Escaped farmed Atlantic salmon - risk assessment and knowledge status 2023
Further genetic introgression of farmed escapees in wild Norwegian salmon
Each year, the Institute of Marine Research (IMR), together with a collaborating partner, the Norwegian Institute of Nature Research (NINA), perform a risk assessment of further introgression of farmed escaped salmon in wild populations. The assessment uses data from a five-year window to inform a set of predicting criteria which in turn projects the probability of rivers in each of the 13 aquaculture production zones in Norway, to be subject to further introgression by farmed escapees. This report is used to assist management decisions regarding the potential impact of farmed escapees from aquaculture on wild populations.
Escaped farmed salmon pose a threat to the genetic integrity and evolutionary trajectory of wild salmon populations. Empirical studies show that there is a connection between the proportion of escaped farmed salmon in a river and the resulting genetic change caused by introgression, measured by genetic markers. This means that the more escaped farmed salmon that are present on the spawning grounds, the higher the probability of genetic change in those wild populations. In addition to the proportion of escaped farmed salmon on the spawning grounds, the robustness of wild salmon populations can affect the level of introgression. This is because large and productive wild salmon populations, without previous introgression of farmed salmon, are presumed to be more robust against introgression of escaped farmed salmon, since the escaped farmed salmon meets greater competition from a larger number of wild individuals that are adapted to those rivers. These factors form the basis for the risk assessment for further genetic change in wild salmon populations due to introgression of farmed escapees.
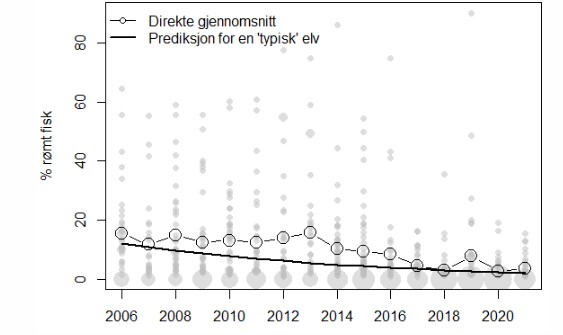
Three factors have been identified as the primary determinants of the probability of the proportion escaped farmed salmon observed on the spawning grounds in each production area: the reported number of escapees from aquaculture, the observed proportion of farmed escapees in the river and removal of escaped farmed salmon from the river. Of these three factors, it is the proportion of escaped farmed salmon in rivers that is weighted most heavily. This is reported annually in an average of 200 rivers through the national monitoring program for escaped farmed salmon in rivers. Although the proportion of escaped farmed salmon in Norwegian rivers have fluctuated between years, there has been a downward trend through the last few years. Official escapement figures are obtained directly from the Norwegian Directorate of Fisheries, and are fraught with uncertainty, while the effect of fishing is assessed based on data from OURO (the farming industry's association for the removal of escaped farmed salmon) and the Monitoring Programme. In rivers with a high concentration of escaped farmed salmon, fishing activities to remove farmed escapees must be carried out, but this is usually done with a delay of one year and may lead to watercourses with a lot of escaped farmed salmon not being covered by OURO.
The robustness of wild populations to “resist” further introgression from farmed escapees present on the spawning grounds is controlled in the risk assessment through the genetic status of the wild population (the degree to which they are already introgressed by farmed escapees), and population abundance (determined as whether each population achieves its set spawning target – i.e., indicating whether there is sufficient or an excess of wild mature salmon on the spawning grounds to fully utilize the river's production potential). Achievement of spawning targets and the level of harvest surplus are surveyed annually by the Norwegian Scientific Advisory Committee for Atlantic Salmon. Introgression of escaped farmed salmon has already led to extensive genetic change in many wild salmon populations, which can weaken the populations and can make them more vulnerable to new introgression. Description of genetic status is published in the report series "Genetic influence of escaped farmed salmon on wild salmon populations" and forms part of the genetic integrity standard of "Quality standard for wild salmon populations (Salmo salar)".
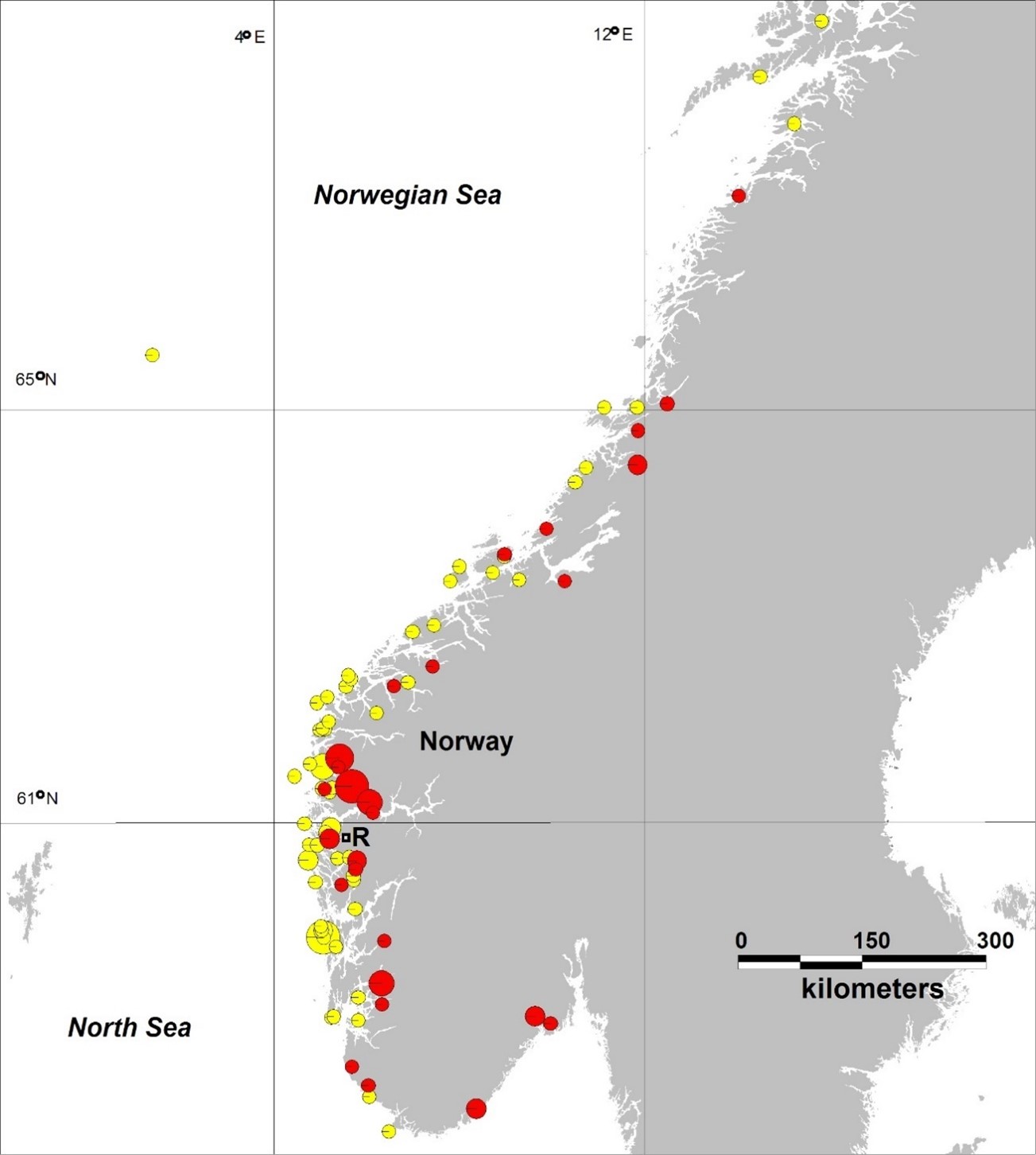
The risk assessment is performed on a regional basis, using Norway´s 13 aquaculture production zones (PZs). The risk assessment performed in 2023 shows that only the two most southerly (PZ1 and PZ2), as well as the most north-eastern aquaculture production zone (PZ13) have a low risk of further genetic change as a result of introgression of escaped farmed salmon. Four of the other production zones (PZ5, 6, 10 and 12) are assessed to have a moderate risk of further genetic change, while six production zones (PZ3, 4, 7, 8, 9 and 11) are assessed to have a high risk of further genetic change as a result of introgression of escaped farmed salmon. In PZ10, the risk level has been reduced from high to moderate, while for the other PZs the assessments are unchanged from the risk assessments performed in 2019, 2021 and 2022. It is therefore concluded that there is a moderate to high risk for further genetic change due to new introgression of escaped farmed salmon into wild populations in most parts of the country, even if the risk level has improved somewhat since the previous assessment. It remains important to note that there are multiple rivers within each production zone (~450 in Norway in total) and as a result there will be individual-river variation in degree of impact from spawning intrusion of domesticated farmed salmon.
This 2023 risk assessment is mainly based on data from 2017-2021, with the exception of genetic status (based upon data from a report published in 2020). The report consists of two parts; the first part which describes the risk assessment per production area, and the second part that describes the status of knowledge relevant for the risk assessment itself, including the consequences of the introgression of escaped farmed salmon into wild stocks.
Sammendrag
Rømming av oppdrettslaks fra oppdrettsanlegg utgjør en trussel mot den genetiske integriteten til de ville laksebestandene. Undersøkelser viser at det er en sammenheng mellom andel rømt oppdrettslaks i et vassdrag og genetisk endring målt som innkryssing med genetiske markører. Det betyr at jo flere rømte oppdrettslaks det er på gyteplassen, desto høyere er sannsynligheten for genetisk endring. I tillegg til andel rømt oppdrettslaks på gyteplassen, kan villaksbestandenes robusthet påvirke nivået av innkryssing. Dette fordi tallrike villaksbestander uten tidligere genetisk innkryssing antakelig er mer robuste overfor rømt oppdrettslaks, siden den rømte oppdrettslaksen der møter større konkurranse fra bedre tilpassete, ville individer. Disse faktorene danner grunnlaget for risikovurderingen for ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks.
Det er i hovedsak tre faktorer som bestemmer sannsynlighet for hvor mye rømt oppdrettslaks det er på gyteplassene i hvert enkelt produksjonsområde: rømming, andel rømt oppdrettslaks i elv og utfisking/fjerning av rømt oppdrettslaks fra elv. Av disse tre faktorene er det andel rømt oppdrettslaks i elv som vektes tyngst. Dette rapporteres årlig i gjennomsnittlig 200 elver gjennom det nasjonale overvåkningsprogrammet for rømt oppdrettslaks i vassdrag. Andelen rømt oppdrettslaks i elvene har endret seg mellom år, og det har vært en synkende tendens i registreringene gjennom de siste årene. Offisielle rømmingstall hentes direkte fra Fiskeridirektoratet, og er heftet med usikkerhet, mens effekten av utfisking vurderes basert på data fra OURO (oppdrettsnæringens sammenslutning for utfisking av rømt oppdrettslaks) og Overvåkningsprogrammet. I elver med høyt innslag av rømt oppdrettslaks skal det gjennomføres utfisking, men dette gjøres som oftest med ett års forsinkelse og kan føre til at vassdrag med mye rømt oppdrettslaks ikke dekkes av OURO.
Hvor robuste bestandene er for innkryssing av rømt oppdrettslaks, påvirkes av genetisk status hos de ville bestandene og villaksens bestandsstatus. Dersom bestandene når sine gytebestandsmål, har de mange nok ville gytelaks på gyteplassene til å kunne utnytte elvens produksjonspotensial. Konkurransen på gyteplassen blir enda sterkere dersom villaksbestanden er høyere enn gytebestandsmålet. Oppnåelse av gytebestandsmål og nivå av høstbart overskudd kartlegges årlig av Vitenskapelig råd for lakseforvaltning (VRL). Innkryssing av rømt oppdrettslaks har allerede ført til omfattende genetisk endring hos mange villaksbestander, noe som kan svekke bestandene og gjøre dem mer sårbar mot ny innkryssing. Beskrivelse av genetisk status publiseres i rapportserien «Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander» og utgjør delnorm genetisk integritet til «Kvalitetsnorm for ville bestander av laks (Salmo salar)».
Risikovurderingen viser at kun de to sørligste (PO1 og PO2), samt det nordøstligste produksjonsområdet (PO13) fortsatt vurderes til å ha lav risiko for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks. Fire av de andre produksjonsområdene (PO5, 6, 10 og 12) vurderes til å ha moderat risiko for ytterligere genetisk endring, mens seks produksjonsområder (PO3, 4, 7, 8, 9 og 11) vurderes til å ha høy risiko for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks. I PO10 er risikobildet justert ned fra høy til moderat risiko, mens for de andre PO-ene er vurderingene uendret fra de tilsvarende risikovurderingene gjort i 2019, 2021 og 2022. Det konkluderes derfor med at det også i de kommende år vil være moderat til høy risiko for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks i ville bestander i store deler av landet, selv om risikobildet er noe forbedret siden forrige vurdering.
Denne risikovurderingen er bygger i hovedsak på data fra 2017-2021. Genetisk status er basert på data publisert i «Genetisk påvirkning av rømt oppdrettslaks på ville bestander – oppdatert status 2020». Risikorapporten består av to hoveddeler; første del som beskriver Risikovurderingen per produksjonsområdet og Kunnskapsstatusen som beskriver bakgrunnskunnskapen for selve risikovurderingen og konsekvenser av innkryssing av rømt oppdrettslaks i ville bestander.
1. Innledning
1.1. Problemstilling
Laksen i norske oppdrettsanlegg stammer opprinnelig fra en rekke ville laksebestander. Tidlig på 1970-tallet ble det etablert flere avlslinjer for å forbedre produksjonsegenskaper som vekst og kjønnsmodning hos oppdrettslaks. 50 år senere har norsk oppdrettslaks gjennomgått omtrent 15 generasjoner med målrettet avl, og anses nå å være delvis domestisert og tilpasset et liv i fangenskap.
Rømt oppdrettslaks er en av hovedutfordringene for en miljømessig bærekraftig oppdrettsnæring. Hvert år rømmer det titusenvis av laks fra norske oppdrettsanlegg, og i enkelte år har antall rapporterte rømt oppdrettslaks vært høyere enn antall voksen villaks som returnerer til elvene for å gyte. De fleste oppdrettslaks som rømmer, forsvinner i havet. Det er sannsynlig at de dør av sult, sykdom eller blir spist av predatorer. Likevel overlever en liten andel etter rømming, og flere tusen vandrer opp i elvene hvert år. Innkryssing av rømt laks svekker den genetiske integriteten til norsk villaks. I 2021 ble villaksen ført opp på rødlisten under kategorien nær truet og mange bestander er alt redusert. Både forvaltning og næring har en nullvisjon for rømt oppdrettslaks og for ytterligere genetisk endring i de ville bestandene som følge av innkryssing av rømt oppdrettslaks.
Rømt oppdrettslaks har en dårligere gytesuksess enn vill laks, særlig hannfisken, men noen klarer å gyte med andre oppdrettslaks eller med villaks. Når oppdrettslaks gyter med villaks, fører dette til genetiske endringer i de ville laksebestandene. I Norge er det dokumentert og/eller indikert innkryssing i rundt to tredjedeler av 227 undersøkte villaksbestander, og i nærmere 30 % av de undersøkte bestandene er innkryssing av oppdrettslaks vist å være over 10 %. Forskning viser at avkommet til oppdrettslaks, og kryssinger med villaks, har en lavere overlevelse i naturen enn avkom fra villaks. Endringer i livshistorie hos både ungfisk og voksenfisk er også dokumentert i villaksbestander som følge av innkryssing av rømt oppdrettslaks. Innkryssing av rømt oppdrettslaks forandrer egenskapene til de ville laksebestandene, svekker bestandenes evne til å tilpasse seg endringer i miljøet og kan føre til en reduksjon i antall villaks som produseres. Innkryssing av rømt oppdrettslaks vil derfor både kunne svekke bestandene, og i tillegg gjøre dem mindre motstandsdyktige mot framtidig innkryssing av rømt oppdrettslaks.
1.2. Mål og omfang
Målsettingen med denne risikovurderingen er å vurdere risiko for ytterligere genetisk endring av rømt oppdrettslaks i villaksbestander som følge av at rømt oppdrettslaks gyter i elvene.
«Ytterligere genetisk endring» er her definert som videre genetisk endring som følge av innkryssing av rømt oppdrettslaks, framover i tid. Risikovurderingen vil dermed ikke vurdere risiko for at innkryssing som allerede har skjedd, vedvarer gjennom avkom av oppdrettslaks klekket i naturen (oppdrett-vill hybrider eller tilbakekrysninger). Status for genetisk innkryssing blir vurdert gjennom Kvalitetsnormen for villaks. Normen er retningsgivende for vurdering av behov for tiltak mot den samlete effekten av både tidligere genetisk innkryssing og ytterligere innkryssing som følge av rømt oppdrettslaks.
I tidligere utgaver av risikovurderingen, utgitt i perioden 2011-2018, har vi brukt andel rømt oppdrettslaks i vassdragene for å vurdere om det var lav, moderat eller høy sannsynlighet for ytterligere genetisk endring vurdert for ett år om gangen. Denne tilnærmingen var basert på data fra det nasjonale overvåkningsprogrammet for rømt oppdrettslaks og undersøkelser som viser at det er en sammenheng mellom andel rømt oppdrettslaks i et vassdrag og genetisk endring målt som innkryssing med genetiske markører. Dette betyr at jo flere rømte oppdrettslaks det er i et vassdrag, desto høyere er sannsynligheten for genetisk endring grunnet innkryssing.
Den rømte oppdrettslaksens biologiske status ved rømming påvirker evnen til å overleve i naturen, vandre opp i vassdragene og gyte sammen med villaks. Dette omfatter for eksempel livsfase, kjønn, kjønnsmodning, tidspunkt for rømming, lysregime på anlegget før rømming, sykdomsstatus, størrelse, alder og tid i det fri. Siden det finnes lite kunnskap om hvordan oppdrettslaksens biologiske status ved rømming varierer mellom produksjonsområder, har vi per nå ikke inkludert disse faktorene i den nåværende risikovurderingen.
Fra 2019 har vi i risikovurderingen i tillegg til andel rømt oppdrettslaks i vassdragene, i større grad tatt hensyn til villaksbestandenes robusthet, som et estimat for motstandsdyktighet for ny innkryssing. Som indikator for robusthet benyttes kunnskap om genetisk status med tanke på tidligere innkryssing og oppnåelse av gytebestandsmål (mengde hunnlaks som trengs for å utnytte elvens produksjonspotensial) og høstingspotensial til bestandene. Dette begrunnes med at tallrike villaksbestander uten tidligere genetisk innkryssing antakelig er mer robuste overfor rømt oppdrettslaks, som møter stor konkurranse fra bedre tilpassete, ville individer. Motsatt, i svekkede villaksbestander som er tallmessig svake og allerede er genetisk innkrysset med rømt oppdrettslaks, vil rømt oppdrettslaks ha mindre konkurranse fra villaks og dermed større sjanse til å få avkom.
I denne rapporten vurderes alle villaksbestandene innen et produksjonsområde samlet, istedenfor å vurdere risikoen for de enkelte bestandene. En del av risikofaktorene er knyttet til fysisk karakteristikk av det enkelte vassdrag og biologisk karakteristikk av den enkelte bestand. Fordi det innenfor hvert produksjonsområde vil være vassdrag og laksebestander som er ulike med hensyn til disse faktorene, vil aggregering av bestandene innenfor et produksjonsområde ikke nødvendigvis gi et godt uttrykk for tilstand og risiko for enkeltbestander. Der det er behov, har vi trukket fram tilstanden i enkeltvassdrag for å gi et mer nyansert risikobilde i det aktuelle produksjonsområdet. Vurderingen er i hovedsak gjort basert på tilgjengelige data fra perioden 2017-2021.
En utdypning av de ulike risikopåvirkende faktorene inkludert data og faglige referanser finnes i kunnskapsstatus i slutten av rapporten.
2. Metodikk for risikovurdering
I denne risikovurderingen kartlegges og analyseres usikkerhet knyttet til ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks. Risikoanalysen inngår som en del av forvaltningens beslutningsunderlag, og skal bidra til å sikre en bærekraftig utvikling av norsk fiskeoppdrett i tråd med norske og internasjonale bærekraftsmål.
Metodikken som ligger til grunn for risikoresultatene presentert i denne rapporten er utviklet av Havforskningsinstituttet og publisert i Andersen mfl. (2022). I dette kapittelet oppsummeres hovedpunktene i metodikken med fokus på å forstå resultatene av risikovurderingen, hvordan de er fremkommet og hva de bygger på. For detaljer rundt fundamentale forhold og hvordan risikovurderingen gjennomføres henviser vi til artikkelen.
I takt med forskningsfronten innen risikofaget finnes ingen objektive og korrekte risikotall som utgjør en form for fasit, det finnes kun usikkerhet knyttet til hva som ligger foran oss. Og det er nettopp denne usikkerheten fagekspertene sier noe om i denne rapporten. Målet med risikovurderingen er å skape risikoforståelse og risikoerkjennelse hos forvaltningen og andre interessenter som utgangspunkt for prioriteringer og beslutninger om veivalg og tiltak. Det er samtidig avgjørende at beslutningstakere og andre som skal benytte resultatene fra risikovurderingen, forstår at ufullstendig informasjon, utilstrekkelig kunnskap, hypoteser og antakelser er en del av, og i stor grad karakteriserer en slik analyse. Vi har valgt å visualisere resultatet av risikovurderingene i form av grafiske hierarkiske strukturer (risikokart) som er ment å gi en hurtig og intuitiv forståelse for risikokilder, hendelser og mulige konsekvenser med tilhørende usikkerhet knyttet til næringens aktivitet. Det er imidlertid argumentasjonen som ligger til grunn for risikokartene som skal skape tillit og eventuelt overbevise leseren om at risikovurderingen gir mening.
Fundamentale forhold knyttet til risikoanalysefaget som terminologi, definisjoner og metodisk tilnærming forankres i; Society for Risk Analysis, glossary (SRA, 2018); "Risk, Surprises and Black Swans – Fundamental Ideas and Concepts in Risk Assessment and Risk Management» (Aven,2014) og NS-ISO 31000:2018 – Risk management guidelines.
Risiko defineres i tråd med (SRA, 2018) som følger:
Med risiko menes konsekvenser (C) av aktiviteten med tilhørende usikkerhet (U)
Risikobegrepet introduserer dermed «konsekvenser» og «usikkerhet» - (C, U) som to sentrale aspekter som henger sammen. Med «usikkerhet» menes her en kunnskapsbasert usikkerhet (epistemisk) som knyttes til det å ikke vite fremtidige konsekvenser. Risikovurderingene i denne rapporten måler og beskriver usikkerhet ved hjelp av sannsynligheter og kunnskapsstyrke knyttet til ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks. Vi måler og beskriver risiko ved (C’, Q, K) der C’ er spesifikke konsekvenser av aktiviteten, Q er et verktøy for å måle usikkerhet, og K er bakgrunnskunnskapen som vurderinger av C’ og Q bygger på. Vi velger å splitte opp de spesifikke konsekvensene C’, i risikokilder RS’, og hendelser A’. Risiko beskrives da ved de fem variablene (RS’, A’, C’, Q, K). Vi måler usikkerheten Q ved hjelp av subjektive sannsynligheter P og vurderinger av kunnskapsstyrke SoK (Strength of Knowledge). Betraktninger om P og SoK hviler på bakgrunnskunnskapen K, og vi skriver Q (P, SoK, K).
For denne aktuelle problemstillingen relateres usikkerheten eksempelvis til:
Hvor mye oppdrettslaks rømmer årlig?
Hvordan sprer den rømte oppdrettslaksen seg etter at den har rømt fra et anlegg?
Hvor stor gytesuksess har den rømte oppdrettslaksen på gyteplassen?
Hvordan påvirker tidligere innkryssing av rømt oppdrettslaks de ville bestandenes robusthet mot ny innkryssing?
Usikkerheten knyttes til hva som kan skje, hvor ofte, samt til omfang og alvorlighetsgrad av konsekvensene. Vi kan måle denne usikkerheten ved hjelp av sannsynlighetsbetraktninger og forventningsverdier forankret i historiske data. Imidlertid vil en tilnærming der vi utelukkende bruker sannsynligheter som mål på usikkerhet, kunne føre til uheldige forenklinger og tap av viktige nyanser. I denne risikovurderingen søker forskerne å legge all tilgjengelig kunnskap til grunn, samt karakterisere denne for å øke innsikt og forståelse. Søkelys på usikkerhet, herunder manglende kunnskap knyttet til mulige kausale sammenhenger og konsekvenser fremover i tid, bidrar til effektiv utnyttelse av all tilgjengelig innsikt og stimulerer til videre utvikling av kunnskapsgrunnlaget.
Styrken til bakgrunnskunnskapen (i forhold til RS’, A’, og C’) vurderes eksplisitt ved SoK, det vil si en vurdering av hvorvidt kunnskapen som danner grunnlaget for en bestemt konklusjon, er sterk eller svak. Resultater som bygger på svak kunnskap, kan skjule deler av risikobildet gjennom eksempelvis feilaktige antakelser. Ved å inkludere en redegjørelse for kunnskapsstyrken i analysen, gis beslutningstakere mulighet til å ta stilling til hvorvidt kunnskapen på et område er tilstrekkelig som fundament for strategiske og operasjonelle veivalg. I motsatt fall vil det være naturlig å vurdere en føre-var strategi og diskutere tiltak som bidrar til å styrke kunnskapsgrunnlaget.
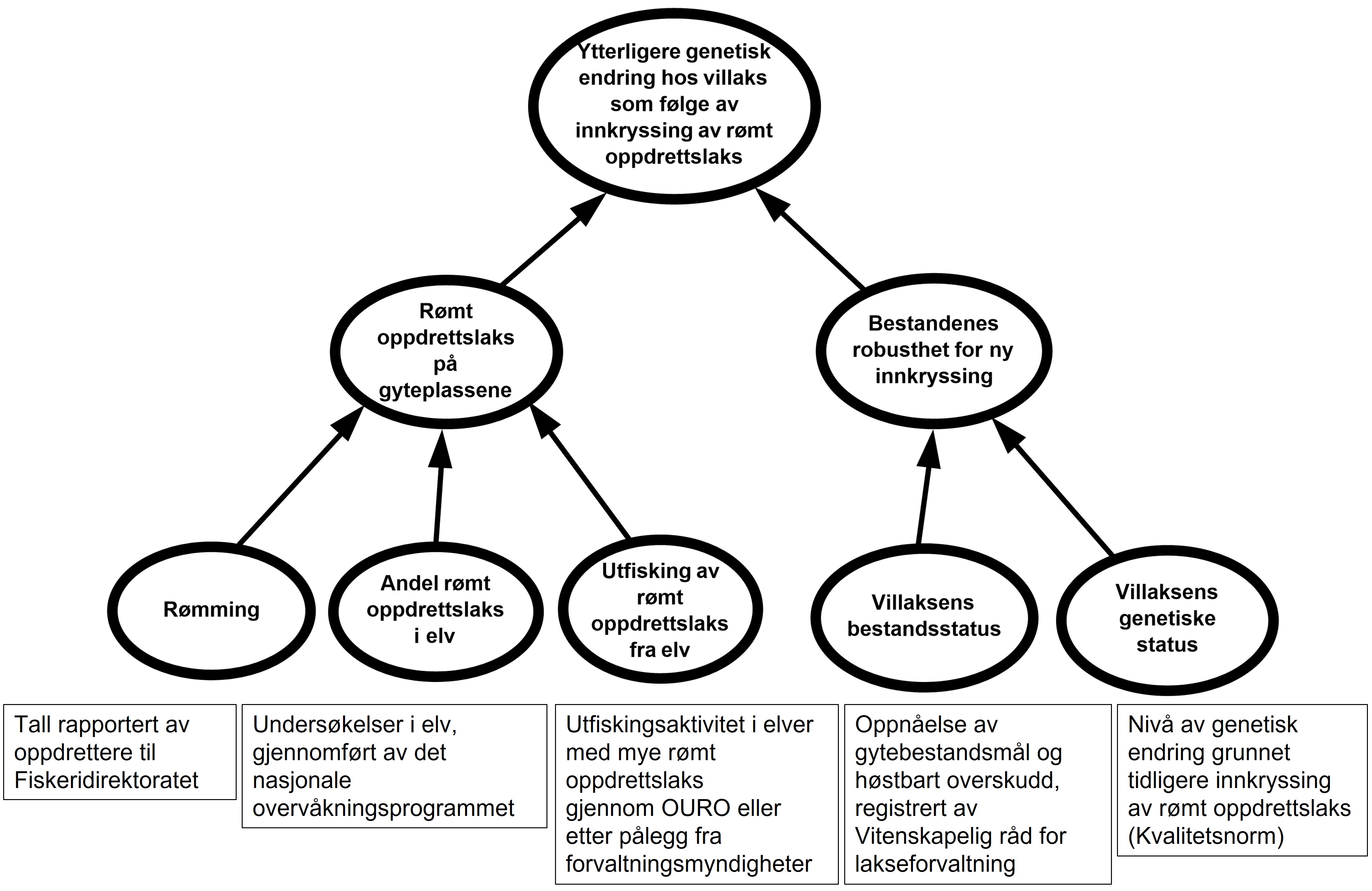
I arbeidet med denne rapporten har vi systematisert og strukturert risikofaktorene (RS’, A’, C’) knyttet til ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks. Det er lagt vekt på å inkludere all tilgjengelig kunnskap som utgangspunkt for å uttrykke og formidle usikkerhet (figur 2.1). Kunnskapsgrunnlaget varierer mellom de ulike risikofaktorene, avhengig av om det er gjennomført mye vitenskapelig forskning og om det eksisterer gode rapporteringssystemer og/eller overvåkningsprogrammer.
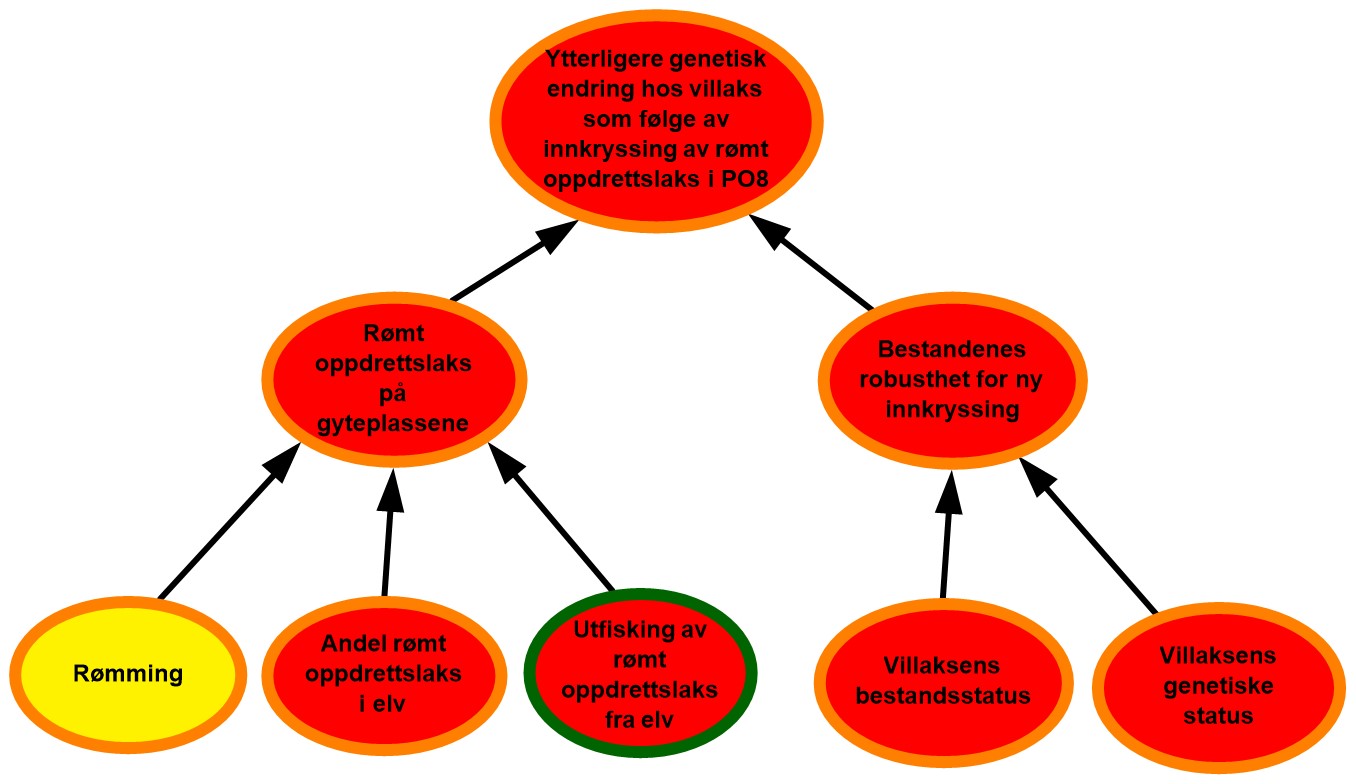
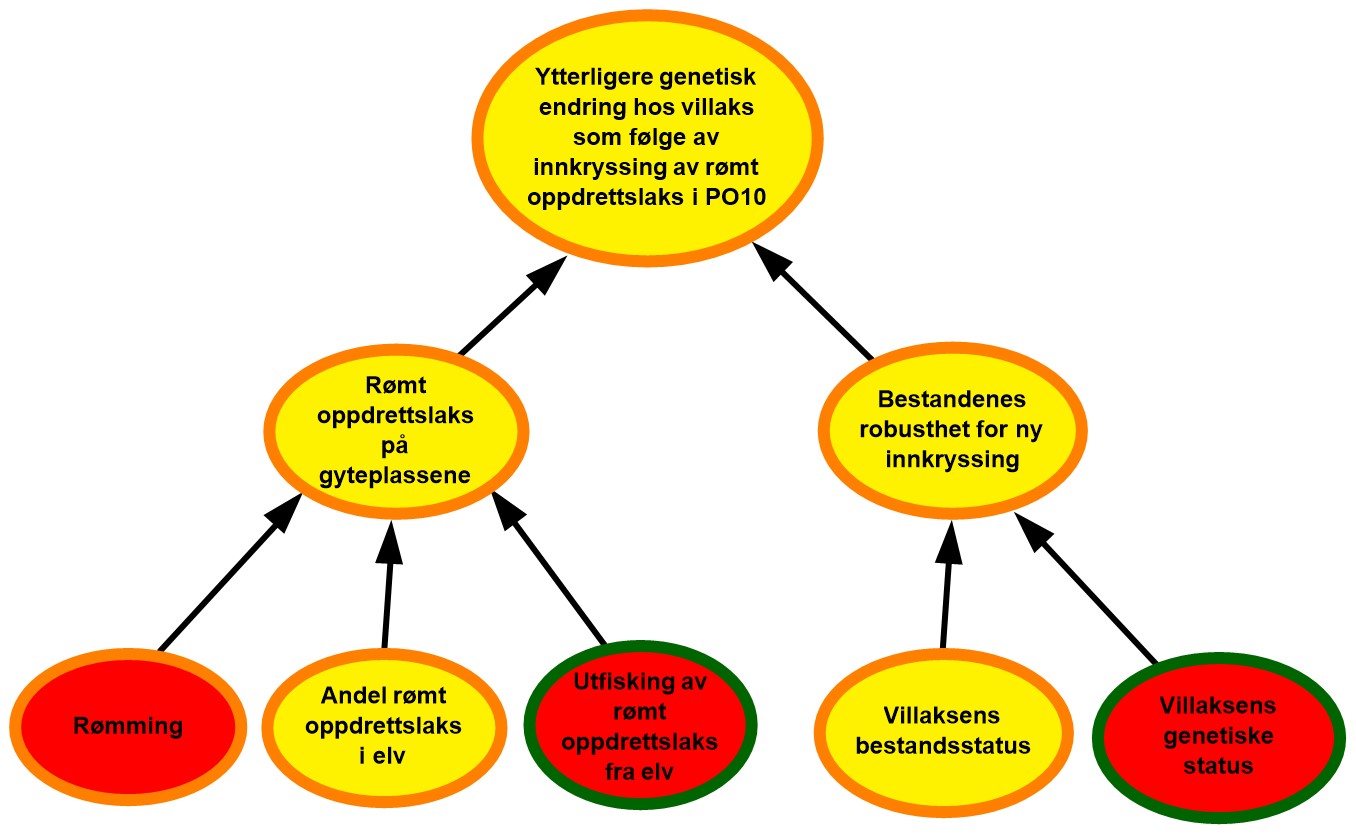
Figur 2.1. Eksempel på systematisering av risikofaktorene RS’, A’ og C’, samt eksempler på data og andre kilder til bakgrunnskunnskap.
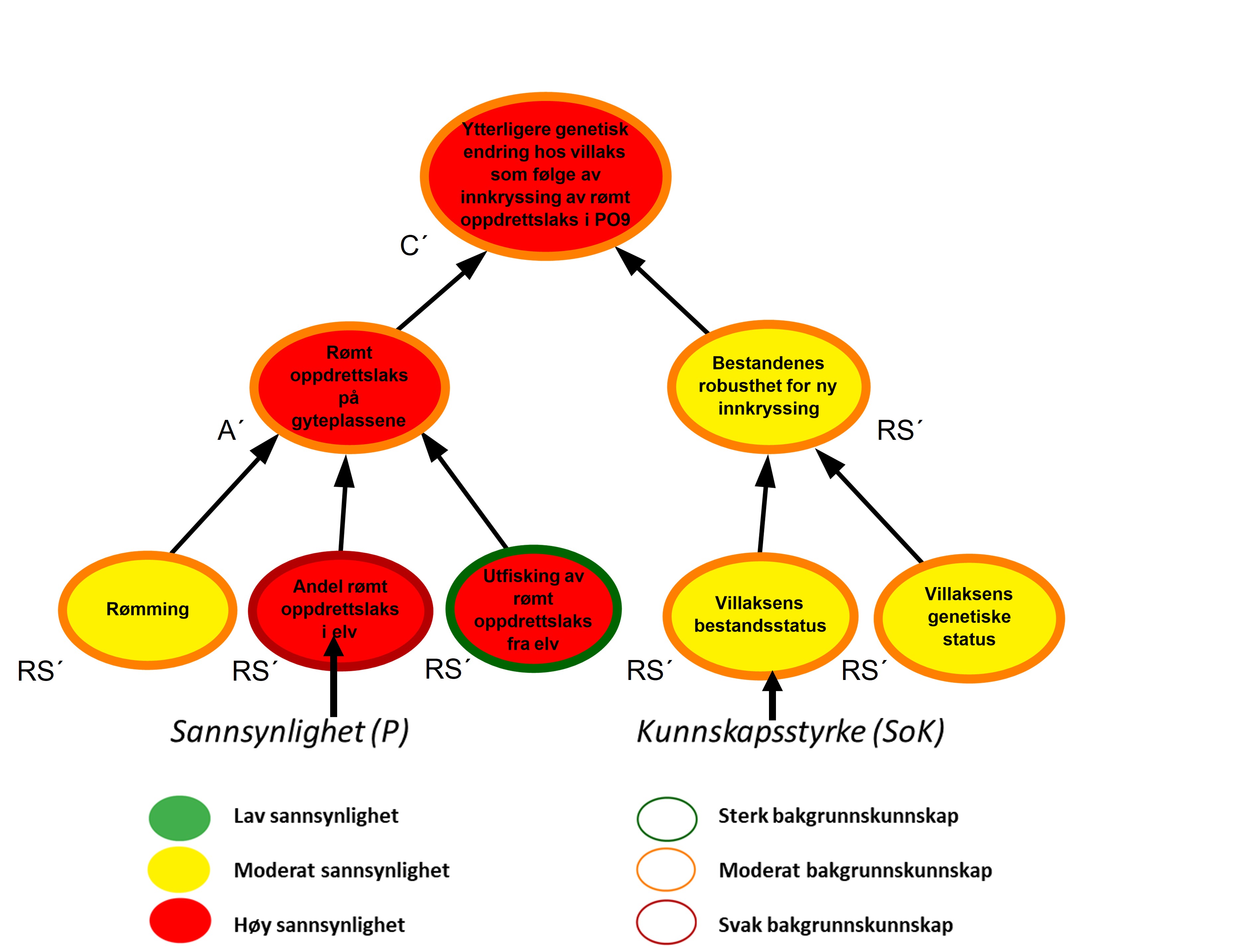
Bayesianske nettverk er valgt som et kvalitativt verktøy for å visualisere risiko, herunder; kausale sammenhenger RS, A’ og C’; sannsynligheter P knyttet til hvorvidt disse inntreffer og styrken på kunnskapen SoK disse vurderingene bygger på (figur 2.1). Hensikten er at de grafiske strukturene (kalt risikokart) og tilhørende argumentasjon skal bidra til hurtig og intuitiv risikoforståelse, også blant dem som ikke har bakgrunn fra oppdrettsnæringen.
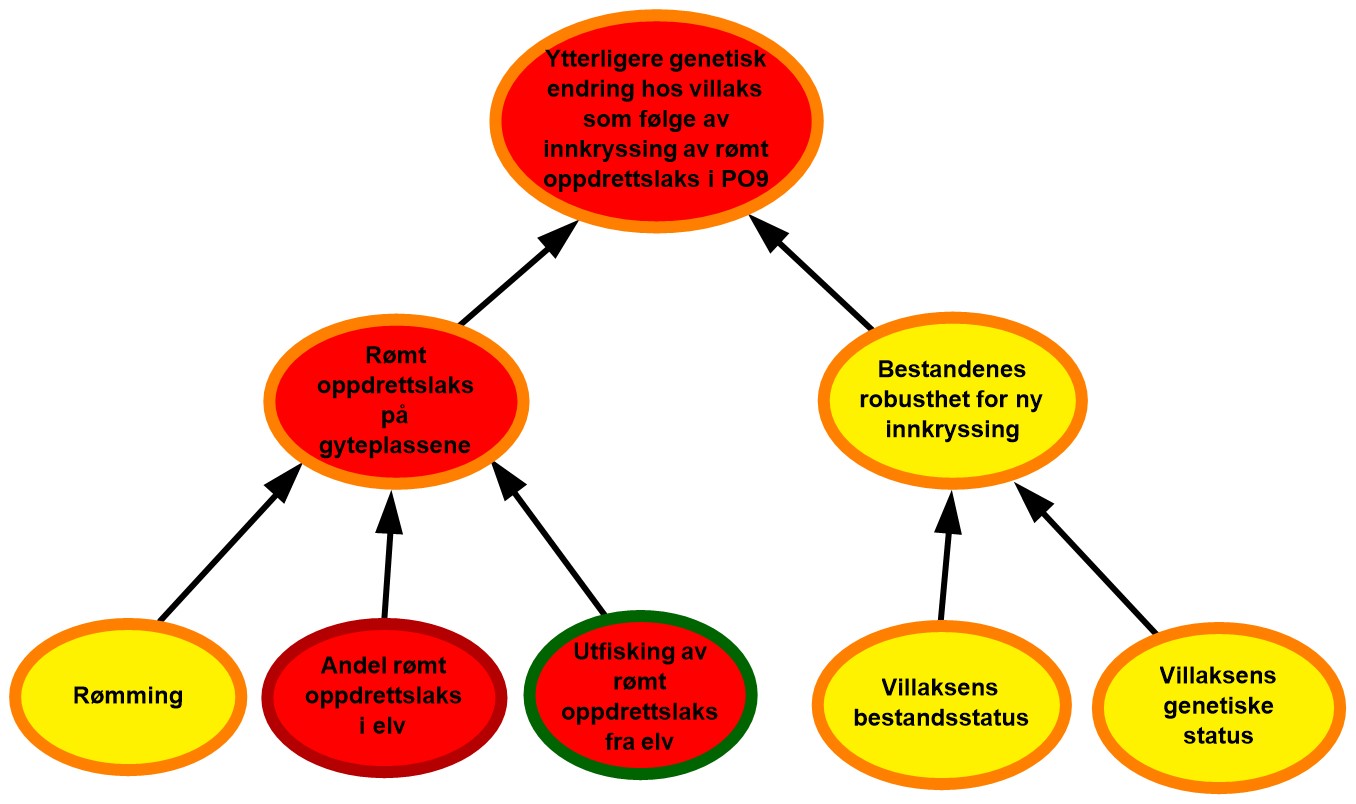
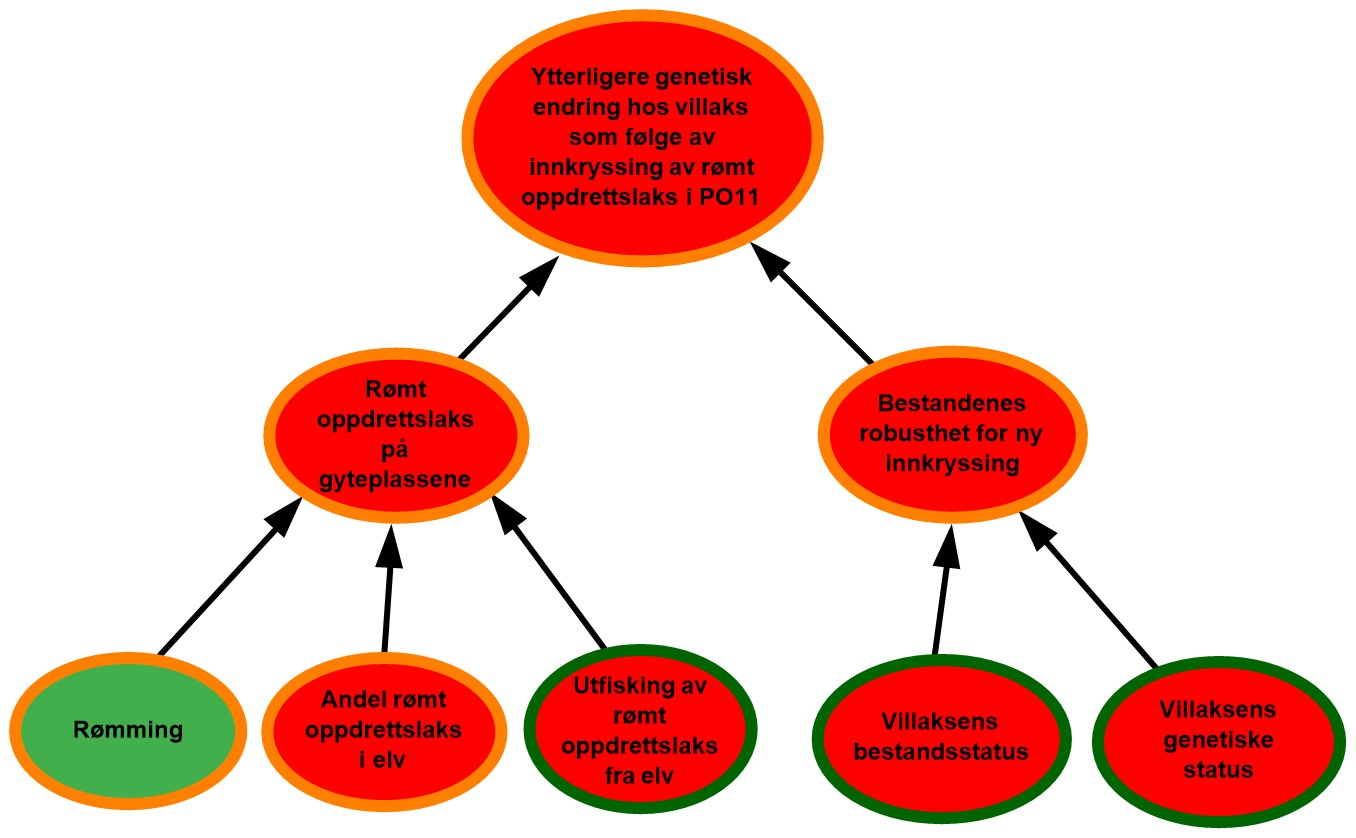
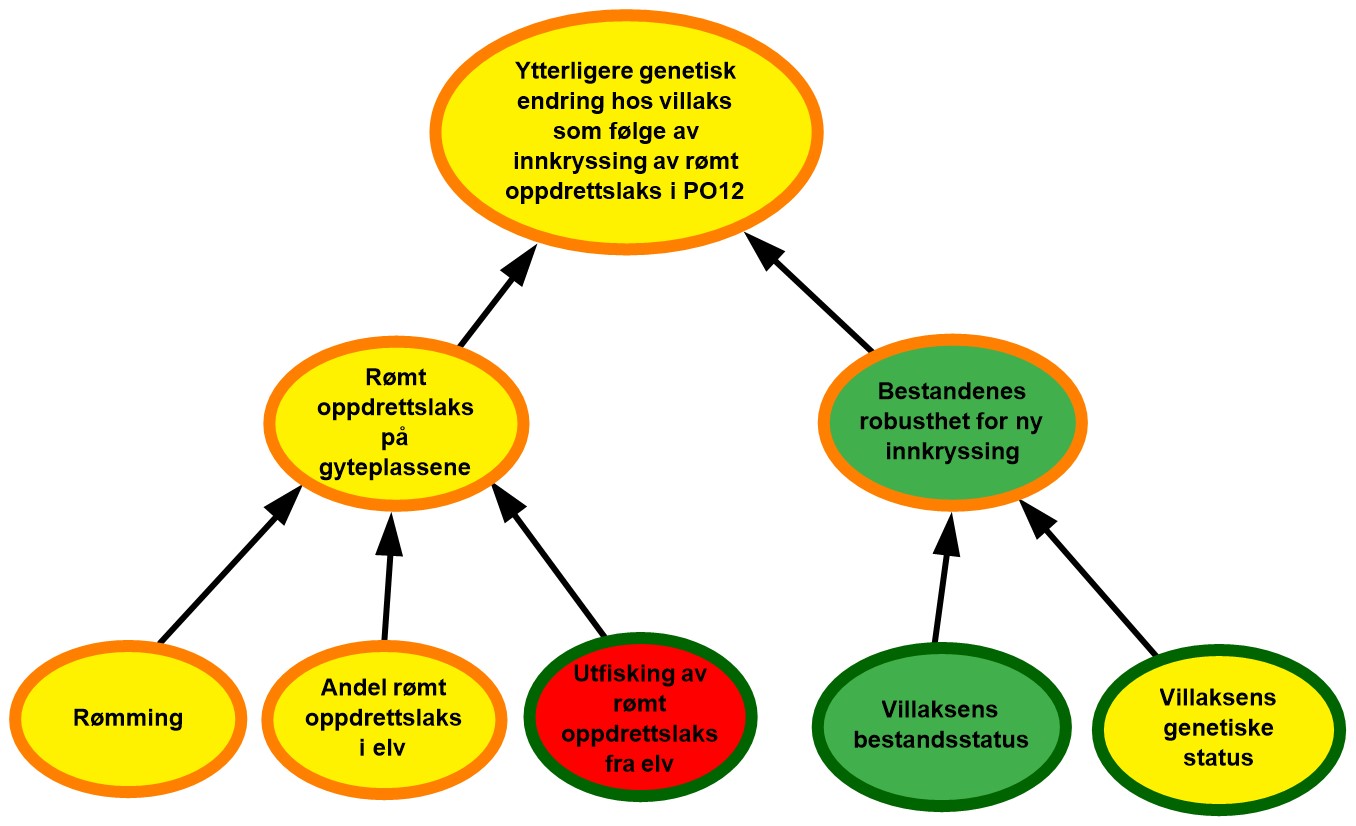
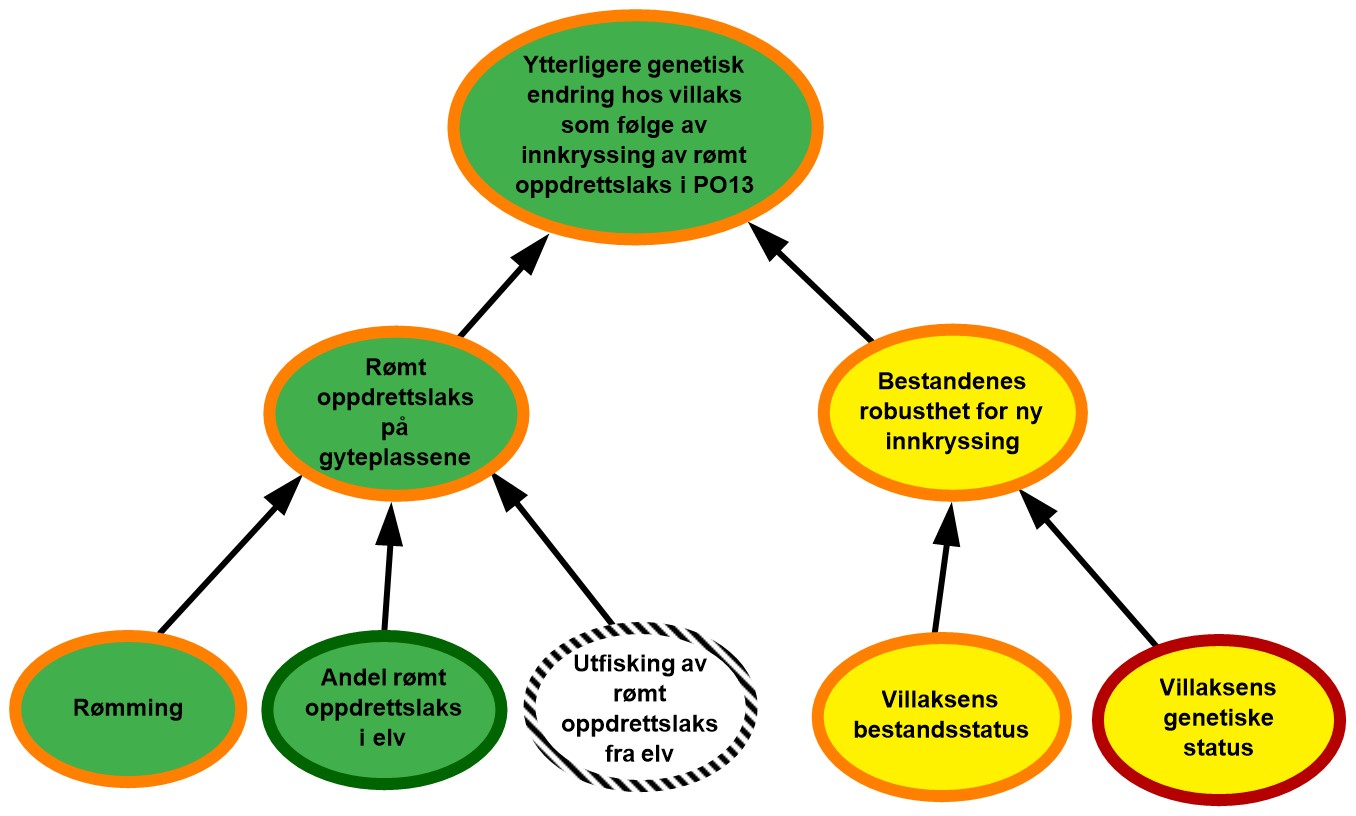
Figur 2.2. Eksempel på visualisering av risiko i PO9. Risikokart bestående av risikokilder, hendelser og konsekvenser på ulike kausale nivå med fargekoder som uttrykk for sannsynlighet og kunnskapsstyrke.
Risikokartene består av spesifikke risikokilder RS’, hendelser A’, og konsekvenser C’ (kalt noder) og piler som illustrerer årsak–virkning. Usikkerhet knyttet til disse måles ved tripletten (P, SoK, K). P er subjektive sannsynligheter som sier noe om hvorvidt RS’, A’, og C’ inntreffer. I risikokartene benyttes fargekodene grønn, gul og rød farge på en node for å illustrere høy, moderat og lav sannsynlighet. Fargen på sirkelen rundt noden illustrere hvorvidt denne vurderingen bygger på sterk, moderat eller svak kunnskap (SoK).
Forskerne benytter ofte en ønsket tilstand som referansepunkt for å støtte arbeidet med å fastsette sannsynligheter knyttet til RS’, A’ og C’. I figur 2.2 vil eksempelvis ønsket tilstand for risikokilden «Rømming» være lite eller ingen rømming av oppdrettslaks. I eksempelet vurderes tilstanden til denne risikokilden som moderat nært ønsket tilstand og fargekode gul benyttes for å visualisere moderat sannsynlighet for rømming i området. Forskerne argumenterer også for at sannsynlighetsvurderingen av risikokilden «Rømming» bygger på moderat kunnskap, og fargekoden oransje benyttes på sirkelen rundt noden.
Et annet eksempel som vist i figur 2.2 er risikokilden «utfisking av rømt oppdrettslaks fra elv» som påvirker i hvilken grad rømt oppdrettslaks fjernes fra elvene før den kan bidra til innkryssing på gyteplassene. Ønsket tilstand for denne risikokilden blir da at det årlig gjennomføres et effektivt uttaksfiske i alle elver med mye rømt oppdrettslaks. I eksempelet over er det høy sannsynlighet for at det ikke gjennomføres tilstrekkelig med uttaksfiske, noe som er langt fra vår ønsket tilstand. Sannsynligheten vurderes derfor som høy for at ikke all rømt oppdrettslaks fjernes fra eleven, og fargekode rød benyttes.
Hendelsen «Rømt oppdrettslaks på gyteplassene» påvirkes i all hovedsak av tre underliggende risikokilder der sannsynligheten for at disse skal inntreffe varierer mellom høy og moderat. For å forstå hvordan hendelsen «Rømt oppdrettslaks på gyteplassene » da kan vurderes som høy sannsynlig må man lese forskernes argumentasjon. I dette tilfellet er forklaring at det finnes mye rømt oppdrettslaks i eleven og at utfiskning er lite effektivt. Dette veier da tyngre enn den siste risikokilden. På samme vis må leseren studere forskernes argumentasjon for å forstå hvordan en samlet vurdering av kunnskapsstyrken knyttet til underliggende risikofaktorer summeres opp til kunnskapsstyrken for en overliggende hendelse.
Effekten av å evaluere bakgrunnskunnskapen blir spesielt tydelig i forbindelse med analyser av potensialet for overraskelser, såkalte «sorte svaner». Begrepet ble første gang presentert i en risikostyringssammenheng av Taleb (2007) og videreutviklet i bl.a. Aven (2014) som beskriver slike hendelser som ekstremt alvorlige, og som dukker opp som totale overraskelser sett i forhold til vår kunnskap og hva vi tror vil kunne skje. Fagmiljøene ved både Havforskningsinstituttet og andre som er involvert i denne risikovurderingen, beskriver potensialet for slike overraskelser der det eksempelvis finnes kombinasjoner av utilstrekkelig kunnskap, hypoteser og svakt funderte antakelser som kan bidra til å skjule risiko. Der risikokilder eller hendelser vurderes å kunne gi opphav til slike overraskelser er disse markert med et «sort svane» merke i risikokartene.
3. Faktorer som medfører genetisk endring hos villaks på grunn av innkryssing
Faktorer som påvirker omfanget av ytterligere genetiske endringer ved innkryssing av oppdrettslaks i villaksbestander, er i første rekke sannsynlighet for å få rømt oppdrettslaks på gyteplassene og bestandenes robusthet for ny innkryssing. Det er i hovedsak følgende tre faktorer som bestemmer sannsynlighet for hvor mye rømt oppdrettslaks det er på gyteplassene: rømming, andel rømt oppdrettslaks i elv og utfisking/fjerning av rømt oppdrettslaks fra elv. Hvor robuste bestandene er for innkryssing av rømt oppdrettslaks, påvirkes av genetisk status hos de ville bestandene og villaksens bestandsstatus (figur 2.1). Hvordan disse faktorene virker inn på risiko for ytterligere genetiske endringer hos villaks, utdypes i teksten under:
Rømt oppdrettslaks på gyteplassene. Det er godt dokumentert at det er en sammenheng mellom andel rømt oppdrettslaks på gyteplassene over tid og graden av genetisk innkryssing. Andel rømt oppdrettslaks som står på gyteplassene under gytingen forklares i all hovedsak av rømming, andel rømt oppdrettslaks i elv, samt utfisking av rømt oppdrettslaks fra elv. Andel rømt oppdrettslaks i elv og utfisking av rømt oppdrettslaks i elvene er lagt mest vekt på i vurderingen av sannsynlighet for rømt oppdrettslaks på gyteplassene.
Ønsket tilstand vil være få eller ingen rømte oppdrettslaks på gyteplassene.
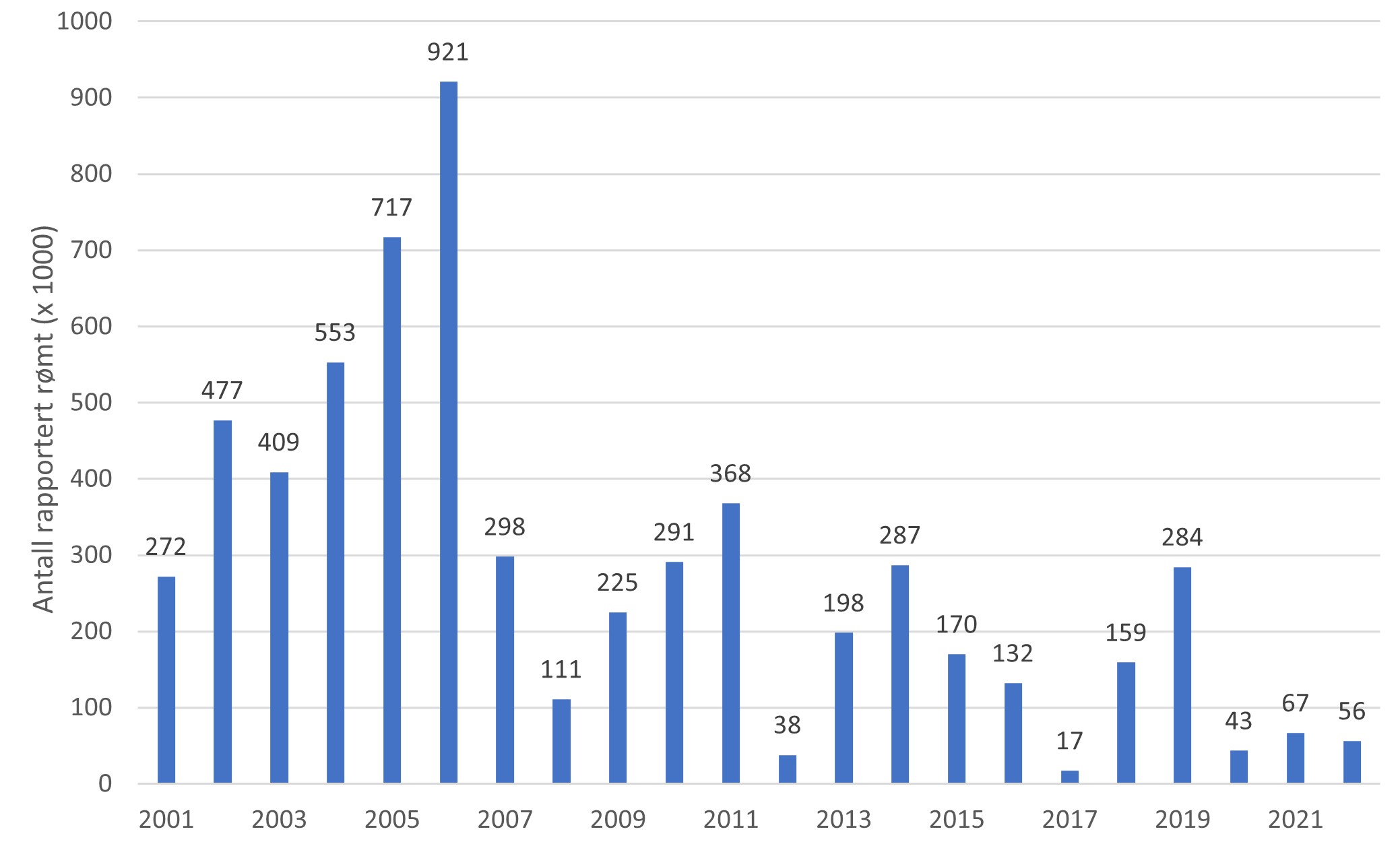
Rømming. Rømming fra oppdrettsanlegg skal rapporteres til Fiskeridirektoratet, og en oversikt over offisielle rømmingstall er tilgjengelig på deres nettsider. De offisielle rømmingstallene er med all sannsynlighet et underestimat av reell rømming, noe som er vist både i merkestudier og ved gjentatte hendelser hvor en finner rømt oppdrettslaks uten at det er rapportert om rømming. Det er også stokastisk usikkerhet knyttet til resultatet av estimering av antall rømt oppdrettslaks grunnet store mengder laks i merdene. En forsinkelse i tallene kan oppstå grunnet tid mellom rømmingstidspunkt og utslakting. Selv om det er usikkerhet i de offisielle rømmingstallene, har vi gjort en vurdering basert på årlig gjennomsnittlig rapportert rømming i perioden 2017–2021 (se tabell 6.1 i kunnskapsstatus). I dette tidsrommet ble det meldt om totalt 573 128 rømte oppdrettslaks.
Produksjonsområder med inntil noen hundre rapporterte rømte oppdrettslaks per år i perioden 2017–2021 er kategorisert som områder med lite eller ingen rømming (fargekode grønn), områder med noen tusen rapporterte rømte oppdrettslaks per år i perioden 2017–2021 er moderat (fargekode gul) og der årlig rapporterte gjennomsnitt i samme perioden er på titusenvis av rømt fisk, anses dette som områder med mye rømming (fargekode rød). Kunnskapsstyrken for de reelle rømmingstallene er usikker og vurderes som moderat (fargekode gul) for samtlige produksjonsområder på grunn av forhold beskrevet over (figur 2.1). Det er også usikkerhet knyttet til påvirkning fra rømming fra andre produksjonsområder. Det er påvist en sammenheng mellom observasjon av rømt oppdrettslaks i elv innen et gitt produksjonsområde og rapportert antall rømt oppdrettsfisk fra det aktuelle, samt de nærliggende produksjonsområdene.
Ønsket tilstand vil være lite eller ingen rømming av oppdrettslaks.
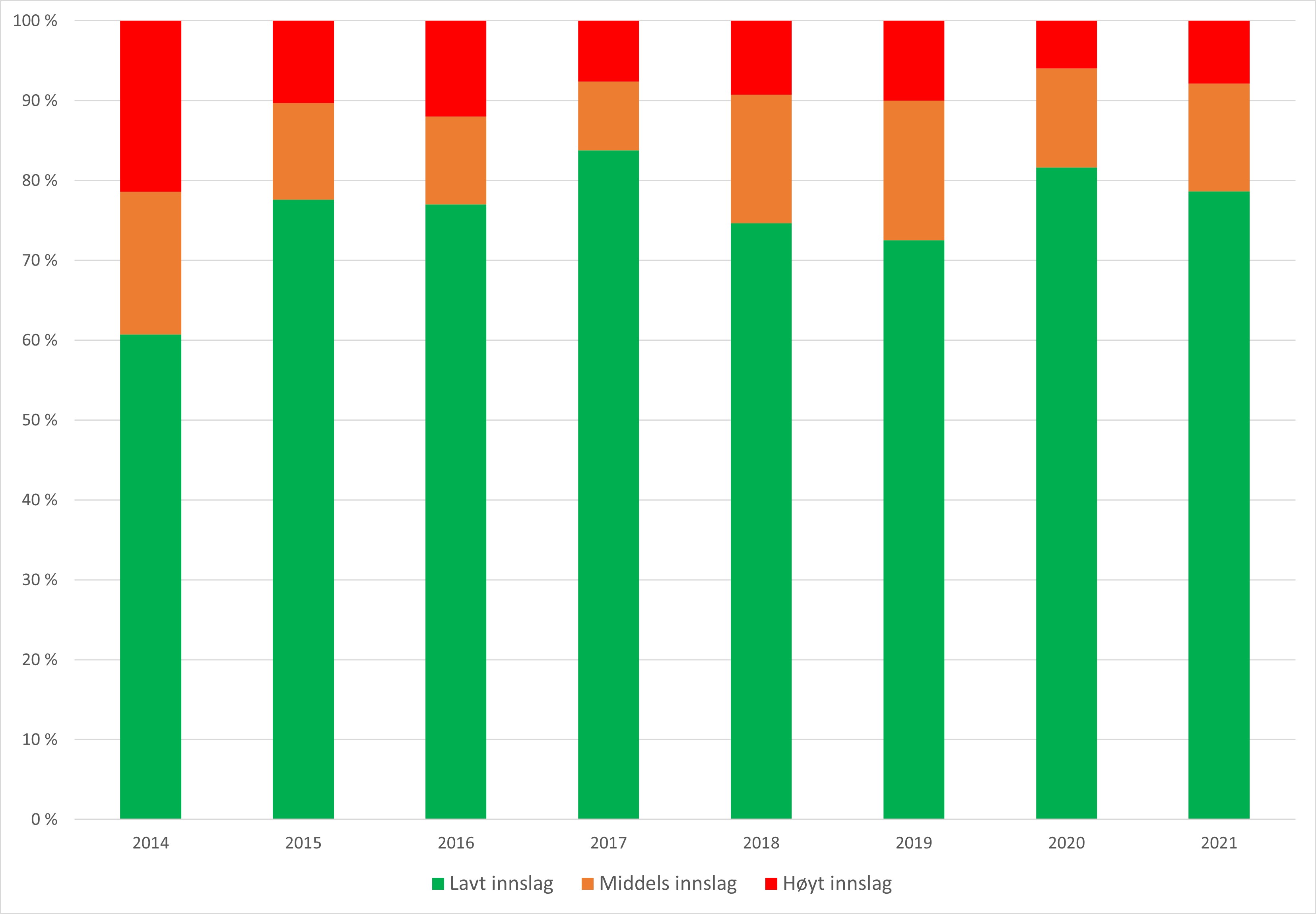
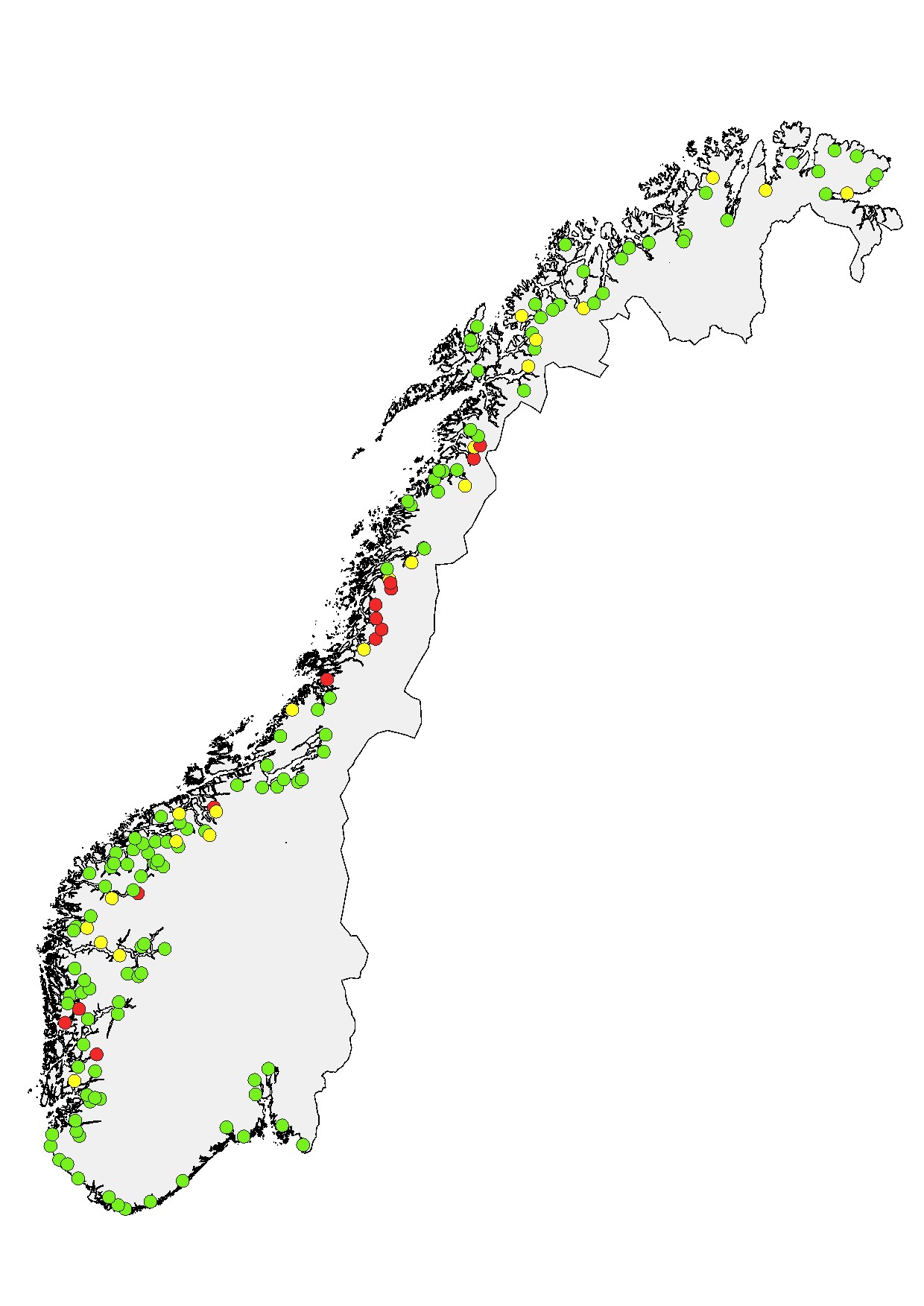
Andel rømt oppdrettslaks i elv. Det er dokumentert en positiv sammenheng mellom den observerte andelen rømt oppdrettslaks i en elv over tid, og graden av genetisk innblanding. Andel rømt oppdrettslaks i elv anses som den mest bestemmende av de tre faktorene som påvirker hvor høy sannsynlighet det er for å finne mye rømt oppdrettslaks på gyteplassene. Andel rømt oppdrettslaks blir registrert årlig i omtrent 200 vassdrag gjennom det nasjonale overvåkningsprogrammet for rømt oppdrettslaks. Det foreligger derfor gode tall for denne faktoren i mange av produksjonsområdene.
Å klassifisere tilstanden med hensyn til andel rømt oppdrettslaks i et produksjonsområde ut fra gjennomsnittet av de estimerte andelene i undersøkte vassdrag, vil ikke være helt korrekt eller gi en god beskrivelse av tilstanden. Overvåkningsprogrammets vurdering og klassifisering er basert på forskjellige observasjonsmetoder som måler andelen rømt oppdrettslaks i vassdrag på ulike måter, og estimatene fra disse metodene kan dermed gi noe forskjellige andeler. Dette aspektet er grundigere diskutert i rapportene «Rømt oppdrettslaks i vassdrag». I perioden 2014-2017 vurderte prosjektgruppen i overvåkningsprogrammet vassdragene ut fra et forenklet system der vassdragene ble kategorisert til å ha lav til moderat sannsynlighet for ytterligere genetisk endring dersom andel rømt laks var <10 %, moderat sannsynlighet for ytterligere genetisk endring dersom andel rømt oppdrettslaks var rundt 10 %, og høy sannsynlighet når andel rømt oppdrettslaks var >10 %. Siden 2018 har overvåkningsprogrammet klassifisert vassdragene til å ha høyt innslag av rømt oppdrettslaks dersom andelen estimert er >10 %, middels innslag ved estimater mellom 4 og 10 %, og til lavt innslag dersom estimatet er under 4 %. I praksis har denne endringen i klassifisering ikke endret vurderingen av vassdragene i særlig grad. Vurderingene utføres for hvert vassdrag for hvert år og baserer seg på alle tilgjengelige data fra ulike metoder og ekspertkunnskap hos forskerne i prosjektgruppen.
I risikovurderingen har vi brukt klassifiseringene fra overvåkningsprogrammet for de enkelte vassdragene i perioden 2017–2021 innenfor hvert av produksjonsområdene og vurdert disse samlet for å kategorisere tilstanden innenfor hvert produksjonsområde. Dette datagrunnlaget (se tabell 6.2 i kunnskapsstatus) er deretter vurdert opp mot klassifiseringsreglene beskrevet nedenfor. Det er viktig å påpeke at i enkelte tilfeller, der et produksjonsområde kommer ut i grenseland mellom kategoriene, er regelen overstyrt der tilgjengelig kunnskap tilsier at dette vil være riktig. Slike tilfeller er nærmere beskrevet og begrunnet under hvert produksjonsområde.
Lav andel rømt oppdrettslaks i et produksjonsområde (fargekode grønn) defineres som:
Andelen vassdrag i produksjonsområdet vurdert som «lav» av overvåkningsprogrammet i perioden 2017–2021 må være minst 90 %.
I tillegg må ingen vassdrag i produksjonsområdet være vurdert som «>10 % rømt oppdrettslaks» i perioden.
Høy andel rømt oppdrettslaks i et produksjonsområde (fargekode rød) defineres som:
Andelen vassdrag i produksjonsområdet vurdert som «lav» i perioden 2017-2021 er mindre enn 50 %.
Eller andelen vassdrag i produksjonsområdet vurdert som «>10 % rømt oppdrettslaks» av overvåkningsprogrammet i perioden 2017–2021 er større enn 10 %.
For produksjonsområder som ikke faller innenfor disse klassifiseringene defineres andelen som moderat (fargekode gul).
Kunnskapsstatus med hensyn til rømt oppdrettslaks innenfor hvert produksjonsområde er vurdert ut fra to faktorer som er vektet likt: andel elver i et produksjonsområde som er undersøkt og klassifisert av overvåkningsprogrammet, og andel av samlet gytebestandsmål som overvåkes i programmet. Det blir også tatt hensyn til om de to faktorene (andel elver og andel av gytebestand) gir lik eller ulik vurdering av andel rømt oppdrettslaks i produksjonsområdet.
Ønsket tilstand vil være få eller ingen rømte oppdrettslaks i elvene.
Utfisking og fjerning av rømt oppdrettslaks fra elv. Utfisking av rømt oppdrettslaks gjennomføres i hovedsak i regi av oppdrettsnæringens sammenslutning for utfisking av rømt oppdrettsfisk (OURO) i vassdrag der det nasjonale overvåkningsprogrammet har påvist høye andeler året før, men også etter direkte pålegg fra forvaltningsmyndigheter ved konkrete rømmingshendelser og i vassdrag hvor mye oppdrettslaks blir observert. I risikovurderingen bruker vi antall rømt oppdrettslaks fjernet per produksjonsområde, og tallene hentes fra overvåkningsprogrammet og fra OURO direkte. I perioden 2017 – 2021 er det årlig gjennomført utfiskingstiltak i minst 58-94 vassdrag for å redusere andelen rømt oppdrettslaks. Antall elver med utfisking kan også være noe høyere, da aktivitet i vassdrag som ikke rapporteres til overvåkningsprogrammet kan forekomme. Elver hvor utfisking har vært gjennomført, uten observasjoner av rømt fisk, er også inkludert i datagrunnlaget for utregning av antall elver med utfisking per produksjonsområde. I overvåkningsprogrammet vil kun utfisket oppdrettslaks som er bekreftet gjennom skjellanalyser inngå i datagrunnlaget. Det reelle antallet oppdrettslaks som tas ut vil derfor kunne være noe høyere grunnet manglende eller uleselige skjellprøver.
Utfisking og fjerning av rømt oppdrettslaks er et viktig, og i mange vassdrag et til dels effektivt tiltak, for å redusere risiko for genetisk endring. Tiltaket har likevel en del begrensninger som medfører at risikoen for ytterligere genetisk endring fra rømt oppdrettslaks ikke vil kunne fjernes helt i et produksjonsområde.
Begrensende faktorer er:
Effekten av utfiskingen vil variere med værforhold og vassdragets vannføring og topografi
Utfisking er mest effektivt i små vassdrag med god sikt, og ikke i de største laksevassdragene
All rømt oppdrettslaks fjernes ikke, og det kan heller ikke utelukkes at det kommer inn mer rømt oppdrettslaks etter utfiskingsperioden, men før gytingen er over
Utfiskingen er rettet mot vassdrag med høy andel rømt oppdrettslaks det foregående året, og derfor kan det i praksis være høy andel rømt oppdrettslaks i et vassdrag uten at det gjennomføres tiltak samme år
Det gjennomføres få utfiskingstiltak i vassdrag som ikke er en del av overvåkningsprogrammet, og disse vassdragene har en ukjent andel rømt oppdrettslaks
Utfisking må gjennomføres på en måte som ikke får negative konsekvenser for vill laksefisk i elven
Disse begrensningene i effekten av utfisking er tatt med i vurderingen av denne påvirkningsfaktoren.
Ingen av produksjonsområdene har full dekning gjennom overvåkningsprogrammet, og det faktum at utfiskingen ikke fjerner all rømt oppdrettslaks som kommer til vassdragene sesongen igjennom, gjør at ingen områder vurderes til å ha stor effekt av utfisking (fargekode grønn). I produksjonsområder hvor minst 90 % av vassdragene med høyt innslag også rapporterte om utfisking samme år vurderes effekten av utfiskingen som moderat (fargekode gul). I produksjonsområder hvor under 90 % av vassdragene med høyt innslag også rapporterte om utfisking samme år vurderes effekten av utfiskingen å være lav (fargekode rød). I områder med vassdrag som egner seg dårlig for utfisking kan regelen overstyres, da effektiviteten av utfisking vil reduseres. Slike tilfeller er nærmere beskrevet og begrunnet under hvert produksjonsområde. Kriteriene for vurdering av effekten av utfisking er som i 2020, da kriteriene ble endret fra tidligere vurderinger.
Kunnskapsstyrken settes til god (fargekode grønn) for samtlige produksjonsområder fordi utfiskingstallene anses som sikre (figur 2.1). Datagrunnlaget for vurdering av denne faktoren er tilgjengelig (se tabell 6.3 i kunnskapsstatus).
Ønsket tilstand vil være at det årlig gjennomføres effektiv utfisking i alle elver der det finnes rømt oppdrettslaks.
Bestandenes robusthet for ny innkryssing. Hvor stor gytesuksess voksen rømt oppdrettslaks har på gyteplassene er avhengig av hvor mange ville konkurrenter de har. Bestandenes robusthet for ny innkryssing avhenger av villaksens bestandsstatus og villaksens genetiske status.
Dersom bestandene når sine gytebestandsmål, har de mange nok vill gytelaks på gyteplassene til å kunne utnytte elvens produksjonspotensial og har dermed god bestandsstatus. Konkurransen på gyteplassen blir enda sterkere dersom villaksbestanden er høyere enn gytebestandsmålet. Bestander med høyt høstingspotensial (vesentlig flere voksen hunnlaks kommer tilbake enn det som er nødvendig for å nå gytebestandsmålet) har også større sannsynlighet for å nå gytebestandsmålet i årene som kommer, selv om overlevelsen i havet skulle bli redusert. Konkurransen mellom ungfiskene i elven vil også være større hvis det er mange laks som gyter i vassdraget, og avkom av rømt oppdrettslaks vil gjøre det relativt sett dårligere hvis det er mange ville ungfisk å konkurrere med.
Når villaksen gyter med den rømte oppdrettslaksen vil det oppstå genetiske endringer i avkommene som avviker fra den genetiske strukturen til den enkelte villaksebestanden. Rømt oppdrettslaks har antakeligvis større gytesuksess i konkurranse med innkryssede individer enn med ikke-innkrysset villaks. Avkom av rømt oppdrettslaks har sannsynligvis høyere overlevelse (og større relativ konkurransestyrke) når individene de konkurrerer med er innkrysset. Det er derfor sannsynlig at høy innkryssing av oppdrettslaks i bestandene også vil gjøre dem mindre robuste for innkryssing av rømt oppdrettslaks i framtiden enn bestander som har liten grad av innkryssing.
Vi antar derfor at bestander som både når gytebestandsmålet, har et høyt produksjonspotensial og har liten grad av innkryssing er mer robuste mot ytterligere genetisk endring som følge av rømt oppdrettslaks enn bestander som har lite produksjonspotensial og/eller som ikke når gytebestandsmålet og/eller har høy grad av genetisk endring.
Kunnskapsstyrken om konkurranseforholdene beskrevet ovenfor, og den kombinerte effekten av bestandsstatus og genetisk status, er begrenset til tross for at vi har relativt god kunnskap om de to underliggende faktorene hver for seg. Styrken på kunnskapen knyttet til «bestandenes robusthet for ny innkryssing» vurderes derfor som moderat i alle produksjonsområdene (fargekode gul) (figur 2.1).
Ønsket tilstand vil være at bestandene er så robuste som mulig mot ny innkryssing.
Villaksens bestandsstatus. Vurdering av villaksens bestandsstatus i produksjonsområdene er basert på beregninger av måloppnåelse for gytebestandsmål og høstingspotensial for den enkelte villaksbestand i perioden 2017–2021. Gjennomsnittlig gytebestandsmåloppnåelse og gjennomsnittlig høstingspotensial ble beregnet for hvert produksjonsområde (se tabell 6.4 i kunnskapsstatus). Grunnlaget for dette er antall bestander i hver av kategoriene fra Vitenskapelig råd for lakseforvaltning (VRL) sin vurdering innenfor hvert produksjonsområde.
Gjennomsnittene ble beregnet både som uveide gjennomsnitt der hvert vassdrag teller likt uavhengig av størrelse, og som veide gjennomsnitt der vassdragenes gytebestandsmål bestemmer vektingen. I den vektede vurderingen vil vassdrag med store gytebestandsmål telle mer enn vassdrag med små gytebestandsmål i den samlede vurderingen innenfor et produksjonsområde. Hvert produksjonsområde ble så gitt en vurdering som om det var et stort vassdrag. For vassdrag som ikke er vurdert etter delnormen, talte vi opp hvilken forenklet vurdering de ble gitt i Vitenskapelig råd for lakseforvaltning sin rapport fra 2021. Den forenklede vurderingen gjelder for 2015–2019, og ble bare tillagt vekt dersom en liten del av vassdragene i produksjonsområdet er gitt en full vurdering, eller der veide og uveide vurderinger ga forskjellig resultat.
I denne risikovurderingen blir kategoriene «god» og «svært god» kvalitet i delnorm gytebestandsmål og høstingspotensial fra «Kvalitetsnorm for ville bestander av laks (Salmo salar)» vurdert som god tilstand (fargekode grønn). Tilsvarende blir «moderat» kvalitet etter kvalitetsnormen kategorisert som moderat (fargekode gul), og «dårlig» og «svært dårlig» kvalitet etter normen blir her kategorisert som dårlig tilstand (fargekode rød).
Kunnskapsstyrken vurderes ut ifra andel av gytebestandsmålet som er vurdert innenfor hvert produksjonsområde. Det blir også tatt hensyn til om klassifiseringen blir den samme uavhengig av hvilken klassifiseringsmåte man legger til grunn.
Ønsket tilstand vil være en bestand som oppnår sitt gytebestandsmål og har normalt eller høyt høstingspotensial.
Villaksens genetiske status. Innkryssing av rømt oppdrettslaks har allerede ført til omfattende genetisk endring hos mange villaksbestander. For å identifisere avkom av rømt oppdrettslaks som er klekket i naturen, og har opphav i gyting et eller flere år tidligere, må man gjøre molekylærgenetiske analyser av individene. De molekylærgenetiske metodene beregner graden av genetisk påvirkning av oppdrettslaks på villaksbestander, og hvorvidt den beregnete påvirkningen er statistisk signifikant. Status for genetisk innkryssing fra rømt oppdrettslaks på ville laksebestander er basert på en beskrivelse av genetisk status i 227 ville laksebestander, utført av Norsk institutt for naturforskning og Havforskningsinstituttet i fellesskap. Beskrivelsen publiseres i rapportserien «Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander» og utgjør delnorm genetisk integritet til «Kvalitetsnorm for ville bestander av laks (Salmo salar)». Siste rapport i serien ble publisert i 2020, og ble hensyntatt i risikovurderingen publisert i 2021 og 2022. Årets risikovurdering er derfor basert på samme datagrunnlag som den foregående vurderingen for denne faktoren. En ny rapport i serien er forventet publisert i 2023, og vil bli hensyntatt ved neste risikovurdering.
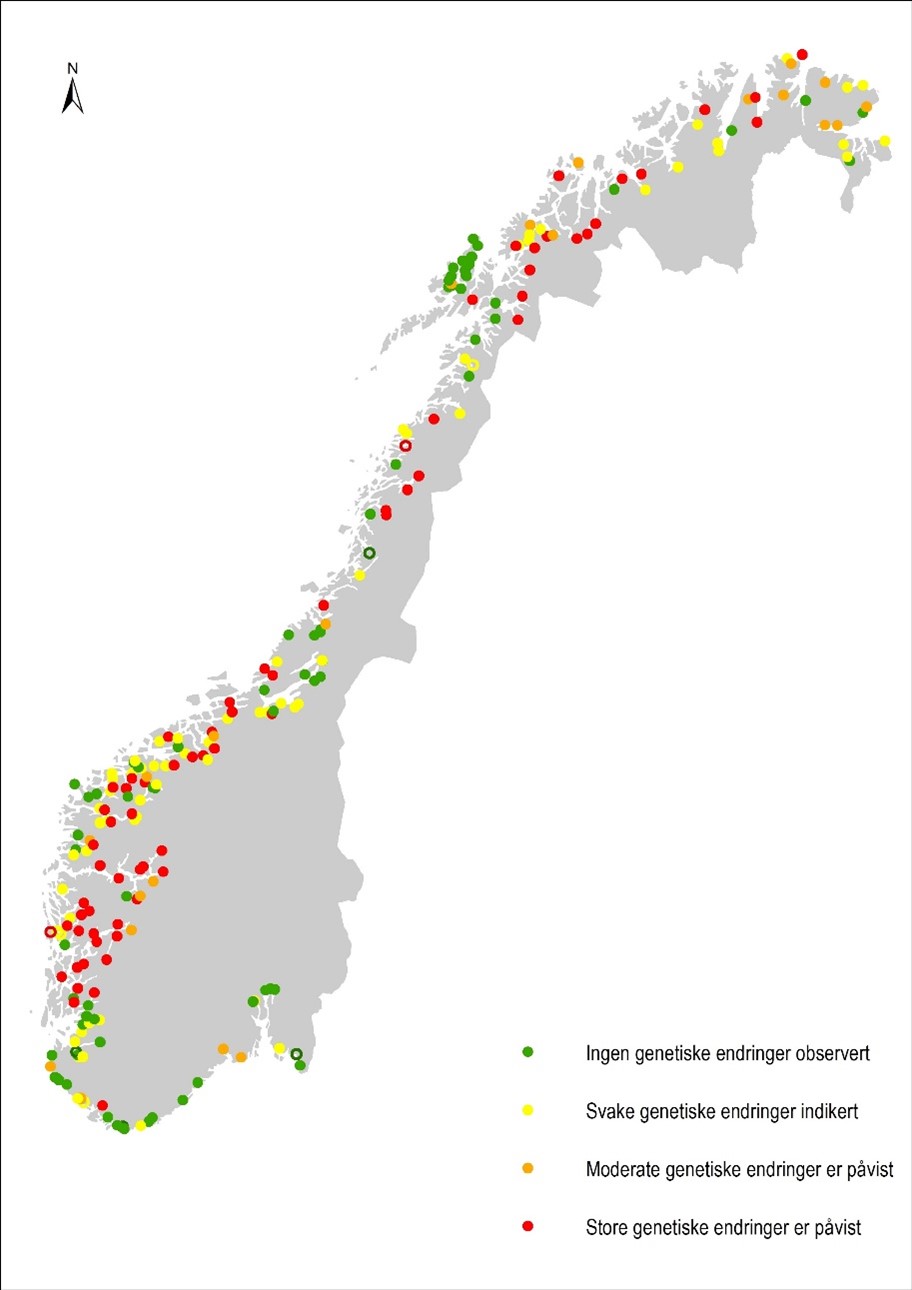
I kvalitetsnormen er den genetiske påvirkningen av rømt oppdrettslaks på de ulike bestandene av villaks karakterisert som tilstandsklasse «svært god/god» hvis ingen genetiske endringer er observert, «moderat» hvis svake genetiske endringer er indikert (estimert innkryssing på 1 - 4 %), «dårlig» hvis moderate genetiske endringer er påvist (estimert innkryssing på 4 -10 %) eller «svært dårlig» hvis store genetiske endringer er påvist (estimert innkryssing på 10 % eller mer). Det er utarbeidet kvalitative og kvantitative kriterier for hver av de fire tilstandsklassene, og i alt er det undersøkt om lag 52 000 villaks for å beskrive genetisk innkryssing i ville laksebestander. De 227 ville laksebestandene som er undersøkt per 2020 representerer om lag 94 % av den samlede villaksressursen i Norge, som i denne sammenhengen er definert som det totale gytebestandsmålet i Norges 448 lakseelver.
I denne risikovurderingen blir den genetiske påvirkningen av rømt oppdrettslaks på ville laksebestander oppsummert per produksjonsområde (se tabell 6.5 i kunnskapsstatus). Bidragene fra genetisk innkryssing i hver av de undersøkte bestandene i et produksjonsområde blir både gitt som en uveid andel (dvs. alle bestandene teller likt) og som andel vektet slik at store bestander teller mer enn små bestander.
Produksjonsområder med totalvurdering ingen genetiske endringer observert (tilstandsklasse «svært god/god») blir her kategorisert som god status (fargekode grønn).
Produksjonsområder med totalvurdering svake genetiske endringer indikert (tilstandsklasse «moderat», tilsvarende 1-4 % estimert innkryssing) blir her kategorisert som moderat status (fargekode gul).
Produksjonsområder med totalvurdering moderate eller store genetiske endringer påvist (tilstandsklasse «dårlig» og «svært dårlig», tilsvarende estimert innkryssing på 4 % eller mer) blir her kategorisert som dårlig status (fargekode rød).
Kunnskapsstyrken for vurderingene av genetisk status er først og fremst knyttet til hvor stor andel av den samlede villaksressursen i hvert produksjonsområde som er representert ved de undersøkte villaksbestandene i området.
Ønsket tilstand vil være at få eller ingen genetiske endringer er observert.
4. Risikovurdering av ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i de 13 produksjonsområdene
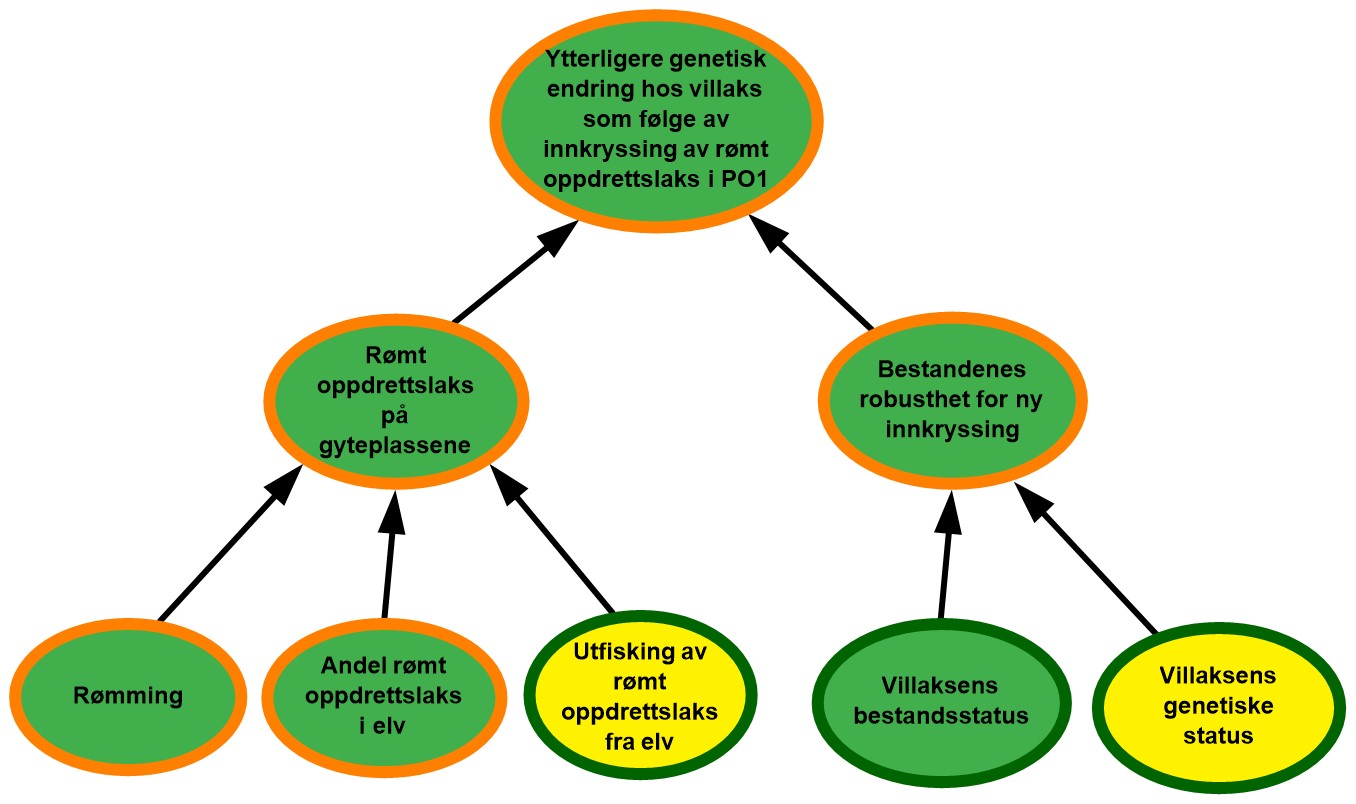
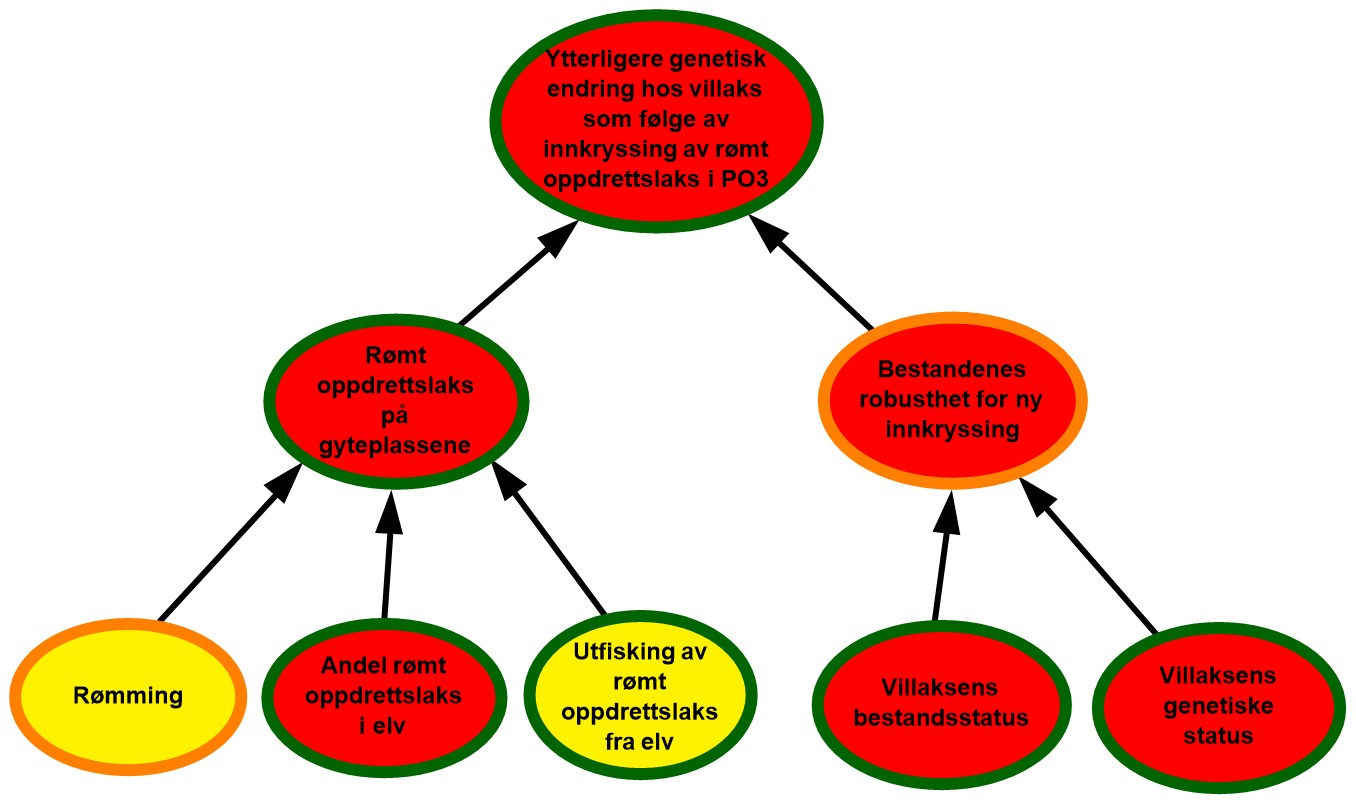
4.1. Produksjonsområde 1, Svenskegrensen til Jæren
Figur 4.1. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 1 (PO1), Svenskegrensen til Jæren.
Rømming. Det ble rapportert om 102 rømte oppdrettslaks i PO1 i perioden 2017–2021, og det vurderes at området har lave rømmingstall. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. Det var i snitt ca. 45 % av vassdragene i området (18 av 40) som årlig inngikk i overvåkningsprogrammet i perioden 2017-2021, noe som representerte 83 % av gytebestandsmålet. De fleste (98 %) registreringer i vassdrag i ulike år er vurdert å vise et lavt innslag av rømt oppdrettslaks, mens det i 2020 var to forekomster av vassdrag med høye innslag. Siden de rapporterte rømmingstallene i området har vært lave over tid kan de høye innslagene i 2020 enten forklares med urapporterte rømminger eller at rømt oppdrettsfisk fra andre produksjonsområder har funnet veien opp i vassdragene. Til tross for enkelttilfellene i 2020, ble det ikke registrert vassdrag med høye innslag i perioden 2017-2019, og det ble heller ikke registrert nye tilfeller i 2021. Basert på dette vurderes det derfor å være lav sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Selv om en høy andel av gytebestandsmålet dekkes av overvåkningen, er andelen vassdrag som dekkes, noe lav. Det kan dermed finnes vassdrag i området med ukjent innslag av rømt oppdrettslaks, og kunnskapsstyrken vurderes derfor som moderat.
Utfisking av rømt oppdrettslaks fra elv. Det ble årlig gjennomført utfiskingstiltak i 1-4 vassdrag i overvåkningsperioden 2017-2021 og det ble fjernet totalt 62 oppdrettslaks. Det er få elver med høyt innslag av rømt oppdrettslaks i området, og høye innslag ble kun registrert i ett av årene. I 2020 var det to vassdrag med høyt innslag av oppdrettslaks og det ble gjennomført utfisking i begge disse vassdragene samme år. Effekten av utfisking vurderes som moderat ettersom utfisking kun gjennomføres i en begrenset tidsperiode og det vurderes derfor at det er moderat sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. I perioden 2017-2021 har det totalt sett vært lite rømming i området og lav sannsynlighet for å få rømt oppdrettslaks i elvene. Utfisking av rømt oppdrettslaks vurderes å være moderat i samme periode. Basert på dette vurderes det å være lav sannsynlighet for å finne mye rømt oppdrettslaks på gyteplassene i PO1. Det er moderat kunnskap om hvor mye rømt oppdrettslaks det er i elvene, da det er en del elver som ikke dekkes av overvåkningsprogrammet. Utfiskingstallene ansees som gode, men det er usikkerhet knyttet til hvor mye oppdrettslaks som har rømt fra anlegg i området. Totalt sett vurderes kunnskapsstyrken derfor som moderat.
Villaksens bestandsstatus. Vassdragene med full vurdering utgjør 85 % av det samlede gytebestandsmålet i produksjonsområdet. Gytebestandsmålet blir nådd for de fleste vassdragene i produksjonsområdet i perioden og det høstbare overskuddet er også godt i de fleste vassdragene. Tilstanden for villaksbestandene i området vurderes som god. Klassifiseringen blir den samme uavhengig av hvilken klassifiseringsmåte man legger til grunn (veid eller uveid), og kunnskapsstyrken for klassifiseringen vurderes derfor som god.
Villaksens genetiske status. Det er ikke observert genetisk endring grunnet innkryssing av rømt oppdrettslaks i to tredjedeler av de 24 undersøkte bestandene. Stor genetisk endring er observert i én bestand, og moderat genetisk endring er vist i den største bestanden (Numedalslågen). Villaksens genetiske status i området vurderes totalt sett til å være i moderat tilstand (svake genetiske endringer indikert). Vurderingen er basert på 24 bestander som til sammen utgjør 97 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som god.
Bestandenes robusthet for ny innkryssing. Tilstanden til villaksbestandene i området vurderes som god og selv om den genetiske statusen vurderes som moderat, vurderes bestandenes robusthet mot innkryssing totalt sett som god. Høy tetthet og konkurranse på gyteplassen antas å gi den rømte oppdrettslaksen lav gytesuksess og vektes derfor tyngre enn den genetiske statusen. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO1. Det antas å være lav sannsynlighet for forekomst av rømt oppdrettslaks på gyteplassene i området og bestandenes robusthet mot ny innkryssing vurderes som god. Totalt sett vurderes derfor villaksbestander i PO1 å ha lav sannsynlighet for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks. I 2020 ble det observert to mindre vassdrag med høyt innslag av rømt laks. Selv om det ikke ble observert vassdrag med høye innslag i 2021, så tyder moderat genetisk status på villaksen i området på at det tidligere har skjedd innkryssing av rømt laks i området. Det er foreløpig en svært begrenset oppdrettsproduksjon i området, men det er noe usikkerhet knyttet til påvirkning fra rømming i andre produksjonsområder med større produksjon. Det er også noe usikkerhet knyttet til hvor mye rømt oppdrettslaks det er på gyteplassene, siden få elver i området er dekket av overvåkningsprogrammet. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
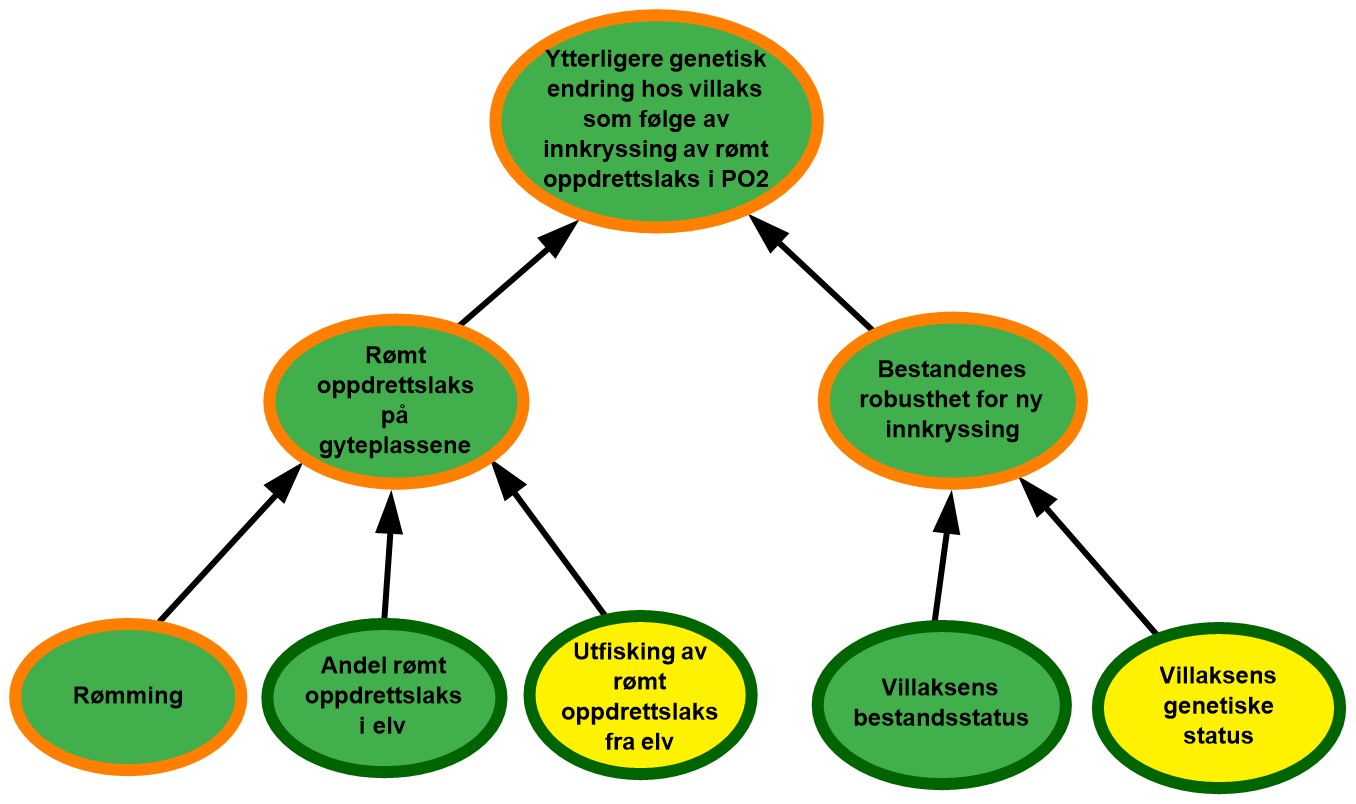
4.2. Produksjonsområde 2, Ryfylke
Figur 4.2. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 2 (PO2), Ryfylke.
Rømming. Det ble årlig rapportert fra 1 til 51 rømte oppdrettslaks i PO2 i perioden 2017–2021 med et årlig gjennomsnitt på 18. Området vurderes derfor å ha hatt lave rømmingstall i perioden. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. I området ble i gjennomsnitt 72 % av vassdragene (16 av totalt 22) overvåket årlig i perioden 2017-2021, noe som representerte 95 % av gytebestandsmålet i området. Innslaget av oppdrettslaks var lavt i 96 % av de undersøkte vassdragene, og ingen vassdrag hadde et høyt innslag i perioden 2017–2021. Vi vurderer derfor at det er lav sannsynlighet for at det er mye rømt oppdrettslaks i elver i området. Et høyt antall vassdrag overvåkes og utgjør mesteparten av gytebestandsmålet i området, og kunnskapsstyrken vurderes derfor som god.
Utfisking av rømt oppdrettslaks fra elv. I perioden 2017 til 2021 ble det årlig gjennomført utfisking i 4-8 vassdrag i området. Det var ingen vassdrag med høyt innslag, og i alle tre vassdrag med middels innslag av rømt oppdrettslaks ble det gjennomført utfisking samme år. Effekten av utfiskingen for området vurderes som moderat ettersom utfisking kun gjennomføres i en begrenset tidsperiode, og det vurderes derfor at det er moderat sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. I PO2 er det lite rømming, lav andel rømt oppdrettslaks i elvene og moderat effekt av utfisking av rømt oppdrettslaks fra elv de siste fem årene. Det vurderes derfor å være lav sannsynlighet for å finne mye rømt oppdrettslaks på gyteplassene i området. Til tross for at det er god kunnskap om to av de tre underliggende faktorene (andel rømt oppdrettslaks i undersøkte elver og effekten av utfisking), finnes ingen fullstendig oversikt over hvor mye rømt oppdrettslaks det er på gyteplassene i alle vassdragene. Kunnskapsstyrken vurderes derfor som moderat.
Villaksens bestandsstatus. Gytebestandsmålet blir nådd og det høstbare overskuddet er høyt for de fleste vassdragene i produksjonsområdet. Klassifiseringen blir den samme uavhengig av hvilken klassifiseringsmåte man legger til grunn (veid eller uveid), og tilstanden for bestandsstatus i området vurderes som god. Vassdragene med full vurdering utgjør 94 % av det samlede gytebestandsmålet i produksjonsområdet, og kunnskapsstyrken vurderes derfor som god.
Villaksens genetiske status. I halvparten av bestandene i området er det ikke observert noen genetisk endring, inkludert de to med høyest gytebestandsmål. Stor genetisk endring er observert i to bestander og tilstanden for området er derfor vurdert totalt sett som moderat (svake genetiske endringer indikert). Vurderingen er basert på 16 bestander som til sammen utgjør 97 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som god.
Bestandenes robusthet for ny innkryssing. Tilstanden for villaksens bestandsstatus vurderes som god mens tilstanden for villaksens genetiske status vurderes som moderat. Totalt sett vurderes likevel tilstanden for bestandenes robusthet mot ny innkryssing som følge av rømt oppdrettslaks som god. Vi antar at høy tetthet og konkurranse på gyteplassen gir den rømte oppdrettslaksen lav gytesuksess i produksjonsområdet. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO2. Sannsynlighet for forekomst av rømt oppdrettslaks på gyteplassene vurderes å være lav i området og bestandenes robusthet vurderes som god. Totalt sett vurderes risiko for ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks som lav i PO2. Til tross for at vi har relativt god kunnskap om flere av de underliggende faktorene hver for seg, mangler det kunnskap knyttet til hvor mye rømt oppdrettslaks det er på gyteplassene og hvor robuste bestandene er for ny innkryssing. Det er også noe usikkerhet knyttet til påvirkning fra rømming i andre produksjonsområder der rømmingstallene er høyere. Kunnskapsstyrken vurderes derfor som moderat.
4.3. Produksjonsområde 3, Karmøy til Sotra
Figur 4.3. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 3 (PO3), Karmøy–Sotra.
Rømming. Det ble årlig rapportert fra 5 til 8 765 rømte oppdrettslaks i PO3 i perioden 2017–2021 med et årlig gjennomsnitt på 3 343. Området vurderes derfor å ha hatt rømming av moderat omfang i perioden. Området har i tidligere år hatt høye rømmingstall, men moderate rømmingstall de siste årene gjør at status for rømming justeres ned fra høy til moderat. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. I området ble i gjennomsnitt 87 % av vassdragene overvåket årlig (16 av totalt 18 vassdrag) i perioden 2017-2021, noe som representerer 93 % av gytebestandsmålet i området. 37 % av de undersøkte vassdragene i området har høye andeler av rømt oppdrettslaks, og det vurderes å være høy sannsynlighet for at det er mye rømt oppdrettslaks i elvene i produksjonsområdet. Et høyt antall elver overvåkes og store deler av gytebestandsmålet dekkes. Derfor vurderes kunnskapsstyrken som god.
Utfisking av rømt oppdrettslaks fra elv. Det er mye rømt oppdrettslaks i elvene i området, og dermed stort behov for utfisking (9-17 elver med utfisking årlig) i perioden 2017-2021. Utfisking ble gjennomført i 90 % (26 av 29) av vassdrag der en høy andel rømt laks ble observert samme år, samt i 69 % (9 av 13) av vassdrag med middels innslag samme år. Til sammen er 856 rømte oppdrettslaks fjernet og verifisert fra vassdragene i området i perioden 2017-2021. En stor andel er fanget i fiskefellen i Etne, der det er dokumentert at utfiskingen er svært effektiv. Likevel er det enkelte vassdrag i området hvor utfisking ikke fungerer optimalt på grunn av vannføring, forekomst av innsjø eller generelt vanskelige forhold. På bakgrunn av dette vurderes det at det er moderat sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. I PO3 er det høy sannsynlighet for rømming og for å få rømt oppdrettslaks i elvene. Utfisking er delvis effektiv. Det vurderes derfor å være høy sannsynlighet for å finne mye rømt oppdrettslaks på gyteplassene i enkelte av vassdragene i enkelte år. Det er god kunnskap både om utfisking av rømt oppdrettslaks fra elv og andel rømt oppdrettslaks i elv, og kunnskapsstyrken vurderes derfor som god.
Villaksens bestandsstatus. Gytebestandsmålet blir nådd for mange av vassdragene i produksjonsområdet. Det høstbare overskuddet er lavt, noe som gjør bestandene utsatt for å komme under gytebestandsmålene i framtiden. To av de større vassdragene i produksjonsområdet, Oselva og Etneelva, har imidlertid høyere høstbart overskudd, noe som trekker opp vurderingen veid med gytebestandsmål. At de små vassdragene har lite høstbart overskudd, trekker totalvurderingen ned og tilstanden for villfiskens bestandsstatus i området vurderes derfor som dårlig. Mange av vassdragene i produksjonsområdet er stengt for fiske, så vassdragene med full vurdering utgjør 69 % av det samlede gytebestandsmålet i produksjonsområdet. Kunnskapsstyrken vurderes totalt sett som god.
Villaksens genetiske status. Stor genetisk endring er påvist i 10 av 12 bestander, deriblant Etneelva. Kun én bestand er uten genetisk endring (Oselva). Villaksens genetiske status i området vurderes til å være i dårlig tilstand (store eller moderate genetiske endringer påvist). Vurderingen er basert på 12 bestander som til sammen utgjør 99 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som god.
Bestandenes robusthet mot innkryssing. Tilstanden for både villaksens bestandsstatus og genetisk status vurderes å være dårlig, og tilstanden for bestandenes robusthet mot innkryssing vurderes derfor som dårlig. Lav konkurranse på gyteplassen kan gi den rømte oppdrettslaksen høyere gytesuksess i produksjonsområdet. Kunnskapen knyttet til villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO3. Høy sannsynlighet for rømt oppdrettslaks på gyteplassene kombinert med dårlig robusthet mot innkryssing gjør at PO3 fortsatt vurderes å ha høy risiko for ytterligere genetisk endring hos villaksen som følge av innkryssing av rømt oppdrettslaks. Denne konklusjonen støttes av at tilstanden for villaks i PO3 har vært svak så lenge at regjeringen i 2013 bestemte at truete villaksbestander i Hardangerfjorden skulle tas vare på i levende genbank for laks. Innsamling av rogn og melke i elvene i regionen startet høsten 2015. Det ble gjort avtaler for midlertidig oppbevaring av dette materialet fra 2017, og sommeren 2020 ble et bygg for levende genbank ferdigstilt. Sju laksestammer fra Hardangerfjorden er lagt inn i den nye genbanken på Ims, og ytterligere to bestander fra regionen er i genbank andre steder. Det er god kunnskap om alle underliggende faktorer utenom rømming, og selv om det knyttes noe usikkerhet til bestandenes robusthet, vurderes kunnskapsstyrken som god.
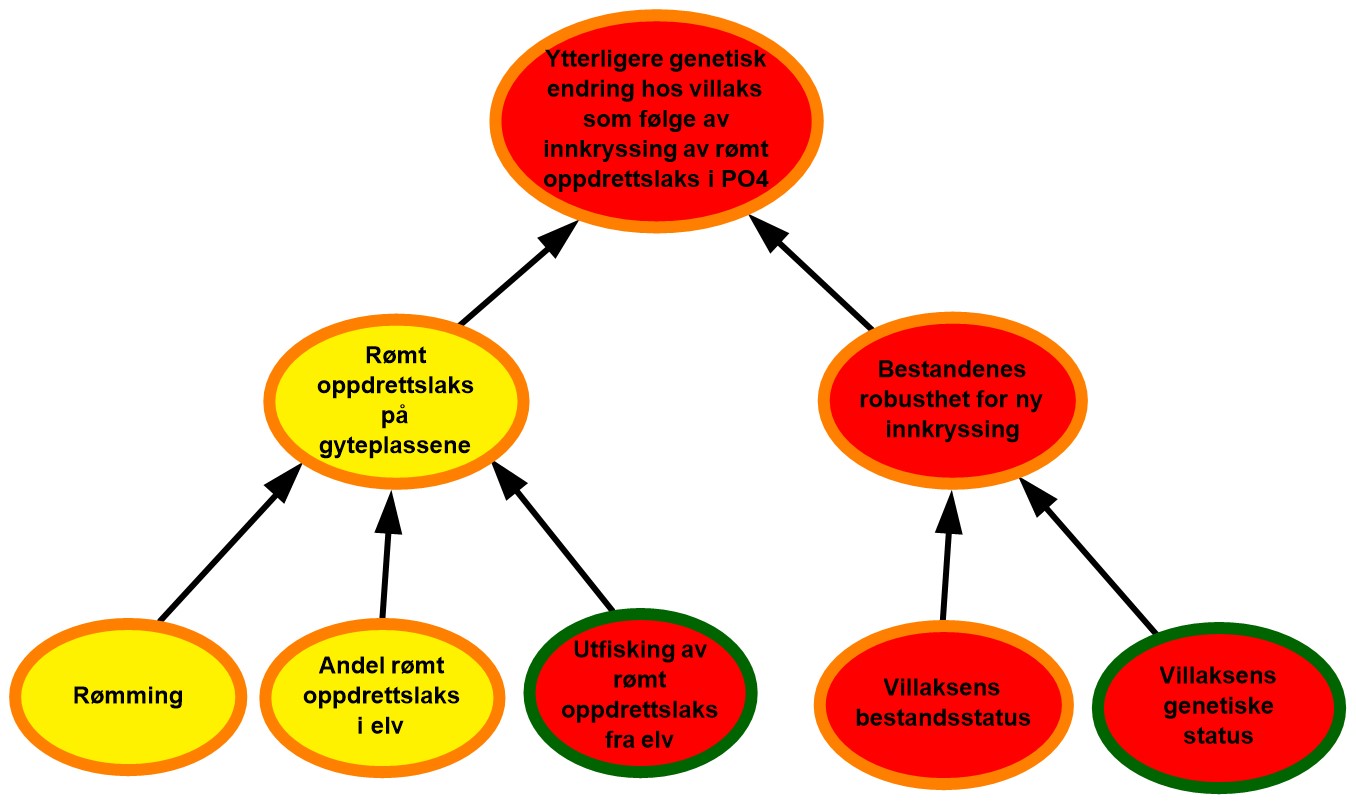
4.4. Produksjonsområde 4, Nordhordland til Stadt
Figur 4.4. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 4 (PO4), Nordhordland til Stadt.
Rømming. Det ble årlig rapportert fra 15 til 17 256 rømte oppdrettslaks i PO4 i perioden 2017–2021 med et årlig gjennomsnitt på 4 851. Området vurderes derfor å ha hatt moderate rømmingstall i perioden. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. I området ble i gjennomsnitt 71 % av vassdragene (31 av 43) overvåket årlig i perioden 2017-2021, som representerte 78 % av det samlede gytebestandsmålet. Det er til dels mye rømt oppdrettslaks i elvene i området (17 % og 5 % av undersøkte elver med henholdsvis moderat og høyt innslag). Basert på dette vurderes det derfor å være moderat sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Både andel elver som overvåkes og dekning av gytebestandsmålet i området er noe redusert siden forrige vurdering og kunnskapsstyrken justeres fra god til moderat for området.
Utfisking av rømt oppdrettslaks fra elv. Utfisking er gjennomført i 12-18 vassdrag årlig i perioden 2017 til 2021. Av vassdrag med høyt og middels innslag av rømt laks ble det samme år gjennomført utfisking i gjennomsnitt 71 % (5 av 7) og 58 % (15 av 26) av vassdragene. Effekten av utfisking og tilstanden for området ansees som dårlig, ettersom det er elver med høyt og middels innslag hvor det ikke har vært utfisking. I tillegg er det vassdrag i området hvor utfisking ikke vil fungere optimalt på grunn av stor vannføring, innsjøer eller generelt vanskelige forhold. Det vurderes derfor at det er høy sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. I PO4 er det moderat med rømming, moderat sannsynlighet for å få rømt oppdrettslaks i elvene, samtidig som at utfisket er lite effektivt. Det vurderes å være moderat sannsynlighet for å finne mye rømt oppdrettslaks på gyteplassene. Til tross for at det er god kunnskap om to av de tre underliggende faktorene (andel rømt oppdrettslaks i undersøkte elver og effekten av utfisking), finnes ingen fullstendig oversikt over hvor mye rømt oppdrettslaks det er på gyteplassene i alle vassdragene. Kunnskapsstyrken vurderes derfor som moderat.
Villaksens genetiske status. Gytebestandsmålet blir nådd for de fleste vassdragene i produksjonsområdet. Noen av de større vassdragene (Lærdalselva, Jølstra og Nausta) har lavt høstbart overskudd, noe som trekker vurderingen ned dersom vi veier med gytebestandsmål. Lærdalselva ble stengt for fiske i 2021. Mange av vassdragene i Sognefjorden har hatt lite eller ikke noe høstbart overskudd de seinere årene i vurderingsperioden. Samlet vurdering for området er derfor dårlig tilstand. Vassdragene med full vurdering utgjør 80 % av det samlede gytebestandsmålet i produksjonsområdet. Selv om dette er mye, varierer den samlede vurderingen avhengig av om vassdragene blir veid med gytebestandsmål eller ikke, og det er betydelig variasjon i vurderingen mellom vassdragene i regionen. Kunnskapsstyrken vurderes derfor fortsatt til å være moderat.
Villaksens genetiske status. Stor genetisk endring er påvist i nær halvparten av bestandene. Store eller moderate endringer er påvist i de tre største bestandene. Villaksens genetiske status i området vurderes til å være i dårlig tilstand (store eller moderate genetiske endringer påvist). Vurderingen er basert på 33 bestander som til sammen utgjør 94 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som god.
Bestandenes robusthet mot innkryssing. Tilstanden for både villaksens bestandsstatus og genetisk status vurderes å være dårlig, og tilstanden for bestandenes robusthet mot innkryssing vurderes derfor som dårlig. Lav konkurranse på gyteplassen kan gi den rømte oppdrettslaksen høyere gytesuksess i produksjonsområdet. Kunnskapen knyttet til villaksens bestandsstatus vurderes som moderat, mens kunnskapen knyttet til genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er også begrenset og vurderes derfor som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO4. Det er moderat sannsynlighet for forekomst av rømt oppdrettslaks på gyteplassene og robustheten vurderes som dårlig for området. Selv om andel rømt oppdrettslaks på gyteplassene er kategorisert som moderat, er risikoen for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks vurdert til å være høy. Dette skyldes den dårlige robustheten av bestandene i området i kombinasjon med at det de tre siste årene har vært elver med høye innslag av rømt oppdrettslaks. Til tross for at vi har relativt god kunnskap om flere av de underliggende faktorene hver for seg, mangler det kunnskap knyttet til hvor mye rømt oppdrettslaks det er på gyteplassene og hvor robuste bestandene er for ny innkryssing. Kunnskapsstyrken vurderes derfor som moderat.
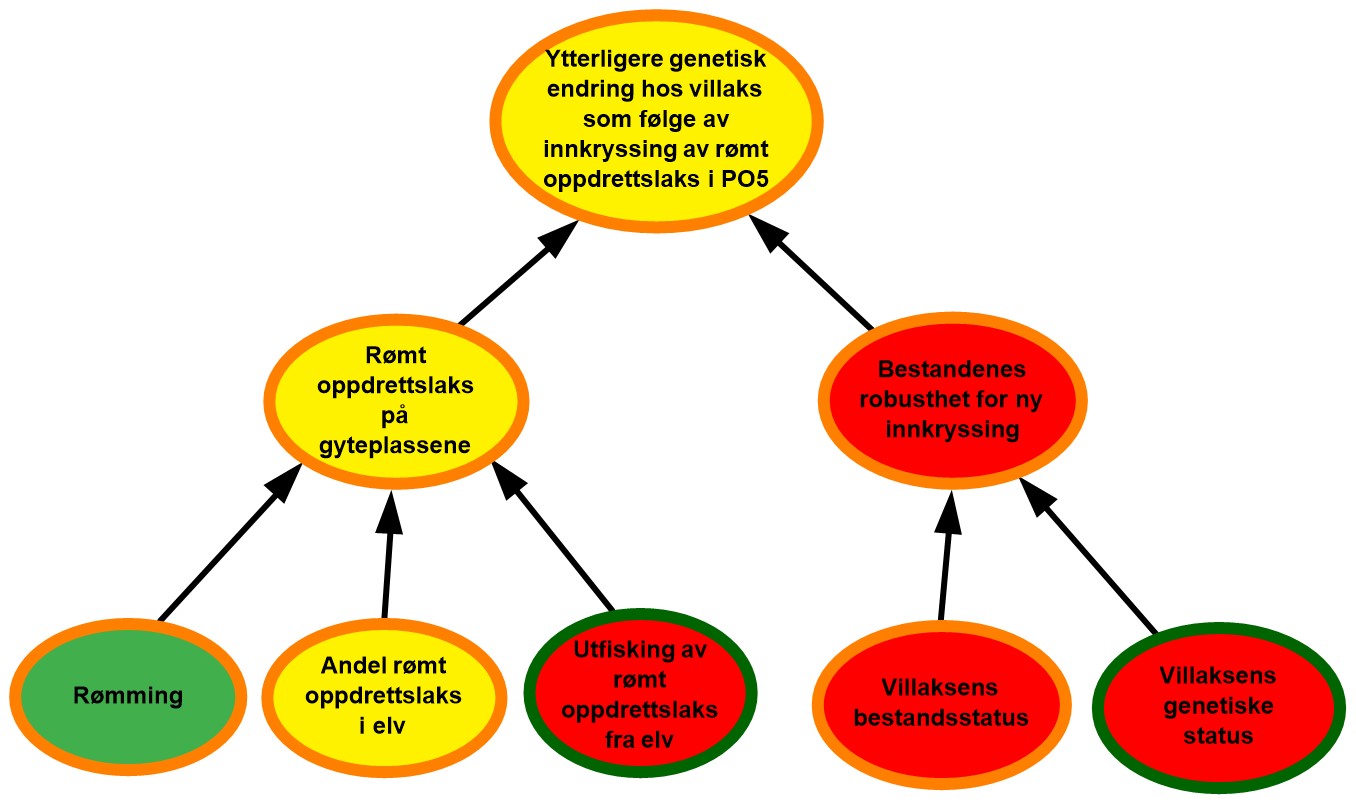
4.5. Produksjonsområde 5, Stadt til Hustadvika
Figur 4.5. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 5 (PO5), Stadt–Hustadvika.
Rømming. Det ble årlig rapportert fra 1 til 2200 rømte oppdrettslaks i PO5 i perioden 2017–2021 med et årlig gjennomsnitt på 825. Området vurderes derfor å ha hatt lave rømmingstall i perioden og tilhørende lav sannsynlighet for rømming. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. Det var i snitt 47 % av vassdragene i området (22 av 46) som inngikk i overvåkningsprogrammet, noe som representerte 66 % av gytebestandsmålet. Totalt plasseres 84 %, 13 % og 3 % av vurderte vassdrag henholdsvis i kategorien lavt, moderat og høyt innslag av rømt oppdrettslaks. Det var ingen vassdrag med høyt innslag av rømt oppdrettslaks de to siste årene (2020-221). Basert på dette vurderes det derfor å være moderat sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Basert på den lave andelen vassdrag som er vurdert i området samt noe lav dekningsgrad på gytebestandsmålet, vurderes kunnskapsstyrken som moderat.
Utfisking av rømt oppdrettslaks fra elv. Utfisking er gjennomført årlig i 4-8 vassdrag i området i perioden 2017 til 2021. Andelen vassdrag med høyt eller middels innslag av rømt oppdrettslaks har ligget lavt i dette området, men var noe høyere i 2019. I 2021 var det ingen vassdrag med høy andel, og ett vassdrag med moderat andel rømt laks. Av vassdrag med høyt og middels innslag av rømt laks ble det samme år gjennomført utfisking i gjennomsnitt 67 % (2 av 3) og 29 % (4 av 14) av vassdragene. Effekten av utfiskingen vurderes som dårlig, ettersom det er elver med høyt og middels innslag hvor det ikke har vært utfisking. Det vurderes derfor at det er høy sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. Det har totalt sett vært lite rømming i området og lav sannsynlighet for å finne mye rømt oppdrettslaks i elvene. Effektiviteten av utfisking av rømt oppdrettslaks er lav i dette området. Det vurderes totalt sett å være moderat sannsynlighet for å finne mye rømt oppdrettslaks på gyteplassene i PO5. Det er moderat kunnskap både om rømming og andel rømt oppdrettslaks i elv og god kunnskap om antallet oppdrettslaks som er fjernet fra elvene under utfisking, og kunnskapsstyrken vurderes totalt sett som moderat.
Villaksens bestandsstatus. Gytebestandsmålet blir nådd for de fleste vassdragene i produksjonsområdet. Det høstbare overskuddet er ytterligere redusert eller nesten borte i flere vassdrag i den sørlige delen av produksjonsområdet, spesielt de siste årene, og den samlede vurderingen justeres fra moderat til dårlig tilstand i området. Vassdragene i Raumaregionen er under reetablering etter behandling mot Gyrodactylus salaris, og er derfor ikke gitt en full vurdering ennå. Vassdragene med full vurdering utgjør kun 55 % av det samlede gytebestandsmålet i produksjonsområdet. Dette skyldes i hovedsak at vassdragene i Raumaregionen ikke blir vurdert fordi de er under oppbygging. Tilstanden blir nå vurdert likt avhengig av om vassdragene blir veid med gytebestandsmål eller ikke, med kunnskapsstyrken er fortsatt moderat for området siden vurderingen kun dekker i overkant av halvparten av det samlede gytebestandsmålet i området.
Villaksens genetiske status. Stor genetisk endring er påvist i nær en fjerdedel av bestandene, inkludert de største bestandene. Villaksens genetiske status i området vurderes til å være i dårlig tilstand (store eller moderate genetiske endringer påvist). Vurderingen er basert på 27 bestander som til sammen utgjør 86 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som god.
Bestandenes robusthet mot innkryssing. Tilstanden for bestandsstatus blir vurdert som dårlig, samtidig som genetisk status også vurderes som dårlig. Den totale vurderingen av tilstanden for bestandenes robusthet mot innkryssing justeres derfor fra moderat til dårlig. Denne justeringen samsvarer med forrige risikovurdering hvor vi påpekte at bestandsstatus var blitt svekket de siste årene i dette området, og at en videre utvikling i samme retning kunne føre til at bestandenes robusthet i området også blir svekket på sikt, slik vi nå ser. Det vurderes derfor å være høy sannsynlighet for redusert robusthet hos villbestandene i området i tiden fremover. Bestandene i Raumaregionen er ikke tatt med i vurderingen, men høy genetisk påvirkning ble funnet i et ungfiskmateriale fra Rauma i 2017.
Ytterligere genetisk endring hos villaks som følge av rømt innkryssing av oppdrettslaks i PO5. Det antas å være moderat sannsynlighet for forekomst av rømt oppdrettslaks på gyteplassene i området. Bestandenes robusthet mot innkryssing er svekket siden forrige vurdering, og vurderes nå som dårlig. Utviklingen de siste årene (2020-2021) viser at ingen vassdrag har hatt høye forekomster av rømt oppdrettslaks de siste to årene, og risikoen for ytterligere genetisk endring i PO5 som følge av innkryssing av rømt oppdrettslaks vurderes fortsatt som moderat. Til tross for at vi har relativt god kunnskap om noen av de underliggende faktorene hver for seg, så mangler det kunnskap knyttet til hvor mye rømt oppdrettslaks det er på gyteplassene ettersom få elver i området er dekket av overvåkningsprogrammet. Det mangler også kunnskap knyttet til hvor robuste bestandene er for ny innkryssing. Kunnskapsstyrken vurderes derfor som moderat.
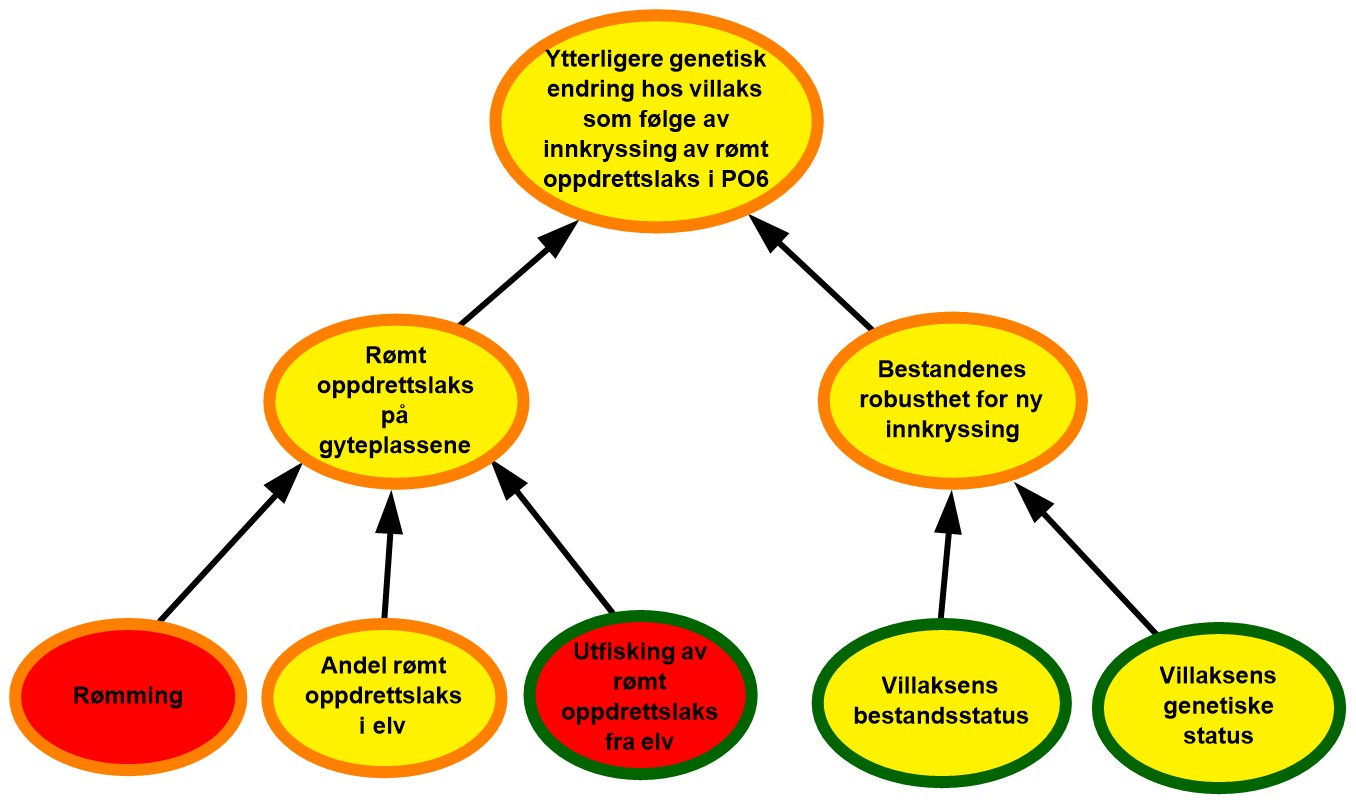
4.6. Produksjonsområde 6, Nordmøre og Sør-Trøndelag
Figur 4.6. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 6 (PO6), Nordmøre til Sør-Trøndelag.
Rømming. Det ble årlig rapportert fra 893 til 28 481 rømte oppdrettslaks i PO6 i perioden 2017–2021 med et årlig gjennomsnitt på 11 862. Området vurderes derfor å ha mye rømming. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. Det var i snitt 24 % av vassdragene (18 av 76) i området som ble overvåket årlig, noe som utgjorde 89 % av det samlede gytebestandsmålet i området. Av alle vurderte vassdrag plasseres 83 %, 14 % og 3 % i kategorien lavt, moderat og høyt innslag av rømt oppdrettslaks. Basert på dette vurderes det derfor å være moderat sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. En lav andel vassdrag er overvåket og selv om dette dekker storparten av gytebestandsmålet i området, kan det være flere vassdrag som kan ha moderat eller høyt innslag av rømt oppdrettslaks uten at dette oppdages. Kunnskapsstyrken vurderes derfor som moderat.
Utfisking av rømt oppdrettslaks fra elv. Antallet elver hvor utfisking er gjennomført har variert fra 2-4 i perioden 2017 til 2021. Av vassdrag med høyt og middels innslag av rømt laks ble det samme år gjennomført utfisking i gjennomsnitt 67 % (2 av 3) og 62 % (8 av 13) av vassdragene, men det er kun tatt ut 30 oppdrettslaks i hele perioden. Effekten av utfiskingen i området vurderes derfor som dårlig, og at det er høy sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. Det er mye rømming i området, og sannsynlighet for at det er mye rømt laks i elvene i området er moderat. Utfisket er lite effektivt i dette området. På grunn av en fortsatt rapportering fra overvåkningsprogrammet om moderat andel rømt oppdrettslaks i elv, vurderes det fortsatt å være moderat sannsynlighet for å finne mye rømt oppdrettslaks på gyteplassene i PO6. Til tross for at det er god kunnskap om utfiskingen i området, er det moderat kunnskap om hvor mye oppdrettslaks som rømmer og hvor stor andel av disse som går opp i elvene. Det finnes heller ingen fullstendig oversikt over hvor mye rømt oppdrettslaks det er på gyteplassene i alle vassdragene. Kunnskapsstyrken vurderes derfor som moderat.
Villaksens bestandsstatus. Gytebestandsmålet blir nådd for mange av vassdragene i produksjonsområdet, likevel har noen av de større vassdragene i produksjonsområdet ikke nådd gytebestandsmålet i enkelte av de siste årene (Orkla og Gaula). Det høstbare overskuddet har også vært lavt, men har bedret seg de senere årene i vurderingsperioden. Vassdragene i Drivaregionen er fortsatt infisert av Gyrodactylus salaris. Den totale vurderingen for området er fortsatt moderat tilstand. Vassdragene med full vurdering utgjør 84 % av det samlede gytebestandsmålet i produksjonsområdet. Tilstanden blir vurdert likt avhengig av om vassdragene blir veid med gytebestandsmål eller ikke, så kunnskapsstyrken er god.
Villaksens genetiske status. Stor genetisk endring er påvist i nær en tredjedel av bestandene, mens i de største bestandene er det ikke observert (eller kun funnet indikasjoner på) genetisk endring. Villaksens genetiske status i området vurderes til å være i moderat tilstand (svake genetiske endringer indikert). Vurderingen er basert på 29 bestander som til sammen utgjør 96 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som god.
Bestandenes robusthet mot innkryssing. Både genetisk status og villaksens bestandsstatus vurderes å ha moderat tilstand. Vurderingen av bestandenes robusthet mot ny innkryssing vurderes derfor som moderat tilstand i PO6. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av rømt innkryssing av oppdrettslaks i PO6. Det vurderes å være moderat sannsynlighet for tilstedeværelse av rømt oppdrettslaks på gyteplassene og robustheten av populasjonene i området vurderes som moderat. Risikoen for ytterligere genetisk endring hos villaksbestandene som følge av innkryssing av rømt oppdrettslaks vurderes derfor som moderat i PO6. Til tross for at vi har relativt god kunnskap om tre av de underliggende faktorene så mangler det kunnskap knyttet til hvor mye rømt oppdrettslaks det er på gyteplassene, siden få elver i området er dekket av overvåkningsprogrammet. Det mangler også kunnskap om hvor robuste bestandene er for ny innkryssing. Kunnskapsstyrken vurderes derfor som moderat.
4.7. Produksjonsområde 7, Nord-Trøndelag med Bindal
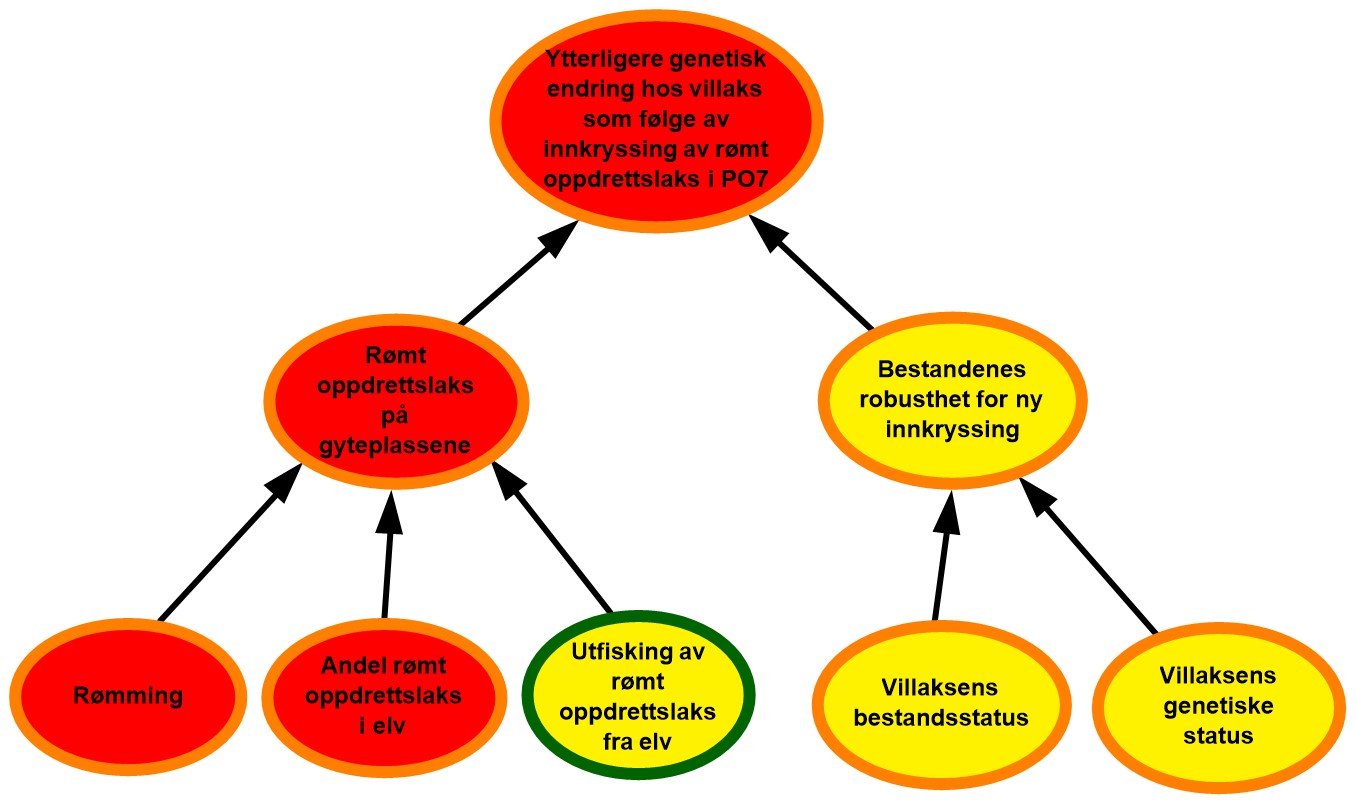
Figur 4.7. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 7 (PO7), Nord-Trøndelag med Bindal.
Rømming. Det ble årlig rapportert om 0 til 107 635 rømte oppdrettslaks i PO7 i perioden 2017–2021 med et årlig gjennomsnitt på 41 025. Området vurderes derfor å ha hatt høye rømmingstall. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. Det var i snitt 37 % av vassdragene i området (9 av 24) som ble overvåket årlig, noe som representerte 91 % av gytebestandsmålet i området. Totalt falt 68 %, 9 % og 23 % av vurderte vassdrag i kategorien lavt, moderat og høyt innslag av rømt oppdrettslaks. Basert på dette vurderes det derfor å være høy sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Overvåkningsprogrammet dekker bare i overkant av en tredjedel av vassdragene i området, og selv om dette utgjør en høy andel av gytebestandsmålet, så vurderes kunnskapsstyrken som moderat.
Utfisking av rømt oppdrettslaks fra elv. Utfisking er gjennomført årlig i 3-5 vassdrag i området i perioden 2017 til 2021. Av vassdrag med høyt og middels innslag av rømt laks ble det samme år gjennomført utfisking i gjennomsnitt 100 % (10 av 10) og 75 % (3 av 4) av vassdragene, og det ble tatt ut 284 oppdrettslaks i hele perioden. Effekten av utfiskingen for området vurderes som moderat ettersom utfisking kun gjennomføres i en begrenset tidsperiode, og det vurderes derfor at det er moderat sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. Det er rapportert mye rømming i området og andel rømt oppdrettslaks i elvene er høy. Samtidig er utfisket delvis effektivt i området. Det vurderes derfor å være høy sannsynlighet for å fortsatt finne mye rømt oppdrettslaks på gyteplassene i PO7 og tilstanden for området vurderes som dårlig. Kunnskapsstyrken vurderes som god knyttet til utfisket, mens det er moderat kunnskap om rømming og andel rømt oppdrettslaks i elv da det er elver som ikke dekkes av overvåkningsprogrammet. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
Villaksens bestandsstatus. Gytebestandsmålet blir nådd for de fleste vassdragene i regionen, men noen av vassdragene har et redusert høstbart overskudd. Den største bestanden i produksjonsområdet (Namsenvassdraget) har også et relativt stort høstbart overskudd, derfor blir vurderingen ulik om man veier med gytebestandsmål eller ikke. De fleste vassdragene med forenklet vurdering i produksjonsområdet har moderat vurdering. Samlet sett vurderes derfor tilstanden som moderat. Vassdragene med full vurdering utgjør 96 % av det samlede gytebestandsmålet i produksjonsområdet. Vurderingen er avhengig av om det veies med gytebestandsmål eller ikke og kunnskapsstyrken vurderes derfor som moderat.
Villaksens genetiske status. Moderate genetiske endringer er påvist i den desidert største bestanden, Namsen, mens tre andre bestander viser ingen observert endring og én bestand viser store endringer. Villaksens genetiske status i området vurderes til å være i moderat tilstand, Tilstanden i området vurderes derfor som moderat. Vurderingen er basert på kun 6 av 24 bestander, men disse utgjør 92 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som moderat.
Bestandenes robusthet mot innkryssing. Både genetisk status og villaksens bestandsstatus vurderes å ha moderat tilstand og bestandenes robusthet mot innkryssing vurderes derfor også til å ha moderat tilstand. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som moderat. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er også begrenset og kunnskapsstyrken vurderes som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO7. Det antas å være høy sannsynlighet for forekomst av rømt oppdrettslaks på gyteplassene i området, og selv om robustheten vurderes som moderat vurderes risikoen for ytterligere genetisk endring grunnet innkryssing av rømt oppdrettslaks som høy for PO7. Det er moderat usikkerhet knyttet til de fleste av de underliggende faktorene for både rømt oppdrettslaks på gyteplassene og hvor robust bestandene er for ny innkryssing. Kunnskapsstyrken vurderes derfor som moderat.
4.8. Produksjonsområde 8, Helgeland til Bodø
Figur 4.8. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 8 (PO8), Helgeland til Bodø.
Rømming. Det ble årlig rapportert fra 18 til 7 032 rømte oppdrettslaks i PO8 i perioden 2017–2021 med et årlig gjennomsnitt på 2 327. Området vurderes derfor å ha moderat rømming i perioden. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. I gjennomsnitt ble 54 % av vassdragene (16 av 30) i området overvåket årlig i perioden 2017-2021, noe som utgjorde 87 % av gytebestandsmålet i området. Totalt falt 72 %, 19 % og 10 % av vurderte vassdrag i kategorien lavt, moderat og høyt innslag av rømt oppdrettslaks. Elver med høy andel rømt oppdrettslaks i denne perioden utgjorde totalt 17% av det vurderte gytebestandsmålet i området. Andelen vassdrag med observert høyt innslag av rømt oppdrettslaks har dermed økt sammenlignet med overvåkningsperioden 2016-2020. Andel rømt oppdrettslaks observert i elv justeres derfor fra moderat til høy. Basert på dette vurderes det derfor å være høy sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Overvåkningsprogrammet dekker kun litt over halvparten av elvene i området, og selv om dette dekker en høy andel av gytebestandsmålet i området, så vurderes kunnskapsstyrken som moderat.
Utfisking av rømt oppdrettslaks fra elv. Utfisking er gjennomført årlig i 3-7 vassdrag i området i perioden 2017 til 2021. Av vassdrag med høyt og middels innslag av rømt laks ble det samme år gjennomført utfisking i gjennomsnitt 38 % (3 av 8) og 40 % (6 av 15) av vassdragene, og det ble tatt ut 150 oppdrettslaks i hele perioden. Effekten av utfiskingen for området vurderes som dårlig og det vurderes derfor at det er høy sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. Nivå på rapporterte rømmingstall er moderat, mens andel rømt oppdrettslaks i elv er justert fra moderat til høy. Utfisking har lav effekt. Basert på dette vurderes det derfor at sannsynlighet for at det er mye rømt oppdrettslaks på gyteplassene i PO8 justeres fra moderat til høy. Kunnskapsstyrken vurderes som god knyttet til utfisket, mens det er moderat kunnskap om rømming og andel rømt oppdrettslaks i elv. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
Villaksens bestandsstatus. Gytebestandsmålene blir nådd for mange av vassdragene i produksjonsområdet. Det høstbare overskuddet er imidlertid lavt i mange vassdrag, noe som gjør dem sårbare for at de kan komme under gytebestandsmålene i framtiden. Ni vassdrag i regionen er enten under reetablering- eller under friskmelding etter behandling mot Gyrodactylus salaris. Disse vassdragene er ikke gitt noen vurdering. Den samlede vurderingen gir derfor dårlig tilstand. Vassdragene med full vurdering utgjør bare 39 % av det samlede gytebestandsmålet i regionen og tilstanden blir i tillegg vurdert ulikt avhengig av om vassdragene blir veid med gytebestandsmål eller ikke. Kunnskapsstyrken vurderes derfor til å være moderat.
Villaksens genetiske status. Stor genetisk endring er påvist i nær halvparten av bestandene, inkludert de fleste store bestandene. Villaksens genetiske status i området vurderes til å være i dårlig tilstand (store eller moderate genetiske endringer påvist). Vurderingen er basert på 11 av 30 bestander som samlet utgjør 81 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som moderat.
Bestandenes robusthet mot innkryssing. Tilstanden for både villaksens bestandsstatus og genetisk status vurderes å være dårlig, og tilstanden for bestandenes robusthet mot innkryssing vurderes derfor som dårlig. Lav konkurranse på gyteplassen kan gi den rømte oppdrettslaksen høyere gytesuksess i produksjonsområdet. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO8. Når det tidligere vært vurdert å være moderate mengder rømt oppdrettslaks på gyteplassene, har den gjennomgående dårlige robustheten hos villaksbestandene veid tyngre. I årets vurdering er sannsynlighet for tilstedeværelse av rømt oppdrettslaks på gyteplassene justert opp til høy, og risikoen for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks vurderes derfor fortsatt som høy i PO8. Det er moderat usikkerhet knyttet til de fleste av de underliggende faktorene for både rømt oppdrettslaks på gyteplassene og hvor robust bestandene er for ny innkryssing. Kunnskapsstyrken vurderes derfor som moderat.
4.9. Produksjonsområde 9, Vestfjorden og Vesterålen
Figur 4.9. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 9 (PO9), Vestfjorden til Vesterålen.
Rømming. Det ble årlig rapportert fra 3 til 20 480 rømte oppdrettslaks i PO9 i perioden 2017–2021 med et årlig gjennomsnitt på 4 372. Området vurderes derfor å ha hatt moderate rømmingstall i perioden. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. I gjennomsnitt ble 21 % av vassdragene i området (12 av 57) overvåket, noe som utgjorde 37 % av det samlede gytebestandsmålet i området. To tredjedeler av de vurderte vassdragene er vurdert til å ha lavt og 20 % til å ha moderat innslag. Andelen vurderte vassdrag med høyt innslag av rømt oppdrettslaks var 12 %. Basert på dette vurderes det derfor å være moderat sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Overvåkningsprogrammet dekker få elver i området og elvene dekker kun en tredjedel av gytebestandsmålet. Kunnskapsstyrken vurderes derfor som svak.
Utfisking av rømt oppdrettslaks fra elv. Utfisking er gjennomført årlig i 5-7 vassdrag i området i perioden 2017 til 2021. Av vassdrag med høyt og middels innslag av rømt laks ble det samme år gjennomført utfisking i gjennomsnitt 100 % (7 av 7) og 75 % (9 av 12) av vassdragene, og det ble tatt ut 101 oppdrettslaks i hele perioden. Det er få vassdrag i området som er dekket av overvåkningsprogrammet og det er vassdrag i området hvor utfisking ikke vil fungere optimalt på grunn av vannføring, forekomst av innsjø eller generelt vanskelige forhold. Effekten av utfisking vurderes av den grunn som lav, på tross av gjennomført utfisking i alle elver med høyt innslag. Det vurderes derfor at det er høy sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. Rømming er moderat, men det er vurdert å være høy sannsynlighet for at det er rømt oppdrettslaks i elvene. Utfisking er lite effektivt i området. Sannsynligheten for å finne mye rømt oppdrettslaks på gyteplassene vurderes derfor fortsatt som høy for PO9. Det er god kunnskap om utfiskingen i området, men det er moderat kunnskap om hvor mye oppdrettslaks som rømmer, og lite kunnskap om hvor stor andel av disse som går opp i elvene. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
Villaksens bestandsstatus. Gytebestandsmålene blir nådd for mange av vassdragene i produksjonsområdet. Det høstbare overskuddet er fortsatt lavt i en del av vassdragene og vurderingen av området er moderat tilstand. Vurderingen blir den samme enten man veier med gytebestandsmål eller ikke. De fleste småvassdragene som er gitt en forenklet vurdering har moderat eller dårlig status, antall vassdrag med god status ved forenklet vurdering har imidlertid økt fra ett ved forrige vurdering til syv. Vassdragene med full vurdering utgjør 65 % av det samlede gytebestandsmålet i regionen. Dette er et område med mange småvassdrag som har fått en forenklet vurdering. Det er imidlertid gytefisktellinger i mange av disse som ligger til grunn for vurderingen, slik at usikkerheten i de forenklede vurderingene er mindre enn i andre produksjonsområder. Kunnskapsstyrken vurderes derfor samlet sett som moderat.
Villaksens genetiske status. Ingen genetisk endring er observert i fire femtedeler av bestandene, inklusive den største bestanden. Det er påvist stor genetisk endring i to bestander, deriblant den nest største bestanden. Villaksens genetiske status i området vurderes til å være i moderat tilstand (svake genetiske endringer indikert). Vurderingen er basert på 20 bestander som til sammen utgjør 62 % av produksjonsområdets totale gytebestandsmål. Siden mange bestander ikke er vurdert, anses kunnskapsstyrken som moderat.
Bestandenes robusthet mot innkryssing. Tilstanden for bestandsstatus og genetisk status blir begge vurdert som moderat, og den totale vurderingen av bestandenes robusthet mot innkryssing vurderes derfor som moderat. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO9. Det antas å være stor sannsynlighet for at det kan forekomme en høy andel rømt oppdrettslaks på gyteplassene, samtidig som bestandenes robusthet for innkryssing er moderat. Risikoen for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks vurderes derfor som høy i PO9, ettersom den høye andelen av rømt oppdrettslaks veier tyngre. Det er derimot svært mange elver som ikke dekkes av overvåkningsprogrammet, som bidrar med usikkerhet til risikobildet. Det er moderat usikkerhet knyttet til de fleste resterende underliggende faktorene for både rømt oppdrettslaks på gyteplassene og hvor robuste bestandene er for ny innkryssing, og kunnskapsstyrken vurderes derfor som moderat.
4.10. Produksjonsområde 10, Andøya til Senja
Figur 4.10. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 10 (PO10), Andøya til Senja.
Rømming. Det ble årlig rapportert fra 2 til 182 417 rømte oppdrettslaks i PO10 i perioden 2017–2021 med et årlig gjennomsnitt på 37 525. Området vurderes derfor å ha hatt høye rømmingstall i perioden. Én enkelt rømmingsepisode på 179 491 ungfisk (13 g) fra et ferskvannsanlegg i juli 2019 påvirker sterkt de observerte rømmingstallene. I etterkant av denne rømmingsepisoden ble det rapportert om en gjenfangst på 104 753 (justert for gjenfangst, har området da hatt et årlig gjennomsnitt på 16 574 rapporterte rømte oppdrettslaks). Dødelighet etter rømming er større for oppdrettslaks som rømmer som ung, i forhold til oppdrettslaks som rømmer som kjønnsmoden eller nær kjønnsmoden, ettersom ungfisken må overleve lengre i naturen før den er kjønnsmoden og kan oppsøke gyteplassene.
Andel rømt oppdrettslaks i elv. I snitt ble 53 % av vassdragene i området (14 av 26) overvåket, noe som utgjorde 72 % av gytebestandsmålet i området. Andelen vurderte vassdrag med høyt og moderat innslag av rømt oppdrettslaks er henholdsvis 26 % og 7 %. I de to siste årene (2020 og 2021) er det ingen vassdrag med høyt innslag av rømt oppdrettslaks. Dette kan tyde på at overlevelse hos ungfisken som rømte i 2019 har vært lav, og/eller at den kan ha oppsøkt gyteplasser i andre områder, og/eller at den ikke har nådd kjønnsmoden alder enda. Ettersom det har vært en nedgang i antall vassdrag med høye og moderate andeler av rømt oppdrettslaks i området, og det i perioden 2017-2021 samlet sett er under 10 % av undersøkte vassdrag med høyt innslag, blir den totale vurderingen av observert andel rømt oppdrettslaks i elv justert fra dårlig til moderat tilstand i PO10. Basert på dette vurderes det derfor å være moderat sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Selv om i underkant av tre fjerdedeler av gytebestandsmålet er dekket av overvåkningsprogrammet, er andelen av vassdrag som overvåkes noe lav, og det er dermed vassdrag med ukjent innslag av rømt oppdrettslaks. Kunnskapsstyrken vurderes derfor som moderat.
Utfisking av rømt oppdrettslaks fra elv. Utfisking er gjennomført årlig i 3-16 vassdrag i området i perioden 2017 til 2021. Av vassdrag med høyt og middels innslag av rømt laks ble det samme år gjennomført utfisking i gjennomsnitt 80 % (4 av 5) og 83 % (15 av 18) av vassdragene, og det ble tatt ut 114 oppdrettslaks i hele perioden. Det er også vassdrag i området som ikke dekkes av overvåkningsprogrammet og/eller hvor utfiskingstiltak ikke kan gjennomføres på en optimal måte på grunn av vannføring, forekomst av innsjø eller andre kompliserende forhold. Effekten av utfiskingen for området vurderes som dårlig og det vurderes derfor at det er høy sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. De rapporterte rømmingstallene for området er høye, men i hovedsak knyttet til én enkel rømmingsepisode av ungfisk i 2019, med lav forventet overlevelse. Andel rømt oppdrettslaks i elv har gått ned, og sannsynlighet for forekomst av rømt oppdrettslaks i elver i området er justert fra høy til moderate. Utfisking i området er lite effektivt. Det vurderes derfor å være moderate sannsynlighet for å finne mye rømt oppdrettslaks på gyteplassene, en nedjustering av risikobildet fra forrige vurdering. Ettersom det er mange vassdrag som ikke inngår i overvåkningsprogrammet vurderes kunnskapsstyrken totalt sett som moderat.
Villaksens bestandsstatus. Gytebestandsmålene blir nådd for mange av vassdragene i produksjonsområdet, men det uveide høstbare overskuddet er lavt i enkelte vassdrag, noe som gjør at den samlede vurderingen av området er moderat tilstand. Det største vassdraget i regionen (Målselv) har normalt høstbart overskudd og trekker vurderingen opp når vi veier med gytebestandsmål. I det nest største vassdraget, Salangselva, er laksebestanden så lav at fiske etter laks har vært forbudt i perioden. Vassdragene som blir vurdert med forenklet vurdering, domineres av dårlig og moderat kvalitet. Det er stor spredning i tilstanden i vassdragene i regionen. Vassdragene med full vurdering utgjør 92 % av det samlede gytebestandsmålet i produksjonsområdet. Vurderingen er ulik om vi vekter med gytebestandsmål eller ikke og kunnskapsstyrken vurderes derfor som moderat.
Villaksens genetiske status. Stor genetisk endring er påvist i en tredjedel av bestandene, inklusive de to største bestandene (Målselva og Salangselva). Villaksens genetiske status i området vurderes til å være i dårlig tilstand (store eller moderate genetiske endringer påvist). Vurderingen er basert på 15 bestander som til sammen utgjør 92 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som god.
Bestandenes robusthet mot innkryssing. Tilstanden for villaksens bestandsstatus er moderat, samtidig som genetisk status vurderes å være dårlig. Tilstanden for bestandenes robusthet mot innkryssing vurderes å være moderat, ettersom bestandsstatus tillegges mer vekt enn genetisk status. Høy konkurranse på gyteplassen kan gi den rømte oppdrettslaksen lavere gytesuksess i produksjonsområdet. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO10. Observerte andeler rømt oppdrettslaks i elv er redusert gjennom vurderingsperioden og det antas å være moderat sannsynlighet for å finne mye rømt oppdrettslaks på gyteplassene. Det er rapportert om høye rømmingstall i området, som i all hovedsak er knyttet til en rømmingsepisode i 2019 hvor et stort antall ungfisk rømte fra et landanlegg, og hvor overlevelse etter rømming er antatt å være lav. Bestandenes robusthet for innkryssing er moderat. Totalt sett vurderes sannsynligheten for ytterligere genetisk endring i PO10 som følge av innkryssing av rømt oppdrettslaks som moderat, redusert fra høy ved forrige vurdering. Det er manglende kunnskap både om rømt oppdrettslaks på gyteplassene og bestandenes robusthet mot innkryssing. Kunnskapsstyrken vurderes derfor som moderat.
4.11. Produksjonsområde 11, Kvaløya til Loppa
Figur 4.11. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 11 (PO11), Kvaløya til Loppa.
Rømming. Det ble årlig rapportert om 0 til 641 rømte oppdrettslaks i PO11 i perioden 2017–2021 med et årlig gjennomsnitt på 130. Området vurderes derfor å ha hatt lave rømmingstall i perioden. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode. Det mangler også kunnskap knyttet til påvirkning fra rømming i andre produksjonsområder og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. I gjennomsnitt ble 53 % av vassdragene i området (11 av 20) overvåket i perioden 2017-2021, noe som utgjorde 90 % av det samlede gytebestandsmålet. Andelen undersøkte vassdrag med høyt innslag av rømt oppdrettslaks var 15 %. Basert på dette vurderes det derfor å være høy sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Selv om en stor andel av gytebestandsmålet er dekket av overvåkningen, er kun i overkant av halvparten av vassdragene vurdert, noe som betyr at en ikke kan utelukke at vassdrag som ikke er vurdert kan ha moderat og lavt innslag av rømt oppdrettslaks. Kunnskapsstyrken vurderes derfor som moderat.
Utfisking av rømt oppdrettslaks fra elv. Utfisking er gjennomført årlig i 2-9 vassdrag i området i perioden 2017 til 2021. Av vassdrag med høyt og middels innslag av rømt oppdrettslaks ble det samme år gjennomført utfisking i gjennomsnitt 88 % (7 av 8) av vassdragene henholdsvis, og det ble fjernet 84 oppdrettslaks i hele perioden. Det er vassdrag i området der utfisking ikke fungerer optimalt på grunn av vannføring, forekomst av innsjø eller andre vanskelige forhold. Effekten av utfiskingen vurderes av den grunn som dårlig og at det er høy sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. Rapportert rømming i området er lav, mens andel rømt oppdrettslaks i undersøkte elver er høy. Effekten av utfiskingstiltak anses å være lav. Det vurderes derfor at det vil være høy sannsynlighet for å finne mye rømt oppdrettslaks på gyteplassene i området. Det kan synes som om rømt oppdrettslaks fra nærliggende produksjonsområder går opp i elvene i dette området, siden rapportert rømming i området er lav. Til tross for at det er god kunnskap om utfiskingen i området, er det moderat kunnskap om hvor mye oppdrettslaks som rømmer og hvor stor andel av disse som går opp i elvene. Kunnskapsstyrken vurderes derfor som moderat.
Villaksens bestandsstatus. Gytebestandsmålene blir nådd for mange av vassdragene i produksjonsområdet. Det høstbare overskuddet er imidlertid lavt i mange vassdrag. Det største vassdraget i produksjonsområdet (Reisaelva) har dårligere status enn de små. Vassdragene i Skibotnregionen er under reetablering etter behandling mot Gyrodactylus salaris og blir derfor ikke gitt noen full vurdering. Samlet vurderes tilstanden som dårlig for området. Vassdragene med full vurdering utgjør 61 % av det samlede gytebestandsmålet i regionen. Vurderingen blir den samme enten vi veier med gytebestandsmål eller ikke og kunnskapsstyrken vurderes derfor som god.
Villaksens genetiske status. Store genetiske endringer er påvist i to tredjedeler av bestandene, inklusive flere store bestander. I den største bestanden er det ikke observert genetiske endringer. Villaksens genetiske status i området vurderes til å være i dårlig tilstand (store eller moderate genetiske endringer påvist). Vurderingen er basert på ni bestander som til sammen utgjør 88 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som god.
Bestandenes robusthet mot innkryssing. Tilstanden for både villaksens bestandsstatus og genetisk status vurderes å være dårlig, og tilstanden for bestandenes robusthet mot innkryssing vurderes derfor som dårlig. Lav konkurranse på gyteplassen kan gi den rømte oppdrettslaksen høyere gytesuksess i produksjonsområdet. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO11. Det er høy sannsynlighet for at det er mye rømt oppdrettslaks på gyteplassene i området, og bestandenes robusthet for innkryssing er vurdert som dårlig. Risikoen for ytterligere genetisk endring i PO11 som følge av innkryssing av rømt oppdrettslaks vurderes derfor som høy. Det mangler kunnskap knyttet til hvor mye rømt oppdrettslaks det er på gyteplassene da overvåkningsprogrammet ikke dekker alle elver i området, og det er usikkerhet knyttet til rømmingstallene. Det er også usikkerhet knyttet til hvor robuste bestandene er for ny innkryssing. Kunnskapsstyrken vurderes derfor totalt sett som moderat.
4.12. Produksjonsområde 12, Vest-Finnmark
Figur 4.12. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 12 (PO12), Vest-Finnmark.
Rømming. Det ble årlig rapportert fra 0 til 26 813 rømte oppdrettslaks i PO12 i perioden 2017–2021 med et årlig gjennomsnitt på 8 324. PO12 vurderes derfor å ha hatt moderate rømmingstall i perioden. De rapporterte rømmingstallene er heftet med usikkerhet, både med tanke på antall rømmingsepisoder og antall rømt oppdrettslaks per episode, og kunnskapsstyrken vurderes derfor som moderat.
Andel rømt oppdrettslaks i elv. I snitt ble 37 % av vassdragene i området (10 av 26) overvåket i perioden 2017-2021. Disse utgjorde 87 % av det samlede gytebestandsmålet i området. Totalt falt 83 %, 17 % og 0 % av vurderte vassdrag i kategorien lavt, moderat og høyt innslag av rømt oppdrettslaks. Basert på dette vurderes det derfor å være moderat sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Overvåkningsprogrammet dekker kun i overkant av en tredjedel av elvene i området, og selv om dette dekker en høy andel av gytebestandsmålet i området, så vurderes kunnskapsstyrken som moderat.
Utfisking og fjerning av rømt oppdrettslaks fra vassdrag. Utfisking er gjennomført årlig i 1-4 vassdrag i området i perioden 2017 til 2021. Det er ingen vassdrag med høyt innslag av rømt laks i området, mens det i vassdrag med middels innslag ble rapportert om utfisking i 75 % (6 av 8) av vassdragene det samme året. Totalt er det i hele perioden kun fjernet 14 rømte oppdrettslaks fra området og effekten av utfisking vurderes av den grunn som dårlig. Det vurderes derfor at det er høy sannsynlighet for at utfiskingen som gjennomføres i området ikke er tilstrekkelig til å fjerne all rømt oppdrettslaks.
Rømt oppdrettslaks på gyteplassen. Rømming og andelen rømt oppdrettslaks i elv vurderes som moderat og utfiskingstiltak vurderes å ha lav effekt. Det vurderes samlet sett å være moderat sannsynlighet for tilstedeværelse av rømt oppdrettslaks på gyteplassene i PO12. Til tross for at det er god kunnskap om utfiskingen i området, er det moderat kunnskap om hvor mye oppdrettslaks som rømmer og hvor mye rømt oppdrettslaks det er i elvene da overvåkningsprogrammet ikke dekker alle elver i området. Kunnskapsstyrken vurderes derfor som moderat.
Villaksens bestandsstatus. Gytebestandsmålene blir nådd i de fleste vassdragene i produksjonsområdet. Det høstbare overskuddet er også godt i de fleste vassdragene. Samlet vurderes tilstanden som god. Vassdragene med full vurdering utgjør 95 % av det samlede gytebestandsmålet i produksjonsområdet. Vurderingen er den samme uavhengig av vurderingsmetode og kunnskapsstyrken vurderes derfor som god.
Villaksens genetiske status. Svak genetisk endring er indikert i nær halvparten av bestandene, inklusive de tre største bestandene. Stor genetisk endring er påvist i tre bestander, mens én er uten observert genetisk endring. Villaksens genetiske status i området vurderes derfor samlet sett til å være i moderat tilstand (svake genetiske endringer indikert). Vurderingen er basert på ni bestander som til sammen utgjør 95 % av produksjonsområdets totale gytebestandsmål, så kunnskapsstyrken anses som god.
Bestandenes robusthet mot innkryssing. Siden bestandsstatus har god tilstand, vurderes også bestandenes robusthet mot innkryssing til god tilstand, selv om den genetiske statusen vurderes som moderat. Høy tetthet og konkurranse på gyteplassen antas å gi den rømte oppdrettslaksen lav gytesuksess og vektes derfor tyngre enn den genetiske statusen. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO12. Selv om bestandenes robusthet mot innkryssing vurderes som god, er det moderat sannsynlighet for at det finnes rømt oppdrettslaks på gyteplassene. Sannsynlighet for ytterligere genetisk endring hos villaksbestandene som følge av rømt oppdrettslaks vurderes fortsatt som moderat i PO12. Til tross for at vi har relativt god kunnskap om flere av de underliggende faktorene, så mangler det kunnskap knyttet til hvor mye rømt oppdrettslaks det er på gyteplassene og hvor robuste bestandene er for ny innkryssing. Vurderingen av kunnskapsstyrken vurderes derfor som moderat for området.
4.13. Produksjonsområde 13, Øst-Finnmark
Figur 4.13. Visualisering av risikobildet for ytterligere genetisk endring i villaksbestandene som følge av innkryssing av rømt oppdrettslaks i produksjonsområde 13 (PO13), Øst-Finnmark.
Rømming. Det ble rapportert én rømmingsepisode med 19 rømte oppdrettslaks i PO13 i perioden 2017–2021 med et årlig gjennomsnitt på 4. Området vurderes derfor å ha hatt lave rømmingstall i perioden. Det mangler kunnskap knyttet til omfang av rømming og påvirkning fra rømming i andre produksjonsområder, og kunnskapstyrken vurderes som moderat.
Andel rømt oppdrettslaks i elv. I gjennomsnitt ble 44 % av vassdragene i området (9 av 20) overvåket årlig i perioden 2017-2021, men verken Tana eller Neiden er inkludert. Dette utgjorde kun 13 % av gytebestandsmålet i området. Det er gjennom perioden 2017–2021 ingen vassdrag i kategorien høyt innslag av rømt oppdrettslaks i dette området. Basert på dette vurderes det derfor å være lav sannsynlighet for at det er mye rømt oppdrettslaks i elvene i området. Vi vurderer kunnskapsstyrken som god, fordi det foreligger estimater av andel rømt oppdrettslaks fra andre undersøkelser utført av finske forskere som viser lave andeler i to elver (Tana og Neiden) som ikke er inkludert i overvåkningsprogrammet.
Utfisking av rømt oppdrettslaks fra elv. Utfisking er ikke registrert i området, og det er ikke registrert vassdrag med høyt innslag av rømt laks. Det er totalt sett registrert to vassdrag med moderat innslag av rømt oppdrettslaks, ett i 2017 og ett i 2021. Vi har derfor valgt å ikke vurdere tilstanden for denne risikofaktoren i PO 13.
Rømt oppdrettslaks på gyteplassen. Det er lite rømming og lav andel rømt oppdrettslaks observert i elvene i PO13. Den totale vurderingen er derfor at det lav sannsynligvis for forekomst av rømt oppdrettslaks på gyteplassene i PO13. Det finnes ingen fullstendig oversikt over hvor mye rømt oppdrettslaks som finnes på gyteplassene og rømt oppdrettslaks fra omkringliggende områder kan også gå opp i elvene i området. Kunnskapsstyrken vurderes derfor som moderat.
Villaksens bestandsstatus. Gytebestandsmålene blir nådd i de fleste vassdragene i produksjonsområdet. Det høstbare overskuddet er også godt i de fleste vassdragene. Imidlertid har den største bestanden i produksjonsområdet (Tanavassdraget) redusert gytebestandsmåloppnåelse og redusert høstbart overskudd, trolig som følge av overbeskatning gjennom en årrekke. Siden Tanavassdraget er den desidert største bestanden i området, gjør dette at vurderingene blir svært forskjellig avhengig av om vi veier med gytebestandsmål eller ikke. Siden Tana er det viktigste laksevassdraget i Norge, tillegges det stor vekt, og vi vurderer den samlede tilstanden til å være moderat for området. Vassdragene med full vurdering utgjør 96 % av det samlede gytebestandsmålet i produksjonsområdet. Det er stort sprik i den samlede vurderingen avhengig av hvordan Tanavassdraget vektlegges. Den samlede vurderingen for regionen er derfor ikke robust, selv om vurderingene av de enkelte vassdragene er relativt sikre, og kunnskapsstyrken vurderes derfor som moderat.
Villaksens genetiske status. Tanavassdraget, som er den dominerende bestanden i regionen, er uten observert genetisk endring. I resten av regionen er det en jevn fordeling av bestander med store, moderate, svake og ingen observerte genetiske endringer. Villaksens genetiske status i området vurderes totalt sett til å være i moderat tilstand (svake genetiske endringer indikert). Vurderingen er basert på 16 bestander som til sammen utgjør 99 % av produksjonsområdets totale gytebestandsmål. Vi har svært begrensede prøver fra Tanavassdraget, med kun 41 voksne laks fra perioden 2005–2010 og en analyse av smolt fra 2014. Da det ikke foreligger prøver fra Tanavassdraget fra tiden etter 2014, justeres kunnskapsstyrken ned fra moderat til dårlig.
Bestandenes robusthet mot innkryssing. Både genetisk status og villaksens bestandsstatus vurderes å ha moderat tilstand og bestandenes robusthet mot innkryssing vurderes derfor også til å ha moderat tilstand. Kunnskapen knyttet til både villaksens bestandsstatus og genetiske status vurderes som god. Kunnskap knyttet til den kombinerte effekten av bestandsstatus og genetisk status er derimot begrenset, så på tross av god kunnskap om begge de underliggende faktorene vurderes kunnskapsstyrken som moderat.
Ytterligere genetisk endring hos villaks som følge av innkryssing av rømt oppdrettslaks i PO13. Selv om bestandenes robusthet i området vurderes som moderat, vektes den lave andelen av rømt oppdrettslaks tyngre og sannsynlighet for ytterligere genetisk endring i PO13 vurderes som lav. Vurderingen for dette produksjonsområdet er noe kompleks i og med at bestandsstatus er annerledes i Tanavassdraget enn i de andre vassdragene i området. Det foreligger heller ikke norske undersøkelser av forekomsten av rømt oppdrettslaks i Tana, men undersøkte prøver viser ingen genetisk endring som følge av innkryssing. Det er også usikkerhet i forhold til utviklingen i akvakultur på russisk side av grensen, hvor det har vært ambisjoner om økning i produksjonen. Det er moderat usikkerhet knyttet de fleste av de underliggende faktorene for både rømt oppdrettslaks på gyteplassene og hvor robust bestandene er for ny innkryssing. Kunnskapsstyrken vurderes derfor samlet sett som moderat.
5. Konklusjon
Risikovurderingen viser at kun de to sørligste (PO1 og PO2), samt det nordøstligste produksjonsområdet (PO13) fortsatt vurderes til å ha lav risiko for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks. Fire av de andre produksjonsområdene (PO5, 6, 10 og 12) vurderes til å ha moderat risiko for ytterligere genetisk endring, mens seks produksjonsområder (PO3, 4, 7, 8, 9 og 11) vurderes til å ha høy risiko for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks. I PO10 er risikobildet justert ned fra høy til moderat risiko, mens for de andre POene er vurderingene uendret fra risikovurderingene gjort i 2019, 2021 og 2022.
I de siste årene har de offisielle rømmingstallene fra industrien vist en nedgang, og overvåkningsprogrammet for rømt oppdrettslaks har vist lavere andel rømt oppdrettslaks i elvene de siste årene. Denne utviklingen gjenspeiles delvis i året risikorapport, hvor en nedgang i andel rømt oppdrettslaks i elvene i PO10 har resultert i en nedjustering av risikobildet. Det er likevel verdt å merke seg at i PO8 så vurderes andel rømt oppdrettslaks som høy, kontra moderat ved forrige vurdering. I områder med lite rømt oppdrettslaks kan én stor rømming forandre denne situasjonen raskt. Andel rømt oppdrettslaks i gytebestandene er også påvirket av endringer i antall villaks.
Så lenge oppdrettslaks produseres med dagens teknologi som hovedsakelig er åpne merder i sjø, vil risikoen for større rømningshendelser være til stede, og tall fra 2019 viser at nærmere 300 000 laks rømte det året. I 2021 ble det innrapportert nærmere 67 000 rømt oppdrettslaks, mens det i 2022 er blitt rapportert om i overkant av 56 000. Det er derfor stor sannsynlighet for at en også i de neste årene vil kunne få større rømningsepisoder. Det konkluderes derfor med at det også i de kommende år vil være moderat til høy risiko for ytterligere genetisk endring som følge av innkryssing av rømt oppdrettslaks i ville laksebestander i store deler av landet.
Tilgjengelig kunnskap tilsier at genetisk endring i ville laksebestander som følge av innkryssing av rømt oppdrettslaks, kan føre til redusert produksjon av genetisk villaks samt forandringer i viktige biologiske egenskaper i bestander som for eksempel alder og størrelse kjønnsmodning, smoltalder og endringer i utvandringstidspunkt for smolt. Dette kan resultere i mindre robuste bestander med redusert evne til å tilpasse seg til fremtidige utfordringer. Videre tilsier all tilgjengelig kunnskap at nivået av negative biologiske forandringer i de ville bestandene i stor grad vil være bestemt av nivået av genetisk innblanding fra rømt oppdrettslaks. Enkelt forklart betyr mer innkryssing større risiko for negative biologiske konsekvenser.
Sterke villaksbestander vil kunne redusere innkryssing av rømt oppdrettslaks ved økt konkurranse og gjennom en uttynningseffekt. Forvaltning etter gytebestandsmål, hvor man tar sikte på å ha nok vill gytelaks til stede om høsten til at elvene når sitt produksjonspotensial, vil bidra til å minske risikoen for at rømt oppdrettslaks fører til ytterligere genetisk endring. Flere tiltak er iverksatt både fra forvaltningen og næringen selv for å redusere rømming. Økt innsats for å gjøre tiltakene så effektive som mulig i å hindre rømt oppdrettslaks å komme til gyteplassene, vil også redusere risikoen for ytterligere innkryssing. Bruk av steril laks i oppdrett vil hindre videre innkryssing, men det vil imidlertid kunne gå flere år før produksjon av steril laks er klar for oppskalering.
Denne risikovurderingen er gjort på produksjonsområdenivå med mål om å gi et overordnet bilde av risiko knyttet til ytterligere genetisk endring i ville laksebestander som en konsekvens av innkryssing av rømt oppdrettslaks. Siden det innenfor hvert produksjonsområde vil være vassdrag og laksebestander som er ulike med hensyn til disse faktorene, vil aggregering av bestandene innenfor et produksjonsområde ikke nødvendigvis gi et godt uttrykk for tilstand og risiko for enkeltbestander. Neste steg kan være å gjøre en risikovurdering på elvenivå eller lokalisere mindre geografiske områder der risikoen vurderes å være høy, og gjøre en mer detaljert vurdering
6. Kunnskapsstatus
I denne todelte kunnskapsstatusen presenterer vi først en utvidet gjennomgang av faktorene knyttet til selve risikovurderingen; andel rømt oppdrettslaks på gyteplassene (rømming, andel rømt oppdrettslaks i elv og utfisking/ fjerning av rømt oppdrettslaks fra elv) og bestandenes robusthet for ny innkryssing (bestandsstatus og genetisk status). Vi drøfter også ulike biologiske faktorer ved den rømte fisken som kan påvirke dens gytesuksess i naturen, men som på nåværende tidspunkt ikke er inkludert i risikovurderingen. Videre presenterer vi en oversikt over konsekvenser av at rømt oppdrettslaks krysser seg med villaks og hvilke faktorer som forklarer variasjon i andel rømt oppdrettslaks i elv og grad av innkryssing.
6.1. Faktorer inkludert i risikovurderingen
Rømt oppdrettslaks på gyteplassene
Rømming
De offisielle innrapporterte rømmingstallene for laks (www.fiskeridir.no) viser at det siden 2001 årlig har rømt flere hundre tusen oppdrettslaks de fleste år (figur 6.1). I 2017 var det kun rapportert 17 000 rømt oppdrettslaks som er det laveste nivået i hele perioden. Det høyeste rapporterte tallet var i 2006 da 921 000 oppdrettslaks ble meldt rømt. Disse tallene er minimumsestimater og de faktiske rømmingstallene er sannsynligvis høyere enn det som rapporteres, både grunnet urapporterte hendelser og usikkerhet knyttet til fastsetting av antall rømt fisk ved store rømmingsepisoder. Havforskningsinstituttets DNA-identifisering av urapportert rømt oppdrettslaks (Glover 2010; Zhang mfl. 2013), og en større studie med utsetting av merket laks og modellering viste at de faktiske rømmingstallene for perioden 2005–2011 sannsynligvis var 2–4 ganger høyere enn den offisielle statistikken (Skilbrei mfl. 2015a). Det er ikke gjort tilsvarende studier for seinere år, og en ny estimering av forskjellen mellom rapporterte og faktiske rømmingstall trengs for å dokumentere effekten av ytterligere tiltak iverksatt for å forhindre rømming i etterkant av 2011. Selv om det er usikkerhet i de offisielle rømmingstallene, er risikovurderingen basert på årlig gjennomsnittlig rapportert rømming per produksjonsområde i perioden 2017–2021 (tabell 6.1). I dette tidsrommet ble det meldt om totalt 573 128 rømte oppdrettslaks.
Figur 6.1. Antall rømt oppdrettslaks rapportert årlig til Fiskeridirektoratet i perioden 2001–2021 (per 12.12.2022). I 2022 er det rapportert 56 174 rømte oppdrettslaks per 17.01.23. Kilde: www.fiskeridir.no.
Tabell 6.1. Rapportert antall rømt oppdrettslaks fra norsk lakseoppdrett i perioden 2017-2021 for produksjonsområde 1-13.
Biologisk status ved rømming påvirker oppdrettslaksens evne til å overleve i naturen, vandre opp i vassdragene og gyte sammen med villaks. Dette omfatter for eksempel livsfase, kjønn, kjønnsmodning, tidspunkt for rømming, lysregime på anlegget før rømming, sykdomsstatus, størrelse, alder og tidsforløp i det fri. Biologisk status påvirker også hvor langt den rømte oppdrettslaksen kan spre seg. Disse faktorene diskuteres her, men er på nåværende tidspunkt ikke inkludert i risikovurderingen da det finnes lite kunnskap om oppdrettslaksens biologiske status ved rømming og i hvor stor grad den sprer seg fra der den rømmer og til andre produksjonsområder.
Det er stor variasjon i spredning og overleving hos rømt oppdrettslaks, og rømt oppdrettslaks kan spre seg over store områder (Hansen mfl. 1993; Hansen 2006a, b; Jensen mfl. 2013; Quintela mfl. 2016). Årstid, rømmingslokalitet (eks. fjord vs. kyst), størrelse og alder ved rømming ser ut til å være avgjørende for hvor den rømte fisken svømmer og i hvilken grad de overlever.
Villaks legger ut på lange vandringer, fra elven til storhavet som liten smolt og tilbake som kjønnsmoden laks. Vandringene er synkronisert med årstidene, der smolt vandrer ut til havet i perioden april-juli, mens kjønnsmoden laks vandrer tilbake til elvene fra sent om våren til utpå høsten.
Også laks i oppdrett har disse vandringsinstinktene, men både årstid, størrelse og kjønnsmodningsstatus til fisken når den rømmer, har betydning for hvilken atferd den får i frihet. I noen tilfeller vil rømt oppdrettslaks spre seg hurtig og være lite fangbare, mens den under andre forhold vil holde seg lenge i området og kanskje søke opp i nærliggende elver.
Havforskningsinstituttet har tidligere gjennomført en serie med eksperimentelle slipp av oppdrettslaks for å studere ulike atferdsmønstre. Resultatene fra disse forsøkene er satt sammen med kunnskap fra andre kilder som også beskriver slipp av merket oppdrettslaks (Hansen 2006a, b; Chittenden mfl. 2011). Smolt og postsmolt som rømmer den første sommeren etter at de er satt ut i merder i sjøen, vil normalt vandre hurtig mot havet (Skilbrei 2010; Skilbrei mfl. 2015a). Dette innebærer at gjenfangst blir nærmest umulig. Denne fisken vokser opp i de åpne havområdene sammen med villaksen, og en liten andel (0,4 % gjenfangst fra våre utslipp) kommer tilbake til kysten ett til tre år senere som kjønnsmoden fisk. Våre undersøkelser viser at en høy andel av disse vil søke tilbake mot ferskvannskilder i området de rømte fra som smolt, men mange vandrer likevel opp i elver spredd over et område på flere hundre kilometer (Skilbrei mfl. 2015a; figur 6.2). Nyere upubliserte analyser at data fra disse eksperimentelle slippene viser at smoltrømming har potensialet til å spre seg mer enn 120 mil fra slipplokasjonen ved retur til ferskvann 1-3 år senere.
Vandringsviljen til postsmolt som rømmer om høsten reduseres i takt med at dagene blir kortere. Derfor øker også den lokale gjenfangsten av postsmolt som rømmer utover høsten (13 % gjenfangst fra våre utslipp), blant annet fordi de er blitt store nok til å bli fanget i garn og av sportsfiskere (Skilbrei mfl. 2015a).
Dersom fisk rømmer kort tid etter at de har blitt flyttet fra kar på et settefiskanlegg til merd i sjøen, blir vandringsmotivasjonen påvirket av lysregimet fisken var utsatt for på settefiskanlegget. Kontinuerlig belysning svekker vandringsviljen og kan få stor settefisk til å holde seg i fjorden uvanlig lenge, selv om de rømmer om sommeren (Skilbrei mfl. 2014). I produksjon av høstsmolt kan bruk av kunstig økt daglengde på settefiskanlegget få laksen til å legge ut på vandring mot havet selv om den rømmer fra merden seint om høsten. Sjansen for at disse overlever vinteren i havet er imidlertid lav (Skilbrei 2013), da slipp av høstsmolt ga lavere gjenfangst som voksen (0,004 %) en slipp av vårsmolt (0,17 %). Det er dermed lavere sannsynlighet for at fisk fra denne type rømming overlever fram til kjønnsmodning og vandrer opp i elv.
Gjenfangst av voksen laks avhenger av rømmingsområdet. Slipp av voksen laks fra anlegg på kysten har gitt lavere gjenfangst (4–7 %) enn slipp i fjorder der fisken oppholder seg lenger og er mer eksponert for fiskeredskaper (7–33 %). Slipp fra anlegg helt ute i havgapet har ikke gitt gjenfangster (Skilbrei mfl. 2015a). Flesteparten av gjenfanget voksen laks ble fanget i månedene etter at de ble satt ut, i nærheten av utslippsområdet. Nyere upubliserte analyser at data fra disse eksperimentelle slippene viser likevel at disse voksenrømlingene spredte seg på det meste mer enn 400 km fra slipplokasjonen. Kun 0,09 % ble gjenfanget 1–2 år etter utslippet (Skilbrei mfl. 2015a). En medvirkende årsak til dette er muligens at voksen laks som rømmer har vansker med å lære seg å fange naturlig føde. Undersøkelser av mageinnhold til voksen rømt oppdrettslaks fanget nær norskekysten viser vanligvis at de aller fleste er tomme (Olsen & Skilbrei 2010). Rømt oppdrettslaks fanget under fisket ved Færøyene har derimot samme diett som villaksen der (Jacobsen & Hansen 2001).
Figur 6.2. Gjenfangst av voksen laks i sjø (gule sirkler) og elv (rød) fra slipp av smolt fra forskningsstasjonen Matre (R) i 2005–2010. Fisk gjenfanget < 6 km fra utsettingsstedet (49 % av rapportert gjenfangst) er ikke vist. Størrelsen på sirklene angir antall fisk fra 1 til 5 individer.
Rømminger om høsten, uavhengig av alder på laksen, kjennetegnes ofte av at mange fisk søker mot ferskvannskilder, både elver og kraftverk som slipper ut turbinvann. Dette er som ventet når det gjelder kjønnsmodnende fisk, men det har vist seg at mange umodne laks også søker inn mot elveosene og at noen også går opp i nedre del av elven (Madhun mfl. 2015). Det er kjent at umoden laks kan gå opp i ferskvann (Webb mfl. 2007), men denne atferden er ikke godt kartlagt. Det er mulig at noen av de umodne fiskene svømmer ut av elven igjen etter en stund, mens andre kan stå i elven til de blir kjønnsmodne.
Som nevnt over har merkestudiene vist at smolt og postmolt som rømmer kan komme tilbake som gyteklar laks etter 1–3 år i havet, og at voksen rømt oppdrettslaks som gjenfanges som oftest blir gjenfanget i løpet av det året de rømte (Skilbrei mfl. 2015a). Undersøkelser av et pigment i kjøtt som reflekterer ulik diett mellom oppdrettsmiljøet og naturen, viste tidlig på 90-tallet at om lag halvparten av den undersøkte rømte oppdrettslaksen fanget i elv hadde rømt nylig mens den resterende halvparten hadde tilbrakt mer enn ett år i naturen (Lura & Sægrov 1994). Senere har fettsyreanalyser av voksen rømt oppdrettslaks i elv vist at flertallet nylig hadde rømt, dette basert på at de hadde fettsyreprofiler som var svært lik oppdrettsfôret. En mindre andel på rundt 10–30 % av den voksne rømte oppdrettslaksen hadde fettsyreprofiler som tydet på at den hadde beitet i havet i lang tid og dermed sannsynligvis hadde rømt som smolt eller postsmolt. Det var færre observasjoner av voksen laks som hadde spist begge typer mat; som hadde rømt som relativt stor laks og deretter klart å finne ville byttedyr (Skilbrei mfl. 2015b; Anon. 2018). Dette støtter antakelsen fra merkestudiene om at en stor del av den umodne voksne oppdrettslaksen som rømmer sannsynligvis ikke overlever fram til den blir kjønnsmoden.
Tilgjengelige data tyder på at sannsynligheten for at en rømt oppdrettslaks overlever fram til kjønnsmodning og vandrer opp i elv er størst hvis fisken rømmer som smolt om sommeren, eller det samme året den blir kjønnsmoden. Sannsynligheten er minst for smolt som rømmer om høsten, og umoden laks som ikke blir kjønnsmoden før tidligst neste år. Et mulig neste steg for denne risikovurderingen vil derfor være å bruke rømmingstallene, i kombinasjon med biologisk data om oppdrettslaks ved rømming, til å vurdere sannsynlighet for overlevelse etter rømming og geografisk spredning.
Andel rømt oppdrettslaks i elv
Siden oppstarten av lakseoppdrett i Norge har flere millioner oppdrettslaks rømt fra en rekke anlegg langs norskekysten. De fleste av disse fiskene «blir borte» i det marine miljøet uten videre spor, men noen vil vandre opp i lakseelver. Det nasjonale overvåkningsprogrammet for rømt oppdrettslaks i vassdrag, som ble utformet og etablert på oppdrag fra Fiskeridirektoratet etter føringer fra Nærings- og fiskeridepartementet i 2014, beregnet innslag av rømt oppdrettslaks i henholdsvis 140 vassdrag ved oppstart (Anon. 2015a). I dag beregnes innslag av rømt oppdrettslaks i gjennomsnittlig 200 vassdrag årlig, 218 på det meste, og antallet vil variere da årlig variasjon i miljømessige forhold vil påvirke datakvaliteten (Anon. 2016a, 2017a, 2018, 2019a, Aronsen mfl. 2020, Wennevik mfl. 2021). I 2021 ble 178 vassdrag vurdert (Wennevik mfl. 2022).
Vassdragene som er overvåket er valgt ut fra en rekke kriterier; god geografisk spredning, inkludering av de nasjonale laksevassdragene, representasjon av vassdrag av ulik størrelse samt å bygge videre på vassdrag med tidsserier og med gode lokale nettverk, som er etablert i ulike prosjekter finansiert av Norges forskningsråd og miljømyndighetene siden 1989 (Fiske mfl. 2006; Diserud mfl. 2019a). Data blir samlet inn fra sportsfiske om sommeren, høstfiske, stamfiske og drivtellinger om høsten. De tre førstnevnte metodene er i hovedsak basert på stangfiske og skiller mellom rømt oppdrettslaks og villaks ved å undersøke fiskens skjell, som gir et bilde av fiskens vekstbetingelser gjennom i livet. Drivtellinger innebærer at snorklere foretar en visuell inspeksjon av fisk i elven, teller opp og karakteriserer vill og rømt oppdrettet laks på basis av utseende og atferd (Mahlum mfl. 2019). I mange av elvene som blir undersøkt blir mer enn én metode benyttet. Innsamlete data går gjennom en kvalitetssikringsprosess og er blitt enkeltvis vurdert i henhold til en rekke kriterier for å få en total vurdering av dataenes representativitet. Innslaget av rømt oppdrettslaks for hver elv presenteres i en årlig rapport fra overvåkningsprogrammet som prosentandelene registrert ved de ulike metodene, samt som en årsprosent som beregnes fra andel oppdrettslaks i sportsfiske og/eller høstfiske/stamfiske (Fiske mfl. 2006). Årsprosenten tar hensyn til at sportsfisket sannsynligvis gir et for lavt, og høstfisket sannsynligvis et for høyt estimat av innslaget av rømt oppdrettslaks. De ulike metodene som blir benyttet i de forskjellige elvene har sine styrker og svakheter, både i forhold til prøvestørrelsene og sikker identifikasjon av rømt oppdrettslaks. At innslaget av rømt oppdrettslaks i vassdragene kan endre seg i løpet av sesongen og at rømt oppdrettslaks til dels har en annen atferd enn villaks, gjør det krevende både å innhente gode data og å sammenligne data innhentet med de ulike metodene. Ved utregning av et estimat for prosentvis andel oppdrettslaks i elven, kommer det i tillegg en statistisk usikkerhet på anslaget som avhenger av prøvestørrelsen og innslaget av rømt oppdrettslaks. Ulike kilder til usikkerhet i dataene fra overvåkningsprogrammet blir diskutert i rapporten (Wennevik mfl. 2022).
Elver blir klassifisert i kategoriene <4 %, 4–10 % og >10 %, tilsvarende systemet foreslått av Taranger mfl. (2012). Klassifiseringen følger ikke årsprosent slavisk, men er basert på en samlet vurdering av alle datakildene:
Lavt innslag: innslag av rømt oppdrettslaks er estimert til under 4 %.
Moderat innslag: innslag av rømt oppdrettslaks er estimert til mellom 4 % og 10 %.
Høyt innslag: innslag av rømt oppdrettslaks er estimert til over 10 %.
Resultatene fra overvåkningsprogrammet er presentert på to nivåer. Hovedrapporten representerer en oppsummering av hovedresultatene, og viser hvilke metoder som er lagt til grunn (Wennevik mfl. 2022) og er publisert elektronisk sammen med vedleggsdokumenter som viser detaljerte resultater for hvert vassdrag. Disse vedleggsdokumentene er organisert fylkesvis. I risikovurderingen har vi brukt klassifiseringene overvåkningsprogrammet har gjort for de enkelte vassdrag i perioden 2017–2021 innenfor hvert av produksjonsområdene og vurdert disse samlet for å kategorisere tilstanden innenfor hvert produksjonsområde (tabell 6.2).
Det ble i 2021 beregnet årsprosent for 102 elver, og det presenteres i rapporten fra overvåkningsprogrammet data fra drivtellinger fra 105 elver. Resultatene fra overvåkningsprogrammet for 2021 viste at til sammen 140 elver (79 %) ble vurdert til å ha lavt innslag av rømt oppdrettslaks (mindre enn 4 %), 24 vassdrag (14 %) ble vurdert til å ha moderat innslag (mellom 4 og 10 %), mens 14 (8 %) vassdrag ble vurdert til å ha et høyt innslag av rømt oppdrettslaks. Dette var en svak oppgang fra 2020, hvor 178 elver (82 %) ble vurdert til å ha lavt innslag av rømt oppdrettslaks (mindre enn 4 %), 27 vassdrag (12 %) ble vurdert til å ha moderat innslag (mellom 4 og 10 %), mens 13 (6 %) vassdrag ble vurdert til å ha et høyt innslag av rømt oppdrettslaks. Det var derimot en nedgang i både antall og andel elver med høyt innslag i 2020 sammenlignet med 2019 (20 vassdrag, 10 %) og 2018 (19 vassdrag, 9 %). I 2017 var det 15 vassdrag (8 %) med høyt innslag av rømt oppdrettslaks. Antall vassdrag med høyt innslag av rømt oppdrettslaks i perioden 2017-2021 har dermed lagt på 6 -10 %, mens det i 2016 var 12 % av undersøkte vassdrag som hadde et høyt innslag av rømt oppdrettslaks (figur 6.3).
Figur 6.3. Andel vassdrag som er klassifisert til å ha lav, middels eller høyt innslag av rømt oppdrettslaks som klassifisert av det nasjonale overvåkningsprogrammet for rømt laks i vassdrag siden oppstart i 2014 og fram til 2021.
Innslaget av rømt oppdrettslaks i 2021 varierte langs norskekysten, for eksempel ved at Vestland, som i tidligere år, hadde flere av vassdragene med høyt innslag av rømt oppdrettslaks (4). Det var også vassdrag med høyt innslag av rømt oppdrettslaks i Møre og Romsdal (1), Trøndelag (1) og Nordland (8). I Nordland var det en dobling i antall vassdrag med høye innslag, i forhold til året før. Ingen vassdrag med høye innslag av rømt oppdrettslaks ble registrert i Troms og Finnmark, mot ett vassdrag i 2020. Tilstanden var også god, med lave innslag av rømt oppdrettslaks, på hele strekningen fra Østfold til Rogaland. De to elvene med høye innslag i dette området i 2020 hadde i 2021 lave innslag av rømt oppdrettslaks (figur 6.4).
Tabell 6.2. Andel rømt oppdrettslaks i elv basert på en forenklet klassifiseringsmetode fra det nasjonale overvåkningsprogrammet i produksjonsområde 1-13. Betegnelsen "Nasjonal lakseelv" betyr at vassdraget har blitt tildelt en spesiell vernestatus.
Produksjonsområde
Antall nasjonale lakseelver
Antall lakse-vassdrag
Andel av totalt antall vassdrag i Norge (%)
Samlet gyte-bestandsmål
Prosent av totalt gytebestands-mål
Gjennomsnitt antall elver overvåket
Gjennomsnitt andel elver overvåket (%)
Gjennomsnitt andel vassdrag (%) vurdert å ha lav andel rømt opprettslaks i 2017-2021
Gjennomsnitt andel vassdrag (%) vurdert å ha moderat andel rømt opprettslaks i 2017-2021
Gjennomsnitt andel vassdrag (%) vurdert å ha høy andel rømt opprettslaks i 2017-2021
Gjennomsnitt andel av samlet gytebestand overvåket (%)
Andel (%) av gytebestand vurdert til å ha lav andel rømt oppdrettslaks 2017-2021
Andel (%) av gytebestand vurdert til å ha moderat andel rømt oppdrettslaks 2017-2021
Andel (%) av gytebestand vurdert til å ha høy andel rømt oppdrettslaks 2017-2021
1 - Svenskegrensen til Jæren
6
40
9
49,686
14
18
45
98
0
2
83
100
0
0
2 - Ryfylke
2
22
5
9,006
2
16
72
96
4
0
95
96
4
0
3 - Karmøy til Sotra
1
18
4
3,666
1
16
87
46
17
37
93
38
18
43
4 - Norhordland til Stadt
11
43
10
21,027
6
31
71
78
17
5
78
82
13
4
5 - Stadt til Hustadvika
2
46
10
17,760
5
22
47
84
13
3
66
70
22
8
6 - Nordmøre og Sør-Trøndelag
11
76
17
87,520
24
18
24
83
14
3
89
94
6
1
7 - Nord-Trøndelag med Bindal
2
24
5
27,600
8
9
37
68
9
23
91
95
2
4
8 - Helgeland til Bodø
3
30
7
18,303
5
16
54
72
19
10
87
57
26
17
9 - Vestfjorden og Vesterålen
0
57
13
6,634
2
12
21
68
20
12
37
68
19
13
10 - Andøya til Senja
2
26
6
12,434
3
14
53
67
26
7
72
51
44
5
11 - Kvaløya til Loppa
2
20
4
8,550
2
11
53
70
15
15
90
76
10
14
12 - Vest-Finnmark
5
26
6
26,802
7
10
37
83
17
0
87
97
3
0
13 - Øst-Finnmark
6
20
4
75,305
21
9
44
95
5
0
13
95
5
0
Sum
53
448
100
364,293
100
200
Kilde: Rømt oppdrettslaks i vassdrag i 2017, 2018, 2019, 2020 og 2021. Rapporter fra det Nasjonale Overvåkningsprogrammet. Fisken og havet Særnummer 2b 2018, Fisken og havet 2019-4, Fisken og havet 2020-3, Rapport fra havforskningen 2021-27, Rapport fra havforskningen 2022-27.
Figur 6.4. Lokalisering av elvene der innslaget av rømt oppdrettslaks i 2021 er vurdert av overvåkningsprogrammet til å være lavt (< 4 %, grønne sirkler), moderat (4 – 10 %, gule sirkler), eller høyt (>10 %, røde sirkler). Se teksten for nærmere forklaring av de tre kategoriene. Fra Wennevik mfl. (2022).
Andelen rømt oppdrettslaks i elvene har endret seg mellom år, og det har vært en synkende tendens i registreringene gjennom de siste årene (figur 6.5). Nivået av rømt oppdrettslaks nådde i 2020 det laveste nivået siden 2006, og størstedelen av nedgangen har skjedd etter 2013. Se Diserud mfl. (2019a) for en oppsummering av resultatene fra overvåkingen før 2014, og Glover mfl. (2019) for en beskrivelse av det nåværende overvåkningsprogrammets aktiviteter og resultater.
Figur 6.5. Gjennomsnittlig % rømt oppdrettslaks (o) i høstundersøkelsene for årene 2006–2021 for totalt 69 elver (Fiske 2013, Fiske mfl. 2014; Anon. 2015a, 2016a, 2017a, 2018, 2019a, Aronsen mfl. 2020, Wennevik mfl. 2021, 2022), vist for hele Norge. Prediksjoner basert på modell er vist med tykk linje (signifikant nedgående trend). Se Wennevik mfl. (2022), for mer detaljer.
Utfisking/fjerning av rømt oppdrettslaks fra elv
Data fra det nasjonale overvåkningsprogrammet kommer til direkte anvendelse gjennom utfiskingsforskriften som Nærings- og fiskeridepartementet vedtok i 2015 (Forskrift om fellesansvar for utfisking mv. av rømt oppdrettsfisk), der oppdrettsnæringens sammenslutning for utfisking av rømt oppdrettsfisk (OURO) er hjemlet. Her er det fastsatt at det skal planlegges tiltak for å redusere mengde rømt oppdrettslaks i elver med mer enn 10 % innslag av oppdrettslaks, dokumentert gjennom overvåkningsprogrammet. Tiltakene iverksettes derfor ett år etter at det er observert høye andeler rømt oppdrettslaks i elvene. Utføring av pålagte oppgaver fra forskriften ble satt i verk av OURO i 2016. Det ble gjennom OURO utført tiltak i minst 37 elver i 2016, 51 elver i 2017, 60 elver i 2018, 48 elver i 2019, 50 elver i 2020 og 30 elver i 2021 (tallene inkluderer også uttak i kilenøter). Det gjennomføres således utfisking i flere elver enn de som er vurdert til å ha > 10 % innslag av rømt oppdrettslaks. I tillegg til utfiskingstiltak organisert gjennom OURO, organiserer også Fiskeridirektoratet og andre forvaltningsmyndigheter uttaksfiske som avbøtende tiltak ved akutte rømmingsepisoder og i vassdrag hvor mye oppdrettslaks blir observert. For nærmere informasjon om utfiskingstiltakene og gjennomføring i de ulike vassdragene se også rapporter fra aktører som har deltatt i fisket på OURO sine nettsider (www.utfisking.no).
I risikovurderingen ser vi på alle elver per produksjonsområde hvor utfisking etter rømt oppdrettslaks har vært gjennomført, og tallene hentes fra overvåkningsprogrammet, fra OURO eller andre utfiskingsaktører direkte (gjelder kun elver med uttaksaktivitet, men hvor det ikke tas ut noe rømt oppdrettslaks, ettersom dette da ikke nødvendigvis rapporteres til Overvåkningsprogrammet). Effekten av utfiske i et produksjonsområde vurderes basert på andel vassdrag med høyt og middels innslag av rømt oppdrettslaks hvor det også ble rapportert om utfisking samme år.
I perioden 2017 – 2021 ble det årlig gjennomført utfiskingstiltak i 58-94 vassdrag for å redusere andelen rømt oppdrettslaks. Antall elver er noe oppjustert fra fjorårets Kunnskapsstatus grunnet tilgang til rapporter fra flere utfiskingsaktører (gjelder 2017 og 2018). I overvåkningsprogrammet vil kun utfisket oppdrettslaks som er bekreftet gjennom skjellanalyser inngå i datagrunnlaget. Det reelle antallet oppdrettslaks som tas ut vil derfor kunne være noe høyere grunnet manglende eller uleselige skjellprøver. Antall elver med utfisking kan også være noe høyere, da aktivitet i vassdrag som ikke rapporteres til overvåkningsprogrammet kan forekomme. Elver hvor vi er kjent med at utfisking har vært gjennomført av OURO eller andre aktører, uten observasjoner av rømt fisk, er dermed inkludert i datagrunnlaget ved utregning av antall elver med utfisking per produksjonsområdet.
Antall elver med utfisking og antall rømt oppdrettslaks fjernet fra elver av OURO og andre aktører i hvert produksjonsområde, som rapportert til Overvåkningsprogrammet for rømt laks i vassdrag, i perioden 2017–2021 utgjør datagrunnlaget for vurdering av denne faktoren i risikovurderingen (tabell 6. 3).
Tabell 6.3A. Antall elver med utfiskingsaktivitet og antall rømt oppdrettslaks fjernet fra elver i perioden 2017- 2021 for produksjonsområde 1-13.
2017-2021
2017
2018
Høyt innslag
Middels innslag
Høyt innslag
Middels innslag
Høyt innslag
Middels innslag
Produksjonsområde
Totalt antall elver med årlig uttak
Totalt antall rømt oppdrettslaks fjernet
Totalt antall elver vurdert til høyt innslag
Antall elver med høyt innslag og uttak
Andel elver med høyt innslag og uttak %
Totalt antall elver vurdert til middels innslag
Antall elver med middels innslag og uttak
Andel elver med middels innslag og uttak
Totalt antall elver med uttak
Totalt antall rømt oppdrettslaks fjernet
Totalt antall elver vurdert til høyt innslag
Antall elver med høyt innslag og uttak
Andel elver med høyt innslag og uttak %
Totalt antall elver vurdert til middels innslag
Antall elver med middels innslag og uttak
Andel elver med middels innslag og uttak
Totalt antall elver med uttak
Totalt antall rømt oppdrettslaks fjernet
Totalt antall elver vurdert til høyt innslag
Antall elver med høyt innslag og uttak
Andel elver med høyt innslag og uttak %
Totalt antall elver vurdert til middels innslag
Antall elver med middels innslag og uttak
Andel elver med middels innslag og uttak
1 - Svenskegrensen til Jæren
12
62
2
2
100
0
0
1
0
0
0
0
0
2
2
0
0
0
0
2 - Ryfylke
27
83
0
0
3
3
100
5
4
0
0
0
0
4
7
0
0
0
0
3 - Karmøy til Sotra
68
856
29
26
90
13
9
69
17
173
9
9
100
1
1
100
16
183
7
6
86
6
4
67
4 - Norhordland til Stadt
78
564
7
5
71
26
15
58
15
77
0
0
2
1
50
18
40
0
0
9
6
67
5 - Stadt til Hustadvika
30
119
3
2
67
14
4
29
8
91
1
1
100
1
0
0
4
11
1
0
0
1
0
0
6 - Nordmøre og Sør-Trøndelag
15
30
3
2
67
13
8
62
3
3
0
0
1
1
100
4
5
2
2
100
1
1
100
7 - Nord-Trøndelag med Bindal
20
284
10
10
100
4
3
75
3
24
1
1
100
0
0
3
12
1
1
100
0
0
8 - Helgeland til Bodø
26
150
8
3
38
15
6
40
7
13
0
0
4
1
25
6
9
3
1
33
3
1
33
9 - Vestfjorden og Vesterålen
29
101
7
7
100
12
9
75
6
24
2
2
100
2
1
50
7
13
0
0
2
1
50
10 - Andøya til Senja
44
114
5
4
80
18
15
83
16
46
1
1
100
2
2
100
12
25
2
2
100
6
5
83
11 - Kvaløya til Loppa
30
84
8
7
88
8
7
88
9
20
1
0
0
1
1
100
8
36
3
3
100
3
3
100
12 - Vest-Finnmark
13
14
0
0
8
6
75
4
6
0
0
2
2
100
2
2
0
0
2
0
0
13 - Øst-Finnmark
0
0
0
0
2
0
0
0
0
0
0
1
0
0
0
0
0
0
0
0
Sum
392
2461
82
68
83
136
85
63
94
481
15
14
93
17
10
59
86
345
19
15
79
33
21
64
Kilde: Rømt oppdrettslaks i vassdrag i 2017, 2018, 2019, 2020 og 2021. Rapporter fra det Nasjonale Overvåkningsprogrammet. Fisken og havet Særnummer 2b 2018, Fisken og havet 2019-4, Fisken og havet 2020-3, Rapport fra havforskningen 2021-27, Rapport fra havforskningen 2022-27. Rapporter fra OURO og andre aktører er listet under Referanser (kapittel 8).
Tabell 6.3B. Antall elver med utfiskingsaktivitet og antall rømt oppdrettslaks fjernet fra elver i perioden 2017- 2021 for produksjonsområde 1-13.
2019
2020
2021
Høyt innslag
Middels innslag
Høyt innslag
Middels innslag
Høyt innslag
Middels innslag
Produksjonsområde
Totalt antall elver med uttak
Totalt antall rømt oppdrettslaks fjernet
Totalt antall elver vurdert til høyt innslag
Antall elver med høyt innslag og uttak
Andel elver med høyt innslag og uttak %
Totalt antall elver vurdert til middels innslag
Antall elver med middels innslag og uttak
Andel elver med middels innslag og uttak
Totalt antall elver med uttak
Totalt antall rømt oppdrettslaks fjernet
Totalt antall elver vurdert til høyt innslag
Antall elver med høyt innslag og uttak
Andel elver med høyt innslag og uttak %
Totalt antall elver vurdert til middels innslag
Antall elver med middels innslag og uttak
Andel elver med middels innslag og uttak
Totalt antall elver med uttak
Totalt antall rømt oppdrettslaks fjernet
Totalt antall elver vurdert til høyt innslag
Antall elver med høyt innslag og uttak
Andel elver med høyt innslag og uttak %
Totalt antall elver vurdert til middels innslag
Antall elver med middels innslag og uttak
Andel elver med middels innslag og uttak
1 - Svenskegrensen til Jæren
2
5
0
0
0
0
4
54
2
2
100
0
0
3
1
0
0
0
0
2 - Ryfylke
8
64
0
0
2
2
100
6
7
0
0
0
0
4
1
0
0
1
1
100
3 - Karmøy til Sotra
15
262
7
6
86
3
3
100
11
125
3
3
100
3
1
33
9
113
3
2
67
0
0
4 - Norhordland til Stadt
17
383
4
4
100
7
3
43
16
47
2
1
50
3
1
33
12
17
1
0
0
5
4
80
5 - Stadt til Hustadvika
6
12
1
1
100
7
3
43
7
2
0
0
4
1
25
5
3
0
0
1
0
0
6 - Nordmøre og Sør-Trøndelag
3
7
0
0
3
2
67
2
3
0
0
4
2
50
3
12
1
0
0
4
2
50
7 - Nord-Trøndelag med Bindal
4
38
2
2
100
1
1
100
5
101
3
3
100
1
0
0
5
109
3
3
100
2
2
100
8 - Helgeland til Bodø
5
15
0
0
4
3
75
5
6
1
0
0
1
1
100
3
107
4
2
50
3
0
0
9 - Vestfjorden og Vesterålen
5
39
2
2
100
3
2
67
5
14
1
1
100
3
3
100
6
11
2
2
100
2
2
100
10 - Andøya til Senja
7
35
2
1
50
3
3
100
6
1
0
0
5
4
80
3
7
0
0
2
1
50
11 - Kvaløya til Loppa
6
23
3
3
100
1
1
100
5
0
1
1
100
2
1
50
2
5
0
0
1
1
100
12 - Vest-Finnmark
3
2
0
0
1
1
100
1
0
0
0
1
1
100
3
4
0
0
2
2
100
13 - Øst-Finnmark
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1
0
0
Sum
81
885
21
19
90
35
24
69
73
360
13
11
85
27
15
56
58
390
14
9
64
24
15
63
Kilde: Rømt oppdrettslaks i vassdrag i 2017, 2018, 2019, 2020 og 2021. Rapporter fra det Nasjonale Overvåkningsprogrammet. Fisken og havet Særnummer 2b 2018, Fisken og havet 2019-4, Fisken og havet 2020-3, Rapport fra havforskningen 2021-27, Rapport fra havforskningen 2022-27. Rapporter fra OURO og andre aktører er listet under Referanser (kapittel 8).
Bestandenes robusthet for ny innkryssing
Villaksens bestandsstatus
Antall gytelaks (kilo hunnlaks) som trengs for å utnytte elvens produksjonspotensial kalles gytebestandsmål (Hindar mfl. 2007). Bestander som både når gytebestandsmålet og har et høyt produksjonspotensial er trolig mer robuste mot innkryssing av rømt oppdrettslaks enn bestander som har lite produksjonspotensial og/eller som ikke når gytebestandsmålet.
Oppnåelse av gytebestandsmål og høstingspotensial til bestandene beregnes årlig av Vitenskapelig råd for lakseforvaltning (VRL), og brukes her som en indikator for villaksbestandens robusthet for ny innkryssing av rømt oppdrettslaks. I «Kvalitetsnorm for ville bestander av laks (Salmo salar)», delnorm gytebestandsmål og høstingspotensial, blir bestandene av VRL delt inn i kategoriene: «svært god», «god», «moderat», «dårlig» og «svær dårlig» kvalitet (Anon. 2019b).
Dersom bestandene når sine gytebestandsmål, har de mange nok ville gytelaks på gyteplassene til å kunne utnytte elvens produksjonspotensial. Konkurransen på gyteplassen blir enda sterkere dersom flere villaks enn gytebestandsmålet er til stede på gyteplassen. Bestander med høyt høstingspotensial (vesentlig flere voksenlaks kommer tilbake enn det som er nødvendig for å nå gytebestandsmålet) har også større sannsynlighet for å nå gytebestandsmålet i årene som kommer, selv om overlevelsen i havet skulle bli redusert. Konkurransen mellom ungfiskene i elven vil også være større hvis det er mange fisk som gyter i vassdraget, og avkom av rømt oppdrettslaks vil gjøre det relativt sett dårligere hvis det er mange ville ungfisk å konkurrere med (Skaala mfl. 2012). Det antas derfor at bestander som både når gytebestandsmålet og har et høyt produksjonspotensial (overgår gytebestandsmålet) er mer robuste mot innkryssing av rømt oppdrettslaks enn bestander som har lite produksjonspotensial og/eller som ikke når gytebestandsmålet.
I risikovurderingen er vurderingen av villaksens bestandsstatus i produksjonsområdene basert på beregninger av måloppnåelse for gytebestandsmål og høstingspotensial (Anon. 2018b og 2019b, 2020, 2021,2022) for den enkelte villaksbestand i perioden 2016–2020 (tabell 6.4).
Tabell 6.4. Villaksens bestandsstatus for produksjonsområde 1-13.
Produksjonsområde
Antall laksevassdrag
Antall elver undersøkt
Prosent gytebestandsmål evaluert
Gjennomsnittlig gytebestandsmåloppnåelse i prosent - uveid
Gjennomsnittlig høstingspotensiale (som prosent av normalt høstingspotensiale for området) - uveid
Gytebestandsmål og høstingspotensiale - uveid
Gjennomsnittlig gytebestandsmåloppnåelse i prosent - veid
Gjennomsnittlig høstingspotensiale (som prosent av normalt høstingspotensiale for området) - veid
I 2021 ble villaksen ført opp på rødlistet som nært truet (Artsdatabanken, 2021). I perioden 1983-2019 har antall voksne villaks som kommer tilbake fra havet blitt redusert med hele 51%. Det er denne nedgangsraten som er lagt til grunn når villaksens nedgang over tre laksegenerasjoner (15-18 år) er beregnet til mellom 21-25 %, og dermed faller inn under kategorien nært truet. I risikovurderingen er det gytebestanden som inngår i estimatet på bestandsstatus, og dette estimatet har ikke vist samme nedgang i denne perioden, mye grunnet reduksjon i fiske.
Villaksens genetiske status
Rømt oppdrettslaks og deres avkom har antakeligvis større suksess i konkurranse med innkryssede individer enn med ikke-innkrysset villaks. Det er derfor sannsynlig at høy innkryssing av oppdrettslaks i bestandene vil gjøre dem mindre robuste for innkryssing av rømt oppdrettslaks i framtiden enn bestander som har liten grad av innkryssing.
NINA og Havforskningsinstituttet har i samarbeid produsert et omfattende sett med estimater for tidligere genetisk innkryssing av rømt oppdrettslaks i 239 vassdrag, hvorav 227 er klassifisert som laksbestander av VRL (Diserud mfl. 2020), som representerer om lag 94 % av villaksressursene i Norge (beregnet som andel av det totale gytebestandsmålet). Resultatene viser at oppdrettslaks er krysset inn i et stort antall elver: store genetiske forandringer er påvist i 68 vassdrag (> 10 % endring), moderate genetiske forandringer er påvist i 22 vassdrag (4-10 % endring), svake genetiske forandringer er indikert i 69 vassdrag (1-4 % endring) og ingen genetiske forandringer er observert i 80 vassdrag (< 1 % endring) (figur 6.6). Dette viser at genetisk innkryssing forekommer i en stor del av elvene i Norge da 2/3 deler av bestandene er plassert i kategoriene svake genetisk forandringer indikert til store genetisk forandringer dokumentert.
Det er utarbeidet kvalitative og kvantitative kriterier for hver av de fire tilstandsklassene, og i alt er det undersøkt om lag 52 000 villaks for å beskrive genetisk innkryssing i ville laksebestander. Beskrivelsen av genetisk status utgjør delnorm genetisk integritet til «Kvalitetsnorm for ville bestander av laks (Salmo salar)» og første statusrapport ble publisert for 125 elver i 2016 (Diserud mfl. 2016).
Over tid vil innkryssing av rømt oppdrettslaks kunne forandre egenskapene til de ville laksebestandene, redusere antall villaks som produseres og svekke bestandenes evne til å tilpasse seg endringer i miljøet. Innkryssing av rømt oppdrettslaks vil derfor kunne svekke bestandene og gjøre dem mindre robuste mot framtidig innkryssing av rømt oppdrettslaks. I risikovurderingen blir den genetiske påvirkningen av rømt oppdrettslaks på ville laksebestander oppsummert per produksjonsområde (tabell 6.5).
Tabell 6.5. Villaksens genetiske status i produksjonsområde 1-13. Grønn, gul, oransje og rød refererer til de fire kategoriene fra Diserud mfl. 2020 for klassifisering av elver med hensyn til genetisk integritet.
Grønn
Gul
Oransje
Rød
Produksjonsområde
Antall lakse-vassdrag
Antall vurderte
Prosent av laksevassdrag vurdert
Samlet gyte-bestandsmål (kg hunnfisk)
Gyte-bestandsmål vurderte (kg)
Prosent av gytebestandsmål vurdert
Antall
Andel
Gyte-bestandsmål vektet
Antall
Andel
Gyte- bestandsmål vektet
Antall
Andel
Gyte-bestandsmål vektet
Antall
Andel
Gyte-bestandsmål vektet
Genetisk innkryssing
Genetisk innkryssing - GBM veid
1 - Svenskegrensen til Jæren
40
24
60
49,686
47,981
97
16
0.667
0.576
4
0.167
0.060
3
0.125
0.325
1
0.042
0.039
0.016
0.020
2 - Ryfylke
22
16
73
9,006
8,714
97
8
0.500
0.676
6
0.375
0.219
0
0.000
0.000
2
0.125
0.104
0.034
0.013
3 - Karmøy til Sotra
18
12
67
3,666
3,621
99
1
0.083
0.117
0
0.000
0.000
1
0.083
0.118
10
0.833
0.765
0.162
0.148
4 - Norhordland til Stadt
43
33
77
21,104
19,771
94
4
0.121
0.095
11
0.333
0.163
3
0.091
0.394
15
0.455
0.348
0.092
0.073
5 - Stadt til Hustadvika
46
27
59
17,760
15,256
86
8
0.296
0.160
12
0.444
0.220
1
0.037
0.047
6
0.222
0.573
0.056
0.118
6 - Nordmøre og Sør-Trøndelag
76
29
38
87,002
83,820
96
7
0.241
0.392
12
0.414
0.505
1
0.034
0.058
9
0.310
0.045
0.074
0.022
7 - Nord-Trøndelag med Bindal
24
6
25
27,215
25,120
92
3
0.500
0.188
1
0.167
0.038
1
0.167
0.743
1
0.167
0.032
0.053
0.042
8 - Helgeland til Bodø
30
11
37
18,303
14,763
81
3
0.273
0.020
3
0.273
0.185
0
0.000
0.000
5
0.455
0.796
0.097
0.120
9 - Vestfjorden og Vesterålen
57
20
35
6,192
3,838
62
16
0.800
0.743
1
0.050
0.039
1
0.050
0.030
2
0.100
0.187
0.022
0.030
10 - Andøya til Senja
26
15
58
12,434
11,476
92
4
0.267
0.124
4
0.267
0.147
2
0.133
0.037
5
0.333
0.692
0.078
0.083
11 - Kvaløya til Loppa
20
9
45
8,550
7,523
88
1
0.111
0.485
1
0.111
0.057
1
0.111
0.024
6
0.667
0.434
0.223
0.156
12 - Vest-Finnmark
26
9
35
26,426
25,168
95
1
0.111
0.109
4
0.444
0.813
1
0.111
0.004
3
0.333
0.073
0.075
0.035
13 - Øst-Finnmark
20
16
80
75,305
74,902
99
3
0.188
0.816
6
0.375
0.073
6
0.375
0.106
1
0.063
0.004
0.045
0.007
Sum
448
227
51
362,649
341,953
94
76
0.335
64
0.282
21
0.093
66
0.291
Kilde: Diserud mfl. 2020. Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander – oppdatert status 2020. NINA Rapport 1926. Norsk institutt for naturforskning.
Figur 6.6. Genetisk status i 239 vassdrag, hvorav 227 regnes som laksebestander, i forhold til kvalitetselementet genetisk integritet. Ingen genetisk endring ble observert i 76 laksebestander (grønne sirkler), svake genetiske endringer ble indikert i 64 laksebestander (gule sirkler), moderate genetiske endringer ble påvist i 21 laksebestander (oransje sirkler), mens store genetiske endringer ble påvist i 66 laksebestander (røde sirkler). I de tolv vassdragene med forekomst av laks, men som ikke er definerte som egne laksebestander (elleve ikke klassifisert som laksevassdrag og ett sidevassdrag) ble ingen genetisk endring ble observert i 4 bestander (grønne åpne sirkler), svake genetiske endringer ble indikert i 5 bestander (gule åpne sirkler), moderate genetiske endringer ble påvist i 1 bestand (oransje åpen sirkel), mens store genetiske endringer ble påvist i 2 bestander (røde sirkler). For mer utfyllende forklaring av de fire kategoriene se Diserud mfl. (2020) der figuren er hentet fra.
Så langt er det beregnet innkryssingsnivå av rømt oppdrettslaks i 227 laksebestander i Norge (Diserud mfl. 2020). I tillegg er det undersøkt og dokumentert forekomster av laksunger med slektskap til oppdrettslaks i en rekke mindre vassdrag/sidebekker (Diserud mlf 2020; Pulg mfl. 2021). Det er også dokumentert at norsk oppdrettslaks har krysset seg inn i svenske bestander (Palm mfl. 2021).
6.2. Konsekvenser av innkryssing av rømt oppdrettslaks
Konsekvensen av at oppdrettsfisken rømmer og gyter sammen med villfisk, er at det oppstår en genetisk endring i de ville bestandene av laks. Gjennom flere internasjonale arbeid er det godt dokumentert at rømt oppdrettslaks kan gyte og etterlate seg avkom i naturen (f.eks. Sægrov mfl. 1997; Clifford mfl. 1998a; Crozier 2000; Glover mfl. 2013; Karlsson mfl. 2016; Sylvester mlf. 2018, 2019; Wringe mfl. o2018). Det er også dokumentert at innkryssing av rømt oppdrettslaks reduserer den genetiske variasjonen som finnes naturlig mellom bestandene (Skaala mfl. 2006; Glover mfl. 2013).
Atlantisk laks, en art med genetisk forskjellige bestander
Gjennom de siste 40 årene har det vokst fram en omfattende vitenskapelig litteratur om laks som dokumenterer en geografisk bestandsstruktur med store genetiske forskjeller mellom bestander i Nord-Amerika og Europa, og med regionale og lokale forskjeller innenfor kontinentene (Webb mfl. 2007; Bourret mfl. 2013, Ozerov mfl. 2017; Wennevik mfl. 2019). Geografisk oppdeling av en art, og variasjoner i livsmiljø, bidrar til utvikling av genetiske forskjeller mellom bestander. Siden vi ikke vet hvilke gener som nå eller i fremtiden er viktige for individer og bestander, er det et mål å bevare den naturlige genetiske variasjonen både innen og mellom bestander.
De siste årene har den vitenskapelige produksjonen som dokumenterer genetiske forskjeller mellom laksebestander økt betraktelig, delvis som følge av den raske utviklingen innenfor molekylærbiologi og statistikk. Etter hvert er det også vist og modellert at avkom av oppdrettslaks, både utplantet og naturlig produsert i elv, har lavere overlevelse i naturen og at oppdrettslaks derfor kan påvirke bestandene av villaks negativt der de krysser seg inn (Hindar mfl. 1991; Bourke mfl. 1997; McGinnity mfl. 1997; Verspoor 1997; Fleming mfl. 2000; Koljonen mfl. 2002; Fraser mfl. 2011; Skaala mfl. 2012; Besnier mfl. 2015; Reed mfl. 2015; Bolstad mfl. 2017, Wringe mfl. 2018, Skaala mfl. 2019; Wacker mfl. 2021; Bolstad mfl. 2021; Besnier mfl. 2022).
Hvor ulik er villaks og oppdrettslaks?
Den genetiske påvirkningen fra rømt oppdrettslaks på villaks er kompleks, og er avhengig av mange faktorer som varierer i tid og rom. Viktige, eller sannsynligvis viktige faktorer, inkluderer blant annet andel rømt oppdrettslaks i de ville bestandene (Glover mfl. 2013; Heino mfl. 2015; Karlsson mfl. 2016; Diserud mfl. 2022), deres gytesuksess (Fleming mfl. 1996, 2000), graden av genetisk forskjell mellom oppdrettet og vill laks (Fraser mfl. 2010; Islam mfl. 2021a, 2021b; Diserud mfl. 2022) og status for den ville bestanden (Glover mfl. 2012, 2013; Heino mfl. 2015; Diserud mfl. 2022). Det er også grunn til å tro at responsen hos de ville bestandene som følge av innkryssing av rømt oppdrettslaks vil variere fra bestand til bestand (Normandeau mfl. 2009; Fraser mfl. 2010; Harvey mfl. 2016a).
Genetiske forskjeller mellom vill- og oppdrettslaks kan oppstå gjennom tilfeldige prosesser (for eksempel begrenset antall stamfisk i hver generasjon, deres opprinnelse og tilpasning til oppdrettsmiljøet) og som resultat av målrettet seleksjon av egenskaper i avlsarbeidet. I Norge har man domestisert laksen i 50 år, med tidlig oppstart av målrettet avl (Gjedrem mfl. 1991; Gjøen & Bentsen 1997; Gjedrem 2010) for å endre kommersielt viktige egenskaper som tilvekst, kjønnsmodning, fettfordeling og sykdomsresistens. Seleksjon for en mer økonomisk produktiv oppdrettslaks foregår i avlsprogram som opprinnelig var basert på vill laks fanget i en rekke norske elver tidlig på 1970-tallet (Gjedrem mfl. 1991; Gjøen & Bentsen 1997). Under kontrollerte forhold blir de mest produktive familier og individer selektert basert på produksjonskriterier (f.eks. tilvekst), og disse individene blir benyttet til å føre stammen videre. På denne måten oppnår man en gradvis domestisering av laksen der viktige kommersielle trekk blir forandret i ønsket retning.
Glover mfl. (2017) oppsummerte den omfattende kunnskapen som nå er etablert om de genetiske forskjellene mellom oppdrettslaks og villaks. Oppdrettet og vill laks har vært sammenlignet med ulike metoder i en lang rekke vitenskapelige arbeider, og disse omfatter studier av genetisk variasjon med molekylære markører, eksperimentelle studier i laboratorium og kar hvor en har sammenlignet atferd, morfologi og fysiologi, og studier av overlevelse og vekst i et naturlig miljø. Noen eksperimentelle studier er også supplert med analyse av genuttrykk og fysiologisk målinger.
Sammenligning av genetisk variasjon og diversitet i oppdrettslinjer og ville laksebestander har blitt gjennomført over lengre tid med en rekke molekylære markører. De tidligste studiene var hovedsakelig basert på analyser av proteinkodende gen (Verspoor 1988; Cross & Challanain 1991; Mjølnerød mfl. 1997; Skaala mfl. 2005), der det er blitt vist genetisk forskjell mellom oppdrettslaks og de ville utgangsbestandene, og redusert nivå av genetisk variasjon i oppdrettslaks, målt som allelisk diversitet (antall varianter av et gen) og heterozygoti (andel individer med to ulike varianter av et gen, kontra individer med to like varianter av genet).
Seinere har DNA-markører blitt brukt til å sammenligne oppdrettslinjer og vill laks, for eksempel med mini- og mikrosatellittmarkører (Mjølnerød mfl. 1997; Clifford mfl. 1998a, b; Norris mfl. 1999; Skaala mfl. 2004), mikrosatellittmarkører kombinert med både mitokondrie DNA (mtDNA) (Karlsson mfl. 2010), og «single nucleotide polymorphism» (SNP) markører (Rengmark mfl. 2006; Karlsson mfl. 2011). Selv om resultatene fra disse studiene varierer noe, støtter de opp om konklusjonene fra tidligere analyser basert på proteinkodende gen; det er redusert genetisk variasjon hos den enkelte oppdrettslinje sammenlignet med de ville laksebestandene.
I en sammenligning av fem avlslinjer av oppdrettslaks med fire villaksbestander fra Neiden, Namsen, Vosso og Loneelva, viste alle de 12 undersøkte DNA-mikrosatellittmarkørene redusert allelisk variasjon i samtlige avlslinjer sammenlignet med de ville bestandene (Skaala mfl. 2004). I gjennomsnitt hadde avlslinjene 58 % av den alleliske variasjonen sammenlignet med prøver av villaks, og dette kan forklares av begrenset effektiv bestandsstørrelse i oppdrettslinjene. Samtidig var estimatene for genetisk forskjell flere ganger høyere mellom de ulike avlslinjene enn mellom de ville laksebestandene. Andre studier har vist at tap av genetisk diversitet i oppdrettslinjer er mer komplekst enn tidligere antatt (Karlsson mfl. 2010), men det kan likevel konkluderes med at oppdrettslaks har redusert genetisk variasjon i forhold til ville laksebestander. Dette samsvarer med tilsvarende observasjoner fra andre domestiserte organismer (se Araki & Schmid 2010), og kan forklares med at det er et begrenset antall familier/individer som bidrar til hver generasjon i et avlsprogram. Den effektive bestandsstørrelsen i norske oppdrettslinjer er tidligere blitt estimert til 33–125 individer (Mork mfl. 1999), noe som teoretisk sett skal føre til lav til moderat innavl over den aktuelle avlsperioden. Avlsprogrammene i dag tar derfor hensyn til genetisk slektskap mellom individene som bidrar til neste generasjon, med det mål å forhindre innavl.
På grunn av et målrettet avlsarbeid er det ikke uventet at oppdrettslaks vokser bedre enn villaks i et oppdrettsmiljø (Einum & Fleming 1997; Thodesen mfl. 1999; Fleming mfl. 2002; Glover mfl. 2009; Solberg mfl. 2013a, b; Harvey mfl. 2016a, b, c, d). I et oppdrettsmiljø vil vekstforholdet mellom oppdrettslaks og villaks være 2-5:1 (Glover mfl. 2017). Også i et naturlig miljø kan oppdrettslaks ha større vekst enn villaks (Johnsson & Björnsson1994; Einum & Fleming 1997; McGinnity mfl. 1997, 2003; Skaala mfl. 2012, 2019; Jonsson & Jonsson 2017; Bolstad mfl. 2017, 2019), men ikke i samme grad som i oppdrettsmiljø (typisk vekstforhold mellom oppdrettslaks og villaks i naturen er 1,0-1,3:1). Oppdrettslaks har et forhøyet vekstpotensial, relativt til villaks, som den ikke får utnyttet i naturen, og det er en kombinasjon av plastisitet (ulik mattilgang mellom disse miljøene) og naturlig seleksjon mot hurtigvoksende oppdrettslaks (økt dødelighet) som er årsaken til de små vekstforskjellene mellom oppdrettslaks og villaks i naturen (Glover mfl. 2018). Vekst hos hybrider mellom oppdrettslaks og villaks, inkludert tilbakekrysninger (Perry mfl. 2020), er ansett å være additivt (mellomliggende til vekst hos vill og oppdrett), men ikke-additive genetiske effekter knyttet til vekst er nylig dokumentert i Atlantisk laks (Besnier mfl. 2020).
Mange egenskaper som ikke inngår direkte i avlsarbeidet, som aggresjon, stress- og temperaturtoleranse, kan også bli endret hos oppdrettslaksen gjennom avlsprosessen (Fleming & Einum 1997; Houde mfl. 2010; Debes & Hutchings 2014; Solberg mfl. 2016). Årsaken er at målrettet seleksjon for blant annet tilvekst påvirker både aggresjon og andre egenskaper, for eksempel hormonregulering og atferd. I eksperimentelle studier er det vist at tilførsel av veksthormon øker appetitten (Johnsson & Björnsson 1994; Jönsson mfl. 1996), aggresjon og aktivitet (Jönsson mfl. 1998), altså atferd som sannsynligvis påvirker overlevelse i naturen (Johnsson mfl. 1996; Jönsson mfl. 1996; Martin-Smith mfl. 2004). Det er derfor ikke overraskende at oppdrettslaks er ulik villaks i flere egenskaper som påvirker overlevelse i naturen, slik som tilvekst, aggresjon, dominans og antipredatoratferd (Einum & Fleming 1997; Fleming & Einum 1997; Johnsson mfl. 2001; Fleming mfl. 2002; Houde mfl. 2010). I tillegg er det avdekket genetiske forskjeller mellom vill- og oppdrettslaks i egenskaper som kjøttfarge, kjønnsmodning og fettinnhold (Glover mfl. 2009), reaksjonsnormer (Darwish & Hutchings 2009; Solberg mfl. 2013a, b), morfologi (Fleming & Einum 1997; Perry mfl. 2019), stresstoleranse (Solberg mfl. 2013a) og en rekke andre egenskaper (Glover mfl. 2017). Selv om rømt oppdrettslaks rapporteres å ha mer prikker en villaks, er det derimot ikke avdekket genetiske arvelige forskjeller i prikkemengde mellom dem (Jørgensen mfl. 2018), og det antas å være forhold i oppdrettsmiljøet som gjør at rømt oppdrettslaks har mer prikker en villaks i naturen.
Det er også dokumentert genetiske forskjeller i genuttrykk mellom laks av ville- og oppdrettsforeldre, samt hybrider av disse (Roberge mfl. 2006, 2008; Solberg mfl. 2012; Bicskei mfl. 2014, 2016). Resultatene er også interessante fordi hybridene ikke alltid hadde et genuttrykk som lå mellom foreldrebestandene. Hybridene hadde i noen tilfeller genuttrykk som lå høyere enn verdiene målt for vill- og oppdrettsfisk, noe som tyder på ikke-additive genetiske effekter. Dokumentasjon av ikke-additive genetiske effekter betyr i praksis at innkryssing av oppdrettsfisk i ville bestander i noen tilfeller kan gi uventede effekter. Hvilken endring man får i genuttrykk hos hybrider av vill- og oppdrettslaks er avhengig av hvilke ville bestander som krysses inn (Normandeau mfl. 2009; Fraser mfl. 2010).
Det er ikke avdekket noen store forskjeller i resistens mot lakselus, ILA (infeksiøs lakseanemi) eller furunkulose mellom vill- og oppdrettslaks (Glover & Skaala 2006; Glover mfl. 2006a, b). Det er heller ikke avdekket genetiske forskjeller i deformiteter hos smolt av oppdretts- og villaks (Fjelldal mfl. 2009). For infeksiøs pankreasnekrose er det funnet genetiske markører (såkalt QTL – Quantitative Trait Locus) som forklarer en stor grad av toleransen for sykdommen (Houston mfl. 2008; Moen mfl. 2009) og denne kunnskapen er inkludert i avlsarbeid hos flere avlsselskaper. Seleksjon for sykdomsresistens har derimot vært praktisert ulikt for de ulike oppdrettslinjene, og dette vanskeliggjør sammenligning mellom linjer.
Det er vist at avkom av rømt oppdrettslaks har lavere overlevelse i naturen enn avkom av villaks, men mekanismene bak dette er komplekse og krevende å avdekke. Tidligere forsøk i naturen (Skaala mfl. 2014) og under eksperimentelle forhold (Fleming & Einum 1997; Debes & Hutchings 2014; Solberg mfl. 2015) har ikke påvist forskjeller i predasjonsdødelighet. I nyere tid er det dokumentert gjennom forsøk i et semi-naturlig miljø at avkom av rømt oppdrettslaks er et lettere bytte for predatorer som større ørret, noe som underbygger deres lavere overlevelse i naturen (Solberg mfl. 2020). Dette resultatet støttes av det faktum at redusert antipredatoratferd (Einum & Fleming 1997; Johnsson mfl. 2001; Houde mfl. 2010), i tillegg til økt toleranse for predasjonsrelatert stress (Debes & Hutchings 2014), tidligere har blitt dokumentert i eksperimentelle forsøk. Det er også dokumenter forskjeller i kromosomale strukturvarianter mellom oppdrettslaks og villaks, linket til gener uttrykt i hjernen som er knyttet til nevrologiske lidelser og endret atferd (Bertolotti mfl. 2020). En annen mulig forklaring hvorfor oppdrettslaks er et lettere bytte for predatorer er at de ikke ser like godt som villaksen, og en reduksjon i øyestørrelse hos oppdrettslaks kontra villaks er observert under eksperimentelle forhold (Perry mfl. 2021).
En oppsummering av vitenskapelig litteratur viser at det er til dels store genetiske forskjeller mellom vill- og oppdrettslaks i kvantitative egenskaper som har direkte eller indirekte betydning for overlevelsen av laks i naturen, og som kan gjøre innkryssede laksebestander mer sårbare mot framtidige klimaendringer (Debes mfl. 2021). Det er grunn til å tro at de genetiske forskjellene kommer til å øke for hver avlsgenerasjon. Dette har blitt observert for tilvekst, der forskjellen mellom vill- og oppdrettslaks under oppdrettsbetingelser økte ytterligere fra generasjon 8 til 10 (Glover mfl. 2009; Solberg 2013a, b). Genetiske endringer i villaks som følge av økologisk interaksjon med oppdrettslaks, gjennom f.eks. overføring av sykdom og parasitter, eller gjennom konkurranse er også en aktuell problemstilling. Blant annet har tilstedeværelse av hybrider vist seg å ha en negativ påvirkning på overlevelse til villfisk (Robertsen mfl. 2018). Observert og potensielle genetiske endringer grunnet økologiske interaksjoner ble oppsummert av Bradbury mfl. (2020), og forfatterne identifiserer flere eksempler på endret seleksjonspress på villaks grunnet oppdrettsaktivitet, inkludert eksempler på genetiske endringer grunnet overføring av sykdom og parasitter.
Rask veksthastighet hos avkom av oppdrettslaks kan også ha konsekvenser for økosystemnivå. Nylig viste Cucherousset mfl. (2021) at laksunger med kunstig økt vekstpotensiale hadde andre effekter i et bekkeøkosystem enn vanlige laksunger. Laksunger med økt vekstpotensiale viste endringer i habitatbruk, kroppsform og ekskresjonsrate, og disse endringene var forbundet med effekter på invertebratsamfunnet og endringer i økosystemfunksjoner som primærproduksjon og nedbryting (Cucherousset mfl. 2021). Dette tyder på at rømming av oppdrettslaks med økt vekstpotensiale kan ha effekter på økosystemnivå i tillegg til effekter på villaks.
Genetisk påvirkning fra rømt oppdrettslaks – hva forteller empiriske data oss?
Ved hjelp av ulike biokjemiske og molekylærgenetiske metoder er det vist at rømt oppdrettslaks gyter i elver. Ved undersøkelser av et pigment i rogn og yngel, som reflekterer ulik diett hos villaks og oppdrettslaks, fant Lura & Sægrov (1991) at rømt oppdrettslaks faktisk produserte levedyktig avkom i en elv. I en skotsk undersøkelse fant Webb mfl. (1993) et pigment fra rømt oppdrettslaks i 14 av 16 undersøkte elver, med et gjennomsnittlig innslag på 5,1 % fra rømt fisk. I Vosso ble bidraget fra rømt oppdrettslaks estimert til opp mot 80 % ved denne metoden (Sægrov mfl. 1997). Bevis for at rømt oppdrettslaks produserte levedyktig avkom ble også funnet i Irland ved hjelp av genetiske markører (Clifford mfl. 1998a; Crozier 1993, 2000). Også langt utenfor det naturlige utbredelsesområdet til den atlantiske laksen, i British Columbia, Canada, er det vist at rømt oppdrettslaks kan produsere levedyktig avkom (Volpe mfl. 2000).
Det første genetiske studiet for å undersøke om norske villaksbestander har endret seg genetisk over tid som følge av innkryssing av rømt oppdrettslaks ble publisert av Skaala mfl. (2006). Her ble det laget DNA-profiler for de sju laksebestandene Namsen, Etne, Opo, Vosso, Granvin, Eio og Håelva. Det ble benyttet gamle skjellprøver og materiale innsamlet i nyere tid, etter lengre tids innslag av rømt oppdrettslaks. Håelva på Jæren ligger i en region der det nesten ikke er lakseoppdrett, og andelen rømt oppdrettslaks i villaksbestanden har vært lav, trolig under 5 %. I Håelva ble det ikke funnet endring i de genetiske profilene. I tre andre bestander, Opo, Vosso og Eio i Hordaland, ble det funnet signifikante endringer i de genetiske profilene over tid. Mer overraskende var det likevel at det ikke ble funnet endringer i Etneelva, Namsen eller Granvinelva, som alle hadde hatt høye andeler rømt oppdrettslaks i gytebestandene, permanent eller periodisk
Som en videreføring av dette arbeidet (Skaala mfl. 2006) ble det gjort en mer omfattende analyse av 21 bestander der historiske og nye prøver ble analysert for flere mikrosatellittmarkører. Undersøkelsen omfattet elver fra hele landet (Glover mfl. 2012) og påviste genetiske forandring over tid i 6 av 21 elver, mens i 15 av bestandene ble det ikke funnet genetiske forandringer. Som i den første undersøkelsen utført av Skaala og kolleger (2006), var det noen bestander med høye innslag av rømt oppdrettslaks på gyteplassene der det ikke ble påvist forandringer.
I de seks bestandene der det ble påvist forandring, har det vært registrert rømt oppdrettslaks i større eller mindre grad. I tillegg ble det funnet nye genvarianter som indikerer at forandringene i disse seks elvene hovedsakelig skyldes genflyt fra andre kilder. Den genetiske forskjellen mellom disse seks bestandene er også blitt redusert over tid. Basert på alle data, ble det konkludert med at innkryssing av rømt oppdrettslaks er hovedårsaken til forandringene. Dette er i tråd med simuleringer fra modeller som viser at innkryssing av rømt oppdrettslaks vil redusere genetisk differensiering mellom bestander over tid (Mork 1991; Besnier mfl. 2011).
Det er kjent at mikrosatellittmarkører i noen tilfeller vil underestimere innkryssing av rømt oppdrettslaks i ville bestander på grunn av signalstøy når en villaksbestand mottar oppdrettsfisk fra flere ulike avlsbestander (Besnier mfl. 2011). Det betyr at antall elver som er påvist å være genetisk påvirket i denne studien (Glover mfl. 2012), og omfanget av de genetiske forandringene i disse elvene, må betraktes som et minimumsestimat. For å få et mer presist svar på omfanget av innkryssing av rømt oppdrettslaks ble en studie gjennomført basert på SNP-markører utviklet for å kunne identifisere oppdrettslaks og skjelne dem fra villaks (Karlsson mfl. 2011). Disse SNP-markørene gir mer presis informasjon om genetiske forandringer forårsaket av rømt oppdrettslaks, og er i mindre grad påvirket av problematikken ved at genetisk forandring i den enkelte villaksstamme er vanskelig å påvise når innkryssing skjer via oppdrettslaks fra forskjellige avlslinjer (Besnier mfl. 2011).
I en studie av 20 laksebestander langs hele norskekysten (Glover mfl. 2013) ble disse SNP-markørene brukt til å estimere prosent innkryssing av rømt oppdrettslaks. Resultatene viste at det genetiske bidraget til noen bestander var nesten 50 %, mens estimert innkryssing av oppdrettslaks var mye lavere i de fleste undersøkte elvene. Arbeidet støttet opp om konklusjonene til de tidligere publikasjonene med andre markørtyper (Skaala mfl. 2006; Glover mfl. 2012). I tillegg til at det ble dokumentert genetiske forandringer i noen villaksbestander på grunn av innkryssing av rømt oppdrettslaks, viser alle disse tre studiene (Skaala mfl. 2006; Glover mfl. 2012, 2013) at den genetiske forskjellen mellom noen ville bestander er blitt mindre over tid. Dette kan tilskrives at de ville bestandene som har hatt en betydelig innkryssing av rømt oppdrettslaks, blir mer lik oppdrettslaksen – og dermed også mer lik hverandre.
Parallelt med dette arbeidet utarbeidet Karlsson mfl. (2014) en statistisk metode der prosentvis innkryssing kunne estimeres i enkeltindivider og uavhengig av om det fantes en historisk prøve av den aktuelle bestanden. Metoden brukte et estimat av andelen «villgenom» i bestanden (omtalt som P(wild) = mengde arvestoff som stammer fra ville foreldre kontra oppdrettsforeldre) for å beregne innkryssingen av rømt oppdrettslaks (Karlsson mfl. 2014).
Denne metodeutviklingen gjorde det mulig å estimere innkryssing i et stort antall ville laksebestander. Karlsson mfl. (2016) studerte 147 laksebestander som til sammen representerer tre fjerdedeler av de ville lakseressursene i Norge, og analyserte genetisk mer enn 20 000 laks som var klekket i naturen og derfor så ut som villaks. Den gjennomsnittlige genetiske innkryssingen i bestandene av voksen laks var 6,4 % med et spenn fra 0 % til over 40 % innkryssing i enkelte bestander. Forskerne fant også at den genetiske innkryssingen generelt var mindre i nasjonale lakseelver og nasjonale laksefjorder enn i bestander uten denne særlige beskyttelsen (Karlsson mfl. 2016). I et materiale av 109 bestander der kun voksen laks klekket i naturen ble inkludert i datamaterialet, var den genetiske innkryssingen signifikant forskjellig fra 0 i omtrent halvparten av bestandene (Karlsson mfl. 2016). Det samme resultatet fikk Karlsson mfl. (2016) om de også tok med et større antall elver, der enkelte estimater var basert på prøver av ungfisk.
Data fra de 20 bestandene som var publisert av HI i Glover mfl. (2013), samt data fra NINA for et større antall bestander (Karlsson mfl. 2016), ble vurdert sammen for å dokumentere genetisk status for ville laksebestander til «Kvalitetsnorm for ville bestander av atlantisk laks (Salmo salar)», som er en forskrift av 20. september 2013 hjemlet i Naturmangfoldloven. I henhold til Kvalitetsnormen vurderte forskere fra HI og NINA i 2016 genetisk status i 125 laksebestander, basert på estimater for innkryssing av rømt oppdrettslaks i bestanden (Diserud mfl. 2016). I senere utgaver av denne rapporten ble antallet bestander økt til 175 i 2017 og deretter til 225 norske laksebestander i 2019 (Diserud mfl. 2017, 2019b). Den hittil mest omfattende vurderingen av genetisk innkryssing omfatter 227 bestander i 2020 og ga følgende resultat (pr. Diserud mfl. 2020): ingen genetiske forandringer (76 vassdrag), svake genetiske forandringer (64 vassdrag), moderat genetiske forandringer (21 vassdrag) og store genetiske forandringer (66 vassdrag). Vurderingen av genetisk innkryssing i disse 227 bestandene dokumenterer at 2/3 av de ville laksebestandene har blitt påvirket av rømt oppdrettslaks. Arbeidet dekker flere tidligere undersøkte elver (Skaala mfl. 2006), og blant annet er det nå påvist store genetiske endringer i Etne og Granvin, og moderate genetisk endringer i Namsen. I tillegg ble tolv vassdrag med forekomst av laks, men som ikke er definerte som egne laksebestander (elleve ikke klassifisert som laksevassdrag og ett sidevassdrag) undersøkt: ingen genetisk endring ble observert i fire vassdrag, svake genetiske endringer ble indikert i fem vassdrag, moderate genetiske endringer ble påvist i ett vassdrag, mens store genetiske endringer ble påvist i to vassdrag. En annen undersøkelse av 11 mindre kystvassdrag på Vest- og Sørlandet viste i enkelte tilfeller svært høye nivåer av innkryssing (> 90%), og tyder på at mindre vassdrag kan fungere som et formeringssted for rømt laks (Pulg mfl. 2021). Det er også dokumentert at norsk oppdrettslaks har krysset seg inn i svenske bestander (Palm mfl. 2021). Tilsvarende estimater fra andre land med lakseoppdrett foreligger fra Canada (Sylvester 2018, 2019; Wringe 2018) og Skottland (Gilbey mfl. 2021). I Skottland er genetisk integritet til ungfisk fra 237 lokaliteter nylig klassifisert, og tegn på innkryssing av rømt oppdrettslaks ble dokumentert i laks fra 23% av lokalitetene.
Eksperimenter i naturen har vist seleksjon mot avkom av rømt oppdrettslaks, både i Irland (McGinnity mfl. 1997, 2003) og Norge (Fleming mfl. 2000; Skaala mfl. 2012, 2019). En skulle derved forvente at når én og samme årsklasse (dvs. laks fra samme klekkeår) studeres, så vil et materiale av ungfisk vise høyere genetisk påvirkning av oppdrettslaks enn et materiale av voksen laks. Dette er også det Karlsson mfl. (2016) fant gjennom analyser av ungfisk som var innsamlet noen år før et materiale av voksen laks fra de samme elvene: ungfisken hadde i gjennomsnitt 2,5 % høyere grad av genetisk innkryssing enn den voksne laksen. Seleksjon mot avkom av rømt oppdrettslaks har også blitt dokumentert i etterkant av en større rømmingsepisode på Newfoundland i Canada (Wringe mfl. 2018; Sylvester mfl. 2019) og ved å studere gyteårsklasser av ungfisk ved ulik alder i Altaelva (Wacker mfl. 2021).
Det foreligger et solid grunnlag for å hevde at rømt oppdrettslaks kan påvirke villfiskbestander negativt (Glover mfl. 2017). Den beste dokumentasjonen av effekter på overlevelse og viktige egenskaper kommer fra eksperimentelle studier utført i tre kontrollerte elver (McGinnity mfl. 1997, 2003; Fleming mfl. 2000; Skaala mfl. 2012, 2019), samt nyere studier som viser sammenhenger mellom molekylært nivå på innkryssing og viktige livshistorietrekk i ville bestander (Bolstad mfl. 2017, 2021; Wacker mfl. 2021; Besnier mfl. 2022).
En direkte og informativ tilnærming til å forstå effektene av at rømt oppdrettslaks krysser seg inn i villaksbestander er å sammenligne tilvekst, atferd og overlevelse hos sammenblandede familiegrupper av oppdrettslaks, villaks og hybrider under like forhold, såkalte «common garden-studier», i et naturlig miljø. Dette kan innebære utplanting av lakserogn eller utsetting av ungfisk fra DNA-identifiserbare familier av oppdrettslaks, villaks og hybrider dem imellom (McGinnity mfl. 1997, 2003; Skaala mfl. 2012, 2019; Crowley mfl. 2022) eller utsetting av kjønnsmodne individer med kjente genetiske profiler (Fleming mfl. 2000) i naturlig elvemiljø, der alle avkom i ulike livsstadier fra rogn til kjønnsmodning i ettertid kan identifiseres ved genetiske markører. En annen tilnærming vil være å genotype alle anadrome foreldre i en bestand og deretter en representativ del av den utvandrende smolten. Dersom den genetiske profilen på foreldrefisk omfatter grad av innkryssing, vil man kunne beregne individuell reproduktiv suksess hos individer med ulik grad av innkryssing. En slik tilnærming forutsetter god kontroll på både oppvandrende laks og utvandrende smolt, som for eksempel Imsa og Guddalselva.
Det første common garden prosjektet som er gjennomført på dette feltet, ble utført i Burrishoole, Irland (McGinnity mfl. 1997, 2003; Ferguson mfl. 2002). I dette prosjektet ble tilvekst, overlevelse og populasjonsdynamikk hos villaks, oppdrettslaks og hybrider undersøkt gjennom to generasjoner. Et stort antall individer fra mange familier av villaks, oppdrettslaks, første- og andregenerasjonshybrider og førstegenerasjonshybrider tilbakekrysset til henholdsvis villaks og oppdrettslaks, ble plantet ut i tre årsklasser som øyerogn ovenfor fiskefellen i Burrishoole. Tilsvarende grupper ble satt ut som smolt i elven for å studere vekst og overlevelse i sjøfasen.
En omfattende innsats med innsamling og genotyping for å identifisere opphavet til alle individ ble gjennomført fra yngel til gytefisk som kom tilbake fra havet etter ett og to år i sjø. I alle tre årsklassene hadde oppdrettslaksen signifikant lavere representasjon enn villaksen i prøver av 0+ parr. Ikke overraskende viste det seg at oppdrettslaksen vokste bedre enn villaksen, og at den større oppdrettsparren fortrengte den ville parren nedover elva gjennom konkurranse. Selv om ungfisk av oppdrettslaks vokste bedre og fortrengte en del av den ville ungfisken, var smoltproduksjonen av oppdrettslaks bare henholdsvis 34, 34 og 55 % sammenlignet med villaksen i de tre årsklassene. Den gjennomsnittlige gjenfangsten etter sjøoppholdet var 0,3 % for oppdrettslaks og 8 % for villaks utsatt som smolt. Overlevelse hos hybridene viste seg ofte å ligge mellom villaks og oppdrettslaks.
Et lignende prosjekt ble gjennomført ved NINAs feltstasjon på Ims (Fleming mfl. 2000) der alle ned- og oppvandrende fisk kontrolleres i en toveis fiskefelle i elven Imsa. Her ble det satt ut kjønnsmodne villaks fra Imsa og oppdrettslaks med kjente genetiske profiler ovenfor fiskefellen. De to gruppene hadde lignende vandringsmønster og valgte de samme gyteplassene i elven. Vill hannlaks var mer aktive i kurtisering av hunnlaksen enn oppdrettshannene var, og hadde dessuten mindre restgonader etter gyting enn oppdrettshannene hadde. Gytesuksessen var mye lavere hos oppdrettslaksen både for hanner (24 %) og hunner (32 %) sammenlignet med villaksen i et samtidig forsøk i store gytebassenger med steinbunn. Gjennom ferskvannsfasen endret andelen av genotyper seg ytterligere i disfavør av oppdrettslaksen, og hoveddelen av oppdrettsbidraget var representert i form av hybrider, produsert av oppdrettshunner og ville hanner. Studier av dietten viste betydelige overlapp i næringsvalg, noe som viser næringskonkurranse mellom oppdrettsavkom og villaksyngel. Den totale smoltproduksjonen for elven var 28 % lavere enn forventet ut fra rognmengde og det observerte forholdet det normalt har vært i Imsa mellom mengde egg og antall smolt (Jonsson mfl. 1998). Det var også en tilsvarende reduksjon i forventet smoltproduksjon av den ville gytefisken. Oppdrettslaksen smoltifiserte og vandret ut tidligere og ved lavere alder enn villaksen. I motsetning til resultatene fra Burrishooleprosjektet med utsatt smolt, fant en i Imsaprosjektet ingen forskjell mellom gruppene i sjøoverlevelse. Senere eksperimenter på Ims med utsetting av smolt av villaks, oppdrettslaks og førstegenerasjons krysninger mellom dem (1996-1998), viste signifikant lavere gjenfangst av oppdrettslaks (Hindar mfl. 2006). Utsatt smolt av oppdrettslaks har også vist høyere feilvandringsrate enn utsatt smolt av Imsalaks på Ims (Jonsson mfl. 2003; Jonsson & Jonsson 2017). Hybrider mellom oppdrettslaks og villaks hadde også høyere feilvandringsrate sammenlignet med den ville Imsalaksen (Jonsson & Jonsson 2017). Retningen på hybridiseringen hadde betydning ved at hybrider med vill mor hadde lavere feilvandring enn hybrider med vill far (Jonsson & Jonsson 2017).
Ved Havforskningsinstituttets feltstasjon i Guddalselva i Hardanger ble det initiert et prosjekt basert på oppsettet for Burrishooleprosjektet. Der ble definerte familiegrupper av vill og oppdrettet laks, og hybrider mellom disse, plantet ut som rogn (Skaala mfl. 2012). All foreldrefisk var genotypet med DNA-mikrosatellittmarkører. Dermed kunne alle individ som var satt ut som øyerogn i seks årsklasser, ca. 150 familier i et «common garden-studie», identifiseres til familie. Det ble samlet inn juvenil laks av alle årsklassene fra elvehabitatet, og tilvekst, overleving og diettvalg ble undersøkt for hver familie.
Resultatene fra de tre første årsklassene viste en overlevelse (fra utplantet egg til smolt) som varierte mellom 0,17 og 6,4 % for de 69 forskjellige familiene (Skaala mfl. 2012). Resultatene viste at eggstørrelsen hadde stor betydning for overlevelsen. I dette studiet hadde oppdrettslaksene større egg enn villaksene. Dette bidro til at noen oppdrettsfamilier hadde en høy overlevelse fra egg til smolt i forsøket. Da det ble kontrollert for eggstørrelse ved å sammenligne overlevelse til en oppdrettsfamilie og dens halvsøskenfamilie der fars bidrag kom fra en vill hann, viste det seg at 16 av 18 halvsøskenfamilier hadde høyere overlevelse med bidrag fra vill far kontra oppdrettsfar. Dette viser at det er additiv genetisk variasjon for overlevelsen i et naturlig miljø og at villaks har høyere overlevelse, selv om bildet kan være noe mer komplisert når det tas hensyn til eggstørrelse. I tillegg til lavere overlevelse når det tas hensyn til eggstørrelse, hadde smolt av oppdrettsforeldre noe høyere vekst enn hybrider og villaks i elven (forholdstall oppdrett:vill = 1,0-1,3:1).
Resultatene fra de tre siste årsklassene i perioden 2008–2016 (Skaala mfl. 2019) har i stor grad støttet resultater fra de tre første årsklassene, og det ble observert langt lavere overlevelse hos avkom hos oppdretts- og hybridlaks i forhold til villaks i ferskvann (1,8 % overlevelse hos oppdrettslaks mot 3.8 % overlevelse hos villaks). I tillegg ble det satt ut to årsklasser med oppdretts-, hybrid- og villsmolt i elven for å studere sjøoverlevelsen. Her fant man lavere sjøoverlevelse hos avkom av oppdrettslaks enn hos avkom fra hybrider (gjennomsnittlig sjøoverlevelse) og villaks (0.41 % overlevelse hos oppdrettslaks mot 0.94 % overlevelse hos villaks). Smolt med oppdrettsforeldre var litt større og vandret ut tidligere i sesongen enn villsmolt. Siden en del avkom av oppdrettslaks også overlever gjennom ferskvannsfasen, vil disse konkurrere om tilgjengelige næringsressurser, som er begrenset i ferskvann. Derfor vil avkom av rømt oppdrettslaks også kunne bidra til en redusert produksjon av villsmolt gjennom næringskonkurranse i tillegg til redusert overlevelse på grunn av innkryssing og endrede fitnessrelaterte egenskaper.
I en nyere studier fra Canada ble yngel av oppdrett, vill og hybrid bakgrunn sluppet ut ved tre lokaliteter i Newfoundland i 2018 og gjenfanget som 0+ parr etter deres første sommer (Crowley mfl. 2022). Overlevelsen var generelt høy i alle gruppene, men lavere overlevelse ble observert hos ungfisk med oppdrettsmødre, i forhold til ungfisk med ville mødre. Den ville ungfisken hadde tilnærmet lik overlevelse som deres maternale halvsøsken (hybrider med vill mor). Lavest overlevelse ble sett hos hybrider av oppdrettsmødre, og ikke hos ungfisk med to oppdrettsforeldre.
Når avkom fra oppdrettslaks, villaks og hybridene konkurrerer om de samme byttedyrene i elven, vil innkryssing av oppdrettslaks kunne redusere den naturlige produksjonen av villsmolt på grunn av konkurranse (Fleming mfl. 2000; Skaala mfl. 2019). Når vi samtidig ser at avkom av oppdrettslaks også har lavere sjøoverlevelse, vil det i sum også kunne redusere antall laks som kommer tilbake til elven. Gyting mellom oppdrettslaks som resulterer i avkom med 100% oppdrettsbakgrunn, vil også kunne påvirke den naturlige produksjonen av villsmolt negativt gjennom næringskonkurranse i elven, til tross for at dette ikke fører til genetisk innkryssing. Produksjon av avkom med 100% oppdrettsbakgrunn ble observert etter at kjønnsmoden oppdrett- og villaks ble satt ut i Imsa (Fleming mfl. 2020), og det er også observert i flere Canadiske elver i etterkant av en større rømmingsepisode (Sylvester mfl. 2018; Wringe mfl. 2018). Avkom av rømt oppdrettslaks som kjønnsmodnes i elv som dverghanner (både avkom med 100% oppdrettsbakgrunn, men også hybrider), kan føre til at innkryssing av oppdrettslaks og effekten av innkryssing framskyndes (Holborn mfl. 2022).
Utviklingen innenfor genomforskningen de siste årene gir stadig bedre muligheter for å vurdere de biologiske konsekvensene av innkryssing av oppdrettslaks i ville bestander. Man vil kunne identifisere hvilke gener som er involvert i de biologiske forskjellene man observerer mellom oppdretts- og villaks. En relativt ny studie identifiserte områder i genomet/arvestoffet som var knyttet til overlevelse i naturen, selv om det er uvisst hvilke gener som var involvert (Besnier mfl. 2015). Det er forventet at nye resultater fra QuantEscapeII-prosjektet (NFR-prosjekt 2016-2020) vil bidra til å kartlegge hvordan endringer i genomet oppstår som følge av innkryssing, og hvilke biologiske konsekvenser dette kan få for ville bestander.
Inntil nylig forelå det ikke dokumentasjon av forandringer i livshistorietrekk og demografi i villaksbestander som konsekvens av genetisk innkryssing. Dette kan skyldes at en ikke har hatt tilstrekkelig gode verktøy til å studere slike endringer, i tillegg til at det er først relativt nylig at man har klart å dokumentere og kvantifisere innkryssing av rømt oppdrettslaks i bestander med bruk av genetiske markører. En annen årsak er at varierende forhold i både ferskvann og i havet, påvirker både livshistorie og demografiske parametere – noe som gjør at det er krevende å identifisere og kvantifisere den relative betydningen av de ulike påvirkningsfaktorene. Betydningen av slike endringer i havklima for laksens tilvekst er nylig dokumentert av Vollset mfl. (2022) og av Harvey mfl. (2020) som identifiserte både sjøtemperatur, mengde zooplankton og intensiteten av lakselusinfestasjon som viktige påvirkningsfaktorer for laksens tilvekst i havet.
Det første arbeid som dokumenterte forandringer i livshistorietrekk hos villaks ble publisert i 2017 og viste endringer i tilvekst og alder ved kjønnsmodning i et stort antall bestander som følge av genetisk innkryssing av rømt oppdrettslaks (Bolstad mfl. 2017). Bolstad mfl. (2017) studerte 62 ville laksebestander med individer av ulik grad av genetisk slektskap til oppdrettslaks. Det genetiske slektskapet ble målt ved bruk av molekylærgenetiske metoder (Karlsson mfl. 2011, 2014). Studien viser at individer med høy grad av genetisk slektskap til oppdrettslaks har endret sjøalder og størrelse ved kjønnsmodning. Disse endringene var forskjellige mellom kjønn og for ulike typer laksestammer. I storlakselver (elver med overvekt av flersjøvinterlaks) ble det observert en økning i antall hunnlaks som returnerer fra havet etter to vintre (tosjøvinterlaks), og en nedgang i antall som returnerer etter en og tre vintre, med økende grad av genetisk slektskap til oppdrettslaks. For hannlaks i de samme elvene var det en økning i ensjøvinterlaks og en tilsvarende nedgang i to- og tresjøvinterlaks. I smålakselver (elver dominert av ensjøvinterlaks) ble det ikke observert slike endringer. Derimot ble det observert en større økning i aldersbestemt størrelse for laksen i smålakselvene enn for laksen i storlakselvene med økende grad av slektskap til oppdrettslaks. I smålakselvene økte vekten med 19 % fra 0 til 100 % genetisk slektskap til oppdrettslaks.
Studien undersøkte også effekten på laks i Finnmark (Bolstad mfl. 2017). Laksen i Finnmark er mer i slekt med laksen på Kolahalvøya i Russland enn med resten av den norske laksen. På grunn av lavere utvalgsstørrelse var resultatene for Finnmark mindre sikre enn for resten av landet, men det kan med sikkerhet fastslås at det er ulik effekt av genetisk slektskap til oppdrettslaks i de to regionene. Spesielt ble det observert en kraftig effekt på antall vintre i sjøen for hannlaks i smålakselvene i Finnmark, som i økende grad ble kjønnsmodne etter to vintre i istedenfor etter en. Det ble også observert en dramatisk økning i aldersbestemt størrelse i smålakselvene i denne regionen, med 24 % vektøkning fra 0 til 50 % genetisk slektskap til oppdrettslaks. Det er viktig å påpeke at de målte endringene i denne studien er underestimat (se Bolstad mfl. 2017 for utdypende forklaring).
I 2021 publiserte Bolstad mfl. en utvidet studie hvor 105 laksebestander ble undersøkt. Studien viser at økende nivå av innkrysning fører til økt vekstrate gjennom hele laksen livssyklus, i tillegg til lavere alder både ved smoltifisering og kjønnsmodning. Kun små forskjeller i vekstrate ble observert i smolt med økende grad av innkrysning, men kombinert med lavere alder ved smoltifisering, tyder studien på at økt vekst bidrar til at individer med økende grad av slektskap til oppdrettslaks forlater elven tidligere enn villaksen (noe som samsvarer med tidligere common garden studier (Skaala m fl 2019)). I sjø var effekten av innkryssing forskjellig mellom ulike bestander og det var i de saktevoksende bestandene at økende grad av slektskap til oppdrettslaks førte til størst økning i vekst. Studien viser at i hurtigvoksende bestander kan innkryssing derimot føre til lavere vekst.
I en studie som viste at økende grad av innkrysning fører til lavere overlevelse hos ungfisk, fant Wacker mfl. (2021) også kun en moderat økning i tilvekst med økende nivå av innkryssing. Studien fra Alta-elven var basert på samme molekylærgenetiske metode som studiene til Bolstad mfl.
Ytterligere én norsk studie har så langt sett på endringer i livshistorie grunnet innkrysning (Besnier mfl. 2022). Ved Havforskningsinstituttet sin forskningsplattform i Etne-elven er all oppvandrende voksenfisk genotypet siden 2013. Samtidig har all rømt oppdrettslaks blitt sortert ut basert på utseende og vekstmønstre i skjell. Bestanden hadde fram til 2013 et gjennomsnittlig nivå av innkryssing av rømt oppdrettslaks på 24 %. Studien av Besnier mfl. baser seg på historiske prøver fra villaks fra starten av 80-tallet (før innkryssing hadde skjedd), prøver fra oppdrettslaks (1989-2012), og prøver av villaks etter at innkrysning hadde skjedd (2013-2016). Prøvene fra villaks før innkryssing, og prøver fra rømt laks tatt i elven i perioden 1989-2012, ble brukt for å utvikle genetiske markører som beregner grad av innkryssing i villaks fra perioden etter 2013. Resultatene viser at økende nivå av innkryssing fører til økende kroppsvekt, både hos smolt og voksenfisk. For hanner ble det i tillegg observert en nedgang i alder ved smoltifisering og ved kjønnsmodning. Studien viser også at individer med økende grad av slektskap til oppdrettslaks returnerte til elven senere i sesongen enn ville individer uten slektskap til oppdrettslaks. Forskjeller i oppvandringstidspunkt kan muligens gi innkrysset laks en fordel, da det reduserer perioden de er utsatt for å bli fanget under sportsfiskesesongen.
Siden tilnærmet all rømt laks har vært fjernet fra Etne-elven siden 2013, gir Etne-elven en unik mulighet til å undersøke hva som skjer i en innkrysset bestand, dersom videre innkryssing av rømt oppdrettslaks opphører (for eksempel på grunn av bruk av steril fisk). På sikt vil resultater fra systemet i Etne kunne belyse i hvilken grad naturlig seleksjon kan rekonstruere laksebestandens egenskaper.
Utenfor Norge er det så langt én studie som har undersøkt forskjeller mellom vill og innkrysset laks (Perriman mfl. 2022). I 2015 og 2016 ble ungfisk fra 18 elver i Newfoundland, Canada, samlet inn og i etterkant klassifisert som vill, oppdrett eller hybrid basert på genetiske markører (Wringe mfl. 2018). Kun små forskjeller ble observert og en morfometrisk analyse viste blant annet at naturlig produserte første generasjons hybrider hadde noe ulik form sammenlignet med den unge villaksen, men ikke den naturlig produserte oppdrettslaksen.
I nyere tid er det også gjennomført flere eksperimentelle forsøk for å undersøke potensielle forskjeller mellom avkom av villaks og laks med ulik grad av genetisk slektskap til oppdrettslaks, klekket i naturen. Resultatene fra disse vil kunne gi oss mer kunnskap om konsekvens av genetisk innkryssing med tanke på forandringer i fenotypiske og livshistorietrekk.
Modellering
Modellering av konsekvenser av innkryssing av rømt oppdrettslaks gir en mulighet til å estimere tidsforløp og omfang av genetisk forandringer i ville laksebestander. Her gjennomgås to publiserte modelleringsarbeid. Det første arbeidet av Hindar mfl. (2006) var basert på best tilgjengelig kunnskap om fitness-forskjeller mellom rømt oppdrettslaks, villaks og deres første- og andregenerasjons avkom midt på 2000-tallet, og estimerte andelen av en villaksbestand som er genetisk innkrysset med rømt oppdrettslaks. Dette ble studert ved forskjellige nivå av rømming. Arbeidet viste at ved 20 % innslag av rømt oppdrettslaks på gyteplassene, fikk en store forskjeller i sammensetningen av bestanden i løpet av 10 laksegenerasjoner (ca. 40 år). Forfatterne viste også at jo mer rømt oppdrettslaks i gytebestanden, desto større andel av den ville bestanden ble genetisk påvirket av rømt oppdrettslaks over tid. Modellen ble senere videreutviklet med en populasjonsdynamisk modell for bestandsstørrelse og en økonomisk modell for verdien av fiske etter laks (Liu mfl. 2013). Modellen hadde ingen fitness-funksjon på den ville bestanden og kunne ikke predikere økologiske konsekvenser av innkryssing annet enn i bestandsstørrelse.
En ny modell (IBSEM – Individual based eco-genetic model) for å studere genetisk innkryssing av rømt oppdrettslaks i villaksbestander er publisert av Castellani mfl. (2015). Modellen ble basert på best tilgjengelig kunnskap om fitnessforskjeller mellom avkom av oppdrettslaks og villaks midt på 2010-tallet. IBSEM er en individbasert modell, og inneholder en realistisk genetisk komponent. Den kan gi et estimat på eventuelle forandringer i ville bestander over tid for parametere som antall yngel, parr, smolt og voksen laks i bestanden, individuell vekst i de forskjellige livsstadier og kjønnsmodning. Ved bruk av modellen har vi dermed en mulighet til å estimere forandringer i ville bestander over tid som følge av forekomst av rømt oppdrettslaks på gyteplassene.
Analyser utført med IBSEM (Castellani mfl. 2018) viste først og fremst at jo flere rømt oppdrettslaks i gytebestanden, desto større genetisk innkryssing og genetiske forandringer i den ville bestanden (slik modellen til Hindar mfl. 2006 også viste). Modellen viste imidlertid at ved lav til moderat innslag av rømt oppdrettslaks er forandringen i den ville bestanden relativt liten. For eksempel, ved 5–10 % innslag av rømt oppdrettslaks på gyteplassene, viste de fleste fenotypiske og livshistorietrekk kun svake forandringer i en villaksbestand etter 50 år med genetisk innkryssing. Kun når innslag av rømt oppdrettslaks på gyteplassene ble økt til 30–50 % ble genetiske forandringer i bestandens gjennomsnittlige fenotype og livshistorietrekk tydelige etter 50 år, selv om det på individnivå kan forekomme større endringer. Forfatterne konkluderte med at dette skyldes at (1) rømt oppdrettslaks har en lavere gytesuksess enn villaks i naturen (Fleming mfl. 1996, 2000), (2) fordi det er en sterk seleksjon mot avkom av oppdrettslaks i naturen (dvs., de har høyere dødelighet enn villaks og påvirker dermed bestandens «karakteristikk» mindre enn dersom de hadde overlevd i større grad), og (3) fordi mange livshistorietrekk i ville bestander er svært plastiske, og tetthetsavhengige. For eksempel, det er godt dokumentert at til tross for at oppdrettslaks vokser langt hurtigere enn vill laks under oppdrettsforhold (Glover mfl. 2017), er det kun funnet svake eller moderate forskjeller i vekst mellom disse gruppene i naturen (McGinnity mfl. 1999; Skaala mfl. 2014; Reed mfl. 2015; Jonsson & Jonsson 2017; Glover mfl. 2018). Modellen indikerer derfor at moderat innslag av rømt oppdrettslaks på gyteplassene kun fører til svake eller moderate forandringer i bestandens gjennomsnittlige fenotype og livshistorietrekk.
IBSEM indikerer likevel at den største målbare effekten av innkryssing av rømt oppdrettslaks er at det kommer færre fisk tilbake fra havet (se også Hutchings 1991 og Liu mfl. 2013). En mulig forklaring er tetthetsavhengig dødelighet, og at en del av elvens produksjonskapasitet brukes til å produsere avkom av rømt og genetisk-påvirket laks, med høyere dødelighet i havet. Dermed reduseres antall laks som vandrer tilbake til elven. Det er likevel viktig å påpeke at modellen, under ulike scenarioer, viser at vedvarende innkryssing av rømt oppdrettslaks vil på sikt føre til en svekket bestand med redusert produksjon av laks av vill avstamning (Castellani mfl. 2018; Sylvester mfl. 2019; Bradbury mfl. 2020a). Dette er forenlig med all tilgjengelig kunnskap om dette temaet (Glover mfl. 2017). Det er likevel viktig å påpeke at modeller ikke kan ta hensyn til ukjente faktorer, og kan derfor har sine begrensninger.
Faktorer som forklarer variasjon i andel rømt oppdrettslaks i elv og grad av innkryssing
Det er mange biologiske faktorer (bestandens størrelse, tetthet, livshistoriekarakteristikk og timing av kritiske hendelser som gyting) og fysiske faktorer (elvegradient og lengde, temperaturforhold, substrat, vandringshinder) som, i tillegg til forekomst av rømt oppdrettslaks, vil påvirke graden av genetisk innkryssing og konsekvensene for den ville bestanden.
Innslag av rømt oppdrettslaks i norske elver registreres årlig gjennom overvåkningsprogrammet i gjennomsnittlig 200 vassdrag, og nivå på innkryssing av rømt oppdrettslaks er estimert for nærmere 250 bestander. En omfattende studie har ved bruk av disse to storskala datasettene undersøkt sammenhengen mellom en rekke biologiske, fysiske og menneskeskapte påvirkninger og andel rømt oppdrettslaks og nivå av genetisk innkryssing i norske vassdrag i perioden 2006-2018. Studien av Diserud mfl. (2022) viser at andel rømt oppdrettslaks øker med økende oppdrettsintensitet i regionen, og at økende vannføring tiltrekker seg rømlinger. Samtidig viser studien at en økning i bestandsstørrelse reduserer andelen rømt oppdrettslaks i bestanden, til tross for at det øker det faktiske antallet. Videre viser studien at selv om oppdrettslaks på et overordnet nivå trekkes mot de største bestandene, vil det i områder med bare små bestander være de relativt største bestandene som tiltrekker seg den rømte oppdrettslaksen.
Den nye studien bygger på og samsvarer med resultatene fra undersøkelsene gjort i forbindelse med 5 og 10-årsevalueringen av ordningen med nasjonale laksevassdrag, og -fjorder, hvor en bl.a. så på hvilke typer elver som tiltrakk seg rømt fisk (Fiske mfl. 2013; Hindar mfl. 2018). Disse undersøkelsene viste også at økt vannføring, større laksebestand og økt oppdrettsintensitet i regionen, økte antallet rømt oppdrettslaks i elven. Andelen rømt oppdrettslaks økte også med økt vannføring og oppdrettsintensitet i regionen, men avtok med økende bestandsstørrelse av villaks. At antall/andel rømt oppdrettslaks i et vassdrag har en sammenheng med nærhet til og omfanget av oppdrett sammenfaller også med tidligere analyser (Fiske mfl. 2006) og er også dokumentert i Canada (Keyser mfl. 2018) og Skottland (Gilbey mfl. 2021). Høy vannføring (og stor laksebestand) ansees å være viktig fordi det tiltrekker rømt oppdrettslaks (Mahlum mfl. 2020). I studien til Mahlum mfl. 2020 ble det også dokumentert en positiv sammenheng mellom mengde rømt oppdrettslaks observert i 54 elver på Vestlandet og oppdrettsintensitet i området. Tilsvarende, i en rapport fra Skottland var innkryssing av rømt oppdrettslaks konsentrert rundt områder med oppdrettsanlegg i sjø og ferskvannanlegg tilknyttet smoltproduksjon, mens det utenfor disse områdene var få tegn til innkryssing av rømt oppdrettslaks (Gilbey mfl. 2021).
Hvor stor gytesuksess voksen rømt oppdrettslaks har på gyteplassene er trolig avhengig av hvor mange ville konkurrenter de har. Rømt oppdrettslaks har generelt sett en lavere gytesuksess enn villaks (Fleming mfl. 1996, 2000), og et gitt innslag av rømt oppdrettslaks på gyteplassene vil derfor ikke automatisk føre til en tilsvarende prosentvis genetisk innkryssing. Det er derimot grunn til å tro at oppdrettslaksens gytesuksess vil variere i tid og rom, avhengig av blant annet hvor lenge den har vært i havet (Fleming mfl. 1996, 1997) og konkurransen den møter på gyteplassen med vill fisk (Glover mfl. 2012). Derfor vil også genetisk innkryssing kunne variere i tid og mellom bestander i ulike vassdrag. Likevel er det dokumentert en viss sammenheng mellom observert andel rømt oppdrettslaks og beregnet genetisk innkryssing på vassdragsnivå. I et arbeid med 20 vassdrag, fant Glover mfl. (2013) en sammenheng mellom beregnet forekomst av rømt oppdrettslaks over tid og beregnet innkryssing. De fant at 47 % av variansen i genetisk innkryssing ble forklart av andel rømt oppdrettslaks observert (R2 = 0,47). Dette ble bekreftet i en mer omfattende analyse av det samme datagrunnlaget av Heino mfl. (2015) som også tok hensyn til bestandsstørrelse (R2 = 0,51).
I 2016 publiserte Karlsson mfl. (2016) et arbeid som viste at forekomst av rømt oppdrettslaks i perioden 1989–2012 (gjennomsnittlig årsprosent, etter Diserud mfl. 2013) forklarte 24 % av variansen i genetisk innkryssing i 77 undersøkte vassdrag. Da forfatterne utførte analysen på et regionalt nivå, økte forklaringsstyrken til 56 %. Oppsummert viser disse analysene at selv om det er en sammenheng mellom andel rømt oppdrettslaks i bestanden og genetisk innkryssing, så er det også andre faktorer i elven som har betydning. En tydelig sammenheng mellom forekomst av rømt laks og nivå av innkryssing ble også bekreftet i studien til Diserud mfl. (2022), som til dags dato er den mest omfattende analysen gjort på dette området. Som nevnt over ble en rekke biologiske, fysiske og menneskeskapte påvirkninger undersøkt (se Diserud mfl. 2022 for mer detaljer), og på et overordnet nivå er grad av genetisk innkryssing i hovedsak styrt av andel rømt laks observert i vassdraget. På elvenivå vil likevel fysiske hindringer i elvemunningen kunne føre til at nivå av innkryssing er lavere enn forventet, fordi den rømte laksen ikke klarer ta seg fram til egnede gyteområder. De nordlige bestandene som er mer i slekt med laksen på Kolahalvøya i Russland enn med resten av den norske laksen, og dermed mindre i slekt med oppdrettslaksen, er mer motstandsdyktig mot innkryssing enn laks fra de sørlige bestandene (men viser også større endringer i livshistorie etter innkryssing; Bolstad mfl. 2017). Bestandene i overgangssonen mellom disse to fylogenetiske gruppene er ekstra sårbar for innkryssing.
7. Takk
Takk til Lasse Berg Andersen, Ellen Sofie Grefsrud, Terje Svåsand og Nina Sandlund for deres bidrag med en generell beskrivelse av metodikken brukt i denne risikovurderingen (Kapittel 2). Takk til Mikko Heino, Geir Bolstad og Terje Svåsand for bidrag til tidligere versjoner av kunnskapsstatusen (Kapittel 6).
8. Referanser
Abrantes KG, Lyle JM, Nichols PD, Semmens JM. 2011. Do exotic salmonids feed on native fauna after escaping from aquaculture cages in Tasmania, Australia? Canadian Journal of Fisheries and Aquatic Sciences 68, 1539–51.
Andersen LB, Grefsrud ES, Svåsand T, Sandlund N. 2022. Risk understanding and risk acknowledgement: a new approach to environmental risk assessment in marine aquaculture. ICES Journal of Marine Science.
Anon. 2015a. Rømt oppdrettslaks i vassdrag i 2014 - rapport fra det nasjonale overvåkningsprogrammet. Fisken og havet, særnr. 2b–2015.
Anon. 2016a. Rømt oppdrettslaks i vassdrag i 2015 - rapport fra det nasjonale overvåkningsprogrammet. Fisken og havet, særnr. 2b–2016.
Anon. 2017a. Rømt oppdrettslaks i vassdrag i 2016 - rapport fra det nasjonale overvåkningsprogrammet. Fisken og havet, særnr. 2b–2017.
Anon. 2018. Rømt oppdrettslaks i vassdrag i 2017 - rapport fra det nasjonale overvåkningsprogrammet. Fisken og havet, særnr. 2–2018.
Anon. 2019a. Rømt oppdrettslaks i vassdrag i 2018 - rapport fra det nasjonale overvåkningsprogrammet. Fisken og havet, nr. 2019–4.
Anon. 2017b. Status for norske laksebestander i 2017. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 10.
Anon. 2018b. Status for norske laksebestander i 2018. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 11.
Anon. 2019b. Status for norske laksebestander i 2019. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 12.
Anon. 2020. Status for norske laksebestander i 2020. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 15.
Anon. 2021. Status for norske laksebestander i 2021. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 16.
Anon. 2022. Status for norske laksebestander i 2022. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 17.
Aven, T. 2014. Risk, Surprises and Black Swans: Fundamental Ideas and Concepts in Risk Assessment and risk management.Routledge, London. 276 s.
Araki H, Schmid C. 2010. Is hatchery stocking a help or harm? Evidence, limitations and future directions in ecological and genetic surveys. Aquaculture, 38 (Supp. 1): 2–11.
Aronsen mfl. 2020. Rømt oppdrettslaks i vassdrag i 2019 - rapport fra det nasjonale overvåkningsprogrammet. Fisken og havet, nr. 2020–3.
Artsdatabanken. 2021. Norsk rødliste for arter 2021. Laks Salmo salar Linnaues, 1758. https://artsdatabanken.no/lister/rodlisteforarter/2021/8149
Besnier F, Glover KA, Skaala Ø. 2011. Investigating genetic changes in wild populations: modelling gene-flow from farm escapees. Aquaculture Environment Interactions 2: 75–86.
Besnier F, Glover KA, Lien S, Kent M, Hansen MM, Shen X, Skaala Ø. 2015. Identification of quantitative genetic components of fitness variation in farmed, hybrid and native salmon in the wild. Heredity 00:1-9.
Besnier F, Solberg MF, Harvey AC, Carvalho GR, Bekkevold D, Taylor MI, Creer S, Nielsen EE, Skaala Ø, Ayllon F, Dahle G, Glover KA. 2020. Epistatic regulation of growth in Atlantic salmon revealed: a QTL study performed on the domesticated-wild interface. BMC Genetics 21.
Besnier F, Ayllon F, Skaala Ø, Solberg MF, Fjeldheim PT, Anderson A, Knutar S, Glover KA. 2022. Introgression of domesticated salmon changes life history and phenology of a wild salmon population. Evolutionary Applications.
Bertolotti AC, Layer RM, Gundappa MK, Gallagher MD, Pehlivanoglu E, Nome T, Robledo D, Kent MP, Røsæg LL, Holen MM, Mulugeta TD, Ashton TJ, Hindar K, Sægrov H, Florø-Larsen B, Erkinaro J, Primmer CR, Bernatchez L, Martin SAM, Johnston IA, Sandve SR, Lien S, Macqueen DJ. 2020. The structural variation landscape in 492 Atlantic salmon genomes. Nature Communications 11:5176.
Bicskei B, Bron J, Glover KA, Taggart J B. 2014. A comparison of gene transcription profiles of domesticated and wild Atlantic salmon (Salmo salar L.) at different life stages, reared under controlled conditions. BMC Genomics 15:884.
Bicskei B, Taggart JB, Glover KA, Bron JE. 2016. Comparing the transcriptomes of embryos from domesticated and wild Atlantic salmon (Salmo salar L.) stocks and examining factors that influence heritability of gene expression. Genetics Selection Evolution, 48(1):1-16.
Bolstad GH, Hindar K, Robertsen G, Jonsson B, Sægrov H, Diserud OH, Fiske P, Jensen AJ, Urdal K, Næsje TF. 2017. Gene flow from domesticated escapees alters the life history of wild Atlantic salmon. Nature Ecology, Evolution, 1: 0124.
Bolstad GH, Karlsson S, Hagen I, Fiske P, Urdal K, Sægrov H, Florø-Larsen B, Sollien VP, Østborg G, Diserud OH, Jensen A, Hindar K. 2021. "Introgression from farmed escapees affects the full life cycle of wild Atlantic salmon." Science Advances 7(52).
Bourke EA, Coughlan J, Jansson H, Galvin P, Cross TF. 1997. Allozyme variation in populations of Atlantic salmon located throughout Europe: diversity that could be compromised by introductions of reared fish. ICES Journal of Marine Science 54: 974–985.
Bourret V, Kent MP, Primmer CR, Vasemägi A, Karlsson S, Hindar K, McGinnity P, Verspoor E, Bernatchez L, Lien S. 2013. SNP-array reveals genome-wide patterns of geographical and potential adaptive divergence across the natural range of Atlantic salmon (Salmo salar). Molecular Ecology 22: 532–551.
Bradbury IR, Duffy S, Lehnert SJ, Johannsson R, Fridriksson JH, Castellani M. Burgetz I, Sylvester E, Messmer A, Layton K, Kelly N, Dempson JB, Fleming I.A. 2020a. Model-based evaluation of the genetic impacts of farm-escaped Atlantic salmon on wild populations. Aquaculture Environment Interactions 12, 45–59.
Bradbury IR, Burgetz I, Coulson MW, Verspoor E, Gilbey J, Lehnert SJ, Kess T, Cross T, Vasemagi A, Solberg MF, Fleming I, McGinnity P. 2020b. Beyond hybridization: the genetic impacts of non-reproductive ecological interactions of salmon aquaculture on wild populations. Aquaculture Environment Interactions 12: 429-445.
Castellani M, Heino M, Gilbey J, Araki H, Svåsand T, Glover KA. 2015. IBSEM: An individual-based Atlantic salmon population model. PLOS One 10(9): e0138444.
Castellani M, Heino M, Gilbey J, Araki H, Svåsand T, Glover K. 2018. Modeling fitness changes in wild Atlantic salmon populations faced by spawning intrusion of domesticated escapees. Evolutionary Applications, 1–16.
Chittenden CM, Rikardsen AH, Skilbrei OT, Davidsen JG, Halttunen E, Skardhamar J, McKinley RS. 2011. An effective method for the recapture of escaped farmed salmon. Aquaculture Environ Interact 2011, 1(3):215-224.
Clifford SL, McGinnity P, Ferguson A. 1998a. Genetic changes in Atlantic salmon (Salmo salar) populations of northwest Irish rivers resulting from escapes of adult farm salmon. Canadian Journal of Fisheries and Aquatic Sciences 55: 358–363.
Clifford SL, McGinnity P, Ferguson A. 1998b. Genetic changes in an Atlantic salmon population resulting from escaped juvenile farm salmon. Journal of Fish Biology 52: 118–127.
Cross TF, Challanain DN. 1991. Genetic characterisation of Atlantic salmon (Salmo salar) lines farmed in Ireland. Aquaculture 98: 209–216.
Crowley SE, Bradbury I, Messmer A, Duffy SJ, Islam SS, Fleming IA. 2020. Common-garden comparison of relative survival and fitness-related traits of wild, farm, and hybrid Atlantic salmon Salmo salar parr in nature. Aquaculture Environment Interactions 14: 35-52.
Crozier WW. 1993. Evidence of genetic interaction between escaped farmed salmon and wild Atlantic salmon (Salmo salar L) in a Northern Irish river. Aquaculture 113(1-2):19-29.
Crozier WW. 2000. Escaped farmed salmon, Salmo salar L., in the Glenarm River, Northern Ireland: genetic status of the wild population 7 years on. Fisheries Management and Ecology, 7: 437–446.
Cucherousset J, Sundt-Hansen LE, Buoro M, Závorka L, Lassus R, Bækkelie KAE, Fleming IA, Björnsson BT, Johnsson JI, Hindar K. 2021. Growth-enhanced salmon modify stream ecosystem functioning. Journal of Fish Biology, 99: 1978-1989.
Darwish TL, Hutchings JA. 2009. Genetic variability in reaction norms between farmed and wild backcrosses of Atlantic salmon (Salmo salar). Canadian Journal of Fisheries and Aquatic Sciences 66: 83–90.
Debes PV, Hutchings JA. 2014. Effects of domestication on parr maturity, growth, and vulnerability to predation in Atlantic salmon. Canadian Journal of Fisheries and Aquatic Sciences 71, 1371–84.
Debes PV, Solberg MF, Matre IHm Dyrhovden L, Glover KA. 2021. Genetic variation for upper thermal tolerance diminishes within and between populations with increasing acclimation temperature in Atlantic salmon. Heredity 127(5): 455-466.
Diserud OH, Fiske P, Hindar K. 2013. Forslag til kategorisering av laksebestander som er påvirket av rømt oppdrettslaks – Oppdatering for perioden 1989–2012. – NINA Rapport 976. 22 s.
Diserud OH, Hindar K, Karlsson S, Glover K, Skaala Ø. 2016. Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander. Vedlegg Notat NINA/HI, s. 53–85 i Anon. 2016. Klassifisering av 104 laksebestander etter kvalitetsnorm for villaks. Temarapport nr. 4. Vitenskapelig råd for lakseforvaltning.
Diserud, OH, Hindar, K, Karlsson, S, Glover, KA, Skaala, Ø. 2017. Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander – status 2017. NINA Rapport 1337. 55 s.
Diserud OH, Fiske P, Sægrov H, Urdal K, Aronsen T, Lo H, Barlaup BT, Niemela E, Orell P, Erkinaro J, Lund RA, Økland F, Østborg GM, Hansen LP, Hindar K. 2019a Frequency of escapees in Norwegian rivers 1989–2013. Ices Journal of Marine Science 76, 1140–50.
Diserud OH, Hindar K, Karlsson S, Glover KA, Skaala Ø. 2019b. Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander – oppdatert status 2019. NINA Rapport 1659. 72 s.
Diserud OH, Hindar K, Karlsson S, Glover KA, Skaala Ø. 2020. Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander – oppdatert status 2020. NINA Rapport 1926. Norsk institutt for naturforskning. ISBN: 978-82-426-4702-3
Diserud OH, Fiske P, Karlsson S, Glover KA, Næsje T, Aronsen T, Bakke G, Barlaup BT, Erikanõ J, Florø-Larsen B, Foldvik A, Heino M, Kanstad- Hanssen Ø, Lo H, Lund RA, Muladal R, Niemelã E, Økland F, Østborg GM, Otterå H, Skaala Ø, Skoglund H, Solberg MF, Sollien VP, Sægrov H, Urdal K,Wennevik V, Hindar K. 2022. Natural and anthropogenic drivers of escaped farmed salmon occurrence and introgression into wild Norwegian Atlantic salmon populations. ICES Journal of Marine Science, akseptert for publisering.
Einum S, Fleming IA. 1997. Genetic divergence and interactions in the wild among native, farmed and hybrid Atlantic salmon. Journal of Fish Biology 50: 634–651.
Ferguson A, McGinnity P, Baker N, Cotter D, Hynes R, O’Hara B, O’Maoileidigh N, Prodöhl P, Rogan G. 2002. A two-generation experiment comparing the fitness and life-history traits of native, ranched, non-native, farmed, and hybrid Atlantic salmon under natural conditions. ICES CM 2002/T:04.
Fiske P, Lund, R A, Hansen, LP. 2006. Relationships between the frequency of farmed Atlantic salmon, Salmo salar L., in wild salmon populations and fish farming activity in Norway 1989–2004. ICES J Mar Sci 63: 1182–1189.
Fiske P, Diserud,OH, Robertsen G, Foldvik A, Skilbrei OT, Heino M, Helland IP, Hindar K. 2013. Midtveisvurdering av nasjonale laksevassdrag og nasjonale laksefjorder. Rømt oppdrettslaks og bestandsstatus. NINA Minirapport, 470: 1–24.
Fiske P. 2013. Overvåking av rømt oppdrettslaks i elv om høsten 2010–2012. NINA Rapport 989.
Fiske P, Aronsen T, Hindar K. 2014. Overvåking av rømt oppdrettslaks i elver om høsten 2013. NINA rapport 1063. 44 s.
Fjelldal PG, Glover KA, Skaala Ø, Imsland A, Hansen TJ. 2009. Vertebral body mineralization and deformities in cultured Atlantic salmon (Salmo salar L.): Effects of genetics and off-season smolt production. Aquaculture 296: 36–44.
Fleming IA, Jonsson B, Gross MR, Lamberg A. 1996. An experimental study of the reproductive behaviour and success of farmed and wild Atlantic salmon (Salmo salar). J Appl Ecol 33:893–905.
Fleming IA, Lamberg A, Jonsson B. 1997. Effects of early experience on the reproductive performance of Atlantic salmon. Behav Ecol 8:470–480.
Fleming IA, Einum S. 1997. Experimental tests of genetic divergence of farmed from wild Atlantic salmon due to domestication. ICES Journal of Marine Science 54: 1051–1063.
Fleming I, Hindar K, Mjølnerød IB, Jonsson B, Balstad T, Lamberg A. 2000. Lifetime success and interactions of farm salmon invading a native population. Proceedings of the Royal Society of London B. 267: 1517–1523.
Fleming IA, Agustsson T, Finstad B, Johnsson JI, Björnsson BT. 2002. Effects of domestication on growth physiology and endocrinology of Atlantic salmon (Salmo salar). Canadian Journal of Fisheries and Aquatic Sciences 59:1323-1330.
Forskrift om fellesansvar for utfisking mv. av rømt oppdrettsfisk. 2015. Lovdata: FOR-2015-02-05-89. Hentet fra: https://lovdata.no/dokument/SF/forskrift/2015-02-05-89.
Fraser DJ, Weir LK, Bernatchez L, Hansen MM, Taylor EB. 2011. Extent and scale of local adaptation in salmonid fishes: review and meta-analysis. Heredity 106: 404–420.
Fraser DJ, Houde ALS, Debes PV, O’Reilly P, Eddington JD, Hutchings JA. 2010. Consequences of farmed-wild hybridization across divergent wild populations and multiple traits in salmon. Ecological Applications 20: 935–953.
Gilbey J, Sampayo J, Cauwelier E, Malcolm I, Millidine K, Jackson F, Morris DJ. 2021. A national assessment of the influence of farmed salmon escapes on the genetic integrity of wild Scottish Atlantic salmon populations Scottish Marine and Freshwater Science Vol 12 No 12
Gjedrem T. 2010. The first family-based breeding program in aquaculture. Reviews in Aquaculture 2, 2–15.
Gjøen HM, Bentsen HB. 1997. Past, present, and future of genetic improvement in salmon aquaculture. ICES Journal of Marine Science 54: 1009–1014.
Glover KA, Skaala, Ø. 2006. Temporal stability of sea louse Lepeophtheirus salmonis Krøyer populations on Atlantic salmon Salmo salar L. of wild, farm and hybrid parentage. Journal of Fish Biology 68: 1795–1807.
Glover KA, Skar C, Christie KE, Glette J, Rudra H, Skaala Ø. 2006a. Size-dependent susceptibility to infectious salmon anemia virus (ISAV) in Atlantic salmon (Salmo salar L.) of farm, hybrid and wild parentage. Aquaculture 254: 82–91.
Glover KA, Bergh Ø, Rudra H, Skaala, Ø. 2006b. Juvenile growth and susceptibility to Aeromonas salmonicida subsp. salmonicida in Atlantic salmon (Salmo salar L.) of farmed, hybrid, and wild parentage. Aquaculture 254: 72–81.
Glover KA, Otterå H, Olsen RE, Slinde E, Taranger GL, Skaala Ø. 2009. A comparison of farmed, wild and hybrid Atlantic salmon (Salmo salar L.) reared under farming conditions. Aquaculture 286: 203–210.
Glover KA. 2010. Forensic identification of farmed escapees: a review of the Norwegian experience. Aquaculture Environment Interactions 1: 1–10.
Glover KA, Quintela, M, Wennevik V, Besnier F, Sørvik AGE, Skaala Ø. 2012. Three decades of farmed escapees in the wild: A spatio- temporal analysis of Atlantic salmon population genetic structure throughout Norway. PloS One 7: e43129.
Glover KA, Pertoldi C, Besnier F, Wennevik V, Kent M, Skaala Ø. 2013. Atlantic salmon populations invaded by farmed escapees: quantifying genetic introgression with a Bayesian approach and SNPs. BMC Genetics, 14.
Glover KA, Solberg MF, McGinnity P, Hindar K, Verspoor E, Coulson MW, Hansen MM, Araki H, Skaala Ø, Svåsand T. 2017. Half a century of genetic interaction between farmed and wild Atlantic salmon: Status of knowledge and unanswered questions. Fish and Fisheries 1–38.
Glover KA, Urdal K, Næsje T, Skoglund H, Florø-Larsen B, Otterå H, Fiske P, Heino M, Aronsen T, Sægrov H, Diserud O, Barlaup BT, Hindar K, Bakke G, Solberg I, Lo H, Karlsson S, Skaala Ø, Lamberg A, Kanstad-Hanssen Ø, Muladal R, Skilbrei OT, Wennevik V. 2019. Domesticated escapees on the run: the second-generation monitoring program reports the numbers and proportions of farmed Atlantic salmon in >200 rivers annually Ices Journal of Marine Science 76, 1151–61.
Glover KA, Wennevik V, Hindar K. Skaala Ø, Fiske P, Solberg MF, Diserud OH, Svåsand T, Karlsson S, Andersen L.B, Grefsrud ES. 2020. The future looks like the past: Introgression of domesticated Atlantic salmon escapees in a risk assessment framework. Fish and Fisheries, akseptert.
Grefsrud ES, Svåsand T, Glover KA, Husa V, Kupka-Hansen P, Samuelsen OB, Sandlund N, Stien LH. 2019. Risikorapport norsk fiskeoppdrett 2019. Fisken og Havet 2019–5.
Grefsrud ES, Svåsand T, Glover KA, Husa V, Kupka-Hansen P, Samuelsen OB, Sandlund N, Stien LH. 2021. Risikorapport norsk fiskeoppdrett 2021. Fisken og Havet 2019–5.
Hansen LP, Jacobsen JA, Lund RA. 1993. High numbers of farmed Atlantic salmon, Salmo salar L., observed on oceanic waters north of the Faroe Islands. Aquaculture and Fisheries Management 24: 777–781.
Hansen LP. 2006a. Migration and survival of farmed Atlantic salmon (Salmo salar L.) released from two Norwegian fish farms. ICES J Mar Sci 63, 1211–1217.
Hansen LP. 2006b. Vandring og spredning av rømt oppdrettslaks. NINA Rapport 162: 1–21.
Harvey A, Glover KA, Taylor MI, Creer S, Carvalho GR. 2016a. A common garden design reveals population-specific variability in potential impacts of hybridization between populations of farmed and wild Atlantic salmon, Salmo salar L. Evolutionary Applications 9, 435–49.
Harvey A, Juleff G, Carvalho G, Taylor M, Solberg MF, Dyrhovden L, Matre IH, Glover KA. 2016b. Does density influence relative growth performance of farm, wild and F1 hybrid Atlantic salmon in semi-natural and hatchery common garden conditions? Royal Society Open Science 3:16152.
Harvey A, Solberg MF, Troianou E, Carvalho GR, Taylor M, Creer S, Dyrhovden L, Matre IH, Glover KA. 2016c. Plasticity in growth of farmed and wild Atlantic salmon: is the increased growth rate of farmed salmon caused by evolutionary adaptations to the commercial diet? BMC Evolutionary Biology 16:264.
Harvey A, Solberg MF, Glover KA, Taylor MI, Creer S, Carvalho GR. 2016d. Plasticity in response to feed availability: Does feeding regime influence the relative growth performance of domesticated, wild and hybrid Atlantic salmon Salmo salar parr? Journal of Fish biology 89:3
Harvey AC, Glover KA, Wennevik V, Skaala Ø. 2020. Atlantic salmon and sea trout display synchronized smolt migration relative to linked environmental cues. Scientific reports 10: 3529.
Heino M, Svåsand T, Wennevik W, Glover KA. 2015. Genetic introgression of farmed salmon in native populations: quantifying the relative influence of population size and frequency of escapees. Aquaculture Environment Interactions 6: 185–190.
Hindar K, Ryman N, Utter F. 1991. Genetic effects of cultured fish on natural fish populations. Canadian Journal of Fisheries and Aquatic Sciences 48: 945–57.
Hindar K, Fleming IA, McGinnity P, Diserud O. 2006. Genetic and ecological effects of salmon farming on wild salmon: modelling from experimental results. ICES J. Marine Sci. 63: 1234–1247.
Hindar K, Diserud OH, Fiske P, Forseth T, Jensen A J, Ugedal O, Jonsson N, Sloreid S.-E, Arnekleiv JV, Saltveit SJ, Sægrov H, Sættem LM. 2007. Gytebestandsmål for laksebestander i Norge. NINA Rapport 226. 78 s.
Hindar K, Diserud OH, Fiske P, Karlsson S, Bolstad GH, Foldvik A, Wennevik V, Bremset G og Rosten C. 2018. Evaluering av nasjonale laksevassdrag og nasjonale laksefjorder: Rømt oppdrettslaks, genetisk innkryssing og bestandsstatus. NINA Rapport 1461. Norsk institutt for naturforskning.
Holborn MK, Crowley SE, Duffy S J, Messmer AM, Kess T, Dempson JB, Wringe BF, Fleming I, Bentzen P og Bradbury I. 2022. Precocial male maturation contributes to the introgression of farmed Atlantic salmon into wild populations. Aquaculture Environment Interactions, 14: 205-218.
Houde ALS, Fraser DJ, Hutchings JA. 2010. Reduced anti-predator responses in multigenerational hybrids of farmed and wild Atlantic salmon (Salmo salar L.). Conservation Genetics 11: 785–794.
Houston RD, Haley CS, Hamilton A. 2008 Major quantitative trait loci affect resistance to infectious pancreatic necrosis in Atlantic salmon (Salmo salar). Genetics 178: 1109–1115.
Hutchings JA. 1991. The threat of extinction to native populations experiencing spawning intrusions by cultured Atlantic salmon. Aquaculture, 98: 119–132.
ICES. 2016. Report of the Workshop to address the NASCO request for advice on possible effects of salmonid aquaculture on wild Atlantic salmon populations in the North Atlantic (WKCULEF), 1–3 March, Copenhagen, Denmark. ICES CM 2016/ACOM 42: 44 pp.
Islam SS, Wringe BF, Bøe K, Bradbury IR, Fleming IA. 2021. Early-life fitness trait variation among divergent European and North American farmed and Newfoundland wild Atlantic salmon populations. Aquaculture Environment Interactions 13: 323-337.
Islam SS, Xue Xm Cabarello-Solares A, Bradbury IR, Rise M, Fleming IA. 2021. Distinct early life stage gene expression effects of hybridization among European and North American farmed and wild Atlantic salmon populations. Molecular Ecology.
Jacobsen JA, Hansen LP. 2001. Feeding habits of wild and escaped farmed Atlantic salmon, Salmo salar L., in the Northeast Atlantic. Ices Journal of Marine Science 58, 916–33.
Jensen AJ, Karlsson S, Fiske P, Hansen LP, Hindar K, Østborg G. 2013. Escaped farmed Atlantic salmon in the Arctic Ocean. Aquaculture Environment Interactions 3: 223–229.
Johnsson JI, Björnsson BT. 1994. Growth hormone increases growth rate, appetite and dominance in juvenile rainbow trout, Oncorhynchus mykiss. Animal Behaviour 48: 177–186.
Johnsson JI, Petersson E, Jönsson E, Björnsson BT, Järvi T. 1996. Domestication and growth hormone alter antipredator behaviour and growth patterns in juvenile brown trout. Salmo trutta. Canadian Journal of Fisheries and Aquatic Sciences 53: 1546–1554
Johnsson JI, Höjesjö J, Fleming IA. 2001. Behavioural and heart rate response to predation risk in wild and domesticated Atlantic salmon. Canadian Journal of Fisheries and Aquatic Sciences 58: 788–794.
Jonsson N, Jonsson B, Hansen LP. 1998. The relative role of density-dependent and densityindependent survival in the life cycle of Atlantic salmon Salmo salar. Journal of Animal Ecology 67: 751–762.
Jonsson B, Jonsson N, Hansen LP. 2003. Atlantic salmon straying from the River Imsa. Journal of Fish Biology 62: 641–657.
Jonsson B, Jonsson N. 2017. Maternal inheritance influences homing and growth of hybrid offspring between wild and farmed Atlantic salmon. Aquaculture Environment Interactions, 9:231-238.
Jönsson E, Johnsson JI, Björnsson BT. 1996. Growth hormone increases predation exposure of rainbow trout. Proceedings of the Royal Society of London, Series B, 263: 647–651.
Jönsson E, Johnsson JI, Björnsson BT. 1998. Growth hormone increases aggressive behavior in juvenile rainbow trout. Hormones and Behaviour 33: 9–15.
Jørgensen KM, Solberg MF, Besnier F, Thorsen A, Fjelldal PG, Skaala O, Malde K, Glover KA. 2018. Judging a salmon by its spots: environmental variation is the primary determinant of spot patterns in Salmo salar. Bmc Ecology 18.
Kanstad-Hanssen Ø, Bjørnbet S, Lamberg A. 2018. Overvåking av elver og uttak av rømt oppdrettslaks – tiltak etter rømming fra Rauma stamfisk AS i 2017. Rapport nr. 2018-01. 11 s.
Kanstad-Hanssen Ø, Gjertsen V, Bentsen V, Jamtfall E. 2021. Uttak av rømt oppdrettslaks i 18 elver – et oppdrag for OURO i 2020. Rapport nr. 2021-03. 23 s.
Kanstad-Hanssen Ø, Gjertsen V, Bentsen V, Lamberg A. 2020. Uttak av rømt oppdrettslaks i 17 elver – et oppdrag for OURO i 2019. Rapport nr. 2020-04. 23 s.
Kanstad-Hanssen Ø, Gjertsen V, Lamberg A. 2018. Overvåking av elver og uttak av rømt oppdrettslaks – tiltak etter smoltrømming fra Akvafarm AS i 2016. Rapport nr. 2018-03. 15 s.
Kanstad-Hanssen Ø, Gjertsen V, Lamberg A. 2019. Uttak av rømt oppdrettslaks i 25 elver – et oppdrag for OURO i 2018. Rapport nr. 2019-02. 29 s.
Kanstad-Hanssen Ø, Gjertsen V. 2019. Overvåking av elver og uttak av rømt oppdrettslaks – tiltak etter smoltrømming fra Akvafarm AS i 2016 og rømming av fisk av samme årsklasse fra Wilsgård Fiskeoppdrett AS i 2017. Rapport nr. 2019-03. 13 s.
Kanstad-Hanssen Ø, Lamberg A. 2018. Uttak av rømt oppdrettslaks i 20 elver – et oppdrag for OURO i 2017. Rapport nr. 2018-04. 31 s.
Kanstad-Hanssen Ø, Strand R, Bentsen V, Jamtfall E. 2022. Uttak av rømt oppdrettslaks i 13 elver – et oppdrag for OURO i 2021. SNA-rapport 04/2022. 28 s.
Karlsson S, Moen T, Hindar K. 2010. Contrasting patterns of gene diversity between microsatellites and mitochondrial SNPs in farm and wild Atlantic salmon. Conservation Genetics 11: 571–582.
Karlsson S, Moen T, Lien S, Glover KA, Hindar K. 2011. Generic genetic differences between farmed and wild Atlantic salmon identified from a 7K SNP-chip. Molecular Ecology Resources 11: 247–253.
Karlsson S, Diserud OH, Moen T, Hindar K. 2014. A standardized method for quantifying unidirectional genetic introgression. Ecology and Evolution 4, 3256–63.
Karlsson S, Diserud O H, Fiske P, Hindar K. 2016. Widespread genetic introgression of escaped farmed Atlantic salmon in wild salmon populations. Ices Journal of Marine Science, 73: 2488–2498.
Keyser F, Wringe BF, Jeffery NW, Dempson JB, Duffy S, Bradbury IR. 2018. Predicting the impacts of escaped farmed Atlantic salmon on wild salmon populations. Canadian Journal of Fisheries and Aquatic Sciences, 1–7.
Koljonen M-L, Tähtinen J, Säisä M, Koskiniemi J. 2002. Maintenance of genetic diversity of Atlantic salmon (Salmo salar) by captive breeding programmes and the geographic distribution of microsatellite variation. Aquaculture 212: 69–9.
Liu YJ, Diserud OH, Hindar K, Skonhoft A. 2013. An ecological-economic model on the effects of interactions between escaped farmed and wild salmon (Salmo salar). Fish and Fisheries 14, 158–73.
Lura H, Sægrov H. 1991. Documentation of successful spawning of escaped farmed female Atlantic salmon, Salmo salar, in Norwegian rivers. Aquaculture 98: 151–159.
Lura H, Økland F. 1994. Content of synthetic astaxanthin in escaped farmed Atlantic salmon, Salmo salar L., ascending Norwegian rivers. Fisheries Management and Ecology, 1: 205–216.
Madhun AS, Karlsbakk E, Isachsen CH, Omdal LM, Sørvik AGE, Skaala Ø, Wennevik V, Glover KA. 2015. Potential disease interaction reinforced: Double-virus infected escaped farmed Atlantic salmon, Salmo salar L., recaptured in a nearby river. Journal of Fish Diseases 38: 209–219.
Mahlum S, Skoglund H, Wiers T, Norman ES, Barlaup BT, Wennevik V, Glover KA, Urdal K, Bakke G, Volsett KW. 2019. Swimming with the fishes: validating drift diving to identify farmed Atlantic salmon escapees in the wild. Aquaculture Environment Interactions 11: 417-427.
Mahlum S, Vollset KW, Barlaup BT, Skoglund H, Velle G. 2020. Salmon on the lam: Drivers of escape farmed fish abundance in rivers. Journal of Applied Ecology.
Martin-Smith KM, Armstrong JD, Johnsson JI, Björnsson BT. 2004. Growth hormone increases growth and dominance of wild juvenile Atlantic salmon with affecting space use. Journal of Fish Biology 65, Suppl. A: 156–172.
McGinnity P, Stone C, Taggart JB, Cooke DD, Cotter D, Hynes, R, McCamley C, CrossT, Ferguson A. 1997. Genetic impact of escaped farmed Atlantic salmon (Salmo salar L.) on native populations: use of DNA profiling to assess freshwater performance of wild, farmed, and hybrid progeny in a natural river environment. ICES Journal of Marine Science 54: 998–1008.
McGinnity P, Prodöhl P, Ferguson A, Hynes R, Ó Maoiléidigh N, Baker N, Cotter D, O’Hea B, Cooke D, Rogan G, Taggart J, Cross T. 2003. Fitness reduction and potential extinction of wild populations of Atlantic salmon, Salmo salar, as a result of interactions with escaped farm salmon. Proceedings of the Royal Society, London, Series B, 270: 2443–2450.
Mjølnerød IB, Refseth UH, Karlsen E, Balstad T, Jakobsen KS, Hindar K. 1997. Genetic differences between two wild and one farmed population of Atlantic salmon (Salmo salar) revealed by three classes of genetic markers. Hereditas 127: 239–248.
Moen T, Baranski M, Sonesson A K, Kjøglum S. 2009. Confirmation and fine-mapping of a major QTL for resistance to infectious pancreatic necrosis in Atlantic salmon (Salmo salar): population-level associations between markers and trait. BMC Genomics 10: 368.
Mork J. 1991. One-generation effects of farmed fish immigration on the genetic differentiation of wild Atlantic salmon in Norway. Aquaculture 98: 267–276.
Mork OI, Bjerkeng B, Rye M. 1999. Aggressive interactions in pure and mixed groups of juvenile farmed and hatchery-reared wild Atlantic salmon Salmo salar L. in relation to tank substrate. Aquaculture Research 30: 571–578.
Muladal R. 2018. Overvåking og uttak av oppdrettslaks i Troms og Finnmark 2017. Rapport 2‑2018. 26 s.
Muladal R. 2019. Overvåking og uttak av oppdrettslaks i Troms og Finnmark 2018. Rapport 1-2019. 26 s.
Muladal R, Wiersbinski G, Fagard P. 2020. Overvåking og uttak av oppdrettslaks i Troms og Finnmark 2019. Rapport 2-2020. 31 s.
Muladal R, Wiersbinski G, Fagard P. 2021. Overvåking og uttak av oppdrettslaks i Troms og Finnmark 2020. Rapport 2-2020. 31 s.
Muladal R, Wiersbinski G, Fagard P. 2022. Overvåking og uttak av oppdrettslaks i Troms og Finnmark 2021. Rapport 2-2022. 31 s.
Normandeau E, Hutchings JA, Fraser DJ, Bernatchez L. 2009. Population-specific gene expression responses to hybridization between farm and wild Atlantic salmon. Evolutionary Applications 2, 489–503.
Olsen RE, Skilbrei OT. 2010. Feeding preference of recaptured Atlantic salmon Salmo salar following simulated escape from fish pens during autumn. Aquaculture Environment Interactions 1, 167–74.
Ozerov M, Vähä J-P, Wennevik V, Svenning M-A, Vasemägi A, Diaz Fernandez R, Unneland L, Haapanen K, Niemelä E, Falkegård M, Prusov S, Lyzhov I, Rysakova K, Kalske T, Christiansen B. 2017. Comprehensive microsatellite baseline for genetic stock identification of Atlantic salmon (Salmo salar L.) in northernmost Europe. ICES Journal of Marine Science, 74(8): 2159–2169.
Palm S. Karlsson S, Diserud OH. 2021. Genetic evidence of farmed straying and introgression in Swedish wild salmon populations. Aquaculture Environment Interactions 13: 505-513.
Perriman B, Betntzen P, Wringe BF, Duffy S, Islam SS, Fleming IA, Solberg MF, Bradbury IR. 2022. Morphological consequences of hybridization between farm and wild Atlantic salmon Salmo salar under both wild and experimental conditions. Aquaculture Environment Interactions. Akseptert for publikasjon, sammendrag publisert.
Perry WB, Solberg MF, Besnier F, Dyrhovden L, Matre HH, Fjelldal PG, Ayllon F, Creer S, Llewellyn M, Taylors MI, Carvalhol G, Glover KA. 2019. Evolutionary drivers of kype size in Atlantic salmon (Salmo salar): domestication, age and genetics. Royal Society Open Science 6, 14.
Perry WB, Solberg MF, Brodie C, Medina AC, Pillay KG, A. E, Harvey AC, Creer S, Llewellyn M, Taylor AC et al: Disentangling the effects of sex, life history and genetic background in Atlantic salmon: growth, heart and liver under common garden conditions. R Soc Open Sci 2020, 7( 200811)
Perry WB, Kaufamnn J, Solberg MF, Brodie C, Medina AMC, Pillay K, Egerton A, Harvey AC, Philips KP, Coughlan J, Egan F, Geralis R, Hutton S, Leseur F, Ryan S, Poole R, Rogan G, Ryder E, Schaal TP, Waters C, Wynne R, Taylor T, Prodohl P, Creer S, Llewellyn, McGinnity P, Carvalho G, Glover KA. 2021. Domestication-induced reduction in eye size revealed in multiple common garden experiments: The case of Atlantic salmon (Salmo salar L.). Evolutionary Applications 14(9): 2319-2332.
Pulg U, Karlsson S, Diserud OH, Postler C, Stranzl S, Espedal EO, Lennox JR. 2021. Laks i sjøørretbekker – villaks eller oppdrettslaks? NORCE LFI rapport 376. Norwegian Research Center, Bergen
Reed TE, Prodohl P, Hynes R, Cross T, Ferguson A, McGinnity P. 2015. Quantifying heritable variation in fitness-related traits of wild, farmed and hybrid Atlantic salmon families in a wild river environment. Heredity 115, 173–84.
Rengmark AH, Slettan A, Skaala O, Lie O, Lingaas F. 2006. Genetic variability in wild and farmed Atlantic salmon (Salmo salar) strains estimated by SNP and microsatellites. Aquaculture, 253: 229–237.
Roberge C, Einum S, Guderley H, Bernatchez L. 2006. Rapid parallell evolutionary changes of gene transcription profiles in farmed Atlantic salmon. Molecular Ecology 15: 9–20.
Roberge C, Normandeau E, Einum S, Guderley H, Bernatchez, L. 2008. Genetic consequences of interbreeding between farmed and wild Atlantic salmon: insights from the transcriptome. Molecular Ecology 17: 314–324.
Robertsen G, Reid D, Einum S, Aronsen T, Fleming I, Sundt-Hansen L, Karlsson S, Kvingedal E, Ugedal O, Hindar K. 2019. Can variation in standard metabolic rate explain contextdependent performance of farmed salmon offspring? Ecology and Evolution 9, 212–22.
Skaala Ø, Høyheim B, Glover KA, Dahle G. 2004. Microsatellite analysis in domesticated and wild Atlantic salmon (Salmo salar L.): allelic diversity and identification of individuals. Aquaculture 240: 131–143.
Skaala Ø, Taggart JB, Gunnes K. 2005. Genetic differences between five major domesticated strains of Atlantic salmon and wild salmon Journal of Fish Biology 67: 118–128.
Skaala Ø, Wennevik V, Glover KA. 2006. Evidence of temporal genetic change in wild Atlantic salmon (Salmo salar L) populations affected by farmed escapees. ICES J. Marine Science 63: 1224–1233.
Skaala Ø, Glover KA, Barlaup BT, Svåsand T, Besnier F, Hansen MM, Borgstrøm, R. 2012. Performance of farmed, hybrid and wild Atlantic salmon families in a natural river environment. Canadian Journal of Fisheries and Aquatic Sciences 69: 1994–2006.
Skaala Ø, Glover KA, Barlaup B, Borgstrøm R. 2014. Microsatellite DNA used for parentage identification of partly digested Atlantic salmon (Salmo salar) juveniles in a natural rivner environment. Marine Biology Research 10: 323–328.
Skaala Ø, Besnier F., Borgstrom R., Barlaup B., Sorvik A.G., Normann E., Østebø B.I., Hansen M.M., Glover K.A. 2019. An extensive common-garden study with domesticated and wild Atlantic salmon in the wild reveals impact on smolt production and shifts in fitness traits. Evolutionary Applications 12, 1001–16.
Skilbrei OT. 2013. Migratory behaviour and ocean survival of escaped out-of-season smolts of farmed Atlantic salmon Salmo salar. Aquaculture Environment Interactions 3:213-221.
Skilbrei OT, Skulstad OF, Hansen T. 2014. The production regime influences the migratory behavior of escaped farmed Atlantic salmon (Salmo salar L.). Aquaculture 424–425, 146–150.
Skilbrei OT, Heino M, Svåsand T. 2015a. Using simulated escape events to assess the annual numbers and destinies of escaped farmed Atlantic salmon of different life stages, from farms sites in Norway. ICES Journal of Marine Science, 72: 670–685.
Skilbrei OT, Normann E, Meier S, Olsen RE. 2015b. Use of fatty acid profiles to monitor the escape history of farmed Atlantic salmon. Aquaculture Environment Interactions 7:1-13.
Skoglund H, Kambestad M, Wiers T, Normann ES, Hellen BA, Lehmann GB, Landro Y, Urdal K. 2018. Utfisking av rømt oppdrettsfisk på oppdrag for OURO i utvalgte vassdrag i Sør-Norge høsten 2017. LFI-rapport nr: 303. 23 s.
Skoglund H, Kambestad M, Wiers T, Normann ES, Hellen BA, Urdal K. 2019. Utfisking av rømt oppdrettsfisk på oppdrag for OURO i utvalgte vassdrag i Sør-Norge høsten 2018. LFI-rapport nr: 335. 23 s.
Skoglund H, Kambestad M, Wiers T, Normann ES, Hellen BA, Urdal K. 2020. Utfisking av rømt oppdrettsfisk på oppdrag for OURO i utvalgte vassdrag i Sør-Norge høsten 2019. LFI-rapport nr: 370. 21 s.
Skoglund H, Wiers T, Normann ES, Furset TT, Hellen BA, Urdal K. 2021. Utfisking av rømt oppdrettsfisk på oppdrag for OURO i utvalgte vassdrag i Sør-Norge høsten 2020. LFI-rapport nr: 404. 21 s.
Skoglund H, Wiers T, Normann ES, Furset TT, Urdal K. 2022. Utfisking av rømt oppdrettsfisk på oppdrag for OURO i utvalgte vassdrag i Sør-Norge høsten 2021. LFI-rapport nr: 436. 20 s.
Solberg MF, Kvamme O, Nilsen F, Glover KA. 2012. Effects of environmental stress on mRNA expression levels of seven genes related to oxidative stress and growth in Atlantic salmon Salmo salar L. of farmed, hybrid and wild origin. BMC Research Notes 5:672.
Solberg MF, Skaala Ø, Nilsen F, Glover KA. 2013a. Does domestication cause changes in growth reaction norms? A study of farmed, wild and hybrid Atlantic salmon families exposed to environmental stress. PLoS ONE 8(1): e54469.
Solberg MF, Zhang Z, Nilsen F, Glover KA. 2013b. Growth reaction norms of domesticated, wild and hybrid Atlantic salmon families in response to differing social and physical environments. BMC Evolutionary Biology 13:234.
Solberg MF, Zhang Z, Glover KA. 2015. Are farmed salmon more prone to risk than wild salmon? Susceptibility of juvenile farm, hybrid and wild Atlantic salmon Salmo salar L. to an artificial predator. Applied Animal Behaviour Science 162: 67–80.
Solberg MF, Dyrhovden L, Matre IH, Glover KA. 2016. Thermal plasticity in farmed, wild and hybrid Atlantic salmon: no indication of domestication-driven divergence in lower thermal tolerance during early development. BMC Evolutionary Biology 16:38.
Solberg MF, Robertsen G, Sundt-Hansen LE, Hindar K, Glover KA. 2020. Domestication leads to increased predation susceptibility. Scientific Reports 10: 1929.
Sylvester EVA, Wringe BF, Duffy SJ, Hamilton LC, Fleming IA, Bradbury IR. 2018. Migration effort and wild population size influence the prevalence of hybridization between escaped farmed and wild Atlantic salmon. Aquaculture Environment Interactions 10, 401–11.
Sylvester EVA, Wringe BF, Duffy SJ, Hamilton LC, Fleming IA, Castellani M, Bentzen P, Bradbury IR. 2019. Estimating the relative fitness of escaped farmed salmon offspring in the wild and modelling the consequences of invasion for wild populations. Evolutionary Applications 12, 705–17.
Sægrov H, Hindar K, Kålås S, Lura H. 1997. Escaped farmed Atlantic salmon replace the original salmon stock in the River Vosso, western Norway. ICES Journal of Marine Science 54: 1166–1172.
Taleb, NN. 2007. The Black Swan: The Impact of the Highly Improbable, Penguin Books, Limited. London 366 s.
Taranger GL, Svåsand T, Bjørn PA, Jansen PA, Heuch PA, Grøntvedt RN, Asplin L. 2012. Forslag til førstegenerasjons målemetode for miljøeffekt (effektindikatorer) med hensyn til genetisk påvirkning fra oppdrettslaks til villaks, og påvirkning av lakselus fra oppdrett på viltlevende laksefisk. Fisken og havet, 13-2010, Havforskningsinstituttet; Veterinærinstituttets rapportserie Nr. 7–2012.
Taranger GL, Karlsen O, Bannister RJ, Glover KA, Husa V, Karlsbakk E, Kvamme BO, Boxaspen KK, Bjørn PA, Finstad B, Madhun AS, Morton HC, Svåsand T. 2015. Risk assessment of the environmental impact of Norwegian Atlantic salmon farming. Ices Journal of Marine Science 72, 997–1021.
Thodesen J, Grisdale-Helland B, Helland SJ, Gjerde B. 1999. Feed intake, growth and feed utilization of offspring from wild and selected Atlantic salmon (Salmo salar). Aquaculture 180:237-246.
Vollset KW, Urdal K, Utne K, Thorstad EB, Sægrov H, Raunsgard A, Skagseth Ø, Lennox RJ, Østborg GM, Ugedal O, Jensen AJ, Bolstad GH, Fiske P. 2022. Ecological regime shift in the Northeast Atlantic Ocean revealed from the unprecedented reduction in marine growth of Atlantic salmon. Sciences advances 8:9.
Verspoor E. 1988. Reduced genetic variability in first-generation hatchery populations of Atlantic salmon (Salmo salar). In: Verspoor, E., Stradmeyer, L., Nielsen J.L. (Eds.) The Atlantic salmon: genetics, conservation and management. Blackwell Publishing, Oxford. Canadian Journal of Fisheries and Aquatic Sciences 45: 1686–1690.
Verspoor E. 1997. Genetic diversity among Atlantic salmon (Salmo salar L.) populations. ICES Journal of Marine Science 54: 965–973.
Volpe JP, Taylor EB, Rimmer DW, Glickman BW. 2000. Evidence of natural reproduction of aquaculture escaped Atlantic salmon (Salmo salar) in a coastal British Columbia river. Conservation Biology 14: 899–903.
Wacker S, Aronsen T, Karlsson S, Ugedal, Diserud OH, Ulvan EM, Hindar K, Næsje T. 2021. Selection against individuals from genetic introgression of escaped farmed salmon in a natural population of Atlantic salmon. Evolutionary Applications, 14: 1450-1460.
Wargelius A, Leininger S, Skaftnesmo KO, Kleppe L, Andersson E, Taranger GL, Schulz RW, Edvardsen RB. 2016. Dnd knockout ablates germ cells and demonstrates germ cell independent sex differentiation in Atlantic salmon. Scientific Reports 6:21284.
Webb JH, McLaren IS, Donaghy MJ, Youngson AF. 1993. Spawning of farmed Atlantic salmon, Salmo salar L., in the second year after their escape. Aquaculture and Fisheries Management 24: 557–561.
Webb JH, Verspoor E, Aubin-Horth N, Romakkaniemi A, Amiro P. 2007. The Atlantic Salmon. Chapter 2. In: The Atlantic salmon: genetics, conservation and management. Verspoor E., Stradmeyer, L.& Nielsen J.L.(Eds). Blackwell Publishing, Oxford, pp. 17–56.
Wennevik V, Quintela M, Skaala Ø, Verspoor E, Prusov S, Glover KA. 2019. Population genetic analysis reveals a geographically limited transition zone between two genetically distinct Atlantic salmon lineages in Norway. Ecology and Evolution 9(12):6901-6921.
Wennevik mfl. Rømt oppdrettslaks i vassdrag i 2020 - rapport fra det nasjonale overvåkningsprogrammet. Rapport fra Havforskningen 2021-27.
Wennevik mfl. Rømt oppdrettslaks i vassdrag i 2021 - rapport fra det nasjonale overvåkningsprogrammet. Rapport fra Havforskningen 2022-21.
Wringe BF, Jeffery NW, Stanley RRE, Hamilton LC, Anderson EC, Fleming IA, Grant C, Dempson JB, Veinott G, Duffy SJ, Bradbury IR. 2018. Extensive hybridization following a large escape of domesticated Atlantic salmon in the Northwest Atlantic. Communications Biology 1.
Zhang Z, Glover KA, Wennevik V, Svåsand T, Sørvik AGE, Fiske P, Karlsson S, Skaala Ø. 2013. Genetic analysis of Atlantic salmon captured in a netting station reveals multiple escapement events from commercial fish farms. Fish Manage Ecol 20:42-51.