Felles norsk-russisk overvåking av 0-gruppe fisk på høsttokt i Barentshavet, 1965-2023
1 - Introduction
A joint Norwegian-Russian survey of 0-group fish (here defined as fish hatched earlier in the same calendar year) in the Barents Sea was started in September 1965 with the motivation to provide initial information on year class strength of commercially important fish stocks (ICES 1965, Eriksen and Prozorkevich 2011). The survey initially used echosounders to record 0-group fish combined with trawl sampling to identify the composition of the acoustic backscatter (Dragesund and Olsen 1965). The joint 0-group survey was continued the following years with participation also by the United Kingdom from 1966 to 1976. Intercalibration of the echosounders was done before the start of the survey to improve comparability of results obtained by different research vessels (Dragesund 1970, Dragesund et al. 2008).
The acoustic information was used in a semiquantitative manner by classifying the echo-sounder paper recordings into 5 categories from no (0) to very dense (4) recordings (Dragesund et al. 2008). The number of fish caught in supporting trawl catches was additionally used to distinguish between scattered and dense concentrations on distribution maps (Haug and Nakken 1977). While trawling in the first period was guided by the echo-sounder results, ICES advised in 1980 on a standardized trawling procedure (stepwise in the upper 60 m; see later section) which has been followed from 1981 onwards. At the same time, the 0-group survey shifted from a combined acoustic-trawl survey to a standardized trawl survey (Dragesund et al. 2008, Eriksen and Prozorkevich 2011). From 1981 onwards, all vessles have used the same type of trawl, a fine-meshed commercial trawl (‘Harstad’) designed to catch capelin (Nakken and Raknes 1996, Dragesund et al. 2008). This trawl has a rectangular opening of about 20 by 20 m.
The results from the survey have been calculated and expressed as a set of 0-group fish abundance indices of the main commercial species of fish found in the Barents Sea (Dingsør 2005, Eriksen et al. 2009, Eriksen and Prozorkevich 2011). The abundance values have also been converted to 0-group fish biomass by multiplying numbers with mean weight of the 0-group fish that are recorded routinely during the surveys (Eriksen et al. 2011, 2017b).
0-group fish play dual roles in the ecosystem. They are the recruiting life stages of fish stocks that are of great ecological and economic importance, and variation in recruitment, as reflected at the 0-group stage, plays large roles for the dynamics of the fish stocks as well as the wider ecosystem through trophic interactions. In addition, the 0-group fish are planktonivorous and constitute a substantial component among the pelagic fish in the ecosystem. This is the case not only for true pelagic species, such as capelin and herring, but also for demersal species, such as cod and haddock, before they settle to near the seafloor later in autumn.
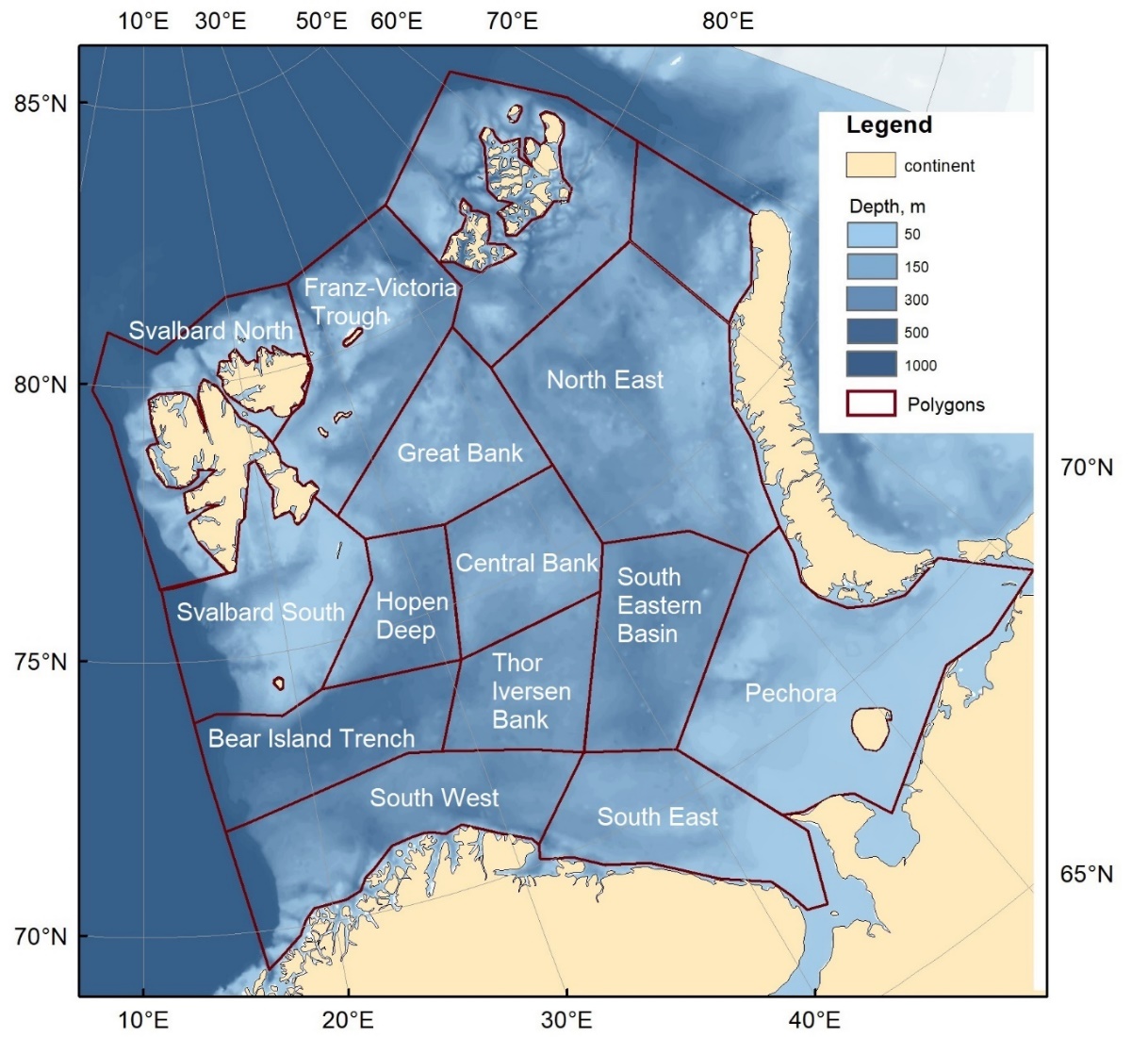
Time series of 0-group abundance and biomass have been used in descriptions and analyses of the Barents Sea ecosystem (e.g. Eriksen et al. 2017, ICES WGIBAR 2018). We are currently expanding these analyses to address in more detail the roles of 0-group fish in relation to recruitment variability and stock dynamics of major fish species, and for the structure and energy flow in food webs of the Barents Sea ecosystem. In the project ‘Trophic Interactions in the Barents Sea: steps towards Integrated Ecosystem Assessment’ (TIBIA) and ICES working group “Integrated ecosystem assessment in the Barents Sea” (WGIBAR), we were using a subdivision of the Barents Sea into 15 subregions (polygons) (Fig. 1). We are using this subdivision (but with 13 polygons only due to lack of coverage in two northeastern polygons) to provide spatially resolved estimates of biomass of major ecosystem components, such as zooplankton, benthos, and fish, including 0-group.
Figure 1. Map showing subdivision of the Barents Sea into 15 WGIBAR-subareas (regions) used to calculate estimates of 0-group abundance based on the Barents Sea autumn surveys (including the Barents Sea ecosystem survey (BESS) since 2004).
In this communication, we provide an updated overview of the joint Norwegian-Russian 0-group investigations in the Barents Sea. We describe the procedures of sampling, analyses, and calculation of results, and discuss associated sources of error. One particular source of error with trawl sampling of small fish is the catchability: to what extent do the 0-group fish escape through the meshes of the trawl as function of fish length, what are the roles of herding, and how is low and variable catchability corrected for (Eriksen et al. 2009). We have used the TIBIA/WGIBAR subdivision to provide spatially resolved estimates of 0-group abundance of major species of fish collected in the 0-group survey. The new abundance estimates by TIBIA polygons are compared with the previous set of abundance indices as reported by Eriksen et al. (2009) and Eriksen and Prozorkevitch (2011). Eriksen and Prozorkevitch (2011) provided distribution maps of four species of fish (capelin, herring, cod and haddock) for each year from 1980 to 2008. Here we provide a new and updated set of distribution maps from 1980 to 2023 for the same 4 species as well as for polar cod and redfish ( Sebastes spp.) (included here in part 7. Spatial distribution). We consider the spatial and temporal coverage of the surveys and note years where incomplete coverage or timing could have influenced the results (Part 6. Survey area and coverage).
2 - Development of the 0-group monitoring
2.1 - From acoustics to trawl-based survey
The international 0-group survey in the Barents Sea shifted from an acoustic survey, where trawling was used to identify the species of 0-group fish in the acoustic layers, to a standardized trawl survey where acoustic records are used mainly to guide sampling (e.g. add extra steps in the vertical if acoustic records suggest that 0-group fish are distributed below 60 m depth) (Dragesund et al. 2008). A study performed in autumn 1963 on abundance and distribution of 0-group fish from acoustic records in the Barents Sea, suggested that it would be feasible to carry out an 0-group survey in autumn based on acoustic methodology (Dragesund and Olsen 1965). At this time, it was known that 0-group fish were abundant in the surface layers of the Barents Sea, stemming from spawning at ‘up-stream’ spawning grounds further south. An echo integrator had also been constructed, which facilitated the treatment of the acoustic recordings (Dragesund et al. 2008). Based on the initial investigation in 1963, and follow-up studies in 1964, it was decided to start a joint international 0-group survey in autumn 1965. The results and experiences from the first four years of the survey (1965-1968) were reported as an ICES publication in 1970 (Dragesund 1970).
The feasibility of an acoustic survey of 0-group fish in the Barents Sea was at the time considered positively, being an early and inspirational case of the general development of fisheries acoustics, where abundance of fish is estimated from acoustic records combined with trawl catches to help identify the acoustic scatterers and allocate the acoustic signals among them (Dragesund et al. 2008). However, it became apparent that use of the acoustic method for 0-group fish was a challenge due to the commonly mixed occurrences of the different species as well as abundant presence of other scatterers such as krill and jellyfishes, as well as 1-group capelin. This led to a shift in emphasis from acoustics to trawling as the basis for the survey.
2.2 - Standardized trawling procedure
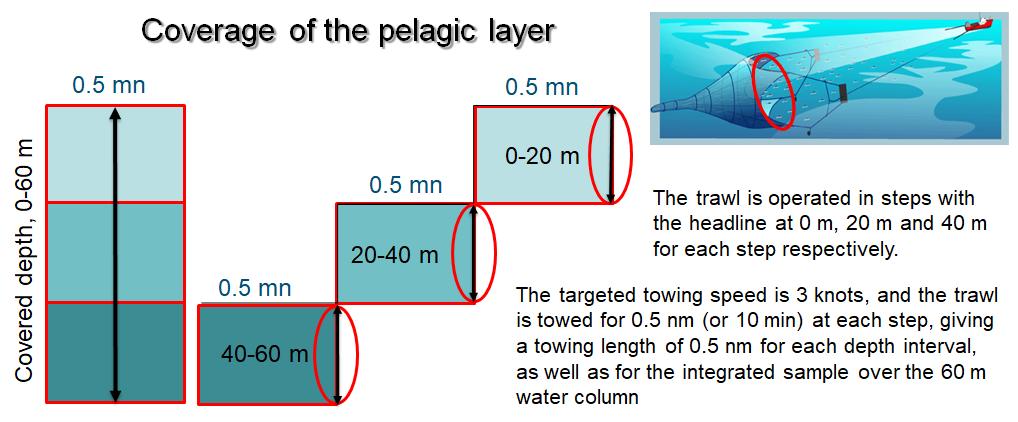
The “Harstad” trawl is designed to capture small fish and has been the standard equipment since around 1980 for the 0-group fish survey, the capelin survey, and later the ecosystem survey (Anon. 1980, Eriksen and Gjøsæter 2013). In the first years of the survey, pelagic trawl hauls were taken frequently, usually no more than 40 nautical miles (nm) apart, targeting acoustic scattering layers to help identify and quantify the contribution by 0-group fish (Dragesund et al. 2008). In addition, some trawl hauls in the surface layer were also taken in areas where there were no clear acoustic records of 0-group fish. Based on advice from ICES, a new trawling procedure was introduced in 1980. This has since been the standard trawling procedure where the trawl is operated in steps with the headline at 0 m, 20 m and 40 m. With a nominal trawl opening of 20x20 m, this provides an integrated sample from the upper 60 m of the water column. The trawling procedure prescribes a towing speed of 3 knots and a tow distance of 0.5 nm for each depth interval (Fig. 2). Additional tows with the headline at 60 and 80 m, and with distance of 0.5 nm, were made when dense concentration was recorded deeper than 60 m on the echo-sounder.
Figure 2. Schematic representation of a pelagic trawling (standard 0-group trawling), indicating three depth steps with headlines at surface (0 m), 20 m and 40 m. With a theoretic trawl opening of 20x20m, this provides an integrated sampling over the upper 60m water column .
Standardization has been an important aspect of the joint 0-group survey in the Barents Sea since its beginning in 1965. The same echo sounders were used on Norwegian and Soviet/Russian vessels in the early years, and inter-ship acoustic calibrations were carried out by comparing results from the same areas (Dragesund et al. 2008). The survey has been a large-scale, multi-ship operation with 3-6 vessels taking part annually. The vessels used in the first years were built as side-trawlers, being gradually replaced between 1970 and 1979 with larger stern-trawlers, better equipped and capable of operating larger trawls (Dragesund et al. 2008). From 1980 all participating vessels have used the same small-meshed sampling ‘standard’ trawl – the ‘Harstad’ trawl. This trawl is constructed with seven panels, with mesh size (un-stretched) decreasing from 100 mm in the first (front) panel to 30 mm in the last panel and 8 mm in a codend (Godø et al. 1993). While the trawl is considered standard and has been used on both Norwegian and Russian vessels, there have been adaptations and differences in rigging due to the Norwegian vessels initiated towing at the surface and the Russian vessels initiated towing at depth.
2.3 - Sample processing and analyses
When the trawl comes on deck, the trawl is shaken well, to allow for fish adhering to the trawl meshes to fall back into the trawl cod end or to the deck. This is to ensure that the calculated biomass and numbers of individuals are as accurate as possible, and to avoid fish from earlier hauls contaminating later samples. The problem of fish being trapped between trawl meshes is greatest at stations with a lot of 0-group capelin. The part of the catch that falls to the deck, usually in poor condition, is collected and processed separately. The sample from the deck is identified to species and weighed per species. The weight of the deck sample is added to the rest of the sample on a species basis to give the total sample weight for each species.
Catch processing in the fish laboratory starts with all jellyfish and larger fish (such as lumpfish) being sorted out to make the rest easier to handle. Jellyfish and larger fish are weighed separately. Sometimes it is necessary to remove excess of water so that the sample weight is affected as little as possible by the water. In the case of large catches, a sub-sample is taken. When sub-sampling, a conversion factor is used to calculate the total weight of all groups in the catch. A factor is calculated as the total weight divided by the sub-sample weight. The samples from the trawl are processed immediately after the catch is removed from the trawl. 0-group fish of different species, as well as other components of the catch (e.g. krill and pelagically distributed small non-commercial fishes), are sorted into groups that are weighed separately. The total weight of the catch is determined as the sum weight of the components. The extra variance introduced by subsampling has not been studied formally but is believed to be low compared to the high variance associated with the trawl samples of 0-group fish.
The 0-group fish are determined to the species level, while some of the small non-commercial species (families Agonidae, Stichaeidae, Cottidae and Myctophidae) could be determined to genus or family level (due to taxonomic difficulties, available expertise, and time constraint). Before 2014, 100 individuals of each species/group (if available) were weighed and separately length measured (to nearest mm on Norwegian and 0.5 mm on Russian vessels). The length sample weight and total catch weight are used to calculate the total number of fish caught. From 2014, the number of fish that were length measured was reduced to 30 individuals (based on statistical considerations described in Pennington and Helle, 2013).
3 - Calculation of abundance indices and quality control of databases
Various ways of calculating abundance indices have been used during the history of the survey. In the early years of the survey, from 1965, the echo abundance was subjectively evaluated from the paper recordings (echograms) on a scale from 0 to 4 (0 - no recording, 1 - very scattered, 2 - scattered, 3 - dense, 4 - very dense). This information was then used during the first 6 years (1965-1970) to classify year-class strength as poor, average, or strong by expert judgement (Dragesund et al. 2008).
3.1 - Area index
The acoustic information was subsequently used to construct a quantitative (or semi-quantitative) abundance index, the so-called area index (Haug and Nakken 1977). Maps of distribution of various 0-group species had been prepared for the annual reports based on the 0 - 4 scale visual grading of paper echograms, guided by results on the 0-group fish counts in the supporting trawl hauls. Classification of the acoustic records was done for every nautical mile sailed along survey lines, with three density grades used to plot the results onto maps: absent, scattered, and dense (Dragesund et al. 2008). Haug and Nakken (1977) established empirical relationships between trawl catches and the 4 density grades (very scattered, scattered, dense, very dense). They noted some inconsistencies in the grading between vessels and years, and established criteria in terms of number of fishes per haul to help standardize the distinction between scattered and dense records of 0-group fish of four species (cod, capelin, redfish, and polar cod).
Haug and Nakken (1977) used the criteria to draw new distribution maps for the four species of 0-group fish for the years 1965-1972. The area index was calculated as the sum of the integrated area on the map with low abundance (scattered), plus the area with high abundance (dense) multiplied by factor 10. This factor was an approximation based on the empirical data (Haug and Nakken 1977). The area index was calculated for six species (cod, capelin, haddock, redfish, polar cod, and long rough dab) for the years 1965-1972. Average index values were used to reclassify year-class strength in each year in this (relatively short) period as average, poor, or strong (Haug and Nakken 1977).
The area index was calculated in subsequent years as one of two methods (the other was the logarithmic index; see below) used to produce time series from the 0-group survey (Dragesund et al. 2008). It became apparent that the area index had shown an increasing trend from 1965 until the early 1990s. Nakken and Raknes (1996) provided a correction to the area index time series by assuming that capture efficiency had increased proportional to the size of the trawls (trawl opening (“mouth”) area) used in the survey. They used the arithmetic mean trawl opening for the survey participating vessels (and trawls) each year, which they considered a rough approximation since differences in geography and catches among the vessels were not taken into account (which would have required much work). The correction represented more than a doubling of the area index values between 1970 and 1984 (Nakken and Raknes 1996). Nakken and Raknes (1996) also attempted an alternative method for correction, using the trend in the sum of index values for cod, haddock and redfish as an expression for the trend in overall capture efficiency. However, this depended strongly on an increasing trend for redfish, and it was uncertain how much of this increase was due to increased capture efficiency.
The corrected area index time series was updated annually and reported in the annual report from the 0-group survey to ICES. Nakken and Raknes (1996) provided corrections for cod, haddock, and redfish. Subsequently, similar corrections were made for Greenland halibut, long rough dab, and polar cod. The area index for herring was calculated by Toresen (1985) for the period 1965-1984. Dragesund et al. (2008) provided a graphical representation (in their Fig. 6.6, page 127) of the area indices for the 1965-2000 period for 7 species of 0-group fish (cod, haddock, herring, redfish, capelin, and polar cod,) (based on ICES 2003).
3.2 - Logarithmic index
The logarithmic index was developed by Randa (1984). The catch in numbers of 0-group fish at each station was log-transformed (natural logarithm, ln), and mean densities (catch rates per nautical mile) were calculated for 17 strata (geographical areas) of the 0-group survey area in the Barents Sea. The overall abundance index for a species was then calculated as the area-weighted mean logarithmic abundance, adjusted for the proportion of hauls with no catch. The method is based on the log-normal theory, and it allows confidence intervals to be calculated based on normal theory (Randa 1984). Randa (1982) showed that log-transformation normalized the catch data for 0-group cod (for the 1965-1979 period).
Randa (1984) took into account the different trawls used in the early years of the joint 0-group survey by estimating ‘relative fishing power’ (relative to R/V “G.O. Sars”, 1971-1979) for each of the participating vessels.
The logarithmic index was calculated by Randa (1984) for cod and haddock, and by Toresen (1985) for herring. These indices were updated and included in the annual reports to ICES from the joint 0-group survey.
The logarithmic index was further developed as one of two alternative indices by Dingsør (2005; the other was an arithmetic index based on stratified sample mean; see below), which he called the ‘Pennington estimator’ (Pennington 1996). While the 0-group data largely follow a log-normal distribution, they usually have many low values close to zero which may bias log-normal-based estimators. A cut level for low values (set at 20 % of the average abundance density in each stratum) was used to reduce the bias from low values and achieve better fit to log-normal distribution for the remaining values above the cut level (Folmer and Pennington 2000). Dingsør (2005) calculated time series of the logarithmic ‘Pennington estimator’ (with standard errors) for cod, haddock, capelin, redfish, and herring for the years 1980-2002. The index was calculated both with and without correction for capture efficiency (see section ‘Capture efficiency’) for cod and haddock. Dingsør (2005) compared the ‘Pennington estimator’ index with the old area index and the previous logarithmic index. He found similar trends but also some discrepancies, notably for some of the species in the 1980s (see Figs 4 and 5 in Dingsør 2005).
Dingsør (2005) recommended using the ‘Pennington estimator’ as the most appropriate method and new standard for presenting 0-group abundance indices in the Barents Sea. However, the arithmetic abundance index based on the ‘stratified sample mean’ method turned out to be the preferred index for routine use. With the start of the joint ecosystem survey (where the 0-group survey became an integral part) in 2004, the arithmetic (total abundance) index was used, and the logarithmic index was no longer calculated after 2004.
3.3 - Total abundance indices
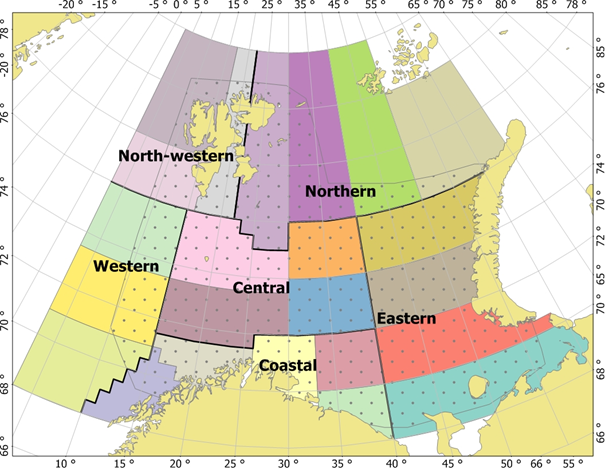
At the transition to the joint ecosystem survey in 2004, a new abundance index was developed by Gjert E. Dingsør and Dmitry Prozorkevich and used for the 0-group results from the survey in 2004 (Anon. 2005, Dingsør 2005). The index was based on a stratified sample mean estimator, reflecting the mean areal density of 0-group fish in the survey area. The density of fish in length groups (number of fish per nm2 ) was calculated for each trawl station, and mean density was calculated for each of 23 strata of the total survey area of the Barents Sea (Fig. 3; Dingsør (2005) used a division into only 4 larger strata). The stratified sample mean estimator of abundance was then calculated as the overall mean density of 0-group fish, by weighting the strata means by the proportion of the survey area in each stratum. The area covered with survey stations within each stratum was determined using GIS software.
The 23 0-group strata were combined into larger areas (north-western, northern, western, central, eastern and coastal; Fig. 3) used in Eriksen at al. 2009, 2011, 2012, and 2014. Later, in the project TIBIA, the Barents Sea was divided into 15 subareas (polygons, see Fig. 1). The division is based on topography and oceanography and is a modification (with some subdivision) of the system used by Eriksen et al. (2017) in a summary analysis of distribution of pelagic biomasses in the Barents Sea. At the ICES WGIBAR meeting in 2018 (ICES WGIBAR 2018), the division of the Barents Sea into 15 polygons was presented and adopted for use in reporting status and changes in the ecosystem.
Figure 3. The Barents Sea 0-group strata system, consisting of 23 strata shown in different colours. 0-group survey coverage area is shown by dots at 0-group strata system. The 23 strata have been used for compilation of data in Barents Sea fish stock assessments.
The stratified sample mean estimator was expressed as a total abundance index by using the total area covered in the survey (sum of polygon mean density of fish, per nm2, multiplied by polygons coverage in nm2 ). The total abundance index was calculated both without and with length correction for low capture efficiency for small fish (see section ‘Capture efficiency’). These two sets of indices (corrected and non-corrected) were calculated back to 1980 for capelin, cod, haddock, herring, saithe, and polar cod, as were uncorrected values for redfish, Greenland halibut, and long rough dab (Tables 2.2 and 2.3 in Anon. 2005). The new total abundance index is calculated with variance and confidence intervals based on the variation in 0-group abundance among sampling stations. At the time it was agreed that the new total abundance index without correction would be the ‘official’ one, while the corrected index was ‘additional’.
Dingsør (2005) showed that the stratified sample mean estimator corresponded closely to the log-normal based ‘Pennington estimator’, with both showing similar temporal patterns from 1980 to 2002 (for cod, haddock, capelin, herring and redfish; see his Table 2).
The total abundance index was used for 0-group data for the next years of the ecosystem survey with some adjustments of the time series (in 2005 and 2007). The former logarithmic index was discontinued in 2005, while the old area index former reported in parallel to the new set of indices (total abundance, corrected and uncorrected) until (and including) 2007 when it also was discontinued.
An ‘overhaul’ of the total abundance index was done in 2009. It had become apparent that there were many mistakes and errors in the data (e.g., punching errors when data were transferred from paper sampling sheets to the computer), and inconsistencies between the data held in data bases of the two institutions conducting the surveys (IMR and PINRO). A major effort was therefore made over a three-year period to check the quality of the data, using cruise logbooks and original data records dating back to 1980.
New sets of total abundance indices based on the quality assured data were calculated and reported by Eriksen et al. (2009). This work included indices corrected for capture efficiency and uncorrected indices for cod, haddock, capelin, herring, saithe, and polar cod, and uncorrected indices for redfish, Greenland halibut, and long rough dab, from 1980 onwards (see Table 2 in Eriksen et al. 2009). The corrections were from slight to substantial in some cases (species and years). However, the broad temporal patterns and trends in 0-group year-class strength did not change much, reflecting that the amplitude of changes in abundance was generally much larger than the corrections (Eriksen et al. 2009). The estimation was carried out in SAS software and the indices of fish abundance for the 0-group are presented in part 9.1.
Eriksen et al. (2009) showed that the revised set of total abundance indices were positively correlated with the old area index for cod, haddock, capelin, and herring (r = 0.80-0.89). The abundance indices were also positively correlated with estimated abundances of the year classes as 1-group for capelin (r = 0.81-0.82), and 3-year old for haddock (r = 0.43-0.49).
Abundance and biomass estimates were calculated by different software during the last four decades: SAS (for the new 23 fisheries subareas, 1980-2017, 0-group strata and WGIBAR polygons ) and MatLab (for the new 15 WGIBAR- polygons ( for the period between 1980 and 2018, ICES WGIBAR 2018) and R (for the new 15 WGIBAR-subareas (2003-202 3 ). Due to software upgrading (which led to challenges with script running in SAS) and personal resource limitation (MatLab), it was decided to develop R-scripts (R core Team, 2023) for estimation of abundance and biomass indices. Two data sets (abundance and biomass indices calculated by R and SAS) were analyzed for similarities and were found to be highly significantly correlated (for capelin r=0.95, cod r=0.99, haddock r=0.94, herring r=0.98 and polar cod r=0.94).
During development of R scripts for abundance and biomass estimation, some errors in the IMR database were detected, that most likely occurred when all historical data were converted from an old to the new "Biotic" format. Apparently, some algorithms failed, which created duplicate rows of existing fish observations and recalculated total weight or abundance. A new quality check was carried out on the data in the new data format, which was corrected back to 2004. The older data (1980-2003) in the IMR database. have not been checked and corrected, and it is uncertain how many errors there are in this part. We note that the data compiled and used in this report were extracted from the database at an earlier stage and are not affected by these errors.
The last “official” updated time series of the abundance and biomass of the 0-group fish are reported in the BESS report 2023 (available at https://www.hi.no/hi/nettrapporter/imr-pinro-en-2024-2) and in Part 9.4 of this report.
4 - Capture efficiency
Small juvenile fish, especially herring, pass through the meshes of the first panels of the Harstad trawl. This gives a low capture efficiency of the trawl when catch is referenced to the mouth opening of the trawl (Godø et al. 1993). The effect is inevitable due to the low maximum swimming speed of small 0-group fish relative to the mesh size and speed of the trawl. This was clearly demonstrated in experiments in the early 1990s, comparing catches of 0-group fish in the standard trawl with catches obtained with a specially designed 0-group trawl with finer meshes (Godø et al. 1993, Hylen et al. 1995).
The experimental trawl was smaller with mouth opening of 30 m2 (compared to 300 m2 for the standard trawl for a specific configuration of 20 m x 15 m), and mesh size decreased from 200 mm in the front panel to 10 mm in the cod end (Godø et al. 1993, Valdemarsen and Misund 1995). Experiments comparing the standard trawl and the experimental trawl were done in the Barents Sea in August 1991 (Godø et al. 1993), and during the 0-group survey in August/September 1992 and 1993 (Hylen et al. 1995). Both studies gave consistent results, with sampling efficiency (comparing density of 0-group fish in numbers per nm2 ) around 3-4 times higher for the experimental trawl compared to the standard trawl for 0-group cod and haddock. Furthermore, there was a clear size selection, where juveniles smaller than 5 cm were captured to very low extent with the standard trawl (Godø et al. 1993). The capture efficiency was strongly size-dependent, increasing from around 10 % for 5 cm long juveniles to nearly 100 % for 10-cm long fish for the standard trawl relative to the experimental trawl (Hylen et al. 1995). For even larger juveniles (>10 cm), there were evidence that they were more effectively captured with the standard trawl, suggesting that they were either herded into the larger trawl or having some avoidance of the smaller experimental trawl (Godø et al. 1993, Hylen et al. 1995).
In addition to a size effect, Hylen et al. (1995) found indication of a considerable effect of density of 0-group fish on capture efficiency. Using acoustic recordings as reference, they found a clear and significant positive effect of fish density (as reflected in trawl catches) on capture efficiency (trawl catch relative to acoustically recorded density). Hylen et al. (1995) explained this relationship by density-dependent herding, with increasing degree of herding (either in front of or inside the trawl) with increasing density of fish.
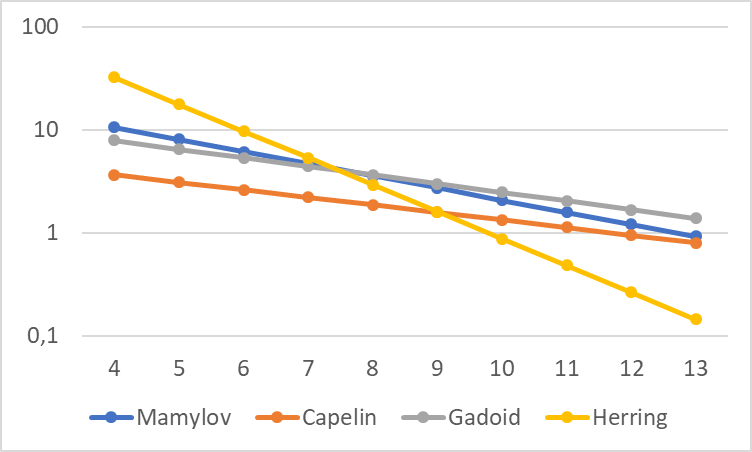
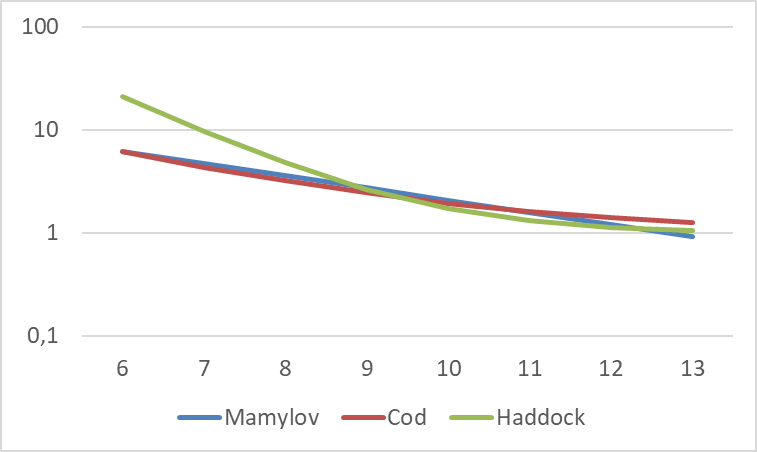
Mamylov (1999) developed a theoretic model of capture efficiency by trawl. The model assumed that the lowest capture efficiency for small fish (4.5 cm and 12.5cm) was equal to the ratio between the cross-sectional area of the cod-end and the mouth opening of the trawl, which he set at 0.1, corresponding to a maximum correction factor of 10. He assumed the capture efficiency of large 0-group fish was equal to 1, i.e. all fish that passed the mouth opening were collected in the cod-end. The equation is Keff = 31.177*exp (-0.2708*L) and illustrated graphically in Figure 4.
Later, PINRO carried out several investigations, and of 1205 analysed trawl catches, 131 trawl catches were selected in which mainly one species was present (Mamylov 2004, Prozorkevich 2010, 2012). The trawl catches in terms of numbers and size of 0-group fish were converted (using target strength relationship) and expressed in units of acoustic backscattering. The acoustic data were scrutinized, and selected portions of the data were regressed against the trawl data expressed in the same units. The equations give very high factors for fish smaller than 4 cm (because of linear extrapolation), and therefore the maximum Keff (gadoids=8, herring =30 and capelin =4) was used for these small fish. The results from these experiments were close to the theoretical model, but they varied between species.
The correction curve for herring is very different, being much steeper than the lines for gadoids and capelin shown in Figure 4. For juvenile herring <6 cm long, the correction factor is higher than 10 (30 at 4 cm length), which is a theoretical maximum. For juvenile herring >10 cm, the correction factor is <1, corresponding to capture efficiency >1 (>100 %). This would imply active herding by doors and bridles in front of the trawl. While this cannot be ruled out, the very low capture efficiency in the low end, and the high capture efficiency in the high end, suggest that the steepness of the herring curve may be an artefact.
Figure 4. Correction factors (Keff) for capture efficiency as a function of length (L in cm) of 0-group fish on log scale. The equation from theoretic model is Keff = 31.177*exp(-0.2708*L) (Mamylov 1999). The equations for capelin (Keff = 7.2075*exp(-0.1688*L), gadoids (Keff = 17.065*exp(-0.1932*L), and herring (Keff = 357.23*exp(-0.6007*L) are from Prozorkevich (2012).
Hylen et al. (1995) provided similar empirical relationships for capture efficiency and correction factors for cod and haddock, using the experimental trawl as a reference for the catches obtained with the standard trawl. The relations from Hylen et al. (1995) have been plotted in Figure 5 using equations (2 and 3) from Dingsør (2005). The lines for cod and haddock are curvilinear on this log-scale plot because the equation is of a different form (declines exponentially to 1 rather than to zero). Apart from this, the line for cod from Hylen et al. (1995) is very similar to the Mamylov line. The haddock line is also close to the Mamylov line for fish in the size range from 8.5 to 13 cm. The haddock line swings upwards at low fish length, to values over 10 for fish <7 cm; again, this is possibly an artefact due to large variation in the underlying data (see Hylen et al. 1995, their tables 3 and 6).
Figure 5. Correction factors (Keff) for capture efficiency as a function of length (L - cm) of 0-group cod and haddock from Hylen et al. (1995) with equations from Dingsør (2005): cod - Keff = 1 + exp(4.158-0.422*L), haddock - Keff = 1 + exp(8.031-0.838*L). The relation from Mamylov (2004) is the same as in Figure 4.
The correction factors for gadoids, capelin and herring in Fig. 4 were used to correct the total abundance indices from the 0-group survey by Dingsør (2005) and Eriksen et al. (2009). Corrections were done for cod, haddock, saithe, and polar cod using the equation for gadoids, and for capelin and herring with their respective equations. The time series of abundance of 0-group fish of redfish, Greenland halibut, and long rough dab were not corrected, and uncorrected indices were used by Eriksen et al. (2009). The corrected abundance time series were used by Eriksen et al. (2011, 2017) where abundance was converted to biomass of 0-group fish.
In 2013-2016, several experiments were performed to study escapement of 0-group fish through the trawl panels and clogging of 0-group fish (BESS reports for 2013-2016, available at https://www.hi.no/hi/nettrapporter?y=2024&query=&serie=imr-pinro&fast_serie= ) with the aim to develop a new 0-group fish trawl. The trawl is designed to obtain constant trawl geometry independent of warp length and to obtain reduced clogging and escapement compared to the standard Harstad trawl. Unfortunately, the newly developed 0-group trawl with fine inner nets and constant opening was too heavy to be towed by the old Russian vessel. It was therefore decided that, for the time being, the Harstad trawl would be used as the standard trawl on all vessels participating in the BESS.
5 - Vertical distribution
The timing and general design of the 0-group fish survey is to allow sampling of the 0-group part of populations of the different species while they still are in the upper pelagic zone. The early studies that used acoustic recordings, showed that the 0-group fishes were generally distributed in the upper 60 m water layer in early autumn, where they are feeding on zooplankton. This observation was the basis for the standard trawling procedure with three steps covering the 0-60 m depth interval (Fig. 2). The procedure is also to include one or two additional deeper steps (to 80 or 100 m) if the acoustic records show deeper distribution of 0-group fish.
There is little information in the literature about when cod change from pelagic life-stage to a demersal life-stage in the Barents Sea. Several studies from other areas have shown that there is no clear relationship between fish age (in days) and fish length (in mm), and that fish of similar length settle at different times (Hussy et al. 2003; Anon. 2009). Boitsov et al. (1996) found that the transition (settlement) is a rather long process that occurs in September-October in the Spitsbergen area and in October-November in the southern Barents Sea. The settlement of cod and their food items occurs gradually and it is likely to be connected with a convection mixing of water layers and deepening of the thermocline layer (Ozhigin et al. 1999). It is assumed that haddock follows a similar pattern to cod, with the transition occurring gradually during the autumn (Dingsør 2005; Anon. 2006, 2009).
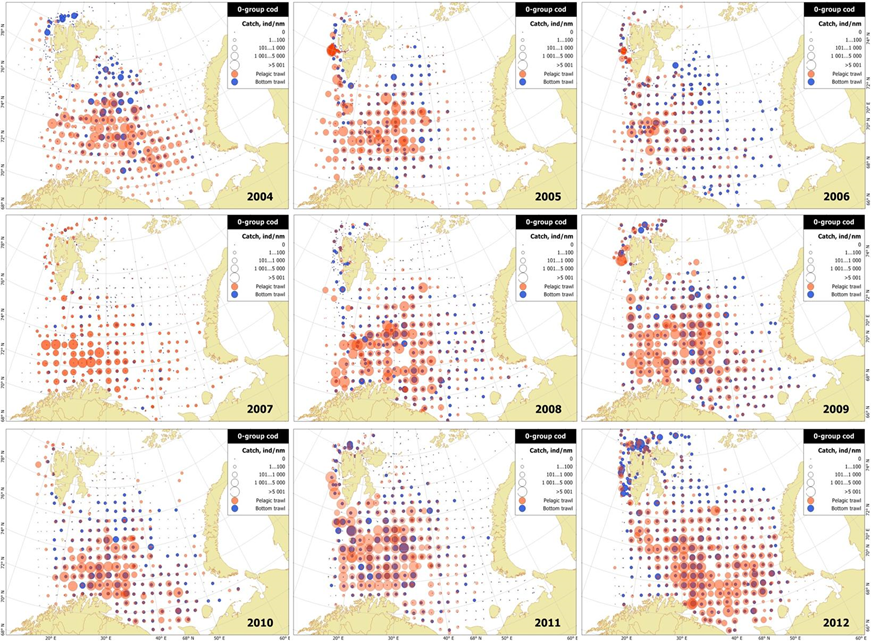
When the 0-group survey became a part of the ecosystem survey (in 2004), bottom trawl samples were also taken. Some 0-group cod were collected by the bottom trawl indicating most likely cod settlement, although ‘contamination’ by 0-group cod from the water column when the bottom trawl was retrieved may also have contributed to the catch. The data indicated varied settlement pattern between years and areas. Prozorkevich and Eriksen (2013) examined 0-group cod distribution based on pelagic and bottom trawl for the years 2005-2012 (Figure 6). They found that numbers of cod taken by demersal trawl were generally low, varying between 0.2 and 1.1%, suggesting that the settled part of the 0-group of cod population is too small to influence 0-group abundance indices markedly. The study suggested that there was no strong relationship between fish settlement and year class strength. However, during some of the most recent years, 0-group fish, notably cod, haddock and capelin, were found to be abundant in the 100-150 m depth layer possibly reflecting early descent from the upper pelagic layer.
Figure 6. 0-group catches of cod taken by pelagic (red) and bottom (blue) trawl in 2005-2012. Higher demersal records than pelagic records at ecosystem survey stations may indicate cod settlement (Figure 4 in Prozorkevich and Eriksen 2013).
6 - Survey area and coverage
0-group fish of the different commercial species, taken together, occupy much of the area of the Barents Sea. Capelin and cod are most widely distributed, haddock and redfish are distributed mainly in the western and central areas, herring in the southern, central and western areas, while polar cod is distributed in the eastern and northern Barents Sea (see maps in Part 7).
The survey area has included the western, southern, and central Barents Sea during the whole survey period. The survey has been operated with 4-6 research vessels each year (Table 1). The vessels have covered different parts of the surveyed area, and cruise lines with sampling stations have been planned so that sampling effort is spread out more or less evenly over the survey area. One reason for this is the aim to monitor distribution and abundance of 0-group fish of several species that have different distribution patterns. The 0-group investigations have also been integrated with other survey elements, into what was called multi-species surveys from the late 1980s, and ecosystem survey from 2004 (Eriksen et al. 2018). Due to the many different purposes of the cruises, a stratified sampling design with higher effort in core areas of 0-group distribution and lower effort elsewhere, has not been used. The distance between trawl stations was about 30 miles until 1994 and 35 miles thereafter (Eriksen et al. 2018).
Year
Vessel name
Start of the survey
End of the survey
1965
Akademik Knipovich
03.09
17.09
1965
Jastreb
03.09
17.09
1965
Johan Hjort
03.09
17.09
1965
G.O. Sars
03.09
17.09
1966
Akademik Knipovich
27.08
10.09
1966
Fridtjof Nansen
27.08
10.09
1966
Johan Hjort
27.08
10.09
1966
G.O. Sars
27.08
10.09
1966
Ernest Holt
27.08
10.09
1967
Akademik Knipovich
24.08
09.09
1967
Fridtjof Nansen
24.08
09.09
1967
Johan Hjort
24.08
09.09
1967
G.O. Sars
24.08
09.09
1967
Ernest Holt
24.08
09.09
1968
Akademik Knipovich
25.08
09.09
1968
Fridtjof Nansen
25.08
09.09
1968
Johan Hjort
25.08
09.09
1968
G.O. Sars
25.08
09.09
1968
Ernest Holt
25.08
09.09
1969
Akademik Knipovich
24.08
07.09
1969
Fridtjof Nansen
24.08
07.09
1969
Johan Hjort
24.08
07.09
1969
G.O. Sars
24.08
07.09
1969
Ernest Holt
24.08
07.09
1970
Akademik Knipovich
23.08
11.09
1970
Fridtjof Nansen
23.08
11.09
1970
Johan Hjort
23.08
11.09
1970
G.O. Sars
23.08
11.09
1971
Akademik Knipovich
20.08
11.09
1971
Fridtjof Nansen
20.08
11.09
1971
G.O. Sars
20.08
11.09
1971
Johan Hjort
20.08
11.09
1971
Cirolana
20.08
11.09
1972
Akademik Knipovich
26.08
10.09
1972
Fridtjof Nansen
26.08
10.09
1972
Poisk
26.08
10.09
1972
Johan Hjort
26.08
10.09
1972
G.O. Sars
26.08
10.09
1973
Fridtjof Nansen
26.08
12.09
1973
Poisk
26.08
12.09
1973
Johan Hjort
26.08
12.09
1973
G.O. Sars
26.08
12.09
1973
Cirolana
26.08
12.09
1974
Akademik Knipovich
27.08
12.09
1974
Poisk
27.08
12.09
1974
G.O. Sars
27.08
12.09
1974
Havdrøn
27.08
12.09
1974
Cirolana
27.08
12.09
1975
Fridtjof Nansen
25.08
07.09
1975
Poisk
25.08
07.09
1975
Johan Hjort
25.08
07.09
1975
G.O. Sars
25.08
07.09
1975
Cirolana
25.08
07.09
1976
Odissey
25.08
07.09
1976
Fridtjof Nansen
25.08
07.09
1976
Johan Hjort
25.08
07.09
1976
G.O. Sars
25.08
07.09
1976
Cirolana
25.08
07.09
1977
G.O. Sars
22.08
11.09
1977
Johan Hjort
20.08
11.09
1977
Odissey
31.08
11.09
1977
Fridtjof Nansen
26.08
11.09
1977
Poisk
25.08
11.09
1978
G.O. Sars
25.08
10.09
1978
Johan Hjort
20.08
10.09
1978
Poisk
25.08
10.09
1978
Fridtjof Nansen
25.08
08.09
1979
Johan Hjort
26.08
14.09
1979
G.O. Sars
19.08
14.09
1979
Poisk
29.08
14.09
1979
Akhill
01.09
03.09
1980
Johan Hjort
16.08
07.09
1980
G.O. Sars
16.08
07.09
1980
Michael Sars
16.08
08.09
1980
Poisk
22.08
08.09
1981
Johan Hjort
21.08
05.09
1981
G.O. Sars
14.08
04.09
1981
Michael Sars
12.08
04.09
1981
Persey III
22.08
06.09
1981
Akhill
23.08
01.09
1982
Johan Hjort
18.08
05.09
1982
G.O. Sars
18.08
05.09
1982
Michael Sars
21.08
11.09
1982
Persey III
31.08
05.09
1982
Poisk
23.08
05.09
1982
Protsion
28.08
30.08
1982
Protsion
11.09
14.09
1983
Eldjarn
21.08
08.09
1983
G.O. Sars
21.08
05.09
1983
Michael Sars
21.08
05.09
1983
Persey III
22.08
05.09
1983
Poisk
24.08
03.09
1983
Alaid
20.08
26.08
1984
Eldjarn
12.08
05.09
1984
G.O. Sars
19.08
03.09
1984
Håkon Mosby
19.08
05.09
1984
Persey III
20.08
30.08
1984
Poisk
26.08
29.08
1984
Alaid
20.08
27.08
1984
Kokshaysk
27.08
02.09
1985
Eldjarn
19.08
04.09
1985
G.O. Sars
19.08
03.09
1985
Håkon Mosby
20.08
02.09
1985
Michael Sars
17.08
19.08
1985
Kokshaysk
23.08
02.09
1985
Vilnyus
25.08
01.09
1986
Eldjarn
20.08
04.09
1986
G.O. Sars
11.08
04.09
1986
Håkon Mosby
20.08
03.09
1986
Kokshaysk
21.08
01.09
1986
Vilnyus
20.08
02.09
1987
Eldjarn
17.08
03.09
1987
G.O. Sars
17.08
03.09
1987
Håkon Mosby
20.08
03.09
1987
Artemida
18.08
28.08
1987
Vilnyus
20.08
01.09
1988
Eldjarn
22.08
06.09
1988
G.O. Sars
22.08
07.09
1988
Håkon Mosby
20.08
03.09
1988
Artemida
21.08
02.09
1988
Professor Marty
26.08
04.09
1989
Eldjarn
22.08
11.09
1989
G.O. Sars
21.08
11.09
1989
Michael Sars
22.08
11.09
1989
Professor Marty
20.08
08.09
1989
PINRO
20.08
09.09
1990
Eldjarn
21.08
05.09
1990
G.O. Sars
21.08
05.09
1990
Michael Sars
16.08
05.09
1990
Professor Marty
16.08
04.09
1990
PINRO
20.08
04.09
1991
Johan Hjort
08.08
09.09
1991
G.O. Sars
19.08
09.09
1991
Michael Sars
15.08
09.09
1991
Professor Marty
15.08
06.09
1991
Fridtjof Nansen
18.08
06.09
1992
Johan Hjort
17.08
03.09
1992
G.O. Sars
18.08
07.09
1992
Michael Sars
13.08
07.09
1992
Professor Marty
17.08
28.08
1992
Fridtjof Nansen
24.08
05.09
1992
Akhill
13.08
15.08
1992
Akhill
05.09
06.09
1993
Johan Hjort
16.08
08.09
1993
G.O. Sars
17.08
07.09
1993
Professor Marty
22.08
08.09
1993
PINRO
23.08
06.09
1994
Michael Sars
16.08
20.08
1994
Johan Hjort
17.08
06.09
1994
G.O. Sars
20.08
07.09
1994
Professor Marty
02.09
08.09
1994
Atlantida
24.08
08.09
1994
Fridtjof Nansen
27.08
08.09
1995
Michael Sars
22.08
09.09
1995
Johan Hjort
25.08
10.09
1995
G.O. Sars
16.08
10.09
1995
Professor Marty
05.09
11.09
1995
Fridtjof Nansen
26.08
11.09
1996
Michael Sars
22.08
10.09
1996
Johan Hjort
24.08
10.09
1996
G.O. Sars
17.08
10.09
1996
Atlantida
15.08
10.09
1996
Persey III
24.08
10.09
1997
Johan Hjort
20.08
08.09
1997
G.O. Sars
19.08
08.09
1997
Atlantida
21.08
06.09
1997
Persey III
15.08
06.09
1998
Fridtjof Nansen
19.08
05.09
1998
Atlantida
08.08
03.09
1998
G.O. Sars
26.08
07.09
1998
Johan Hjort
25.08
08.09
1998
M. Sars
25.08
04.09
1999
Atlantniro
15.08
02.09
1999
G.O. Sars
27.08
06.09
1999
Johan Hjort
22.08
07.09
1999
Persey 4
22.08
03.09
2000
Atlantniro
22.08
01.09
2000
Fridtjof Nansen
19.08
03.09
2000
G.O. Sars
20.08
03.09
2000
Johan Hjort
18.08
07.09
2001
G.O. Sars
16.06
08.09
2001
Johan Hjort
20.08
08.09
2001
Atlantniro
10.08
03.09
2001
Fridtjof Nansen
12.08
03.09
2002
G.O. Sars
16.06
08.09
2002
Johan Hjort
24.08
08.09
2002
Atlantniro
10.08
08.09
2002
Fridtjof Nansen
29.08
08.09
2003
Johan Hjort
05.08
02.10
2003
G.O. Sars
27.07
01.09
2003
Jan Mayen
01.09
16.09
2003
Tsivilsk
07.09
02.10
2003
Smolensk
25.08
02.10
2004
Jan Mayen
04.08
01.10
2004
Johan Hjort
01.08
04.10
2004
Smolensk
06.08
02.10
2004
Fridtjof Nansen
07.08
02.10
2005
G.O. Sars
06.08
30.09
2005
Johan Hjort
01.08
08.09
2005
Jan Mayen
04.08
04.09
2005
Smolensk
09.08
26.09
2005
Fridtjof Nansen
17.08
26.09
2006
G.O. Sars
18.08
28.09
2006
Johan Hjort
14.08
20.09
2006
Jan Mayen
08.08
17.08
2006
Jan Mayen
11.09
29.09
2006
Smolensk
16.08
29.09
2006
Fridtjof Nansen
11.08
05.10
2007
G.O. Sars
14.08
30.09
2007
Johan Hjort
01.08
31.08
2007
Johan Hjort
14.09
26.09
2007
Jan Mayen
10.09
27.09
2007
Smolensk
07.08
28.09
2007
Vilnyus
06.08
23.09
2008
G.O. Sars
19.08
30.09
2008
Johan Hjort
01.09
16.09
2008
Jan Mayen
08.09
24.09
2008
Vilnus
08.08
26.09
2008
Atlantic star
01.08
10.08
2009
G.O. Sars
20.08
05.09
2009
Johan Hjort
23.08
03.09
2009
Jan Mayen
10.09
27.09
2009
Vilnus
07.08
29.09
2010
G.O. Sars
24.08
11.09
2010
Johan Hjort
29.08
22.09
2010
Helmar Hanssen
26.08
12.09
2010
Vilnus
14.08
21.09
2011
Chriastine E.
27.08
17.09
2011
Johan Hjort
31.08
05.10
2011
Helmar Hanssen
09.08
24.08
2011
Vilnus
11.08
02.10
2012
G.O. Sars
18.08
12.09
2012
Johan Hjort
16.08
30.09
2012
Helmar Hanssen
06.08
05.09
2012
Vilnus
08.08
29.09
2013
G.O. Sars
23.08
19.09
2013
Johan Hjort
04.08
01.10
2013
Helmar Hanssen
19.08
01.09
2013
Vilnus
09.08
01.11
2014
G.O. Sars
23.08
19.09
2014
Johan Hjort
14.08
01.10
2014
Helmar Hanssen
19.08
01.09
2014
Vilnus
09.08
03.10
2015
G.O. Sars
11.09
09.10
2015
Johan Hjort
13.08
04.10
2015
Helmar Hanssen
17.08
07.09
2015
Vilnus
19.08
09.10
2016
Eros
17.08
20.09
2016
Johan Hjort
19.08
30.09
2016
Helmar Hanssen
24.09
05.10
2016
Fridtjof Nansen
09.08
30.09
2017
G.O.Sars
24.08
28.09
2017
Johan Hjort
21.08
04.10
2017
Helmar Hanssen
21.08
07.09
2017
Vilnyus
24.08
17.10
2017
G.O.Sars
24.08
28.09
2017
Johan Hjort
21.08
04.10
2017
Helmar Hanssen
21.08
07.09
2017
Vilnyus
24.08
17.10
2018
G.O.Sars
07.09
27.09
2018
Johan Hjort
21.08
29.09
2018
Helmar Hanssen
14.09
29.09
2018
Vilnyus
24.08
29.09
2019
G.O.Sars
14.08
09.09
2019
Johan Hjort
21.08
29.09
2019
Helmar Hanssen
22.09
02.10
2019
Vilnyus
16.08
29.09
2020
G.O.Sars
12.08
05.09
2020
Johan Hjort
21.08
28.09
2020
Kronprins Haakon
15.09
08.10
2020
Vilnyus
29.09
11.11
2021
G.O.Sars
21.08
09.10
2021
Johan Hjort
19.08
25.09
2021
Helmar Hanssen
13.09
30.09
2021
Vilnyus
12.08
25.09
2022
G.O.Sars
16.08
09.09
2022
Johan Hjort
19.08
03.10
2023
G.O.Sars
20.08
14.09
2023
Johan Hjort
25.08
30.09
2023
Kronprins Haakon
16.09
30.9
2023
Vilnyus
13.08
24.09
Table 1. Overview of participating vessels and dates for the annual 0-group surveys in the Barents Sea, 1965-2023. Note that the north-eastern-most part of the Barents Sea (polygons Franz-Josef Land and St. Anna Trough, see Fig. 1) have never been covered. For area covered each year, see maps in section 7.
The survey has generally been run from south to north in the Barents Sea; that is, the research vessels have started in south and worked their way northwards. This is a broad pattern and there are many exceptions in specific years. Maps with cruise lines and station positions for the different research vessels are included in annual cruise reports that are available electronically (Table 2). The cruise lines are generally placed either in S-N or W-E directions, although zig-zag or more irregular patterns have also sometimes been used to obtain a good coverage of the survey area within the limits of time and ship availability.
A change in survey lines was made in the mid-1990s. From 1980 and up to 1994 (and also in 1997), the S-N survey lines followed longitudes and the E-W lines followed latitudes. From 1995 onwards (except 1997), the survey lines were placed equidistant (35 nm apart). The grid was oriented true North along the 30o E longitude, while it deviated in NW direction in the western Barents Sea, and in NE direction in the eastern Barents Sea. A consequence of this was an opening of the sampling grid in the northern end, compared to when the lines followed longitudes.
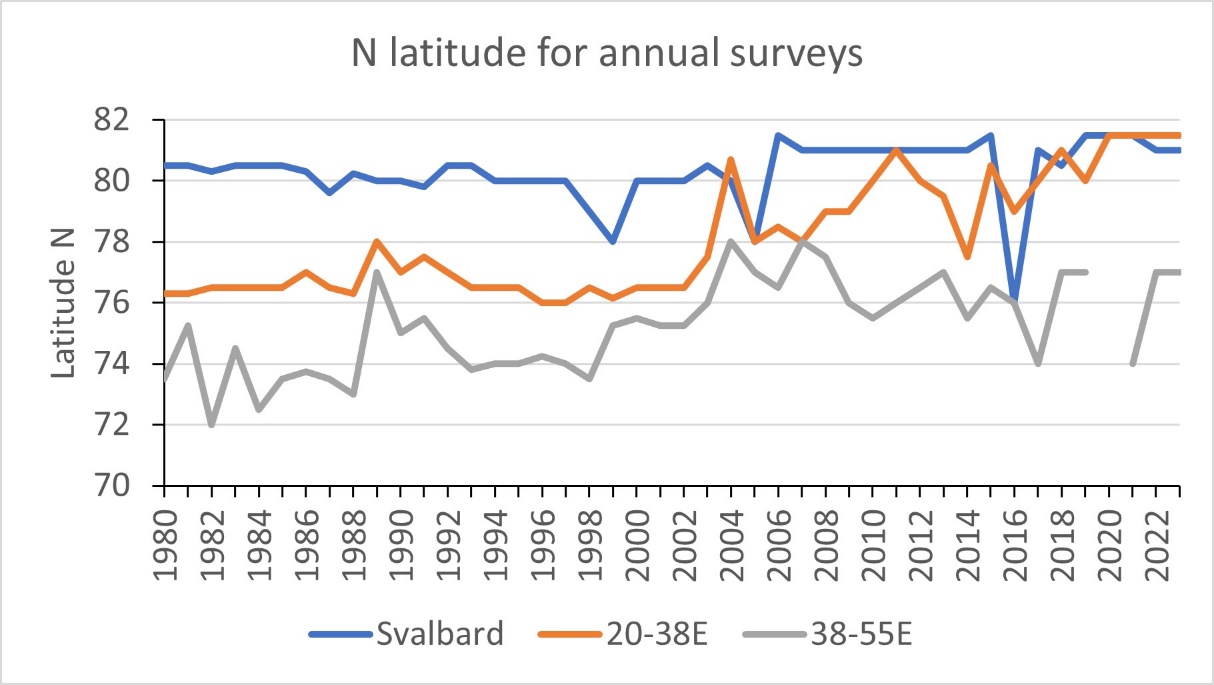
The surveyed area has expanded northward in concert with reduction of sea ice in the Barents Sea. This can be seen from the maps with station locations in Part 7. A summary of the northern boundary of the survey area in three sectors is illustrated in Fig. 7.
In the Svalbard (Spitsbergen Archipelago) sector, the survey area has extended up along the west side of Spitsbergen to around 80-81oN. Up to 2004, the survey extended north to 80-80.5oN, while from 2006 it extended north to 81oN or beyond (Fig. 7). The northwestern corner of Spitsbergen lies just south of 80oN. With the northward extension from 2006, there was also an eastward extension to cover the waters north of Svalbard (Spitsbergen Archipelago) , east to 20-35oE. In three of the years, the waters west of Spitsbergen was either not sampled (2016) or only partially sampled (north to 78 o N; 1999 and 2005).
In a sector through the central Barents Sea, east of Svalbard (Spitsbergen Archipelago) and east to about 38oE, the sampling extended north to 76-77 oN in the years up to 2002 (except for two years, 1989 and 1991), while from 2004 the survey area extended north to 78 oN or beyond (Fig. 7). The northward shift reflects a change to less sea ice and more open water in the northern Barents Sea, while the large variability in recent year reflects variable ice conditions. A similar northward extension is seen for the area east of 38o E, but with considerable variation among years reflecting variable sea ice conditions as well as vessel availability (Fig. 7).
Figure 7. The northern boundary of the survey area in three sectors: Svalbard (Spitsbergen Archipelago) sector, area between 20 and 38°E and area between 38-55°E.
The survey is semi-synoptic since it takes about 3-4 weeks, or in some cases longer, to complete the survey of 0-group distribution. The 0-group survey typically started in mid-August (10-20 August) and ended in early September (5-15 September). This was the case during the 1980s and 90s when the 0-group investigations were done as a separate cruise, or as the first part of a combined multispecies cruise. This pattern with a main part of sampling in the second half of August and the first part of September has continued after 2004 when the 0-group survey became part of the ecosystem survey, although there has been an extension of sampling later in September as the survey has extended northward (described in the following).
Survey year
Author
Year
Title
ICES
IMR/PINRO Joint Report Series
Pages
1965
Anon.
1965
Preliminary Report of the joint Soviet-Norwegian investigations in the Barents Sea and adjacent waters September 1965
CM 1965/No. 161
1966
Anon.
1966
Preliminary Report of the joint international 0-group fish survey in the Barents Sea and adjacent waters August/Sept 1966
CM 1966/H:23
17
1967
Anon.
1967
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August/September 1967
C.M. 1967/H:31
18
1968
Anon.
1968
Preliminary Report of the 0-group fish survey in the Barents Sea and adjacent waters August-September 1968
C.M. 1968/H:25
12
1969
Anon.
1969
Preliminary Report of the 0-group fish survey in the Barents Sea and adjacent waters August-September 1969
C.M. 1969/F:34
14
1970
Anon.
1970
Preliminary Report of joint Soviet-Norwegian 0-group fish survey in the Barents Sea and adjacent waters August-September 1970
C.M. 1970/H:34
13
1971
Anon.
1971
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1971
C.M. 1971/H:32
14
1972
Anon.
1972
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1972
C. M.1973/H:15
16
1973
Anon.
1973
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1973
C.M. 1973/H:25
28
1974
Anon.
1974
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1974
C.M. 1974/H:33
23
1975
Anon.
1975
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1975
C.M. 1975/H:48
23
1976
Anon.
1976
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1976
C.M. 1976/H:43
26
1977
Anon.
1977
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1977
C.M. 1977/H:45
26
1978
Anon.
1978
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1978
CM 1978/H:33
26
1979
Anon.
1979
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1979
CM 1979/H:65
26
1980
Anon.
1980
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1980
CM 1980/G:53
26
1981
Anon.
1981
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1981
CM 1981/G:78
28
1982
Anon.
1982
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1982
CM 1982/G:44
28
1983
Anon.
1983
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1983
CM 1983/G:35
28
1984
Anon.
1984
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1984
C.M. 1984/H:36
28
1985
Anon.
1985
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1985
C.M. 1985/G:75
28
1986
Anon.
1986
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1986
C.M. 1986/G:78
28
1987
Anon.
1987
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1987
C.M. 1987/G:38
32
1988
Anon.
1988
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1988
C.M. 1988/G:45
38
1989
Anon.
1989
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1989
C.M. 1989/G:40
40
1990
Anon.
1990
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1990
C.M. 1990/G:46
36
1991
Anon.
1991
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1991
C.M. 1991/G:50
34
1992
Anon.
1992
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1992
C.M. 1992/G:82
33
1993
Anon.
1994
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1993
C.M. 1994/G:3
38
1994
Anon.
1995
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1994
C.M. 1995/G:xx
36
1995
Anon.
1996
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1995
C.M. 1996/G:xx
36
1996
Anon.
1996
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1996
C.M. 1996/G:31
38
1997
Anon.
1997
Preliminary Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1997
25
1998
Anon.
2001
Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1998
No. 2/2001
26
1999
Anon.
2001
Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 1999
No. 3/2001
27
2000
Anon.
2001
Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 2000
No. 4/2001
26
2001
Anon.
2001
Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 2001
No. 8/2001
26
2002
Anon.
2002
Report of the international 0-group fish survey in the Barents Sea and adjacent waters August-September 2002
No. З/2002
28
2003
Anon.
2003
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea, August – October 2003.
No. 2/2003
55
2004
Anon.
2004
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea, August – October 2004, Volume 1
No. 3/2004
71
2005
Anon.
2005
Survey report from the Joint Norwegian/Russian ecosystem survey in the Barents Sea August-October 2005, Volume 1
No. 3/2005
99
2006
Anon.
2006
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-October 2006 (vol.1).
No. 2/2006
97
2007
Anon.
2007
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-October 2007 (vol.1).
No. 4/2007
97
2008
Anon.
2009
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-October 2008 volume 1.
No. 1/2009
103
2009
Anon.
2009
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-October 2009 (adopted vol.)
No. 2/2010
118
2010
Anon.
2010
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-September 2010.
No. 4/2010
108
2011
Anon.
2011
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-October 2011
No. 3/2011
118
2012
Eriksen
2012
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-October 2012
No. 2/2012
139
2013
Prokhorova
2013
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea and adjacent waters, August-October 2013
No. 4/2013
131
2014
Eriksen
2015
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea and adjacent waters, August-October 2014
No. 1/2015
153
2015
Prozorkevich and Sunnanå
2016
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea and adjacent waters, August-October 2015
No. 1/2016
77
2016
Prozorkevich and Sunnanå
2017
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea and adjacent waters, August-October 2016
No. 2/2017
101
2017
Prozorkevich et al.
2018
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea and adjacent waters, August-October 2017
No. 2/2018
97
2018
van der Meeren and Prozorkevich
2019
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea and adjacent waters, August-October 2018
No. 2/2019
85
2019
Prozorkevich and van der Meeren
2020
Survey report from the joint Norwegian/ Russian ecosystem survey in the Barents Sea and adjacent waters August-October 2019.
No. 1/2020
93
2020
van der Meeren and Prozorkevich
2021
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea and adjacent waters, August-November 2020
No. 1/2021
123
2021
Prozorkevich and van der Meeren
2022
Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea and adjacent waters, August-September 2021
No. 2/2022
111
2022
van der Meeren and Prozorkevich
2023
Survey report from the joint Norwegian/Russian Ecosystem Survey in the Barents Sea and the adjacent waters August-December 2022
No. 2023-10
2023
Prozorkevich and van der Meeren
2024
Survey report (Part 1) from the joint Norwegian/Russian Ecosystem Survey in the Barents Sea and the adjacent waters August-October 2023
No. 2024-2
Table 2. Reports from Joint Norwegian-Russian (IMR-PINRO) 0-group cruises in the Barents Sea, 1965-20 23 . The annual survey reports for 1965-1996 are available as ICES Council Meeting Reports, while the annual survey reports for 1998-2023 are available in the IMR/PINRO Joint Report Series from the IMR web page ( Rapporter Havforskningsinstituttet (hi.no).
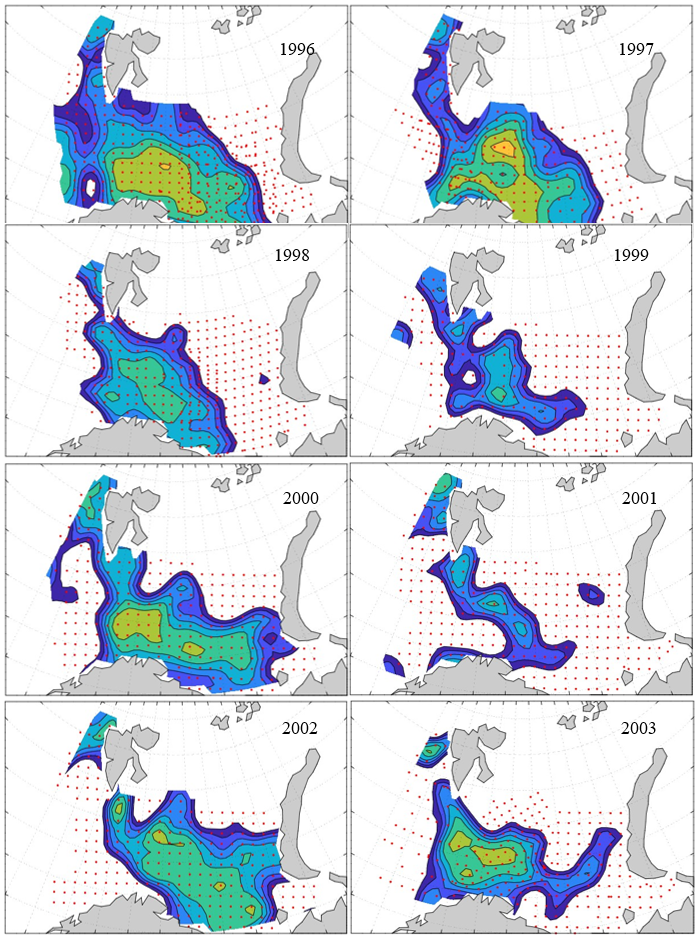
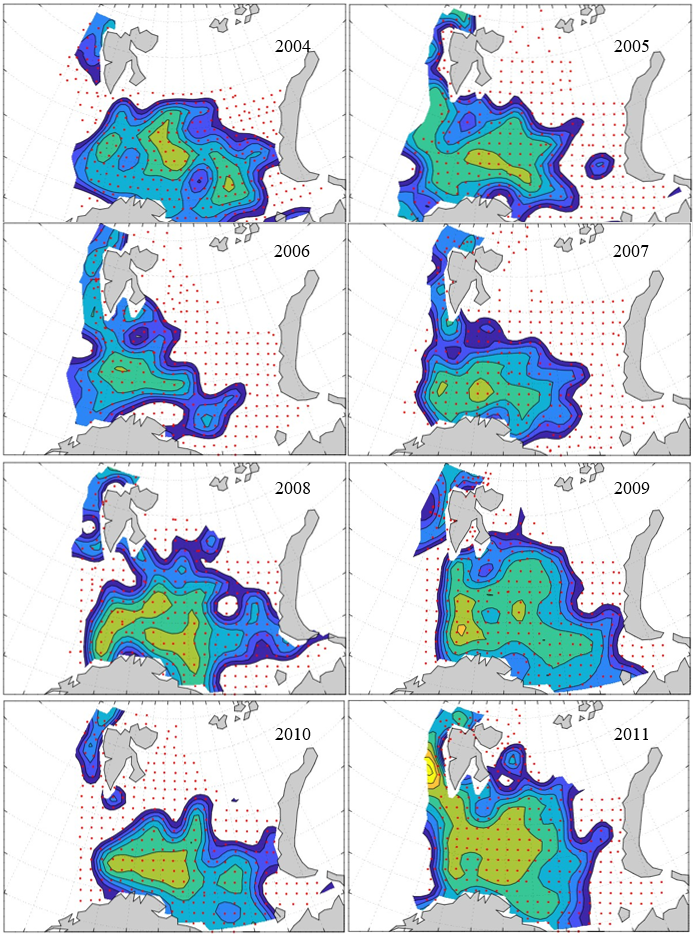
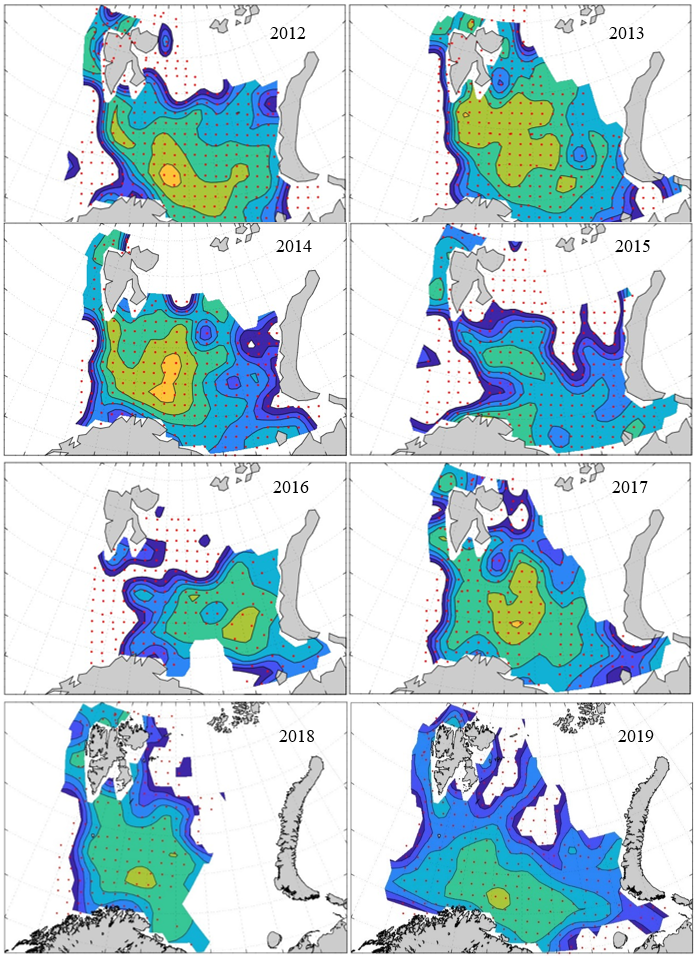
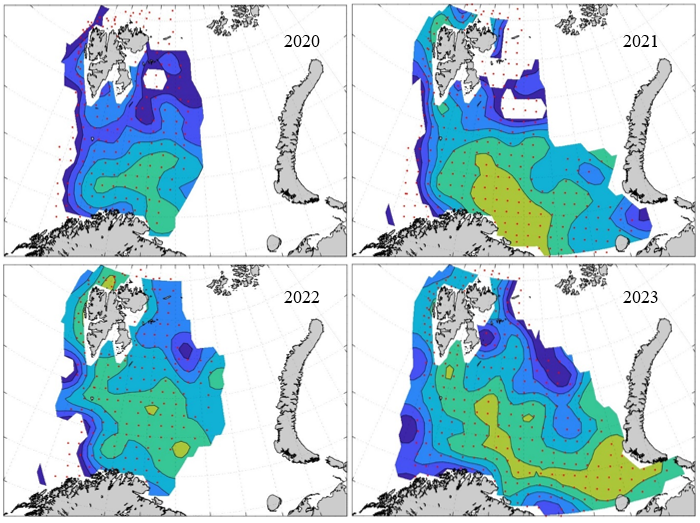
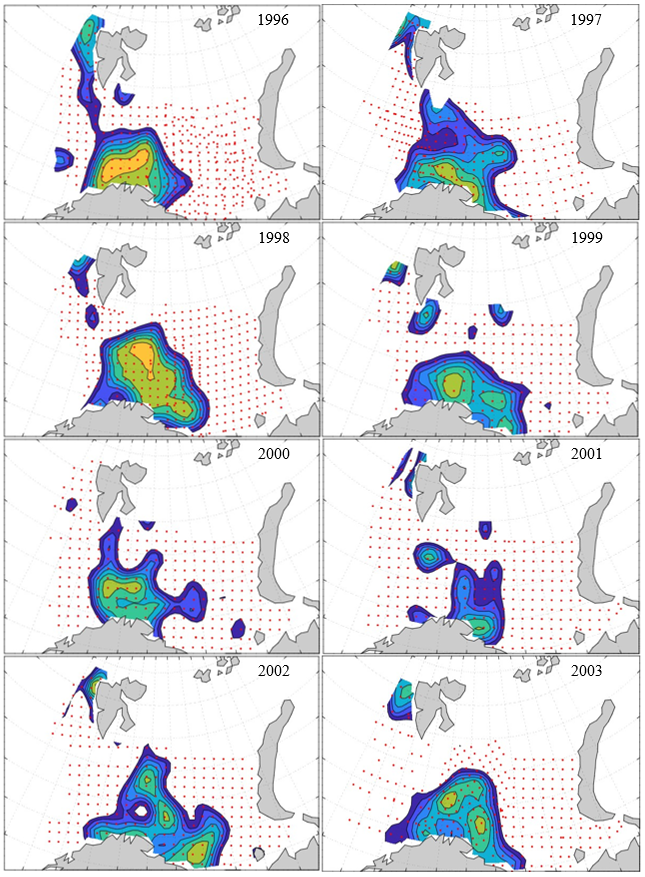
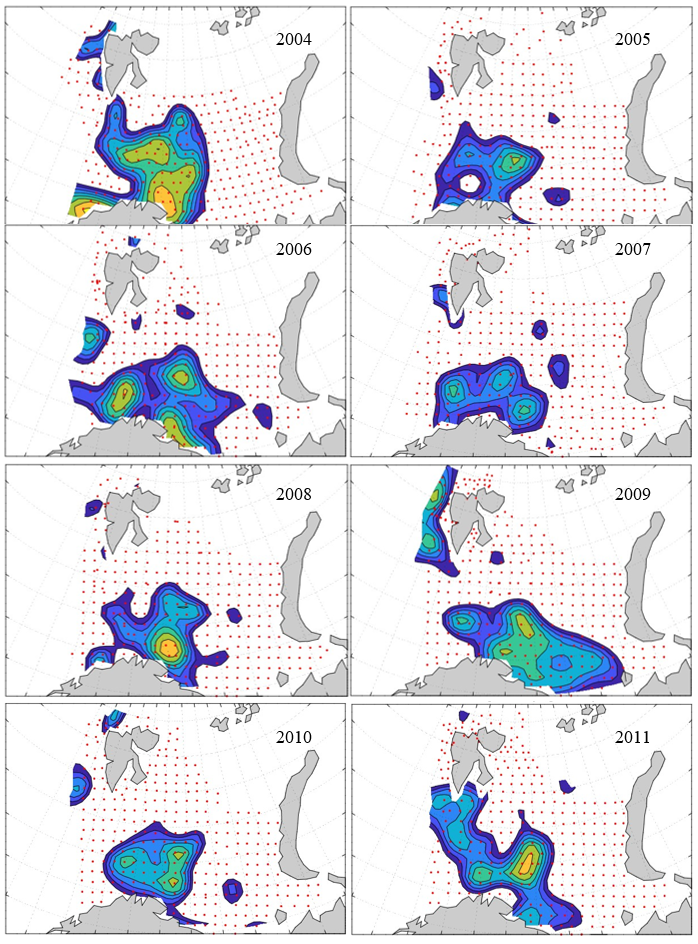
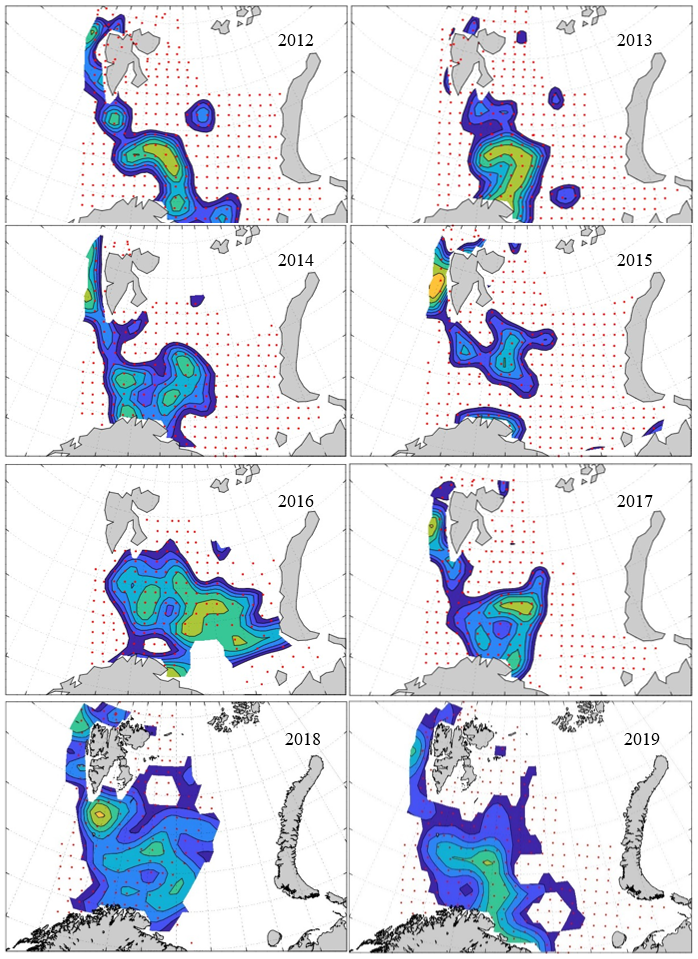
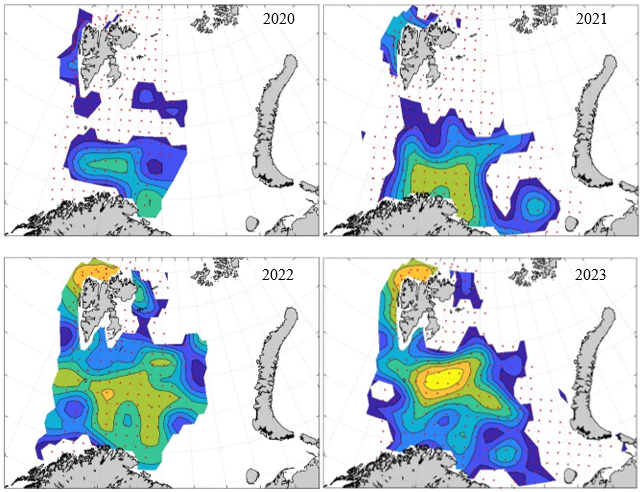
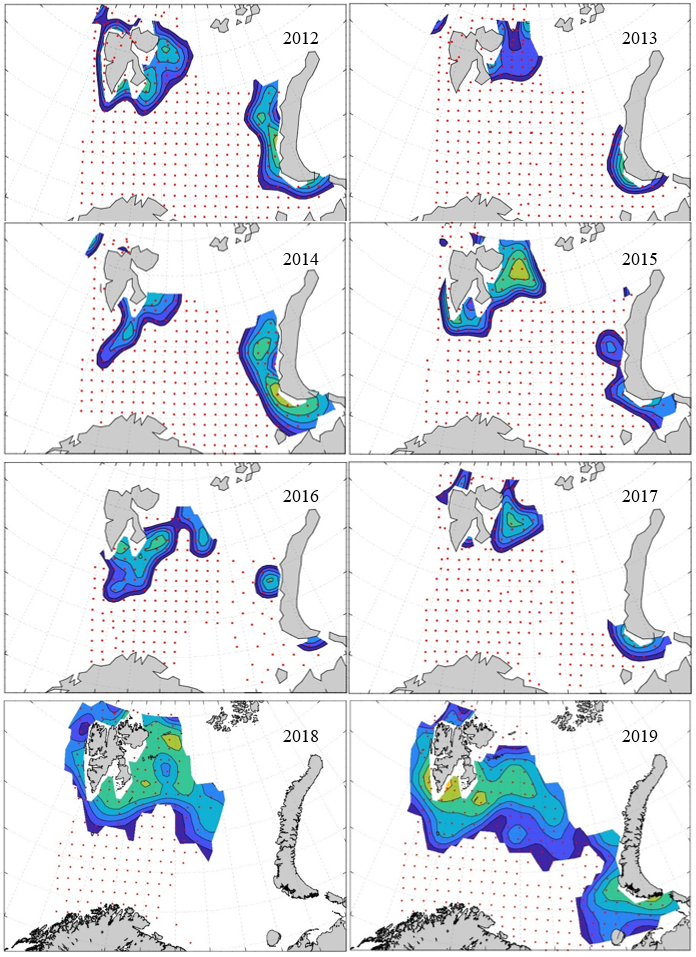
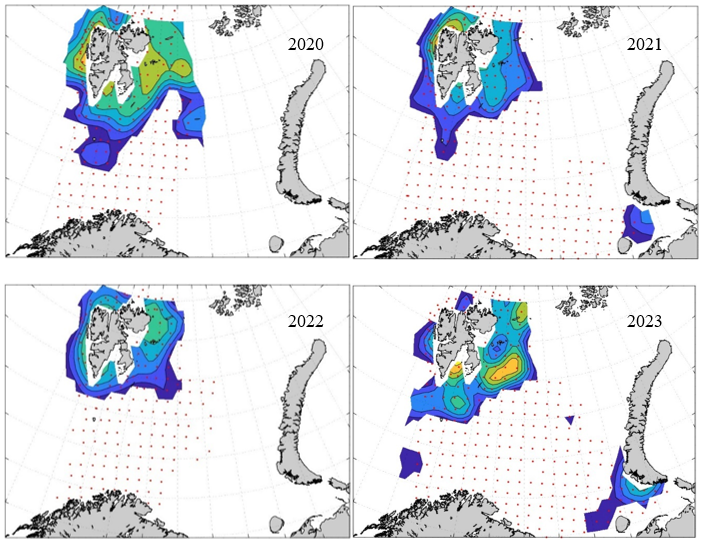
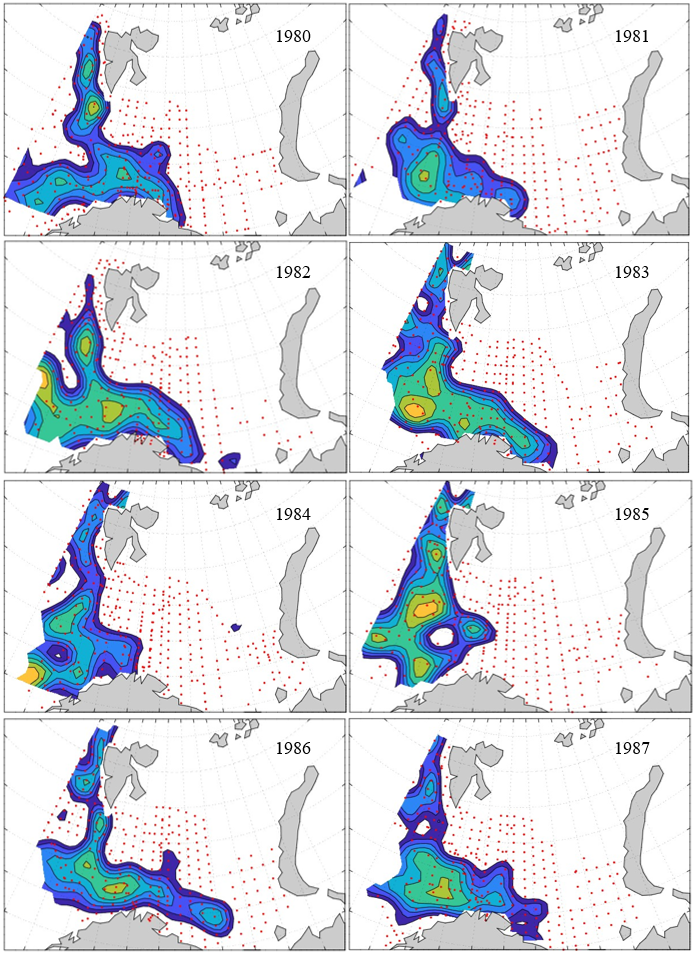
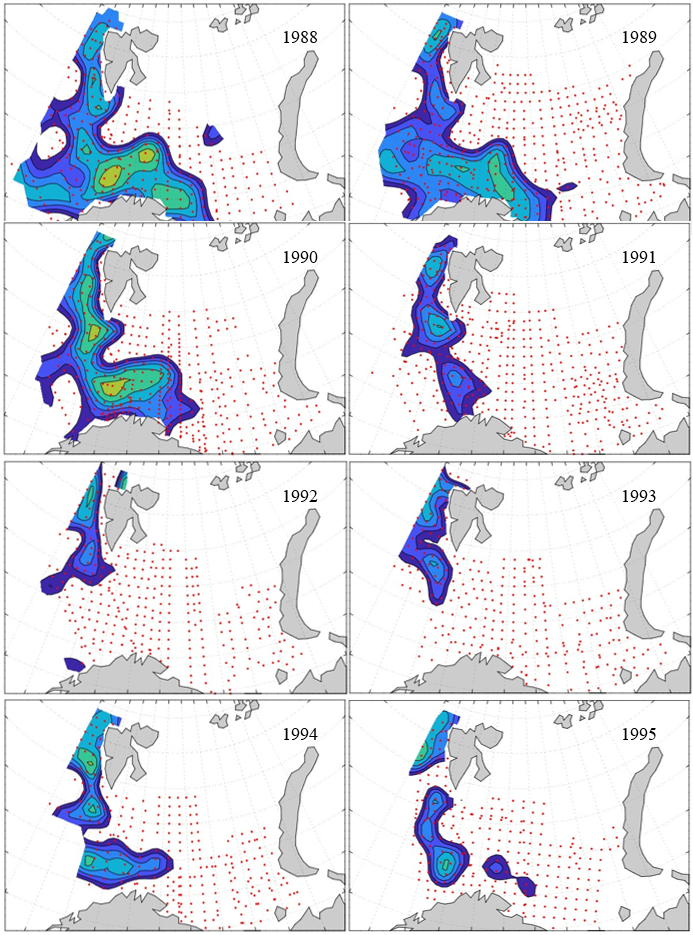
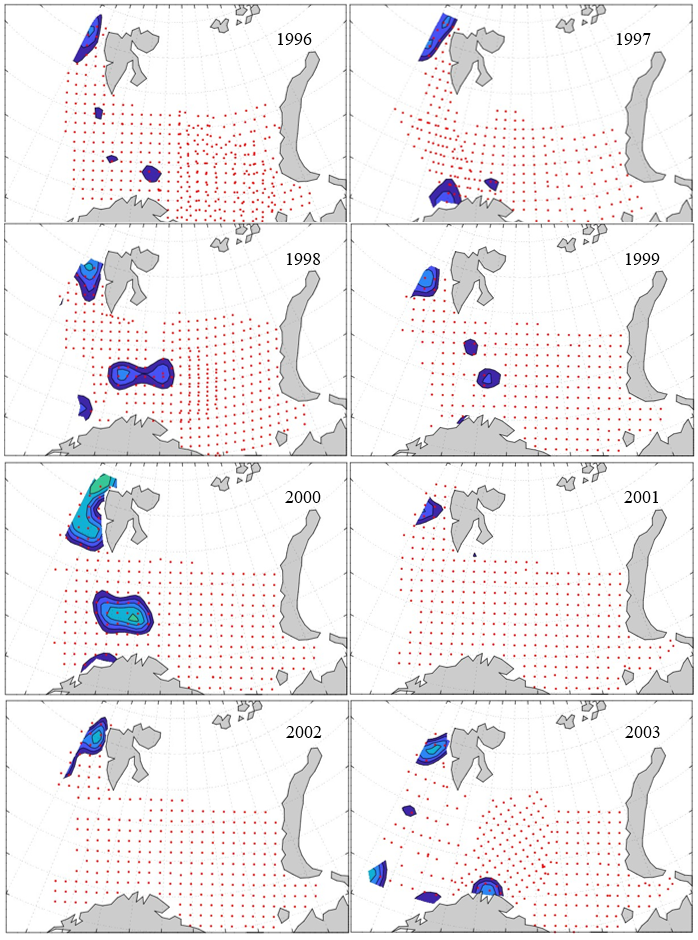
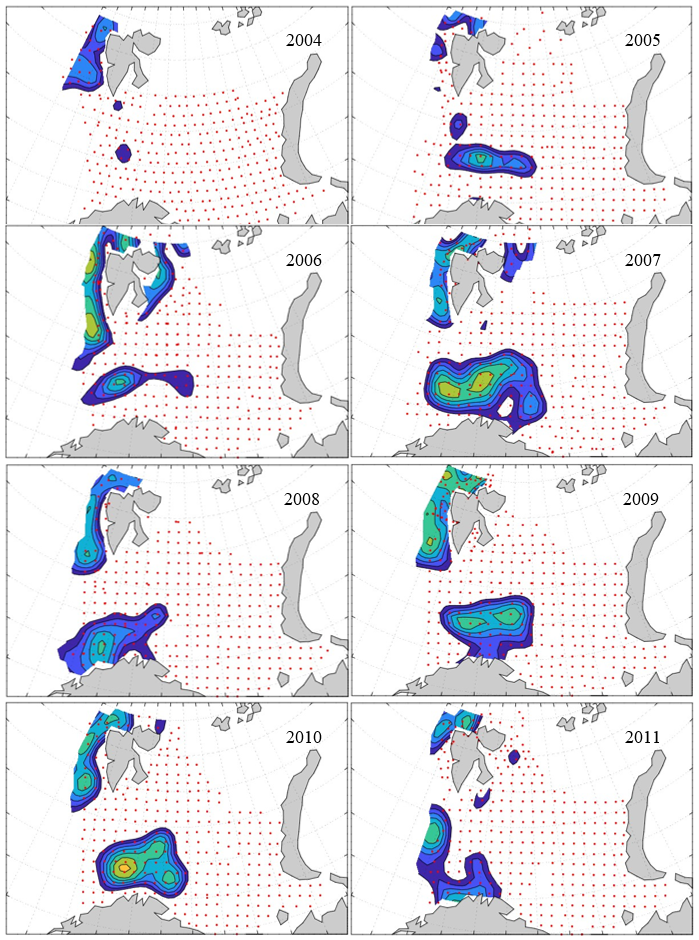
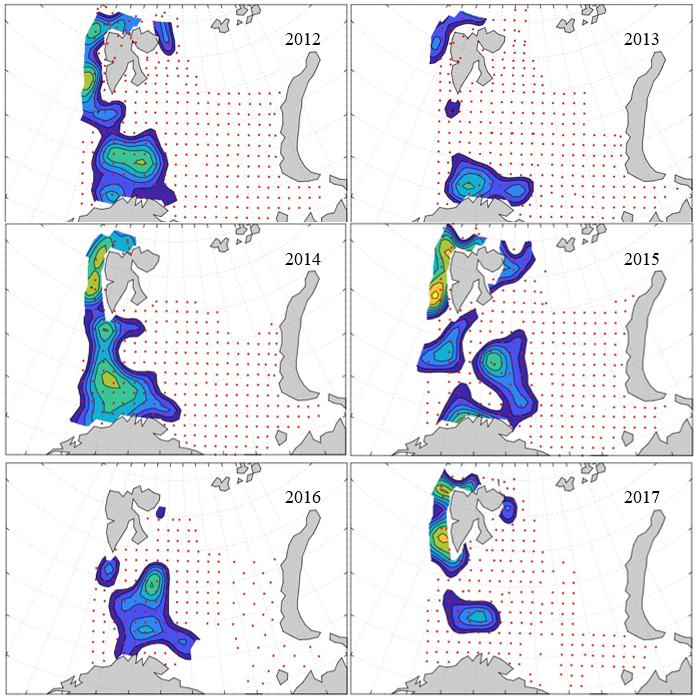
7 - Spatial distribution of 0-group fish in 1980-2023
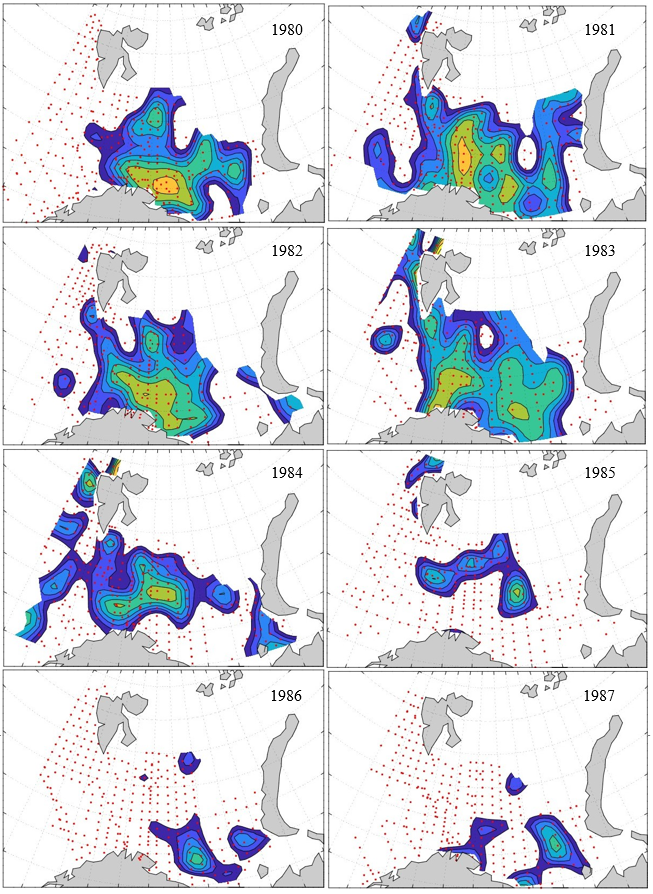
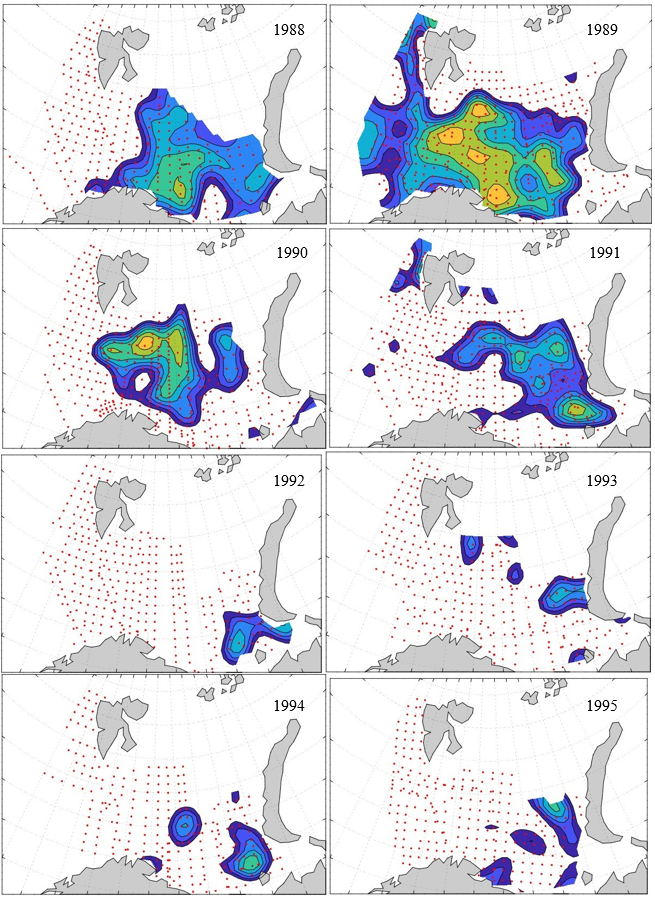
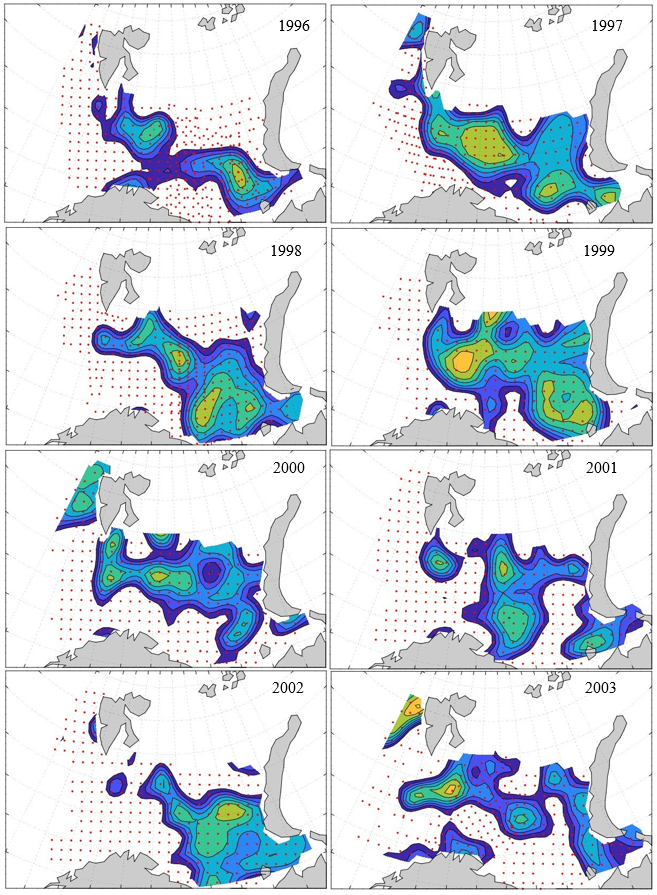
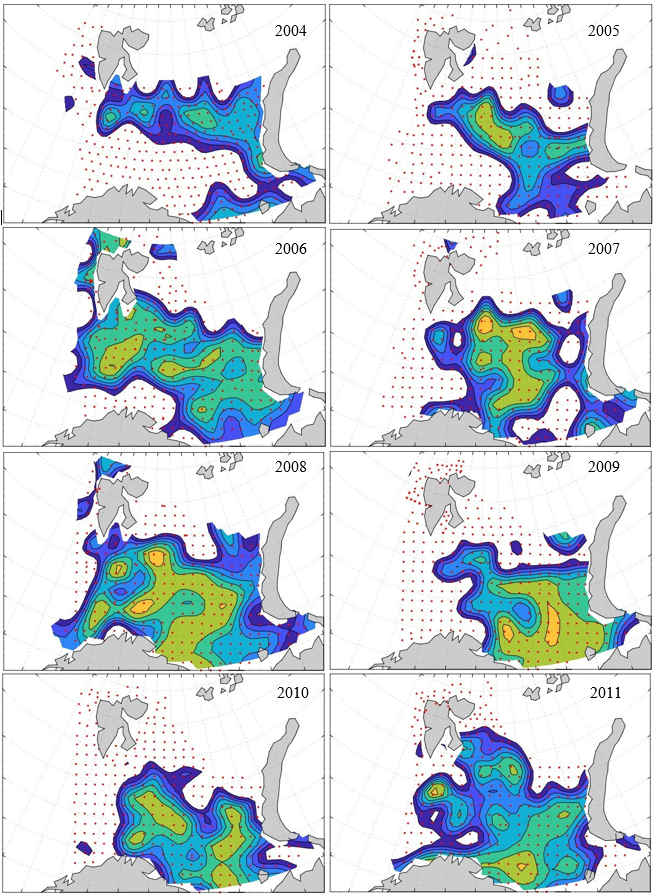
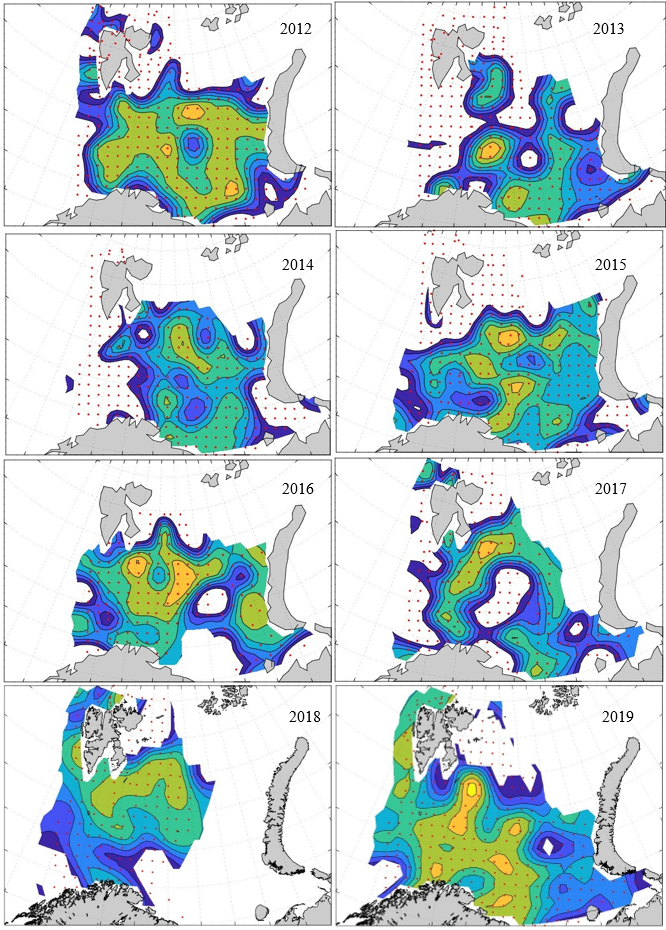
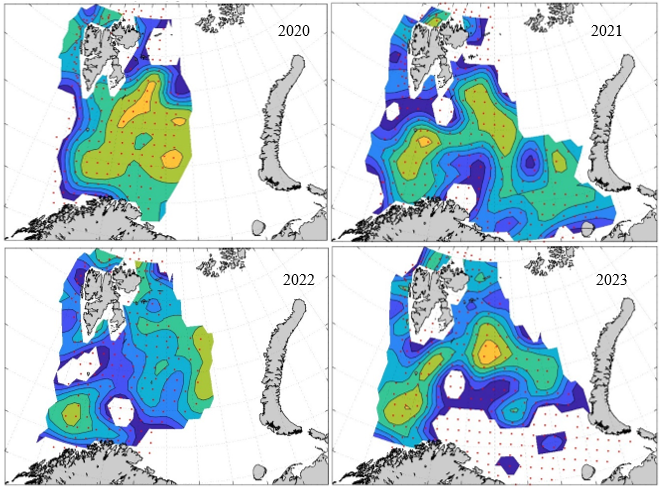
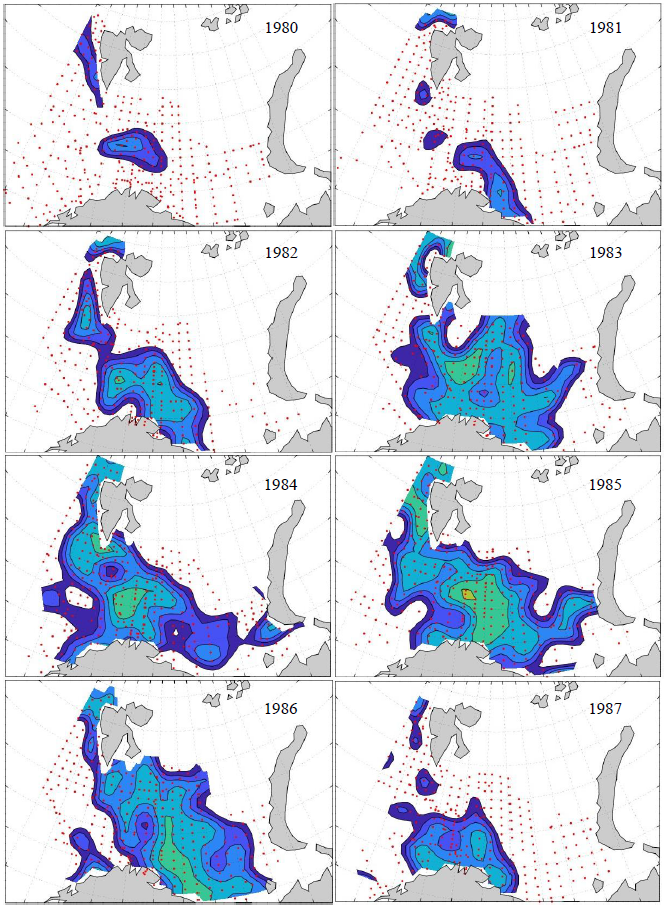
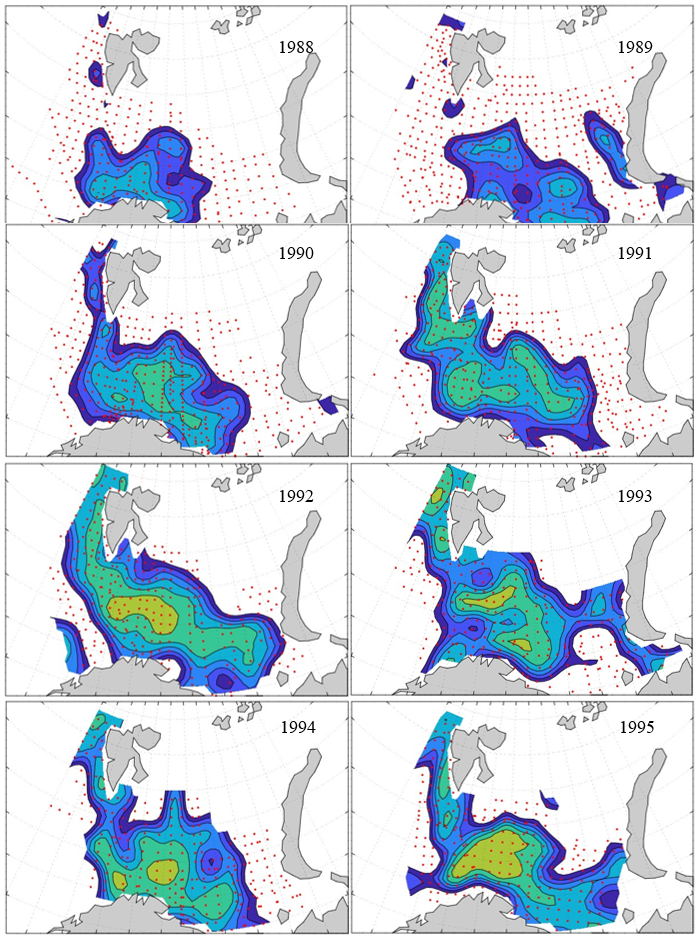
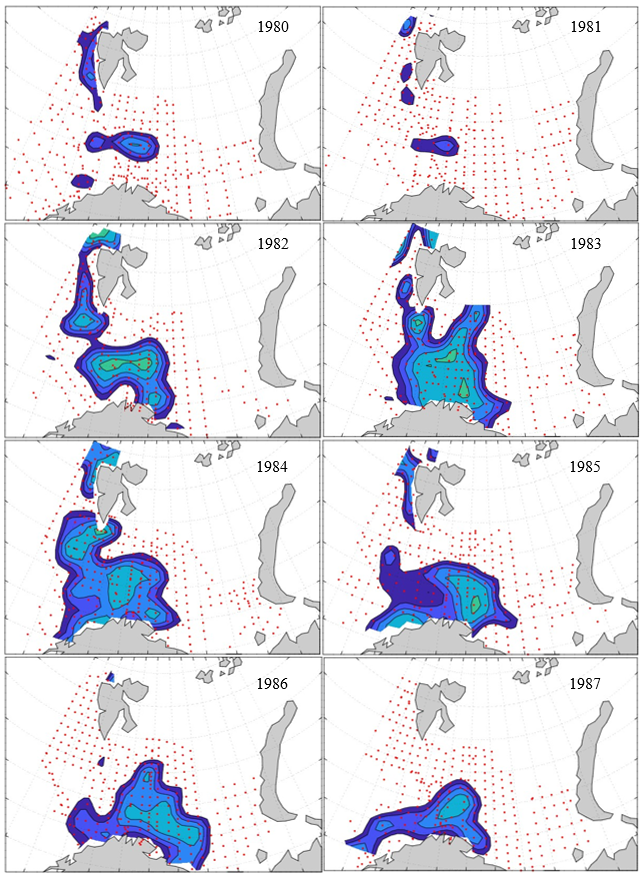
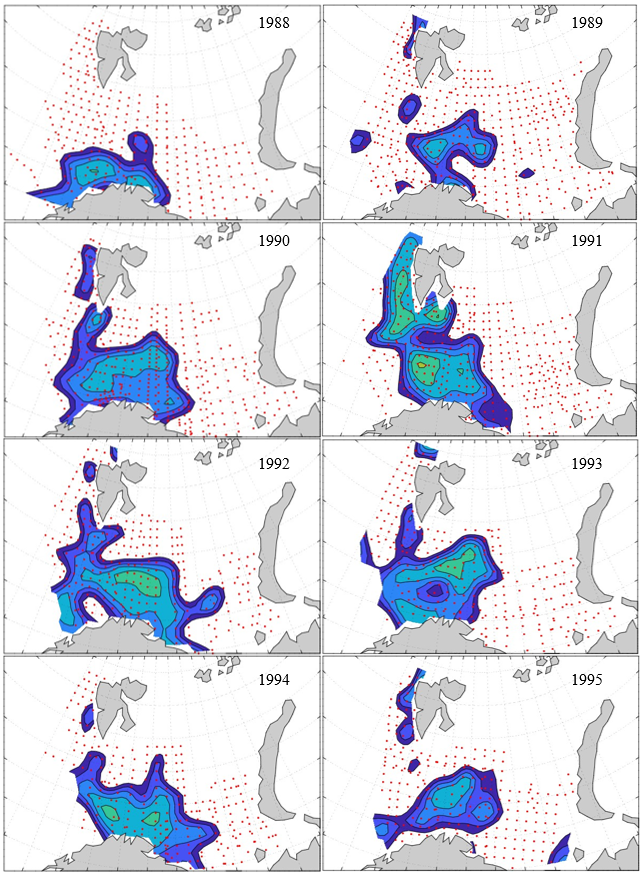
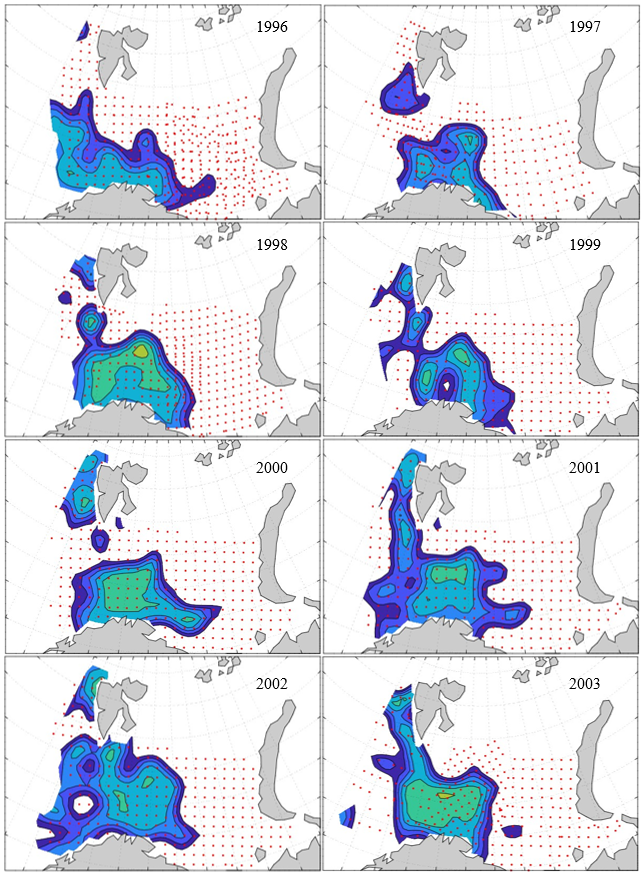
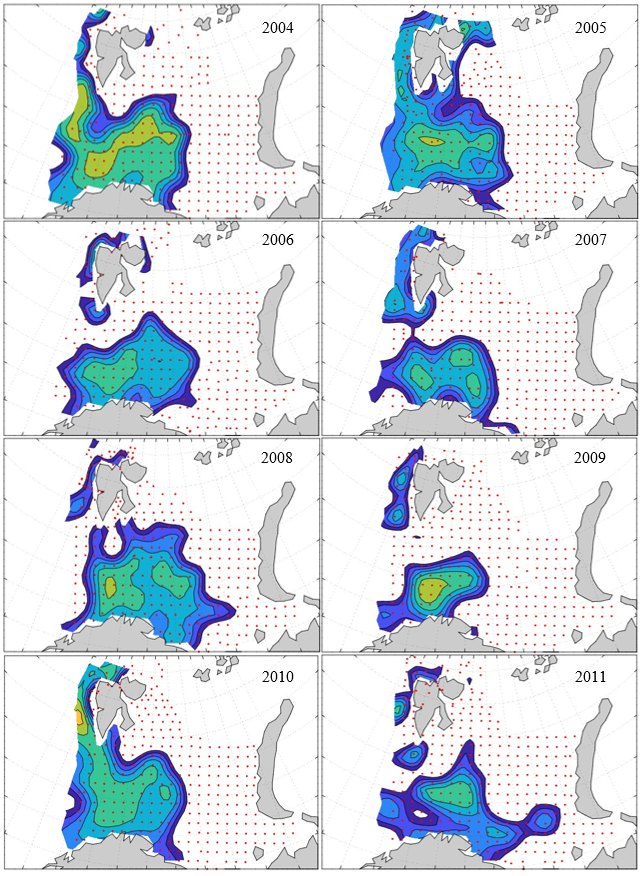
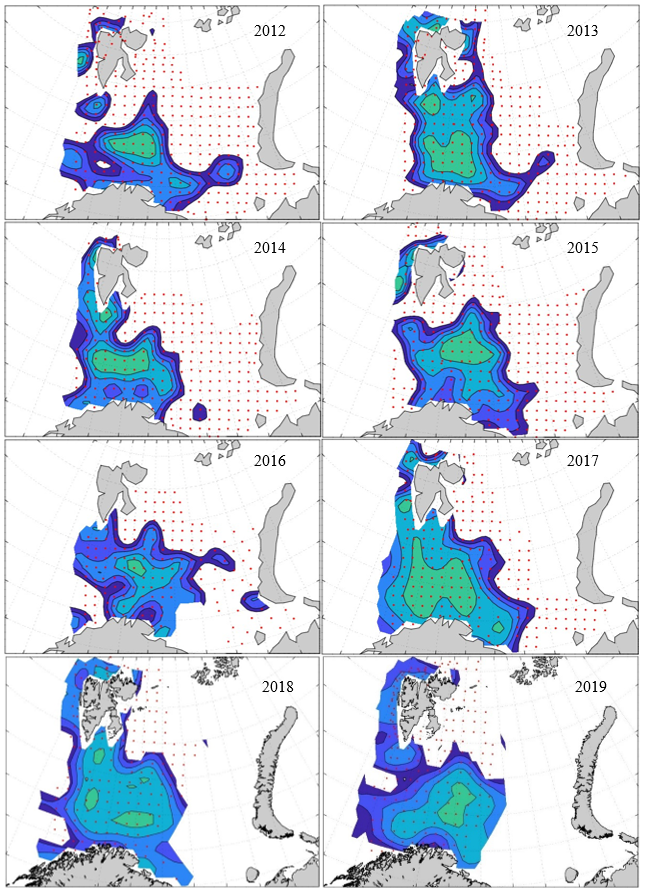
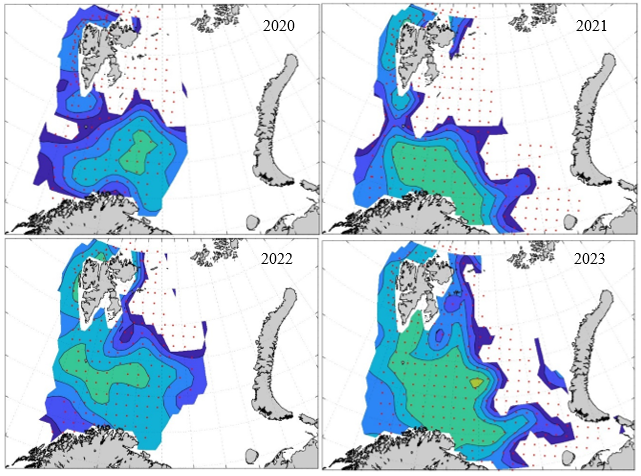
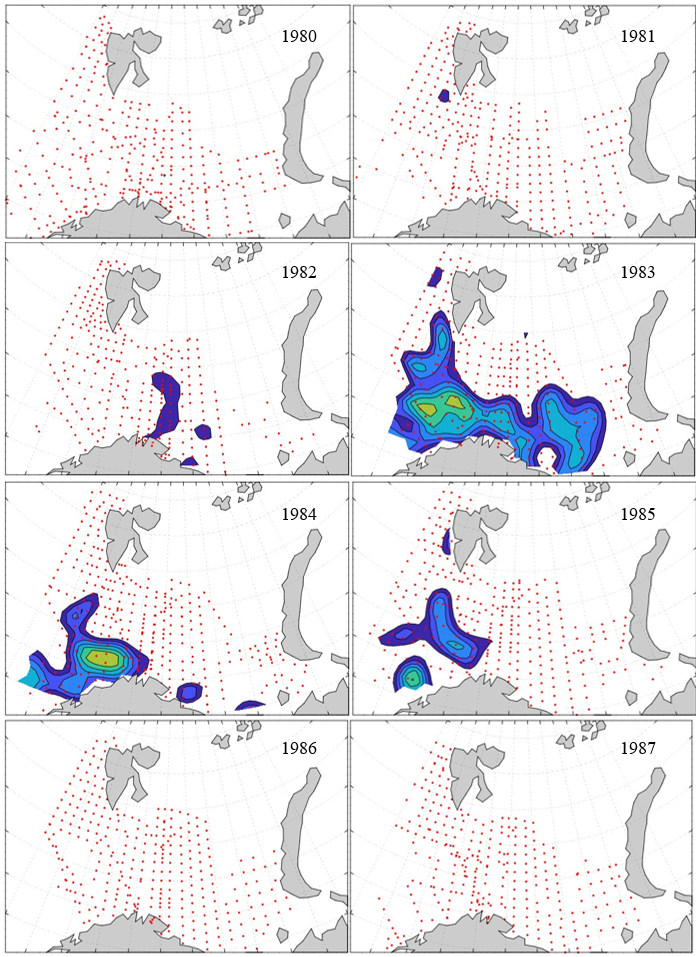
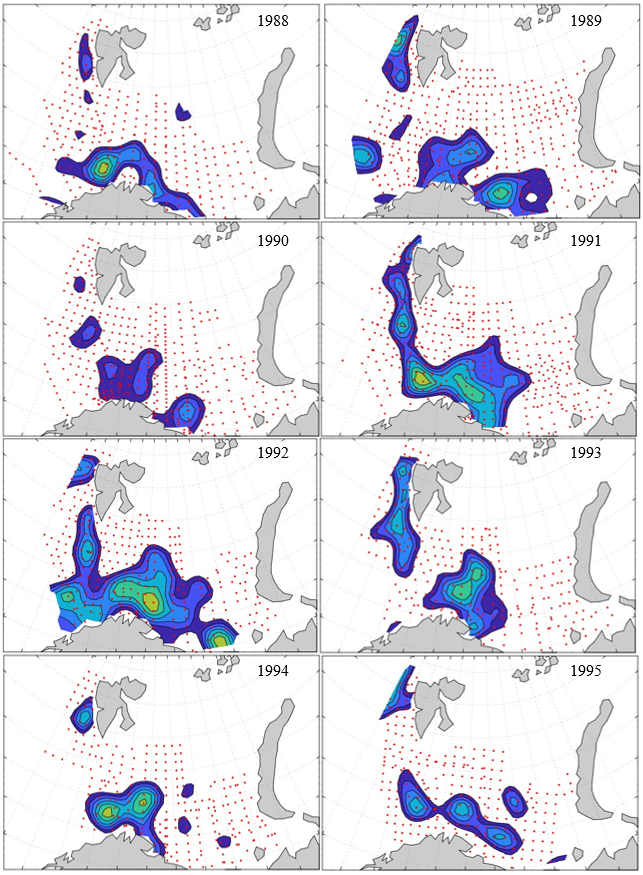
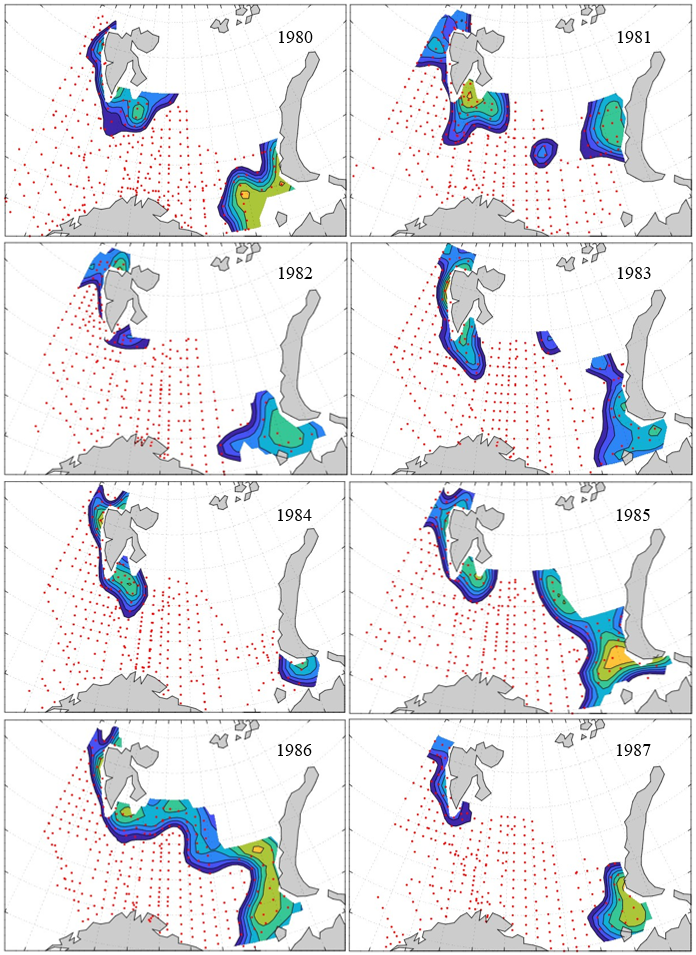
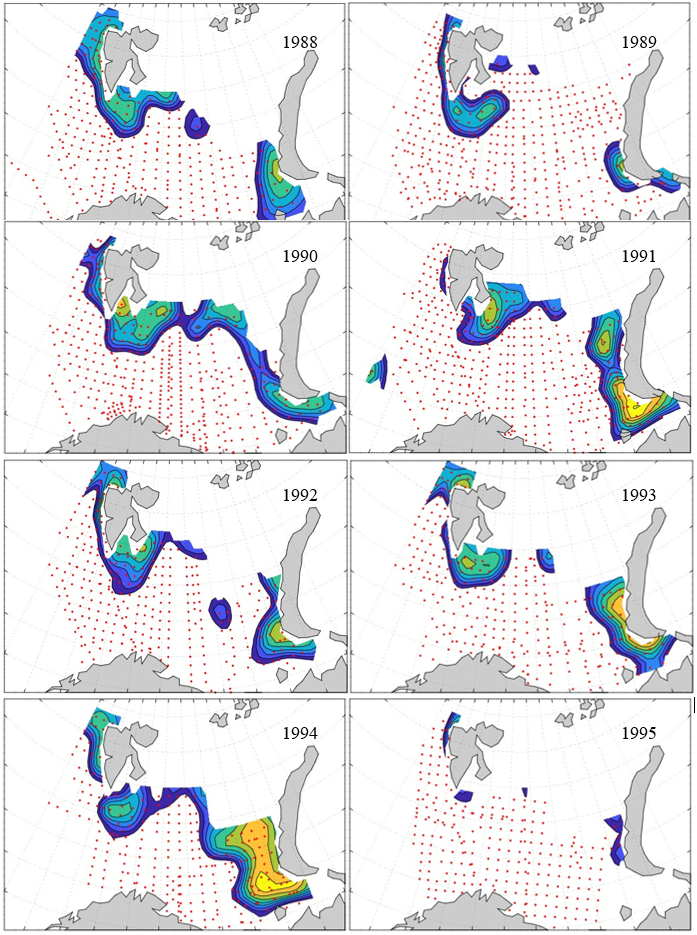
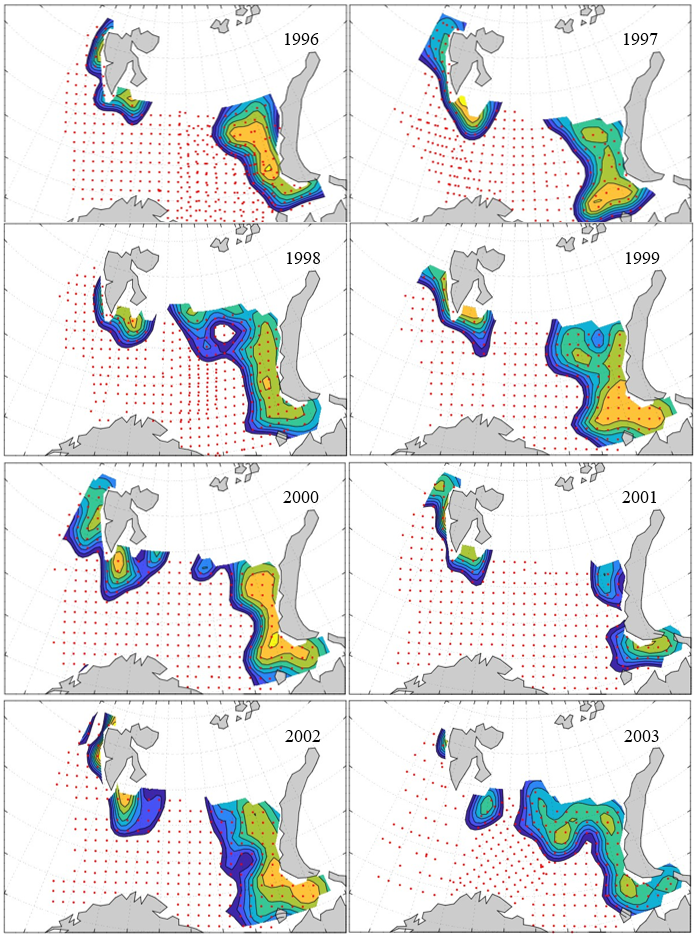
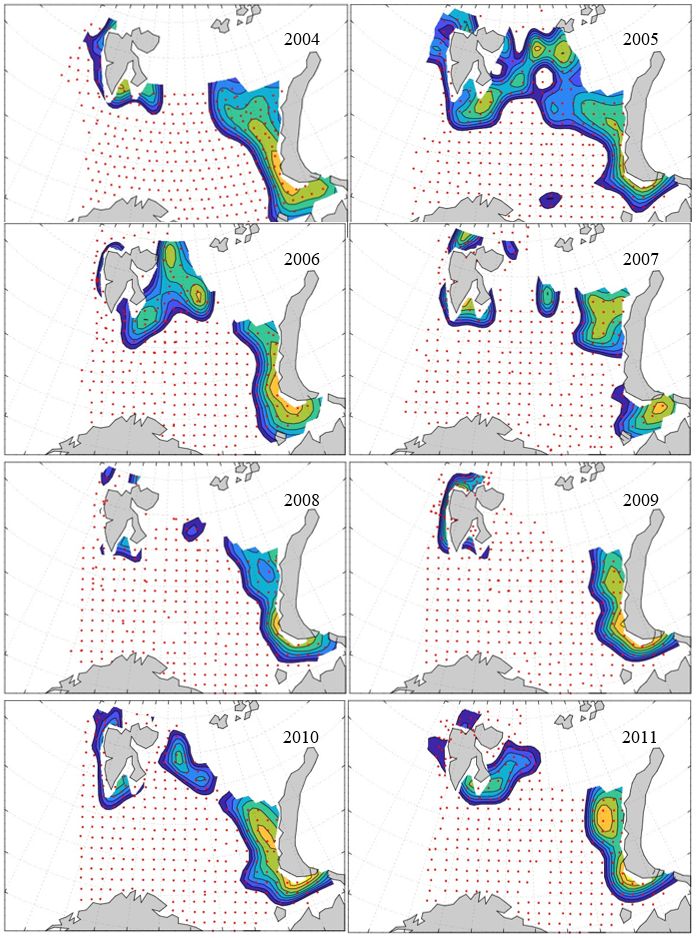
Figure 8. Distribution of 0-group capelin in 2016.
In this section, maps of spatial distribution of 0-group density of six species are shown, based on log transformed abundance per station (colored). The species are: Atlantic capelin Mallotus villosus , Atlantic cod Gadus morhua , haddock Melanogrammus aeglefinus , Atlantic herring Clupea harengus , polar cod Boreogadus saida , and redfish (Sebastes spp.).
Species abundance at stations have been estimated, based on species catches at station, by standard methods (Eriksen et al. 2009), taking into account the opening of trawl, vessels speed, towing distance, and number of depths layers covered (see section 2.3). The abundances have been corrected for size-dependent catch efficiency for all species except redfish Sebastes spp. (section 4). Abundances are given as areal density of 0-group, expressed as number of individuals per square nautical miles.
Abundance is shown by color, where light yellow indicates highest value, and dark blue lowest value. The scale is log10, with steps of one log10 unit, corresponding to factor 10. It ranges from >100 million individuals per (nautical miles)2 to <100 individuals per (nautical miles)2 . Stations are shown by red dots.
0-group capelin are generally widespread in the Barents Sea (except 1985-86 and 1992-1995) and occurrence area has varied from 116 to 1130 thousand km2. Distribution of 0-group herring was widest in 1983, 1992, 2016 and in 2022-2023, and it seems that larger occupation area was not related to occurrence of strong year classes. 0-group cod are generally widely distributed and abundant year classes seem to be observed on larger area. Haddock were widespread in 2004, 2008, 2017 and 2023, and varied from 62 to 630 thousand km2. Redfish and polar cod 0-group distributions are generally more restricted than for cod, capelin, haddock, and herring. The widest distribution was observed in 1982-83 (515 thousand km2, redfish) and 1999 and 2005 (560 thousand km2 , polar cod).
7.1 - Capelin
Figur 9. Distribution of 0-group capelin between 1980-1987. Figur 10. Distribution of 0-group capelin between 1988-1995. Figur 11. Distribution of 0-group capelin between 1996-2003. Figur 12. Distribution of 0-group capelin between 2004-2011. Figur 13. Distribution of 0-group capelin between 2012-2019. Figur 14. Distribution of 0-group capelin between 2020-2023.
7.2 - Cod
Figur 15. Distribution of 0-group cod between 1980-1987. Figur 16. Distribution of 0-group cod between 1988-1995. Figur 17. Distribution of 0-group cod between 1996-2003. Figur 18. Distribution of 0-group cod between 2004-2011. Figur 19. Distribution of 0-group cod between 2012-2019. Figur 20. Distribution of 0-group cod between 2020-2023.
7.3 - Haddock
Figur 21. Distribution of 0-group haddock between 1980-1987. Figur 22. Distribution of 0-group haddock between 1988-1995. Figur 23. Distribution of 0-group haddock between 1996-2003. Figur 24. Distribution of 0-group haddock between 2004-2011. Figur 25. Distribution of 0-group haddock between 2012-2019. Figur 26. Distribution of 0-group haddock between 2020-2023.
7.4 - Herring
Figur 27. Distribution of 0-group herring between 1980-1987. Figur 28. Distribution of 0-group herring between 1988-1995. Figur 29. Distribution of 0-group herring between 1996-2003. Figur 30. Distribution of 0-group herring between 2004-2011. Figur 31. Distribution of 0-group herring between 2012-2019. Figur 32. Distribution of 0-group herring between 2020-2023.
7.5 - Polar cod
Figur 33. Distribution of 0-group polar cod between 1980-1987. Figur 34. Distribution of 0-group polar cod between 1988-1995. Figur 35. Distribution og 0-group polar cod between 1996-2003. Figur 36. Distribution of 0-group polar cod between 2004-2011. Figur 37. Distribution of 0-group polar cod between 2012-2019. Figur 38. Distribution of 0-group polar cod between 2020-2023.
7.6 - Redfish
Figur 39. Distribution of 0-group redfish between 1980-1987. Figur 40. Distribution of 0-group redfish between 1988-1995. Figur 41. Distribution of 0-group redfish between 1996-2003. Figur 42. Distribution of 0-group redfish between 2004-2011. Figur 43. Distribution of 0-group redfish between 2012-2017.
8 - Abundance and biomass indices
Abundance and biomass estimates were calculated by different software during the last for decades: SAS (for the 23 strata, see Fig. 3, 1980-2017), MatLab (for the new 15 TIBIA/WGIBAR- polygons (see Fig. 1, 1980- 2018, WGIBAR 2018) and R (for the 15 WGIBAR- polygons (2003-2023).
8.1 - Indices calculated in SAS
Table 3. 0-group abundance indices (in millions) with 95% confidence limits, not corrected for capture efficiency. These indices have been reported to ICES WG groups (AFWG, WGWIDE and WGIBAR).
Year
Capelin
Cod
Haddock
Herring
Redfish
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
1980
197278
131674
262883
72
38
105
59
38
81
4
1
8
277873
0
701273
1981
123870
71852
175888
48
33
64
15
7
22
3
0
8
153279
0
363283
1982
168128
35275
300982
651
466
835
649
486
812
202
0
506
106140
63753
148528
1983
100042
56325
143759
3924
1749
6099
1356
904
1809
40557
19526
61589
172392
33352
311432
1984
68051
43308
92794
5284
2889
7679
1295
937
1653
6313
1930
10697
83182
36137
130227
1985
21267
1638
40896
15484
7603
23365
695
397
992
7237
646
13827
412777
40510
785044
1986
11409
98
22721
2054
1509
2599
592
367
817
7
0
15
91621
0
184194
1987
1209
435
1983
167
86
249
126
76
176
2
0
5
23747
12740
34755
1988
19624
3821
35427
507
296
718
387
157
618
8686
3325
14048
107027
23378
190675
1989
251485
201110
301861
717
404
1030
173
117
228
4196
1396
6996
16092
7589
24595
1990
36475
24372
48578
6612
3573
9651
1148
847
1450
9508
0
23943
94790
52658
136922
1991
57390
24772
90007
10874
7860
13888
3857
2907
4807
81175
43230
119121
41499
0
83751
1992
970
105
1835
44583
24730
64437
1617
1150
2083
37183
21675
52690
13782
0
36494
1993
330
125
534
38015
15944
60086
1502
911
2092
61508
2885
120131
5458
0
13543
1994
5386
0
10915
21677
11980
31375
1695
825
2566
14884
0
31270
52258
0
121547
1995
862
0
1812
74930
38459
111401
472
269
675
1308
434
2182
11816
3386
20246
1996
44268
22447
66089
66047
42607
89488
1049
782
1316
57169
28040
86299
28
8
47
1997
54802
22682
86922
67061
49487
84634
600
420
780
45808
21160
70455
132
0
272
1998
33841
21406
46277
7050
4209
9890
5964
3800
8128
79492
44207
114778
755
23
1487
1999
85306
45266
125346
1289
135
2442
1137
368
1906
15931
1632
30229
46
14
79
2000
39813
1069
78556
26177
14287
38068
2907
1851
3962
49614
3246
95982
7530
0
16826
2001
33646
0
85901
908
152
1663
1706
1113
2299
844
177
1511
6
1
10
2002
19426
10648
28205
19157
11015
27300
1843
1276
2410
23354
12144
34564
130
20
241
2003
94902
41128
148676
17304
10225
24383
7910
3757
12063
28579
15504
41653
216
0
495
2004
16901
2619
31183
19408
14119
24696
19372
12727
26016
136053
97442
174664
862
0
1779
2005
42354
12517
72192
21789
14947
28631
33637
24645
42630
26531
1288
51774
12676
511
24841
2006
168059
103577
232540
7801
3605
11996
11209
7413
15005
68531
22418
114644
20403
9439
31367
2007
161594
87683
235504
9896
5993
13799
2873
1820
3925
22319
4517
40122
156548
46433
266663
2008
288799
178860
398738
52975
31839
74111
2742
830
4655
15915
4477
27353
9962
0
20827
2009
189747
113135
266360
54579
37311
71846
13040
7988
18093
18916
8249
29582
49939
23435
76443
2010
91730
57545
125914
40635
20307
60962
7268
4530
10006
20367
4099
36636
66392
3114
129669
2011
175836
3876
347796
119736
66423
173048
7441
5251
9631
13674
7737
19610
7026
0
17885
2012
310519
225728
395311
105176
37917
172435
1814
762
2866
26480
299
316769
58535
0
128715
2013
94673
28224
161122
90108
62788
117428
7235
4721
9749
70972
8393
133550
928
310
1547
2014
48933
5599
92267
102977
72975
132980
4185
2217
6153
16674
5671
27677
77658
35010
120306
2015
147961
87971
207951
8744
3008
14479
6005
2816
9194
11207
0
25819
101653
40258
163048
2016
274050
157185
390915
16872
9942
23801
4029
1952
6107
32956
15793
50119
12941
1713
24168
2017
72486
36535
108438
69371
46841
91901
9205
6081
12329
32112
11180
53045
43561
0
97558
Mean
93511
30280
4442
28586
60307
Median
62721
17088
1760
19641
22075
Table 4. 0-group abundance indices (in millions) with 95% confidence limits, not corrected for capture efficiency. These indices have been reported to ICES WG groups (AFWG, WGWIDE and WGIBAR).
Year
Saithe
Gr halibut
Long rough dab
Polar cod (east)
Polar cod (west)
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
1980
3
0
6
111
35
187
1273
883
1664
28958
9784
48132
9650
0
20622
1981
0
0
0
74
46
101
556
300
813
595
226
963
5150
1956
8345
1982
143
0
371
39
11
68
1013
698
1328
1435
144
2725
1187
0
3298
1983
239
83
394
41
22
59
420
264
577
1246
0
2501
9693
0
20851
1984
1339
407
2271
31
18
45
60
43
77
127
0
303
3182
737
5628
1985
12
1
23
48
29
67
265
110
420
19220
4989
33451
809
0
1628
1986
1
0
2
112
60
164
6846
4941
8752
12938
2355
23521
2130
180
4081
1987
1
0
1
35
23
47
804
411
1197
7694
0
17552
74
31
117
1988
17
4
30
8
3
13
205
113
297
383
9
757
4634
0
9889
1989
1
0
3
1
0
3
180
100
260
199
0
423
18056
2182
33931
1990
11
2
20
1
0
2
55
26
84
399
129
669
31939
0
70847
1991
4
2
6
1
0
2
90
49
131
88292
39856
136727
38709
0
110568
1992
159
86
233
9
0
17
121
25
218
7539
0
15873
9978
1591
18365
1993
366
0
913
4
2
7
56
25
87
41207
0
96068
8254
1359
15148
1994
2
0
5
39
0
93
1696
1083
2309
267997
151917
384078
5455
0
12032
1995
148
68
229
15
5
24
229
39
419
1
0
2
25
1
49
1996
131
57
204
6
3
9
41
2
79
70134
43196
97072
4902
0
12235
1997
78
37
120
5
3
7
97
44
150
33580
18788
48371
7593
623
14563
1998
86
39
133
8
3
12
27
13
42
11223
6849
15597
10311
0
23358
1999
136
68
204
14
8
21
105
1
210
129980
82936
177023
2848
407
5288
2000
206
111
301
43
17
69
233
120
346
116121
67589
164652
22740
14924
30556
2001
20
0
46
51
20
83
162
78
246
3697
658
6736
13490
0
28796
2002
553
108
998
51
0
112
731
342
1121
96954
57530
136378
27753
4184
51322
2003
65
0
146
13
0
34
78
45
110
11211
6100
16323
1627
0
3643
2004
1400
865
1936
72
29
115
36
20
52
37156
19040
55271
341
101
581
2005
55
37
74
10
4
15
200
109
291
6545
3202
9888
3231
1283
5178
2006
139
56
221
11
2
21
707
434
979
26016
9997
42036
2112
465
3760
2007
53
6
100
1
0
2
262
46
479
25883
8494
43273
2533
0
5135
2008
45
22
69
6
0
13
956
410
1502
6649
845
12453
91
0
183
2009
22
0
46
7
4
10
115
51
179
23570
9661
37479
21433
5642
37223
2010
402
126
678
14
8
20
128
18
238
31338
13644
49032
1306
0
3580
2011
27
0
59
20
11
29
58
23
93
37431
15083
59780
627
26
1228
2012
69
2
135
30
16
43
173
0
416
4173
48
8298
17281
0
49258
2013
3
1
5
21
13
28
5
0
14
1634
0
4167
148
28
268
2014
1
0
2
10
3
16
309
89
528
2779
737
4820
746
79
1414
2015
47
0
101
27
2
52
575
361
789
128
18
237
6074
2001
10146
2016
3
0
7
6
1
12
601
0
1267
258
0
624
1180
128
2231
2017
127
2
252
8
1
14
72
27
117
43
0
106
1009
0
2795
Mean
161
26
514
30388
7850
Median
54
14
190
9453
3932
Table 5. 0-group abundance indices (in millions) with 95% confidence limits, corrected for capture efficiency. These indices have been reported to ICES WG groups (AFWG, WGWIDE and WGIBAR).
Year
Capelin
Cod
Haddock
Herring
Saithe
Polar cod (east)
Polar cod (west)
Polar cod Tot
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
Confidence limit
Abundance index
1980
740289
495187
985391
276
131
421
265
169
361
77
12
142
21
0
47
203226
69898
336554
82871
0
176632
286097
1981
477260
273493
681026
289
201
377
75
34
117
37
0
86
0
0
0
4882
1842
7922
46155
17810
74500
51037
1982
599596
145299
1053893
3480
2540
4421
2927
2200
3655
2519
0
5992
296
0
699
1443
154
2731
10565
0
29314
12008
1983
340200
191122
489278
19299
9538
29061
6217
3978
8456
195446
69415
321477
562
211
912
1246
0
2501
87272
0
190005
88518
1984
275233
161408
389057
24326
14489
34164
5512
3981
7043
27354
3425
51284
2577
725
4430
871
0
2118
26316
6097
46534
27187
1985
63771
5893
121648
66630
32914
100346
2457
1520
3393
20081
3933
36228
30
7
53
143257
39633
246881
6670
0
13613
149927
1986
41814
642
82986
10509
7719
13299
2579
1621
3537
93
27
160
4
0
9
102869
16336
189403
18644
125
37164
121513
1987
4032
1458
6607
1035
504
1565
708
432
984
49
0
111
4
0
10
64171
0
144389
631
265
996
64802
1988
65127
12101
118153
2570
1519
3622
1661
630
2693
60782
20877
100687
32
11
52
2588
59
5117
41133
0
89068
43721
1989
862394
690983
1033806
2775
1624
3925
650
448
852
17956
8252
27661
10
0
23
1391
0
2934
164058
15439
312678
165449
1990
115636
77306
153966
23593
13426
33759
3122
2318
3926
15172
0
36389
29
4
55
2862
879
4846
246819
0
545410
249681
1991
169455
74078
264832
40631
29843
51419
13713
10530
16897
267644
107990
427299
9
4
14
823828
366924
1280732
281434
0
799822
1105262
1992
2337
250
4423
166276
92113
240438
4739
3217
6262
83909
48399
119419
326
156
495
49757
0
104634
80747
12984
148509
130504
1993
952
289
1616
133046
58312
207779
3785
2335
5236
291468
1429
581506
1033
0
2512
297397
0
690030
70019
12321
127716
367416
1994
13898
70
27725
70761
39933
101589
4470
2354
6586
103891
0
212765
7
1
12
2139223
1230225
3048220
49237
0
109432
2188460
1995
2869
0
6032
233885
114258
353512
1203
686
1720
11018
4409
17627
415
196
634
6
0
14
195
0
390
201
1996
136674
69801
203546
280916
188630
373203
2632
1999
3265
549608
256160
843055
430
180
679
588020
368361
807678
46671
0
116324
634691
1997
189372
80734
298011
294607
218967
370247
1983
1391
2575
463243
176669
749817
341
162
521
297828
164107
431550
62084
6037
118131
359912
1998
113390
70516
156263
24951
15827
34076
14116
9524
18707
476065
277542
674589
182
91
272
96874
59118
134630
95609
0
220926
192483
1999
287760
143243
432278
4150
944
7355
2740
1018
4463
35932
13017
58848
275
139
411
1154149
728616
1579682
24015
3768
44262
1178164
2000
140837
6551
275123
108093
58416
157770
10906
6837
14975
469626
22507
916746
851
446
1256
916625
530966
1302284
190661
133249
248072
1107286
2001
90181
0
217345
4150
798
7502
4649
3189
6109
10008
2021
17996
47
0
106
29087
5648
52526
119023
0
252146
148110
2002
67130
36971
97288
76146
42253
110040
4381
2998
5764
151514
58954
244073
2112
134
4090
829216
496352
1162079
215572
36403
394741
1044788
2003
340877
146178
535575
81977
47715
116240
30792
15352
46232
177676
52699
302653
286
0
631
82315
42707
121923
12998
0
30565
95313
2004
53950
11999
95900
65969
47743
84195
39303
26359
52246
773891
544964
1002819
4779
2810
6749
290686
147492
433879
2892
989
4796
293578
2005
148466
51669
245263
72137
50662
93611
91606
67869
115343
125927
20407
231447
176
115
237
44663
22890
66436
25970
9987
41953
70633
2006
515770
325776
705764
25061
11469
38653
28505
18754
38256
294649
102788
486511
280
116
443
182713
73645
291781
15965
3414
28517
198678
2007
480069
272313
687825
42628
26652
58605
8401
5587
11214
144002
25099
262905
286
3
568
191111
57403
324819
22803
0
46521
213914
2008
995101
627202
1362999
234144
131081
337208
9864
1144
18585
201046
68778
333313
142
68
216
42657
5936
79378
619
25
1212
43276
2009
673027
423386
922668
185457
123375
247540
33339
19707
46970
104233
31009
177458
62
0
132
168990
70509
267471
154687
37022
272351
323677
2010
318569
201973
435166
135355
68199
202511
23669
14503
32834
117087
32045
202129
1066
362
1769
267430
111697
423162
12045
0
33370
279474
2011
594248
58009
1130487
448005
251499
644511
19114
14209
24018
83051
48024
118078
96
0
225
249269
100355
398183
4924
218
9629
254193
2012
988600
728754
1248445
410757
170242
651273
5281
2626
7936
177189
35046
2111493
229
5
453
25026
1132
48920
125306
0
357381
150332
2013
316020
127310
504731
385430
269640
501219
16665
11161
22169
289391
67718
511064
11
4
18
11382
0
29002
1011
262
1760
12393
2014
163630
31980
295280
464124
323330
604919
11765
6160
17371
136305
42164
230447
4
0
9
17349
5184
29515
5298
500
10096
22647
2015
457481
274631
640331
37474
17244
57704
15089
6204
23973
82749
0
190673
406
0
930
795
107
1484
49584
15385
83784
50380
2016
778784
479130
1078438
53796
30970
76622
5504
2791
8216
79439
38415
120464
10
0
21
1544
0
3718
9288
459
18117
10832
2017
213787
112459
315115
233275
150239
316310
19484
12902
26067
153763
34713
272813
379
18
740
64390
48
256
6580
0
18044
70971
LTM
314184
114452
11740
163247
471
252074
65130
317204
8.2 - Indices calculated in MatLab
Table 6. TIBIA indices (spatial abundance indices, for description see the ICES WGIBAR report 2018). See Fig. 1 for geographical location of the polygons.
Cod
South West
Bear Island Trench
Thor Iversen Bank
Hopen Deep
Svalbard South
Svalbard North
South East
Pechora
Southeastern Basin
Central Bank
Great Bank
Franz-Victoria Trough
North East
1980
14
105
17
9
23
77
308
2
1
5
6
0
10
1981
60
76
21
4
40
36
58
9
2
3
6
0
9
1982
681
821
831
84
79
158
280
78
118
140
77
0
286
1983
3834
9218
2158
2378
1963
607
419
261
837
1732
765
0
512
1984
2831
10005
620
277
3207
3450
153
407
241
217
459
0
1888
1985
8493
18649
17373
2496
1768
4114
870
3293
1656
3265
763
0
1702
1986
1924
915
2458
992
350
828
3255
731
458
698
135
377
336
1987
651
142
62
4
17
30
78
11
4
5
18
0
45
1988
1695
193
84
57
35
23
203
28
31
72
52
0
142
1989
220
132
106
289
88
24
884
226
859
135
17
21
177
1990
11078
7475
2794
537
331
172
1681
141
174
416
220
968
564
1991
4695
11160
5986
2011
9673
2687
437
529
3562
2047
346
631
558
1992
8054
89406
22281
15198
27081
9933
1243
8158
3896
5905
2017
5568
4243
1993
8305
52603
6285
27191
12251
23728
1145
1017
670
2707
2138
0
4051
1994
35784
25071
25861
592
1638
1312
5783
1294
1741
1551
1326
0
2724
1995
39488
127900
41212
32416
13179
6412
4924
8697
2971
15622
10903
0
7529
1996
109606
79362
67857
19478
3007
1135
36476
5406
7629
6828
9611
0
3835
1997
109864
90949
72791
33075
2304
1194
40852
3210
8427
20233
6739
0
10934
1998
8241
9301
3688
2357
1580
375
3358
212
353
157
302
0
351
1999
3386
799
712
298
419
135
94
25
44
71
69
0
73
2000
50934
50590
15260
719
3150
1511
4827
3318
4164
999
1625
0
1932
2001
100
436
148
978
566
3285
25
15
30
25
139
0
95
2002
4144
4638
16823
4247
2240
3641
7749
13484
20847
15820
1765
0
2260
2003
21667
35319
15673
2070
3316
3853
173
331
445
1752
1287
0
1507
2004
4068
13373
33009
9136
3233
411
1520
2453
7767
6521
566
2526
857
2005
11557
31115
18420
6372
3130
1095
238
292
381
7543
246
210
692
2006
2257
17266
2165
1088
3105
670
80
245
112
109
57
95
371
2007
23416
18297
4271
638
927
170
392
243
134
560
133
195
432
2008
100687
81327
39620
15985
2162
387
13314
1267
5033
10458
1894
2229
3116
2009
62546
48804
33402
12376
10941
3214
4518
1790
11189
16441
1144
1552
2327
2010
55299
38258
48624
2931
693
236
5952
6106
2243
5514
440
687
2325
2011
60634
80339
56483
49804
79384
80504
6668
1276
1068
25466
3089
1866
5089
2012
113561
53296
187437
10932
8784
2381
40446
16052
33580
10790
1330
1308
6598
2013
14183
24469
86007
47430
124718
11069
4150
5500
22959
66783
31063
4039
13993
2014
107234
99179
200391
57055
21080
3377
2089
2347
4358
89155
5270
11998
11240
2015
11742
3756
9561
7827
1116
1176
2677
5026
1395
10733
429
216
639
2016
1355
235
11134
395
409
1244
725
7694
40237
6791
341
526
10660
Table 7. TIBIA indices (spatial abundance indices, for description see the ICES WGIBAR report 2018). See Fig. 1 for geographical location of the polygons.
Haddock
South West
Bear Island Trench
Thor Iversen Bank
Hopen Deep
Svalbard South
Svalbard North
South East
Pechora
Southeastern Basin
Central Bank
Great Bank
Franz-Victoria Trough
North East
1980
38
118
14
2
58
51
1072
3
1
3
5
0
10
1981
8
35
1
715
10
17
1078
1
1036
1101
1
0
2
1982
327
1182
513
86
111
253
22
56
31
88
68
0
268
1983
1873
2247
537
368
553
236
102
53
37
31
215
0
153
1984
1625
1462
131
66
997
710
12
47
34
38
114
0
390
1985
904
448
598
30
18
166
44
22
11
40
25
0
67
1986
868
512
383
282
48
15
350
54
99
56
33
134
86
1987
353
182
73
9
14
6
9
8
3
8
15
0
35
1988
1052
339
31
11
16
13
13
19
15
12
33
0
90
1989
138
233
104
76
23
27
20
3
5
47
3
7
5
1990
821
1114
446
100
214
80
100
12
12
175
27
97
77
1991
2222
4126
381
89
3886
1501
54
45
10
42
83
166
200
1992
933
2036
1854
260
262
121
98
61
78
240
53
140
113
1993
435
1885
188
970
344
123
5
12
7
177
74
0
111
1994
2609
1115
981
51
141
52
183
42
31
74
65
0
168
1995
242
555
235
266
69
94
4
33
10
83
69
0
40
1996
1416
227
149
19
36
16
130
29
27
12
60
0
31
1997
1183
484
271
139
168
27
120
22
7
85
99
0
86
1998
6910
5457
1602
1969
1527
692
220
82
22
169
152
0
179
1999
1434
732
1041
175
91
147
72
13
26
218
56
0
48
2000
5270
5635
919
105
393
586
216
68
82
101
157
0
194
2001
904
2339
541
238
463
386
20
10
19
54
67
0
76
2002
605
1626
516
414
745
2324
16
19
10
207
133
0
86
2003
7663
15314
2229
479
1089
4835
46
61
28
441
433
0
491
2004
22112
15476
3325
2387
1828
187
2015
217
217
1349
294
1210
457
2005
27758
31640
14462
10324
4741
704
359
311
366
6014
209
276
698
2006
3194
15846
5466
1649
2193
1448
42
271
145
2917
133
521
438
2007
4467
2866
779
386
78
79
59
42
66
324
24
54
86
2008
5858
4253
531
272
92
92
93
31
22
142
31
86
124
2009
16348
10001
4044
1310
804
215
434
167
734
1215
116
245
379
2010
10937
12642
1286
140
104
203
33
90
18
45
55
115
391
2011
4643
6451
1127
1076
2939
2531
38
56
12
416
49
74
217
2012
662
2580
1648
334
191
82
95
41
54
148
16
20
55
2013
4061
3407
1381
892
4613
1713
125
38
49
296
189
177
277
2014
1595
8233
704
143
1349
147
20
44
9
60
57
291
202
2015
2145
5922
3850
5585
584
238
110
66
121
3310
231
80
150
2016
2109
1548
1221
664
234
126
208
77
18
159
41
82
109
2017
8552
6048
1509
758
1841
669
608
270
148
241
102
119
790
Table 8. TIBIA indices (spatial abundance indices, for description see the ICES WGIBAR report 2018). See Fig. 1 for geographical location of the polygons.
Herring
South West
Bear Island Trench
Thor Iversen Bank
Hopen Deep
Svalbard South
Svalbard North
South East
Pechora
Southeastern Basin
Central Bank
Great Bank
Franz-Victoria Trough
North East
1980
18
15
35
2
3
2
1
1596
1
3
2
0
3
1981
1151
2
490
0
13
13
0
2596
419
447
1
0
2009
1982
406
104
110
20
17
17
128
66
48
38
81
0
330
1983
81526
87079
22081
771
7840
5925
3989
1717
1135
1133
2489
0
4212
1984
12914
2402
349
42
307
75
13756
1160
461
84
387
0
1333
1985
7324
2300
116
38
385
196
20
351
346
32
216
0
432
1986
33
21
11
3
3
0
8
2
0
3
1
3
3
1987
48
3
515
1
1314
885
443
954
1211
627
1
0
2
1988
37876
15593
3902
145
2189
3749
1399
745
409
553
1080
0
3578
1989
4937
662
1332
125
2409
1531
5718
349
1130
96
71
196
142
1990
13379
1646
368
57
201
63
1332
196
213
54
128
574
367
1991
62320
123652
21113
2187
13193
19045
1447
841
3696
8229
1191
3103
3372
1992
35154
17679
22565
2311
4092
5047
6661
7300
1035
1484
1300
3355
1890
1993
40044
20483
47686
9024
25873
62637
1380
1183
1885
7553
5655
0
9242
1994
86872
48930
36841
1204
2083
588
1822
1112
584
1343
1683
0
3693
1995
3010
3380
2242
39
2005
736
307
229
1044
152
203
0
256
1996
373641
143713
62166
4324
13345
3385
20896
5092
2992
3106
11385
0
6253
1997
274891
134164
34564
24021
52196
2930
22821
5065
1159
11658
24899
0
20226
1998
117297
182691
166423
103930
12641
3333
36894
4519
14277
17689
7885
0
7581
1999
27826
2592
3995
159
3689
1858
3671
882
3441
176
785
0
728
2000
201135
308221
24741
5107
8166
1934
1015
2148
1447
2108
7296
0
8517
2001
1387
1033
4233
1676
1497
1139
1022
35
216
3941
971
0
311
2002
10266
6416
13498
8486
848
67397
27990
35085
10364
6668
4004
0
3211
2003
43422
89450
11789
10012
2742
4585
1493
396
517
2268
3172
0
3194
2004
591985
93707
99155
31259
5919
2498
24668
3491
8575
49049
4530
18476
7462
2005
2719
30373
83041
5670
565
212
17214
1587
2368
23288
473
393
1055
2006
159046
27844
9887
5862
1640
411
63346
4410
3440
6781
736
1268
4301
2007
109637
58283
6862
303
1201
218
407
731
186
617
757
694
1439
2008
111704
40955
73806
2601
1972
241
22669
932
2257
13705
664
1980
2776
2009
45142
29609
13432
867
3424
5298
5614
818
1277
1566
276
775
1208
2010
39173
11617
68658
3704
417
205
5975
3116
8890
21038
577
616
2236
2011
12779
15358
40843
1715
14451
1954
2897
434
2034
5758
234
320
890
2012
9421
51674
96660
3162
887
1152
4495
569
2503
4391
433
445
1397
2013
98059
107419
90108
19283
19963
1059
3941
881
2367
10482
1427
1067
4949
2014
47341
49054
22133
1079
20780
789
425
678
2820
2841
709
3686
2547
2015
1612
6480
1521
4820
5584
64878
1769
347
83
1656
803
1357
674
2016
3557
3508
21370
31989
2029
1328
15129
5647
19492
14496
792
1536
2591
2017
32094
25604
74644
7661
3392
4243
39586
2619
7120
22677
1065
842
10760
Table 9. TIBIA indices (spatial abundance indices, for description see the ICES WGIBAR report 2018). See Fig. 1 for geographical location of the polygons.
Capelin
South West
Bear Island Trench
Thor Iversen Bank
Hopen Deep
Svalbard South
Svalbard North
South East
Pechora
Southeastern Basin
Central Bank
Great Bank
Franz-Victoria Trough
North East
1980
364057
19370
129488
13978
3758
2857
101385
14751
41147
17258
18113
33470
1981
131645
140672
107016
53307
8333
3296
38752
6861
15247
7021
13815
9798
1982
132237
104173
108715
4864
90832
84577
17285
14751
12054
9951
13296
62035
1983
83436
35909
39443
6132
57022
119756
10863
24418
9222
5519
21425
7880
1984
24047
56188
40605
49989
20949
29477
584
3343
4400
41343
0
24060
1985
1379
8474
24383
4722
13902
2295
467
2663
30189
18497
1904
3205
1986
465
39
696
59
105
94
32401
17473
4451
136
420
1180
1148
1987
164
20
67
4
12
12
1813
1689
369
20
60
148
1988
27671
704
20842
647
210
147
3848
2139
17246
1417
1523
2779
1989
126506
163575
141517
93926
25478
10234
140295
20742
124670
61079
20491
15242
14456
1990
5868
4218
29128
38217
9220
344
1882
6835
6455
22492
1989
0
2948
1991
2565
31482
21068
55350
6162
14659
1850
39458
30821
16634
6865
1276
6258
1992
4
2
13
3
10
4
88
4801
184
30
20
52
107
1993
7
5
23
183
162
3
22
209
617
88
297
467
1994
186
35
220
12
74
58
2279
14807
1590
49
227
517
1995
58
14
537
11
16
11
66
6509
3122
1201
78
5124
1996
8944
18713
12424
54779
7436
580
2109
27808
10671
25741
678
2412
1997
9479
77357
76240
34157
25159
1291
1236
21153
4541
21493
0
9961
1998
1541
2551
19546
21383
6923
781
18067
12252
25675
25269
10206
2496
1999
7448
26474
27599
94005
15695
1256
1442
45430
23226
93421
81672
10394
2000
2104
38763
10218
44904
47327
8013
327
4521
1073
15692
11981
3556
2001
1111
4549
6367
1181
8089
315
3675
103599
2583
4888
885
1918
2002
219
45
2267
753
278
509
3986
22367
42700
1590
1105
23779
2003
12980
80026
7903
84031
181125
155421
343
1797
1783
12401
5320
7190
2004
560
8618
664
2772
9806
213
6172
4091
1322
27994
1660
0
3351
2005
505
1592
41975
32251
1166
268
635
3346
19786
114292
3783
653
9150
2006
6009
86042
134696
127174
71731
62415
5899
21063
39835
194431
38931
5556
25538
2007
4182
5599
78865
76100
5944
432
13247
7522
94184
213841
202201
17158
30967
2008
80336
156283
165986
234694
129445
998
126790
28499
161568
87937
43054
19621
16258
2009
18867
9497
136231
7702
1109
668
90446
230294
196396
142990
5122
5626
19561
2010
22033
8776
51375
80889
5073
347
13991
59533
85162
50067
4046
2393
32136
2011
23430
101645
7853
66186
327507
2426
114432
47185
35867
14342
11168
3769
11343
2012
128379
109680
112339
154320
80582
1574
32590
67242
110011
204616
51967
4194
83382
2013
14984
33424
40398
17442
3185
527
57468
9943
30800
18902
13835
96970
60037
2014
4424
2256
10236
12352
4691
342
21403
11851
14246
111662
31741
17991
24530
2015
95523
57795
55254
67510
7843
832
20493
9549
22958
174445
20572
2764
37988
2016
13236
55564
101165
170358
200812
44444
18323
55396
20403
176653
53015
23308
46963
2017
9625
12616
996
43687
47318
2181
12779
4024
5814
3717
90039
6629
33622
Table 10. TIBIA indices (spatial abundance indices, for description see the ICES WGIBAR report 2018). See Fig. 1 for geographical location of the polygons.
Polar cod
South West
Bear Island Trench
Thor Iversen Bank
Hopen Deep
Svalbard South
Svalbard North
South East
Pechora
Southeastern Basin
Central Bank
Great Bank
Franz-Victoria Trough
North East
1980
1059
691
2010
359
25200
70776
3182
402493
66633
1786
2916
0
31617
1981
160
313
187
2218
53535
4795
44
1791
466
232
2945
0
16132
1982
11
22
13
22
1130
19923
21
4364
128
83
309
0
1932
1983
93
186
13
113
4360
81925
45
3289
201
256
1651
0
2398
1984
214
572
75
449
32973
10106
34
1563
283
162
603
0
2865
1985
296
163
904
211
15640
1854
956
219598
15787
2666
3292
0
27873
1986
161
710
202
689
38668
9807
537
92519
35368
262
6189
55686
234492
1987
36
13
55
22
596
648
260
227020
707
262
850
0
2962
1988
105
955
56
650
29169
12734
17
6811
214
353
8296
0
6126
1989
539
4464
285
11148
139423
30072
43
2426
135
878
4200
17784
1460
1990
569
763
129
3339
168245
119581
76
3527
84
341
5928
81795
15758
1991
1222
4304
1298
20097
291732
4333
2736
679190
26712
3042
65316
229395
70623
1992
663
5033
174
638
30695
45903
252
76429
1266
548
1609
7402
12074
1993
849
478
739
444
36069
88545
934
487154
25248
1409
7511
0
312687
1994
1807
847
638
237
12575
2608
941
1427208
140936
716
8405
0
831542
1995
1
1
0
2
41
365
4
24
1
3
32
0
44
1996
1999
2979
733
3946
140281
7550
1621
409651
185865
1705
15803
0
541952
1997
976
2004
196
1086
163485
12405
2225
402750
9831
1388
7601
0
97562
1998
765
1708
158
612
87848
43166
192
80580
13851
591
7636
0
84570
1999
1889
555
1174
1497
23754
53460
1542
1114824
106162
2929
22481
0
244323
2000
1410
4899
660
5510
281318
13505
1083
446544
73195
2136
15733
0
654006
2001
233
525
57
270
24232
171716
122
29364
348
142
2787
0
7025
2002
1060
1664
742
1676
108699
188194
1416
540371
46915
1178
10147
0
197939
2003
152
51
2483
221
2590
39582
218
35127
21740
7428
2566
0
125724
2004
649
104
328
200
3751
2669
490
322929
20269
284
2438
16014
87158
2005
59
263
46
238
15306
4727
38
25272
550
593
5260
4834
41523
2006
395
70
92
403
1791
1317
274
230604
4760
578
37889
10892
50832
2007
222
88
76
31
7250
12064
299
230416
774
195
1363
1268
129501
2008
47
6
39
10
353
285
63
38481
742
89
187
454
40875
2009
472
127
178
158
4056
119499
152
132900
11445
207
1258
4792
128138
2010
311
61
390
307
7788
376
258
220614
22075
469
1341
1763
153662
2011
290
112
758
276
6980
245
310
134038
8363
2045
2136
1746
250135
2012
192
466
74
161
3699
50191
73
9844
4557
139
592
1945
34284
2013
46
16
18
7
23
54
32
14379
295
10
63
950
587
2014
37
180
29
1387
3893
69
30
17948
1147
57
996
2130
4899
2015
77
559
38
173
16399
9887
24
577
52
74
2186
23051
855
2016
212
1276
48
729
8826
11370
228
203
266
45
383
1290
3206
2017
12
33
9
121
4458
253
6
382
26
16
273
1230
174
Table 11. TIBIA indices (spatial abundance indices, for description see the ICES WGIBAR report 2018). See Fig. 1 for geographical location of the polygons.
Redfish
South West
Bear Island Trench
Thor Iversen Bank
Hopen Deep
Svalbard South
Svalbard North
South East
Pechora
Southeastern Basin
Central Bank
Great Bank
Franz-Victoria Trough
North East
1980
3279
7631
1968
176
19944
139044
116
1657
726
1308
3177
0
6564
1981
5457
11813
304
1528
133417
8471
98
2225
210
611
3069
0
3517
1982
28644
26794
649
282
6883
2537
264
1538
632
461
1869
0
7662
1983
25543
64300
494
659
41303
3545
1676
1410
357
558
2889
0
4073
1984
13706
1708
76
42
2575
1338
45
421
301
186
1018
0
3536
1985
6769
34507
586
589
20750
195800
170
4301
919
711
6329
0
13674
1986
41955
52036
6686
713
4064
6530
342
887
234
857
1342
3210
2918
1987
11053
9935
60
69
407
249
40
258
64
111
467
0
1087
1988
19695
21508
1342
2373
10042
20357
2291
1349
628
1060
2871
0
6878
1989
3118
2360
523
66
722
5402
609
50
51
40
72
221
139
1990
13521
18884
2216
373
25169
23422
63
285
78
120
769
3456
2137
1991
1668
2501
46
165
14412
4119
12
122
11
17
179
545
553
1992
20
75
7
6
394
10542
10
141
15
43
156
403
299
1993
11
19
2
3
88
4438
4
22
9
21
114
0
198
1994
3944
11547
120
80
4261
12948
32
312
47
139
602
0
1446
1995
2441
2433
37
28
1425
598
15
210
28
42
274
0
391
1996
16
6
3
0
2
5
0
0
0
0
1
0
0
1997
145
4
2
1
2
5
2
2
0
1
6
0
7
1998
61
731
76
21
35
58
1
4
1
25
9
0
10
1999
17
6
1
1
12
8
0
0
0
0
1
0
1
2000
3652
5549
391
111
207
2166
36
34
15
73
124
0
132
2001
0
0
0
0
7
1
0
0
0
0
0
0
0
2002
0
0
0
0
2
146
0
1
0
0
2
0
3
2003
34
1
1
0
1
182
0
0
0
0
2
0
3
2004
35
720
12
16
34
222
1
4
1
3
6
12
10
2005
647
12430
916
62
468
225
15
34
7
13
10
38
87
2006
1466
7544
735
120
4943
4000
14
182
29
219
51
165
304
2007
51685
103056
10833
10604
1225
3317
124
754
170
3979
536
713
1534
2008
4224
3759
141
81
622
270
9
28
8
46
31
95
125
2009
1905
6981
2005
669
3764
31543
53
291
58
528
231
632
1048
2010
34467
31929
2346
400
776
2276
71
244
64
177
172
322
1128
2011
279
696
16
135
6934
576
3
20
2
11
94
94
78
2012
3271
41675
13845
2954
5010
7904
195
237
371
2058
251
237
693
2013
920
56
120
5
15
25
8
3
8
6
3
4
17
2014
6128
21389
107
229
12071
36182
36
297
22
44
404
1907
1410
2015
7332
9943
409
5334
10212
57542
103
336
43
263
289
514
857
2016
491
1057
385
9595
2901
510
325
132
52
182
152
321
247
2017
290
1532
19
34
4094
57588
77
752
214
14
242
304
2197
8.3 - Abundance indices calculated in SAS (1980-2017) and R (2018-2023)
Table 12. The modern official 0-group abundance indices (corrected for capture efficiency, KEFF) calculated in SAS (1980-2017) and R (since 2018). Abundance is given as total number of individuals in millions. Redfish abundance is given as both uncorrected numbers and numbers corrected with a KEFF of 3.8. These indices have been reported to the ICES WG groups (AFWG, WGWIDE and WGIBAR).
Years
Cod
Haddock
Capelin
Polar cod
Herring
Redfish
Redfish_KEFF
1980
276
265
740289
286097
77
277873
1055917
1981
289
75
477260
51037
37
153279
582460
1982
3480
2927
599596
12008
2519
106140
403332
1983
19299
6217
340200
88518
195446
172392
655090
1984
24326
5512
275233
27187
27354
83182
316092
1985
66630
2457
63771
149927
20081
412777
1568553
1986
10509
2579
41814
121513
93
91621
348160
1987
1035
708
4032
64802
49
23747
90239
1988
2570
1661
65127
43721
60782
107027
406703
1989
2775
650
862394
165449
17956
16092
61150
1990
23593
3122
115636
249681
15172
94790
360202
1991
40631
13713
169455
1105262
267644
41499
157696
1992
166276
4739
2337
130504
83909
13782
52372
1993
133046
3785
952
367416
291468
5458
20740
1994
70761
4470
13898
2188460
103891
52258
198580
1995
233885
1203
2869
201
11018
11816
44901
1996
280916
2632
136674
634691
549608
28
106
1997
294607
1983
189372
359912
463243
132
502
1998
24951
14116
113390
192483
476065
755
2869
1999
4150
2740
287760
1178164
35932
46
175
2000
108093
10906
140837
1107286
469626
7530
28614
2001
4150
4649
90181
148110
10008
6
23
2002
76146
4381
67130
1044788
151514
130
494
2003
81977
30792
340877
95313
177676
216
821
2004
65969
39303
53950
293578
773891
862
3274
2005
72137
91606
148466
70633
125927
12676
48168
2006
25061
28505
515770
198678
294649
20403
77531
2007
42628
8401
480069
213914
144002
156548
594884
2008
234144
9864
995101
43276
201046
9962
37857
2009
185457
33339
673027
323677
104233
49939
189768
2010
135355
23669
318569
279474
117087
66392
252289
2011
448005
19114
594248
254193
83051
7026
26698
2012
410757
5281
988600
150332
177189
58535
222432
2013
385430
16665
316020
12393
289391
928
3528
2014
464124
11765
163630
22647
136305
77658
295099
2015
37474
15089
457481
50380
82749
101653
386283
2016
53796
5504
778784
10832
79439
12941
49175
2017
233275
19484
213787
70971
153763
43561
165530
2018
79914
9313
657267
112278
57572
60889
60889
2019
67629
2460
1630176
196153
48254
240553
240553
2020
19740
6314
1811118
429165
11636
799276
799276
2021
206432
25387
364610
135895
205373
85864
85864
2022
71863
20988
152869
352153
2015797
72597
72597
2023
231081
50403
574441
7614233
3624101
101995
101995
LTM
116924
12926
387024
469259
276287
83019
228852
Red font identifies years with lack of coverage of the eastern Barents Sea.
8.4 - Biomass indices calculated in SAS (1980-2017) and R (2018-2023)
Table 13. The modern official 0-group biomass indices (thousand tonnes) calculated in SAS (1980-2017) and R (since 2018). These indices have been reported to ICES WG groups (AFWG, WGWIDE and WGIBAR).
Year
Capelin
Cod
Haddock
Herring
Polar cod
Redfish
Total biomass
1980
92
0
1
0
124
356
574
1981
61
0
0
0
22
197
281
1982
117
5
14
3
5
136
280
1983
86
54
47
554
38
221
1000
1984
100
60
37
132
12
107
447
1985
24
158
16
69
65
611
943
1986
6
16
12
0
53
122
208
1987
1
1
2
0
28
12
44
1988
16
4
9
105
19
161
315
1989
183
8
4
47
72
21
335
1990
32
84
31
69
108
210
536
1991
68
131
88
820
480
49
1635
1992
2
533
42
340
57
28
1002
1993
0
478
40
686
159
4
1367
1994
9
292
45
172
950
102
1571
1995
1
1053
13
16
0
19
1102
1996
48
795
29
742
276
0
1890
1997
37
656
15
629
156
0
1493
1998
26
88
161
947
84
1
1307
1999
67
18
31
156
511
0
784
2000
25
274
66
659
481
9
1514
2001
56
9
47
11
64
0
187
2002
15
215
53
397
454
0
1132
2003
56
159
188
458
41
0
902
2004
16
245
525
1997
127
1
2911
2005
37
287
859
295
31
22
1530
2006
175
112
309
782
86
33
1498
2007
173
100
74
279
93
362
1081
2008
197
554
72
210
19
8
1060
2009
119
738
346
243
140
168
1755
2010
65
548
174
233
121
213
1355
2011
136
1449
201
156
110
11
2063
2012
304
1193
45
295
65
109
2012
2013
77
895
202
871
5
1
2051
2014
42
987
107
218
10
151
1514
2015
151
99
160
137
22
160
729
2016
358
245
116
275
5
39
1039
2017
87
845
235
359
31
91
1649
2018
164
80
9
58
56
20
387
2019
526
132
22
28
95
69
872
2020
453
20
6
12
215
266
972
2021
577
365
172
95
138
20
1367
2022
38
72
21
2016
176
24
2347
2023
233
440
367
1587
681
31
3339
Mean
115
330
114
390
147
95
1140
9 - References
Anonymous 1980. Preliminary report of the international 0-group fish survey in the Barents Sea and adjacent waters in August–September 1980. Annal Biol, Cons Int Explor Mer 37: 259-266
Anonymous 2004. Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea in August-October 2004 (vol.2). IMR/PINRO Joint Report Series, No. 1/2005. ISSN 1502-8828.
Anonymous 2005. Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-October 2005. Volume 1, IMR/PINRO Joint Report Series, No. 3/2005. ISSN 1502-8828. 99 pp.
Anonymous 2006. Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-October 2006. IMR/PINRO Joint Report Series, No. 2/2006. ISSN 1502-8828, 97 pp
Anonymous 2009. Survey report from the joint Norwegian/Russian ecosystem survey in the Barents Sea August-September 2009. IMR/PINRO Joint Report Series, No. 2/2010. ISSN 1502-8828. 118 pp.
Boitsov VD, Dolgov AV, Krysov AI, Seliverstova EI, Shevelev MS (eds) (2013) Polar cod of the Barents Sea. PINRO Press, Murmansk (in Russian)
Dingsør G.E. 2005. Estimating abundance indices from the international 0-group fish survey in the Barents Sea. Fish Res 72:205-218
Dragesund, O. and Olsen, S. 1965. On the possibility of estimating yearclass strength by measuring echo-abundance of 0-group fish. Reports on Norwegian fishery and marine investigations. Fiskeridirektoratet, Bergen 1965, 48-75.
Dragesund, O. (ed.). 1970. International O-group fish surveys in the Barents Sea 1965-1968. Coop.res.rep.int.Coun.Explor. Sea, 1970, Ser. A(18): 1-81
Dragesund, O., Hylen, A., Olsen, S., Nakken, O. 2008. The Barents Sea 0-group surveys; a new concept of pre-recruitment studies. In: Nakken, O. (ed.) Norwegian spring-spawning herring and Northeast Arctic cod – 100 Years of Research and Management, Tapir Academic Press, Trondheim, pp 119-136
Eriksen, E., Prozorkevich, D., Dingsør, G., 2009. An evaluation of 0-group abundance indices of Barents Sea fish stocks, The Open Fish Science Journal, 2: 6-14
Eriksen, E. and Prozorkevich, D. 2011. 0-group survey. In: Jakobsen T and Ozhigin, V. (ed) The Barents Sea ecosystem, resources, management. Half a century of Russian-Norwegian cooperation. Tapir Academic Press, Trondheim, pp 557-569.
Eriksen, E., Bogstad, B., Nakken, O. 2011. Ecological significance of 0-group fish in the Barents Sea ecosystem. Polar Biology 34:647–657, https://doi.org/10.1007/s00300-010-0920-y
Eriksen, E., Ingvaldsen, R., Stiansen, J.E., and Johansen, G.O. 2012. Thermal habitat for 0-group fishes in the Barents Sea; how climate variability impacts their density, length and geographical distribution. ICES Journal of Marine Science, 69(5): 870–879, https://doi.org/10.1093/icesjms/fsr210
Eriksen, E. and Gjøsæter, H. (ed) 2013. A monitoring strategy for the Barents Sea - Report from project no. 14256 Survey strategy for the Barents Sea. Bergen: Havforskningsinstituttet, 73 pp.
Eriksen, E., Durif, C.M.F., and Prozorkevich, D. 2014. Lumpfish (Cyclopterus lumpus) in the Barents Sea: development of biomass and abundance indices, and spatial distribution. ICES Journal of Marine Science; doi:10.1093/icesjms/fsu059
Eriksen, E., Skjoldal, H.R. Gjøsæter, H. and Primicerio R. 2017. Spatial and temporal changes in the Barents Sea pelagic compartment during the recent warming. Progress in Oceanography 151: 206-226, http://dx.doi.org/10.1016/j.pocean.2016.12.009
Eriksen, E., Gjøsæter, H., Prozorkevich, D., Shamray, E., Dolgov, A., Skern-Mauritzen, M., Stiansen, J.E., Kovalev, Yu., Sunnanaa K. 2018. From single species surveys towards monitoring of the Barents Sea ecosystem. Progress in Oceanography, 166: 4-14, http://dx.doi.org/10.1016/j.pocean.2017.09.007
Folmer, O., & Pennington, M. (2000). A statistical evaluation of the design and precision of the shrimp trawl survey off West Greenland. Fisheries Research , 49 (2), 165-178.
Godø, O.R., Valdemarsen, J.W., Engås, A. 1993. Comparison of efficiency of standard and experimental juvenile gadoid sampling trawls, ICES Mar Sci 196:196-201
Haug A and Nakken O. Echo abundance indices of 0-group fish in the Barents Sea 1965–1972. Rapports et Procés-verbaux des Réunions du Conseil International pour l’Exploration de la Mer, 1977; 170: 259–264.
Hylen, A., Korsbrekke, K., Nakken, O. and Ona, E. 1995. Comparison of the capture efficiency of O-group fish in pelagic trawls. Pp. 145-156 in Hylen, A.(ed.): Precision and relevance of prerecruit studies for fishery management related to fish stocks in the Barents Sea and adjacent waters. Proceedings of the sixth IMR-PINRO symposium, Bergen, 14-17 June 1994. Institute of Marine Research, Bergen, Norway.
Hussy K, Mosegaard H, Hinrichsen HH, Bottcher U. 2003. Using otolith microstructure to analyse growth of juvenile Baltic cod Gadus morhua. Mar Ecol Prog Ser 258:233-241
ICES 1965. Preliminary Report of the joint Soviet-Norwegian investigations in the Barents Sea and adjacent waters September 1965. ICES CM documents;1965 No. 161
ICES WGIBAR 2018. Interim Report of the Working Group on the Integrated Assessments of the Barents Sea (WGIBAR), including Annex 5: the state and trends of the Barents Sea ecosystem in 2017. ICES Expert Group reports (until 2018). Report. https://doi.org/10.17895/ices.pub.8267
Mamylov, V. S., 1999: Methodical aspects of trawl-acoustic surveys on redfish stock in the Irminger Sea. Work.Doc. 3 in Report of the study group on redfish stocks. ICES CM 1999/G:9.
Mamylov, V.S. 2004. About the comparison of fish distribution densities estimated using trawl and acoustic methods. In: Chernook, V. (ed) Improvement of instrumental methods for stock assessment of marine organisms, Proceedings of the Russian-Norwegian Workshop, Murmansk 2003. PINRO Press, Murmansk, pp 114-32
Nakken, 0., Hylen, A. and Ona, E. 1995. Acoustic estimates of O-group fish abundance in the Barents Sea and adjacent waters in 1992 and 1993. Pp. 187-197 in Hylen, A. (ed): Precision and relevance ofprerecruit studies for fishery management related to fish stocks in the Barents Sea and adjacent waters. Proceedings ofthe ixth IMRPINRO symposium, Bergen, 14-17 June 1994. Institute ofMarine Research, Bergen, Norway.
Nakken, O. and Raknes, A. 1996. Corrections of indices of abundance of 0-group fish in the Barents Sea for varying capture efficiency. ICES Document CM 1996/G:12. 10 pp.
Ozhigin V.K., Tretyak V.L., Yaragina N.A., and V.A. Ivshin. 1999. Oceanographic conditions of the Barents Sea and their effect on survival and development of juveniles of North-East Arctic cod. – Murmansk: PINRO Press, 88 pp
Randa K. 1984. Abundance and distribution of 0-group Arcto-Norwegian cod and haddock 1965–1982. In: Godø, O.R. and Tilseth, S., Reproduction and Recruitment of Arctic Cod, Proceedings of the First Soviet-Norwegian Symposium, Leningrad, 1983. Institute of Marine Research, Bergen, Norway,1984; pp. 189–209.
R Core Team (2023) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
Pennington, M. 1996. Estimating the mean and variance from highly skewed marine data. Fishery Bull., 94 (3) (1996), pp. 498-505
Pennington, M. and Helle, K. 2013. Evaluation of the sampling strategy for the Norwegian-Russian 0-group component of the ecosystem summer survey. Proceedings of the 16th Norwegian-Russian Symposium, Sochi, 10–12 September 2013: Assessments for Management of Living Marine Resources in the Barents Sea and Adjacent Waters – a Focus on Methodology (2013), pp. 45-52
Prozorkevich, D.V. 2010. Instrumental assessment of fish abundance at early stages of ontogenesis in large marine ecosystems (in Russian). Rybnoje khozyjstvo. 4: 49-53.
Prozorkevich D.V. 2012. Stock formation of the main Barents Sea fish species at the early stages of their development: Cand. Sci. (in Russian) Petrozavodsk, 166 с.
Prozorkevich, D. and Eriksen, E. 2013. Significance of cod settlement on 0-group cod abundance indices. Proceedings of the 16th Norwegian-Russian symposium, Sochi, 10-12 September 2013: Assessments for management of living marine resources in the Barents Sea and adjacent waters – a focus on methodology, pp 34-39
Toresen, R. 1985. Recruitment indices of Norwegian springspawning herring for the period 1965–1984 based on the International 0-group Fish Surveys. ICES CM 1985/H: 54
Valdemarsen, J.W and Misund, O.A. 1995.Trawl designs and techniques used by Norwegian research vessels to sample fish in the pelagic zone. In Hylen, A. (ed) Proceedings of the sixth PRlW-PEW0 symposium, Bergen, 14-17 June 1994. Institute of Marine Research, Bergen, Norway. ISBN 82-7461-039-3.