvirkningsfaktorer
2011
2018
Lophelia pertusa
Korallrev
VU
NT
Påvirkning på habitat
Radicipes sp.
Grisehalekorall-skogbunn
VU
EN
Påvirkning på habitat
Geodia svamper
Ostur i Barentshavet sør
-
NT
Påvirkning på habitat
Saccharina latissima
Nordlig sukkertareskog
-
EN
Påvirkning fra beiting av kaldtvannsarten grønn kråkebolle som i sin tur påvirkes av temperatur og rovdyr/krabbe
Laminaria digitata
Nordlig fingertarebunn
-
VU
Påvirkning fra beiting av kaldtvannsarten grønn kråkebolle som i sin tur påvirkes av temperatur og rovdyr/krabbe
Laminaria hyperborea
Nordlig stortareskog
-
NT
Påvirkning fra beiting av kaldtvannsarten grønn kråkebolle som i sin tur påvirkes av temperatur og rovdyr/krabbe
Mytilus edulis
Eksponert blåskjellbunn
-
VU
Kan påvirkes av klimatiske endringer i vanntemperatur, forurensing, samt påvirkning fra stedegne rovdyr og patogener/parasitter
Polar havis
-
CR
Påvirkes av klimabetinget temperaturendring
Kaldtvanns-bassenger
-
EN
Påvirkning av klimabetinget temperaturendring. Kan i fremtiden påvirkes av at snøkrabbe etablerer seg.
Tabell 4.2. Tabellen viser truede og nær truede naturtyper i Barentshavet og områdene utenfor Lofoten. Den viser også hva slags menneskelig og naturlig påvirkning som kan ha betydning for artene. De som er markert gul og grønn hadde en forbedring i utviklingen fra 2010-2015. De som er farget med rødt hadde en forverring i utviklingen. Grønn markering betyr at den er vurdert å ikke være truet. Kilde: Artsdatabanken (2018). Norsk rødliste for naturtyper 2018. Hentet (23.01.20) fra https://www.artsdatabanken.no/rodlistefornaturtyper.
4.8 - Forurensning, inkludert ” Trygg sjømat”
Tilførsler og nivåer av miljøgifter og radioaktiv forurensning er stort sett stabile eller nedadgående. Målinger i sedimenter har vist forholdsvis høye nivåer av polyaromatiske hydrokarboner nær Svalbard som trolig skyldes naturlige kullforekomster. Nivåene av forurensede stoffer i fisk og skalldyr er generelt lave og godt innenfor krav til trygg sjømat. For de fleste målte miljøgifter er de også under tilgjengelige grenseverdier for god miljøkvalitet, men vi finner fortsatt høyere nivåer i toppredatorer. Isbjørn har høye nivåer av fettløselige miljøgifter som oppkonsentreres i næringskjeden. Nivåene av mange miljøgifter i isbjørn avtar imidlertid i takt med nedgang i økosystemet ellers, mens nivåene av noen «nye» stoffer øker. Miljøgiftnivåene kan ha skadelige effekter på isbjørn, særlig i kombinasjon med sult.
Faktaboks 4.1. Klassifiseringssystemer for miljøgifter.
1) Miljødirektoratets klassifiseringssystem for sediment er inndelt i fem tilstandsklasser: Bakgrunn (I), God (II), Moderat (III), Dårlig (IV), Svært dårlig (V). Grenseverdier er angitt for ulike metaller og miljøfarlige stoffer i Miljødirektoratets veileder M-608 (Miljødirektoratet, 2016).
2) PROREF klassifiseringssystem: Under Miljødirektoratets MILKYS program (Miljøgifter i norske kystområder) er det blitt etablert en såkalt PROREF (Provisional high reference concentration, eller provisorisk høy referansekonsentrasjon) for torsk og blåskjell. Begrepet «referanse» her kan i hovedsak betraktes som «bakgrunn» for kystområder. PROREF er basert på MILKYS data fra de siste 25 årene, og oppdateres periodevis. Programmet omfatter prøver fra både antatt forurensede steder og fra steder langs kysten langt fra kjente punktkilder. Formålet med systemet er å kunne skille mellom stasjoner med lite forurensning og forholdsvis lave nivåer og stasjoner påvirket av lokale kilder. Fremgangsmåte og resultatene er først beskrevet i MILKYS rapporten for 2017 (Green et al. 2017).
3) Miljøkvalitetsstandarder: Vannforskriften har definert et sett med EU-definerte grenseverdier som kalles miljøkvalitetsstandarder eller EQS (Environmental Quality Standard). Norge har i tillegg definert sine egne miljøkvalitetsstandarder på stoffer og medier som ikke er dekket av EUs liste. Begrepet «miljøkvalitetsstandarder» er brukt i denne rapporten som en felles betegnelse for begge kilder. Miljøkvalitetsstandarder er satt for å beskytte de mest sårbare delene av økosystemet. Selv om det er målt miljøgiftnivåer over miljøkvalitetsstandarden, behøver ikke det å bety at organismene selv tar skade, men det viser at nivået er såpass høyt at andre deler av økosystemet kan ta skade, for eksempel sjøpattedyr. Miljøkvalitetsstandardene er ikke arts- og vevsspesifikk.
4) EUs og Norges grenseverdier for mattrygghet: Disse grenseverdiene angir den maksimale konsentrasjonen av en gitt miljøgift eller radioaktivt cesium som er tillatt i sjømat som omsettes for salg. Siden mennesker spiser langt mindre sjømat enn dyr høyt oppe i den marine næringskjeden, er disse grenseverdiene ofte satt høyere enn miljøkvalitetsstandardene. For enkelte stoffer er imidlertid mattrygghetsgrensen lavere enn PROREF. Grenseverdiene for mattrygghet er forskjellig for ulike typer sjømat. Grenseverdiene er satt for å beskytte befolkningen mot for stort inntak av de farligste miljøgiftene ved å fjerne «verstingene», men garanterer ikke for at man ikke kan få i seg for mye av en type miljøgift dersom man spiser svært mye av en bestemt type fisk eller annen sjømat.
4.8.1 - Tilførsel av forurensende stoffer til Barentshavet
Tilførsel av miljøgifter til Barentshavet skjer med luft- og havstrømmer, tilførsler fra elver, avrenning fra land, og transport med is. Luft- og havstrømmer står for hovedmengden av tilførslene.
Lufttilførsler av miljøgifter Konsentrasjonene av miljøgifter i lufta på Svalbard påvirkes av utslipp av miljøgifter i ulike deler av verden. Europa og Asia står for de største bidragene. Konsentrasjonene påvirkes også av klimatiske forhold som påvirker de atmosfæriske tilførselsveiene. I mange år har det vært nedgang for flere av miljøgiftene som fraktes til Barentshavet med luft- og havstrømmer. Trenden har stagnert de siste årene.
I 2018 var konsentrasjonene av polyklorerte bifenyler (PCB), polyaromatiske hydrokarboner (PAH) og de fleste plantevernmidler som måles i lufta ved målestasjonen Zeppelin på Svalbard litt lavere enn tidligere år, og nivåene viser en fortsatt nedadgående trend. Nivåene av bromerte flammehemmere (polybromerte difenyletere, PBDE) varierer fra år til år, og det er ingen tydelige trender å se. Per- og polyfluorerte alkylstoffer (PFAS) og heksabromsyklodekan (HBCD) er hovedsakelig under deteksjonsgrensen og ingen tydelige trender kan ses. Nivået av plantevernmiddelet heksaklorbenzen (HCB) viste en oppadgående trend fra begynnelsen av 2000-tallet og frem til 2015, men har deretter vist en nedadgående trend i perioden 2016–2018. Fortsatt overvåking er nødvendig for å følge med utviklingen.
Årlig middelkonsentrasjon av PAH i luft hadde en klar nedadgående trend fra 1998 til 2006. Etter dette har nivåene vært relativt stabile, med noen variasjoner fra år til år. Forhøyede nivåer i 2013 og 2014 skyldes i hovedsak enkeltmålinger med høye nivåer. Deretter (2015–2018) har nivåene gått nedover. De fleste av PFAS-forbindelsene som måles på Svalbard er under det analytiske deteksjonsnivået, nivåene er med andre ord så lave at de ikke vises i målingene. Av PFAS er perfluoroktansulfonsyre (PFOS), perfluoroktansulfonsamid (PFOSA) og perfluroktansyre (PFOA) de forbindelsene som i størst grad er målt over deteksjonsnivået, men det er ikke observert entydige trender.
Det er en svak nedadgående trend for i kvikksølvnivåene i lufta på Svalbard siden 2000. Dette samsvarer med observasjoner på lavere breddegrader, som også viser en avtagende trend, skjønt nedgangen har vært tydeligere på lavere breddegrader. Konsentrasjonen av bly har gått ned med 30% ved Zeppelinstasjonen siden målingene startet i 1994. Lavere nivåer av bly har sammenheng med at utslippene er betydelig redusert i Europa og Nord-Amerika, etter at bly i bensin ble forbudt i vestlige land. Utslippene har imidlertid økt sterkt i Asia.
Nivået av kvikksølv som måles i lufta på Svalbard varierer gjennom året. Om vinteren transporteres forurenset luft fra vestlige, sentrale og østlige deler av Europa nordover, noe som fører til høyere nivåer av kvikksølv i lufta på Svalbard. Om sommeren er nivåene også høye. Årsaken er avdamping av kvikksølv fra havet, fordi havisen smelter. Om våren er det episoder hvor kvikksølvnivåene i lufta er sterkt redusert. Dette skyldes prosesser i atmosfæren som omdanner kvikksølv til mer reaktive forbindelser som avsettes på bakken, på is og i havoverflater. Mesteparten av kvikksølvet vil fordampe tilbake til atmosfæren. En liten andel vil ende opp i sedimenter, jord, innsjøer og i havet (AMAP 2011a). Alger og bakterier kan omdanne kvikksølv til det svært giftige metylkvikksølv, og dermed kommer kvikksølvet inn i næringskjeden. Fisk og pattedyr høyt oppe i næringskjeden oppkonsentrerer metylkvikksølv i kroppen gjennom det de spiser.
Klimaendringer med økt temperatur forventes å gi økt spredning av miljøgifter på global skala (UNEP/AMAP 2011). Smelting av havis og tining av permafrost kan forårsake remobilisering og fordamping av miljøgifter til atmosfæren i Arktis (AMAP 2011b; AMAP 2016). Store skogbranner og branner på dyrket mark har vist seg å gi økt tilførsel av organiske miljøgifter til Arktis (Eckhardt et al. 2007). Økt lokal industrietablering (som for eksempel olje- og gassvirksomhet og bergverk) og skipstrafikk i nordområdene kan potensielt bidra til økte tilførsler av noen av de miljøgiftene som måles i lufta på Svalbard.
Det er fortsatt en nedgang i tilførslene og nivåene av flere av miljøgiftene som måles i lufta på Svalbard, men for noen har det vært en svak økning de siste årene. Antatte årsaker kan være fortsatt bruk av miljøgiftene i ulike deler av verden, og at miljøgiftene frigis med økende temperatur fra tidligere avsetninger i miljøet.
Tilførsler av forurensning fra kystnære landområder, elver og kystvann Indikatoren ”tilførsel av forurensninger i elver” viser tilførslene fra land til kystsonen ved forvaltningsplanområdet. Hvert år overvåkes utslipp til norskekysten av næringsstoffer, tungmetaller og organiske miljøgifter. Elveovervåkingsprogrammet er en del av Norges forpliktelser i OSPAR-avtalen, som omfatter tilførsler og utslipp til Nord-Atlanteren. Overvåkingen utføres i til sammen 47 elver. Elver med utløp til Barentshavet som inngår i programmet er Målselva, Altaelva, Tanaelva og Pasvikelva. I tillegg beregnes tilførsler fra umålte felt, og direkteutslipp fra industri, kloakkrenseanlegg og fiskeoppdrett. Tilførslene av fosfor og nitrogen har økt betydelig i perioden 1990-2018. Konsentrasjonen av næringssalter som fraktes med elvene ut i dette havområdet, har vært relativt uforandret i perioden. Den store økningen i tilførslene av næringssalter skyldes derfor utslipp fra kilder i havet, og da hovedsakelig fra fiskeoppdrett.
Figur 4.33. Årlig tilførsel av kobber til den delen av kysten som grenser mot Barentshavet-Lofoten.
Tilførslene av kobber har også økt mye i den samme perioden (figur 4.33). Mye av økningen skyldes økte utslipp fra oppdrettsnæringen, men det har også periodevis vært store tilførsler av kobber med elvene, for eksempel i 2013. Sistnevnte skyldes i hovedsak utslipp fra Kola i Russland (Bohlin-Nizzetto et al. 2019). Russiske utslipp av kobber og nikkel fører til forhøyede konsentrasjoner i norske elver som ligger lengst øst mot grensen. Dette påvirker også transporten fra land til den østlige delen av Barentshavet. Det er store variasjoner i tilførslene av nikkel fra år til år, blant annet som følge av variasjon mellom år i frekvens og intensitet av flommer. Det er imidlertid uklart hvor mye av disse utslippene som transporteres fra kystsonen og inn i selve forvaltningsplanområdet.
Tilførsler av radioaktiv forurensning Det er tilførsler av både antropogene og naturlige radioaktive stoffer i havområdet og det er ulike tilførselsveier for de forskjellige nuklidene. Technetium-99 (Tc-99) kommer i stor grad fra Sellafield-anlegget i Storbritannia, mens cesium-137 (Cs-137) og strontium-90 (Sr-90) kommer med havstrømmer fra områder som ble forurenset etter Tsjernobyl-ulykken, og i noe mindre grad fra Sellafield. Plutonium-239 og -240 (Pu-239, 240) kommer i stor grad fra nedfall etter prøvesprengninger på den nordlige halvkule. Utslipp av radionuklider er regulert og stort sett på et lavere nivå enn tidligere, og tilførsler til Barentshavet fra nye utslipp er relativt små. Tilførsler av radioaktive stoffer i luft overvåkes av et nasjonalt nettverk av målestasjoner og datautveksling med andre europeiske land (Møller et al. 2018).
Fordelingen av radionuklidene i dag er avhengig av parametere som havstrømmer, diffusjon, sedimenteringshastigheter og dynamikk mellom vann og sedimenter. Ulik halveringstid vil også ha betydning: Cs-137 har en fysisk halveringstid på rundt 30 år, mens Tc-99 har en halveringstid på omtrent 210 000 år. Det vil si at mesteparten av Tc-99 som er sluppet ut fortsatt er i miljøet, mens Cs-137 i miljøet fra Tsjernobyl-ulykken nå er halvert.
Det er utslipp av naturlige nuklider fra petroleumsindustrien i Nordsjøen og Norskehavet, og disse har vært ganske stabile siden utslippsrapporteringen startet (figur 4.34). Modellering viser at utslippene kan gi Barentshavet et tilleggsbidrag til de naturlige radionuklidene som allerede finnes i havområdet (Skancke og Nordam 2016).
Figur 4.34. Utslipp (GBq) av radioaktive stoffer (radium-226 og -228) fra norsk olje- og gassindustri fra 2003 til og med 2017. Kilde: Norsk olje og gass.
Strandsøppel på Svalbard Luftskipodden og Brucebukta på Svalbard ryddes årlig av Sysselmannen sine feltinspektører etter OSPAR sin metodikk, i tillegg veies strandsøppelet i Brucebukta. Det er ingen klar trend på noen av strendene for søppelmengde, og mengdene som driver i land er påvirket av lokale vær- og strømforhold.
Fra Sysselmannen sine årlige ryddeaksjoner på Svalbard rapporteres det at plastavfall fra fiskeflåten er den vanligste søppeltypen. De vanligste gjenstandene er notrester og trålposer, garnkuler av plast, bakelitt eller metall, fiskekasser, husholdningsavfall som plastdunker, flasker og glass, samt annen plastemballasje, skosåler og bruksgjenstander.
Søppel i havet stammer fra en lang rekke aktiviteter både til lands og på havet. Det som finnes på ei strand kan både skyldes lokal forsøpling og at søppel føres med havstrømmer og vind fra andre områder. Fiskeflåten i Norskehavet og Barentshavet er trolig den største kilden til strandsøppel på Svalbard, men mye kan også komme fra cruisebåter og andre fartøy.
Store mengder søppel tilføres havene hvert år mens svært lite fjernes. Gjenstander av plast, glass og gummi kan bli værende i naturen i hundrevis til tusener av år. Plastgjenstander brytes også ned til lite synlig «mikroplast» som kan tas opp av organismer. Se for øvrig avsnittet om mikroplast i avsnittet om miljøgifter i sedimentene. I tillegg ble det sammenstilt et spesialkapittel om marin forsøpling i Overvåkingsgruppens statusrapport for Norskehavet (2019), som dekket alle norske havområder.
Radioaktiv forurensning i sjøvann og i tang Radioaktiv forurensning i sjøvann fra Barentshavet overvåkes med prøver fra tokt hvert tredje år, årlige fjordtokt og vannprøver fra kyststasjonene ved Grense Jakobselv, Hillesøy, Bjørnøya, Hopen og Ny Ålesund. Generelt er nivåene av forurensning lave, og på grunn av radioaktiv nedbrytning og minkende tilførsler er også nivåene stabile og til dels synkende.
Nivåene ligger i hovedsak under 2 Bq/m3 for Cs-137. Dette er lavere enn for eksempel nivåene i Nordsjøen, som blant annet påvirkes av utstrømming fra Østersjøen som i større grad enn våre havområder ble påvirket av Tsjernobyl-ulykken. For plutoniumisotopene og americum-241 er nivåene lave.
Blæretang oppkonsentrerer Tc-99, og er en indikator for Tc-99 i Barentshavet. Det tas årlige tangprøver ved Grense Jakobselv, Indre Kiberg og Ingøy og månedlige prøver fra Hillesøy. Det var en stor økning av utslippet av Tc-99 fra Sellafield på 1990-tallet, og nivåene økte i Barentshavet. Det ble målt Tc-99 både i sjøvann og tang, og det er en klar sammenheng mellom utslipp og nivåer i Barentshavet, med en forsinkelse på rundt tre år (Karcher et al. 2004). Utslippet nå er lite og nivåene i sjøvann stort sett under deteksjonsgrensen, men nivåene i tang er fortsatt målbare.
Forurensning i sedimenter Sedimenter i åpne havområder i Barentshavet har lite miljøgifter, men langtransporterte tilførsler av bly og kvikksølv har økt de siste 100 årene. Det er få lokale kilder til forurensning i Barentshavet og utenfor Lofoten. Langtransportert forurensning med hav- og luftstrømmer er hovedkilden til forurensning i området. Det er for øvrig naturlig høye nivåer av enkelte stoffer (hydrokarboner) i nærheten av Svalbard.
Det er ikke nye data for hydrokarboner og barium i områder med letevirksomhet siden før 2017, så vi viser til forrige statusrapport for informasjon om status og trender (Arneberg og Jelmert 2017).
Polyaromatiske hydrokatboner (PAH) kan komme fra olje, men også fra andre kilder, som for eksempel forbrenning. Målinger av hydrokarboner (THC) og PAH i sedimenter utført gjennom MAREANO-programmet i 2006–2018 viste lavest nivåer nordvest for Finnmarkskysten (bakgrunnsnivå). Den sørlige delen, vest for Lofoten, hadde noe høyere nivåer, tilsvarende Klasse II (i følge Miljødirektoratets klassifiseringer for benzo[a]pyren; faktaboks 4.1). Det er også funnet noe høyere nivåer i den østlige delen av Barentshavet, som kan skyldes både naturlige forekomster av enkelte PAH-forbindelser (perylen) og oppkonsentrering av langtransporterte tilførsler i is-nedsmeltingssone. De høyeste nivåene av PAH er målt i områdene rundt Svalbard, hvor naturlige forekomster av kull fører til naturlig høye bakgrunnsnivåer av petrogene PAH (PAH som stammer fra fossile brensler).
Målinger av tungmetaller, arsen, bromerte flammehemmere, klororganiske miljøgifter og nye organiske miljøgifter i overflatesedimenter utføres flere steder langs kysten nord for polarsirkelen. Generelt sett viser målingene lave nivåer av miljøgifter. Unntaket er noe forhøyede verdier av bly og kvikksølv. Det er også funnet forhøyede nivåer av arsen i østlige og nordlige deler av Barentshavet. Prøver tatt i perioden 2016–2018 viser de samme arsennivåene fra Lofoten og nordover som tidligere. I Barentshavet er det markant høyere arsennivåer enn i Norskehavet, svarende til tilstandsklasser III og IV i Miljødirektoratets klassifisering for kyst- og fjordsedimenter (se faktaboks 4.1).
For å undersøke tidstrender er det tatt prøver av sedimentkjerner (30-40 cm lange) i dypere områder av sokkelen på skråningen ved Eggakanten utenfor Lofoten, øst i den norske delen av Barentshavet og i Storfjordrenna sør for Svalbard (figur 4.35).
Figur 4.35. Konsentrasjoner (mg/kg sediment tørrvekt) av kvikksølv, arsen og bly målt i sedimentkjerner prøvetatt ved tre ulike stasjoner, markert med svarte prikker i kartet. Dybden i sedimentkjernene representerer tid, og utvalgte år er markert.
Flere lag nedover i flere sedimentkjerner har blitt analysert og datert. Disse undersøkelsene indikerer at spesielt konsentrasjonene av tungmetallene bly og kvikksølv økte gjennom en periode fra slutten av 1800-tallet og i første del av 1900-tallet. Mens bly økte fra et bakgrunnsnivå på ca. 10 mg/kg sediment til 20–35 mg/kg sediment økte kvikksølv fra 0,01–0,02 mg/kg sediment til 0,03–0,06 mg/kg sediment. For kvikksølv kan det spores en svak nedgang de siste tjue årene i flere sedimentkjerner. Arsen øker inntil tre ganger i de øverste tre centimeter fra bakgrunnsnivå i sedimentkjernen fra Barentshavet øst. Det er usikkert om dette skyldes naturlig høyt bakgrunnsnivå kombinert med anrikningsprosesser, eller om det er knyttet til tilførsler fra menneskeskapte kilder.
Det understrekes at nivåene av både kvikksølv og bly fortsatt er lave, men at langtransportert menneskelig påvirkning kan spores. For andre tungmetaller (kadmium, kobber, krom, nikkel, sink) er nivåene generelt lave og det er liten eller ingen økning i de analyserte sedimentkjernene.
Bromerte flammehemmere (PBDE), PCB og klorerte pesticider, alkylfenoler og nye organiske miljøgifter (PFAS, dekloraner, siloksaner, fosfororganiske flammehemmere og klorparafiner) undersøkes i sedimenter i Barentshavet under MAREANO-programmet. Nivåene som er målt er som regel meget lave, like over målegrensene.
Radioaktiv forurensning i sedimenter i Barentshavet, områdene rundt Svalbard og Vesterålen og Lofoten kommer stort sett langveis fra. I 2018 varierte aktiviteten av Cs-137 i overflateprøver fra Barentshavet fra 0,8 til 4,8 Bq/kg tørrvekt (figur 4.36). Nivåene har vært ganske stabile de siste ti årene. Det er analysert for Cs-137 i 19 overflateprøver fra Barentshavet og i Svalbardfjordene Kongsfjorden og Rijpfjorden i MAREANO-programmet. Nivåene varierer fra under deteksjonsgrensen til 6 Bq/kg sediment tørrvekt. Konsentrasjonene av Cs-137 er ofte høyest langs kysten og i fjorder lenger sør (figur 4.36), områder som mottok mye radioaktivt nedfall etter Tsjernobyl-ulykken.
Området rundt ubåten Komsomolets, som sank sørvest for Bjørnøya i 1989, undersøkes hvert år. Nivåene av Cs-137 i sedimenter rundt vraket har variert fra under deteksjonsgrensen til rundt 9 Bq/kg tørrvekt. I 2018 varierte nivåene fra 4,3 til 6,3 Bq/kg tørrvekt. Dette er sammenlignbart med nærliggende havområder. I 2019 ble det gjort undersøkelser med ROV rundt vraket, og prøvene blir nå analysert. Resultater vil bli klare i løpet av 2020.
Figur 4.36. Cesium-137 (Bq/kg) i overflatesediment fra Barentshavet og utvalgte fjorder i 2018.
Mikroplast i sedimenter er analysert i overflateprøver i forskjellige deler av Barentshavet og i fjorder på Svalbard i MAREANO-programmet. Mikroplast er partikler mindre enn 5 mm. Antall partikler varierer fra 26–560 partikler per kg sediment. Det er ikke gjort analyse av mikroplast dypere i sedimentene utover overflateprøver, så det er ikke mulig å si noe om tidstrendene for mikroplast.
Forurensning i sedimenter påvirkes både av langtransportert forurensning og forurensning fra lokale kilder, samt naturlige kilder. I Barentshavet er langtransportert forurensning av størst betydning for mange miljøgifter, og for noen miljøgifter gir naturlige kilder forhøyet nivå.
Nivået av de fleste miljøgiftene i åpne havområder er lave, og gir ikke grunn til bekymring. Imidlertid er det mulig å spore en økning i tilførsel over tid av bly, kvikksølv og PAH siden slutten av 1800-tallet og begynnelsen av 1900-tallet. Økningen har sammenheng med økt menneskelig påvirkning og langtransportert forurensning. Arsen har relativt høye nivåer i Barentshavet, tilsvarende klasse III og IV (faktaboks 4.1), som kan gi toksiske effekter ved kronisk eksponering. PAH forekommer naturlig i høye konsentrasjoner noen steder rundt Svalbard.
Nivåene av radioaktiv forurensning i sediment er i stor grad på samme nivå eller lavere enn tidligere observert. Nedgangen har flere årsaker, blant annet radioaktiv nedbrytning og mindre utslipp fra viktige kilder som Sellafield og La Hague. Nivåene av naturlig forekommende stoffer i sedimenter kan få et tilleggsbidrag fra utslipp i Nordsjøen og Norskehavet.
Forurensning i blåskjell langs kysten av Nordland, Troms og Finnmark Blåskjell langs kysten av Nordland, Troms og Finnmark er stort sett lite forurenset av miljøgifter. I den grad forskerne ser noen trend for de siste 20 årene går den nedover.
Konsentrasjonene i blåskjell er mindre enn to ganger PROREF (se faktaboks 4.1) for kvikksølv, bly og diklordifenyltrikloretan ( DDT). Nivåene av kadmium, HCB og PCB i skjell fra begge stasjoner i Varangerfjorden var opp til fem ganger PROREF.
I 2018 var konsentrasjonene av PCB over miljøkvalitetsstandarden på begge stasjoner i Varangerfjorden. PBDE ble ikke undersøkt. For kvikksølv, DDT og HCB lå konsentrasjonene under miljøkvalitetsstandarden på samtlige stasjoner. Det er ikke satt miljøkvalitetsstandarder for kadmium og bly i biologisk materiale.
De tre blåskjellstasjonene som overvåkes ligger inne i fjorder, slik at resultatene herfra ikke er direkte relevante for forholdene i forvaltningsplanområdet. Stasjonene er imidlertid trolig ikke nær punktkilder.
Forurensning i reker Miljøgifter i reker analyseres både i hele reker og pillede reker (uten skall og hode). Nivåer av miljøgifter i pillede reker uten skall og hode blir vurdert opp mot grenseverdier gitt for mattrygghet, mens hele reker blir vurdert mot miljøkvalitetsstandarden satt i vanndirektivet.
Det var forholdsvis høye nivåer av kadmium i tre samleprøver av hele reker fra 2011 og 2013 (1,0, 1,6 og 1,7 mg/kg våtvekt). Alle disse prøvene var tatt relativt langt nord i Barentshavet (78 - 80 °N). I 2014–2019 var nivåene av kadmium lavere igjen, med den høyeste verdien 0,74 mg/kg våtvekt i en samleprøve tatt ved 76 °N. Kadmiumnivåene i reker kan se ut til å øke med økende breddegrad. Arseninnholdet i reker er høyt, men foreligger stort sett i lite giftige former. Arseninnholdet er høyere i pillede reker enn i hele reker, mens nivået av kobber er høyest i hele reker. Nivået av kvikksølv i hele reker i Barentshavet er like over miljøkvalitetsstandarden satt i vanndirektivet.
Nivåene av de organiske miljøgiftene dioksiner og dioksinlignende PCB, PCB, PBDE og ulike plantevernmidler er generelt lave i reker, og for mange av forbindelsene er de lavere enn bestemmelsesgrensene. Dette skyldes delvis rekenes lave fettinnhold (opp til 2,0-5,1 prosent i 2018–2019). Nivåene er noe høyere i hele reker enn i pillede reker.
Det kan se ut til å ha vært en nedgang i nivået av PCB i reker de siste årene.
Gjennomsnittsnivået av PBDE i hele reker i 2017 var det laveste siden overvåkingen startet, og nivået har vært relativt lavt de siste fire årene.
På tross av en tilsynelatende nedgang, er nivåene av PCB og PBDE fortsatt over miljøkvalitetsstandardene satt i vanndirektivet for å beskytte de mest sårbare delene av økosystemet.
Flere av plantevernmidlene viste målbare, men lave nivåer både i hele og pillede reker i 2016-2018. Blant disse var p,p'-DDE (diklordifenyldikloreten, nedbrytningsprodukt av DDT), dieldrin, toksafen og HCB. Toksafen viste de høyeste nivåene, med sum toksafen opp til 1,4 µg/kg våtvekt i hele reker, målt i 2016. Resultater for plantevernmidler for 2019 er så langt ikke tilgjengelig. Nivåene av plantevernmidler var under miljøkvalitetsstandardene for de stoffene der disse er satt, med unntak av heptaklor og heptaklorepoksid, som var over denne standarden. Av PFAS-forbindelsene var det målbare nivåer av PFOS i hele reker i 2018–2019, med høyeste målte verdi 1,0 µg/kg våtvekt. Før 2018 var bestemmelsesgrensen i metoden høyere slik at det ikke forelå målbare verdier. Nivået av PFOS er godt innenfor miljøkvalitetsstandarden.
Av målte PAH-forbindelser var det flere med målbare resultat for en eller flere prøver av hele reker i 2018–2019. Forbindelsen med høyest konsentrasjon var krysen med opp til 0,39 µg/kg våtvekt. Ingen rekeprøver hadde nivå av benzo(a)pyren over miljøkvalitetsstandarden satt i vanndirektivet.
Det er lave nivåer av radioaktiv forurensning i reker fra Barentshavet. Nivåene av Cs-137 i hele reker har generelt vært under 1 Bq/kg våtvekt i perioden 1993 til 2018.
Forurensning i lodde Lodde fra Barentshavet har generelt lave miljøgiftnivåer. Alle prøver av lodde hadde svært lave nivåer av kvikksølv og bly i 2013–2019. Nivået av kvikksølv i lodde er gjennomgående lavere enn miljøkvalitetsstandarden for kvikksølv i biota. Kadmiumnivået har vært relativt stabilt siden 2007, og i 2019 var gjennomsnittskonsentrasjonen av kadmium rundt gjennomsnittet for hele perioden.
Nivåene av de organiske miljøgiftene dioksiner og dioksinlignende PCB, sum PCB6 og PBDE i lodde var lave i 2019, som i tidligere år. Det ser ikke ut til å ha skjedd noen endring i nivåene av dioksiner og PCB siden overvåkingen startet i 2007, mens nivået av PBDE er lavere etter 2010. Nivået av PBDE i lodde er likevel høyere enn miljøkvalitetsstandarden. I 2017 og 2019 analyserte vi også for de bromerte flammehemmerne heksabromsyklododekan (HBCD) og tetrabrombisfenol A (TBBP-A). Nivået av α-HBCD var omtrent likt nivået av sum PBDE7, mens TBBP-A var under målbart nivå.
Lodde har målbare nivåer av mange ulike klorerte plantevermidler, blant andre HCB, dieldrin, toksafen, klordan og α-HCH (α-heksaklorsykloheksan). Nivået av HCB var langt under miljøkvalitetsstandarden. Andre klorerte plantevernmidler som det er miljøkvalitetsstandarder for er endosulfan, heptaklor og heptaklorepoksid samt DDT. Nivået av heptaklor og heptakloroksid er over miljøkvalitetsstandarden, mens de øvrige stoffene er godt under.
Blant annet fordi analysemetoden har endret seg, er det for en del plantevernmidler ikke mulig å vurdere om det har vært noen reell endring i innholdet av plantevernmidler i lodde over tid. For cis-klordan har nivåene i lodde fra og med 2014 vært lavere enn tidligere.
Nivået av alle målte PFAS-forbindelser er lavere enn bestemmelsesgrensene Det betyr at alle prøver er godt innenfor miljøkvalitetsstandarden satt for PFOA. I 2013–2019 er det også analysert for PAH-forbindelser i lodde, der bare noen få av stoffene viste målbare, men lave, nivåer.
Nivåene av radioaktiv forurensning i lodde er i stor grad på samme nivå eller lavere enn tidligere observert. I 2018 viste analysene nivåer av Cs-137 under 0,1 Bq/kg våtvekt. Det har vært en generell nedgang i nivåene av radioaktiv forurensning i det marine miljø siden tidlig på 1990-tallet. Se også mattrygghets-kapittelet.
Forurensning i polartorsk Nivåene av bly og kvikksølv i polartorsk var svært lave i 2019, som i tidligere år. De fleste årene vi har data for, har nivåene enten vært under eller nær bestemmelsesgrensene. Kvikksølvnivået var under miljøkvalitetsstandarden. Nivået av arsen i polartorsk har siden 2006 variert mye uten noen økende eller avtakende trend, og nivået i 2019 var rundt gjennomsnittet for hele perioden. Kobbernivået er lavt i polartorsk og i 2019 var nivået blant de laveste som er målt i denne overvåkningen (siden 2006).
Nivåene av organiske miljøgifter i samleprøver av hel polartorsk var svært lave fra 2006 til 2019. Det var ingen klar økende eller avtakende trend i perioden, med mulig unntak av PBDE, som ut fra disse resultatene kan se ut til å ha avtatt. Nivået av sum PBDE7 var imidlertid i 2019 noe høyere enn i årene 2013–2018. Likevel er nivået av PBDE i polartorsk klart over miljøkvalitetsstandarden. I 2017 og 2019 ble polartorsk også analysert for HBCD og TBBP-A. Nivået av α-HBCD var i 2019 noe høyere enn nivået av sum PBDE7, mens TBBP-A var under målbart nivå.
Blant analyserte plantevernmidler har prøvene av polartorsk hatt målbare nivåer av HCB og dieldrin i hele perioden 2006-2018 (ikke analysert i 2010). Nivået av HCB var betydelig høyere i 2017 og 2018 enn tidligere år, men langt under miljøkvalitetsstandarden. Nivået av HCH har vært under eller så vidt over bestemmelsesgrensene, og enkelte klordaner, DDT og toksafener har vært over bestemmelsesgrensene fra og med 2011 (da analysemetoden ble bedre). Heptaklor og heptaklorepoksid var godt over miljøkvalitetsstandarden i hele perioden 2011-2018. Blant annet fordi det er brukt ulike analysemetoder med ulike bestemmelsesgrenser, kan man for en del plantevernmidler ikke sammenligne resultatene direkte fra år til år.
Nivåene av PFAS målt i hel polartorsk var under eller like over bestemmelsesgrensene i hele perioden til og med 2019, og nivået av PFOA var under miljøkvalitetsstandarden satt for dette stoffet.
Polartorsk ble analysert for PAH-forbindelser fra og med 2014. Få av prøvene hadde målbare nivåer av forbindelsene og benzo(a)pyren og fluoranten var langt under miljøkvalitetsstandardene som er satt for disse.
Nivåene av radioaktiv forurensning i polartorsk i 2018 var i stor grad på samme nivå eller lavere enn tidligere observert. Se også kapittelet om mattrygghet.
Forurensning i torsk I Barentshavet finner vi både kysttorsk og nordøstarktisk torsk, og denne rapporten inkluderer resultater for begge typer torsk. Tre av de fire kysttorsk-stasjonene ligger inne i fjorder eller nær punktkilder (områdene ved Tromsø havn og Hammerfest havn, og Kjøfjord i ytre Varangerfjorden), slik at resultatene herfra ikke er direkte relevante for forholdene i forvaltningsplanområdet.
Nivåene av fettløselige miljøgifter som PCB, PBDE og DDT i torsk er høyere i lever enn i filet. Det skyldes at torskefilet har lite fett (< 1 %), mens torskelever har mye fett (rundt 50 %). I kysttorsk måles derfor kvikksølv i filet, mens de fettløselige miljøgiftene måles i lever. Konsentrasjonene av miljøgifter i kysttorsk fra Nordland, Troms og Finnmark er stort sett lave; mindre enn to ganger PROREF (se faktaboks 4.1). I 2018 var konsentrasjonene av kvikksølv, PCB og PBDE likevel over miljøkvalitetsstandarden på samtlige stasjoner, mens for DDT og HCB lå konsentrasjonene under miljøkvalitetsstandarden på samtlige stasjoner.
Det er en oppadgående trend for bly i kysttorsk ved to stasjoner, og ellers er det nedgang for PCB ved en stasjon og for PBDE ved en annen stasjon.
Gjennomsnittsnivået av kvikksølv i nordøstarktisk (NØA) torsk prøvetatt i åpent hav i Barentshavet er under PROREF. I den østlige delen av Barentshavet er kvikksølvnivået i torskemuskel under miljøkvalitetsstandarden, mens i den vestlige delen og totalt for Barentshavet er det over miljøkvalitetsstandarden. Kvikksølvnivået i NØA torsk i Barentshavet har vært stabilt i perioden siden overvåkningen startet i 1994.
Kadmiumnivået i torskelever er høyere enn i torskefilet, fordi kadmium hoper seg opp i organer som lever og nyrer. Kadmiumnivået i lever av torsk lå over PROREF i de nordligste områdene. Nivået av bly er svært lavt i torskelever, og under PROREF på alle stasjoner.
Nivået av sum dioksiner og dioksinlignende PCB i torskelever var nokså stabilt fra 2006 til 2016, og i 2017–2019 noe lavere enn tidligere. De tre siste årene var gjennomsnittsverdiene like under miljøkvalitetsstandarden. I motsetning til lever inneholder den magre torskefileten svært lave konsentrasjoner av dioksiner og dioksinlignende PCB, og filet overvåkes ikke regelmessig for disse stoffene. At nivåene i torskefilet fortsatt er svært lave, ble imidlertid bekreftet da i alt 20 torskeprøver prøvetatt i 2018–2019 ble analysert for dioksiner og dioksinlignende PCB og andre organiske miljøgifter.
Konsentrasjonene av ikke-dioksinlignende PCB (PCB7/PCB6) og PBDE er også generelt mye høyere i torskelever enn i torskefilet. Nivået av PCB7 er mye lavere enn PROREF, men er likevel over miljøkvalitetsstandarden. Det har vært en nedadgående trend i PCB-nivå fra 1990-tallet til i dag. Fra 2006 og fremover har PCB-nivået i torskelever avtatt mindre, men likevel statistisk signifikant i det østligste området. Nivåene av PBDE målt i lever av nordøstarktisk torsk i 2016–2019 var de laveste siden målingene startet i 2006, og det er en nedadgående trend. Den nedadgående trenden er bare signifikant for det østligste området. Nivået av PBDE er godt under PROREF, men høyere enn miljøkvalitetsstandarden.
Torskefilet har svært lave nivåer også av plantevernmidler og PFAS. Siden henholdsvis 2008 og 2009 har kun lever av nordøstarktisk torsk blitt analysert for disse stoffene. I 2018 var det lave nivåer av PFAS i torskelever, med de fleste analyserte forbindelsene under bestemmelsesgrensene i de fleste prøvene, med noen unntak: To av 49 prøver hadde PFOA over bestemmelsesgrensen, en hadde PFTrDA (perfluortridekylkarboksylsyre) og 14 hadde PFUdA (perfluorundekylkarboksylsyre) over bestemmelsesgrensen. De målte nivåene var lave.
Av plantevernmidlene som det er analysert for i lever av NØA torsk, er det DDT og dets metabolitter som forekommer i høyest konsentrasjon. Sum DDT er likevel de siste årene godt under både PROREF og miljøkvalitetsstandarden. Nivået av HCB er noe lavere enn DDT, men likevel over miljøkvalitetsstandarden og en til to ganger PROREF. Mens DDT ser ut til å ha hatt en nedadgående trend siden tidlig på 2000-tallet, har nivået av HCB vært omtrent det samme siden 1992. Andre plantevernmidler som i 2017 og 2018 forekom i målbare konsentrasjoner i de fleste prøver av torskelever fra Barentshavet, var: toksafener, klordaner, nonaklor, cis-heptaklorepoksid, dieldrin, α-HCH, pentaklorbenzen og mirex.
Nivået av γ-HCH (lindan) var både i 2017 og 2018 under bestemmelsesgrensene for de fleste prøver, og høyeste målte verdi var langt under både miljøkvalitetsstandarden og PROREF (se tabell 6.4 for oppsummering).
Når det gjelder radioaktiv forurensning i det marine miljø, har det vært en generell nedgang i nivåene siden 1990-tallet. Dette gjelder også nivåene av radioaktiv forurensning i torsk (se også kapittel 4.8.3 om mattrygghet).
Forurensning i sjøfugl Indikatorene for forurensning i sjøfugl i Barentshavet inkluderer polarlomvi, polarmåke og krykkje, der det er eggene som overvåkes. Det er ikke oppdaterte data på disse indikatorene, og vi henviser til forrige statusrapport fra 2017 for status og trend for miljøgifter i sjøfugl.
Forurensning i ringsel Det er ikke oppdaterte data for forurensning i ringsel, og vi henviser til forrige statusrapport fra 2017 for status og trend for miljøgifter i ringsel (Arneberg og Jelmert 2017).
Forurensning i isbjørn Miljøgiftbelastingen i isbjørn fra Svalbard er dominert av fettløselige organiske miljøgifter, deres nedbrytningsprodukter (metabolitter) og perfluorerte forbindelser, mens nivåer av nye kommersielle kjemikalier er lave hos isbjørn. Isbjørn er utsatt for høye nivåer av miljøgifter som er tungt nedbrytbare og øker i konsentrasjon oppover i næringskjeden. Overvåking av isbjørn viser at konsentrasjonene av fettløselige organiske miljøgifter (PCBer, β-HCH, oksyklordan og PBDE-47) har gått ned mellom 1997 og 2017. Dette er i samsvar med de trendene man ser i flere andre arktiske dyr, og er en bekreftelse på at internasjonale reguleringer av disse stoffene har vært vellykkede. Konsentrasjonene av DDE og HCB gikk ned frem til 2010, og deretter økte konsentrasjonene frem til 2017 (21 % per år for DDE og 11 % per år for HCB). Nivået av PFOS, det perfluorerte stoffet som har vært regulert lengst, avtok med 14 % per år i perioden 2003-2009, men har vært stabilt siden. I motsetning til dette økte nivåene av nyere perfluorerte stoffer med rundt to prosent per år i perioden 2000-2014.
Pelagisk isbjørn fra Barentshavet, som følger isen mot øst når isen rundt Svalbard smelter på sommeren, har høyere inntak av organiske miljøgifter sammenlignet med kystisbjørn som blir ved Svalbard gjennom hele året (Blévin et al. 2020) . Det er flere årsaker til disse forskjellene. For eksempel spiser pelagisk isbjørn en høyere andel marine byttedyr, og på et høyere trofisk nivå enn kystisbjørnene. De har også høyere energikrav og dermed et høyere inntak av byttedyr. I tillegg spiser pelagisk isbjørn en høyere andel byttedyr som de fanger i den marginale issonen og byttedyr som ligger nærmere forurensende utslippskilder/transportveier. Til tross for høyere energiforbruk var pelagisk isbjørn fetere sammenlignet med kystbjørnene. Dette skyldes mest sannsynlig høyt inntak av sel gjennom hele året. Selv om inntaket av miljøgifter er høyere hos pelagisk isbjørn enn hos kystisbjørn, har de like konsentrasjoner av fettløselige miljøgifter. Dette er fordi pelagiske individer har en større mengde fett hvor fettløselige forbindelser lagres.
Konsentrasjonene av de forskjellige miljøgiftene varierer mellom de ulike isbjørnpopulasjonene i Arktis. Isbjørn fra Barentshavet viser høyere nivåer av perfluorerte stoffer enn isbjørn fra øst-Grønland eller Canada. Nivåer av PCBer og klorerte plantevernmidler i isbjørn er i noen områder høyere og i andre områder lavere sammenlignet med Barentshavet. Nivåer av kvikksølv er generelt lave i isbjørn fra Barentshavet.
Nivåene av miljøgifter er fremdeles høye i isbjørn, og isbjørnunger har vist seg å ha over dobbelt så høye nivåer av fettløselige miljøgifter sammenlignet med isbjørnbinner. Effektstudier på isbjørn tyder på at miljøgiftbelastningen kan påvirke aktiviteten av viktige molekyler i hjernen, immunforsvaret og hormoner som er viktige for utviklingsprosesser og energimetabolisme. Miljøgifter har også evne til å forstyrre fettlagrings- og fettforbrenningsprosesser hos isbjørn.
De høye nivåene av miljøgifter i isbjørn, og spesielt i isbjørnunger, er urovekkende da disse stoffene kan påvirke utvikling, samt kan gjøre isbjørn mer mottakelig for smitte og sykdom. Perioder med sult og tæring på kroppsfettet er naturlig for isbjørnen, men kan være kritisk fordi miljøgifter som er lagret i fettvevet blir mer konsentrert og har evne til å påvirke lagring og forbrenning av fett. De blir også frigjort til blodet når fett forbrennes. Miljøgiftene blir da tilgjengelige og tas opp i organer som lever og hjerne.
4.8.2 - Vurdering av nivåer av forurensende stoffer med hensyn til mattrygghet
For å vurdere nivåer av forurensende stoffer med hensyn til mattrygghet vurderes nivåer av miljøgifter og radioaktive stoffer i spiselige deler, som kokte, pillede reker, filet av fisk og lever av mager fisk. Nivåene av forurensende stoffer blir vurdert opp mot grenseverdier for mattrygghet (se faktaboks 4.1). For sjømat finnes det grenseverdier som gjelder tungmetallene bly, kadmium og kvikksølv samt for de fettløselige organiske miljøgiftene dioksiner og dioksinlignende PCB og ikke-dioksinlignende PCB (PCB6).
For filet av magre fiskeslag er det stort sett bare én miljøgift som i noen tilfeller gir problemer i forhold til mattrygghet, og det er kvikksølv. Både kysttorsk fra kysten av Troms og Finnmark og nordøstarktisk torsk fra Barentshavet har vist kvikksølvnivåer godt under grenseverdien for mattrygghet i alle prøver. Andre fiskearter i Barentshavet som i perioden 2016-2019 har vært undersøkt for miljøgifter i forbindelse med to ulike kartleggingsprogrammer, er vanlig uer og snabeluer, samt rødspette prøvetatt i de tilstøtende kyst- og fjordområdene (Frantzen et al. in press; Nilsen et al. 2020). Miljøgifter i sei og blåkveite overvåkes jevnlig (upublisert). Disse artene hadde stort sett nivåer av kvikksølv i filet godt under grenseverdien for mattrygghet. Arten som har vist høyest gjennomsnittlig kvikksølvnivå i filet i Barentshavet er blåkveite, men kun enkeltfisk har hatt konsentrasjoner over grenseverdi i Barentshavet. Generelt har arter på høyt nivå i næringskjeden høyest kvikksølvnivå. Men også andre faktorer, som alder, størrelse og veksthastighet, gir variasjoner i kvikksølvnivå mellom arter.
Kadmium og bly akkumuleres i liten grad i muskel, og nivåene i filet av torsk og de fleste andre fiskeslag er svært lave.
Mager fisk har svært lave nivåer av fettløselige organiske miljøgifter i filet, da disse hoper seg opp i leveren sammen med fettet. Fetere fiskearter som blåkveite, uer og gråsteinbit har litt høyere nivå av organiske miljøgifter i filet enn de helt magre fiskeartene, men ingen prøver av fiskefilet fra Barentshavet analysert de siste årene har hatt nivåer av dioksiner og dioksinlignende PCB eller PCB6 over grenseverdien for mattrygghet (faktaboks 4.1).
Lever av fisk kan ha relativt høye nivåer av organiske miljøgifter på grunn av det høye fettinnholdet og fordi lever kan ha en funksjon som avgiftningsorgan. Siden fiskelever spises i mye mindre grad enn filet, har EU og Norge satt høye grenseverdier for summen av dioksiner og dioksinlignende PCB og PCB6 i fiskelever (EU, 2018). I lever av fisk fra Barentshavet er gjennomsnittsnivåene av disse stoffene under grenseverdiene i alle artene som har vært analysert i perioden 2016-2018.
Krepsdyr som reker og krabbe kan akkumulere høye nivåer av kadmium i fordøyelseskjertelen, og nivåene er høyere i reker fra Barentshavet enn i reker fra Nordsjøen og kysten av Norskehavet. For taskekrabbe er det vist klart høyere kadmiumnivåer langs kysten av Nord-Norge enn lenger sør (Wiech et al. 2020). Hele reker fra Barentshavet har relativt høyt nivå av kadmium, men vi mennesker spiser som oftest bare pillede reker, som er ren muskel, og grenseverdien for mattrygghet gjelder kun for muskel av krepsdyr (EU, 2018). Nivået av kadmium i pillede reker er godt under denne grenseverdien. Også snøkrabbe analysert i 2014 og kongekrabbe analysert i 2012 hadde kadmiumnivåer i muskel fra gangbein under grenseverdien (Frantzen og Måge 2016; Julshamn et al. 2015). Nivåene av kvikksølv og bly er lave og godt under grenseverdiene i både reker, snøkrabbe og kongekrabbe. Også nivåene av organiske miljøgifter er lave og langt under grenseverdiene i reker og kongekrabbe. Organiske miljøgifter ble ikke målt i snøkrabbe (Frantzen og Måge 2016) .
Det er en grenseverdi på 600 Bq/kg for Cs-137 for de fleste matvarer, og nivåene i sjømat ligger langt under dette. Nivåene av naturlige radioaktive stoffer i sjømat, i hovedsak polonium-210, vil gi et større dosebidrag til befolkningen enn radioaktiv forurensning i sjømat (Komperød et al. 2015) .
4.9 - Påvirkning fra aktivitet i forvaltningsplanområdet
Fiskeriene påvirker direkte bestander i Barentshavet. Gjennom høstingsregelen som settes sammen med Russland, bidrar dette til en moderat, og under- grensen grad av fiskedødelighet. Samtidig utvikler trålfiskeriene redskap som i mindre grad skal ødelegge bunnen og teknologi for å vurdere om de skal sette redskap eller ikke. Kvotene blir derfor fisket opp på stadig færre tråltimer og dermed mindre miljøpåvirkning.
4.9.1 - Fiskedødelighet i Barentshavet
Gjennom historien har fiskeriene hatt en klar påvirkning på de kommersielle bestandene med ringvirkninger til økosystemet. Estimert fiskedødelighet, som er den andelen av bestanden som fiskeriene tar ut av totalbestanden, er en viktig faktor i beregningene som ligger til grunn for fiskerirådene. Fiskedødelighet for torsk, hyse og snabeluer i Barentshavet er på bærekraftige nivåer for alle bestandene og har vært det i mange år (figur 4.37).
Figur 4.37. Fiskedødelighet for NØA torsk, NØA hyse og snabeluer. Fisket regnes som bærekraftig så lenge F er negativ i forhold til vedtatt fiskedødelighetsgrense (0). Dataene er hentet fra beregningene fra det internasjonale havforskerrådet ICES (ICES 2020).
4.9.2 - Trålaktivitet i Barentshavet
a)
b)
Figur 4.38. Samlet antall tråltimer (a) og fangst (b) med norske trålere i Barentshavet. Kilde: Fiskeridirektoratet
Figuren over viser det samlede antall tråltimer, målt fra når trålen settes i sjøen til den trekkes samt fangst (figur 4.38 a og b). Den dekker i all hovedsak innsatsen i det norske trålfisket i Barentshavet etter kvotebelagte arter som torsk, sei, hyse, snabeluer og reke, inklusive bifangst av andre arter. Antall tråltimer som norske trålere brukte på fiske i Barentshavet sank fra en topp på vel 480 000 timer i begynnelsen av perioden til vel 81 000 timer i 2013. Innsatsen har økt noe de siste årene og lå i 2016 på et nivå i underkant av 119 000 timer. Innsatsen i trålfisket etter reke kan under noe bedre økonomiske betingelser muligens øke noe .
Nedgangen fra 1987 til 2013 skyldes i første rekke en sterk reduksjon i antall trålere, bl.a. som en følge av strukturtiltak for å redusere overkapasitet i denne flåten. Flåten består i dag av vesentlig større og mer moderne fartøy sammenlignet med 1987. Etter 2000 har nedgangen også sammenheng med redusert innsats i trålfisket etter reke, noe som skyldes redusert lønnsomhet grunnet lavere priser og økte drivstoffutgifter.
Tallene fra 1987 til 2009 er basert på innsendte fangstdagbøker fra norske trålere. Grunnlaget for trålstatistikken ble endret i 2010 ved at den manuelle innsamlingen av fangstdagbøker i løpet av året ble erstattet av elektronisk innsendte fangstopplysninger. Problemer i forbindelse med innføring av elektronisk fangstdagbok i 2010 førte til at datagrunnlaget ble såpass mangelfullt at en har valgt å se vekk fra dette året. Ordningen med elektronisk fangstdagbok omfatter litt flere trålere fordi en del mindre reketrålere som bare sporadisk sendte inn papirutgaver av fangstdagboken, nå er kommet med.
Siden 1987 har det også vært en betydelig teknologisk utvikling når det gjelder leteutstyr (sonarer og ekkolodd). Dette har gitt en betydelig økning i fangsteffektiviteten, og som også har bidratt til å redusere antall tråltimer, ved at det brukes mindre tid til å fiske kvotene.
5 - Særlig verdifulle og sårbare områder
Som et bidrag til arbeidet med vurdering av særlig verdifulle og sårbare områder (SVO), er det her gitt en kort oppsummering av status for de elementene som ligger til grunn for utpeking av området som et SVO og som Overvåkingsgruppen har indikatorer for (tabell 5.1). Oppsummeringene er basert på kapittel 4.
Område
Indikator
Status og endringer i status siden 2017
Havområdene utenfor Lofoten til Tromsøflaket, inkludert eggakanten.
Nordøstarktisk torsk
Bestanden har minket noe siden 2013, men er fortsatt på et høyt nivå. Den er fremdeles utbredt over store deler av Barentshavet.
Norsk vårgytende sild
Bestandsstørrelsen har avtatt siden 2009 på grunn av dårligere rekruttering.
Vanlig uer
Bestanden er på det laveste nivået som noen gang har vært målt. Arten er vurdert som sterkt truet i Rødlisten.
Snabeluer
Bestanden er restituert til bærekraftig nivå og er i vekst.
Krykkje
Arten er i sterk nedgang i kolonier langs fastlandskysten og er vurdert som sterkt truet i Rødlisten. Koloniene på Svalbard har holdt seg stabile eller kun vist en svak nedgang siste 10 år.
Lomvi
Bestanden har minket betydelig og står i fare for å forsvinne som hekkefugl fra store deler av fastlandskysten. Bestanden øker øst i Finnmark og på Bjørnøya.
Lunde
Fra 2009 til 2019 har det vært fullstendig hekkesvikt i kolonien på Røst, og bestanden minker. På Anda har det vært rekruttering, men nedgang i bestanden.
Tromsøflaket
Nordøstarktisk torsk
Bestanden har minket noe siden 2013, men er fortsatt på et høyt nivå. Den er fremdeles utbredt over store deler av Barentshavet.
Norsk vårgytende sild
Bestandsstørrelsen har avtatt siden 2009 på grunn av dårligere rekruttering.
Kystnære områder for øvrig – fra Tromsøflaket til grensen mot Russland.
Krykkje
Arten er i sterk nedgang i kolonier langs fastlandskysten og er vurdert som sterkt truet i Rødlisten.
Lomvi
Bestanden har minket betydelig og står i fare for å forsvinne som hekkefugl fra store deler av fastlandskysten. Bestanden øker øst i Finnmark og på Bjørnøya.
Lunde
Hekkebestanden på Gjesvær har holdt seg stabil, mens den har vært svakt negativ på Hornøya.
Iskanten
Isbjørn
Det er ikke tegn på at bestanden minker. Den kan være i vekst etter fredning, men mot en bæreevne som avtar på grunn av klimaendringene.
Lodde
Bestanden har gått betydelig ned siden 2017 og er på et lavt nivå.
Krykkje
Arten er i sterk nedgang i kolonier langs fastlandskysten og er vurdert som sterkt truet i Rødlisten. Koloniene på Svalbard har holdt seg stabile eller kun vist en svak nedgang siste 10 år.
Lomvi
Bestanden har minket betydelig og står i fare for å forsvinne som hekkefugl fra store deler av fastlandskysten. Bestanden øker øst i Finnmark og på Bjørnøya.
Polarfronten
Lodde
Bestanden har gått betydelig ned siden 2017 og er på et lavt nivå.
Polarlomvi
Bestanden er i nedgang i hele forvaltningsplanområdet, sannsynligvis som følge av dårligere næringstilgang i overvintringsområdet rundt Island.
Lomvi
Bestanden har minket betydelig og står i fare for å forsvinne som hekkefugl fra store deler av fastlandskysten. Bestanden øker øst i Finnmark og på Bjørnøya.
Nordøstarktisk torsk
Bestanden har minket noe siden 2013, men er fortsatt på et høyt nivå. Den er fremdeles utbredt over store deler av Barentshavet.
Havområdene rundt Svalbard, inkludert Bjørnøya.
Krykkje
Koloniene på Svalbard har holdt seg stabile eller kun vist en svak nedgang siste 10 år.
Lomvi
Bestanden har minket betydelig og står i fare for å forsvinne som hekkefugl fra store deler av fastlandskysten. Bestanden øker øst i Finnmark og på Bjørnøya.
Polarlomvi
Bestanden er i nedgang i hele forvaltningsplanområdet, sannsynligvis som følge av dårligere næringstilgang i overvintringsområdet rundt Island.
Tabell 5.1 Kort oppsummering av status for elementer som ligger til grunn for identifisering av de særlig verdifulle og sårbare områdene (SVO) i forvaltningsplanen for Barentshavet og havområdene utenfor Lofoten og som Overvåkingsgruppen har indikatorer for.
6 - Indikatorliste og vurdering av indikatorverdier i forhold til referan severdier og tiltaksgrenser
Dette kapittelet inneholder en vurdering av indikatorverdier i forhold til referanseverdier og tiltaksgrenser der slike er satt. I slutten av kapittelet er det også gitt en liste over alle indikatorene for Barentshavet som er i bruk av Overvåkingsgruppen.
6.1 - Indikatorer for fisk
Navn på indikator
Referanseverdier
Tiltaksgrenser*
Status til henhold til tiltaksgrenser 2019
Lodde i Barentshavet
Føre-vargrense for gytebestand
Hvis beregnet gytebestand er mindre enn føre-vargrensen.
Denne modne delen av bestanden er under føre-vargrensen og bestandsnivået gir ikke grunnlag for å åpne loddefiske i 2020.
Kolmule i Barentshavet
Føre-vargrense for gytebestand
Hvis beregnet gytebestand er mindre enn føre-vargrensen.
Etter 2004 har det vært en klar nedgang. Fra 2008 har mengde kolmule variert på et lavt nivå, men med en topp i 2016.
Nordøstarketisk torsk i Barentshavet
Føre-vargrense for gytebestand
Hvis beregnet gytebestand er mindre enn føre-vargrensen.
Gytebestanden i 2019 er lang over tiltaksgrensen. Gytebestanden er viktig for å sikre god rekruttering.
Blåkveite
Føre-vargrense for bestand
Hvis beregnet bestand er mindre enn føre-vargrensen.
Bestanden er beregnet å forbli under beregnet føre-var nivå de nærmeste årene, selv om trenden i biomasse (fangstbar mengde) har flatet ut de aller siste årene og var svakt nedadgående i 2016.
Vanlig uer
Føre-vargrense for bestand
Hvis beregnet bestand er mindre enn føre-vargrensen.
Det er en klar reduksjon i bestanden av vanlig uer, og bestanden er nå på det laveste nivået som noen gang har vært målt.
Snabeluer
Føre-vargrense for bestand
Hvis beregnet bestand er mindre enn føre-vargrensen.
Snabeluerbestanden har blitt restitutert til et bærekraftig reproduksjonsnivå. Rekrutteringen var svak fra 1996 til 2004, men har blitt klart styrket etter dette.
Fiskedødelighetsrate
Føre-vargrense for fiskedødelighetsrate
Hvis beregnet fiskeridødelighet er mindre enn føre-vargrensen.
Beregnet fiskedødelighet er under, til dels godt under føre-vargrensen.
Tabell 6.1. Vurdering av indikatorverdier for 2019 i forhold til referanseverdier og tiltaksgrenser for indikatorer for fiskebestander i Barentshavet.
6.2 - Indikatorer for sjøfugl
Navn på indikator
Tiltaksgrenser
Status til henhold til tiltaksgrenser 2019
Krykkje i Barentshavet
Det er ikke ønskelig med en nedgang i bestanden på 20% eller mer over fem år, eller at hekking mislykkes fem år på rad.
De siste fem årene har det vært betydelig hekkesvikt i mange kolonier langs fastlandskysten hvor vi også ser en nedgang i bestanden
Polarlomvi i Barentshavet
Det er ikke ønskelig med en nedgang i bestanden på 20% eller mer over fem år, eller at hekking mislykkes fem år på rad.
Flere av de overvåkede koloniene har hatt en nedgang på mer enn 20% de siste fem årene.
Lomvi i Barentshavet
Det er ikke ønskelig med en nedgang i bestanden på 20% eller mer over fem år, eller at hekking mislykkes fem år på rad.
De siste fem årene har det vært betydelig hekkesvikt i mange kolonier langs fastlandskysten hvor vi også ser en nedgang i bestanden
Lunde i Barentshavet
Det er ikke ønskelig med en nedgang i bestanden på 20% eller mer over fem år, eller at hekking mislykkes fem år på rad.
Det har vært hekkesvikt og nedgang bestandene i flere kolonier langs fastlandskysten i de siste fem årene.
Romlig fordeling av sjøfugl i Barentshavet
Det er ikke ønskelig med et avvik på mer enn 10% i forhold til forventet utbredelse.
For noen arter er tyngdepunkt i utbredelse forskjøvet i nordlig retning med mer enn 10% av avstanden mellom sørligste og nordligste tyngdepunkt for alle arter.
Tabell 6.2. Vurdering av indikatorverdier for 2019 i forhold til tiltaksgrenser for indikatorer for sjøfugl i Barentshavet.
6.3 - Indikatorer for sårbare og truede arter
Navn på indikator
Referanseverdier
Tiltaksgrenser*
Status til henhold til tiltaksgrenser 2019
Fremmede arter i Barentshavet
Arter som er identifisert i Rødlisten for sårbare eller truede arter.
Ingen sårbare eller truede arter bør forsvinne fra Barentshavet.
Ingen arter som er identifisert i Rødlisten for sårbare og truede arter er forsvunnet fra Barentshavet.
Tabell 6.3. Vurdring av indikatorverdi for 2019 i forhold til referanseverdier og tiltaksgrenser for indikatoren for truede og sårbare arter i Barentshavet.
6.4 - Indikatorer for forurensende stoffer
Tabell 6.4. Oversikt over de ulike indikatorartene som overvåkes og andel overskridelser ved siste måling av tre ulike typer grenseverdier for ulike stoffer (faktaboks 4.1): Grenseverdier for mattrygghet, miljøkvalitetsstandarder og PROREF (kun torsk og blåskjell). Forklaring på fargekodene er gitt nedenfor.
Aarflot, J. M., H. R. Skjoldal, P. Dalpadado & M. Skern-Mauritzen, 2017. Contribution of Calanus species to the mesozooplankton biomass in the Barents Sea. ICES Journal of Marine Science 75(7):2342-2354 doi:10.1093/icesjms/fsx221.
Aars, J., 2013. Variation in detection probability of polar bear maternity dens. Polar Biology 36(8):1089-1096 doi:10.1007/s00300-013-1331-7.
Aars, J., T. A. Marques, S. T. Buckland, M. Andersen, S. Belikov, A. Boltunov & O. Wiig, 2009. Estimating the Barents Sea polar bear subpopulation size. Marine Mammal Science 25(1):35-52 doi:10.1111/j.1748-7692.2008.00228.x.
Aars, J., T. A. Marques, K. Lone, M. Andersen, Ø. Wiig, I. D. B. Fløystad, S. B. Hagen & S. T. Buckland, 2017. The number and distribution of polar bears in the western Barents Sea. Polar Research 36(1) doi:10.1080/17518369.2017.1374125
Ahonen, H., K. M. Stafford, L. de Steur, C. Lydersen, O. Wiig & K. M. Kovacs, 2017. The underwater soundscape in western Fram Strait: Breeding ground of Spitsbergen's endangered bowhead whales. Mar Pollut Bull 123(1-2):97-112 doi:10.1016/j.marpolbul.2017.09.019.
Ahonen, H., K. M. Stafford, C. Lydersen, L. de Steur & K. M. Kovacs, 2019. A multi-year study of narwhal occurrence in the western Fram Strait - detected via passive acoustic monitoring. . Polar Research 38,doi:1-14 doi:10.33265/polar.v38.3468.
AMAP, 2011a. AMAP Assessment 2011: Mercury in the Arctic. Arctic Monitoring and Assessment Programme (AMAP), Oslo, Norway. xiv + 193 pp.
AMAP, 2011b. Combined Effects of Selected Pollutants and Climate Change in the Arctic Environment. By: R. Kallenborn, K. BorgÃ¥, J.H. Christensen, M. Dowdall, A. Evenset, J.Ã. Odland, A. Ruus, K. Aspmo Pfaffhuber, J. Pawlak, and L.-O. Reiersen. AMAP Technical Report No. 5 (2011). Arctic Monitoring and Assessment Programme (AMAP), Oslo. 108 pp.
AMAP, 2016. Influence of Climate Change on Transport, Levels, and Effects of Contaminants in Northern Areas – Part 2. By: P. Carlsson, J.H. Christensen, K. Borgå, R. Kallenborn, K. Aspmo Pfaffhuber, J.Ø. Odland, L.-O. Reiersen, and J.F. Pawlak. Arctic Monitoring and Assessment Programme (AMAP), Oslo. 52 pp.
Andersen, L. W., C. Lydersen, A. K. Frie, A. Rosing-Asvid, E. Hauksson & K. M. Kovacs, 2011. A population on the edge: genetic diversity and population structure of the world's northernmost harbour seals (Phoca vitulina). Biol J Linnean Soc 102(2):420-439 doi:10.1111/j.1095-8312.2010.01577.x.
Anker-Nilssen, T., 1992. Food supply as a determinant of reproduction and population development in Norwegian Puffins Fratercula arctica. PhD-thesis. University of Trondheim.
Ardyna, M., M. Babin, M. Gosselin, E. Devred, S. Belanger, A. Matsuoka & J. E. Tremblay, 2013. Parameterization of vertical chlorophyll a in the Arctic Ocean: impact of the subsurface chlorophyll maximum on regional, seasonal, and annual primary production estimates. Biogeosciences 10(6):4383-4404 doi:10.5194/bg-10-4383-2013.
Ardyna, M., M. Babin, M. Gosselin, E. Devred, L. Rainville & J.-É. Tremblay, 2014. Recent Arctic Ocean sea ice loss triggers novel fall phytoplankton blooms. Geophys Res Lett 41(17):6207-6212 doi:10.1002/2014gl061047.
Arneberg, P. & A. Jelmert, 2017. Status for miljøet i Barentshavet - rapport fra Overvåkingsgruppen Fisken og Havet. Havforskningsinstituttet.
Arrigo, K. R., D. K. Perovich, R. S. Pickart, Z. W. Brown, G. L. van Dijken, K. E. Lowry, M. M. Mills, M. A. Palmer, W. M. Balch, F. Bahr, N. R. Bates, C. Benitez-Nelson, B. Bowler, E. Brownlee, J. K. Ehn, K. E. Frey, R. Garley, S. R. Laney, L. Lubelczyk, J. Mathis, A. Matsuoka, B. G. Mitchell, G. W. K. Moore, E. Ortega-Retuerta, S. Pal, C. M. Polashenski, R. A. Reynolds, B. Schieber, H. M. Sosik, M. Stephens & J. H. Swift, 2012. Massive Phytoplankton Blooms Under Arctic Sea Ice. Science 336(6087):1408-1408 doi:10.1126/science.1215065.
Arrigo, K. R., D. K. Perovich, R. S. Pickart, Z. W. Brown, G. L. van Dijken, K. E. Lowry, M. M. Mills, M. A. Palmer, W. M. Balch, N. R. Bates, C. R. Benitez-Nelson, E. Brownlee, K. E. Frey, S. R. Laney, J. Mathis, A. Matsuoka, B. Greg Mitchell, G. W. K. Moore, R. A. Reynolds, H. M. Sosik & J. H. Swift, 2014. Phytoplankton blooms beneath the sea ice in the Chukchi sea. Deep Sea Research Part II: Topical Studies in Oceanography 105:1-16 doi: https://doi.org/10.1016/j.dsr2.2014.03.018 .
Arrigo, K. R. & G. L. van Dijken, 2015. Continued increases in Arctic Ocean primary production. Prog Oceanogr 136:60-70 doi:10.1016/j.pocean.2015.05.002.
Assmy, P., M. Fernandez-Mendez, P. Duarte, A. Meyer, A. Randelhoff, C. J. Mundy, L. M. Olsen, H. M. Kauko, A. Bailey, M. Chierici, L. Cohen, A. P. Doulgeris, J. K. Ehn, A. Fransson, S. Gerland, H. Hop, S. R. Hudson, N. Hughes, P. Itkin, G. Johnsen, J. A. King, B. P. Koch, Z. Koenig, S. Kwasniewski, S. R. Laney, M. Nicolaus, A. K. Pavlov, C. M. Polashenski, C. Provost, A. Rosel, M. Sandbu, G. Spreen, L. H. Smedsrud, A. Sundfjord, T. Taskjelle, A. Tatarek, J. Wiktor, P. M. Wagner, A. Wold, H. Steen & M. A. Granskog, 2017. Leads in Arctic pack ice enable early phytoplankton blooms below snow-covered sea ice. Scientific Reports 7 doi:10.1038/srep40850.
Bachiller, E., G. Skaret, L. Nøttestad & A. Slotte, 2016. Feeding Ecology of Northeast Atlantic Mackerel, Norwegian Spring-Spawning Herring and Blue Whiting in the Norwegian Sea. PLoS One 11(2):e0149238 doi:10.1371/journal.pone.0149238.
Balazy, K., E. Trudnowska, M. Wichorowski & K. Błachowiak-Samołyk, 2018. Large versus small zooplankton in relation to temperature in the Arctic shelf region. Polar Research 37(1):1427409 doi:10.1080/17518369.2018.1427409.
Barber, D. G., W. N. Meier, S. Gerland, C. J. Mundy, M. Holland, S. Kern, Z. Li, C. Michel, D. K. Perovich, T. Tamura, J. Berge, J. Bowman, J. S. Christiansen, J. K. Ehn, S. Ferguson, M. A. Granskog, T. Kikuchi, H. Kuosa, B. Light, N. Lundholm, I. A. Melnikov, C. Polashenski, L. H. Smedsrud, G. Spreen, M. Tschudi, T. Vihma, M. Webster & L. Zhang, 2017. Arctic sea ice. Chapter 5 of Snow, Water, Ice and Permafrost in the Arctic (SWIPA). AMAP, Oslo. 103-136.
Barrett, R. T., K. E. Erikstad & T. K. Reiertsen, 2017. Ocean climate and egg investment in the black-legged kittiwake Rissa tridactyla Marine Ecology Progress Series 579:129-137.
Basedow, S. L., A. Sundfjord, W.-J. von Appen, E. Halvorsen, S. Kwasniewski & M. Reigstad, 2018. Seasonal Variation in Transport of Zooplankton Into the Arctic Basin Through the Atlantic Gateway, Fram Strait. Frontiers in Marine Science 5(194) doi:10.3389/fmars.2018.00194.
Belikov, S., I. Oganin & Å. Høines, 2011. Fish. Blue whiting. . In Jakobsen, T. & V. K. Ozhigin (eds) The Barents Sea - Ecosystem, Resources and Management Half a century of Russian-Norwegian cooperation. Tapir Academic Press, Trondheim, 355-363.
Bentsen, M., I. Bethke, J. B. Debernard, T. Iversen, A. Kirkevåg, Ø. Seland, H. Drange, C. Roelandt, I. A. Seierstad, C. Hoose & J. E. Kristjánsson, 2013. The Norwegian Earth System Model, NorESM1-M – Part 1: Description and basic evaluation of the physical climate. Geosci Model Dev 6(3):687-720 doi:10.5194/gmd-6-687-2013.
Bjørge, A., 2017. Rapport fra forskerutvalg om sjøpattedyr, 2017. Havforskningsinstituttet.
Bjørge, A., 2018. Forskerutvalg om sjøpattedyr 2018. Rapport Havforskningsinstituttet. . 56.
Bjørge, A., 2020. Rapport fra forskerutvalg om sjøpattedyr, 2020. Havforskningsinstituttet.
Blanchet, M. A., C. Lydersen, R. A. Ims & K. M. Kovacs, 2015. Seasonal, Oceanographic and Atmospheric Drivers of Diving Behaviour in a Temperate Seal Species Living in the High Arctic. PLoS One 10(7) doi:10.1371/journal.pone.0132686.
Blanchet, M. A., C. Lydersen, R. A. Ims & K. M. Kovacs, 2016. Making it through the first year: Ontogeny of movement and diving behavior in harbor seals from Svalbard, Norway. Marine Mammal Science 32(4):1340-1369 doi:10.1111/mms.12341.
Blanchet, M. A., C. Lydersen, R. A. Ims, A. D. Lowther & K. M. Kovacs, 2014. Harbour seal Phoca vitulina movement patterns in the high-Arctic archipelago of Svalbard, Norway. Aquatic Biology 21(3):167-181 doi:10.3354/ab00580.
Blévin, P., J. Aars, M. Andersen, M.-A. Blanchet, L. Hanssen, D. Herzke, R. M. Jeffreys, E. S. Nordøy, M. Pinzone, C. de la Vega & H. Routti, 2020. Pelagic vs Coastal—Key Drivers of Pollutant Levels in Barents Sea Polar Bears with Contrasted Space-Use Strategies. Environmental Science & Technology 54(2):985-995 doi:10.1021/acs.est.9b04626.
Bogstad, B., H. Gjøsæter, T. Haug & U. Lindstrøm, 2015. A review of the battle for food in the Barents Sea: Cod vs. marine mammals. Frontiers in Ecology and Evolution 3 doi:10.3389/fevo.2015.00029.
Bohlin-Nizzetto, P., W. Aas & V. Nikiforov, 2019. Monitoring of environmental contaminants in air and precipitation. Annual report 2018. NILU report 11/2019, 124 pp.
Boitsov, V., A. Karsakov & A. Trofimov, 2012. Atlantic water temperature and climate in the Barents Sea, 2000-2009. ICES Journal of Marine Science 69:833-840 doi:10.1093/icesjms/fss075.
Buhl-Mortensen, P., M. F. J. Dolan, R. E. Ross, G. Gonzalez-Mirelis, L. Buhl-Mortensen, L. R. Bjarnadóttir & J. Albretsen, 2020. Classification and Mapping of Benthic Biotopes in Arctic and Sub-Arctic Norwegian Waters. Frontiers in Marine Science 7(271) doi:10.3389/fmars.2020.00271.
Bustnes, J. O., S. Bourgeon, E. H. K. Leat, E. Magnusdottir, H. Strom, S. A. Hanssen, A. Petersen, K. Olafsdottir, K. Borga, G. W. Gabrielsen & R. W. Furness, 2015. Multiple Stressors in a Top Predator Seabird: Potential Ecological Consequences of Environmental Contaminants, Population Health and Breeding Conditions. PLoS One 10(7) doi:10.1371/journal.pone.0131769.
Cavalieri, D. J., C. L. Parkinson, P. Gloersen & H. J. Zwally, 1996, updated yearly. Sea Ice Concentrations from Nimbus-7 SMMR and DMSP SSM/I-SSMIS Passive Microwave Data, Version 1. Monthly Mean Gridded Ice Concentration Fields . Boulder, Colorado USA. NASA National Snow and Ice Data Center Distributed Active Archive Center. doi: https://doi.org/10.5067/8GQ8LZQVL0VL .
Chierici, M. & A. Fransson, 2018. Arctic chemical oceanography at the edge: focus on carbonate chemistry. In Wassmann, P. (ed) At the edge. 343.
Chierici, M., I. Skjelvan, M. Norli, E. Jones, K. Y. Børsheim, S. K. Lauvset, H. H. Lødemel, K. Sørensen, A. L. King & T. Johannessen, 2017. Overvåking av havforsuring i norske farvann i 2016, Rapport, Miljødirektoratet, M-776|2017.
Christensen-Dalsgaard, S., R. May & S.-H. Lorentsen, 2018. Taking a trip to the shelf: Behavioral decisions are mediated by the proximity to foraging habitats in the black-legged kittiwake. Ecol Evol 8(2):866-878 doi:10.1002/ece3.3700.
Cury, P. M., I. L. Boyd, S. Bonhommeau, T. Anker-Nilssen, R. J. M. Crawford, R. W. Furness, J. A. Mills, E. J. Murphy, H. Österblom, M. Paleczny, J. F. Piatt, J.-P. Roux, L. Shannon & W. J. Sydeman, 2011. Global Seabird Response to Forage Fish Depletion—One-Third for the Birds. Science 334(6063):1703-1706 doi:10.1126/science.1212928.
Dahne, M., A. Gilles, K. Lucke, V. Peschko, S. Adler, K. Krugel, J. Sundermeyer & U. Siebert, 2013. Effects of pile-driving on harbour porpoises (Phocoena phocoena) at the first offshore wind farm in Germany. Environmental Research Letters 8(2) doi:10.1088/1748-9326/8/2/025002.
Dalpadado, P., K. R. Arrigo, S. S. Hjollo, F. Rey, R. B. Ingvaldsen, E. Sperfeld, G. L. van Dijken, L. C. Stige, A. Olsen & G. Ottersen, 2014. Productivity in the Barents Sea - Response to Recent Climate Variability. PLoS One 9(5) doi:10.1371/journal.pone.0095273.
Dalpadado, P., K. R. Arrigo, G. L. van Dijken, H. R. Skjoldal, E. Bagøien, A. Dolgov, I. Prokopchuk & E. Sperfeld, in press. Climate effects on temporal and spatial dynamics of phytoplankton and zooplankton in the Barents Sea. . Prog Oceanogr.
Dalpadado, P., H. Hop, J. Ronning, V. Pavlov, E. Sperfeld, F. Buchholz, A. Rey & A. Wold, 2016. Distribution and abundance of euphausiids and pelagic amphipods in Kongsfjorden, Isfjorden and Rijpfjorden (Svalbard) and changes in their relative importance as key prey in a warming marine ecosystem. Polar Biology 39(10):1765-1784 doi:10.1007/s00300-015-1874-x.
Dalpadado, P., R. B. Ingvaldsen, L. C. Stige, B. Bogstad, T. Knutsen, G. Ottersen & B. Ellertsen, 2012. Climate effects on Barents Sea ecosystem dynamics. Ices Journal of Marine Science 69(7):1303-1316 doi:10.1093/icesjms/fss063.
De Boer, M., N. Janinhoff, G. Nijs & H. Verdaat, 2019. Encouraging encounters: unusual aggregations of bowhead whales Balaena mysticetus in the western Fram Strait. Endangered Species Research 39:51-62 doi:10.3354/esr00948.
Denisenko, S. G., 2013. Biodiversity and bioresources of macrozoobenthos in the Barents Sea, structure and long-term changes. Nauka, St Petersburg. Pp 284.
Derocher, A. E., 2005. Population ecology of polar bears at Svalbard, Norway. Popul Ecol 47(3):267-275 doi:10.1007/s10144-005-0231-2.
Derocher, A. E., M. Andersen, O. Wiig, J. Aars, E. Hansen & M. Biuw, 2011. Sea ice and polar bear den ecology at Hopen Island, Svalbard. Marine Ecology Progress Series 441:273-279 doi:10.3354/meps09406.
Descamps, S., T. Anker-Nilssen, R. T. Barrett, D. B. Irons, F. Merkel, G. J. Robertson, N. G. Yoccoz, M. L. Mallory, W. A. Montevecchi, D. Boertmann, Y. Artukhin, S. Christensen-Dalsgaard, K. E. Erikstad, H. G. Gilchrist, A. L. Labansen, S. H. Lorentsen, A. Mosbech, B. Olsen, A. Petersen, J. F. Rail, H. M. Renner, H. Strom, G. H. Systad, S. I. Wilhelm & L. Zelenskaya, 2017. Circumpolar dynamics of a marine top-predator track ocean warming rates. Global Change Biology 23(9):3770-3780 doi:10.1111/gcb.13715.
Descamps, S., H. Strom & H. Steen, 2013. Decline of an arctic top predator: synchrony in colony size fluctuations, risk of extinction and the subpolar gyre. Oecologia 173(4):1271-1282 doi:10.1007/s00442-013-2701-0.
Dolgov, A., E. Orlova, P. Renaud, v. Boitsov, I. Prokopchuk & M. Zashihina, 2013. Structure of the macroplankton–pelagic fish–cod trophic complex in a warmer Barents Sea. Marine Biology Research 9:851-866 doi:10.1080/17451000.2013.775453.
Dupont, N., E. Bagoien & W. Melle, 2017. Inter-annual variability in spring abundance of adult Calanus finmarchicus from the overwintering population in the southeastern Norwegian Sea. Prog Oceanogr 152:75-85 doi:10.1016/j.pocean.2017.02.004.
Eckhardt, S., K. Breivik, S. Manø & A. Stohl, 2007. Record high peaks in PCB concentrations in the Arctic atmosphere due to long-range transport of biomass burning emissions. Atmos Chem Phys 7(17):4527-4536 doi:10.5194/acp-7-4527-2007.
Ellingsen, K. E., N. G. Yoccoz, T. Tveraa, K. T. Frank, E. Johannesen, M. J. Anderson, A. V. Dolgov & N. L. Shackell, 2020. The rise of a marine generalist predator and the fall of beta diversity. Global Change Biology 26(5):2897-2907 doi:10.1111/gcb.15027.
Engelsen, O., E. N. Hegseth, H. Hop, E. Hansen & S. Falk-Petersen, 2002. Spatial variability of chlorophyll-a in the Marginal Ice Zone of the Barents Sea, with relations to sea ice and oceanographic conditions. Journal of Marine Systems 35(1):79-97 doi: https://doi.org/10.1016/S0924-7963(02)00077-5 .
Engelsen, O., H. Hop, E. Hegseth, E. Hansen & S. Falk-Petersen, 2004. Deriving phytoplankton biomass in the Marginal Ice Zone from satellite observable parameters. International Journal of Remote Sensing 25:1453-1457 doi:10.1080/01431160310001592436.
Eriksen, E., M. Huserbråten, H. Gjøsæter, F. Vikebø & J. Albretsen, 2019. Polar cod egg and larval drift patterns in the Svalbard archipelago. Polar Biology doi:10.1007/s00300-019-02549-6.
Eriksen, E., R. B. Ingvaldsen, K. Nedreaas & D. Prozorkevich, 2015. The effect of recent warming on polar cod and beaked redfish juveniles in the Barents Sea. Regional Studies in Marine Science 2:105-112 doi: http://doi.org/10.1016/j.rsma.2015.09.001 .
Eriksen, E., H. R. Skjoldal, H. Gjosaeter & R. Primicerio, 2017. Spatial and temporal changes in the Barents Sea pelagic compartment during the recent warming. Prog Oceanogr 151:206-226 doi:10.1016/j.pocean.2016.12.009.
Erikstad, K. E., T. K. Reiertsen, T. Anker-Nilssen, R. T. Barrett, S.-H. Lorentsen, H. Strøm & G. H. Systad, 2007. Levedyktighetsanalyser for norske lomvibestander. NINA Rappport 240. 25.
Erikstad, K. E., T. Moum, J. O. Bustnes & T. K. Reiertsen, 2011. High levels of organochlorines may affect hatching sex ratio and hatchling body mass in arctic glaucous gulls. Functional Ecology 25(1):289-296 doi:10.1111/j.1365- 2435.2010.01771.x.
Falk-Petersen, S., P. Mayzaud, G. Kattner & J. R. Sargent, 2009. Lipids and life strategy of Arctic Calanus. Marine Biology Research 5(1):18-39 doi:10.1080/17451000802512267.
Fall, J. & M. Skern-Mauritzen, 2014. White-beaked dolphin distribution and association with prey in the Barents Sea. Marine Biology Research 10(10):957-971 doi:10.1080/17451000.2013.872796.
Fauchald, P., R. T. Barrett, J. O. Bustnes, K. E. Erikstad, L. Nøttestad, M. Skern-Mauritzen & F. B. Vikebø, 2015. Sjøfugl og marine økosystemer. Status for sjøfugl og sjøfuglenes næringsgrunnlag i Norge og på Svalbard. NINA Rapport 1161. 44.
Fernández-Méndez, M., L. M. Olsen, H. M. Kauko, A. Meyer, A. Rösel, I. Merkouriadi, C. J. Mundy, J. K. Ehn, A. M. Johansson, P. M. Wagner, Å. Ervik, B. K. Sorrell, P. Duarte, A. Wold, H. Hop & P. Assmy, 2018. Algal Hot Spots in a Changing Arctic Ocean: Sea-Ice Ridges and the Snow-Ice Interface. Frontiers in Marine Science 5:75.
Fluhr, J., H. Strom, R. Pradel, O. Duriez, G. Beaugrand & S. Descamps, 2017. Weakening of the subpolar gyre as a key driver of North Atlantic seabird demography: a case study with Brunnich's guillemots in Svalbard. Marine Ecology Progress Series 563:1-11 doi:10.3354/meps11982.
Fossheim, M., R. Primicerio, E. Johannesen, R. B. Ingvaldsen, M. M. Aschan & A. V. Dolgov, 2015. Recent warming leads to a rapid borealization of fish communities in the Arctic. Nature Clim Change 5(7):673-677 doi:10.1038/nclimate2647
Fransson, A., M. Chierici, H. Hop, H. S. Findlay, S. Kristiansen & A. Wold, 2016. Late winter-to-summer change in ocean acidification state in Kongsfjorden, with implications for calcifying organisms. Polar Biology 39(10):1841-1857 doi:10.1007/s00300-016-1955-5.
Frantzen, S. & A. Måge, 2016. Fremmedstoffer i villfisk med vekt på kystnære farvann. Brosme, lange og bifangstarter. Gjelder tall for prøver samlet inn i 2013-2015. Bergen, NIFES: 116.
Frantzen, S., B. M. Nilsen & M. Sanden, in press. Fremmedstoffer i rødspette, breiflabb og lyr. Sluttrapport for kartleggingsprogrammet «Fremmedstoffer i villfisk med vekt på kystnære farvann» 2016-2018. Rapport fra Havforskningen 2020-11. xx s.
Frederiksen, M., B. Moe, F. Daunt, R. A. Phillips, R. T. Barrett, M. I. Bogdanova, T. Boulinier, J. W. Chardine, O. Chastel, L. S. Chivers, S. Christensen-Dalsgaard, C. Clément-Chastel, K. Colhoun, R. Freeman, A. J. Gaston, J. González-Solís, A. Goutte, D. Grémillet, T. Guilford, G. Høj Jensen, Y. Krasnov, S.-H. Lorentsen, M. L. Mallory, M. Newell, B. Olsen, D. Shaw, H. Steen, H. Strøm, G. H. Systad, T. L. Thórarinsson & T. Anker-Nilssen, 2012. Multi-colony tracking reveals the winter distribution of a pelagic seabird on an ocean basin scale. Diversity and Distributions 18:530-542.
Gerland, S., A. H. H. Renner, F. Godtliebsen, D. Divine & T. B. Loyning, 2008. Decrease of sea ice thickness at Hopen, Barents Sea, during 1966-2007. Geophys Res Lett 35(6) doi:10.1029/2007gl032716.
Gjøsæter, H., B. Bogstad & S. Tjelmeland, 2009. Ecosystem effects of the three capelin stock collapses in the Barents Sea. Marine Biology Research 5(1):40-53 doi:10.1080/17451000802454866.
Gjøsæter, H., E. H. Hallfredsson, N. Mikkelsen, B. Bogstad & T. Pedersen, 2015. Predation on early life stages is decisive for year-class strength in the Barents Sea capelin (Mallotus villosus) stock. ICES Journal of Marine Science: Journal du Conseil doi:10.1093/icesjms/fsv177.
Gjøsæter, H., E. H. Hallfredsson, N. Mikkelsen, B. Bogstad & T. Pedersen, 2016. Predation on early life stages is decisive for year-class strength in the Barents Sea capelin (Mallotus villosus) stock. Ices Journal of Marine Science 73(2):182-195 doi:10.1093/icesjms/fsv177.
Granskog, M. A., I. Fer, A. Rinke & H. Steen, 2018. Atmosphere-Ice-Ocean-Ecosystem Processes in a Thinner Arctic Sea Ice Regime: The Norwegian Young Sea ICE (N-ICE2015) Expedition. Journal of Geophysical Research: Oceans 123(3):1586-1594 doi:10.1002/2017jc013328.
Green, N., M. Schøyen, S. Øxnevad, A. Ruus, D. Hjermann, G. Severinsen, T. Høgåsen, B. A. Beylich, J. Håvardstun, E. Lund, L. Tveiten & K. Bæk, 2017. Contaminants in coastal waters of Norway 2016. Miljøgifter i norske kystområder 2016., NIVA. Miljødirektoratet rapport M-856.
Grémillet, D., A. Ponchon, M. Paleczny, M.-L. D. Palomares, V. Karpouzi & D. Pauly, 2018. Persisting Worldwide Seabird-Fishery Competition Despite Seabird Community Decline. Current Biology 28(24):4009-4013.e2 doi: https://doi.org/10.1016/j.cub.2018.10.051 .
Guéry, L., S. Descamps, K. Hodges, R. Pradel, B. Moe, S. Hanssen, K. Erikstad, G. Gabrielsen, H. Gilchrist & S. Jenouvrier, in press. Winter extratropical cyclone influence on seabird survival: variation between and within common eider populations. . Mar Ecol Prog Ser.
Hamilton, C. D., K. M. Kovacs, R. A. Ims, J. Aars & C. Lydersen, 2017. An Arctic predator-prey system in flux: climate change impacts on coastal space use by polar bears and ringed seals. The Journal of animal ecology 86(5):1054-1064 doi:10.1111/1365-2656.12685.
Hamilton, C. D., K. M. Kovacs & C. Lydersen, 2018. Individual variability in diving, movement and activity patterns of adult bearded seals in Svalbard, Norway. Scientific Reports 8(1):16988 doi:10.1038/s41598-018-35306-6.
Hamilton, C. D., C. Lydersen, R. A. Ims & K. M. Kovacs, 2015. Predictions replaced by facts: a keystone species' behavioural responses to declining arctic sea-ice. Biology Letters 11(11) doi:10.1098/rsbl.2015.0803.
Hamilton, C. D., C. Lydersen, R. A. Ims & K. M. Kovacs, 2016. Coastal habitat use by ringed seals Pusa hispida following a regional sea-ice collapse: importance of glacial refugia in a changing Arctic. Marine Ecology Progress Series 545:261-277 doi:10.3354/meps11598.
Hamilton, C. D., J. Vacquié-Garcia, K. M. Kovacs, R. A. Ims, J. Kohler & C. Lydersen, 2019. Contrasting changes in space use induced by climate change in two Arctic marine mammal species. Biology Letters 15(3):20180834 doi:doi:10.1098/rsbl.2018.0834.
Hatun, H., K. Azetsu-Scott, R. Somavilla, F. Rey, C. Johnson, M. Mathis, U. Mikolajewicz, P. CoupeI, J. E. Tremblay, S. Hartman, S. V. Pacariz, I. Salter & J. Olafsson, 2017. The subpolar gyre regulates silicate concentrations in the North Atlantic. Scientific Reports 7 doi:10.1038/s41598-017-14837-4.
Hegseth, E. N. & A. Sundfjord, 2008. Intrusion and blooming of Atlantic phytoplankton species in the high Arctic. Journal of Marine Systems 74(1):108-119 doi: https://doi.org/10.1016/j.jmarsys.2007.11.011 .
Hipfner, J. M., L. K. Blight, R. W. Lowe, S. I. Wilhelm, G. J. Robertson, R. T. Barrett, T. Anker-Nilssen & T. P. Good, 2012. Unintended consequences: how the recovery of sea eagle Haliaeetus spp. populations in the northern hemisphere is affecting seabirds. Marine Ornithology 40:39-52.
Hirche, H. J., 1997. Life cycle of the copepod Calanus hyperboreus in the Greenland sea. Marine Biology 128(4):607-618 doi:10.1007/s002270050127.
Hjelset, A. M., M. Andersen, I. Gjertz, C. Lydersen & B. Gulliksen, 1999. Feeding habits of bearded seals (Erignathus barbatus) from the Svalbard area, Norway. Polar Biology 21(3):186-193 doi:10.1007/s003000050351.
Hjermann, D. O., G. Ottersen & N. C. Stenseth, 2004. Competition among fishermen and fish causes the collapse of Barents Sea capelin. Proc Natl Acad Sci U S A 101(32):11679-11684 doi:10.1073/pnas.0402904101.
Hop, H., P. Assmy, A. Wold, A. Sundfjord, M. Daase, P. Duarte, S. Kwasniewski, M. Gluchowska, J. M. Wiktor, A. Tatarek, J. Wiktor, S. Kristiansen, A. Fransson, M. Chierici & M. Vihtakari, 2019. Pelagic Ecosystem Characteristics Across the Atlantic Water Boundary Current From Rijpfjorden, Svalbard, to the Arctic Ocean During Summer (2010–2014). Frontiers in Marine Science 6(181) doi:10.3389/fmars.2019.00181.
Horvat, C., D. R. Jones, S. Iams, D. Schroeder, D. Flocco & D. Feltham, 2017. The frequency and extent of sub-ice phytoplankton blooms in the Arctic Ocean. Science Advances 3(3):e1601191 doi:10.1126/sciadv.1601191.
Hovinen, J. E. H., J. Welcker, S. Descamps, H. Strom, K. Jerstad, J. Berge & H. Steen, 2014. Climate warming decreases the survival of the little auk (Alle alle), a high Arctic avian predator. Ecol Evol 4(15):3127-3138 doi:10.1002/ece3.1160.
Hurrell, J. W., Y. Kushnir, G. Ottersen & M. Visbeck, 2003. An Overview of the North Atlantic Oscillation. In Hurrell, J. W., Y. Kushnir, G. Ottersen & M. Visbeck (eds) The North Atlantic Oscillation: Climatic Significance And Environmental Impact. American Geophysical Union, Washington D.C., 1-35.
Huserbråten, M. B. O., E. Eriksen, H. Gjøsæter & F. Vikebø, 2019. Polar cod in jeopardy under the retreating Arctic sea ice. Communications Biology 2(1):407 doi:10.1038/s42003-019-0649-2.
ICES, 2018. Interim Report of the Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). WGIBAR 2018 REPORT 9-12 March 2018. Tromsø, Norway. ICES CM 2018/IEASG:04. 210 pp. .
ICES, 2019a. Arctic Fisheries Working Group (AFWG). ICES Scientific Reports. 1:30. . 930.
ICES, 2019b. ICES/NAFO/NAMMCO Working Group on Harp and Hooded Seals (WGHARP). ICES Scientific Reports. 1:72. . 193.
ICES, 2019c. The Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). ICES Scientific Reports. 1:42. . 157.
ICES, 2020. Working Group on the Integrated Assessments of the Barents Sea ( WGIBAR). ICES Scientific Reports. 2:30. . 206.
Ingvaldsen, R. B., L. Asplin & H. Loeng, 2004. Velocity field of the western entrance to the Barents Sea. Journal of Geophysical Research: Oceans 109(C3) doi:10.1029/2003jc001811.
IPCC, 2014. IPCC, 2014: Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, (eds.)]. IPCC, Geneva, Switzerland, 151 pp. . In: Pachauri, R. K. & L. A. Meyer (eds).
IPCC, 2019 - in press. Summary for policy makers. In Pörtner, H.-O., et al. (eds) IPCC Special Report on the Ocean and Cryosphere in a Changing Climate.
Itkin, P., M. Karcher & R. Gerdes, 2014. Is weaker Arctic sea ice changing the Atlantic water circulation? Journal of Geophysical Research-Oceans 119(9):5992-6009 doi:10.1002/2013jc009633.
Jansen, T., S. Post, A. H. Olafsdottir, P. Reynisson, G. J. Óskarsson & K. E. Arendt, 2019. Diel vertical feeding behaviour of Atlantic mackerel (Scomber scombrus) in the Irminger current. Fisheries Research 214:25-34 doi: https://doi.org/10.1016/j.fishres.2019.01.020 .
Jepsen, J. U., P. Arneberg, R. A. Ims, A. Siwertsson & N. G. Yoccoz, 2019. Test av fagsystemet for økologisk tilstand. Erfaringer fra pilotprosjekter for arktisk tundra og arktisk del av Barentshavet. NINA Rapport 1674. Norsk institutt for naturforskning.
Johannesen, E., R. B. Ingvaldsen, B. Bogstad, P. Dalpadado, E. Eriksen, H. Gjøsæter, T. Knutsen, M. Skern-Mauritzen & J. E. Stiansen, 2012. Changes in Barents Sea ecosystem state, 1970–2009: climate fluctuations, human impact, and trophic interactions. ICES Journal of Marine Science 69(5):880-889 doi:10.1093/icesjms/fss046.
Johnsen, G., M. Norli, M. Moline, I. Robbins, C. von Quillfeldt, K. Sørensen, F. Cottier & J. Berge, 2018. The advective origin of an under-ice spring bloom in the Arctic Ocean using multiple observational platforms. Polar Biology 41(6):1197-1216 doi:10.1007/s00300-018-2278-5.
Joint, I. & S. B. Groom, 2000. Estimation of phytoplankton production from space: current status and future potential of satellite remote sensing. J Exp Mar Biol Ecol 250(1-2):233-255 doi:10.1016/s0022-0981(00)00199-4.
Jones, E., M. Chierici, I. Skjelvan, M. Norli, K. Y. Børsheim, H. H. Lødemel, T. Kutti, K. Sørensen, A. L. King, S. Lauvset, K. Jackson, T. de Lange, T. Johannessen & C. Mourgue, 2019. Monitoring ocean acidification in Norwegian seas in 2018, Rapport, Miljødirektoratet, M-1417|2019.
Julshamn, K., S. Valdersnes, A. Duinker, K. Nedreaas, J. H. Sundet & A. Maage, 2015. Heavy metals and POPs in red king crab from the Barents Sea. Food Chem 167:409-17 doi:10.1016/j.foodchem.2014.07.003.
Karcher, M. J., S. Gerland, I. H. Harms, M. Iosjpe, H. E. Heldal, P. J. Kershaw & M. Sickel, 2004. The dispersion of 99Tc in the Nordic Seas and the Arctic Ocean: a comparison of model results and observations. Journal of Environmental Radioactivity 74(1):185-198 doi: https://doi.org/10.1016/j.jenvrad.2004.01.026 .
King, J., G. Spreen, S. Gerland, C. Haas, S. Hendricks, L. Kaleschke & C. Wang, 2017. Sea-ice thickness from field measurements in the northwestern Barents Sea. Journal of Geophysical Research: Oceans 122(2):1497-1512 doi:10.1002/2016JC012199.
Komperød, M., A. L. Rudjord, L. Skuterud & J. E. Dyve, 2015. Stråledoser fra miljøet. Beregninger av befolkningens eksponering for stråling fra omgivelsene i Norge. StrålevernRapport 2015:11. Østerås: Statens strålevern.
Kovacs, K. M., J. Aars & C. Lydersen, 2014. Walruses recovering after 60+ years of protection in Svalbard, Norway. Polar Research 33(1):26034 doi:10.3402/polar.v33.26034.
Kovacs, K. M., B. A. Krafft & C. Lydersen, 2020. Bearded seal (Erignathus barbatus) birth mass and pup growth in periods with contrasting ice conditions in Svalbard, Norway. Marine Mammal Science 36(1):276-284 doi:10.1111/mms.12647.
Kovacs, K. M., C. Lydersen, J. Vacquiè-Garcia, O. Shpak, D. Glazov & M. P. Heide-Jørgensen, innsendt. The endangered Spitsbergen bowhead whales’ secrets revealed after hundreds of years in hiding. Biology Letters.
Krafft, B. A., K. M. Kovacs, M. Andersen, J. Aars, C. Lydersen, T. Ergon & T. Haug, 2006. ABUNDANCE OF RINGED SEALS (PUSA HISPIDA) IN THE FJORDS OF SPITSBERGEN, SVALBARD, DURING THE PEAK MOLTING PERIOD. Marine Mammal Science 22(2):394-412 doi:10.1111/j.1748-7692.2006.00035.x.
Krause, J. W., I. K. Schulz, K. A. Rowe, W. Dobbins, M. H. S. Winding, M. K. Sejr, C. M. Duarte & S. Agustí, 2019. Silicic acid limitation drives bloom termination and potential carbon sequestration in an Arctic bloom. Scientific Reports 9(1):8149 doi:10.1038/s41598-019-44587-4.
Lien, V. & B. Ådlandsvik, 2011. Bottom water formation as a primer for spring-blooms on Spitsbergenbanken? J Mar Syst 130 doi:10.1016/j.jmarsys.2011.11.018.
Lind, S., R. Ingvaldsen & T. Furevik, 2016. Arctic layer salinity controls heat loss from deep Atlantic layer in seasonally ice-covered areas of the Barents Sea. Geophys Res Lett 43 doi:10.1002/2016GL068421.
Lind, S., R. B. Ingvaldsen & T. Furevik, 2018. Arctic warming hotspot in the northern Barents Sea linked to declining sea-ice import. Nature Climate Change doi:10.1038/s41558-018-0205-y.
Loeng, H., 1991. Features of the physical oceanographic conditions of the Barents Sea. Polar Research 10(1):5-18 doi: https://doi.org/10.3402/polar.v10i1.6723 .
Lone, K., B. Merkel, C. Lydersen, K. M. Kovacs & J. Aars, 2017. Sea ice resource selection models for polar bears in the Barents Sea subpopulation. Ecography:n/a-n/a doi:10.1111/ecog.03020.
Lydersen, C., J. Aars & K. M. Kovacs, 2008. Estimating the number of walruses in Svalbard from aerial surveys and behavioural data from satellite telemetry. Arctic 61(2):119-128.
Lydersen, C., A. Martin, K. Kovacs & I. Gjertz, 2001. Summer and autumn movements of white whales Delphinapterus leucas in Svalbard, Norway. Marine Ecology-progress Series - MAR ECOL-PROGR SER 219:265-274 doi:10.3354/meps219265.
Lydersen, C., J. Vaquie-Garcia, E. Lydersen, G. N. Christensen & K. M. Kovacs, 2017. Novel terrestrial haul-out behaviour by ringed seals (Pusa hispida) in Svalbard, in association with harbour seals (Phoca vitulina). Polar Research 36(1):1374124 doi:10.1080/17518369.2017.1374124.
Lyubina, O. S., E. A. Frolova & D. R. Dikaeva, 2012. Current zoobenthos monitoring at the Kola Transect in the Barents Sea. Berichte zur Polarforschung 640:177-189.
Lyubina, O. S., N. A. Strelkova (Anisimova), P. A. Lubin, E. A. Frolova, D. R. Dikaeva, O. L. Zimina, O. Y. Akhmetchina, I. E. Manushin, I. O. Nekhaev, A. A. Frolov, D. V. Zakharov, E. A. Garbul & V. S. Vyaznikova, 2016. Modern quantitative distribution of zoobenthos along the transect Kola Section (In Rusian). Transactions of the Kola Science Centre Series 3 Oceanology 36(2):64-91.
Meier, W. N., F. Fetterer, M. Savoie, S. Mallory, R. Duerr & J. Stroeve, 2017a. NOAA/NSIDC Climate Data Record of Passive Microwave Sea Ice Concentration, Version 3. Boulder, Colorado USA. National Snow and Ice Data Center, doi: https://doi.org/10.7265/N59P2ZTG .
Meier, W. N., F. Fetterer & A. K. Windnagel, 2017b. Near-Real-Time NOAA/NSIDC Climate Data Record of Passive Microwave Sea Ice Concentration, Version 1, Boulder, Colorado USA. National Snow and Ice Data Center, doi: https://doi.org/10.7265/N5FF3QJ6 .
Meredith, M., M. Sommerkorn, S. Cassotta, C. Derksen, A. Ekaykin, A. Hollowed, G. Kofnas, A. Mackintosh, J. Melbourne-Thomas, M. M. C. Muelbert, G. Ottersen, H. Pritchard & E. A. G. Schuur, 2019 - in press. Polar Regions. In Pörtner, H.-O., et al. (eds) IPCC Special Report on the Ocean and Cryosphere in a Changing Climate.
Merkel, B., C. Lydersen, N. G. Yoccoz & K. M. Kovacs, 2013. The World’s Northernmost Harbour Seal Population–How Many Are There? PLoS One 8(7):e67576 doi:10.1371/journal.pone.0067576.
Mundy, C. J., M. Gosselin, J. Ehn, Y. Gratton, A. Rossnagel, D. G. Barber, J. Martin, J.-É. Tremblay, M. Palmer, K. R. Arrigo, G. Darnis, L. Fortier, B. Else & T. Papakyriakou, 2009. Contribution of under-ice primary production to an ice-edge upwelling phytoplankton bloom in the Canadian Beaufort Sea. Geophys Res Lett 36(17) doi:10.1029/2009gl038837.
Mundy, C. J., M. Gosselin, Y. Gratton, K. Brown, V. Galindo, K. Campbell, M. Levasseur, D. Barber, T. Papakyriakou & S. Bélanger, 2014. Role of environmental factors on phytoplankton bloom initiation under landfast sea ice in Resolute Passage, Canada. Marine Ecology Progress Series 497:39-49 doi:10.2307/24893504.
Møller, B., K. Tazmini, J. Drefvelin & T. Gäfvert, 2018. Overvåking av radioaktivitet i omgivelsene 2017. StrålevernRapport 2018:11. Østerås: Statens strålevern.
NAMMCO & Havforskningsinstituttet, 2019. Report of the Status of Harbour Porpoise in the North Atlantic Workshop. Tromsø, Norway.
Nilsen, B. M., S. Frantzen & M. Sanden, 2020. Fremmedstoffer i snabeluer (Sebastes mentella) og vanlig uer (Sebastes norvegicus). Sluttrapport for kartleggingsprogrammet «Miljøgifter i fisk og fiskevarer» 2016-2018. Rapport fra Havforskningen 2020-11. 63 s.
Notz, D., J. Dörr, D. A. Bailey, E. Blockley, M. Bushuk, J. B. Debernard, E. Dekker, P. DeRepentigny, D. Docquier, N. S. Fučkar, J. C. Fyfe, A. Jahn, M. Holland, E. Hunke, D. Iovino, N. Khosravi, F. Massonnet, G. Madec, S. O'Farrell, A. Petty, A. Rana, L. Roach, E. Rosenblum, C. Rousset, T. Semmler, J. Stroeve, B. Tremblay, T. Toyoda, H. Tsujino & M. Vancoppenolle, 2020. Arctic Sea Ice in CMIP6. Geophys Res Lett n/a(n/a):e2019GL086749 doi:10.1029/2019gl086749.
Nybø, S. & M. Evju, 2017. Fagsystem for fastsetting av god økologisk tilstand. Forslag fra et ekspertråd. Ekspertrådet for økologisk tilstand, 247 s.
Olsen, L. M., P. Duarte, C. Peralta-Ferriz, H. M. Kauko, M. Johansson, I. Peeken, M. Rozanska-Pluta, A. Tatarek, J. Wiktor, M. Fernandez-Mendez, P. M. Wagner, A. K. Pavlov, H. Hop & P. Assmy, 2019. A red tide in the pack ice of the Arctic Ocean. Sci Rep 9(1):9536 doi:10.1038/s41598-019-45935-0.
Onarheim, I. H., T. Eldevik, L. H. Smedsrud & J. C. Stroeve, 2018. Seasonal and Regional Manifestation of Arctic Sea Ice Loss. Journal of Climate 31(12):4917-4932 doi:10.1175/jcli-d-17-0427.1.
Onarheim, I. H. & M. Årthun, 2017. Toward an ice-free Barents Sea. Geophys Res Lett 44(16):8387-8395 doi:10.1002/2017gl074304.
Orlova, E. L., A. V. Dolgov, P. E. Renaud, M. Greenacre, C. Halsband & V. A. Ivshin, 2015. Climatic and ecological drivers of euphausiid community structure vary spatially in the Barents Sea: relationships from a long time series (1952–2009). Frontiers in Marine Science 1(74) doi:10.3389/fmars.2014.00074.
Oziel, L., P. Massicotte, A. Randelhoff, J. Ferland, A. Vladoiu, L. Lacour, V. Galindo, S. Lambert-Girard, D. Dumont, Y. Cuypers, P. Bouruet-Aubertot, C. Mundy, J. Ehn, G. Becu, C. Marec, M.-H. Forget, N. Garcia, P. Coupel, P. Raimbault & M. Babin, 2019. Environmental factors influencing the seasonal dynamics of spring algal blooms in and beneath sea ice in western Baffin Bay. Elem Sci Anth 7:34 doi:10.1525/elementa.372.
Paulsen, M. L., H. Doré, L. Garczarek, L. Seuthe, O. Müller, R.-A. Sandaa, G. Bratbak & A. Larsen, 2016. Synechococcus in the Atlantic Gateway to the Arctic Ocean. Frontiers in Marine Science 3(191) doi:10.3389/fmars.2016.00191.
Pavlova, O., V. Pavlov & S. Gerland, 2014. The impact of winds and sea surface temperatures on the Barents Sea ice extent, a statistical approach. Journal of Marine Systems 130:248-255 doi: https://doi.org/10.1016/j.jmarsys.2013.02.011 .
Peacock, E., S. A. Sonsthagen, M. E. Obbard, A. Boltunov, E. V. Regehr, N. Ovsyanikov, J. Aars, S. N. Atkinson, G. K. Sage, A. G. Hope, E. Zeyl, L. Bachmann, D. Ehrich, K. T. Scribner, S. C. Amstrup, S. Belikov, E. W. Born, A. E. Derocher, I. Stirling, M. K. Taylor, Ø. Wiig, D. Paetkau & S. L. Talbot, 2015. Implications of the Circumpolar Genetic Structure of Polar Bears for Their Conservation in a Rapidly Warming Arctic. PLoS One 10(1):e112021 doi:10.1371/journal.pone.0112021.
Polyakov, I. V., A. V. Pnyushkov, M. B. Alkire, I. M. Ashik, T. M. Baumann, E. C. Carmack, I. Goszczko, J. Guthrie, V. V. Ivanov, T. Kanzow, R. Krishfield, R. Kwok, A. Sundfjord, J. Morison, R. Rember & A. Yulin, 2017. Greater role for Atlantic inflows on sea-ice loss in the Eurasian Basin of the Arctic Ocean. Science 356(6335):285-291 doi:10.1126/science.aai8204.
Prop, J., J. Aars, B.-J. Bårdsen, S. A. Hanssen, C. Bech, S. Bourgeon, J. de Fouw, G. Gabrielsen, J. Lang, E. Noreen, T. Oudman, B. Sittler, L. Stempniewicz, I. Tombre, E. Wolters & B. Moe, 2015. Climate change and the increasing impact of polar bears on bird populations. Frontiers in Ecology and Evolution 3:1-12 doi:10.3389/fevo.2015.00033.
Quintela, M., F. Besnier, B. Seliussen, K. A. Glover & U. Lindstrøm, 2020. Population structure of bycaught harbour porpoise (Phocoena phocoena) in Norway. Marine Biology Research:1-7 doi:10.1080/17451000.2020.1729992.
Reiertsen, T. K., K. E. Erikstad, T. Anker-Nilssen, R. T. Barrett, T. Boulinier, M. Frederiksen, J. Gonzalez-Solis, D. Gremillet, D. Johns, B. Moe, A. Ponchon, M. Skern-Mauritzen, H. Sandvik & N. G. Yoccoz, 2014. Prey density in non-breeding areas affects adult survival of black-legged kittiwakes Rissa tridactyla. Marine Ecology Progress Series 509:289-+ doi:10.3354/meps10825.
Reiertsen, T. K., K. E. Erikstad, R. T. Barrett, H. Sandvik & N. G. Yoccoz, 2012. Climate fluctuations and differential survival of bridled and non-bridled Common Guillemots Uria aalge. Ecosphere 3(6) doi:10.1890/es12-00031r.
Reigstad, M., T. Eldevik & S. Gerland, 2019. The Nansen Legacy. Fram forum 2019, 100-105. Fram Centre, Tromsø, Norway. ISSN 8193-5540 (online version).
Renaud, P. E., M. Daase, N. S. Banas, T. M. Gabrielsen, J. E. Søreide, Ø. Varpe, F. Cottier, S. Falk-Petersen, C. Halsband, D. Vogedes, K. Heggland & J. Berge, 2018. Pelagic food-webs in a changing Arctic: a trait-based perspective suggests a mode of resilience. ICES Journal of Marine Science 75(6):1871-1881 doi:10.1093/icesjms/fsy063.
Renaut, S., E. Devred & M. Babin, 2018. Northward Expansion and Intensification of Phytoplankton Growth During the Early Ice-Free Season in Arctic. Geophys Res Lett doi:doi:10.1029/2018GL078995.
Renner, A. H. H., S. Hendricks, S. Gerland, J. Beckers, C. Haas & T. Krumpen, 2013. Large-scale ice thickness distribution of first-year sea ice in spring and summer north of Svalbard. Annals of Glaciology 54(62):13-18 doi:10.3189/2013AoG62A146.
Ressler, P. H., P. Dalpadado, G. J. Macaulay, N. Handegard & M. Skern-Mauritzen, 2015. Acoustic surveys of euphausiids and models of baleen whale distribution in the Barents Sea. Marine Ecology Progress Series 527:13-29 doi:10.3354/meps11257.
Rey, F., 2012. Declining silicate concentrations in the Norwegian and Barents Seas. ICES Journal of Marine Science 69(2):208-212 doi:10.1093/icesjms/fss007.
Rosel, A., P. Itkin, J. King, D. Divine, C. X. Wang, M. A. Granskog, T. Krumpen & S. Gerland, 2018. Thin Sea Ice, Thick Snow, and Widespread Negative Freeboard Observed During N-ICE2015 North of Svalbard. Journal of Geophysical Research-Oceans 123(2):1156-1176 doi:10.1002/2017jc012865.
Routti, H., T. C. Atwood, T. Bechshoft, A. Boltunov, T. M. Ciesielski, J.-P. Desforges, R. Dietz, G. W. Gabrielsen, B. M. Jenssen, R. J. Letcher, M. A. McKinney, A. D. Morris, F. F. Rigét, C. Sonne, B. Styrishave & S. Tartu, 2019. State of knowledge on current exposure, fate and potential health effects of contaminants in polar bears from the circumpolar Arctic. Science of The Total Environment 664:1063-1083 doi: https://doi.org/10.1016/j.scitotenv.2019.02.030 .
Sandvik, H., R. T. Barrett, K. E. Erikstad, M. S. Myksvoll, F. Vikebo, N. G. Yoccoz, T. Anker-Nilssen, S. H. Lorentsen, T. K. Reiertsen, J. Skardhamar, M. Skern-Mauritzen & G. H. Systad, 2016. Modelled drift patterns of fish larvae link coastal morphology to seabird colony distribution. Nat Commun 7 doi:10.1038/ncomms11599.
Sandvik, H., K. E. Erikstad, R. T. Barrett & N. G. Yoccoz, 2005. The effect of climate on adult survival in five species of North Atlantic seabirds. J Anim Ecol 74(5):817-831 doi:10.1111/j.1365-2656.2005.00981.x.
Sandvik, H., K. E. Erikstad & B. E. Saether, 2012. Climate affects seabird population dynamics both via reproduction and adult survival. Marine Ecology-Progress Series 454:273-+ doi:10.3354/meps09558.
Sandvik, H., T. K. Reiertsen, K. E. Erikstad, T. Anker-Nilssen, R. T. Barrett, S. H. Lorentsen, G. H. Systad & M. S. Myksvoll, 2014. The decline of Norwegian kittiwake populations: modelling the role of ocean warming. Climate Research 60(2):91-102 doi:10.3354/cr01227.
Sandø, A., Y. Gao & H. Langehaug, 2014a. Poleward ocean heat transports, sea ice processes and Arctic sea ice variability in NorESM1-M simulations. Journal of Geophysical Research: Oceans 119 doi:10.1002/2013JC009435.
Sandø, A., A. Melsom & W. P. Budgell, 2014b. Downscaling IPCC control and future scenario with focus on the Barents Sea. Ocean Dynamics 64:927-949 doi:10.1007/s10236-014-0731-8.
Sarnocińska, J., J. Teilmann, J. D. Balle, F. M. van Beest, M. Delefosse & J. Tougaard, 2020. Harbor Porpoise (Phocoena phocoena) Reaction to a 3D Seismic Airgun Survey in the North Sea. Frontiers in Marine Science 6(824) doi:10.3389/fmars.2019.00824.
Schlesinger, M. E. & N. Ramankutty, 1994. An oscillation in the global climate system of period 65–70 years. Nature 367(6465):723-726 doi:10.1038/367723a0.
Screen, J. A. & I. Simmonds, 2010. Increasing fall-winter energy loss from the Arctic Ocean and its role in Arctic temperature amplification. Geophys Res Lett 37(16) doi:10.1029/2010gl044136.
Shchepetkin, A. F. & J. C. McWilliams, 2005. The regional oceanic modeling system (ROMS): a split-explicit, free-surface, topography-following-coordinate oceanic model. Ocean Modelling 9(4):347-404 doi: https://doi.org/10.1016/j.ocemod.2004.08.002 .
Sirenko, B. I., 2001. Introduction. In List of species of free-living invertebrates of Eurasian Arctic Seas and adjacent waters. pp. 3-8. Ed by Sirenko B.I. Explorations of the fauna of the seas, 51. Zoological Institute of the Rus.Ac. of Sci.: Saint Petersburg.
Skancke, J. & T. Nordam, 2016. Long-term fate of Ra-226 originating from offshore produced water discharges. : SINTEF 2016 (ISBN 978-82-14-06062-1) 20 s. OCEAN.
Skaret, G., P. Dalpadado, S. S. Hjollo, M. D. Skogen & E. Strand, 2014. Calanus finmarchicus abundance, production and population dynamics in the Barents Sea in a future climate. Prog Oceanogr 125:26-39 doi:10.1016/j.pocean.2014.04.008.
Skern-Mauritzen, M., E. Johannesen, A. Bjorge & N. Oien, 2011. Baleen whale distributions and prey associations in the Barents Sea. Marine Ecology Progress Series 426:289-301 doi:10.3354/meps09027.
Smedsrud, L. H., I. Esau, R. B. Ingvaldsen, T. Eldevik, P. M. Haugan, C. Li, V. S. Lien, A. Olsen, A. M. Omar, O. H. Otterå, B. Risebrobakken, A. B. Sandø, V. A. Semenov & S. A. Sorokina, 2013. THE ROLE OF THE BARENTS SEA IN THE ARCTIC CLIMATE SYSTEM. Reviews of Geophysics 51(3):415-449 doi:10.1002/rog.20017.
Soltwedel, T., E. Bauerfeind, M. Bergmann, A. Bracher, N. Budaeva, K. Busch, A. Cherkasheva, K. Fahl, K. Grzelak, C. Hasemann, M. Jacob, A. Kraft, C. Lalande, K. Metfies, E. M. Nothig, K. Meyer, N. V. Queric, I. Schewe, M. Wlodarska-Kowalczuk & M. Klages, 2016. Natural variability or anthropogenically-induced variation? Insights from 15 years of multidisciplinary observations at the arctic marine LTER site HAUSGARTEN. Ecological Indicators 65:89-102 doi:10.1016/j.ecolind.2015.10.001.
Solvang, H. K., H. Yanagihara, N. Øien & T. Haug, 2017. Temporal and geographical variation in body condition of common minke whales (Balaenoptera acutorostrata acutorostrata) in the Northeast Atlantic. Polar Biology 40(3):667-683 doi:10.1007/s00300-016-1992-0.
Sorteberg, A. & B. Kvingedal, 2006. Atmospheric Forcing on the Barents Sea Winter Ice Extent. Journal of Climate 19(19):4772-4784 doi:10.1175/jcli3885.1.
Stafford, K. M., S. E. Moore, C. L. Berchok, Ø. Wiig, C. Lydersen, E. Hansen, D. Kalmbach & K. M. Kovacs, 2012. Spitsbergen’s endangered bowhead whales sing through the polar night. Endangered Species Research 18(2):95-103.
Stern, H. L. & K. L. Laidre, 2016. Sea-ice indicators of polar bear habitat. The Cryosphere 10(5):2027-2041 doi:10.5194/tc-10-2027-2016.
Stige, L. C., E. Eriksen, P. Dalpadado & K. Ono, 2019. Direct and indirect effects of sea ice cover on major zooplankton groups and planktivorous fishes in the Barents Sea. Ices Journal of Marine Science 76:I24-I36 doi:10.1093/icesjms/fsz063.
Storrie, L., C. Lydersen, M. Andersen, R. B. Wynn & K. M. Kovacs, 2018. Determining the species assemblage and habitat use of cetaceans in the Svalbard Archipelago, based on observations from 2002 to 2014. Polar Research 37(1):1463065 doi:10.1080/17518369.2018.1463065.
Stroeve, J. C. & D. Notz, 2018. Changing state of Arctic sea ice across all seasons. Environmental Research Letters 13 doi:10.1088/1748-9326/aade56.
Torgersen, T. & G. Huse, 2005. Variability in retention of Calanus finmarchicus in the Nordic Seas. Ices Journal of Marine Science 62(7):1301-1309 doi:10.1016/j.icesjms.2005.05.016.
UNEP/AMAP, 2011. Climate Change and POPs: Predicting the Impacts. Report of the UNEP/AMAP Expert Group. Secretariat of the Stockholm Convention, Geneva. 62 pp.
Vacquié-Garcia, J., C. Lydersen, R. A. Ims & K. M. Kovacs, 2018. Habitats and movement patterns of white whales Delphinapterus leucas in Svalbard, Norway in a changing climate. Movement Ecology 6(1):21 doi:10.1186/s40462-018-0139-z.
Vacquié-Garcia, J., C. Lydersen & K. M. Kovacs, 2019. Diving behaviour of adult male white whales (Delphinapterus leucas) in Svalbard, Norway. Polar Research 38(0) doi:10.33265/polar.v38.3605.
Vacquié-Garcia, J., C. Lydersen, T. Marques, J. Aars, H. Ahonen, M. Skern-Mauritzen, N. Øien & K. Kovacs, 2017a. Late summer distribution and abundance of ice-associated whales in the Norwegian High Arctic. Endangered Species Research 32:59-70 doi:10.3354/esr00791.
Vacquié-Garcia, J., C. Lydersen, T. Marques, M. Andersen & K. Kovacs, 2020. First abundance estimate for white whales Delphinapterus leucas in Svalbard, Norway. Endangered Species Research 41:253-263 doi:10.3354/esr01016.
Vacquié-Garcia, J., C. Lydersen, T. A. Marques, J. Aars, H. Ahonen, M. Skern-Mauritzen, N. Øien & K. M. Kovacs, 2017b. Late summer distribution and abundance of ice-associated whales in the Norwegian High Arctic. Endangered Species Research 32:59-70.
Vihtakari, M., J. Welcker, B. Moe, O. Chastel, S. Tartu, H. Hop, C. Bech, S. Descamps & G. W. Gabrielsen, 2018. Black-legged kittiwakes as messengers of Atlantification in the Arctic. Scientific Reports 8(1):1178 doi:10.1038/s41598-017-19118-8.
Wang, Y., H. Bi, H. Huang, Y. Liu, Y. Liu, X. Liang, M. Fu & Z. Zhang, 2019. Satellite-observed trends in the Arctic sea ice concentration for the period 1979–2016. Journal of Oceanology and Limnology 37(1):18-37 doi:10.1007/s00343-019-7284-0.
Wiech, M., S. Frantzen, A. Duinker, J. D. Rasinger & A. Maage, 2020. Cadmium in brown crab Cancer pagurus. Effects of location, season, cooking and multiple physiological factors and consequences for food safety. Science of the Total Environment 703 doi:10.1016/j.scitotenv.2019.134922.
Wisniewska, Danuta M., M. Johnson, J. Teilmann, L. Rojano-Doñate, J. Shearer, S. Sveegaard, Lee A. Miller, U. Siebert & Peter T. Madsen, 2016. Ultra-High Foraging Rates of Harbor Porpoises Make Them Vulnerable to Anthropogenic Disturbance. Current Biology 26(11):1441-1446 doi: https://doi.org/10.1016/j.cub.2016.03.069 .
Wisniewska, D. M., M. Johnson, J. Teilmann, L. Rojano-Doñate, J. Shearer, S. Sveegaard, L. A. Miller, U. Siebert & P. T. Madsen, 2018. Response to “Resilience of harbor porpoises to anthropogenic disturbance: Must they really feed continuously?”. Marine Mammal Science 34(1):265-270 doi:10.1111/mms.12463.
Zhukova, N. G., V. N. Nesterova, I. P. Prokopchuk & G. B. Rudneva, 2009. Winter distribution of euphausiids (Euphausiacea) in the Barents Sea (2000–2005). Deep Sea Research Part II: Topical Studies in Oceanography 56(21):1959-1967 doi: https://doi.org/10.1016/j.dsr2.2008.11.007 .
Årthun, M., T. Eldevik & L. H. Smedsrud, 2019. The Role of Atlantic Heat Transport in Future Arctic Winter Sea Ice Loss. Journal of Climate 32(11):3327-3341 doi:10.1175/jcli-d-18-0750.1.
Årthun, M., T. Eldevik, L. H. Smedsrud, Ø. Skagseth & R. B. Ingvaldsen, 2012. Quantifying the Influence of Atlantic Heat on Barents Sea Ice Variability and Retreat. Journal of Climate 25(13):4736-4743 doi:10.1175/jcli-d-11-00466.1.