In this report Institute of Marine Research IMR has evaluated kelp harvesting in an area in mid Norway from a sustainability perspective based on the best knowledge available. We show how new tools together with environmental monitoring can be used for an ecosystem based and adaptive management of kelp harvesting in Norway.
Bærekraftig taretråling
— Vurdering av bærekraftskriterier ved Vikna
Rapportserie:
Rapport fra havforskningen 2021-46
ISSN: 1893-4536
Publisert: 15.11.2021
Prosjektnr: 14914
Oppdragsgiver(e): Fiskeridirektoratet
Forskningsgruppe(r):
Bunnsamfunn
Tema:
Stortare
Program:
Kystøkosystemer
Approved by:
Research Director(s):
Geir Lasse Taranger, Karin Kroon Boxaspen og Geir Huse
Program leader(s):
Jan Atle Knutsen
English summary
Sammendrag
I denne rapporten har Havforskningsinstituttet brukt kunnskapsgrunnlaget til å vurdere taretråling ved Vikna ut fra et bærekraftspersektiv. Vi viser også hvordan nyutviklede verktøy sammen med miljøovervåkning kan brukes for økosystembasert og adaptiv forvaltning av taretråling i Norge.
1 - Bakgrunn
Viknaområdet har vært stengt for taretråling i flere år ut fra føre-var hensyn. Området har rik tareskog og er oppvekstområde for kysttorsk. Havforskningsinstituttet har gjennomført undersøkelser som kan belyse om denne virksomheten er bærekraftig.
1.1 - Stengning for taretråling ved Vikna
Et område sør av Vikna har vært stengt for taretråling siden 2015 som et føre var-tiltak i påvente av mer kunnskap om tareskogens betydning for gyteaktiviteten til torsk. Viknaområdet har flere viktige gyteområder for kysttorsk. Bruken av føre-var krever oppfølgende undersøkelser og siden stengningen har Havforskningsinstituttet gjennomført flere forskningsprosjekter for å bedre kunnskapsgrunnlaget om viktige effekter av denne aktiviteten og øke forståelsen av ressursgrunnlaget. I denne rapporten bruker vi resultater fra både forskning og overvåkning til å foreslå kriterier for bærekraftig taretråling ved Vikna. Kriteriene danner også grunnlag for bærekraftig, økosystembasert og adaptiv forvaltning av taretråling langs hele Norges kyst.
1.2 - Stortareskog i Norge og trusler mot tareskog i nord
Stortare ( Laminaria hyperborea ) er vår viktigste marine makroalge, og Norge har Europas største bestander av denne arten (Faktaboks 1). Denne taren danner tette skoger i bølgeeksponerte områder langs norskekysten. Tareskogene har høy biomasseproduksjon og skaper et tredimensjonalt miljø som er tilholdssted for en rekke organismer. På hver kvadratmeter tareskog kan det finnes 100 000 små krepsdyr, snegl, muslinger, pigghuder og børstemark (Norderhaug et al. 2002, Christie et al. 2003). Disse dyrene er hovedføden til mange av kystfiskebestandene som bruker tareskogen både som føde- og oppvekstområde (Norderhaug et al. 2005, 2020). Tareskogene huser små kutlinger (Gobiidae), flere arter leppefisk som bergnebb (Ctenolabrus rupestris) som lever stasjonært, bunnfisk som torsk (G adus morhua) og ulker ( Myoxocephalus spp.), ung sei ( Pollachius virens ) som beiter over tareskogen og lyr (Pollachius pollachius) på jakt etter annen fisk.
Tareskogene er også viktige beiteområder for kystnære sjøfugl som storskarv (Phalacrocorax carbo), toppskarv (Phalacrocorax aristotelis) og teist (Cepphus grylle), som jakter på unge årsklasser av sei og andre småfisk med tilknytning til tareskog (Dehnhard et al. 2021, Hillesøy og Lorentsen 2012). Tareskogsområdene ser også ut til å være en nøkkelressurs for de sterkt reduserte bestandene av krykkjer (Rissa tridactyla), alke (Alca torda) og lomvi (Uria aalge) når de mer pelagiske byttedyrene uteblir. Også sjøpattedyr som sel (Bjørge et al. 2002) og nise finner mat i og rundt tareskogen.
|
Tang og tare (fra hi.no)
Norge har Europas største bestander av tang og tare. Ca. 10.000 kvadratkilometer av norskekysten er bevokst med tang og tare, noe som tilsvarer landets areal av dyrket mark. Begrepet tang og tare brukes ofte som en fellesbetegnelse på store, fastvoksende alger (eller makroalger), men i virkeligheten dreier det seg om to ulike ordener innenfor klassen brunalger (Phaeophyceae). Tilstanden i taresamfunnene og effekter av tarehøstingen undersøkes gjennom Havforskningsinstituttets overvåkingsprogram.
Både tang og tare krever et hardt substrat (fjell eller steinbunn) som festepunkt. Tangartene dominerer på hardbunnslokaliteter i fjæresonen, der enkeltartene tilsynelatende opptrer i avgrensede dyp, slik at det dannes horisontale vegetasjonsbelter.
Vegetasjonsbeltene domineres av arter som spiraltang (Fucus spiralis), blæretang (F. vesiculosus), grisetang (Ascophyllum nodosum) og sagtang (F. serratus), i et mønster som går igjen langs store deler av norskekysten. Tareartene tåler i liten grad tørrlegging og vokser nedenfor lavvannsmerket, ned til ca. 30 meters dyp. Det er hovedsakelig stortare (Laminaria hyperborea) som danner tareskogene langs norskekysten, mens arter som sukkertare (Saccharina latissima), butare (Alaria esculenta) og fingertare (Laminaria digitata) kan dominere i enkelte områder.
Tareskogenes betydning for livet langs kysten gjør at tap av tareskog på grunn av kråkebollebeiting har hatt store økologiske konsekvenser. Kråkebollene kan beite ned all tare over store områder, og står for det største biomassetapet av tareskog langs norskekysten, det meste av tareskogene langs kysten av Midt- og Nord-Norge har vært eller er beitet ned. Årsaken til oppblomstringen av kr å keboller var med stor sannsynlighet overfiske frem mot 1970-tallet (Norderhaug et al. 2020). Modernisering av kystfiskeflåten førte til utfisking av rovfisk med kråkeboller på menyen, som torsk, hyse og steinbit. Også tareskogen rundt Vikna har tidligere vært preget av nedbeiting, men på 1990-tallet kom tareskogen tilbake her, og i andre sørlige deler av nedbeitingsområdet. Oppvarming av kystvannet reduserte rekrutteringen av kråkeboller og førte til at nye rovdyr, krabber, ekspanderte nordover (Fagerli 2013, 2014). |

1.3 - Taretråling i Norge
I Norge høstes stortare gjennom tråling for utvinning av alginat, og dette er en av flere goder mennesker nyter godt av fra tareskog (Figur 2). Trålingen foregår på kyststrekningen Rogaland–Trøndelag. Tarehøsting reguleres gjennom fylkesvise forskrifter der kystområdene deles inn i felt som er åpne for høsting hvert femte år. På grunnlag av Havforskningsinstituttets overvåking av stortarevegetasjonens tilstand og kråkebolleforekomstene gjøres vurderinger av feltenes egnethet for tarehøsting som grunnlag for Fiskeridirektoratets forvaltning (Steen 2019, 2020, 2021).

Taretråling tar først og fremst de store tareplantene, mens de små tareplantene som overlever vokser raskere pga. bedre lysforhold, og vil med tiden reetablere tareskogen. På grunn av kuperte bunnforhold, som er lite tilgjengelig for taretrålen, er det vanligvis store partier med uberørt tareskog i områdene der det høstes stortare. Selv om taretrålingen er et avgrenset inngrep, vil likevel tareskogens økologiske funksjon reduseres i flere år avhengig av uttaksgraden og tarevegetasjonens reetableringsevne. I Norge tråles det årlig omtrent 150 000 tonn tare til en verdi av i overkant av 1 milliard kroner. Det er ikke kjent hvor stor total biomasse av stortareskog det finnes langs kysten, men anslag for all tareskog (både stortare- og sukkertareskog) tilsier i størrelsesorden godt over 50 millioner tonn (Gundersen et al. 2021). Per i dag er det bare en operativ aktør i markedet, men det forventes at flere aktører vil høste tare i fremtiden. Det vil derfor bli mer behov for bedre kunnskap om ressursgrunnlaget og hvordan tareskogen påvirkes og restitueres i ulike deler av landet.
1.4 - Oppdraget til Havforskningsinstituttet
Fiskeridirektoratet har bedt Havforskningsinstituttet å vurdere om det er biologisk forsvarlig utifra eksisterende kunnskapsgrunnlag at de deler av Vikna som ble stengt for tarehøsting i 2015 (fra høstefelt 414E til 420A) og som ikke er vernet gjennom annet lovverk (naturmangfoldloven, mm), kan åpnes for en begrenset tarehøsting. Fiskeridirektoratet har også bedt Havforskningsinstituttet utarbeide et forskningsopplegg for å kunne gi råd om hvilke kvantumsbegrensninger som skal settes på tarehøsting i de aktuelle feltene i Vikna. Dette innbefatter bruk av kunnskapsgrunnlaget som er opparbeidet etter at området ble stengt for tarehøsting for å beregne tareressursgrunnlaget på de aktuelle feltene, samt å utarbeide kriterier som skal legges til grunn for å vurdere hva som utgjør et forsvarlig uttak i høstefeltene.
I forskningssammenheng anbefalte Havforskningsinstituttet at det gjeldende høsteforbudet midlertidig ble opprettholdt for innhenting av grunnlagsdata, men at det kan gjennomføres begrenset forsøkstråling av tare innenfor områder nærmere angitt av Havforskningsinstituttet som del av disse studier. For å få bedre kunnskap om tareforekomstene i Vikna-området foreslo Havforskningsinstituttet å:
-
Etablere biomassemodell for tare i Vikna (for å beregne ressursgrunnlaget).
-
Lage kriteriesett for biologisk forsvarlig høsting (bærekraft)
-
Etablere miljøovervåkningsprogram for effektene av taretråling på tareskog ved Vikna (som en viktig del av adaptiv forvaltning).
I denne rapporten presenterer vi forslag til 1-3 over som sammen med annen forskning Havforskningsinstituttet har gjennomført etter 2015 danner grunnlag for råd om bærekraftig taretråling ved Vikna. Kunnskapsgrunnlaget presenteres kapittel for kapittel for å gi en mer kvantitativ forståelse av konsekvensene av taretråling og mulighet for forvaltningen til å kunne vurdere uttak av tare i forhold til ressursgrunnlaget og opp mot andre økosystemtjenester. I kapittel 2 presenteres Viknaområdet og taretråling som har vært gjennomført tidligere, kapittel 3 gjennomgår kunnskapsgrunnlaget for effekter av taretråling og kapittel 4 gjennomgår bærekraftsbegrepet og hvordan det er brukt her. I kapittel 5 presenteres biomassmodellen for stortare som er kjørt for Viknaområdet. Kapittel 6 viser hvordan denne modellen kan danne grunnlaget for et forvaltningsverktøy som viser kvantitativt hvordan ressursgrunnlaget, økosystemet og dets tjenester påvirkers av taretråling. Kapittel 7 presenteres forslag til miljøovervåkningsprogram for Vikna for å overvåke omfang av tråling og restitusjon av tareskogen etter tråling. Havforskningsinstituttets råd og anbefalinger for taretråling ved Vikna gis i kapittel 8, men rådene er ment som grunnlag for tareforvaltning i Norge.
2 - Viknaområdet og taretråling
Viknas skjærgård huser rike tareskogsressurser og er gyteområder for fisk og fødeområder for sjøfuglbestander. Etter 2015 har det ikke vært gjennomført kommersiell taretråling i området.
2.1 - Viknaområdet og høstefelt
Skjærgården utenfor og sør for Vikna er kupert med øyer og skjær. Rundt finnes rike tareskoger. De største skogene finnes i grunne områder ut mot bølgeeksponerte områder i vest. Området huser rike kystbestander og har viktige gyteområder for kysttorsk og hekke- og beiteområder for sjøfugl. Det er etablert mindre sjøfuglreservater for hekkende sjøfugl. I indre, mer beskyttede områder mot øst er taren mindre og mer av bunnen er bløtbunn. Området for taretråling omfatter 7 høstefelt for tare fra 414E til 420A som hver har en utstrekning på en nautisk mil (eller et breddeminutt) i nord-sør retning (Figur 3).
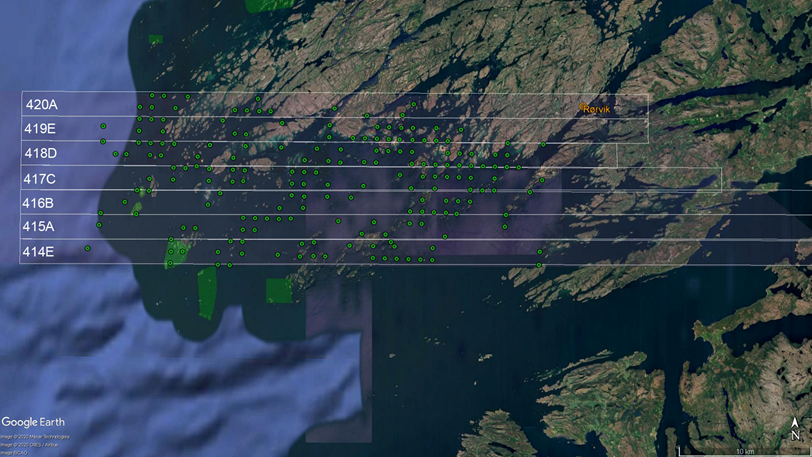
2.2 - Taretråling ved Vikna
Det har tidligere vært gjennomført forsøkshøsting av stortare i hele eller deler av høstefeltene i området i perioden 2010-2014 (Tab. 1, Steen et al. 2014, 2016). Høstefelt 418D var i tillegg åpent for tarehøsting i ca en måned i 2015. Etter dette har området vært stengt for taretråling, bortsett fra forskningstråling av i underkant av 3 000 tonn tare i et avgrenset område på høstefelt 416B i forbindelse med KelpFish-prosjektet i 2018 (Figur 4, Kapittel 3.3, Norderhaug et al. 2020).
|
År Felt |
2010 (tonn) |
2011 (tonn) |
2012 (tonn) |
2013 (tonn) |
2014 (tonn) |
2015 (tonn) |
|---|---|---|---|---|---|---|
| 420A | 3536 | |||||
| 419A | 4129 | |||||
| 418D | 1808 | 3469 | ||||
| 417D | 5383 | |||||
| 416B | 5138 | |||||
| 415A | 107 | |||||
| 414E |

3 - Kunnskapsgrunnlag for effekter av taretråling
De økologiske effektene av taretråling avhenger av hvor mye av den stående biomassen som tråles, hvor store effekter trålingen har på tareskogen der det tråles og hvor raskt økosystemet restitueres etter tråling. Gjennom forskningsprosjekter og miljøovervåkning har Havforskningsinstituttet har undersøkt disse spørsmålene systematisk.
3.1 - Metodeutvikling
Det har vært nødvendig å utvikle nye metoder for å dokumentere effektene av taretråling og dette har vært en sentral del i forskningsprosjektene som har vært gjennomført. De økologiske effektene av taretråling avhenger av hvor mye av den stående biomassen som tråles, hvor store effekter trålingen har på alle nivåer av næringskjeden der det tråles og hvor raskt økosystemet restitueres etter tråling. Det har ikke vært kjent hvor store tareressursene er eller hvor stort uttaket er i forhold til ressursgrunnlaget i høstefeltene. Tarevegetasjonen overvåkes regelmessig, men kunnskap om effekter høyere opp i næringskjeden er liten. En grunn til dette er at studier av fisk og sjøfugl krever forsøk på en stor skala som er ressurskrevende. Utvikling av nye metoder i senere år har gjort det mulig å samle inn gode og sammenlignbare data om fisk og følge sjøfugls adferd på en relevant skala.
3.1.1 - Ressursmodell for stortare
Det kanskje største kunnskapshullet har vært knyttet til beregninger av ressursgrunnlaget. Det har ikke vært kjent hvor store ressursene er innen hvert høstefelt og det har ikke vært satt begrensninger på uttak. Derfor har Havforskningsinstituttet utviklet en romlig biomassemodell for stortare som predikerer hvordan tarens biomasse varierer i områder med ulike miljøforhold (van Son et al. 2019, Norderhaug et al. 2020). Fordi modellen eksplisitt predikerer biomasse direkte, kan man ved bruk av uavhengige valideringsdata få gode mål på usikkerhet og hvor god modellen er. Dette er nødvendig for å gjøre gode vurderinger av hva modellen egner seg til å brukes til. Modellen har vært kjørt for Vikna og er presentert i kapittel 5.
3.1.2 - Stereovideo
Det har skjedd mye innen observasjonsteknologi for vitenskapelige studier i senere år (Duffy et al. 2019) og nye metoder måtte tas i bruk for å sammenligne data fra tett tareskog og åpne trålgater. Prosjektet Kelpfish ble etablert for å undersøke de økologiske effektene av taretråling. En hovedoppgave i prosjektet var å teste nye metoder for å kunne telle og måle fisk. Det finnes lite data om hvordan fisk bruker tareskog og blir påvirket av aktiviteter som taretråling. Dette kommer blant annet av at det er vanskelig å samle sammenlignbare data i og utenfor tareskog fordi tradisjonelle fiskemetoder fisker ulikt i og utenfor tett vegetasjon og visuelle metoder som kamera har utfordringer på grunn av ulik sikt i tett tareskog og åpne trålgater. Stereovideokamera (Figur 5 og 7) er imidlertid en måte å overkomme denne systematiske feilen fordi stereovideofilming gir «dybdesyn» som gjør det mulig å måle avstander og lengder på fisk. Dermed kan man telle fisk i like store vannvolum i tett vegetasjon som i åpent lende. Vi laget derfor rigger for to GoPro-kameraer som var kalibrert for å se i stereoanalyseverktøy etterpå.

3.1.3 - Bunnstående ekkolodd
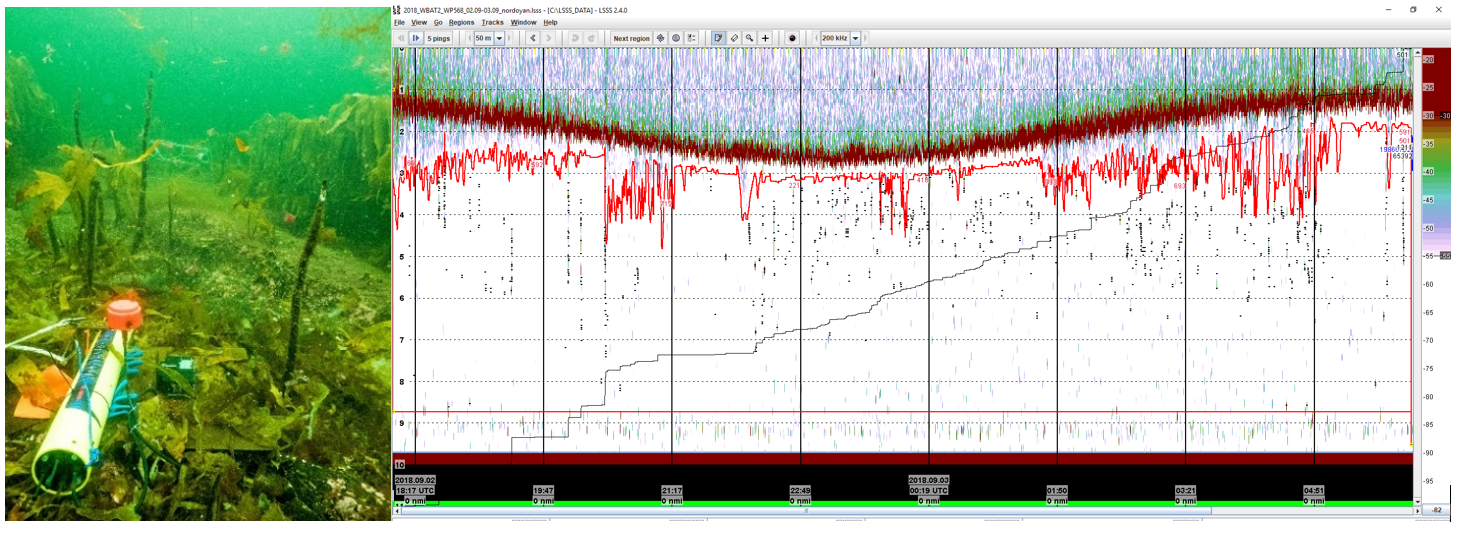
Visuelle metoder som kamera har imidlertid begrensinger i dårlig lys og selv nattlys kan påvirke fisk og andre dyrs adferd. Akustikk er en måte å måle antall eller biomasse av fisk og andre dyr gjennom hele vannsøylen som gir like gode data om dagen som på natten. Vi testet bunnstående ekkolodd med transduser stilt oppover. I 2017 testet vi bruk av akustikk med et Simrad EK15 med 200kHz transduser og kasse med pc og batteri på land. I 2018 brukte vi Simrads undervannsenhet WBAT med 200 kHz transduser der alt er pakket inn i en enhet (Figur 6). Begge år brukte vi oppadrettede GoPro-cameras sammen med transduseren for å verifisere fisk i ekkogrammene mens det var nok lys.
Akustikk har også stort potensiale i overvåkning av tarens biomasse fordi det er en metode som samler data raskt for store områder og som kan analyseres i kartverktøy og brukes i biomassemodellen. Kombinasjonen biomassemodell og overvåkning med bruk av akustikk kan vise seg å bli en effektiv forvaltningsverktøy i fremtiden. Vi har derfor foreslått å teste akustikk til dette formålet i Viknaområdet (kapittel 7).
3.1.4 - Dykking kan ikke erstattes
Dykking er en nødvendig del av vitenskapelig innsamling på grunt vann. Dykking er en metode som ikke kan erstattes av ny teknologi, hverken i dag eller i uoverskuelig fremtid. Tilstedeværelse selv gir mulighet til å observere arter, miljøet og økosystemene direkte og gjennomføre innsamling nøyaktig og representativt (Figur 7). I arbeidet med å utvikle biomassemodellen var dykking nødvendig for å teste om modellens prediksjoner stemte overens med virkeligheten og gi kvantitative tall på hvor god korrelasjonen mellom modellen og virkeligheten var. Modellerte data har liten verdi hvis det ikke kan vises at de predikerer det vi finner når vi går ut i felt.

I Kelpfish-prosjektet var dykkere nødvendige for å samle inn tare i ruter, påvekstalger og evertebrater. Dykkere måtte også brukes for å plassere ekkolodd og stereovideorigger riktig, enten de skulle filme langs trålgatene eller langs kanten av tareskoen rundt. Sensorere og kameraer kan heller ikke erstatte observasjoner gjort på stedet, enten det er på land eller i vann og dykkerens observasjoner var en nødvendig del av helhetsforståelsen i studiet.

3.2 - Ressursgrunnlaget
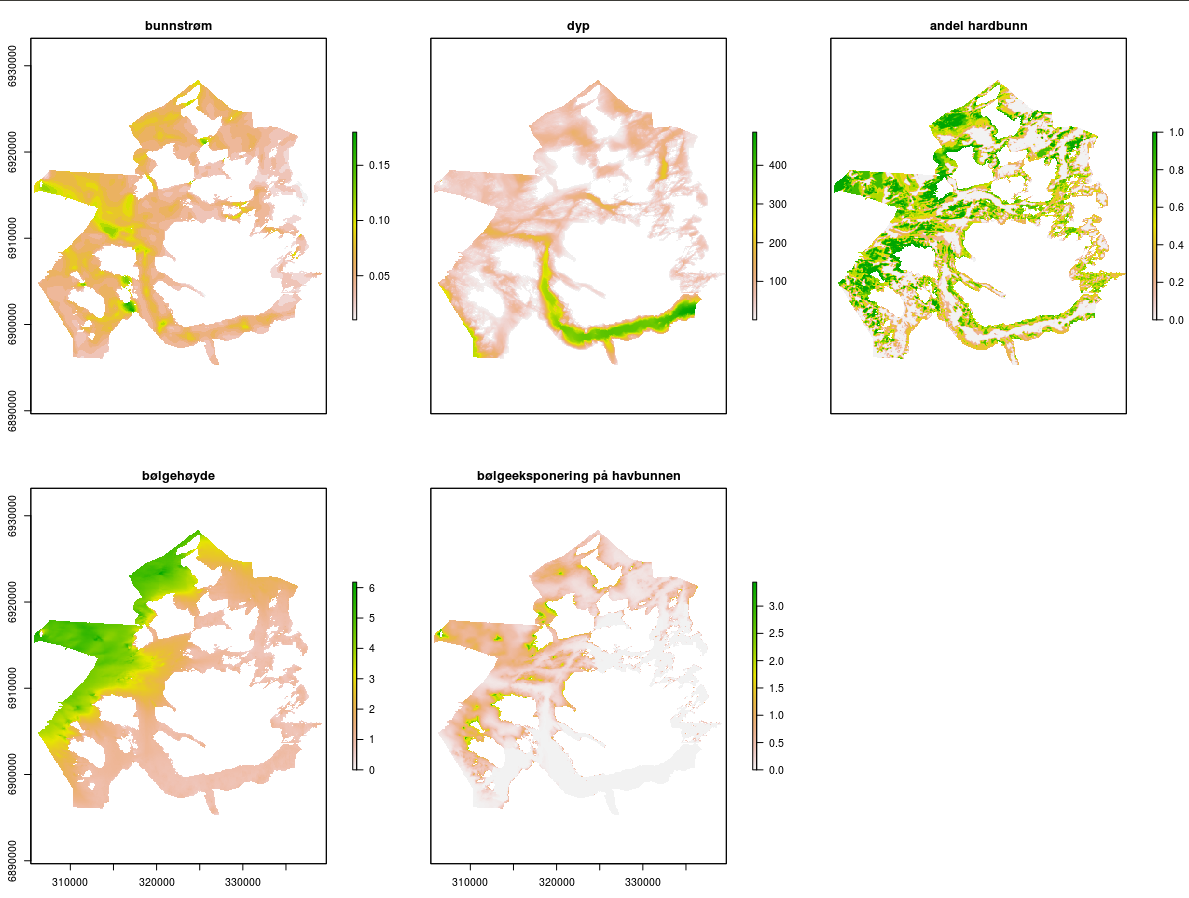
I Ressursmodell for stortare laget Havforskningsinstituttet en romlig biomassemodell for stortare. Denne biomassemodellen ble utviklet i et område i Møre og Romsdal der vi hadde tilgjengelig høyoppløselige kartlag om viktige miljøforhold som bestemmer tarens utbredelse, størrelse og vekt (figur 9). Ved bruk av en statistisk modell kunne vi lage en god modell (van Son et al. 2020) som siden kan brukes som referanse for enklere, grovere modeller for større områder der vi ikke hadde like gode data om miljøforholdene (Norderhaug et al. 2020). Når modellert tarebiomasse ble sammenlignet med høstestatistikken for pilotområdet på Møre var uttaket i løpet av en 5 års høstesyklustotalt cirka 6% av den stående biomassen av stortare. Prosjektet ble finansiert av Nærings- og fiskeridepartementet og resultatene er publisert i det anerkjente vitenskapelige tidsskriftet Frontiers in Marine Science (van Son et al. 2020 https://doi.org/10.3389/fmars.2020.00107).
En viktig del av modellering er å beregne hvor god modellen stemmer med virkeligheten. Mål på usikkerhet viser hvor nøyaktig modellen er og er dermed avgjørende for hva den kan brukes til. Det ble samlet inn uavhengige data, blant annet av dykkere som høstet hele kvadratmetere med tare for å måle vekten, som ble brukt til å teste hvor god modellen predikerte tarens biomasse (Figur 10). De uavhengige valideringsdataene korrelerte 0,85 med modellens prediksjoner, noe som er svært bra for kartmodeller. Uavhengig validering av den enklere modellen ga korrelasjon på 0,69 noe som også gir god relasjon mellom modell og virkelighet. Denne enklere modellen ble brukt i Viknaområdet og er presentert i kapittel 5.

3.3 - Økosystemeffekter
Tråling av tare kan påvirke hele økosystemet tareskog og alle artene som lever der. Det finnes en del kunnskap om hvordan taren selv og samfunn av små alger og dyr påvirkes og restitueres etter tråling, fra Havforskningsinstituttets overvåkning av taretråling (f eks Steen et al. 2020), men også fra eldre studier (Chrstie et al. 1998). Kunnskapen om påvirkning høyere opp i næringsnettet er imidlertid liten. I prosjektet KelpFish- Effekter av taretråling på tareskogens økosystem - brukte Havforskningsinstituttet taretråling som vitenskapelig metode for å undersøke effektene av taretråling på fire nivåer av tareskogens økosystem; primærprodusenter representert ved tare og små påvekstalger på tarens stilk, sekundærkonsumenter i form av evertebratfauna, småfisk og yngel som spiser disse smådyrene og rovfisk som jakter mindre fisk. Ved bruk av nye innsamlingsmetoder som stereovideo og akustikk (kapittel 3.1) i kombinasjon med tradisjonelle metoder som teiner som kan sammenlignes med historiske data, kunne vi samle inn sammenlignbare data fra tett tareskog og åpne trålgater. Prosjektet ble finansiert av Nærings- og fiskeridepartementet og resultatene er publisert i open access i det anerkjente vitenskapelige tidsskriftet Marine Ecology Progress Series (Norderhaug et al. 2020 https://doi.org/10.3354/meps13426).

Studien ble gjennomført som en såkalt BACI-studie (Before-After, Control-Impact), der to områder ble undersøkt før (before) og etter (after) taretråling i det ene området (Impact) og der det andre området ikke ble trålt men fungerte som et kontrollområde (control) for naturlig år til år-variasjon. Et område på ca 15 km2 i skjærgården ved Nordøyan (Figur 11) ble trålt og et like stort område med like miljøforhold ved Sørøyan ble brukt som kontrollområde. I september 2017 ble før-undersøkelsene gjennomført på stasjoner som ble valgt randomisert, tråling av ca 3.000 tonn tare ble gjennomført av tarenæringen (Dupont) inne i trålområdet i mai 2018. Etter-undersøkelser ble gjennomført i september 2018, det vil si i samme måned som før-undersøkelsene og etter at forstyrrelsen som trålingen representerer hadde fått tid til å legge seg.
Teiner og RUVs ble plassert i tett tareskog (før) og i trålgater (etter) tråling ved Nordøyan og i kontrollområdet ved Sørøyan. RUVs ble også plassert langs kanten av trålgatene etter tråling rettet mot tareskogen rundt for å måle kanteffekter på tareskogen som står igjen rundt trålgatene (Figur 12). Ikke alle metoder virker for all arter og de utfyller hverandre med informasjon. Teinene ble agnet og samlet krabber og fisk på fødejakt, mens RUVs er passive observatører. For eksempel ble små fisk som kutlinger observert på video mens teiner ikke egnet seg til å fange disse (for små og agnet står ikke på menyen) eller lyr (for store). Det ble observert mer sei både i teiner og på video inne i trålgatene etter tråling, men ekkolodd som fanger mengden fisk gjennom hele vannsøylen registrerte ingen endring i biomassen av fisk. Dette ble tolket som at det ikke ble mer sei i vannsøylen totalt på grunn av tråling, men at fisken fordelte seg helt ned til bunnen når tareskogen ble åpnet opp. Taskekrabber ble fanget i teiner, men det ble observert for få taskekrabber på video til at dataene kunne behandles statistisk.

Resultatene fra studien viste at taretråling fragmenterer tareskogen og reduserer mengden store tarer i «trekronene», men at 74% av tareskogen sto igjen på trålstasjonene etter tråling. Resultatene fra målinger inne i trålgatene viste effekter over 4 nivåer i næringskjeden og disse er summert i tabell 2. I trålgatene ble det funnet en konsistent reduksjon for alle grupper/arter lavt i næringskjeden. Mengden av store tarer og tarens påvekstalger (makro-og mikrohabitat) ble redusert vesentlig. Med dem forsvant mye av det rike livet med svømmende evertebrater som lever blant påvekstalgene og stedegen fisk som bergnebb som holder seg nær enkeltplanter og beiter på smådyrene. Vi fant også en generell reduksjon i mengde fisk og endret fordeling av fisk i trålgatene og i tareskogen rett ved siden av trålgatene (såkalte kanteffekter) sammenlignet med i tett tareskog. Denne reduksjonen gjaldt både småfisk som kutlinger, juvenile fisk samt større rovfisk som lyr som passerer gjennom skogen. Blant arter høyt i næringskjeden varierte effektene mye avhengig av hvordan disse artene brukte tareskogen i matsøk. Småsei beiter i vannsøylen over tarebladene og ble i liten grad påvirket av åpningene i skogen som ble laget av trålgatene. Lyr som cruiser under taredekket på jakt etter fisk forsvant nesten helt. Krabber som lever på bunnen ble i liten grad påvirket av at taren forsvant. Det er verdt å merke seg at det ble observert svært lite torsk i hele studiet og for få individer ble fanget til å kunne gjennomføre statistiske tester.
| Funksjonell gruppe | Funksjon i tareskogen | Effekt av taretråling | ||
| Nivå 1: Primærprodusenter | ||||
| Stortare | Skogdannende tredimensjonal struktur, primærproduksjon | -0,7 | ||
| Påvekstalger | Mikrohabitat, biomangfold | -0,8 | ||
| Nivå 2: Sekundærkonsumenter | ||||
| Evertebrater | Føde for fisk, biomangfold, primærproduksjon | -1,0 | ||
| Innsamlingsmetode | Teiner trålgate | Stereovideo trålgate | Stereovideo tareskogkant | |
| Taskekrabber | Rovdyr, byttedyr | -0,2 | ||
| Nivå 3: Rovfisk som spiser evertebrater | ||||
| Fiskeyngel (sei og lyr) | Oppvekstområde for fisk | -1,1 | -1,0 | |
| Kutling | Rovdyr, byttedyr | -0,4 | -3,2 | |
| Bergnebb | Stedegen fisk, Rovdyr, byttedyr | -0,9 | -3,2 | -3,3 |
| Sei | Over tareskogen, rovdyr, byttedyr | +0,3 | +0,4 | 0,0 |
| Nivå 4: Rovfisk som spiser fisk | ||||
| Torsk | Rovfisk | -0,1 | ||
| Lyr | Rovfisk | -3,2 | -3,3 | |
Kelpfish viste at taretråling ikke bare har effekter på taren selv men gjennom 4 nivåer i økosystemet opp til rovfisk, og påvirker både tareskogens biomangfold og rolle som primærprodusent og funksjon som oppvekstområde for kystfiskbestander. Prosjektet viste imidlertid også at taretråling representerer et begrenset inngrep og at en del av tareskogen står igjen på trålstasjonene.

3.4 - Miljøovervåkning og tareskogens restitusjon
Gjennom miljøovervåkning av taretråling følger Havforskningsinstituttet med på restitusjon av tareskogen etter tråling langs hele kysten. Prosjektet er finansiert av Nærings- og fiskeridepartementet og resultatene publiseres årlig i Havforskningsinstituttet rapporter (for eksempel Steen 2019, 2020, 2021 Steen et al. 2020). Overvåkingen gjøres primært ved hjelp av undervannsvideo langs transekter på stasjoner i høstefelt og i referanseområder som er stengt for tarehøsting. Overvåkingsstasjonene er valgt ut på basis av dybdekart og innrapporterte sporingsdata etter tidligere tarehøsting, og tarevegetasjonens tilstand på høstefeltstasjonene sammenlignes med tilstanden på referansestasjonene. For detaljerte undersøkelser av tarevegetasjonens aldersstruktur og påvekstsamfunn (epifytter) er det på enkelte stasjoner også samlet inn tareplanter ved hjelp av tareskrape/taretrål. Resultatene viser at tarens biomasse forventes å være restituert etter ca 4 år ved Viknas breddegrad, men at økosystemet tar minst 6 år for å komme tilbake etter tråling (Steen et al. 2016, 2020). Resultater fra overvåkningen med relevans for Vikna-området er brukt i analyser av økosystemets restitusjon i denne rapportens kapittel 6.2.
3.5 - Gyteområder
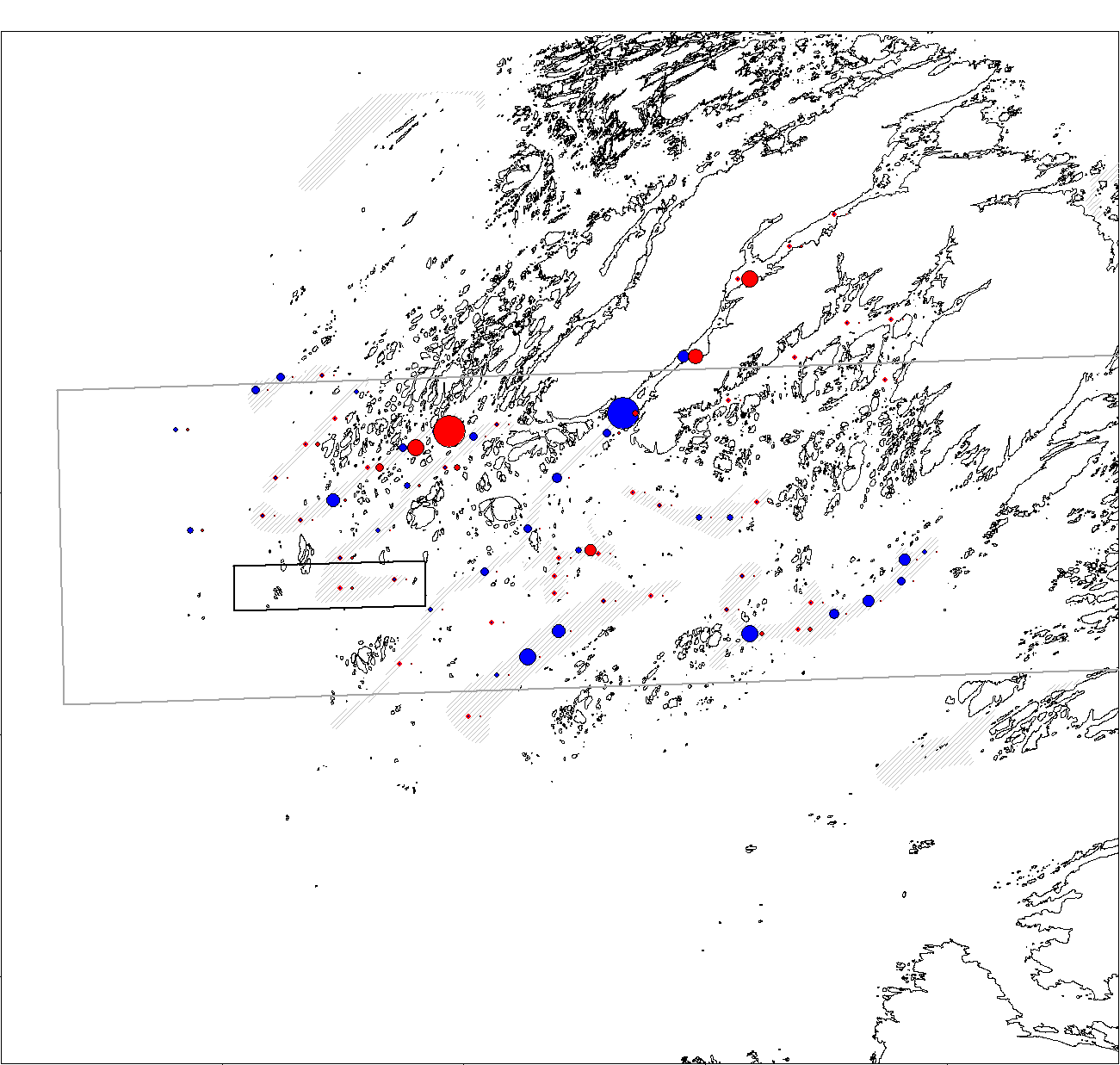
Hjemmelen for stengning av Vikna-området var forankret i et føre-var vedtak for å beskytte viktige gyteområder for torsk. Viknaområdet er et viktig område for rekruttering av kysttorsk. For å undersøke mulige effekter av taretråling på gyting av torsk ble det i forbindelse med Havforskningsinstituttets program for kartlegging av gytefelt og oppvekstområder for kommersielt viktige arter i kystsonen gjennomført innsamling av ny-gytte egg på 60 stasjoner i og rundt forbudsområdet i Vikna, henholdsvis før (mars 2018) og etter (mars 2019) forsøkstrålingen av 3000 tonn stortare i Kelpfish-prosjektet.
Begge årene ble det funnet relativt store mengder egg som ble artsbestemt til å være torskeegg. En forholdsvis stor andel (83%) av disse eggene var helt ny-gytte, det vil si mindre 3-6 dager gamle. Tilsvarende andel i et større område av kysten av Trøndelag i 2018 var 76%, noe som tilsier at tallene på antall egg vil være en god indikator på produksjon av egg.
Resultatene fra gytefeltundersøkelsene viste at fordeling av torskeegg varierte noe mellom 2018 og 2019, men totalmengden av egg var omtrent den samme (793 egg i 2018 mot 876 egg i 2019, figur 14). Det var relativt sett en mindre økning i antall egg fra 2018 til 2019 i høstingsfeltet enn utenfor, men alle forskjeller i antall egg er innenfor det som regnes som tilfeldig variasjon. Denne studien tyder ikke på at taretråling i 2018 påvirket gyting i 2019 men datagrunnlaget med en måling før og en måling etter tråling gir et begrenset konklusjonsgrunnlag
Men hvor blir eggene som gytes ved Vikna av?
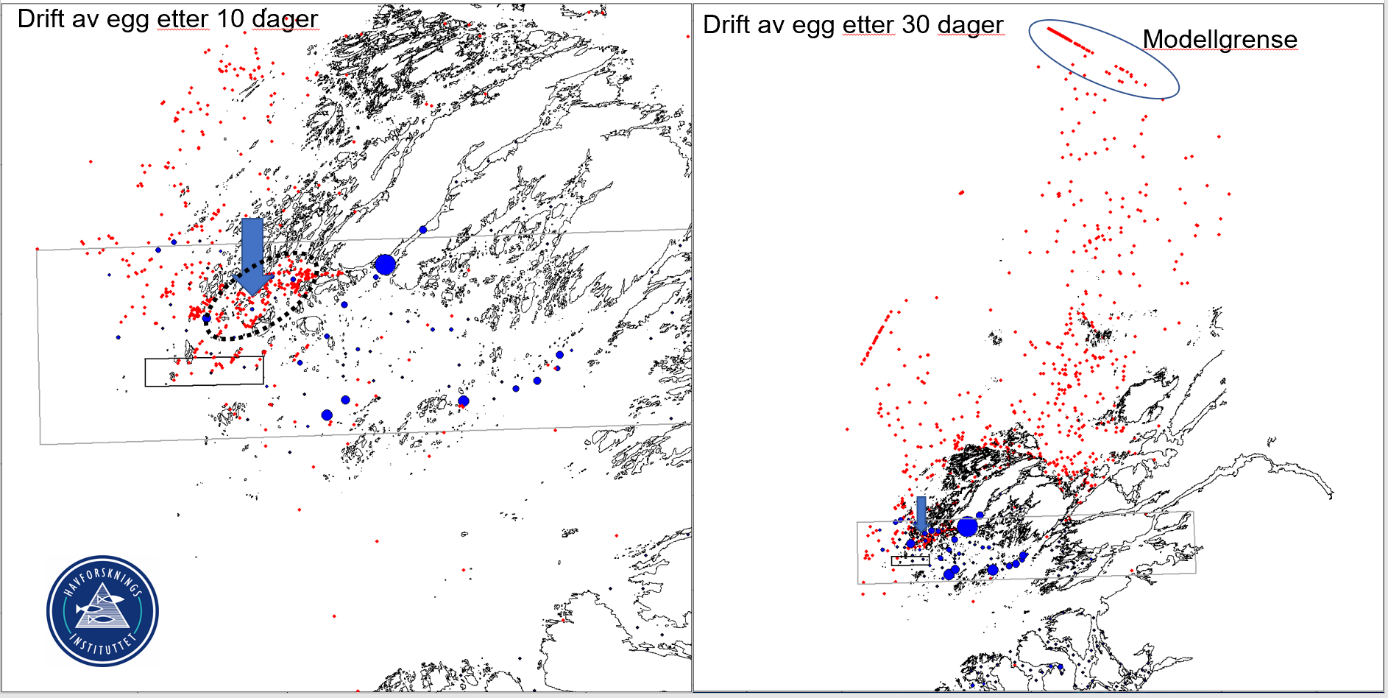
I tillegg til innsamling av egg ble det gjennomført en modelleringsstudie for å predikere spredningen av egg i Viknaområdet (figur 15). Modelleringen baseres seg på havmodellen ROMS (Regional ocean modeling system) med en partikkelspredningsmodell (LADIM) som kjøres som standard metode for kartlegging av gytefelter.
Modellen indikerer at eggene som gytes i forbudssonen ved Vikna i stor grad vil drifte nordover og ut av Viknaområdet. Bare 16% av eggene drev kortere enn 10 km fra opprinnelsesstedet i løpet av 30 dager og hele 25% av eggene drev ut av modellområdet (markert som en linje egg på figur 15). Dette tyder på at driften av egg er mye større enn gytefelter kartlagt i indre deler av kysten. Dette betyr videre at det er lite retensjon av yngel i Vikna område og at yngel som bruker dette område som oppvekstområde sannsynligvis drifter inn fra andre gytefelter lengre sør.
3.6 - Sjøfugl
Det finnes få studier om effekter av taretråling på dyr høyere opp i næringsnettet som for eksempel sjøfugl. Mange sjøfugl bruker tareskogen for å finne mat og om taretråling påvirker sjøfuglene er avhengig av hva som skjer med næringsforholdene, altså gjennom trofiske interaksjoner som fører til en endret tilgang på byttedyr. Som visuelle predatorer kan tråling påvirke fuglens næringssøk på ulike måter: Når tare blir fjernet kan fisken bli et lettere bytte fordi de naturlige gjemmestedene i tareskogen forsvinner. Alternativt kan mattilgangen for sjøfuglene bli redusert fordi mengden av egnet fiskehabitat reduseres og sjøfuglene dermed må dykke dypere eller fly lengre for å få tak i mat. Omfanget av effektene vil altså henge sammen med adferden til fisken etter taretråling samt mengden av alternative beiteområder for de påvirkede sjøfuglene.
I en studie ved Sklinna nord for Vikna ble toppskarv instrumentert med GPS-loggere i hekkesesongen over seks år (2013-2018) for å studere beiteadferd hos hekkende toppskarv i forhold til taretrålingsaktivitet i området (Christensen-Dalsgaard et al. 2020). Toppskarven fra Sklinna spiser mest småsei som står i stimer over tett tareskog (Hillesøy og Lorentsen 2012), og det er vist at toppskarvens hekkesuksess er knyttet til andelen av sei i dietten (Bustnes et al. 2013). Kelpfish prosjektet (kapittel 3.3) viste at når taren tråles bruker småseien hele vannsøylen ned til bunnen slik at toppskarven kanskje må dykke dypere for å fange sei. Samtidig kan ikke seien gjemme seg i «trekornene» når toppskarven dykker. Christensen-Dalsgaard et al. (2020) analyserte overlapp av taretråling og beiteaktivitet til toppskarv i rutenett på 1,5 km2 og dokumenterte derved at tarehøsting i stor grad foregår til samme tid og i de samme områdene som hekkende toppskarv beiter. Dette viser at det potensielt kan være konflikt mellom taretråling og næringssøk for sjøfugl. Resultatene viste imidlertid at når man sammenlignet den totale dykkeadferden i ruter før og etter tarehøsting, var det ingen endring i dykkeadferd til toppskarven. Resultatene viste altså at for større beiteområder vurdert samlet, var det ikke noe som tydet på at sjøfugl bruker trålte områder mer eller mindre til matsøk. Områdene som brukes som beiteområder for hekkende toppskarv fra Sklinna er rike på tareskog (Christensen-Dalsgaard et al. 2017). Det kan altså tenkes at det her er så rikelig tilgang på alternative beiteområder at fuglene bare kan oppsøke tilstøtende områder etter taretråling har funnet sted. I motsetning til dette dokumenterte Lorentsen et al. (2010), i en annen studie av effekten av taretråling, at beitende storskarv tilsynelatende responderte på endret næringstilgang ved å dykke mer i områder hvor det var høstet tare enn ikke-høstede områder.
De to norske studiene har dokumentert at det er et overlapp i skarvenes beiteområder og områder der det høstes tare. Dette medfører stort potensiale for interaksjon mellom næringssøkende skarv og taretråling. Det er imidlertid tvetydig i hvilken grad denne interaksjonen påvirker fuglene i positiv eller negativ grad, eller om det påvirker dem i det hele tatt. Trolig avhenger effekten av taretråling av skalaen undersøkelsene gjøres på, omfanget av trålingen og mengden av alternative beiteområder rundt koloniene. Studiene identifiseres flere spørsmål for videre studier for å kartlegge mulige effekter av taretråling, deriblant behov for eksperimentelle feltstudier som av typen Kelpfish-studiet som ved å inkludere sjøfugl hadde kunnet identifisere finskala endringer i beiteadferd til sjøfugl målt mer direkte.
3.7 - Helhetsforståelse og betydning av skala
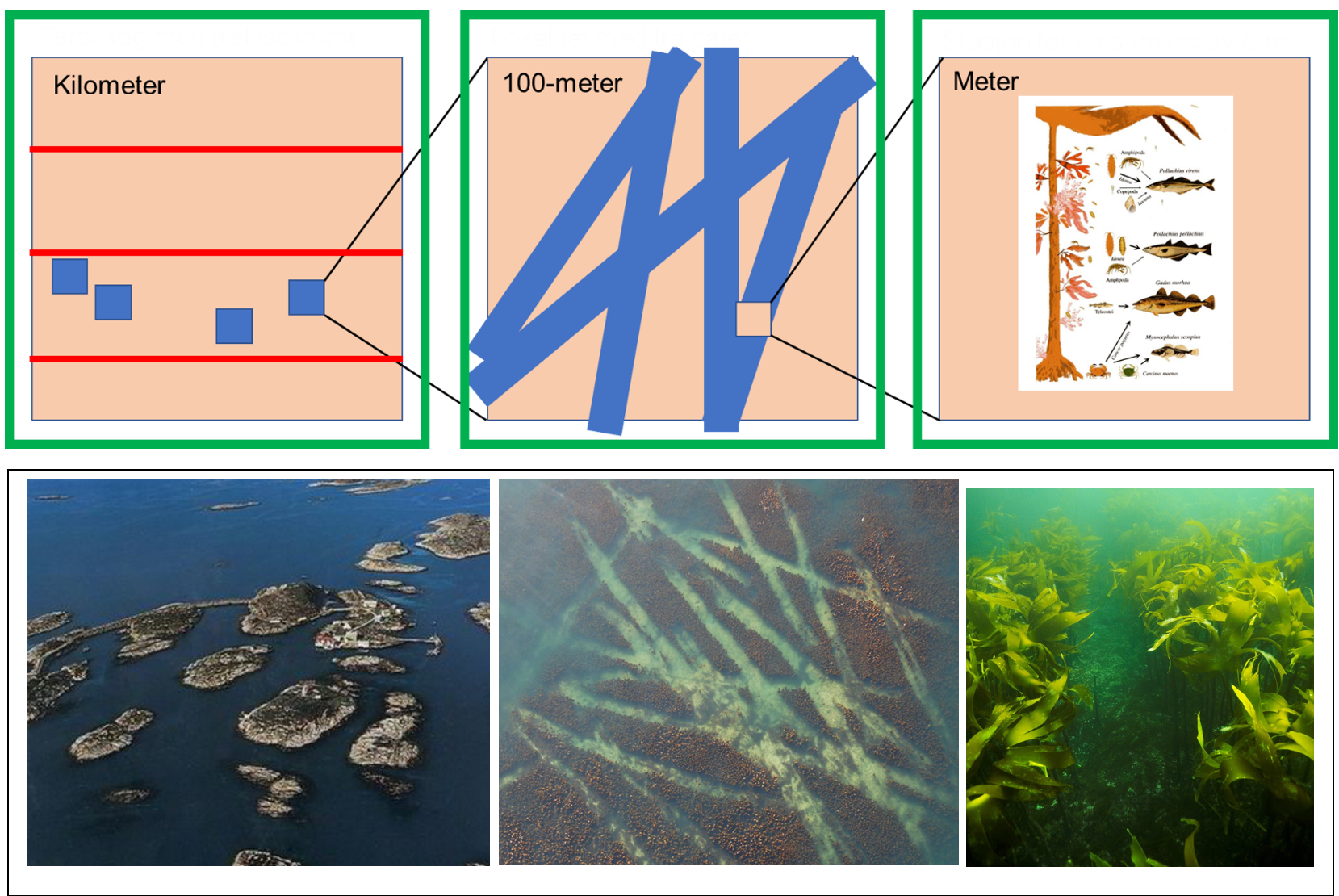
Resultatene fra alle typer undersøkelser om hvordan taretråling påvirker tareskog må forstås på riktig skala for å få en objektiv oppfatning av hva de betyr. Effekter betyr mindre for tareskogen i sin helhet hvis de skjer i en liten enn stor del av skogen. Store effekter inne i trålgatene vil for eksempel være mindre alvorlig hvis bare en liten del av skogen tråles i forhold til hvis en stor del tråles. Samtidig kan effekter, og spesielt små effekter, være vanskeligere å fange opp på stor enn liten skala. Det er for eksempel lettere å måle effekter nøyaktig inne i en trålgate (meter skala) som ble gjort i Kelpfish-prosjektet (kapittel 3.3), enn å måle effekter på dykkeadferd hos sjøfugl eller modellert uttak av et trålfelt (kilometerskala). Derfor er gode uttrykk for usikkerhet viktig og resultatene må tolkes på det nøyaktighetsnivået de kan si noe om.
I figur 16 er viktige skalaer illustrert. Hvert år tråles det i en sektor som har en lengde på et lengdeminutt (1 843 m) og bredde som skjærgården i det aktuelle området (1). Per i dag er det ingen begrensninger i hvor mye som kan tråles i hver sektor. Resultatene fra biomassemodell fra stortare viste at uttaket i høstefeltene i studieområdet på Møre var ca 6% gjennom en høstesyklus på 5 år. Usikkerheten knyttet til resultatet er imidlertid relativt stor med korrelasjon på 0,69.
Trålerne høster på steder innenfor sektoren der de erfaringsmessig finner mye tare (2). Trålen trekkes over bunnen på en 100 meters skala, men hvor lange trålstrekkene er og hvor mye som tråles avhenger av topografi på bunnen og andre faktorer som påvirker effektiviteten av bunntrålingen. I Kelpfish ble i gjennomsnitt 26% av tareskogen på stasjonene trålt. Hvis det meste av tareskogen sto igjen også på Sklinna kan det forklare at det ikke ble observert endringer i dykkeadferd hos skarv i studiet til Christensen-Dalsgaard et al. (2020) men stor usikkerhet kan også ha gjort det vanskelig å finne små effekter.
Inne i trålgatene (3 m brede) viste Kelpfish store effekter og at de fleste store «trekronedannende» tarene var borte og bare små rekrutter sto igjen (Norderhaug et al. 2020). Med de store tarene forsvant også mange av artene alger og smådyr som lever i tareskogen og de fleste fisk med tilknytning til tareskog, både juvenile og adult fisk. Effektene kunne på denne lille skalaen måles nøyaktig og med relativ liten usikkerhet.
Samlet sett viser resultatene fra KelpFish-prosjektet at taretråling har effekter på tareskogens funksjon som oppvekstområde for fisk, men at det meste av tareskogen står igjen med intakt økosystem. Biomassemodellen i Møre og Romsdal tyder videre på at taretråling bare berører en liten del av den stående biomasse tareskog. Dette kan imidlertid variere fra område til område og endre seg dersom høstetrykket i framtiden skulle øke (f.eks hvis flere aktører og/eller ny fangstteknologi kommer til) og/eller tarebestandene reduseres (f.eks som følge av kråkebollebeiting, klimaendringer, o.l.). Overvåkningen av taretråling viser at tarens biomasse restitueres relativt raskt, 3-4 år noe som er kortere enn høstesyklusen. Samtidig tar det lengre tid for det assossierte samfunnet av alger å dyr å restitueres, opp til 6 år (Steen et al. 2020). Bærekraftig taretråling ved Vikna må ta alle disse aspektene inn som del av grunnlaget.
4 - Hva er bærekraftig taretråling?
Taretråling er en av mange økosystemtjenester tareskogen tilbyr. Hva som er bærekraftig uttak avhenger av inngrepets omfang og taresamfunnets restitusjonsevne. I tillegg må denne økosystemtjenesten veies mot andre slike tjenester.
At høsting av de levende marine ressursene skjer på en bærekraftig måte er et sentralt fundament for Nærings- og fiskeridepartementet (Regjeringen, fisheries.no). En forutsetning for bærekraftig høsting er at de marine økosystemene fungerer. Norsk marin forvaltning er basert på vitenskapelige råd og både Fiskeridirektoratet og Havforskningsinstituttet har drevet fiskeriforvaltning og havforskning i over 100 år. Å bidra til sunne økosystemer er derfor et grunnleggende prinsipp for all aktivitet knyttet til fiske, fangst og havbruk.
Hva som er bærekraftig høsting av tareskog avhenger av:
-
Inngrepets omfang eller hvor stor del av tarevegetasjonen som høstes og hvor mye som står igjen. Her er uttak i forhold til ressursgrunnlaget i hvert enkelt høstefelt sentralt.
-
Evnen tareskogen, det vil si både taren og det til hørende økosystemet som yter en rekker goder, har til å restitueres tilbake til referansetilstand etter tråling. Her er høstingssyklusen for høstefeltene sentral.
Inngrepet taretrålingen representerer vil være stort hvis en stor del av biomassen høstes og hvis restitusjonen for taren og resten av økosystemet er langsom. Det vil være mindre hvis en liten del av biomassen høstes og det tar kort tid for tareskogen å restitueres. Det er et vanlig bærekraftkriterium å ikke høste mer enn hva plantene eller algene produserer, men tareskogens goder skal deles av mange, både smådyr, fisk, sjøfugl, sel, hval - og mennesker. Slike goder kalles økosystemtjenester (Faktaboks 2).
Det er derfor viktig for forvaltningen å ha et objektivt grunnlag for å vurdere hva taretråling betyr i reduksjon av andre økosystemtjenester, og vi har derfor i denne rapporten i størst mulig grad forsøkt å lage et kvantitativt grunnlag for forvaltning av taretråling. Kunnskapsgrunnlaget gir mulighet til dette, biomassemodellen for stortare er et verktøy som kan brukes til å kvantifisere ressursgrunnlaget og ved å legge inn overvåkningsdata kan modellen brukes til å vise restitusjon av både ressursene og økosystemet og hvordan denne varierer i ulike områder. Dermed er biomassemodellen et viktig verktøy for økosystembasert og adaptiv forvaltning av taretråling. Hvordan dette kan gjøres i praksis er demonstrert i kapitlene 5-7.
|
Tareskogens økosystemtjenester
Økosystemtjenester er et begrep som brukes for å kvantifisere og verdisette goder eller tjenester et økosystem yter. Man kan snakke om ulike typer slike tjenester og de viktigste økosystemtjenestene som tareskogen yter kan som for andre økosystemer deles i fire, forsynende, regulerende, støttende og kulturelle økosystemtjenester:
Forsynende økosystemtjenester er mat og andre ressurser som utnyttes direkte. Primærproduksjonen i en tareskog er stor Små alger vokser på stilkene til de gamle tarene. Store tarestilker er buskete av påvekstalger. Et vrimmel av smådyr bruker påvekstalger og tarehapterene (tarens festeorgan) som levested og på hver eneste kvadratmeter kan 100 000 smådyr fra over 100 arter finnes (Norderhaug et al. 2002). Tareskogen forsyner disse dyrene med næring men den store produksjonen gjør at bare 10-20% av tareskogens primærproduksjon untyttes i tareskogen (Norderhaug & Christie 2011). Det meste eksporteres ut av tareskogen og forsyner økosystemer i tangvoller og på dypt vann. Tareskogen er dermed viktig i å forsyne livet langs kysten med mat. 
Regulerende tjenester kontrollerer eller påvirker viktige biotiske eller abiotiske prosesser som eksempel klimaregulering. Tareskogen binder store mengder karbon og bidrar i klimaregulering (Norderhaug & Moy 2018). Mye av bladene og avreven tare fraktes bort med bølger og strøm ut på grunt eller dypt vann. I fjæra danner de tangvoller. En ukjent del havner også på dypet på rolig sedimentbunn. Her forsyner de evertebrater med næring og en del av tarematerialet begraves og forsvinner fra det raske karbonkretsløpet. På denne måten fungerer tareskogene som klimapumper som pumper CO2 fra atmosfæren og ned i havbunnen. Tareskogen beskytter kysten mot erosjon ved ekstremvær.
Støttende tjenester er de grunnleggende livsprosessene som fotosyntese. Taren selv og det rike biomangfoldet av påvekstalger på tarens stilk driver fotosyntese for å skaffe energi til primærproduksjonen. Taren skaper makrohabitat for fisk og oppvekstområder for yngel av kystfisk og påvekstalgene mikrohabitat for et myldrende liv av små evertebrater som snegl, krepsdyr og børstemarker.
Kulturelle tjenester er rollen naturen spiller for menneskers opplevelser. Tareskogene er populære dykkemål og attraktive for sportsfiskere, men de kan også være velegnede for opplærings- og utdanningsformål. |
5 - Modellert ressursgrunnlag ved Vikna
Basert på vekt av tare og miljødata ble biomassemodellen for stortare kjørt for Viknaområdet. Biomassen var størst på grunt vann vest i området der bølgeeksponeringen var høy. På grunn av lav kvalitet på tilgjengelige dybdedata overpredikerte modellen. Korrigerte prediksjoner tilsier at det totalt finnes nesten 175 000 tonn tare til sammen i alle trålfeltene og at mengden varierer fra felt til felt fra i overkant av 17 000 tonn til nesten 35 000 tonn tare .
5.1 - Feltarbeid, datagrunnlag og statistikk
En biomassemodell ble brukt for å beregne tareressursgrunnlaget på hostefeltene 414E-420A i Vikna. Datagrunnlaget inkluderte data om tarebiomasse samlet inn i felt. Feltarbeidet ble gjennomført i Vikna i juni 2020. Det ble gjort videoobservasjoner på 203 lokaliteter mellom 0,7 og 47 meters dyp (Fig. 17). På hver lokalitet ble det gjort anslag av andel hardbunn, samt tetthet og høyde av stortarevegetasjonen per kvadratmeter. Vegetasjonsmålingene ble senere omregnet til biomassetetthet av stortare per kvadratmeter ved å multiplisere den observerte plantetettheten med gjennomsnittlig vekt per plante. Stortareplantenes vekt ble beregnet utifra plantehøyden som observeres på videoene ved å benytte en relasjon (korrelasjonskoeffisient = 0,88) mellom plantehøyde og vekt utviklet gjennom målinger av innsamlede stortareplanter (Van Son et al. 2020). En verifisering av videometodens biomasseberegninger er tidligere gjort ved hjelp av dykkerinnsamlinger i et område på Sunnmøre. Dette ga en korrelasjon på 0,77 mellom den videoberegnede tarebiomassen og tarebiomassen innsamlet med dykkere, som anses som svært bra (Norderhaug et al. 2020).
Miljøvariabler
For å analysere sammenhengen mellom tarebiomasse og miljøforhold, ble følgende miljøvariabler innhentet fra studieområdet: dyp, bunnens kurvatur, bunnens skråning og bølgekraft på bunnen. Dybdedata som ble brukt var beste tilgjengelige 50 m oppløsning grid data fra Statens Kartverket. Bunnkurvatur og bunnskråning (50 m oppløsning) ble beregnet fra dybdedata, ved bruk av arbeidspakke raster i R software (R Core Team, 2018). Disse variablene kan brukes som erstatninger for data om hardhet og kornstørrelse i områder kart over hard- og bløtbunn mangler. Tare og andre bunnalger trenger hard bunn for å feste seg. Bølgeeksponering på bunnen (50 m oppløsning) ble beregnet fra SWAN bølgemodell med værdata fra Met.no (http://swanmodel.sourceforge.net) som beskrevet i Van Son et al. (2020). Pearson korrelasjon koeffisienter ble brukt til å undersøke kollinearitet mellom variablene. Alle koeffisienter var <0.6 og derfor kunne disse variabler brukes samtidig i samme modell.
Biomasse modellering
Generalisert Additive Modeller (GAM) ble brukt for å undersøke relasjonen mellom stortare biomasse og miljøvariablene. Observasjonene om tarebiomasse fra felt ble brukt som respons i modellene, mens dyp, bølgeeksponering på bunnen, krumming og skråning ble brukt som forklarende variabler. Siden tarebiomasse bare kan ha verdier ≥ 0 ble en miks Poisson-Gamma distribusjon brukt. Forklaringsvariabler ble modellert som en smooth function ƒ av type «penalized regression splines». Modellene ble kjørt i arbeidspakke mgcv i programmet R. Modelseleksjon ble utført basert på Akaike information criterion (AIC) og følgende modell ble selektert som den beste modellen:
Biomasse i = α + ƒ (Dybde j ) + ƒ ( Bølgeeksponering på bunn ) + ƒ ( bunnskråning ) + ε ij
I modellen er i stasjoner, α er modellinteresept , og ε ij er uavhengig normalfordelt støy. Modelvalidering ble gjort ved å inspisere residualenes fordeling, fordelign av residualene relativt til forklarende variabler, samt romlig korrelasjon i residualene (som anbefalt av Zuur og Leno (2016)). Ingen avvik fra modellforutsetninger ble oppdaget. Til slutt, ble modellen brukt til å predikere tarebiomasse i helle studieområdet, basert på miljøvariabelverdiene i de nye områdene.
5.2 - Ressursmodellens resultater basert på tilgjengelige dybdedata
Biomassen som ble målt på stasjonene er vist i figur 17. De varierer mellom 0 og 31,2 kg/m 2 avhengig av dyp, fysiske forhold og bunnforhold. Blant 203 undersøkt stasjoner var stortare observert i 38 % av disse lokalitetene.
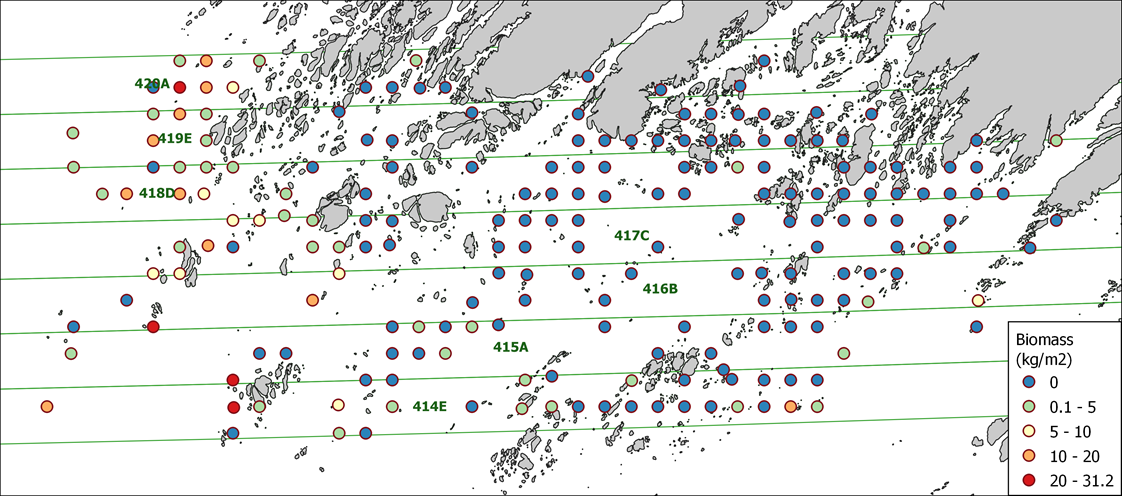
Resultatet viste at biomassen av stortare er betydelig påvirket av dybde og bølgeeksponering (Tabell 3, Figur 18). Biomassen var størst på grunt vann der lysforholdene er gode. Tareskog er tilpasset værharde forhold ytterst på kysten og bølgeeksponering hadde en positiv effekt i modellen, med høyest biomasse observert i områder med bunnbølgeeksponering over 1 m/s. Bølgene holder bladene friske og fri for påvekstorganismer som skygger for lyset og øker hastigheten for næringsopptak over tarebladenes overflate. Skråning har en mindre tydelig effekt på tarebiomasse, noe som kan gjenspeile at skråning bare delvis forklarer hvor bunnen er hard og myk. I tillegg viste tarebiomassen stor variasjon på de få observasjonslokalitetene vi hadde fra områder med bratt bunn.
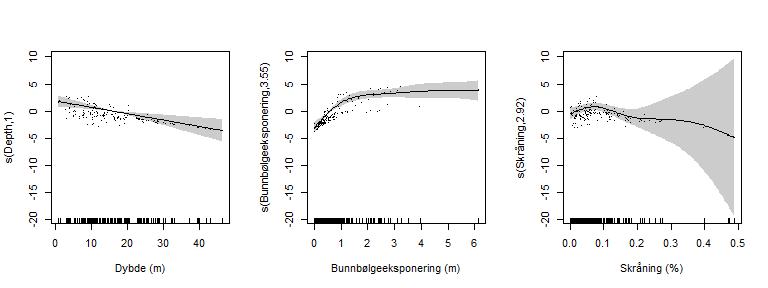
| EDF | F | P | |
| s(Dybde) | 1,0 | 12,2 | < 0,001 |
| s(Bunnbølgeeksponering) | 3,6 | 21,5 | < 0,001 |
| s(Skråning) | 2,9, | 2,3 | 0,051 |
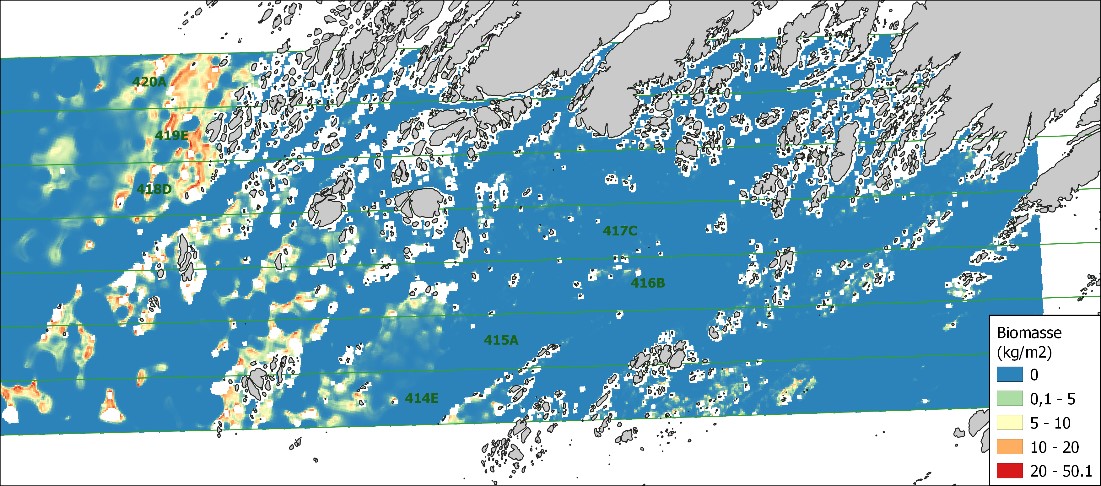
Den romlige fordelingen av biomasse for stortare slik den er predikert av modellen i høstfeltene 414E-420A og basert på miljøvariablene og målt biomasse er vist i figur 19. De størst forekomstene av stortare er predikert i de grunne eksponert områdene i vest, lengre inn i beskyttede områder er tarens biomasse liten.
5.3 - Validering av biomassemodellen
Datainnsamling til validering av biomassemodellen ble gjennomført i juni 2021. Randomisert utvelgelse av stasjoner ble gjennomført på nytt for å skaffe et helt uavhengig datasett for å teste hvor godt modellresultatet var. Dette er den strengeste formen for modellvalidering for å unngå systematiske feil som kan føre til at modellresultatet ser bedre ut enn det er (for eksempel ved bruk av valideringsmetoder som kryssvalidering eller bruk av data fra modelldatasettet til validering, van Son et al. 2019). Figur 20 viser målt og predikert biomasse for området.
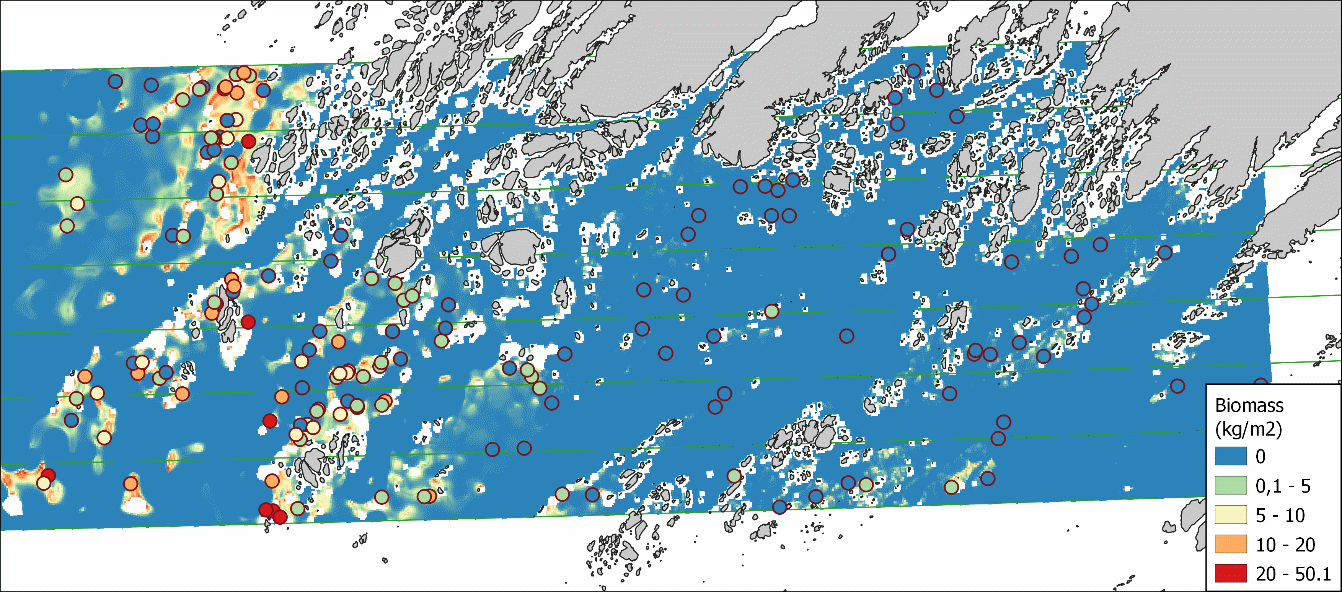
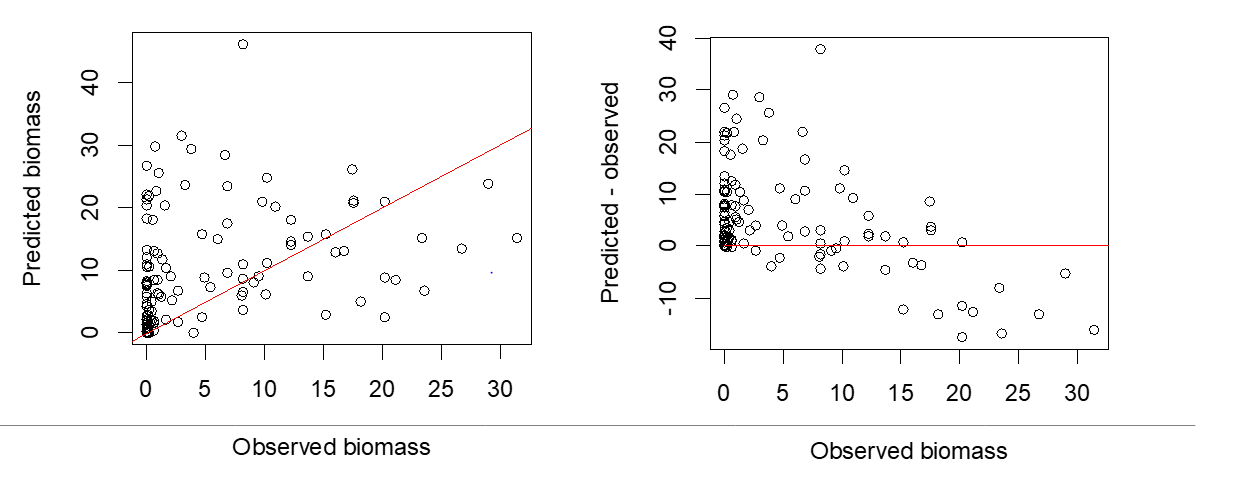
Valideringen viste relativt liten sammenheng mellom de uavhengige observasjonene av tarebiomsse og predikert biomasse fra modellen (Figur 21). korrelasjonen (Pearson correlation) for sammenhengen var 0,42. Dette betyr at det var ganske stor forskjell mellom biomassen for tare som modellen predikerte og hva som ble observert i felt når vi sjekket modellen. Modellen overestimerte mengden tare på steder med lite tare (<10 kg/m 2 ) og underestimerte mengden tare på steder med mye tare (>20 kg/m 2 ).
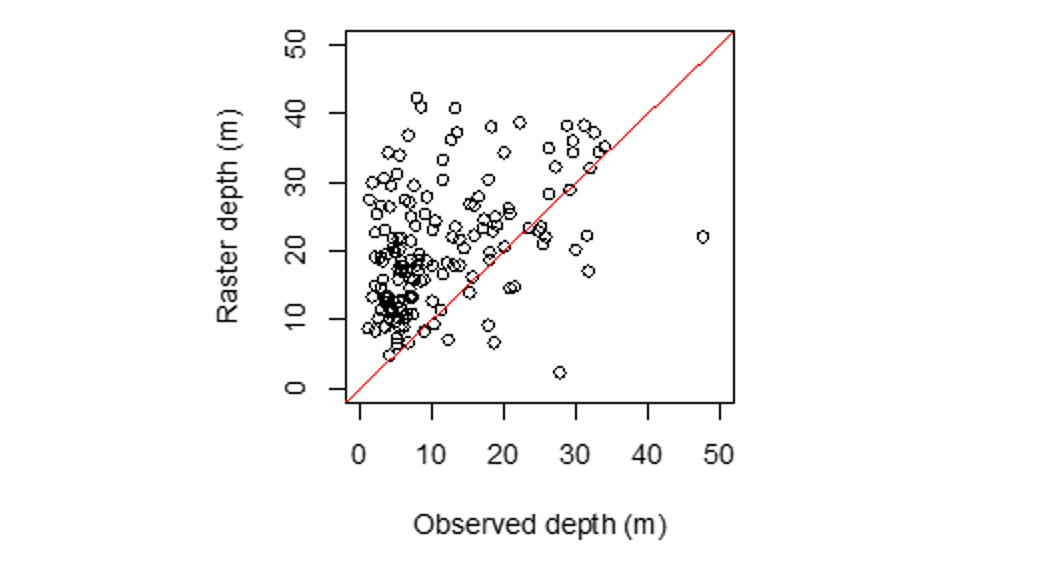
Det observerte dypet på valideringsstasjonene varierte også mye i forhold til det beste tilgengelige dybdegriddet fra Kartverket og som modellen var basert på (Figur 22). Korrelasjonen mellom observert dyp og dyp i dybdegriddet var på 0,31 (Pearson correlation). Vi ønsket derfor å teste biomassemodellen basert på det faktisk målte dypet og uten bruk av dybdegriddet.
5.4 - Modell basert på observerte dyp og andel hardbunn
For å identifisere årsaken til lav korrelasjon mellom biomassemodellens prediksjoner og valideringsdataene modellerte vi biomasse basert på observert dyp (siden det var lav korrelasjon mellom observert dyp i felt og dyp i dybdegriddet), bølgekraft på bunnen og observert andel hardbunn (istedenfor bunnens kurvatur og bunnens skråning) . Modellen viste at dyp, bølgekraft på bunnen og andel hardbunn forklarte hele 84.6% av variasjonen i datasettet (Figur 23, tabell 4). Responskurvene stemmer også godt overens med modellresultater fra et område på Sunnmøre der vi hadde høyoppløselige grunnlagsdata (van Son et al. 2019), noe som styrker resultatene.
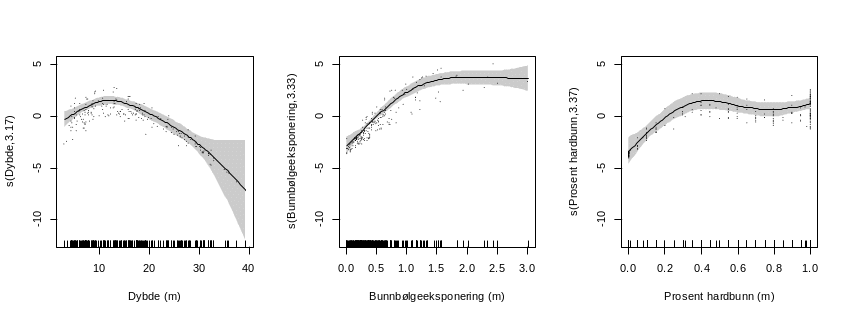
| EDF | F | P | |
| s(Dybde) | 3,17 | 18,52 | < 0,001 |
| s(Bunnbølgeeksponering) | 3,33 | 44,92 | < 0,001 |
| s(ProsentHardbunn) | 3,37 | 8,01 | < 0,001 |
Korrelasjonen mellom observert og predikert biomasse for denne modellen var på 85,6%, noe som er svært høy forklaringsgrad (Figur 24). Dette viser at biomassemodellen predikerer tarens biomasse svært godt når grunnlagsdataene er av god nok kvalitet.
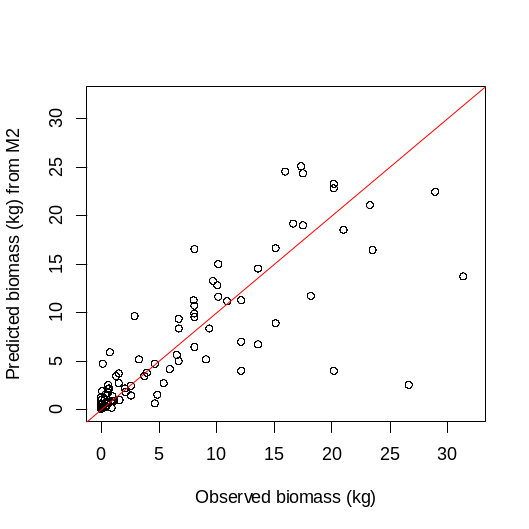
5.5 - Ressursgrunnlaget av tare ved Vikna
Den romlige biomassefordelingen per høstefelt ble beregnet fra modellen og er vist i tabell 4. Ressursmodellen predikerte mest tare i trålfelt 414E med over 67 000 tonn og minst tare i felt 417C der den predikerte biomassen var drøyt 34 000 tonn (tabell 5). Total predikert biomasse i alle trålfeltene fra felt 414E til 420A var ca. 341 000 tonn. Valideringen viste imidlertid at modellen overestimerte biomassen med en faktor på 1,96. Den totale observerte biomassen på alle valideringsstasjoner var 628 kg mens den predikerte biomassen fra taremodellen (basert på beste tilgjengelige dybdegrid) var hele 1 232 kg. Modellen som var basert på observerte dybdedata predikerte totalt 594 kg tare. For beregninger i kapittel 6 er derfor alle modellprediksjoner korrigert ned med en korreksjonsfaktor på 1,96 som konservative estimater for bærekraftvurderinger.
Tabell 5. Predikert biomasse av stortare per høstfelt i Vikna fra biomassemodellen og korrigert estimat (korreksjonsfaktor 1,96) basert på modellen for observerte dyp og som forklarer tarebiomassen svært godt (Pearson correlation 0,86).
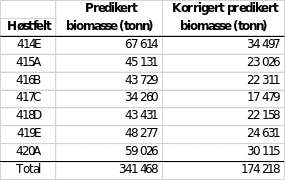
5.6 - Konklusjoner
Biomassemodellen viste at dyp og bølgeeksponering var viktigst for å forklare forekomst og biomasse av stortare ved Vikna. Valideringen viste at modellen overpredikerte tarens biomasse på grunn av lav kvalitet på tilgjengelige dybdedatasett og at i tillegg til de øvrige variablene også bunntype er viktig for å forklare tarebiomassens utbredelse. Med observert dyp og bunnsubstrat i modellen forklarte den nesten 85% av variasjonen i datasettet noe som er svært bra. Korrigert (konservative) verdier viste at det totalt finnes nesten 175 000 tonn tare i alle trålfeltene og at mengden varierer fra felt til felt fra i overkant av 17 000 tonn til nesten 35 000 tonn tare. Samlet sett viser denne studien betydningen av gode grunnlagsdata (spesielt dypbdedata av god kvalitet og god nok oppløsning). I tillegg viser studien viktigheten av uavhengig validering av modellresultatet for å sikre et godt modellresultat og gode estimater for hvor godt modellen stemmer med virkeligheten. Både gode prediksjoner og usikkerhetsestimater er nødvendig kunnskap for å vite hva modellresultatet kan brukes til og ikke, og dermed gi det beste tilgjengelige forvaltningsgrunnlaget.
6 - Bærekraftsvurderinger for Vikna
I dette kapittelet har vi brukt kunnskapsgrunnlaget fra kapittel 3, bærekraftkriterier fra kapittel 4 og ressursmodellen for Vikna presentert i kapittel 5 sammen med enkle regneeksempler for å vise hvordan biomassemodellen kan brukes som forvaltningsverktøy som gir forvaltningen mulighet til å veie ulike økosystemtjenester mot hverandre.
6.1 - Omfang av taretråling
En sammenligning av uttak per høstefelt i forhold til modellert biomasse per høstefelt (og korrigert for observert dyp, kapittel 5.5) er vist i tabell 6. I Viknaområdet er arealene på trålfeltene fra 47 til 73 km 2 . Uttaket varierer fra 0,5% i høstefelt 415A i 2012 til 31% i høstefelt 417C i 2014. Taretråling ble stengt i hele området etter 2015. I Kelpfish forskningsprosjektet ble imidlertid 2 986 tonn høstet i trålfelt 416B i 2018 i samarbeid med Dupont. Det beregnede uttaket av stående biomasse ved Vikna har vært større enn det som har blitt beregnet for Møre og Romsdal i perioden 2013-2017, her var beregnet uttak mellom 1 og 11% per trålfelt (Norderhaug et al. 2020). For hele området ble det trålt i gjennomsnitt 2% av stående biomasse per år for hele området i perioden 2010-2018 (der 2016 og 2017 ikke er tatt med siden det ikke ble trålt disse årene).

Tabell 6 . Omfang av taretråling i Vikna relativt til ressursgrunnlaget (predikert biomasse). Tabellen viser høstefelt, predikert biomasse fra ressursmodellen, hvor mye tare som ble høstet i de ulike feltene per år i tonn og hvor stort uttak dette representerte som % i forhold til modellens prediksjoner.

6.2 - Restitusjon av tarens biomasse og økosystemet
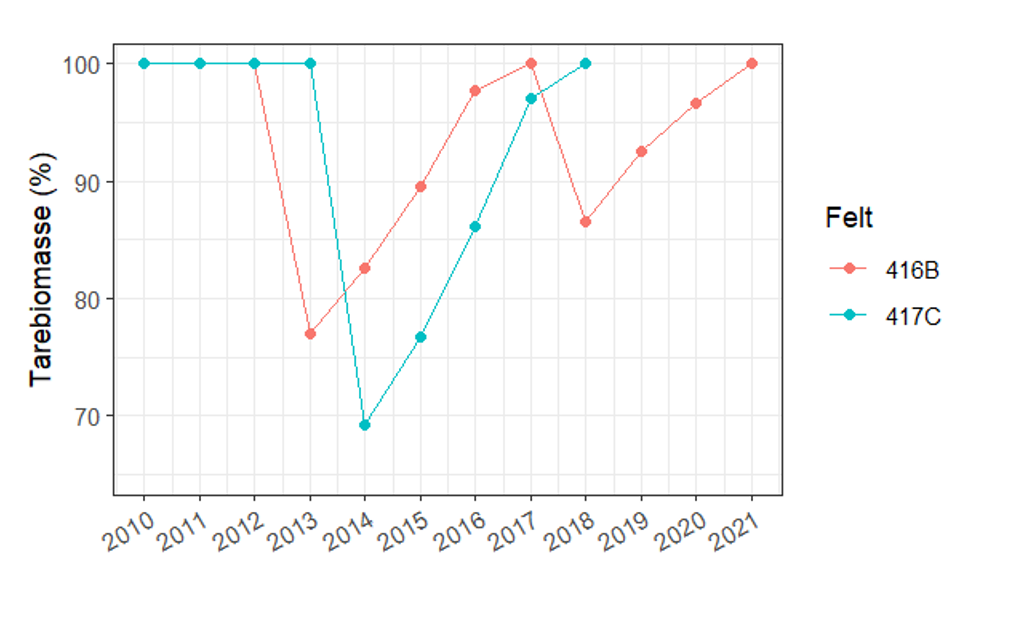
Gjenveksten av tare etter taretråling overvåkes gjennom Havforskningsinstituttets miljøovervåkning av taretråling (kapittel 3.4). Når de store tarene som danner «trekronene» blir høstet med trålen øker veksten til rekrutter som står under dette tresjiktet (figur 26, Steen et al. 2020).

I figur 27 presenteres utviklingen av tarens biomasse i tiden etter tråling med data fra overvåkingsstasjoner i Møre og Romsdal og Trøndelag. De viser at taren vokser raskt i tiden etter tråling og at biomassen er restituert etter cirka 4 år etter tråling.
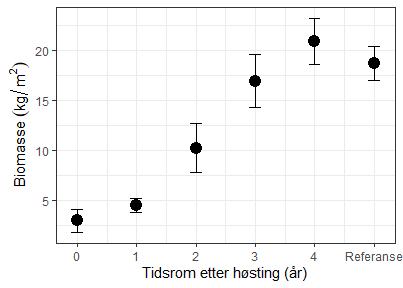
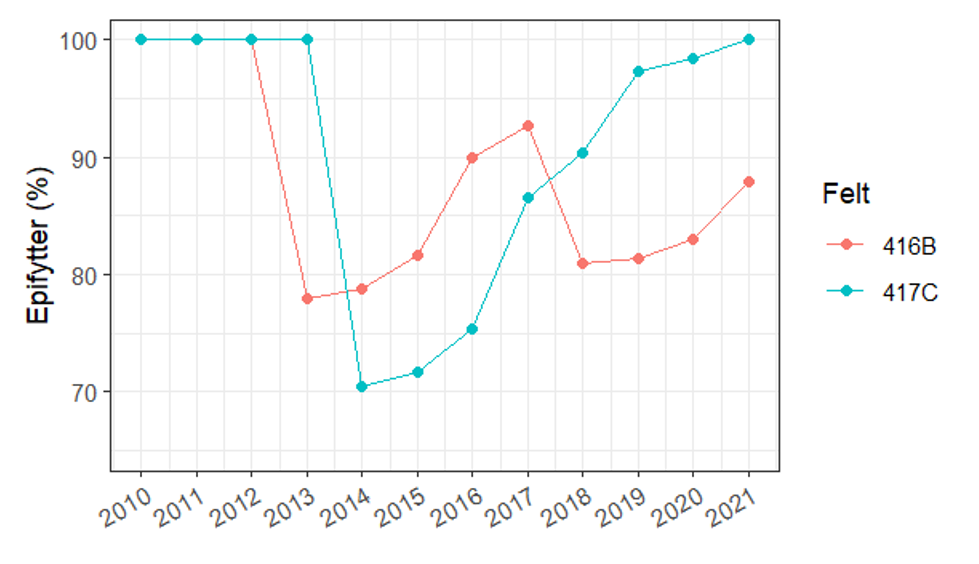
Restitusjon av påvekstalger på tarens stilk trenger lengre tid (figur 28). Taren må først vokse seg stor og få en barkaktig overflate små alger kan feste seg på før samfunnet av epifytter kan utvikle seg. I regneeksempelet under har vi brukt tilgjengelige data fra overvåkning i Møre og Romsdal, Trøndelag og Nordland til å illustrere restitusjonstid for påvekstalger. Vi har bare tilgjengelig data fra en stasjon for mer enn 4 år etter tråling og denne indikerer at det tar 6 år før mengden påvekst er lik mengden i referansefeltet som ikke ble trålt. Overvåkning ved Vikna vil kunne av-/bekrefte restitusjonstiden for påvekstalger. Eksempelet anses uansett som konservativt siden restitusjonstiden forventes å være lengre i Nordland enn ved Vikna (Steen et al. 2020).
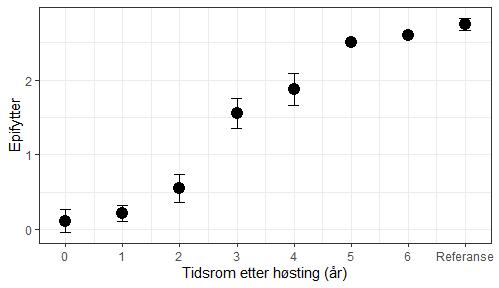
Mengden påvekstorganismer (eller epifytter) på tarestilkene er en god indikator for restitusjon av tareskogens økosystem av flere grunner. Epifyttene er en viktig del av tareskogens biomangfold med over 40 arter grønn-, rød- og brunalger (Marstein 1997) og fastsittende dyr (Christie et al. 2003, Norderhaug et al. 2020). Mengden påvekstorganismer øker med tarens alder og størrelse (figur 28 og 29). Påvekstalgene danner de viktigste mikrohabitatene for et mylder av smådyr som snegl, muslinger, krepdsdyr og børstemark. Volumet av påvekstalger er proposjonalt med antallet smådyr som lever blant dem (Norderhaug et al. 2007) og mengden påvekstalger er dermed en god indikator for mengden av smådyr i skogen. Siden disse dyrene er de viktigste fødedyrene for mye fisken som lever i tareskogen (Norderhaug et al. 2005), er epifyttene også en indikator for tareskogens verdi, eller økosystemfunksjonalitet, som føde- og oppvekstområde for kystfisk. Gjennom miljøovervåkningen av taretråling samler Havforskningsinstituttet inn data om forekomst av påvekstalger (epifytter) fra trålings- og referansefelter som ikke tråles.

Ved å kombinere data om ressursgrunnlaget og uttak i trålfeltene (tabell 6) med data om restitusjonshastighet fra overvåkningen (figur 27 og 28) er den prosentvise mengden av tarens biomasse og påvekstorganismer over tid beregnet for de to trålfeltene 416B og 417C og vist i figurene 30 og 31. Drøyt 5 000 tonn tare ble trålt i felt 416B i 2013 (tabell 6). Den (korrigerte) modellerte biomassen i dette feltet er i overkant av 22 000 tonn tare, slik at uttaket utgjorde 23% av biomassen. Med 4 års restitusjonstid var tarens biomasse restituert i 2017, året før feltet ble trålt på nytt, mens samfunnet av påvekstorganismer ikke ble fullt ut restituert før ny tråling. Nesten 5 400 tonn tare ble trålt i felt 417C i 2014, noe som utgjorde nesten en tredjedel av stående biomasse i feltet. De prosentvise effektene på biomasse og påvekstorganismer etter førstegangstrålingen var følgelig noe større på felt 417C enn på 416B.
6.3 - Økosystempåvirkning på ulikt uttaksnivå
Når bærekraftsindikatorene settes i system kan de brukes til å kvantitativt vise påvirkningen på tareressursene og økosystemet ved ulike grader av uttak. I tabell 7 vises hvor mye biomassen av tare og mengde påvekstorganismer som til enhver tid er redusert på ulikt høstenivå med høstefelt 416B som eksempel og med dagens 5-års høstesyklus. Hvis 1 000 tonn tare høstes i det aktuelle feltet hver gang vil tarebiomassen til enhver tid være 0-5% redusert i forhold til referansetilstanden.
Siden epifytter bruker lengre tid på å restitueres (6 år) enn høstesyklusen (5 år) blir den maksimale påvirkningen større enn ved hvert enkelt uttak. For eksempel, med et uttak på 1000 tonn tare i hver syklus er epifyttene i 2021 redusert 4,5% på grunn av tråling i 2021 pluss 0,4% fra gjenværende effekt fra forrige tråling i 2016, totalt redusert 4,9%. Ved større uttak på henholdsvis 3 000 og 10 000 tonn høstet tare ved hver høstesyklus blir påvirkningen tilsvarende større både på tarebiomassen og mengden epifytter. Den maksimale reduksjonen for tare blir for eksempel 45% og for påvekstalger 49% ved uttak av 10 000 tonn tare hvert 5. år for høstefeltet 416B.
| Uttak tonn | Biomassereduksjon tare % | Reduksjon epifytter % | ||||||||||
| 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | |
| 1 000 | 4,5 | 3,3 | 2 | 0,4 | 0 | 4,5 | 4,5 | 4,1 | 3,5 | 2 | 1,4 | 4,9 |
| 3 000 | 13 | 10 | 6,1 | 1,2 | 0 | 13 | 13 | 12 | 10 | 5,9 | 4,1 | 15 |
| 10 000 | 45 | 33 | 19 | 4,3 | 0 | 45 | 45 | 41 | 35 | 20 | 14 | 49 |
6.4 - Andre økosystemtjenester
Beregningene av effekter av taretråling gjort i kapittel 6.1 til 6.3 kan veies opp mot andre økosystemtjenester som kan måles kvantitativt. Skjebnen til produksjonen i tareskog er ikke godt kjent, men med noen forenklede regnestykker basert på kjent kunnskap kan vi likevel sette et uttak av 3000 tonn tare fra eksemplet i kapittel 6.3 i perspektiv. Vi har brukt tilgjengelige data fra ulike områder. Regnestykkene må sees på som eksempler men er antatt å være konservative fordi produksjonsdata er hentet fra Møre som antas å ha større produksjon enn ved Vikna, og restitusjonsdata blant annet er hentet fra Nordland der vi antar restitusjonstiden er lengre enn ved Vikna.
Uttaket av tare gjennom tråling reduserer både biomassen og habitatet og deres økologiske funksjon. Produksjonen til den viktige mobile evertebratfaunaen som er primære byttedyr for fisk utgjør 3-8% av primærproduksjonen til taren (Norderhaug & Christie 2011, Pedersen et al. 2012). 3 000 tonn tare kunne dermed produsert 90- 240 tonn evertebratfauna som teoretisk kunne gitt i størrelseorden 20-50 tonn fisk med 20% overføringsrate av biomasse fra et økosystemnivå til det neste. En stor del av den produserte taren konsumeres imidlertid ikke men fraktes ut av skogen i form av avrevne tarebiter av ulik størrelse (Wernberg & Filbee-Dexter 2018). All denne taren forsyner andre økosystemer i tangvoller og på dyp sedimentbunn eller begraves i sedimentbunn og lagres der i overskuelig fremtid. På denne måten pumpes karbon fra atmosfæren og ned i havbunnen. Karbon som tas ut fra det raske karbonkretsløpet på denne måten refereres gjerne til som blått karbon (Krause-Jensen & Duarte 2016). Hvis all den trålte taren hadde blitt lagret som blått karbon ville dette kunne binde snaut 1 100 tonn CO 2 (Gundersen et al. 2011).
Tarens vekt mål ved Vikna i Kelpfish-prosjektet var i gjennomsnitt 11,8 kg/m 2 (Norderhaug et al. 2020). 3 000 tonn høstet tare vil dermed påvirke 0,27 km 2 tareskog som habitat. Dette arealet vil gi et tap av mikrohabitat på i overkant av 70 tonn påvekstalger som igjen reduserer mengden svømmende evertebratfauna med i overkant av 10 tonn. Fisk ble påvirket på forskjellig måte på grunn av habitattapet avhengig av hvordan de brukte habitatet. Totalt ble det for eksempel ikke registrert mindre fisk etter tråling inne i trålgatene med bruk av akustikk gjennom hele vannsøylen men antall fisk/time registrert ved bunnen av stereovideo ble redusert med 46% i trålgatene (Norderhaug et al. 2020).
Tapene av primærproduksjon og habitat kan sammenlignes med andre økosystemer fra tareskogen. Hva sjøfugl spiser avhenger blant annet av næringsinnholdet i føden men i et studie er skarv estimert til å spise 430 g fisk per dag (Barett et al. 1990). Til sammenligning ble det i 2018 fisket totalt 2 145 tonn fisk derav 523 tonn torsk i statistisk fiskeriområde 12 (figur 32, Fiskeridirektoratet fangststatistikk). Tapt verdi for opplevelser og turisme er mer usikre og vi har ikke forsøkt å beregne verdi av dette.
7 - Miljøovervåkningsprogram for Vikna
Vi anbefaler å starte miljøovervåkning ved Vikna samtidig med kontrollert forskningstråling for å skaffe førdata, data om uttak og restitusjon av ressursgrunnlaget og økosystemet og dermed avkrefte eller bekrefte vurderingene vi har gjort i denne rapporten. Overvåkningsprogrammet kan med fordel bygge på Havforskningsinstituttets eksisterende tareovervåkning som samler data etter de samme prinsipper som denne rapporten anbefaler og som vil gi et godt sammenligningsgrunnlag med historiske data. I vurdering av kriterier for overvåkningsindikatorene har vi lagt vekt på at de:
-
Kan måles med kamera, sensorer. Dette gjør overvåkningen effektiv.
-
Dekker viktige økosystemtjenester, først og fremst produksjon, biomangfold og funksjon som føde- og oppvekstområde som er viktige økosystemtjenester.
-
Kan etableres som kartlag og modelleres i biomassemodellen for å si noe om effekter og restitusjon i rom. Dermed kan ny kunnskap om restitusjonshastighet etableres.
Under har vi listet kandidater for overvåkningsindikatorer:
-
Tarebiomasse: viser graden av uttak av biomasse og reduksjon i primærproduksjonen i tareskogen over tid. Er også indikator på mengden skogdannende makrohabitat for fisk.
-
Mengde ava påvekstorganismer (indeks 0-3, Steen et al. 2020): Indikator for reduksjon i biomangfold av alger men også av dyr gjennom sin funksjon som mikrohabitat for evertebrater som lever blant dem og som er de viktigste fødedyrene for tareskogens fisk.
-
Hapterstørrelse på taren: indikator for størrelse på tapt mikrohabitat ved bunnen og dermed indikerer mengde og biomangfold av dyr tilknyttet dette mikrohabitatet.
-
Alderssammensetning hos tareplanter som samles inn (vha skrape) på utvalgte stasjoner før og etter tråling og i høstefelt og referansefelt. Det innsamlede materiale kan i tillegg benyttes til detaljstudier av morfologi (f.eks størrelse av tareplantenes blad, stilk og hapter), påvekstorganismer og veksthistorikk (Steen et al. 2020, Steen 2021).
-
Antall, størrelse og arter av fisk. Fiskesamfunnet som viser mengden og biomangfoldet av tredje og fjerde nivå i næringskjeden. Forekomster av yngel viser tareskogens funksjon som oppvekstområde.
-
Antall og størrelse av taskekrabber som lever på bunnen. Ressurs og bunnlevende rovdyr.
-
Områdebruk til hekkende sjøfugl kartlagt vha. langtidsloggende GPS- og dykkeloggere montert på sjøfugl gjennom hele hekkesesongen, for å dokumentere adferd, før-, ved- og rett etter tråling.
Vi foreslår å begynne overvåkningen med indikatorene biomasse av tare og mengde påvekstalger som de viktigste indikatorene. Dette gir indikatorer for de to laveste nivåene i næringsnettet (tare, påveksalger, evertebrater) som danner basis for verdien tareskogen har som oppvekst- og fødeområde for fisk og fugl.
Tarens biomasse, størrelse og grad av påvekst kan estimeres med videokamera med samme metoder som benyttes i Havforskningsinstituttets ordinære overvåkningsprogram (Steen et al. 2020). Vi foreslår i tillegg å teste akustikk som overvåkningsteknologi for biomasse for stortare i tillegg til tradisjonelle metoder som brukes i dag. Akustikk kan bli en effektiv metode for overvåkning i forbindelse med taretråling i fremtiden. Innsamlinger av tareplanter for å studere utvikling av tarevegetasjonens alderssammensetning, morfologi, påvekstorganismer og veksthistorikk etter tråling vil også kunne bidra med forvaltningsrelevant kunnskap for å bl.a vurdere høstesyklus. Krabber og fisk kan overvåkes med krabbe- og leppefiskteiner, akustikk og stereovideokamera (RUV, Norderhaug et al. 2020).
Det anbefales at overvåkningen inkluderer sporingsdata fra taretrålerne og at mengde tare som tråles og posisjoner for hvor det tråles registreres. Slike sporingsdata kan gi mer nøyaktige data om hvor i høstefeltene uttaket skjer. Da kan industrien identifisere høsteområder som ikke utnyttes og forvaltningen analysere hvordan trålingen konsentreres og om det også er behov for mer detaljerte reguleringer innenfor høstefeltene.
8 - Råd om bærekraftig forvaltning av taretråling
Denne rapporten summerer forskningsresultater Havforskningsinstituttet a) har innhentet om økologiske effekter fra taretråling ved Vikna, b) presenterer resultater fra biomassemodellen for stortare som er kjørt for Vikna og c) viser med eksempler hvordan biomassemodellen sammen med overvåkningsdata kan brukes som verktøy for økosystembasert og adaptiv forvaltning av taretråling.
Havforskningsinstituttet vurderer ut fra kunnskapsgrunnlaget at området ved Vikna bør kunne åpnes for begrenset taretråling.
Vi anbefaler imidlertid at det før åpning av kommersiell tråling eventuelt vedtas, bare gjennomføres kontrollert forskningstråling sammen med miljøovervåkningen for å bekrefte eller avkrefte våre vurderinger og som grunnlag for å bestemme omfang av uttaket i hvert trålfelt.
Miljøovevåkningsprogrammet kan bygge på det pågående overvåkningsprogrammet for taretråling til Havforskningsinstituttet som allerede inneholder viktige indikatorer for økosystembasert forvaltning.
Av overvåkningsindikatorene vi har presentert her foreslår vi å starte med tarens biomasse og mengden påvekstorganismer (epifytter). I tillegg anbefaler vi å teste akustikk som effektiv overvåkningsmetode for tarebiomasse. Dersom overvåkning av fisk skal igangsettes anbefaler vi Fiskeridirektoratet om å vurdere om annen virksomhet med direkte påvirkning på fiskebestandene, som for eksempel fiske, bør stanses for å kunne måle en eventuell effekt av taretråling.
Vi anbefaler at metoden for bærekraftig taretråling som er beskrevet i denne rapporten, og inkludert biomassemodellen for stortare (van Son et al. 2020), også vurderes implementert i forvaltning av taretråling i øvrige deler Norge. Vikna er brukt som eksempel men verktøyene som er utviklet kan brukes i hele landet med tilpassede verdier. Dette er viktig siden både tarens størrelse, vekst og mengde og restitusjonshastighet for påvekstalger og fauna varierer med breddegrad (Kain 1967, Christie et al. 2003, Steen et al. 2020).
Vi anbefaler også at det vurderes å etablere høyoppløselige kartlag/datasett om dyp og bunnsubstrat for Vikna og andre prioriterte områder for å kunne beregne ressursene og påvirkning mer nøyaktig. Denne rapporten viser betydningen av gode grunnlagsdata for kartlegging og modellering som grunnlag for å øke kunnskapen om utbredelsen av tare som ressurs og kartlegging av marin natur generelt.

9 - Referanser
Barrett R, Nils RøvJ LoenWilliam A Montevecchi. 1990. Diets of Shags Phalacrocorax aristotelis and Cormorants P. carbo in Norway and possible implications for gadoid stock recruitment. 1990 Marine Ecology Progress Series 66(3):205-218 DOI: 10.3354/meps066205
Bjørge Arne Trine, Bekkby, Edmund Bryant. 2002. Summer home range and habitat selection of harbor seal ( Phoca vitulina ) pups. Marine Mammal Science 18(2):438 - 454
Bustnes JO, Anker-Nilssen T, Erikstad KE, Lorentsen SH, Systad GH (2013) Changes in the Norwegian breeding population of European shag correlate with forage fish and climate. Mar Ecol Prog Ser 489: 235 − 244
Christensen-Dalsgaard S, Mattisson J, Bekkby T, Gundersen H, May, R, Rinde E, Lorentsen S-H (2017) Habitat selection of foraging chick-rearing European shags in contrasting marine environments. Mar Biol 164:196
Christensen-Dalsgaard, S., Mattisson, J., Norderhaug, K.M., Lorentsen S-H. (2020) Sharing the neighbourhood: assessing the impact of kelp harvest on foraging behaviour of the European shag. Mar Biol 167, 136. https://doi.org/10.1007/s00227-020-03739-1
Christie, H., S. Fredriksen & E. Rinde (1998). "Regrowth of kelp and colonization of epiphyte and fauna community after kelp trawling at the coast of Norway." Hydrobiologia375/376: 49-58.
Christie H, Jørgensen NM, Norderhaug KM, Waage-Nielsen E. 2003. Species distribution and habitat exploitation of fauna associated with kelp (Laminaria hyperborea) along the Norwegian coast. J Mar Biol Ass UK 83:687-699
Dehnhard, N., Langset, M., Aglen, A., Lorentsen, S.-H., and Anker-Nilssen, T. Fish consumption by great cormorants in Norwegian coastal waters—a human-wildlife conflict for wrasses, but not gadids. – ICES Journal of Marine Science, doi:10.1093/icesjms/fsab004.
Duffy J. Emmett, Lisandro Benedetti-Cecchi, Joaquin A. Trinanes, Frank Edgar Muller-Karger, Rohani Ambo-Rappe, Christoffer Boström, Alejandro H. Buschmann, Jarrett Byrnes, Robert G Coles, Joel Creed, Leanne Cullen-Unsworth, Guillermo Diaz-Pulido, Carlos M. Duarte, Graham J. Edgar, Miguel Dino Fortes, Gustavo Jorge Goni, Chuanmin Hu, Xiaoping Huang, Catriona L. Hurd, Brenda Konar, Dorte Krause-Jensen, Kira Krumhansl, Peter Ian Macreadie, Helene Marsh, Len J. McKenzie, Nova Mieszkowska, Patricia Miloslavich, Enrique Montes, Masahiro Nakaoka, Kjell Magnus Norderhaug, Lina M. Nordlund, Robert J. Orth, Anchana Prathep, Nathan Freeman Putman, Jimena Samper-Villarreal, Ester A Serrao, Fred Short, Isabel Sousa PInto, Peter David Steinberg, Rick Stuart-Smith, Richard K.F. Unsworth, Mike van Keulen, Brigitta Ine Van Tussenbroek, Mengqiu Wang, Michelle Waycott, Lauren V. Weatherdon, Thomas Wernberg and Siti Maryam Yaakub. 2019. Toward a coordinated global observing system for marine macrophytes. Systematic Review, Front. Mar. Sci. 6:317. doi.org/10.3389/fmars.2019.00317
Fagerli CW, Norderhaug KM, Christie H, Pedersen MF, Fredriksen S. 2014. Predators of the destructive sea urchin grazer ( Strongylocentrotus droebachiensis ) on the Norwegian coast . Mar Ecol Prog Ser 502:207-218.
Fagerli CW, Norderhaug KM, Christie H. 2013. Can lack of sea urchin settlement explain kelp forest recovery in overgrazed areas in Norway? Mar Ecol Prog Ser. Vol. 488: 119–132 .
Freitas, C, Villegas ‐ R í os, D, Moland, E, Olsen, EM. Sea temperature effects on depth use and habitat selection in a marine fish community. J Anim Ecol. 2021; 00: 1– 14. https://doi.org/10.1111/1365 ‐ 2656.13497
Gundersen H, Christie H, de Wit H, Norderhaug KM, Bekkby T, Walday M. 2011. Utredning om CO2-opptak i marine naturtyper. NIVA report 6070. 21 pp.
Gundersen Hege, Eli Rinde, Trine Bekkby, Kasper Hancke, Janne K. Gitmark and Hartvig Christie. 2021. Variation in Population Structure and Standing Stocks of Kelp Along Multiple Environmental Gradients and Implications for Ecosystem Services. Front. Mar. Sci., https://doi.org/10.3389/fmars.2021.578629
Hillersøy, G. & Lorentsen, S-H. 2012. Annual Variation in the Diet of Breeding European Shag (Phalacrocorax Aristotelis) in Central Norway. Waterbirds 35: 420-429.
K Kain JM (1967) Populations of Laminaria hyperborea at vari- ous latitudes. Helgolander wiss Meeresunters 15:489-49
Krause-Jensen Dorte & Carlos M. Duarte. 2016. Substantial role of macroalgae in marine carbon sequestration. Nature Geoscience volume 9, pages 737–742.
Lorentsen, S-H., Sjøtun, K. & Grémillet, D. 2010. Multi-trophic consequences of kelp harvest. Biological Conservation 143: 2054-2062.
Marstein, A.C., 1997. Epiphytic algae on kelp stipes from Vega — an area with varying densities of sea urchins. Blyttia 3, 123–129.
Norderhaug K. M., K. Filbee-Dexter, C. Freitas, S.-R. Birkely, L. Christensen, I. Mellerud, J. Thormar, T. van Son, F. Moy, M. Vázquez Alonso, H. Steen. 2020. Ecosystem-level effects of large-scale disturbance in kelp forests. MEPS :ITRSav4. https://doi.org/10.3354/meps13426
Norderhaug KM, Christie H, Rinde E. 2002. Colonisation of kelp imitations by epiphyte and holdfast fauna; a study of mobility patterns. Mar Biol 141:965-973
Norderhaug KM, Christie H, Fosså JH, Fredriksen S. 2005. Fish-macrofauna interactions in a kelp (Laminaria hyperborea) forest. J Mar Biol Ass UK. 85:1279-1286.
Norderhaug KM, Christie H. 2011. Secondary production in a Laminaria hyperborea kelp forest and variation according to wave exposure. Est Coast Shelf Sci 95:135-144.
Norderhaug KM, Moy F. 2018. Blå skog – en glemt klimaaktør? Klima magasin, Cicero
Norderhaug KM, Nedreaas K, Huserbråten M, Moland E. 2020. Depletion of coastal predatory fish sub-stocks coincided with the largest sea urchin grazing event observed in the NE Atlantic. Ambio Perspectives in review. Ambio Perspectives doi.org/10.1007/s13280-020-01362-4
Norderhaug Kjell Magnus, Thijs Christiaan van Son, Nikolaos Nikolioudakis, Jonas Thormar, Frithjof Moy, Jan Atle Knutsen (HI), Sigrid Elvenes NGU og Henning Steen. 2020. Biomassemodell for stortare — Ressursmodell for fremtidens forvaltning. Havforskningsinstituttet rapport
Norderhaug KM, Christie H. 2011. Secondary production in a Laminaria hyperborea kelp forest and variation according to wave exposure. Est Coast Shelf Sci 95:135-144.
Norderhaug KM, Christie H, Fredriksen S. 2007. Is habitat size an important factor for faunal abundances on kelp (Laminaria hyperborea)? J Sea Res. 58:120-124.
Pedersen MF, Nejrup LB, Fredriksen S, Christie H, Norderhaug KM. 2012. Effects of wave exposure on population structure, demography, biomass and productivity in kelp, Laminaria hyperborea. Mar Ecol Prog Ser 451:45-60.
R_Core_Team (2018) R: A language and environment for statistical computing, Vol. R Foundation for Statistical Computing, ISBN 3-900051-07-0, URL http://www.R-project.org/, Vienna, Austria
Steen H, Moy FE, Bodvin T. 2014. Prøvehøsting av stortare i Nord-Trøndelag – Gjenvekstundersøkelser 2010-2014. Rapport fra Havforskningen Nr. 37-2014.
Steen H, Moy FE, Bodvin T, Husa V. 2016. Regrowth after kelp harvesting in Nord-Trøndelag, Norway. ICES Journal of Marine Science. 73(10): 2708–2720.
Steen H. 2019. Tilstandsvurdering av C-felt for tarehøsting i Rogaland og Sogn og Fjordane i 2019. Rapport fra Havforskningen Nr. 32-2019.
Steen H. 2020. Tilstandsvurdering av høstefelt for stortare i Møre og Romsdal og Trøndelag i 2020. Rapport fra havforskningen 2020-31.
Steen H. 2021. Tilstandsvurdering av høstefelt for stortare i Rogaland og Vestland i 2021. Rapport fra havforskningen 2021-34.
Steen H, Norderhaug KM, Moy F. 2020 Tareundersøkelser i Nordland i 2019. Rapport fra havforskningen 2020-9.
van Son TC, N Nikolioudakis, H Steen, J Albretsen, BR Furevik, S Elvenes, F Moy, KM Norderhaug. 2019. Achieving Reliable Estimates of the Spatial Distribution of Kelp Biomass. Frontiers in Marine Science 7, 107
Vondolia, Godwin K; Chen, Wenting; Armstrong, Claire W.; Norling, Magnus D; (2020) Bioeconomic Modelling of Coastal Cod and Kelp Forest Interactions: Co-benefits of Habitat Services, Fisheries and Carbon Sinks. Environmental and Resource Economics ISSN 0924-6460. Vol 75 No1 s25-48 doi: 10.1007/s10640-019-00387-y
Wernberg, T., Filbee-Dexter, K. Grazers extend blue carbon transfer by slowing sinking speeds of kelp detritus. Sci Rep 8, 17180 (2018). https://doi.org/10.1038/s41598-018-34721-z
Zuur, A.F. & Ieno, E.N. (2016) A protocol for conducting and presenting results of regression-type analyses. Methods in Ecology and Evolution, 7, 636-645.