Rapportserie:
Rapport fra havforskningen 2025-25ISSN: 1893-4536Publisert: 23.04.2025Prosjektnr: 15954Oppdragsgiver(e): Fiskeri og Havbruksnæringens Forskningsfond (FHF)Referanse: 901831 Forskningsgruppe(r):
Smittespredning og sykdomProgram:
Miljøeffekter av akvakultur
Forskningsgruppeleder(e):
Monica F. Solberg (Smittespredning og sykdom)
Godkjent av:
Forskningsdirektør(er):
Geir Lasse Taranger
Programleder(e):
Mari Skuggedal Myksvoll
Assessment of five different Spironucleus salmonicida infection models for post-smolt Atlantic salmon in seawater conditions
SpiroFri Pilot Infection Trial Spring 2023
Five different infection models for Spironucleus salmonicida , a diplomonad parasite associated with severe outbreaks of spironucleosis in Norwegian aquaculture, were evaluated. The parasite, isolated from lesions in infected salmon during an outbreak, was cultured according to established protocols and subsequently administered to post-smolt salmon using five different methods: bath exposure, intraperitoneal injection, oral intubation, anal intubation, and cohabitation. The primary objective of the experiment was to determine which model would be most suitable for inducing infection in seawater for the SpiroFri project.
The experiment lasted for six weeks, during which fish were monitored for growth, clinical signs, and lesions, and tested for infection in blood and intestinal fluid samples using qPCR. After six weeks, only minimal differences in weight gain were observed among the groups, and no clinical signs of spironucleosis were detected. qPCR analysis of blood and intestinal fluid samples showed no S. salmonicida infection in the cohabitant group, and only one positive fish in the anal intubation group. Furthermore, the qPCR results indicated that oral intubation led to a high prevalence of S. salmonicida in the blood at three weeks post-infection, whereas the injection and bath models showed high prevalence at six weeks post-infection.
Based on these findings, and considering the advantages and disadvantages of each infection model, the bath infection model, being a minimally invasive method representative of natural infection routes, was selected for continued work in the project.
Sammendrag
Fem ulike smittemodellar for Spironucleus salmonicida, ein diplomonad parasitt knytt til alvorlege utbrot av spironukleose i norsk akvakultur er vurdert. Parasitten, isolert frå lesjonar hos infisert laks, vart dyrka i henhald til etablerte protokollar og deretter administrert til postsmolt laks ved hjelp av fem ulike metodar: bad, intraperitoneal injeksjon, oral intubasjon, anal intubasjon og kohabitasjon. Hovudmålet for forsøket var å fastslå kva modell som var best å nytte til smitte i sjøvatn i prosjektet SpiroFri.
Forsøket varte 6 veker og fisken vart undersøkt for vekst, kliniske teikn og lesjonar, samt testa for infeksjon i blod ‐ og tarmvæskeprøvar ved hjelp av qPCR. Etter 6 veker var det berre minimale skilnader i vektauke mellom gruppene, og ingen kliniske teikn på spironukleose vart observert. Ved analyse av blod og tarmvæske med qPCR vart det ikkje funne S. salmonicida infeksjon i kohabitantgruppa, og kun ein positiv fisk i den anal-intuberte gruppa. Vidare viste qPCR-resultata at det ved oral intubasjon var høg prevalens av S. salmonicida i blodet 3 veker etter smitte, medan injeksjons- og badmodellane viste høg prevalens først 6 veker etter smitte.
Basert på funna, og den enkelte smittemodells fordelar og ulemper, vart badsmittemodellen, som er ein lite invasiv metode representativ for naturlege infeksjonsvegar, vald som metode for det vidare arbeidet i prosjektet.
1 - Prosjekt
Spironucleus salmonicida infeksjonar og spironukleose i norsk akvakultur:
smitteveier, vertsspekter og forebygging
SpiroFri
FHF-prosjekt 901831
HI-prosjekt 15954
Delrapport 3.1
2 - Introduksjon
Etter omfattande utbrudd av spironucleose i Nord-Norge i 2022 (Sommerset, Wiik-Nielsen et al. 2023, Sommerset, Wiik-Nielsen et al. 2024), lyste FHF ut midler til å undersøke smittevegar og desinfeksjonsstrategiar for parasitten Spironucleus salmonicida . Prosjektet « Spironucleus salmonicida infeksjoner og spironucleose i norsk akvakultur: smitteveier, vertsspekter og forebygging (SpiroFri)» fekk tildelt prosjektmidlar (FHF prosjektnummer 191831). SpiroFri-prosjektet er leia av Havforskingsinstituttet, og gjennomførast i samarbeid med Nofima Tromsø, Veterinærinstituttet, Pure salmon Kaldnes v/ Erik Sterud, og Grieg Seafood ASA. Formålet med prosjektet er å få frem kunnskap for førebygjing og bekjemping av S. salmonicida , både med tanke på laksefisk, men òg rensefisk og villfisk.
Diplomonadar er små anaerobe (mikroaerofile) flagellatar (Adam 2017) som er frittlevande eller symbiontar i fordøyingssystemet til dyr. Nokre av desse flagellatane kan vera skadelige parasittar og forårsake sjukdom. Mest kjend av desse er nok Giardia intestinalis , som kan gje plagsom magesjuke hos menneske. Spironucleus artane lever særleg i tarmen hjå kaldblodige dyr som fisk, amfibiar og skilpadder, men der er og nokre artar i tarmen på fugl og pattedyr som er klassifisert i denne slekta. Fleire av artane forårsakar sjukdom. I fisk er fleire av spironucleus artane patogene, inkludert S. vortens , S. salmonis , og S. salmonicida (Uldal and Buchmann 1996, Paull and Matthews 2001, Poynton, Fard et al. 2004, Jorgensen and Sterud 2006, Fard, Jorgensen et al. 2007).
I Nord-Noreg har S. salmonicida gjenntatte gongar forårsaka sjukdomsutbrot i oppdrett av laks. Arten var først skildra og navngjeven i 2006, og flagellaten vart tidlegare identifisert som Hexamita sp., Hexamita salmonis og som Spironucleus barkhanus. S. salmonicida (då som Hexamita sp.) vart først påvist i 1989, i samband med eit utbrot av spironukleose i oppdrettslaks i Nord-Noreg (Mo, Poppe et al. 1990, Poppe, Mo et al. 1992). Etter 1989, der 6 lakselokalitetar vart affisert, er det påvist utbrot på ein lokalitet med oppdrettslaks i 2001 (Sterud, Poppe et al. 2002), nokre lakselokalitetar i 2007-08 (Skjelstad, Bornø et al. 2008, Johansen, Kongtorp et al. 2009), samt ved ein oppdrettslokalitet for røyr i 2002 (Sterud, Poppe et al. 2003). Det hittil siste, og største, utbrotet av spironukleose i norsk oppdrett var i 2022-2023 der 10-12 matfisklokalitetar vart råka (Lie, Wisløff et al. 2022, Sommerset, Wiik-Nielsen et al. 2023, Sommerset, Wiik-Nielsen et al. 2024). Utbrota av spironukleose har vore alvorlege med store konsekvensar for affiserte anlegg, med tilfeller av destruksjon, høg dødelegheit, redusert produksjon samt nedklassifisering av fisken (Mo, Poppe et al. 1990, Sterud, Poppe et al. 2002, ASA 2023). Utbrotet som starta i 2022 er det hittil største, med estimerte tap for oppdrettar opp mot 900 millionar norske kroner (Grieg Seafood, 2023).
Det er framleis lite kunnskap om livssyklus, smittevegar og infeksjons- og sjukdomsdynamikk for S. salmonicida . Det er publisert resultat frå fire ulike smitteforsøk med S. salmonicida i laks i ferskvatn. Etter eit utbrot av spironukleose på oppdretta kongelaks (chinook salmon, Oncorhynchus tshawytscha ) i sjøvatn i 1991 gjennomførte Kent mfl. (1992) smitteforsøk med kongelaks og atlantisk laks (Kent, Ellis et al. 1992). Laks vart intubert med infektivt materiale, men i løpet av dei ca fem vekene intubasjonssmitteforsøket pågjekk, vart atlantisk laks ikkje påviseleg smitta. Kongelaks vart derimot smitta med fleire ulike smittemodellar i dette forsøket, og det var høg dødelegheit over kort tid (1.5-4.5 veker). Parasitten på kongelaks i British Columbia er S. salmonicida , men er nok ein annan variant enn me har i Noreg. Som oppfølging av spironukleoseutbrotet i Noreg i 1989-1991 gjorde Mo mfl. (1993) kohabitasjonssmitteforsøk med laks, aure, røyr og regnbogeaure i brakkvatn (25 ‰) og ferskvatn (Mo, Poppe et al. 1993). Smittekjelda i forsøket var stor laks med spironukleose. Etter 26 dagar var kun ein laks smitta i brakkvatn. For å undersøke smittevegar og infeksjonsdynamikk gjennomførte Guo og Woo (2004) smitteforsøk med S. salmonicida på laks i ferskvatn (Guo and Woo 2004). Laks nytta som smittespreiarar («sheddarar») vart smitta med injeksjon i bukhola (intraperitonal injeksjon; i.p.), og kohabitantar tilsett. All injisert fisk vart infisert, og 50% av kohabitasjonsfisken var smitta etter 4-6 veker. I løpet av forsøksperioden på inntil 21 veker etter smitte utvikla fisken byllar og sår, og det var høg dødelegheit. I 2018 gjorde Alfjorden smitteforsøk med laks i ferskvatn med to ulike dosar parasitt pipettert direkte ned i magen (oral intubasjon) eller injisert intraperitonalt i fisken (Alfjorden 2018, Alfjorden, Astvaldsson et al. 2019). Vekentlege blodprøvar viste aukande prevalens til 100% i dei 10 vekene forsøket gjekk, men det var lite lesjonar og ingen fisk døydde.
Ved Havforskingsinstituttet vart det i SpiroFri-prosjektet planlagt gjennomført tre større smitteforsøk i sjøvatn med laks, aure, røyr og rognkjeks for å undersøke mottakelegheit, smittevegar og konsekvensar av infeksjon i desse artane i sjøvatn. Som følgje av den manglande kunnskapen om smitteforsøk med S. salmonicida i sjøvatn var det naudsynt å teste ut metodikk. Det vart derfor gjort ei pilotstudie for å få erfaring med gjennomføring av smitteforsøk med S. salmonicida , samt å avgjere kva smittemodell som var mest tenleg å nytte i dei større smitteforsøka. I pilotforsøket skildra her vart parasitten først isolert frå laks med spironukleose, og dyrka etter tidlegare dyrkingsprotokollar. Parasittar frå kulturar vart so nytta til å teste fem ulike smittemodellar: Bad, intraperitonal injeksjon, oral intubasjon, anal intubasjon, og kohabitasjon. Fisken vart undersøkt midtvegs i forsøket og ved slutt, og det vart tatt tarmvæske og blodprøve for påvising av S. salmonicida med qPCR (kvantitativ PCR, sanntids PCR). Ved sluttuttaket vart fisken òg undersøkt for indre og ytre teikn til spironukleose.
3 - Metodar
3.1 - Dyrking av S. salmonicida til smitteforsøk
Spironucleus salmonicida vart isolert frå lesjonar frå laks frå anlegg med utbrot. Antatt infisert laks vart avliva, pakka på is og umiddelbart sendt frå oppdrettslokalitet til Havforskingsinstituttet i Bergen med over-natt transport. I Bergen vart fisken undersøkt for indre og ytre teikn til lesjonar. Der det vart funne lesjonar som truleg skuldast S. salmonicida vart væske frå lesjonen undersøkt i mikroskop. Ved bekrefta infeksjon og tilstadeværelse av S. salmonicida vart væske frå lesjonen aseptisk tatt ut ved hjelp av sprøyte, overført til dyrkingsmedium i tette 15 ml prøverøyr og inkubert ved 4 ° C.
Suksessfulle isolater vart dyrka i henhald til standard dyrkingsbetingar. Modifisert Keister’s medie (ATCC 2695 Medium; Keisters modifiserte Giardia TYI-S-33) utan glukose, og med MEM vitaminløysing (Sigma Aldrich, M6895) i staden for Diamonds vitamintilsetjing (Merck, 58980C) tilsatt, vart nytta som dyrkingsmedie (Keister 1983). Keisters basemedie bestående av trypton, gjærekstrakt, NaCl, K 2 HPO 4 , KH 2 PO 4 og jernammonium citrat vart laga, justert til pH 7.4 og autoklavert. Før bruk vart basemediet tilsatt oksegalle, cystein, vitaminer og antibiotika (antimycotic). Parasitten vart dyrka aseptisk i celledyrkingsflasker med 25, 75 eller 175 cm2 overflate (Nunc EasyFlask Cell Culture Flasks; Thermo Scientific, henholdsvis art. nr 156340, 156472, 159920) ved 4, 6 eller 10 ° C utan risting. Kulturane vart splitta med 3-6 dagars mellomrom avhengig av vekst. Ved dyrking i 75 cm2 celledyrkingsflasker vart 10 ml kultur overført til 30 ml ferskt medium ved splitting. For å halde kulturane oksygenfrie vart mediet «lufta» med nitrogengass kvar gong flasker eller røyr vart opna og utsett for oksygen (luft). Rein nitrogengass vart bobla ned i mediet i ca 10 sekund og deretter vart flasker/røyr raskt lukka. Kulturar vart overvaka med omvendt mikroskop ved 10-20x, for vekst og vitalitet.
Før smitte vart S. salmonicida frå 75 cm2 kulturflasker, 40 ml kultur, sentrifugert ved 500 x G i 5 min ved 4°C. Supernatant, unntatt 3 ml over pellet, vart fjerna, og pellet dernest blanda/løyst løst opp i PBS/basemedium og sentrifugert ein gong til (500 x G, 5 min, 4°C). Supernatanten vart igjen fjerna og pellet løyst opp i PBS. For kvantifisering vart flagellatar immobilisert med glyserol i PBS (Glyserol:PBS 1:3) og talt med hemocytometer (Kova Glasstic, 87144E) i mikroskop i henhold til produsentens skildring. Kvar prøve vart talt tre gongar. Røyra stod på is under arbeidet.
3.2 - Fisk
Laks post-smolt (127 ±14.8 g, 22.5 ±0.9 cm) (Tabell 1) til forsøket vart kjøpt frå Industrilaboratoriet i Bergen (ILAB). Fisken var av Stofnfiskur (Benchmark Genetics Island) stamme. Fisken vart transportert frå ILAB (Thormøhlensgt. 55, Bergen) med bil til Havforskingsinstituttets smittelaboratorie (Nordnesgaten 50, Bergen) 11. januar 2023. Før transport til Havforskingsinstituttet vart fiskegruppa testa for patogene agens i henhold til ILAB sine rutiner.
Etter mottak vart fisken sett i eit 600 L kar med 350 L 24.0±2.3 ‰ saltvatn time-1, ca 9 ° C og 12:12 L:D lysregime fram til smittedagen, 1. februar. Etter smitte vart fisk fordelt i 6 kar à 250L med 30 fisk i kvart kar (Tabell 1), med same vassparametre som før smitten. Gjennom heile forsøksperioden vart fisken fora med Nutra RC3, 3 mm fôr (Skretting AS). Fisken vart røkta minimum to gongar/dag, og temperatur, salthald og oksygen vart målt dagleg.
3.3 - Smitteforsøk
Ved forsøksstart vart det satt opp 6 ulike smittegrupper (Tabell 1). Etter smitte vart all fisken halden i 250L kar med same vasskvalitet, lysregime og foring som i aklimatiseringsperioden. Fisk som vart fysisk handtert under smitteprosedyra (intubasjon og i.p.) vart bedøvd med 100 mg/L Trikainmesilat (Finquel vet., MSD Animal Health) før smitte vart gjennomført. Etter smitte vart bedøvd fisk lagt til oppvakning og overvaka inntil den hadde tilnærma normal symjing.
Forsøksgruppe
N
Snitt (g)
Median (g)
Min (g)
Max (g)
SD (g)
95% c.i.
Kontroll
30
124,2
125,1
101,2
152,6
13,1
120-130
Anal intubering
31
125,0
123,4
99,4
154,8
13,6
127-138
Oral intubering
30
132,5
132,0
109,6
163,8
13,9
119-131
Badsmitte
30
124,7
123,0
96,8
169,0
15,4
119-129
Kohabitasjon
301 (15/kar)
131,4
133,2
82,0
159,4
16,2
125-138
Injeksjon (i.p.)
301 (15/kar)
125,0
128,6
85,4
151,0
14,2
120-130
Tabell 1. Oversikt over forsøksgruppene i forsøket og startvekt i gram (g).
1 Injeksjonssmitta fisk vart nytta som sheddarar for kohabitasjonsmitta fisk.
Kontrollfisk vart håva frå aklimatiseringskaret og over i forsøkskaret utan ytterlegare behandling.
For badsmitte vart ein balje med 30 laks, 50 L vatn, 25 ‰ sjøvatn med lufting tilsett ca 500 millionar S. salmonicida (~106 parasittar L-1). Fisken vart halden i smitten i 60 minutt med kontinuerlig overvaking av oksygennivå (92,2-93,3 % O2 ) og so sett over i forsøkskaret.
Figur 1. Illustrasjon av prosedyre for oral (øvst) og anal (nedst) intubering (foto: Bjørn Olav Kvamme).
Ved oral og anal intubasjon (Figur 1) vart laks bedøvd i mindre grupper (2-4 fisk) for å sikre god kontroll på tid i bedøvinga, og deretter smitta. Ved oral intubasjon vart 104 flagellatar i 100 µL fysiologisk saltvatn deponert i magesekken ved hjelp av ei 1 mL sprøyte og ein mjuk gummislange (fisken halden vertikalt). Ved anal intubasjon vart tilsvarande mengde og dose parasittar deponert 2.5-3 cm inn i baktarm ved hjelp av 1 mL sprøyte påsatt ein mjuk tynn plasttip (Sigma-Aldrich; CAD9928).
Ved injeksjonssmitte (i.p.) vart 104 flagellatar i 100 µL PBS injisert i bukhula på bedøvd fisk. Injeksjon vart gjort med insulinsprøyte med 25G kanyle der stikkdjupn var tilpassa fiskens storleik og lik for kvar fisk. Injeksjonssmitta fisk hadde og funksjon som sheddarar for uttesting av kohabitasjonssmitte. Denne fisken vart difor sett i to kar med 15 injiserte fisk i kvart. For å skilje i.p. smitta frå kohabitantar vart i.p. fisken fettfinneklipt samtidig med smitte.
Kohabitasjonssmitte vart gjennomført ved å kohabitere 15 laks saman med 15 i.p. infisert laks i to kar, totalt 30 kohabitantar. Laksen i kohabitasjonsgruppa vart håva direkte frå aklimatiseringskaret til forsøkskaret dag en etter smitte (2. februar).
Forsøket er godkjent av Forsøksdyrforvaltningens tilsyns- og søknadssystem (FOTS-ID: 29986).
3.4 - Prøvetaking
Det vart teke blodprøve og tarmprøve av alle fisk 3 (midtuttak; 3 VES) og 6 (sluttuttak; 6 VES) veker etter smitte (VES). Ved sluttuttaket vart all fisk i tillegg undersøkt for ytre og indre sjukdomsteikn. Fisken vart avliva, og visuelt undersøkt utvendig for sår og «bulkar» i huda (typisk ved bylldannelse under huda og i muskelen). Det vart og følt med fingrane langs sidene og rundt halerota for usynlege klumpar/byllar. Dernest vart buken åpna og hjerte, lever, milt, tarm, og nyre inspiser for avvik (t.d. lesjonar/teikn på spironukleose). Til sist vart muskulaturen snitta på langs for å sjå etter lesjonar. Ved avvik (moglege lesjonar) vart det mikroskopert på nativpreparat på objektglas for påvising av S. salmonicida . Det vart då og tatt prøvar for histologi. Etter prøveuttak og undersøking 6 VES vart all fisk frosne enkeltvis i posar (-20 ° C) for eventuell seinare prøvetaking.
Blodprøver vart tekne med 1 mL sprøyter og 0.6G kanylespissar, frå dei caudale blodkara. Volumet var 100 µL, tatt forsiktig for å ikkje gje interne skader på fisken, og deretter overført til prøverøyr tilsatt antikoagulant (10 µl 20mM EDTA). Prøva vart halden på is inntil den vart frosen på -20 ° C.
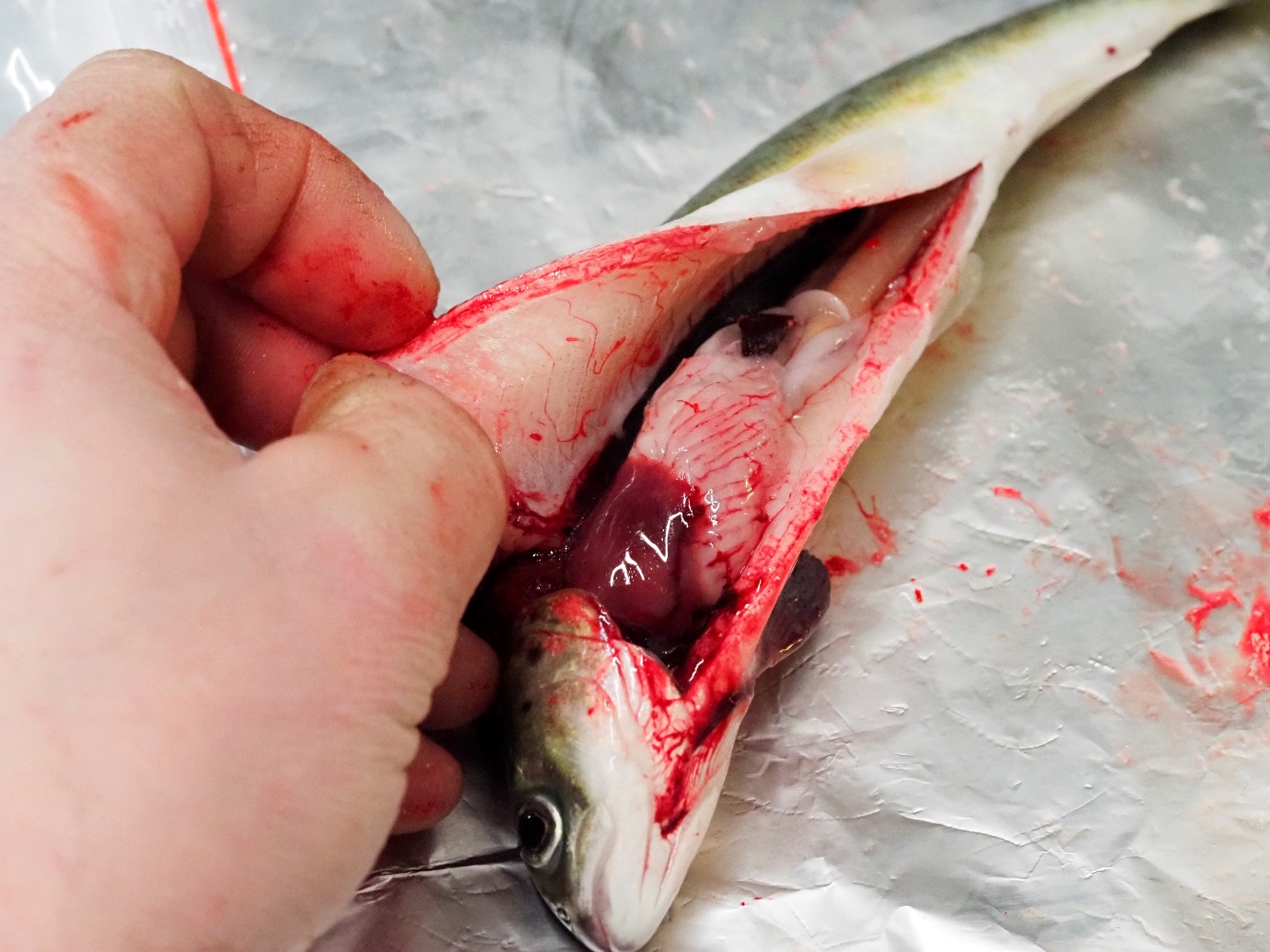
Figur 2. Illustrasjonsfoto (foto: Bjørn Olav Kvamme). All fisk vart undersøkt for ytre og indre teikn til spironukleose 6 VES utan funn.
Tarmprøva (tarmvæske) vart teken med pipette påsatt avrunda spiss. Spissen vart ført forsiktig inn gjennom endetarmsåpningen og 200 µ l PBS forsiktig sprøyta inn i tarmen. Deretter vart utsiden av fisken forsiktig massert for blanding før 100 µl væske vart sugd ut igjen. Tarmvæska vart overført til eppendorfrøyr på is og prøvane frosne ved -20 ° C.
3.5 - Rensing av genomisk DNA
Rensing av DNA frå blodprøvar vart gjort med Genefinder V3 kit (Beckman Coulter) på ein Biomek i5 Automated Workstation (Beckman Coulter) i 96-brønnars brett. Blodprøvane vart tint og deretter homogenisert ved hjelp av risting med stålkuler. Frå den homogeniserte prøven vart 5 µl overført til 195 µl PBS-buffer og DNA rensa i henhold til produsentens anbefalingar. DNA vart eluert i 50 µl elueringsbuffer og lagra på -20 ° C inntil analysert. Genefinder protokollen vart i all hovudsak fulgt med nokre optimaliseringar av det automatiske programmet for arbeidsstasjonen.
Rensing av DNA frå tarmprøvar vart gjort med DNAdvance kit (Beckman Coulter) på ein Biomek i5 Automated Workstation (Beckman Coulter) i 96-brønnars brett. I kortheit vart tarmvæske tint og 40 µl tarmvæske overført til 200 µl PBS, og DNA rensa ihht produsentens anbefalingar. DNA vart eluert i 50µl elueringsbuffer og lagra på -20 ° C inntil analysert.
3.6 - qPCR-analyser
Kvantifisering av S. salmonicida DNA i blod og tarmvæske vart gjort med kvantitativ real-time PCR (qPCR) med EagleTag 2x Universal Master Mix (ROX) (Merck, art.nr. 7260296190) i henhold til produsentens anbefalingar. Rensa DNA frå tarmvæske- og blodprøve vart nytta som templat, enten ufortynna eller fortynna 1:10 med dH2O. Det vart nytta brett med 384 brønnar og totalvolum 8 µl reaksjon-1; 4 µl 2X qPCR Master Mix, 0.4 µl av kvar primer (10 µM), 0.4 µL probe (4 µM), 0.8 µl vatn og 2 µl templat. Primerar og probar er gitt i Tabell 2. Effektiviteten for S. salmonicida qPCR assayet (Tabell 2) vart berekna til mellom 0.87 (Blod) og 1.00 (PBS) (R2 hhv. 0.99 og 0.98) her, og er oppgitt av Miller mfl. (2016) til 0.94-1.03 (R2 0.97-1.00) for ulike analyseplattformer (Miller, Gardner et al. 2016).
All qPCR vart gjennomført i Quantstudio TM 5 Real-Time PCR instrument (384-block) (Applied Biosystems, by Thermo Fisher Scientific) med følgande betingingar: 10m@95°C, 45 syklar 15s@95°C-60s@60°C -1s@72°C, deretter 30s@40°C og ∞@4°C.
Ct-verdiar vart bestemt ved hjelp av QuantStudio TM Design & Analysis Software v1.4.2 (Applied Biosystems), med grenseverdi for berekning av Ct satt til 0.2.
Tabell 2. Primerar og probar nytta for qPCR-analysar. F=forward, R=reverse, P=probe.
§ Glyseraldehyd 3-fosfat dehydrogenase
3.7 - Vassprøvar
Det vil vera nyttig å tidfeste starten på smittefrigjering («shedding») frå den S. salmonicida smitta fisken. For å påvise S. salmonicida i karvatn gjennom infeksjonsforløpet vart vatn pumpa frå nokre kar med fisk smitta med S. salmonicida , filtrert, og analysert ved hjelp av qPCR. Kara var anal intubering, oral intubering, og badsmitte. Frå desse tre forsøkskara vart 2 L vatn pumpa over på plastbehaldarar to gongar kvar veke gjennom forsøket. Som negativ kontroll vart det tatt vatn frå vassinntaket til fiskehelselaboratoriet. Vatnet vart lagra kjølig inntil det vart filtrert. For kvar vassprøve vart 2 L vatn filtrert gjennom eit Millipore Sterivex GP Pressure filter (Millipore, Prod.nr: SVGPL10RC). DNA vart deretter ekstrahert frå filteret ved hjelp av DNeasy PowerWater Sterivex Kit (Qiagen) i henholdt til produsentens anbefalingar og eluert i 100 µl.
4 - Resultat
4.1 - Fysiologiske data
Vekt og lengd vart målt for all fisk ved kvart uttak. Det var kun signifikant forskjell i vekst for heile forsøksperioden (6 veker) for analintubasjon og kohabitasjon (Figur 3, Tabell 3). Det var ingen signifikante forskjellar i vekt mellom dei ulike gruppene ved sluttuttaket 6 veker etter smitte (VES). Det var heller ikkje signifikant forskjell i kondisjonsfaktor innad i smittegruppene for forsøksperioden, eller mellom dei ulike gruppene ved forsøksslutt (6 VES) (ikkje vist).
Fysisk undersøkjing av fisken for lesjonar og mikroskopiering for påvising av S. salmonicida 6 VES resulterte i funn av ein fisk med lysare flekkar i midtnyra. Det vart gjort mikroskopi og teke full prøvepakke av for utvida undersøkjing, men S. salmonicida vart ikkje påvist i blod, tarm, organer eller nyrelesjon ved qPCR. Det var derfor ingen indre eller ytre teikn foreinleg med spironukleose på fisk 6 VES.
Det var ikkje mortalitet knytt til infeksjon med S. salmonicida i forsøksperioden .
Figur 3. Tukey-plot for vekt av alle gruppene ved start (0 VES), midtuttak (3 VES) og sluttuttak (6 VES). Signifikante forskjellar mellom start og sluttuttak vist (P< 0.01 (**), 0.001 (***)). Det var ingen signifikant forskjell i vekt ved forsøksslutt mellom smittegruppene.
Gruppe
VES
Snitt (g)
Median
Min
Max
SD
95% c.i.
SGR
Kontroll
0
124,2
125,1
101,2
152,6
13,1
119-129
0,27
3
130,4
131,5
106,2
166,0
15,0
125-136
6
139,1
138,2
99,0
184,4
20,5
131-147
Anal
0
125,0
123,4
99,4
154,8
13,6
120-130
0,38
3
132,2
130,4
98,0
169,2
16,6
126-138
6
146,4
143,2
110,8
193,6
20,6
139-154
Bad
0
132,5
132,0
109,6
163,8
13,9
127-138
0,25
3
131,4
135,9
16,8
167,0
25,1
131-142
6
147,0
146,6
116,8
183,2
18,0
140-154
Kohab
0
124,7
123,0
96,8
169,0
15,4
119-131
0,33
3
134,3
131,4
108,4
169,4
16,0
128-140
6
143,0
142,3
108,8
186,4
20,0
135-151
I.P.
0
131,4
133,2
82,0
159,4
16,2
125-138
0,22
3
133,6
136,2
82,0
180,8
18,3
126-141
6
143,8
145,2
83,2
198,6
20,9
136-152
Oral
0
125,0
128,6
85,4
151,0
14,2
120-130
0,25
3
127,3
132,5
92,2
156,2
15,6
121-133
6
138,9
137,6
105,6
168,8
16,3
133-145
Tabell 3. Oppsummering vektdata for alle gruppene for 0, 3 og 6 VES. SGR er berekna frå 0 til 6 VES (42 dagar).
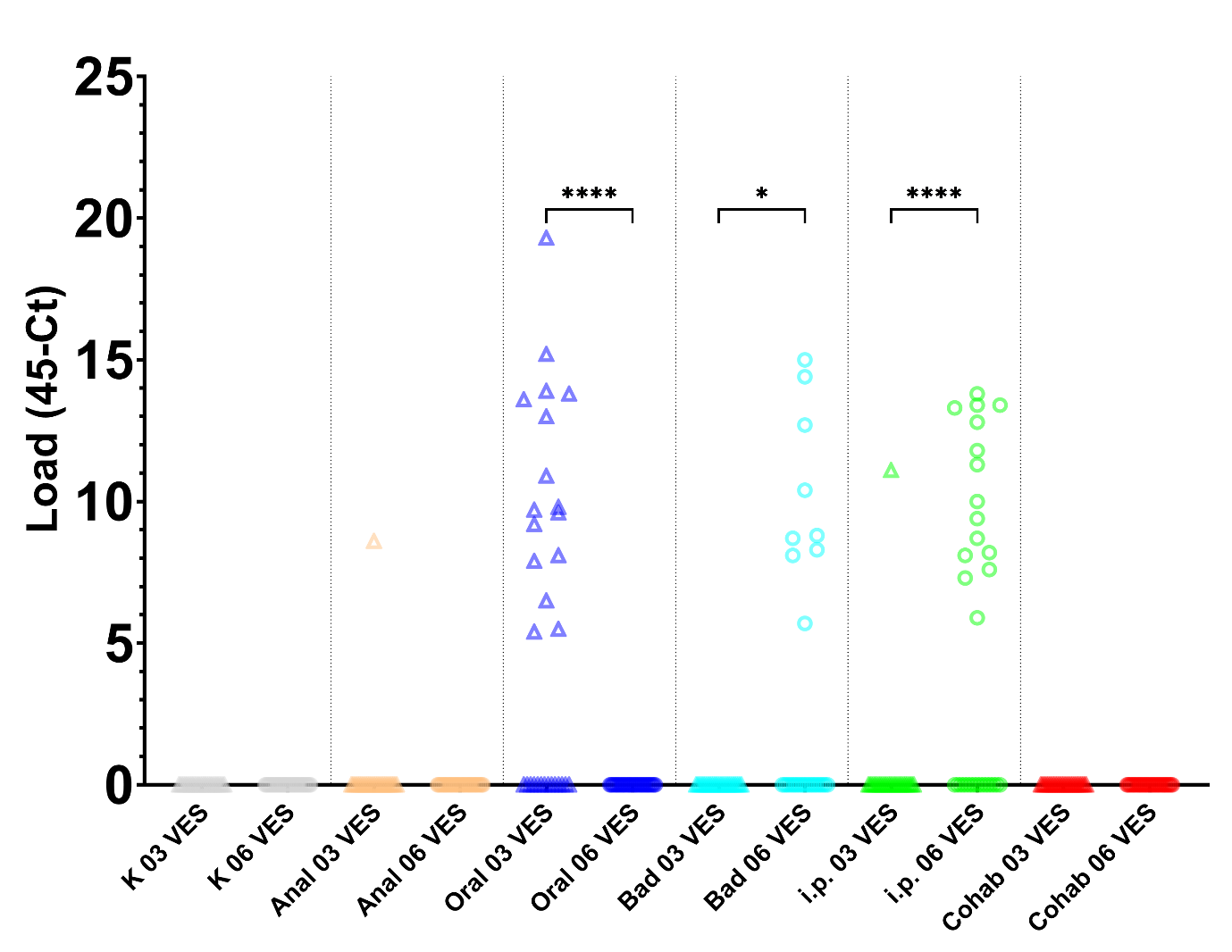
4.2 - Kvantitering av S. salmonicida i blod og tarmvæske med qPCR
qPCR analyser av blod er oppsummert i Figur 4 og 5, og i Tabell 4. Det vart ikkje påvist S. salmonicida i blod eller tarm frå fisk i kontrollgruppa eller kohabitasjonsgruppa ved noko uttak.
I gruppa med anal intubasjonssmitte vart det kun påvist S. salmonicida i blod i ein fisk 3 veker etter smitte. Det var ingen fisk som var positive for S. salmonicida i blod 6 veker etter smitte, og i tarm var det ingen positive fisk ved noko uttak.
Ved oral intubasjonssmitte vart det påvist høg andel (16/24) av fisk med S. salmonicida i blod 3 veker etter smitte. Det vart ikkje funnet fisk med S. salmonicida verken i blod 6 veker etter smitte, eller i tarm for noko uttak.
Badsmitta fisk var negative for S. salmonicida i blod 3 veker etter smitte både i blod og i tarmvæske. 6 veker etter smitte var 9 av 26 fisk positive for S. salmonicida i blod, og 3 av 30 fisk positive i tarmprøva. To av fiskane med positiv tarmprøve hadde ikkje positiv blodprøve, so totalprevalensen var her minst 43% (11/26).
Fisk som vart smitta ved i.p. injeksjon var positive både 3 (1/24) og 6 (15/27) veker etter smitte i blod. Både 3 og 6 veker etter smitte vart det påvist S. salmonicida i tarmen på ein fisk, som hadde negativt blod. Totalprevalensen var difor minst 2/24 og 16/27.
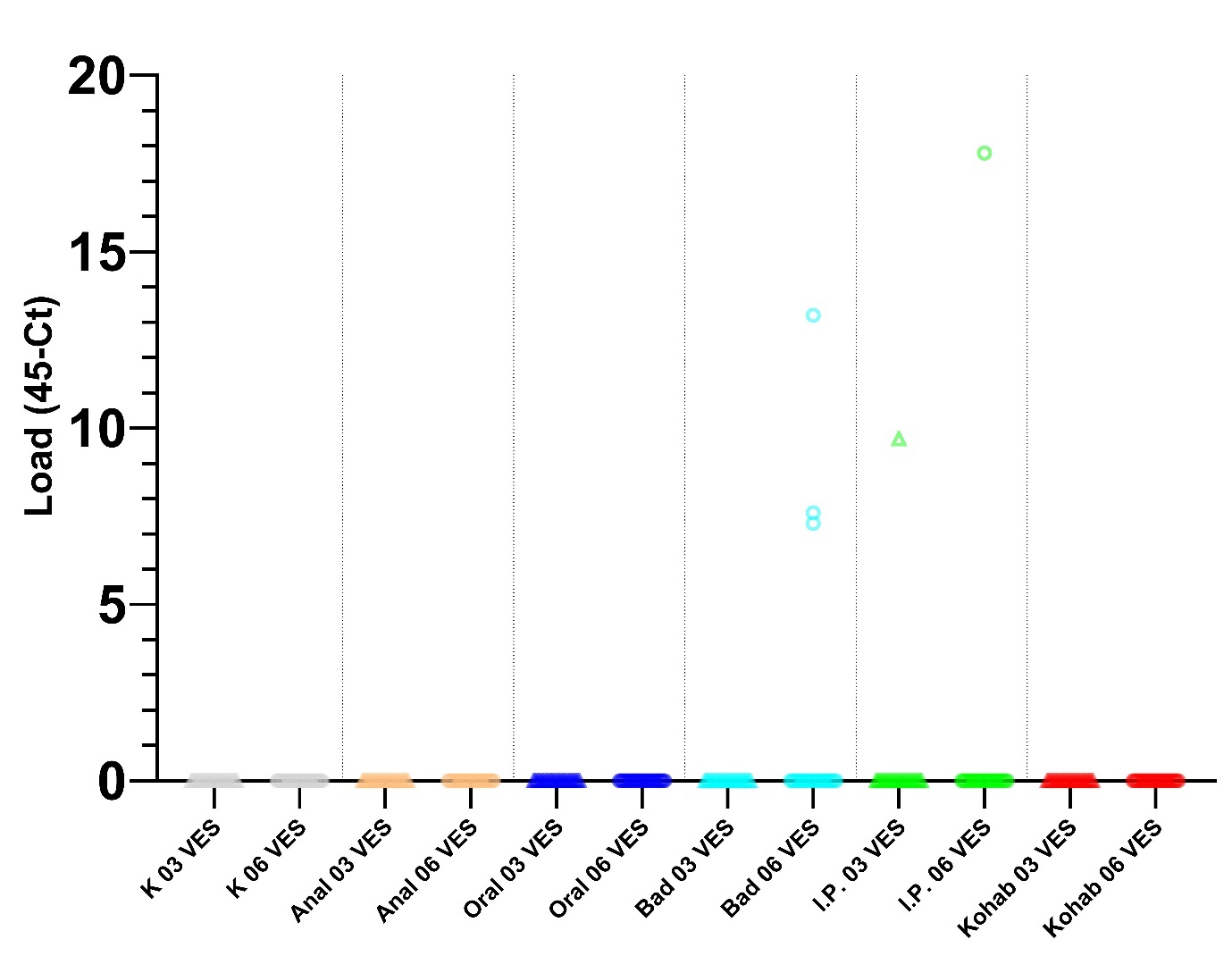
Det var signifikant endring (p<0.05) i gjennomsnittleg Load i blod mellom 3 og 6 VES for Oral intubasjonssmitte, Badsmitte og injeksjonssmitte (i.p.) (Figur 2). For tarm var det ikkje signifikant endringar i gjennomsnittleg load mellom 3 og 6 VES for nokon grupper (Figur 3)
Figur 4. Load (45-Ct) berekna frå qPCR analyser av tarmvæske for alle seks smittegruppene: K-Kontroll, Anal-Anal intubasjon, Oral-Oral intubasjon, Bad-Badsmitte, i.p.-Intraperitonal smitte, og Cohab-Kohabitasjonssmitte. Gruppene er skilt med vertikale linjer. Prøvar tatt 3 veker etter smitte (03 VES), er vist som trekantar, 6 veker etter smitte (06 VES) 0som sirklar. Signifikant endring i gjennomsnittleg Load frå 3 til 6 veker etter smitte er indikert (P< 0.05 (*), 0.0001 (****)).
Figur 5. Load (45-Ct) berekna frå qPCR analyser av tarmvæske for alle seks smittegruppene: K-Kontroll, Anal-Anal intubasjon, Oral-Oral intubasjon, Bad-Badsmitte, I.P.-Intraperitonal smitte, og Cohab-Kohabitasjonssmitte. Gruppene er skilt med vertikale linjer. Prøvar tatt 3 veker etter smitte (03 VES), er markert med trekant, 6 veker etter smitte (06 VES) med sirkel. Det var ikkje signifikant endring i gjennomsnittleg load mellom 3 og 6 VES for noko gruppe.
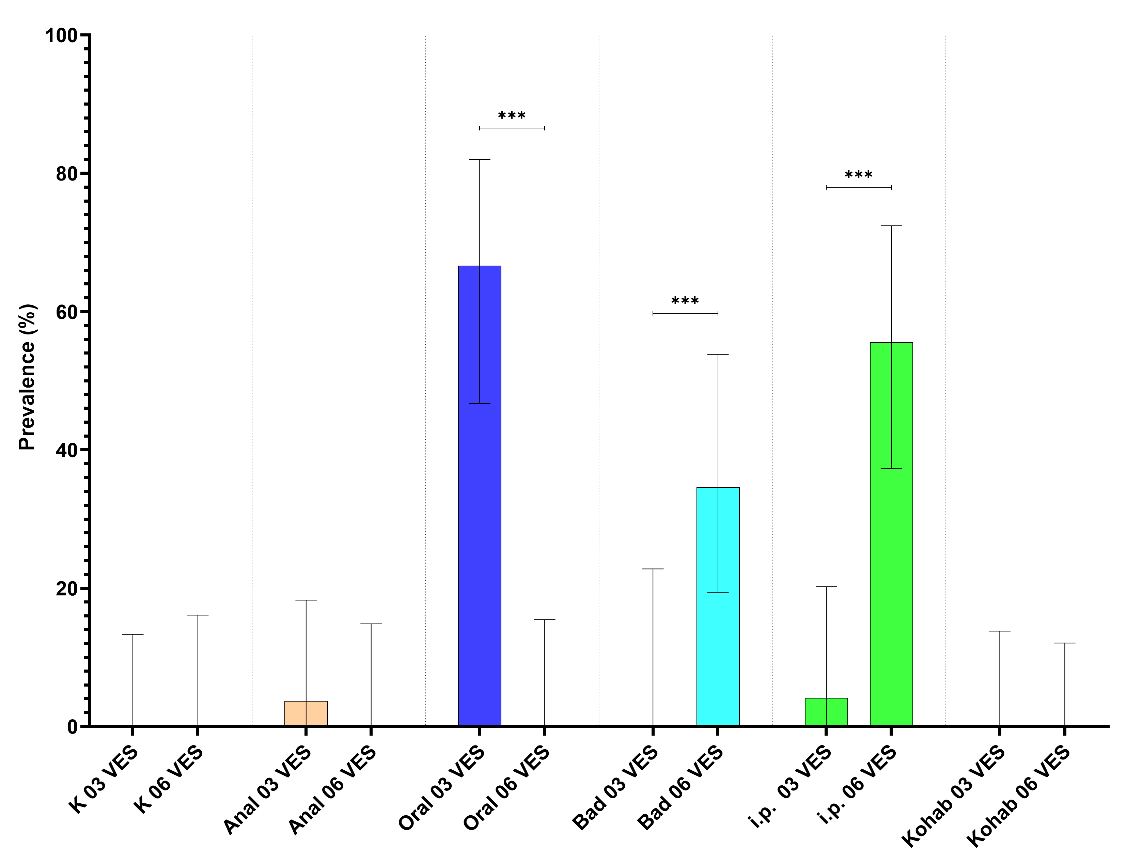
4.3 - Prevalens av S. salmonicida i blod og tarmvæske (qPCR)
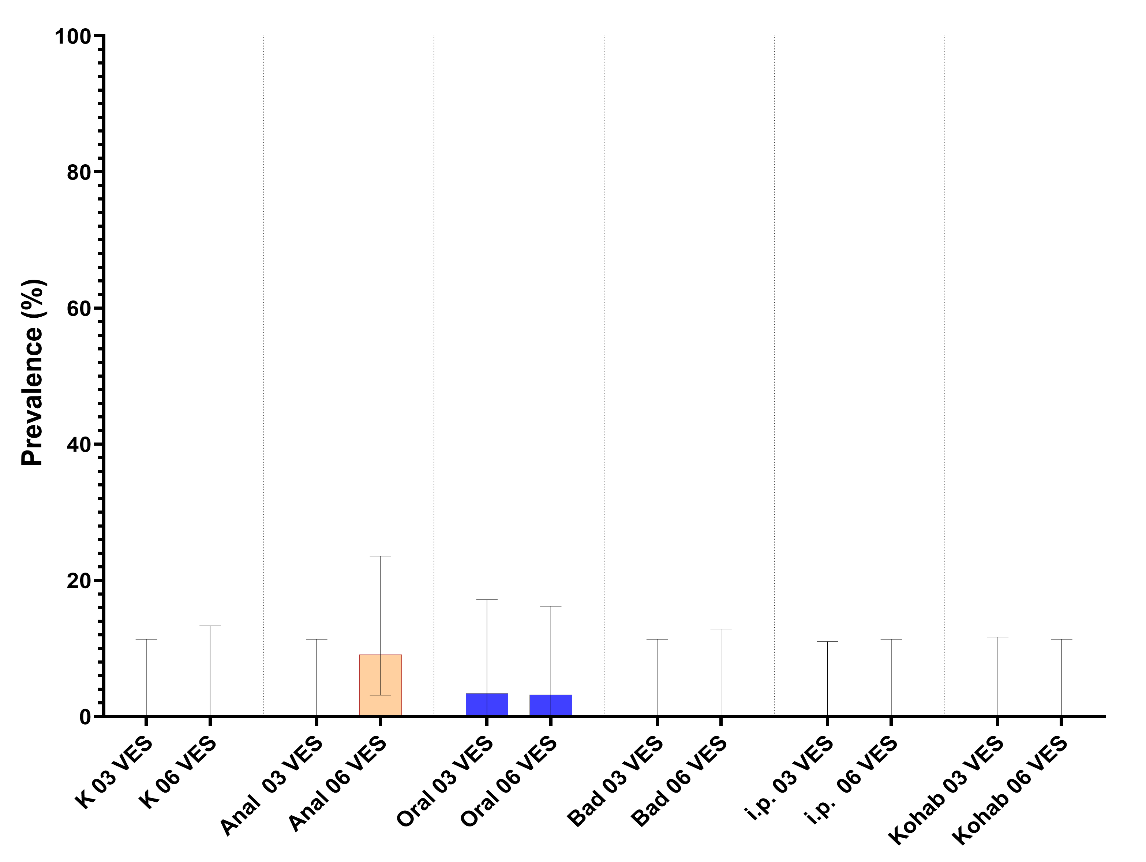
Prevalensen til S. salmonicida basert på blod og tarm analysar med qPCR er vist i Figur 6.
Antall positive / antall analysert*
Tarm
Blod
Veker etter smitte
Smittemodell
GAPDH
Spiro
GAPDH
Spiro
03
Ingen (Kontroll)
30/30
0/30
25/30
0/30 (0/25 * )
Anal intubasjon
30/30
0/30
27/30
1/30 (1/27 * )
Oral Intubasjon
29/30
0/30 (0/29 * )
24/30
16/30 (16/24 * )
Bad
30/30
0/30
13/30
0/30 (0/13 * )
Injeksjon (i.p.)
28/29
1/29 (1/28 * )
24/29
1/29 (1/24 * )
Kohabitering
30/31
0/31
24/31
0/31 (0/24 * )
06
Ingen (Kontroll)
25/30
0/30 (0/25 * )
20/30
0/30 (0/20 * )
Anal intubasjon
30/30
0/26
22/30
0/30 (0/22 * )
Oral Intubasjon
30/30
0/30
21/30
0/30 (0/21 * )
Bad
30/30
3/30
26/30
9/30 (9/26 * )
Injeksjon (i.p.)
30/30
1/30
27/30
15/30 (15/27 * )
Kohabitering
30/30
0/30
28/30
0/30 (0/28 * )
Tabell 4. Oversikt over S. salmonicida positive (Ct<45) fisk i tarm- og blodprøvar. GAPDH (laks) vart nytta som amplifiseringskontroll. Grupper med påvist S. salmonicida er utheva.
1 Antall Spironucleus-positive fisk i forhold til antall amplifiseringskontroll-positive fisk.
Kun oral intubering 3 VES, badsmitte 6 VES og injeksjon 6 VES viste S. salmonicida prevalens signifikant (p<0,001) høgare enn 0.
I gruppa smitta ved oral intubering gjekk prevalens signifikant ned frå 3 til 6 VES (Figur 5). I dei badsmitta og i.p. smitta gruppene auka prevalensen signifikant frå 3 VES til 6 VES (Figur 5).
I tillegg til data vist her er det gjort utvida analyser på blod og vev frå badsmitta fisk (ikkje vist, mastergradsarbeid forventa ferdig juni 2025). Preliminært viser resultat frå fleire qPCR-køyringar på blod og fleire ulike vev 6 VES at faktisk prevalens i badgruppa kan være over 80%.
Figur 6. Prevalens for S. salmonicida i dei eksperimentelle gruppene ved 3 og 6 veker basert på qPCR positive prøvar frå blod (øverst) og baktarmvæske (nederst). Signifikant (P<0,001 (***)) endring i prevalens frå 3 til 6 VES er markert.
4.4 - Lesjonar ved sluttuttaket
Det vart observert moglege lesjonar kun i ein einaste fisk, som hadde eit mindre område med lyse flekkar i midtnyra. Mikroskopi på nativpreparat av prøvar frå fisken, inkludert lesjonane, viste seg negative for S. salmonicida . Histologi gav ikkje svar på årsak.
4.5 - Vassprøvar
Det vart tatt vassprøvar frå kar med anal-, oral- og badinfisert fisk to gongar i veka gjennom forsøket. Metoden brukt viste seg svært arbeidskrevjande, og dei preliminære qPCR-analysene ga begrensa resultat med høg variasjon (ikkje vist). Difor vart det avgjort å ikkje gå vidare med dette arbeidet.
5 - Diskusjon
Målet med smitteforsøka var å etablere ein smittemodell for laks postsmolt i sjø som kunne nyttast seinare for å undersøke kor mottakelege sjøaure, sjørøyr, rognkjeks og eventuelt pukkellaks er for S. salmonicida, med laks som mottakeleg vert (positiv kontroll) i forsøka. Det var ikkje eit mål at fisken skulle utvikle sjukdom, men derimot ein prevalens høg nok til å vise forskjellar mellom artane ( >50% prevalens ynskja).
Det vart brukt låge smittedosar (oral, anal og i.p.), dels fordi tidlegare smitteforsøk ikkje har vist ein klar effekt av dose (Alfjorden 2018; Guo 2007), dels fordi høge dosar mest truleg er lite naturlege. Dette var høg nok dose for å etablere systemiske infeksjonar med i.p., men ikkje nok til å lage sheddarfisk som kunne smitte kohabitantar i løpet av 6 veker. Det var høg nok dose til å etablere infeksjon ved oral intubering, men ikkje ved anal intubering i baktarmen. Det er tidlegare gjort anal intubering av laks med betre resultat (Sommerhaug 1995), då viste enkeltfiskar undersøkt 7, 9 og 11 dagar etter smitte flagellatar i blod eller vev, og dei fleste fiskane var infiserte etter to veker (Sommerhaug 1995). Det vart då sprøyta parasittar heile 9 cm inn frå gattet (til «tynntarmen»). Dette vart ikkje vurdert her av fiskevelferdsgrunnar.
Når det gjeld badsmitte har ein ikkje studiar å samanlikne med. Ein prevalens på ca. 40% etter 6 veker er noko lågt, og høgare dose bør difor brukast for å få høgare prevalens fortare.
Analyser av blod og tarmvæske viste stor variasjon mellom smittemodellane. Det var i tillegg utfordringar med inhibering av PCR-reaksjonen, spesielt i tarmvæske. Dette kan ha påverka resultata noko, og prevalensen påvist her må reknast som ein minimumsprevalens.
Det vart ikkje påvist S. salmonicida infeksjon i kohabitasjonsgruppa. Guo mfl (2004) påviste først S. salmonicida i ein kohabiterande laks fem veker etter smitte i ferskvatn. Resultata her tyder ikkje på raskare smitte i sjø. Men den smitta fisken hadde heller lette infeksjonar, og kan ha frigjeve lite smitte.
Heller ikkje for fisk smitta ved anal intubering vart det påvist S. salmonicida smitte. Dette var ikkje venta. Ei nærliggande forklaring er at sensitiviteten for vår DNA-baserte analyse på tarmvæske er låg, og at ein faktisk infeksjon i tarm i denne modellen ikkje vart påvist, samstundes som infeksjonen ikkje gjekk over i blodet i løpet av forsøksperioden. Ein anna forklaring kan være at smittemetoden, som innebærer å føre ein tynn slange inn i rektum til fisken og deponere parasitten i tarmen, ikkje fungerte optimalt. Dersom parasitten lekker ut igjen, eller fisken tømmer tarmen raskt etter intuberinga, vil det kunne føre til at smittedosen vert for låg til at infeksjon kan etablere seg.
Injeksjon, bad og oral smitte ga alle relativt høg prevalens, enten 3 eller 6 VES.
Oralt smitta fisk hadde høg prevalens allereie 3 VES, og verkar her som å være den smittemodellen som gjev raskast infeksjon i blod. Resultatet stemmer godt med funna gjorde av Alfjorden (2018) (Alfjorden 2018), der oralt infisert fisk var positive etter nokre få veker, med høg prevalens i blod allereie 3 VES.
Injeksjonssmitte ga høg prevalens i dette forsøket først 6 VES. Sidan dette er ein svært direkte smittemodell som omgår mange vernemekanismer mot patogener kunne ein forventa svært rask etablering av infeksjon i denne smittemodellen. Det var derfor uventa at det ikkje vart påvist infeksjon i blod 3 VES i vårt forsøk. Rask påvising og høg prevalens tidleg i forsøket kunne og forventast ut frå resultata til Guo mfl. (2004), der injeksjon av S. salmonicida ga rask parasitemi i blod, og høg prevalens allereie etter 2-4 veker (Guo and Woo 2004). Resultatet her har likevel noko støtte i funna til Alfjorden (2018) (Alfjorden 2018). Her fann ein at i.p. smitta fisk først ga høg prevalens 4 VES. Ei mogleg forklaring på dette er at bukhola ikkje er eit optimalt miljø for etablering av ein S. salmonicida infeksjon. Parasitten kan bruke meir tid til etablere seg og vandre ut i blod, enn ved meir naturleg smitteveg som oral smitte.
For badsmitte vart det påvist mange infiserte fisk først 6 VES. Det er ikkje gjort badsmitte med S. salmonicida tidlegare. At etablering av infeksjon i blodet ville ta lenger tid samanlikna med intubasjon og injeksjonssmitte kunne forventast. Fisken kan smittast ved at fisken drikk vatn (oralt), men moglegvis og ved at parasitten festar seg på overflater på fisken (mukus, auger, gjeller, gatt etc) og deretter unnvik oksygen og trenger inn i vev eller tarm og etablerar infeksjon i blod eller tarm. Smittedosen er derfor avhengig av miljøfaktorar og fiskens adferd, og kan difor variere. Badsmitte kan samanliknast med kohabitasjonssmitte, men der mengda patogen normalt vil vera svært høg, og eksponeringstida mykje kortare og definert. Gitt dette er det naturleg at badsmittemodellen ga høg prevalens raskare enn kohabitasjonsmodellen som vart nytta av Guo mfl (2004), der første fisk med påvist S. salmonicida vart funnet 5 veker etter smittestart (Guo and Woo 2004).
Oppsummert tyder resultata frå 6 VES på at i.p. injeksjon og bading er effektive smittemodellar for S. salmonicida i sjøvatn. Kohabitasjon forventast og å være ein effektivt metode, men som tek ein del lengre tid enn dei andre. Resultat er i tråd med tidlegare smitteforsøk med S. salmonicida (Kent, Ellis et al. 1992, Mo, Poppe et al. 1993, Guo and Woo 2004, Alfjorden 2018).
Av dei testa smittemodellane er det bad og kohabitasjon modellane som gir mest naturleg smitteveg, truleg òg oral intubasjon. Det er fordelar og ulemper med alle metodane. Oral intubasjon er mindre invasiv enn injeksjon, men krever framleis handtering av fisken. Det er ein fordel at smittedosen kan kvantifiserast nøye og at smittetidspunktet er kjent. Det er òg ein fordel, samanlikna med badsmitte, at mengda parasitt som er naudsynt for å gjennomføre oppsmitting er begrensa. Kohabitasjon vil sannsynlegvis være den naturlegaste smittevegen og, med unnatak for sheddarane, den smittemodellen med minst handtering av fisken som skal smittast. Men, smittetidspunktet for kohabitasjonsfisk vil være ukjent, og variere mellom fiskane. Det forventast òg at kohabitasjon er ein modell som vil krevje lengre forsøk og auka kostnadar. Badsmitte er den minst invasive smittemodellen. Som for oral intubasjon vil det då vera eit definert smittetidspunkt, og ein kan gjera samtidig smitte av fleire artar («common garden» liknande eksperiment). Smittedosen i vatnet kan være godt definert, men den faktiske smittedosen for fisken vil være avhengig av fiskens atferd og miljøforholda. Ein vanske med badsmitte er at det krevjer store mengder parasitt for å oppnå høg smittedose, noko som kan være krevjande å få til.
Ut frå resultata frå dette pilotstudiet og smittemodellane sine fordelar og ulemper konkluderte prosjektgruppa med at badsmittemodellen var den beste for følgande smitteforsøk i SpiroFri.
All fisk vart undersøkt 6 VES for ytre og indre teikn til spironukleose. Det vart ikkje funne teikn til sjukdom knytt til S. salmonicida i nokon av fiskegruppene som var eksponert for parasitten. På grunn av den korte forsøksperioden var heller ikkje sjukdomsutvikling forventa. Verken Kent mfl (1992) eller Mo mfl (1993) fann teikn til sjukdom i laks i sine forsøk (Kent, Ellis et al. 1992, Mo, Poppe et al. 1993). I forsøksperioden på inntil 21 veker fann Guo mfl (2004) S. salmonicida i blod frå ei veke etter smitte i dei tre ulike forsøka. Lesjonar og mortalitet vart derimot ikkje påvist før ca sju veker etter smitte (Guo and Woo 2004). Alfjorden (2018) gjennomførte eit 10 veker langt forsøk utan dødelegheit på fisken, og det var kun mindre teikn på sjukdom i eit fåtall fisk. Desse lesjonane vart sett først fem veker etter smitte og utover (Alfjorden 2018). Sommerhaug (1995) såg lesjonar (byllar) først dag 55 dagar (ca 8 VES) etter anal og oral intubering, men hadde kun få smitta fisk (Sommerhaug 1995).
Påvising av S. salmonicida i vatn vil være svært nyttig både eksperimentelt, i overvaking av parasitten i næringa, samt for miljøundersøkjingar. Det var derfor ynskjeleg å teste ein metode for dette i denne piloten. Dessverre viste innleiande arbeid og resultater at den valgte metoden var til dels særs arbeidskrevjande med det utstyret som var tilgjengeleg, og analysene ga begrensa meirdata. På bakgrunn av dette vart metoden ikkje nytta i det vidare arbeidet. Men, dei innleiande resultata viser at det er mogleg å påvise S. salmonicida i sjøvatn ved filtrering og qPCR. Ein optimalisert protokoll for innsamling, bearbeiding og analyse av vatn bør difor utviklast for påvisning av S. salmonicida i vassprøvar. Slike metodar vil vera til hjelp i fleire ulike settingar, mellom anna som eit godt ikkje-invasivt verktøy for å studere infeksjonsdynamikken til S. salmonicida .
Det var kun forskjell i vekst i forsøksperioden i gruppene som var smitta ved analintubering og kohabitasjon. Det var heller ikkje forskjell i vekt mellom gruppene ved slutten av forsøket. Dette er som forventa ut frå forsøkets varigheit, at klinisk spironukleose utviklar seg over tid i laks, samt at all fisk har fått same behandling. Ved kortvarig (6 veker) vil små forskjellar i vekst som følgje av infeksjon vanskeleg påvisast. Sidan infeksjon med spironukleose utviklar seg over litt tid, og kan gå gradvis over i sjukdom, er det rimeleg at det er liten eller ingen negative effekter på fysiologiske parametre som følgje av infeksjonen dei fyste vekene etter smitte. I tidlegare smitteforsøk med S. salmonicida har det i liten grad vore fokusert på vekst hos fisken i forsøksperioden. Av dei fire smitteforsøka som er publisert (Kent, Ellis et al. 1992, Mo, Poppe et al. 1993, Guo and Woo 2004, Alfjorden, Astvaldsson et al. 2019), er det berre Alfjorden (2018) som rapporterte vektutvikling. Over forsøksperioden på 10 veker fann Alfjorden (2018) at det var svakt betre vekst i den PBS-behandla kontrollgruppa, samanlikna med gruppene injisert eller oral intubert med S. salmonicida .
6 - Konklusjonar
Oral intubasjon, bad og injeksjonssmitte er alle effektive smittemodellar.
Resultata presentert her er i samsvar med tidlegare funn.
Badsmitte syntest å vera den beste modellen for vidare smitteforsøk i SpiroFri.
Utvikling av spironukleose tek lang tid, og undersøkjing av sjukdomsdynamikk krever derfor lang forsøkstid.
7 - Referansar
Adam, R. D. (2017). Diplomonadida. Hogbook of the Protists. J. M. Archibald, A. G. B. Simpson, C. H. Slamovits et al. Cham, Springer International Publishing. DOI: 10.1007/978-3-319-32669-6_1-1.
Alfjorden, A. (2018). Experimental Spironucleus infections in Atlantic salmon (Salmo salar) - Hidden secrets of the life cycle of Spironucleus salmonicida revealed. MSc, Uppsala University. Sverige.
Alfjorden, A., Astvaldsson, A., Jannson, E. og Svärd, S.. Experimental challenge of Atlantic salmon (Salmo salar) with the diplomonad parasite Spironucleus salmonicida to characterize the infection cycle. Samandrag for artikkel i Astvaldsson (2019). Henta frå https://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-378925
Astvaldsson, A. (2019). Pathogenesis and Cell Biology of the Salmon Parasite Spironucleus salmonicida. PhD, Uppsala Universitet.
Grieg Seafood ASA (2023). Annual Report 2023. Noreg.
Fard, M. R. S., A. Jorgensen, E. Sterud, W. Bleiss og S. L. Poynton (2007). "Ultrastructure og molecular diagnosis of Spironucleus salmonis (Diplomonadida) from rainbow trout Oncorhynchus mykiss in Germany." Diseases of Aquatic Organisms 75(1): 37-50.
Guo, F. C. og P. T. Woo (2004). "Experimental infections of Atlantic salmon Salmo salar with Spironucleus barkhanus." Dis Aquat Organ 61(1-2): 59-66.
Johansen, R., R. T. Kongtorp, G. Bornø, H. R. Skjelstad, A. B. Olsen, K. Flesjå, D. Colquhoun, I. Ørpetveit, H. Hansen, Å. H. Garseth og B. Hjeltnes (2009). Fiskehelserapporten 2008. Oslo, Veterinærinstituttet.
Jorgensen, A. og E. Sterud (2006). "The marine pathogenic genotype of Spironucleus barkhanus from farmed salmonids redescribed as Spironucleus salmonicida n. sp." Journal of Eukaryotic Microbiology 53(6): 531-541.
Keister, D. B. (1983). "Axenic culture of Giardia lamblia in TYI-S-33 medium supplemented with bile." Trans R Soc Trop Med Hyg 77(4): 487-488.
Kent, M. L., J. Ellis, J. W. Fournie, S. C. Dawe, J. W. Bagshaw og D. J. Whitaker (1992). "Systemic hexamitid (Protozoa, Diplomonadida) infection in seawater pen-reared chinook salmon Oncorhynchus tshawytscha." Diseases of Aquatic Organisms 14(2): 81-89.
Lie, K.-I., H. Wisløff, L. Østevik, M. Alacorn og S. Nylund (2022). "Spironukleose hos atlantisk laks." Norsk Veterinaer Tidsskrift 134(06-2022): 400-401.
Miller, K. M., I. A. Gardner, R. Vogerstichel, T. Burnley, A. D. Schulze, S. Li, A. Tabata, K. H. Kaukinen, T. J. Ming og N. G. Ginther (2016). Report on the Performance Evaluation of the Fluidigm BioMark Platform for High-Throughput Microbe Monitoring in Salmon: 293.
Mo, T. A., T. Poppe og C. Appleby (1993). "Kohabiteringsforsøk med fire ulike arter laksefisk og en encellet parasitt i slekten "Hexamita"." Norsk Veterinærtidsskrift 105(3).
Mo, T. A., T. T. Poppe og L. Iversen (1990). "Systemic hexamitosis in salt-water reared Atlantic salmon (Salmo salar L.)." Bull. eur. ass. Fish Pathol 10(3).
Paull, G. C. og R. A. Matthews (2001). "Spironucleus vortens, a possible cause of hole-in-the-head disease in cichlids." Diseases of Aquatic Organisms 45(3): 197-202.
Poppe, T. T., T. A. Mo og L. Iversen (1992). "Disseminated hexamitosis in sea-caged atlantic salmon Salmo salar." Diseases of Aquatic Organisms 14(2): 91-97.
Poynton, S. L., M. R. S. Fard, J. Jenkins og H. W. Ferguson (2004). "Ultrastructure of Spironucleus salmonis n. comb. (formerly Octomitus salmonis sensu Moore 1922, Davis 1926, og Hexamita salmonis sensu Ferguson 1979), with a guide to Spironucleus species." Diseases of Aquatic Organisms 60(1): 49-64.
Skjelstad, H. R., G. Bornø, K. Flesjå, H. Hansen, H. Nilsen, M. A. Wasmuth og B. Hjeltnes (2008). Fiskehelserapporten 2007. Oslo, Noreg, Veterinærinstituttet.
Sommerhaug, E. (1995). Kultivering av mikroparasitten Hexamita sp. in vitro og in vivo i atlantisk laks (Salmo salar), og påvisning av antistoffaktivitet mot parasitten i atlantisk laks. MSc, Universitetet i Tromsø.
Sommerset, I., J. Wiik-Nielsen, T. Moldal, V. H. S. Oliveira, J. C. Svendsen, A. Haukaas og E. Brun (2024). Fiskehelserapporten 2023. Oslo, Veterinærinstituttet.
Sommerset, I., J. Wiik-Nielsen, V. H. S. Oliveira, T. Moldal, G. Bornø, A. Haukaas og E. Brun (2023). Fiskehelserapporten 2022. Oslo, Veterinærinstituttet.
Sterud, E., T. Poppe og G. Borno (2003). "Intracellular infection with Spironucleus barkhanus (Diplomonadida : Hexamitidae) in farmed Arctic char Salvelinus alpinus." Diseases of Aquatic Organisms 56(2): 155-161.
Uldal, A. og K. Buchmann (1996). "Parasite host relations: Hexamita salmonis in rainbow trout Oncorhynchus mykiss." Diseases of Aquatic Organisms 25(3): 229-231.