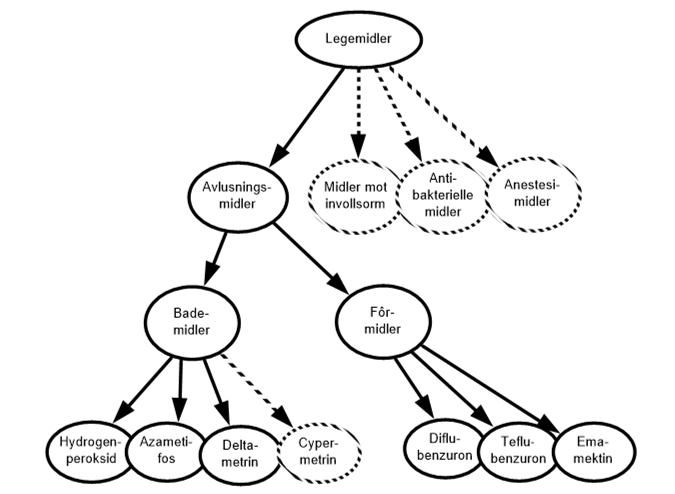
Havforskningsinstituttet har siden 2011 gitt ut en årlig rapport for risikovurdering av miljøeffekter knyttet til norsk fiskeoppdrett. Frem til 2018 var risikorapporten både risikovurdering og kunnskapsstatus i ett. For å tydeliggjøre risikovurderingen har vi fra 2019 valgt å publisere to delrapporter under rapportserien «Risikorapport norsk fiskeoppdrett», en som omfatter selve risikovurderingen, «Risikovurdering - miljøeffekter av norsk fiskeoppdrett» og en som presenterer et oppdatert kunnskapsgrunnlag for risikovurderingen, «Kunnskapsstatus - miljøeffekter av norsk fiskeoppdrett».
Årets risikovurdering omfatter effekter på vill laksefisk, utslipp fra anlegg, fiske og bruk av leppefisk i lakseoppdrett, samt velferd hos oppdrettsfisk i merd i sjø. Kunnskapsgrunnlaget for denne vurderingen utdypes her for de enkelte kapitlene. Under ligger en kort oppsummering av hvert kapittel.
Ytterligere genetisk endring hos villaks som følge av rømt oppdrettslaks
Hvert år rømmer det titusenvis av laks fra norske oppdrettsanlegg, og i enkelte år har antall rapporterte rømt oppdrettslaks vært høyere enn antall voksen villaks som returnerer til elvene for å gyte. De fleste oppdrettslaks som rømmer, forsvinner i havet. Det er sannsynlig at de dør av sult, sykdom, eller blir spist av predatorer. Likevel overlever noen etter rømming, og flere tusen vandrer opp i elvene hvert år og utgjør en risiko for innkryssing og genetisk påvirkning av villaksen. Over tid vil innkryssing av rømt oppdrettslaks kunne forandre egenskapene til de ville laksebestandene, redusere antall villaks som produseres og svekke bestandenes evne til å tilpasse seg endringer i miljøet.
Miljøeffekter som følge av utslipp av løste næringssalter fra fiskeoppdrett
Overgjødsling (eutrofiering) grunnet økte konsentrasjoner av næringssalter i kystvannet, med påfølgende økt produksjon av planteplankton, kan medføre store negative endringer i økosystemet. Økt produksjon av planteplankton fører til økt mengde dyreplankton, økt nedfall til bunn med konsekvenser for oksygenkonsentrasjoner i bunnvann og effekter på dyresamfunn i sedimentene og makroalgesamfunn i fjæresonen. Overgjødsling av kystvann er kjent tidligere fra skagerrakområdet og fra mange deler av verden, mens mesteparten av norskekysten er vist å ha relativt lavt innhold av næringssalter. Norske kystvannsområder er i hovedsak nitrogenbegrensede, det vil si at i sommerhalvåret er det lite nitrogen i vannet.
Miljøpåvirkning på bunn som følge av partikulære organiske utslipp fra fiskeoppdrett
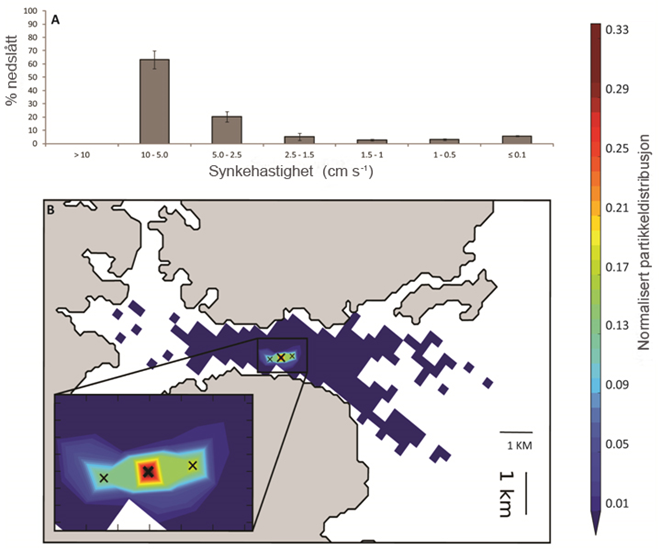
Oppdrettsfisk i Norge produseres i all hovedsak i åpne merdanlegg, og det slippes ut organiske partikler direkte til miljøet i form av fekalier fra fisken, og fôr som ikke spises. Utslippene spres eller akkumuleres på eller i sedimentet, og vil påvirke miljøet i større eller mindre grad rundt oppdrettsanlegget. Utslippene av organiske partikler fra fiskeoppdrett er høye, og påvirkningen på bunnen kan bli stor under produksjonen. Utslippene består imidlertid hovedsakelig av lett nedbrytbare forbindelser, påvirkningen er reversibel, og regenerering av bunnen vil kunne vare fra noen måneder til noen år. For bløtbunn er det satt grenser for hvor stor bunnpåvirkning som aksepteres fra partikulære organiske utslipp både under anleggene og i områdene rundt. For blandingsbunn og hardbunn finnes ennå ingen standardisert overvåkning med grenseverdier.
Miljøeffekter på non-target-arter ved bruk av legemidler
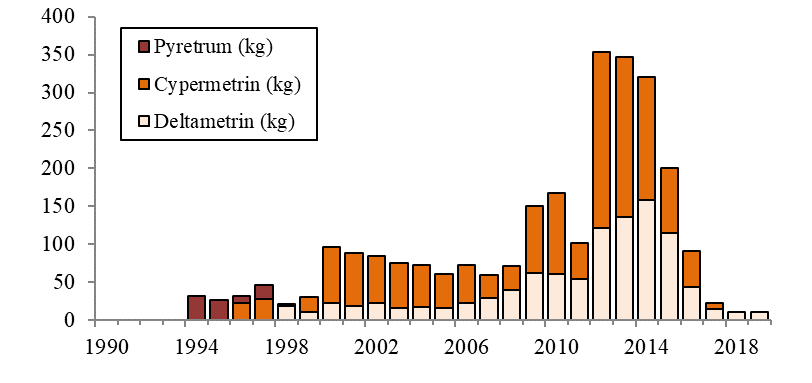
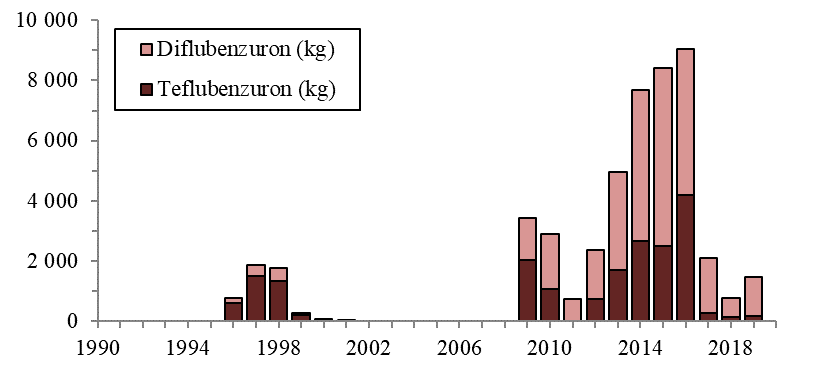
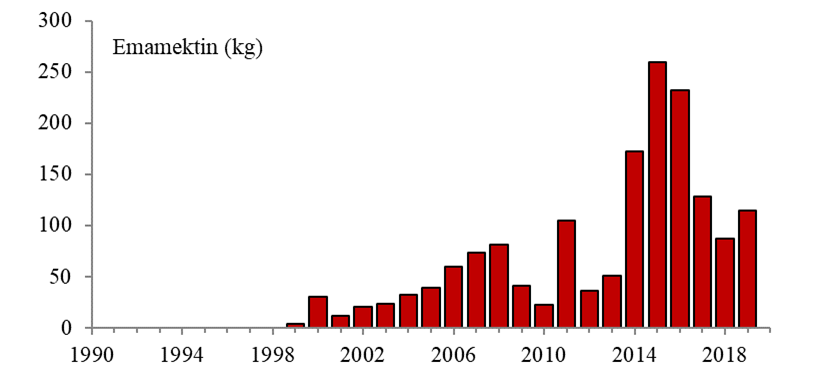
Lakselus tilhører dyregruppen krepsdyr, og medikamenter som dreper lakselusen kan også påvirke andre krepsdyrarter og også andre arter. Andre arter enn lakselus er gitt en generell betegnelse som «non-target-arter», og omfatter arter som lever fritt i vannmassene, på bunnen og i strandsonen. Det er forskjell i måten bademidler og fôrmidler påvirker non-target-arter. Bademidler gir kortvarig effekt, mens fôrmidler vil kunne påvirke non-target-arter over en lengre tidsperiode. Hvilke avlusningsmiddel som brukes, vil også ha stor betydning, siden de ulike avlusningsmidlene har varierende giftighet og effekt på ulike arter og livsstadier.
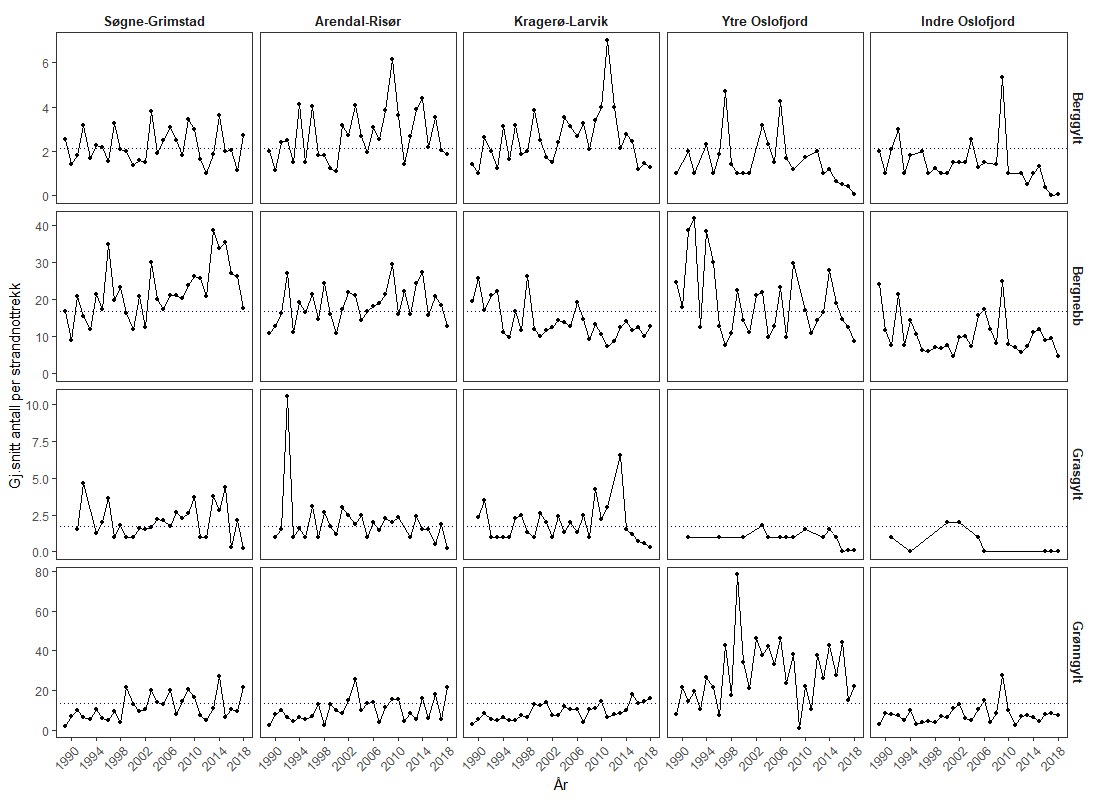
Miljøeffekter ved bruk av villfanget leppefisk i fiskeoppdrett
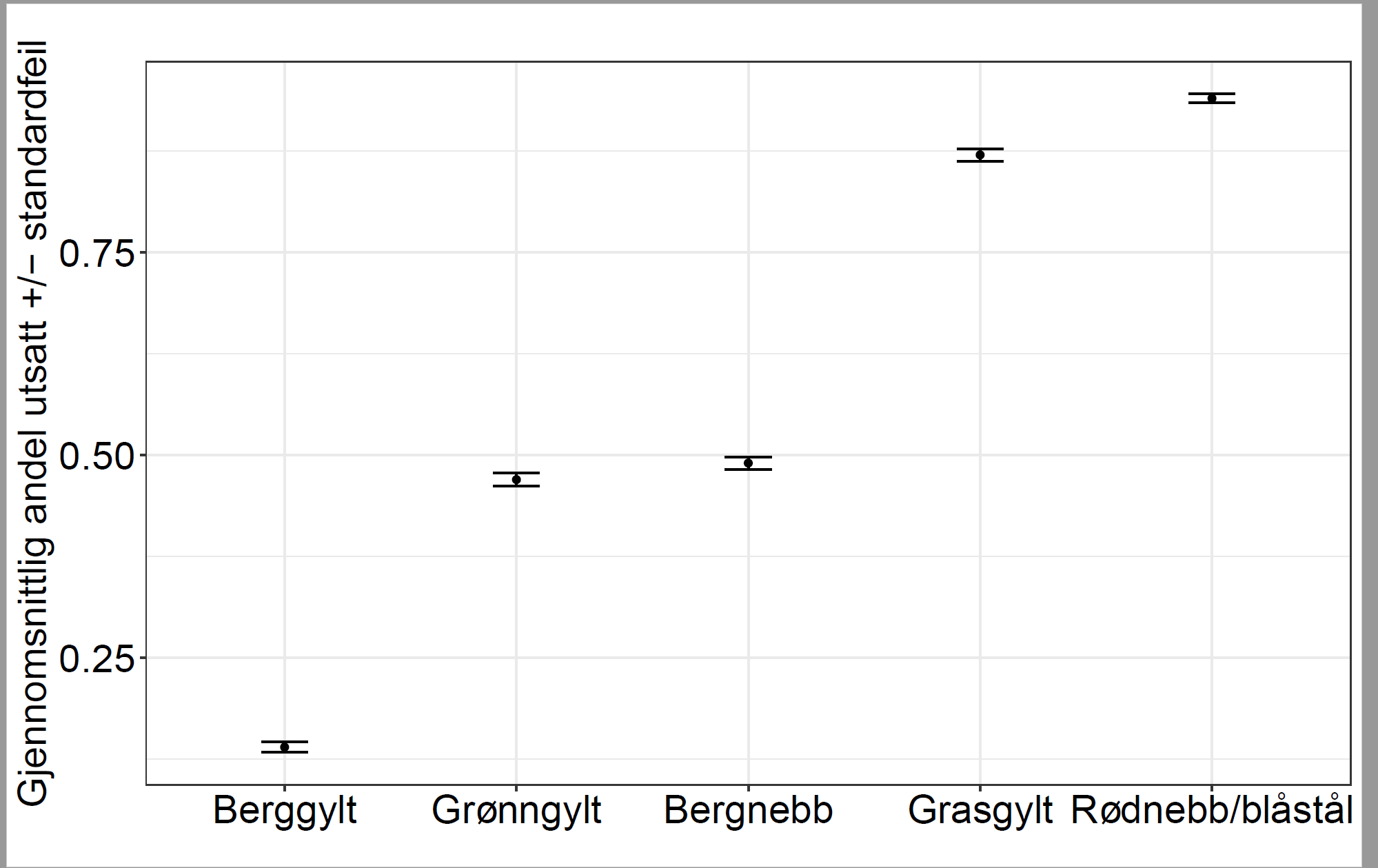
Leppefisk og rognkjeks brukes som rensefisk for å bekjempe lakselus hos laksefisk i oppdrett, og er regnet som et miljøvennlig alternativ til bruk av legemidler. Rensefisk påfører også oppdrettsfisken mindre stress enn avlusningsmetoder som krever håndtering. De vanligste artene som brukes til avlusning er berggylt, grønngylt, bergnebb, gressgylt og rognkjeks. All rognkjeks som brukes som rensefisk, kommer fra oppdrett. Det er et mål at fiske og bruk av rensefisk ikke skal ha vedvarende negative miljøeffekter. Dette innebærer ingen permanent endring i genetisk struktur, ikke endret helsestatus og ingen uønskede, langvarige eller permanente økosystemendringer.
I norske oppdrettsmerder er det til enhver tid 300–400 millioner oppdrettslaks og flere titalls millioner rensefisk. Dette er individer som etter dyrevelferdsloven har krav på å bli holdt i et miljø som gir god velferd ut fra artstypiske og individuelle behov, og mulighet for stimulerende aktivitet, bevegelse, hvile og annen naturlig atferd. Oppdretterne må også sikre at fôret er av god kvalitet og dekker fiskens behov, at oppdrettsfisken er beskyttet mot skade, sykdom og andre farer. Oppdrettsfisken må være robust nok og ha forutsetninger for å tåle oppdrettsforholdene, og de skal ikke utsettes for unødige påkjenninger og belastninger.
1.1 - Status og miljømessig bærekraft i norsk fiskeoppdrett
1.1.1 - Status norsk fiskeoppdrett
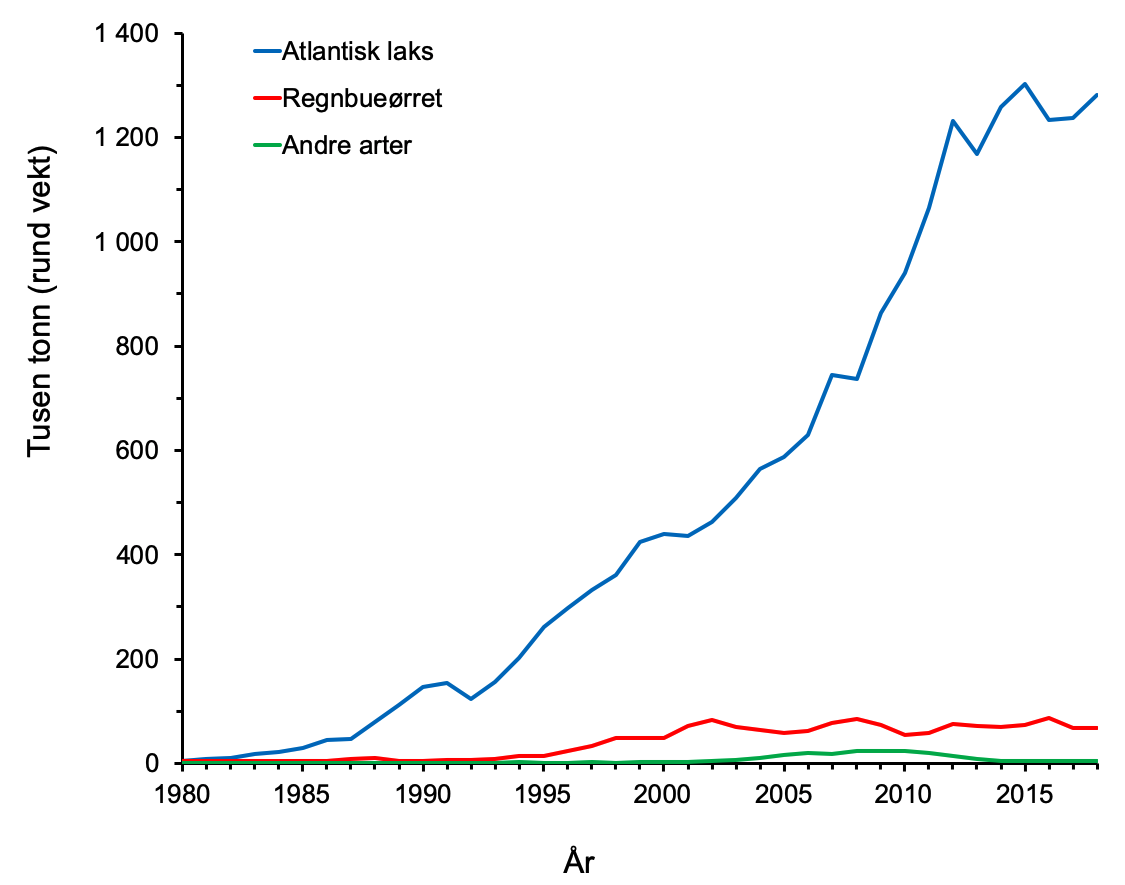
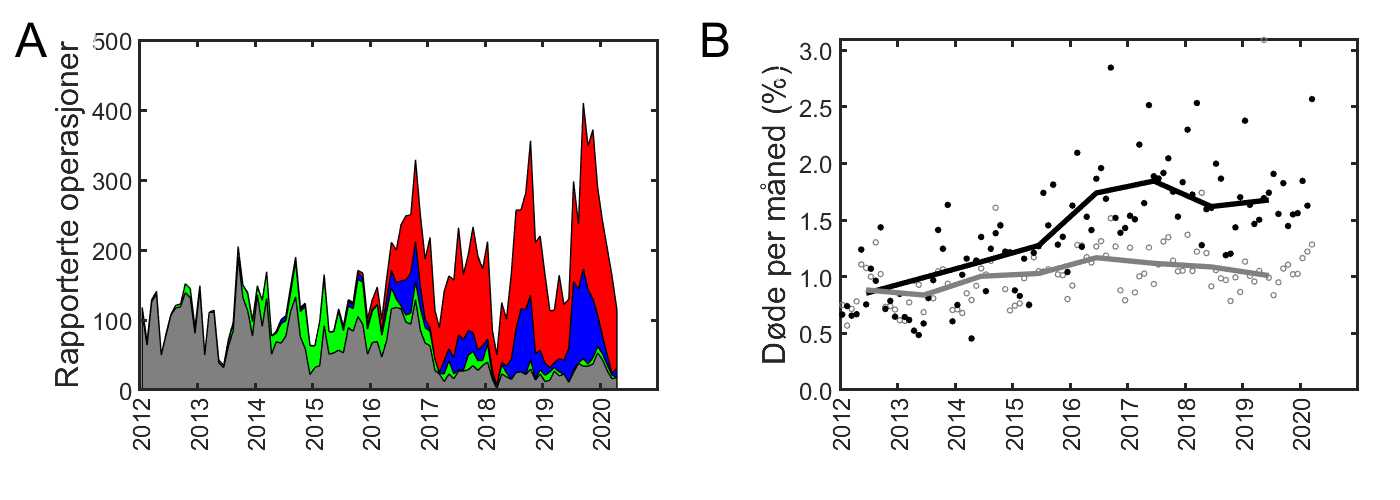
Norge eksporterte i 2018 1,1 millioner tonn oppdrettslaks og 46 400 tonn ørret av en verdi på henholdsvis 67,8 og 3 milliarder norske kroner, og utgjorde 72 % av den samlede eksportverdien av sjømat på 99 milliarder kroner (tall fra Norges sjømatråd). Siden 2012 har oppdrettsproduksjonen ligget på rundt 1,2–1,3 millioner tonn i året, og også for 2018 var produksjonen på samme nivå (figur 1.1). Produksjon av regnbueørret var på 66 723 tonn i 2018, en økning på rundt 5 000 tonn fra 2017. Selv om det arbeides aktivt med andre arter og nye produksjonsformer, vil sannsynligvis laks fortsette å være den viktigste oppdrettsarten i mange år.
Figur 1.1 Akvakulturproduksjon av atlantisk laks, regnbueørret og andre fiskearter i Norge i perioden 1980–2018. Kilde: Fiskeridirektoratet.
1.1.2 - Miljømessig bærekraft og dyrevelferd i norsk fiskeoppdrett
Med rundt 400 millioner oppdrettsfisk stående i åpne merder langs kysten, er det liten uenighet om at aktiviteten i større eller mindre grad påvirker miljøet. Samtidig har det vært dokumentert til dels store utfordringer knyttet til dyrevelferd i norsk fiskeoppdrett. Til tross for omfattende forskning og overvåkning, har det vært vanskelig å få et helhetlig bilde og felles forståelse av hvor store utfordringene knyttet til miljøvirkninger og dyrevelferd har vært i norsk havbruksnæring. Havforskningsinstituttet har derfor årlig gjennomført og publisert en risikovurdering av norsk fiskeoppdrett siden 2011, der vi også i de seinere årene har tatt med oversikter og vurderinger knyttet til dyrevelferden.
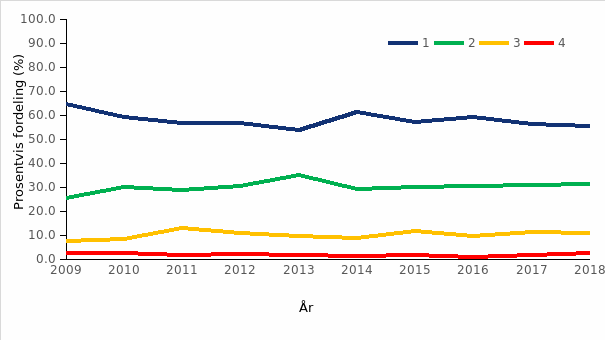
I Stortingsmelding 16 (2014–2015) Forutsigbar og miljømessig bærekraftig vekst i norsk lakse- og ørretoppdrett slås det fast at «Regjeringen mener at miljømessig bærekraft må benyttes som den viktigste forutsetningen for å regulere videre vekst i oppdrettsnæringen». Med bakgrunn i denne stortingsmeldingen ble det i 2017 innført en handlingsregel basert på produksjonsområder og miljøindikatorer som fremtidig system for kapasitetsendringer (trafikklyssystemet). En rekke miljøindikatorer ble vurdert i prosessen, og basert på dagens produksjonsform med oppdrett i åpne merder i sjø, kunnskapsstatus på de ulike indikatorene samt Norges særskilte internasjonale ansvar for å bevare villaksen, ble det enighet om at påvirkning fra lakselus på vill laksefisk var den mest relevante miljøindikatoren for å regulere regional vekst innen oppdrett av laksefisk. I den sammenheng valgte en å dele kysten opp i 13 produksjonsområder basert på vannstrøm og smittepotensial for lakselus.
Trafikklyssystemet innebærer en handlingsregel der miljøstatus, vurdert som lakselusindusert dødelighet hos vill laksefisk for hvert produksjonsområde, skal legges til grunn for om produksjonen får øke, skal være stabil eller må reduseres. Dette systemet fanger imidlertid bare opp én av mange miljøvirkninger av fiskeoppdrett, men er supplert av en rekke andre forvaltningsmessige grep for å regulere havbruksnæringen.
Akvakultur har imidlertid et svært komplekst risikobilde, og det er varierende kunnskapsnivå knyttet til de ulike faktorene. Siden 2011 har Havforskningsinstituttet publisert en samlet rapport som har omfattet både en risikovurdering og en oppdatert kunnskapsstatus for de ulike temaene i rapporten. Fra 2019 har det blitt innført ny risikometodikk, der risikovurderingen rendyrkes og fortsatt publiseres i rapportserien «Risikorapport norsk fiskeoppdrett», mens kunnskapen som ligger til grunn for risikovurderingen publiseres i en ny rapportserie «Kunnskapsstatus - miljøeffekter av norsk fiskeoppdrett». Her vil det gis en oppdatering av kunnskapen for hvert tema fra risikorapporten, en utdypning av de ulike risikofaktorene som ligger til grunn for vurderingen samt oppdaterte faglige referanser.
2 - Ytterligere genetisk endring hos villaks som følge av rømt oppdrettslaks
Forfatter(e):
Kevin Glover
(HI), Kjetil Hindar (*), Vidar Wennevik
, Monica F. Solberg
(HI), Sten Karlsson (*), Mikko Heino (**), Geir Bolstad (*), Ola H. Diserud (*), Øystein Skaala
, Terje Svåsand
(HI), Peder Fiske (*), *Norsk institutt for naturforskning og (**) Universitetet i Bergen/Havforskningsinstituttet
Fra laksefella i Etne. Foto: Erlend A. Lorentzen/Havforskningsinstituttet
2.1 - Innledning
Rømming av oppdrettslaks fra oppdrettsanlegg utgjør en trussel mot den genetiske integriteten til de ville laksebestandene. U ndersøkelser viser at det er en sammenheng mellom andel rømt oppdrettslaks i et vassdrag og genetisk endring målt som innkryssing med genetiske markører. Det betyr at jo flere rømte oppdrettslaks det er på gyteplassen, desto høyere er sannsynligheten for genetisk endring. I tillegg til andel rømt oppdrettslaks på gyteplassen, vil villaksbestandenes robusthet påvirke nivået av innkryssing. Dette fordi tallrike villaksbestander uten tidligere genetisk innkryssing antakelig er mer robuste overfor rømt oppdrettslaks, siden den rømte oppdrettslaksen der møter større konkurranse fra bedre tilpassete, ville individer. Disse faktorene danner grunnlaget for risikovurderingen for ytterligere genetisk endring hos villaks som følge av rømt oppdrettslaks (Grefsrud mfl. 2019; Glover mfl. 2020).
I denne todelte kunnskapsstatusen presenterer vi først en oversikt over konsekvenser av at rømt oppdrettslaks krysser seg med villaks. Videre presenterer vi en utvidet gjennomgang av faktorene knyttet til selve risikovurderingen; andel rømt oppdrettslaks på gyteplassene (rømming, andel rømt oppdrettslaks i elv og utfisking/ fjerning av rømt oppdrettslaks fra elv) og bestandenes robusthet for ny innkryssing (bestandsstatus og genetisk status). Vi drøfter også ulike biologiske faktorer ved den rømte fisken som kan påvirke dens gytesuksess i naturen, men som på nåværende tidspunkt ikke er inkludert i risikovurderingen.
2.2 - Konsekvenser av innkryssing av rømt oppdrettslaks
Konsekvensen av at oppdrettsfisken rømmer og gyter sammen med villfisk, kan være at det oppstår en genetisk endring i de ville bestandene av laks. Gjennom flere internasjonale arbeid er det godt dokumentert at rømt oppdrettslaks kan gyte og etterlate seg avkom i naturen (f.eks. Sægrov mfl. 1997; Clifford mfl. 1998a; Crozier 2000; Glover mfl. 2013; Karlsson mfl. 2016). Det er også dokumentert at innkryssing av rømt oppdrettslaks reduserer den genetiske variasjonen som finnes naturlig mellom bestandene (Skaala mfl. 2006; Glover mfl. 2013).
2.2.1 - Atlantisk laks, en art med genetisk forskjellige bestander
Gjennom de siste 40 årene har det vokst fram en omfattende vitenskapelig litteratur om laks som dokumenterer en geografisk bestandsstruktur med store genetiske forskjeller mellom bestander i Nord-Amerika og Europa, og med regionale og lokale forskjeller innenfor kontinentene (Webb mfl. 2007; Bourret mfl. 2013, Ozerov mfl. 2017; Wennevik mfl. 2019). Geografisk oppdeling av en art, og variasjoner i livsmiljø, bidrar til utvikling av genetiske forskjeller mellom bestander, både i gener av betydning for fitness (tilpasningsdyktighet) og i ikke-selekterte regioner av genomet (arvestoffet). Siden vi ikke vet hvilke gener som nå eller i fremtiden er viktige for individer og bestander, er det et mål å bevare den naturlige genetiske variasjonen både innen og mellom bestander.
De siste årene har den vitenskapelige produksjonen som dokumenterer genetiske forskjeller mellom laksebestander økt betraktelig, delvis som følge av den raske utviklingen innenfor molekylærbiologi og statistikk. Etter hvert er det også vist og modellert at avkom av oppdrettslaks har lavere overlevelse i naturen og at de derfor kan påvirke bestandene av villaks negativt der de krysser seg inn (Hindar mfl. 1991; Bourke mfl. 1997; McGinnity mfl. 1997; Verspoor 1997; Fleming mfl. 2000; Koljonen mfl. 2002; Fraser mfl. 2011; Skaala mfl. 2012; Besnier mfl. 2015; Reed mfl. 2015; Skaala mfl. 2019).
2.2.2 - Hvor ulik er villaks og oppdrettslaks?
Den genetiske påvirkningen fra rømt oppdrettslaks på villaks er kompleks, og er avhengig av mange faktorer som varierer i tid og rom. Viktige, eller sannsynligvis viktige faktorer, inkluderer blant annet andel rømt oppdrettslaks i de ville bestandene (Glover mfl. 2013; Heino mfl. 2015; Karlsson mfl. 2016), deres gytesuksess (Fleming mfl. 1996, 2000), graden av genetisk forskjell mellom oppdrettet og vill laks (Fraser mfl. 2010) og status for den ville bestanden (Glover mfl. 2012, 2013; Heino mfl. 2015). Det er også grunn til å tro at responsen hos de ville bestandene som følge av innkryssing av rømt oppdrettslaks vil variere fra bestand til bestand (Normandeau mfl. 2009; Harvey mfl. 2016a).
Genetiske forskjeller mellom vill- og oppdrettslaks kan oppstå gjennom tilfeldige prosesser (for eksempel begrenset antall stamfisk i hver generasjon, deres opprinnelse og tilpasning til oppdrettsmiljøet) og som resultat av målrettet seleksjon av egenskaper i avlsarbeidet. I Norge har man domestisert laksen i snart 50 år, med tidlig oppstart av målrettet avl (Gjedrem mfl. 1991; Gjøen & Bentsen 1997; Gjedrem 2010) for å endre kommersielt viktige egenskaper som tilvekst, kjønnsmodning, fettfordeling og sykdomsresistens. Seleksjon for en mer økonomisk produktiv oppdrettslaks foregår i avlsprogram som opprinnelig var basert på vill laks fanget i en rekke norske elver tidlig på 1970-tallet (Gjedrem mfl. 1991; Gjøen & Bentsen 1997). Under kontrollerte forhold blir de mest produktive familier og individer selektert basert på produksjonskriterier (f.eks. tilvekst), og disse individene blir benyttet til å føre stammen videre. På denne måten oppnår man en gradvis domestisering av laksen der viktige kommersielle trekk blir forandret i ønsket retning.
Glover mfl. (2017) oppsummerte den omfattende kunnskapen som nå er etablert om de genetiske forskjellene mellom oppdrettslaks og villaks. Oppdrettete og vill laks har vært sammenlignet med ulike metoder i en lang rekke vitenskapelige arbeider, og disse omfatter studier av genetisk variasjon med molekylære markører, eksperimentelle studier i laboratorium og kar hvor en har sammenlignet atferd, morfologi og fysiologi, og studier av overlevelse og vekst i et naturlig miljø. Noen eksperimentelle studier er også supplert med analyse av genuttrykk og fysiologisk målinger.
Sammenligning av genetisk variasjon og diversitet i oppdrettslinjer og ville laksebestander har blitt gjennomført over lengre tid med en rekke molekylære markører. De tidligste studiene var hovedsakelig basert på analyser av proteinkodende gen (Verspoor 1988; Cross & Challanain 1991; Mjølnerød mfl. 1997; Skaala mfl. 2005), der det er blitt vist genetisk forskjell mellom oppdrettslaks og de ville utgangsbestandene, og redusert nivå av genetisk variasjon i oppdrettslaks, målt som allelisk diversitet og heterozygoti.
Seinere har DNA-markører blitt brukt til å sammenligne oppdrettslinjer og vill laks, for eksempel med mini- og mikrosatellittmarkører (Mjølnerød mfl. 1997; Clifford mfl. 1998a, b; Norris mfl. 1999; Skaala mfl. 2004), mikrosatellittmarkører kombinert med både mitokondrie DNA (mtDNA) (Karlsson mfl. 2010), og «single nucleotide polymorphism» (SNP) markører (Rengmark mfl. 2006; Karlsson mfl. 2011). Selv om resultatene fra disse studiene varierer noe, støtter de opp om konklusjonene fra tidligere analyser basert på proteinkodende gen; det er redusert genetisk variasjon hos den enkelte oppdrettslinje sammenlignet med de ville laksebestandene.
I en sammenligning av fem avlslinjer av oppdrettslaks med fire villaksbestander fra Neiden, Namsen, Vosso og Loneelva, viste alle de 12 undersøkte DNA-mikrosatellittmarkørene redusert allelisk variasjon i samtlige avlslinjer sammenlignet med de ville bestandene (Skaala mfl. 2004). I gjennomsnitt hadde avlslinjene 58 % av den alleliske variasjonen sammenlignet med prøver av villaks, og dette kan forklares av begrenset effektiv bestandsstørrelse i oppdrettslinjene. Samtidig var estimatene for genetisk forskjell flere ganger høyere mellom de ulike avlslinjene enn mellom de ville laksebestandene. Andre studier har vist at tap av genetisk diversitet i oppdrettslinjer er mer komplekst enn tidligere antatt (Karlsson mfl. 2010), men det kan likevel konkluderes med at oppdrettslaks har redusert genetisk variasjon i forhold til ville laksebestander. Dette samsvarer med tilsvarende observasjoner fra andre domestiserte organismer (se Araki & Schmid 2010), og kan forklares med at det er et begrenset antall familier/individer som bidrar til hver generasjon i et avlsprogram. Den effektive bestandsstørrelsen i norske oppdrettslinjer er tidligere blitt estimert til 33–125 individer (Mork mfl. 1999), noe som teoretisk sett skal føre til lav til moderat innavl over den aktuelle avls-perioden.
På grunn av et målrettet avlsarbeid er det ikke uventet at oppdrettslaks vokser bedre enn villaks i et oppdrettsmiljø (Einum & Fleming 1997; Thodesen mfl. 1999; Fleming mfl. 2002; Glover mfl. 2009; Solberg mfl. 2013a, b; Harvey mfl. 2016a, b, c). I et oppdrettsmiljø vil vekstforholdet mellom oppdrettslaks og villaks være 2-5:1 (Glover mfl. 2017). Også i et naturlig miljø kan oppdrettslaks ha større vekst enn villaks (Johnsson & Björnsson1994; Einum & Fleming 1997; McGinnity mfl. 1997, 2003; Skaala mfl. 2012, 2019; Jonsson & Jonsson 2017; Bolstad mfl. 2017), men ikke i samme grad som i oppdrettsmiljø (typisk vekstforhold mellom oppdrettslaks og villaks i naturen er 1,0-1,3:1). Oppdrettslaks har et forhøyet vekstpotensial, relativt til villaks, som den ikke får utnyttet i naturen, og det er en kombinasjon av plastisitet (ulik mattilgang mellom disse miljøene) og naturlig seleksjon mot hurtigvoksende oppdrettslaks (økt dødelighet) som er årsaken til de små vekstforskjellene mellom oppdrettslaks og villaks i naturen (Glover mfl. 2018). Vekst hos hybrider mellom oppdrettslaks og villaks er ansett å være additivt (mellomliggende til vekst hos vill og oppdrett), men ikke-additive genetiske effekter knyttet til vekst er nylig dokumentert i Atlantisk laks (Besnier mfl. 2020).
Mange egenskaper som ikke inngår direkte i avlsarbeidet, som aggresjon, stress- og temperaturtoleranse, kan også bli endret hos oppdrettslaksen gjennom avlsprosessen (Fleming & Einum 1997; Houde mfl. 2010; Debes & Hutchings 2014; Solberg mfl. 2016). Årsaken er at målrettet seleksjon for blant annet tilvekst påvirker både aggresjon og andre egenskaper, for eksempel hormonregulering og atferd. I eksperimentelle studier er det vist at tilførsel av veksthormon øker appetitten (Johnsson & Björnsson 1994; Jönsson mfl. 1996), aggresjon og aktivitet (Jönsson mfl. 1998), altså atferd som sannsynligvis påvirker overlevelse i naturen (Johnsson mfl. 1996; Jönsson mfl. 1996; Martin-Smith mfl. 2004). Det er derfor ikke overraskende at oppdrettslaks er ulik villaks i flere egenskaper som påvirker overlevelse i naturen, slik som tilvekst, aggresjon, dominans og antipredatoratferd (Einum & Fleming 1997; Fleming & Einum 1997; Johnsson mfl. 2001; Fleming mfl. 2002; Houde mfl. 2010). I tillegg er det avdekket genetiske forskjeller mellom vill- og oppdrettslaks i egenskaper som kjøttfarge, kjønnsmodning og fettinnhold (Glover mfl. 2009), reaksjonsnormer (Darwish & Hutchings 2009; Solberg mfl. 2013a, b), morfologi (Fleming & Einum 1997; Perry mfl. 2019), stresstoleranse (Solberg mfl. 2013a) og en rekke andre egenskaper (Glover mfl. 2017).
Det er også dokumentert genetiske forskjeller i genuttrykk mellom laks av ville- og oppdrettsforeldre, samt hybrider av disse (Roberge mfl. 2006, 2008; Solberg mfl. 2012; Bicskei mfl. 2014, 2016). Resultatene er også interessante fordi hybridene ikke alltid hadde et genuttrykk som lå mellom foreldrebestandene. Hybridene hadde i noen tilfeller genuttrykk som lå høyere enn verdiene målt for vill- og oppdrettsfisk, noe som tyder på ikke-additive genetiske effekter. Dokumentasjon av ikke-additive genetiske effekter betyr i praksis at innkryssing av oppdrettsfisk i ville bestander i noen tilfeller kan gi uventede effekter. Hvilken endring man får i genuttrykk hos hybrider av vill- og oppdrettslaks er avhengig av hvilke ville bestander som krysses inn (Normandeau mfl. 2009; Fraser mfl. 2010).
Det er ikke avdekket noen store forskjeller i resistens mot lakselus, ILA (infeksiøs lakseanemi) eller furunkulose mellom vill- og oppdrettslaks (Glover & Skaala 2006; Glover mfl. 2006a, b). Det er heller ikke avdekket genetiske forskjeller i deformiteter hos smolt av oppdretts- og villaks (Fjelldal mfl. 2009). For infeksiøs pankreasnekrose er det funnet genetiske markører (såkalt QTL – Quantitative Trait Locus) som forklarer en stor grad av toleransen for sykdommen (Houston mfl. 2008; Moen mfl. 2009) og denne kunnskapen er inkludert i avlsarbeid hos flere avlsselskaper. Seleksjon for sykdomsresistens har derimot vært praktisert ulikt for de ulike oppdrettslinjene, og dette vanskeliggjør sammenligning mellom linjer.
Til tross for at avkom av rømt oppdrettslaks har lavere overlevelse i naturen enn avkom av villaks, har flere tidligere forsøk i naturen (Skaala mfl. 2014) og under eksperimentelle forhold (Fleming & Einum 1997; Debes & Hutchings 2014; Solberg et al. 2015) ikke klart å demonstrere at avkom av oppdrettslaks er utsatt for høyere predasjon. Først nylig er det dokumentert at avkom av rømt oppdrettslaks er et lettere bytte for predatorer som større ørret, noe som underbygger deres lavere overlevelse i naturen (Solberg mfl. 2020). Dette resultatet støttes oppom av det faktum at redusert anti-predator-atferd (Einum & Fleming 1997; Johnsson et al. 2001; Houde et al. 2010), i tillegg til økt toleranse for predasjonsrelatert stress (Debes & Hutchings 2014), tidligere har blitt dokumentert i eksperimentelle forsøk.
En oppsummering av vitenskapelig litteratur viser at det er til dels store genetiske forskjeller mellom vill- og oppdrettslaks i kvantitative egenskaper som har direkte eller indirekte betydning for overlevelsen av laks i naturen. Blant annet har tilstedeværelse av hybrider vist seg å ha en negativ påvirkning på overlevelse til villfisk (Robertsen mfl. 2018). Det er grunn til å tro at de genetiske forskjellene kommer til å øke for hver avlsgenerasjon. Dette har blitt observert for tilvekst, der forskjellen mellom vill- og oppdrettslaks under oppdrettsbetingelser økte ytterligere fra generasjon 8 til 10 (Glover mfl. 2009; Solberg 2013a, b).
2.2.3 - Genetisk påvirkning fra rømt oppdrettslaks – hva forteller empiriske data oss?
Ved hjelp av ulike biokjemiske og molekylærgenetiske metoder er det vist at rømt oppdrettslaks gyter i elver. Ved undersøkelser av et pigment i rogn og yngel, som reflekterer ulik diett hos villaks og oppdrettslaks, fant Lura & Sægrov (1991) at rømt oppdrettslaks faktisk produserte levedyktig avkom i en elv. I en skotsk undersøkelse fant Webb mfl. (1993) et pigment fra rømt oppdrettslaks i 14 av 16 undersøkte elver, med et gjennomsnittlig innslag på 5,1 % fra rømt fisk. I Vosso ble bidraget fra rømt oppdrettslaks estimert til opp mot 80 % ved denne metoden (Sægrov mfl. 1997). Bevis for at rømt oppdrettslaks produserte levedyktig avkom ble også funnet i Irland ved hjelp av genetiske markører (Clifford mfl. 1998a; Crozier 1993, 2000). Også langt utenfor det naturlige utbredelsesområdet til den atlantiske laksen, i British Columbia, Canada, er det vist at rømt oppdrettslaks kan produsere levedyktig avkom (Volpe mfl. 2000).
Det første genetiske studiet for å undersøke om norske villaksbestander har endret seg genetisk over tid som følge av innkryssing av rømt oppdrettslaks ble publisert av Skaala mfl. (2006). Her ble det laget DNA-profiler for de sju laksebestandene Namsen, Etne, Opo, Vosso, Granvin, Eio og Håelva. Det ble benyttet gamle skjellprøver og materiale innsamlet i nyere tid, etter lengre tids innslag av rømt oppdrettslaks. Håelva på Jæren ligger i en region der det nesten ikke er lakseoppdrett, og andelen rømt oppdrettslaks i villaksbestanden har vært lav, trolig under 5 %. I Håelva ble det ikke funnet endring i de genetiske profilene. I tre andre bestander, Opo, Vosso og Eio i Hordaland, ble det funnet signifikante endringer i de genetiske profilene over tid. Mer overraskende var det likevel at det ikke ble funnet endringer i Etneelva, Namsen eller Granvinelva, som alle hadde hatt høye andeler rømt oppdrettslaks i gytebestandene, permanent eller periodisk.
Som en videreføring av dette arbeidet (Skaala mfl. 2006) ble det gjort en mer omfattende analyse av 21 bestander der historiske og nye prøver ble analysert for flere mikrosatellittmarkører. Undersøkelsen omfattet elver fra hele landet (Glover mfl. 2012) og påviste genetiske forandring over tid i 6 av 21 elver, mens i 15 av bestandene ble det ikke funnet genetiske forandringer. Som i den første undersøkelsen utført av Skaala og kolleger (2006), var det noen bestander med høye innslag av rømt oppdrettslaks på gyteplassene der det ikke ble påvist forandringer.
I de seks bestandene der det ble påvist forandring, har det vært registrert rømt oppdrettslaks i større eller mindre grad. I tillegg ble det funnet nye genvarianter som indikerer at forandringene i disse seks elvene hovedsakelig skyldes genflyt fra andre kilder. Den genetiske forskjellen mellom disse seks bestandene er også blitt redusert over tid. Basert på alle data, ble det konkludert med at innkryssing av rømt oppdrettslaks er hovedårsaken til forandringene. Dette er i tråd med simuleringer fra modeller som viser at innkryssing av rømt oppdrettslaks vil redusere genetisk differensiering mellom bestander over tid (Mork 1991; Besnier mfl. 2011).
Det er kjent at mikrosatellittmarkører i noen tilfeller vil underestimere innkryssing av rømt oppdrettslaks i ville bestander på grunn av signalstøy når en villaksbestand mottar oppdrettsfisk fra flere ulike avlsbestander (Besnier mfl. 2011). Det betyr at antall elver som er påvist å være genetisk påvirket i denne studien (Glover mfl. 2012), og omfanget av de genetiske forandringene i disse elvene, må betraktes som et minimumsestimat. For å få et mer presist svar på omfanget av innkryssing av rømt oppdrettslaks ble en studie gjennomført basert på SNP-markører utviklet for å kunne identifisere oppdrettslaks og skjelne dem fra villaks (Karlsson mfl. 2011). Disse SNP-markørene gir mer presis informasjon om genetiske forandringer forårsaket av rømt oppdrettslaks, og er i mindre grad påvirket av problematikken ved at genetisk forandring i den enkelte villaksstamme er vanskelig å påvise når innkryssing skjer via oppdrettslaks fra forskjellige avlslinjer (Besnier mfl. 2011).
I en studie av 20 laksebestander langs hele norskekysten (Glover mfl. 2013) ble disse SNP-markørene brukt til å estimere prosent innkryssing av rømt oppdrettslaks. Resultatene viste at det genetiske bidraget til noen bestander var nesten 50 %, mens estimert innkryssing av oppdrettslaks var mye lavere i de fleste undersøkte elvene. Arbeidet støttet opp om konklusjonene til de tidligere publikasjonene med andre markørtyper (Skaala mfl. 2006; Glover mfl. 2012). I tillegg til at det ble dokumentert genetiske forandringer i noen villaksbestander på grunn av innkryssing av rømt oppdrettslaks, viser alle disse tre studiene (Skaala mfl. 2006; Glover mfl. 2012, 2013) at den genetiske forskjellen mellom noen ville bestander er blitt mindre over tid. Dette kan tilskrives at de ville bestandene som har hatt en betydelig innkryssing av rømt oppdrettslaks, blir mer lik oppdrettslaksen – og dermed også mer lik hverandre.
Parallelt med dette arbeidet utarbeidet Karlsson mfl. (2014) en statistisk metode der prosentvis innkryssing kunne estimeres i enkeltindivider og uavhengig av om det fantes en historisk prøve av den aktuelle bestanden. Metoden brukte et estimat av andelen «villgenom» i bestanden (omtalt som P(wild) = mengde arvestoff som stammer fra ville foreldre kontra oppdrettsforeldre) for å beregne innkryssingen av rømt oppdrettslaks (Karlsson mfl. 2014).
Denne metodeutviklingen gjorde det mulig å estimere innkryssing i et stort antall ville laksebestander. Karlsson mfl. (2016) studerte 147 laksebestander som til sammen representerer tre fjerdedeler av de ville lakseressursene i Norge, og analyserte genetisk mer enn 20 000 laks som var klekket i naturen og derfor så ut som villaks. Den gjennomsnittlige genetiske innkryssingen i bestandene av voksen laks var 6,4 % med et spenn fra 0 % til over 40 % innkryssing i enkelte bestander. Forskerne fant også at den genetiske innkryssingen generelt var mindre i nasjonale lakseelver og nasjonale laksefjorder enn i bestander uten denne særlige beskyttelsen (Karlsson mfl. 2016). I et materiale av 109 bestander der kun voksen laks klekket i naturen ble inkludert i datamaterialet, var den genetiske innkryssingen signifikant forskjellig fra 0 i omtrent halvparten av bestandene (Karlsson mfl. 2016). Det samme resultatet fikk Karlsson mfl. (2016) om de også tok med et større antall elver, der enkelte estimater var basert på prøver av ungfisk.
Data fra de 20 bestandene som var publisert av HI i Glover mfl. (2013), samt data fra NINA for et større antall bestander (Karlsson mfl. 2016), ble vurdert sammen for å dokumentere genetisk status for ville laksebestander til «Kvalitetsnorm for ville bestander av atlantisk laks ( Salmo salar )», som er en forskrift av 20. september 2013 hjemlet i Naturmangfoldloven. I den første versjonen av Kvalitetsnormen av februar 2016 vurderte forskere fra HI og NINA i fellesskap genetisk status i 125 laksebestander, basert på estimater for innkryssing av rømt oppdrettslaks i bestanden (Diserud mfl. 2016). I senere utgaver av Kvalitetsnormen er antallet bestander økt til 175 og deretter til 225 norske villaksbestander (Diserud mfl. 2017, 2019b). Den hittil mest omfattende vurderingen av genetisk innkryssing ga følgende resultat (pr. Diserud mfl. 2019b): ingen genetiske forandringer (75 vassdrag), svake genetiske forandringer (67 vassdrag), moderat genetiske forandringer (16 vassdrag) og store genetiske forandringer (67 vassdrag). Vurderingen av genetisk innkryssing i disse 225 bestandene dokumenterer at 2/3 av de ville laksebestandene har blitt påvirket av rømt oppdrettslaks.
Eksperimenter i naturen har vist seleksjon mot avkom av rømt oppdrettslaks, både i Irland (McGinnity mfl. 1997, 2003) og Norge (Fleming mfl. 2000; Skaala mfl. 2012, 2019). En skulle derved forvente at når én og samme årsklasse (dvs. laks fra samme klekkeår) studeres, så vil et materiale av ungfisk vise høyere genetisk påvirkning av oppdrettslaks enn et materiale av voksen laks. Dette er også det Karlsson mfl. (2016) fant gjennom analyser av ungfisk som var innsamlet noen år før et materiale av voksen laks fra de samme elvene: ungfisken hadde i gjennomsnitt 2,5 % høyere grad av genetisk innkryssing enn den voksne laksen. Seleksjon mot avkom av rømt oppdrettslaks har også blitt dokumentert i etterkant av en større rømmingsepisode på Newfoundland i Canada (Wringe mfl. 2018; Sylvester mfl. 2019).
Som oversiktsartikkelen til Glover mfl. (2017) oppsummerer, så foreligger det et solid grunnlag for å hevde at rømt oppdrettslaks kan påvirke villfiskbestander negativt. Den beste dokumentasjonen av effekter på overlevelse og viktige egenskaper kommer fra eksperimentelle studier utført i tre kontrollerte elver (McGinnity mfl. 1997, 2003; Fleming mfl. 2000; Skaala mfl. 2012, 2019), samt en studie som viser sammenhenger mellom et molekylært mål på innkryssing (P(wild)) og viktige livshistorietrekk i ville bestander (Bolstad mfl. 2017). Det arbeides videre med å kvantifisere økologiske effekter av innkryssingen av oppdrettslaks i ville bestander i NFR-prosjektet QuantEscape II.
Det er gjennomført få empiriske studier som evaluerer de genetiske effektene av at rømt oppdrettslaks krysser seg inn i villaksbestander, selv om det foreligger en omfattende litteratur om populasjonsgenetisk teori, og om de grunnleggende evolusjonskreftene (mutasjon, naturlig seleksjon, genetisk drift og migrasjon) som påvirker og former den genetiske sammensetningen i bestander.
En direkte og informativ tilnærming til problematikken er å sammenligne tilvekst, atferd og overlevelse hos sammenblandede familiegrupper av oppdrettslaks, villaks og hybrider under like forhold, såkalte «common garden-studier», i et naturlig miljø. Dette kan innebære utplanting av lakserogn fra DNA-identifiserbare familier av oppdrettslaks, villaks og hybrider dem imellom (McGinnity mfl. 1997, 2003; Skaala mfl. 2012, 2019), eller utsetting av kjønnsmodne individer med kjente genetiske profiler (Fleming mfl. 2000) i naturlig elvemiljø, der alle avkom i ulike livsstadier fra rogn til kjønnsmodning i ettertid kan identifiseres ved genetiske markører.
Det første prosjektet som er gjennomført på dette feltet, ble utført i Burrishoole, Irland (McGinnity mfl. 1997, 2003; Ferguson mfl. 2002). I dette prosjektet ble tilvekst, overlevelse og populasjonsdynamikk hos villaks, oppdrettslaks og hybrider undersøkt gjennom to generasjoner. Et stort antall individer fra mange familier av villaks, oppdrettslaks, første- og andregenerasjonshybrider og førstegenerasjonshybrider tilbakekrysset til henholdsvis villaks og oppdrettslaks, ble plantet ut i tre årsklasser som øyerogn ovenfor fiskefellen i Burrishoole. Tilsvarende grupper ble satt ut som smolt i elven for å studere vekst og overlevelse i sjøfasen.
En omfattende innsats med innsamling og genotyping for å identifisere opphavet til alle individ ble gjennomført fra yngel til gytefisk som kom tilbake fra havet etter ett og to år i sjø. I alle tre årsklassene hadde oppdrettslaksen signifikant lavere representasjon enn villaksen i prøver av 0+ parr. Ikke overraskende viste det seg at oppdrettslaksen vokste bedre enn villaksen, og at den større oppdrettsparren fortrengte den ville parren nedover elva gjennom konkurranse. Selv om ungfisk av oppdrettslaks vokste bedre og fortrengte en del av den ville ungfisken, var smoltproduksjonen av oppdrettslaks bare henholdsvis 34, 34 og 55 % sammenlignet med villaksen i de tre årsklassene. Den gjennomsnittlige gjenfangsten etter sjøoppholdet var 0,3 % for oppdrettslaks og 8 % for villaks utsatt som smolt. Overlevelse hos hybridene viste seg ofte å ligge mellom villaks og oppdrettslaks.
Et lignende prosjekt ble gjennomført ved NINAs feltstasjon på Ims (Fleming mfl. 2000) der alle ned- og oppvandrende fisk kontrolleres i en toveis fiskefelle i elven Imsa. Her ble det satt ut kjønnsmodne villaks fra Imsa og oppdrettslaks med kjente genetiske profiler ovenfor fiskefellen. De to gruppene hadde lignende vandringsmønster og valgte de samme gyteplassene i elven. Vill hannlaks var mer aktive i kurtisering av hunnlaksen enn oppdrettshannene var, og hadde dessuten mindre restgonader etter gyting enn oppdrettshannene hadde. Gytesuksessen var mye lavere hos oppdrettslaksen både for hanner (24 %) og hunner (32 %) sammenlignet med villaksen i et samtidig forsøk i store gytebassenger med steinbunn. Gjennom ferskvannsfasen endret andelen av genotyper seg ytterligere i disfavør av oppdrettslaksen, og hoveddelen av oppdrettsbidraget var representert i form av hybrider, produsert av oppdrettshunner og ville hanner. Studier av dietten viste betydelige overlapp i næringsvalg, noe som viser næringskonkurranse mellom oppdrettsavkom og villaksyngel. Den totale smoltproduksjonen for elven var 28 % lavere enn forventet ut fra rognmengde og det observerte forholdet det normalt har vært i Imsa mellom mengde egg og antall smolt (Jonsson mfl. 1998). Det var også en tilsvarende reduksjon i forventet smoltproduksjon av den ville gytefisken. Oppdrettslaksen smoltifiserte og vandret ut tidligere og ved lavere alder enn villaksen. I motsetning til resultatene fra Burrishoole-prosjektet, fant en i Imsaprosjektet ingen forskjell mellom gruppene i sjøoverlevelse. Senere eksperimenter på Ims med utsetting av smolt av villaks, oppdrettslaks og førstegenerasjons krysninger mellom dem (1996-1998), viste signifikant lavere gjenfangst av oppdrettslaks (Hindar mfl. 2006). Utsatt smolt av oppdrettslaks har også vist høyere feilvandringsrate enn utsatt smolt av Imsalaks på Ims (Jonsson mfl. 2003; Jonsson & Jonsson 2017). Hybrider mellom oppdrettslaks og villaks hadde også høyere feilvandringsrate sammenlignet med den ville Imsalaksen (Jonsson & Jonsson 2017). Retningen på hybridiseringen hadde betydning ved at hybrider med vill mor hadde lavere feilvandring enn hybrider med vill far (Jonsson & Jonsson 2017).
Ved Havforskningsinstituttets feltstasjon i Guddalselva i Hardanger ble det initiert et prosjekt basert på oppsettet for Burrishoole-prosjektet. Der ble definerte familiegrupper av vill og oppdrettet laks, og hybrider mellom disse, plantet ut som rogn (Skaala mfl. 2012). All foreldrefisk var genotypet med DNA-mikrosatellittmarkører. Dermed kunne alle individ som var satt ut som øyerogn i seks årsklasser, ca. 150 familier i et «common garden-studie», identifiseres til familie. Det ble samlet inn juvenil laks av alle årsklassene fra elvehabitatet, og tilvekst, overleving og diettvalg ble undersøkt for hver familie.
Resultatene fra de tre første årsklassene viste en overlevelse (fra utplantet egg til smolt) som varierte mellom 0,17 og 6,4 % for de 69 forskjellige familiene (Skaala mfl. 2012). Resultatene viste at eggstørrelsen hadde stor betydning for overlevelsen. I dette studiet hadde oppdrettslaksene større egg enn villaksene. Dette bidro til at noen oppdrettsfamilier hadde en høy overlevelse fra egg til smolt i forsøket. Da det ble kontrollert for eggstørrelse ved å sammenligne overlevelse til en oppdrettsfamilie og dens halvsøskenfamilie der fars bidrag kom fra en vill hann, viste det seg at 16 av 18 halvsøskenfamilier hadde høyere overlevelse med bidrag fra vill far kontra oppdrettsfar. Dette viser at det er additiv genetisk variasjon for overlevelsen i et naturlig miljø og at villaks har høyere overlevelse, selv om bildet kan være noe mer komplisert når det tas hensyn til eggstørrelse. I tillegg til lavere overlevelse når det tas hensyn til eggstørrelse, hadde smolt av oppdrettsforeldre noe høyere vekst enn hybrider og villaks i elven (forholdstall oppdrett:vill = 1,0-1,3:1).
Resultatene fra de tre siste årsklassene i perioden 2008–2016 (Skaala mfl. 2019) har i stor grad støttet resultater fra de tre første årsklassene, og det ble observert langt lavere overlevelse hos avkom hos oppdretts- og hybridlaks i forhold til villaks i ferskvann (1.8 % overlevelse hos oppdrettslaks mot 3.8 % overlevelse hos villaks). I tillegg ble det satt ut to årsklasser med oppdretts-, hybrid- og villsmolt i elven for å studere sjøoverlevelsen. Her fant vi lavere sjøoverlevelse hos avkom av oppdrettslaks enn hos avkom fra hybrider (gjennomsnittlig sjøoverlevelse) og villaks (0.41 % overlevelse hos oppdrettslaks mot 0.94 % overlevelse hos villaks).
Når avkom fra oppdrettslaks, villaks og hybridene konkurrerer om de samme byttedyrene i elven, vil innkryssing av oppdrettslaks kunne redusere den naturlige produksjonen av villsmolt på grunn av konkurranse. Når vi samtidig ser at avkom av oppdrettslaks også har lavere sjøoverlevelse, vil det i sum også kunne redusere antall laks som kommer tilbake til elven.
Utviklingen innenfor genomforskningen de siste årene gir stadig bedre muligheter for å vurdere de biologiske konsekvenser av innkryssing av oppdrettslaks i ville bestander på genomnivå. Man vil kunne identifisere hvilke gener som er involvert i de biologiske forskjellene man observerer mellom oppdretts- og villaks. En relativt ny studie identifiserte områder i genomet/arvestoffet som var knyttet til overlevelse i naturen, selv om det er uvisst hvilke gener som var involvert (Besnier mfl. 2015). Det er forventet at nye resultater fra QuantEscapeII-prosjektet (NFR-prosjekt 2016-2020) vil bidra til å kartlegge hvordan endringer i genomet oppstår som følge av innkryssing, og hvilke biologiske konsekvenser dette kan få for ville bestander.
Inntil nylig forelå det ikke dokumentasjon av forandringer i livshistorietrekk og demografi i villaksbestander som konsekvens av genetisk innkryssing. Dette kan skyldes at en ikke har hatt tilstrekkelig gode verktøy til å studere slike endringer, i tillegg til at det er først relativt nylig at man har klart å dokumentere og kvantifisere innkryssing av rømt oppdrettslaks i bestander med bruk av genetiske markører. En annen årsak er at varierende forhold i havet kan påvirke både livshistorie og demografiske parametere – noe som gjør at gradvise forandringer i bestander kan være krevende å kvantifisere på kort sikt. Likevel er det nylig publisert et arbeid som for første gang dokumenterer forandringer i livshistorietrekk hos villaks i et stort antall bestander som følge av genetisk innkryssing av rømt oppdrettslaks (Bolstad mfl. 2017).
Bolstad mfl. (2017) studerte 62 ville laksebestander med individer av ulik grad av genetisk slektskap til oppdrettslaks. Det genetiske slektskapet ble målt ved bruk av molekylærgenetiske metoder (Karlsson mfl. 2011, 2014). Studien viser at individer med høy grad av genetisk slektskap til oppdrettslaks har endret sjøalder (figur 2.1) og størrelse ved kjønnsmodning. Disse endringene var forskjellige mellom kjønn og for ulike typer laksestammer. I storlakselver (elver med overvekt av flersjøvinterlaks) ble det observert en økning i antall hunnlaks som returnerer fra havet etter to vintre (tosjøvinterlaks), og en nedgang i antall som returnerer etter en og tre vintre, med økende grad av genetisk slektskap til oppdrettslaks. For hannlaks i de samme elvene var det en økning i ensjøvinterlaks og en tilsvarende nedgang i to- og tresjøvinterlaks. I smålakselver (elver dominert av ensjøvinterlaks) ble det ikke observert slike endringer. Derimot ble det observert en større økning i aldersbestemt størrelse for laksen i smålakselvene enn for laksen i storlakselvene med økende grad av slektskap til oppdrettslaks. I smålakselvene økte vekten med 19 % fra 0 til 100 % genetisk slektskap til oppdrettslaks.
Figur 2.1 Sjøalderfordeling (%) ved kjønnsmodning ved ulik grad av genetisk slektskap til oppdrettslaks for begge kjønn i ulike typer elver. Sjøalder er oppgitt som ensjøvinter (blå), tosjøvinter (grønn) og tresjøvinter eller eldre (rosa). Vertikal strek representerer +/- en standardfeil (symmetrisk på logitskala). Det ble ikke observert laks med 100 % genetisk slektskap til oppdrettslaks i Finnmark. Fra Bolstad mfl. (2017).
Studien undersøkte også effekten på laks i Finnmark (Bolstad mfl. 2017). Laksen i Finnmark er mer i slekt med laksen på Kolahalvøya i Russland enn med resten av den norske laksen. På grunn av lavere utvalgsstørrelse var resultatene for Finnmark mindre sikre enn for resten av landet, men det kan med sikkerhet fastslås at det er ulik effekt av genetisk slektskap til oppdrettslaks i de to regionene. Spesielt ble det observert en kraftig effekt på antall vintre i sjøen for hannlaks i smålakselvene i Finnmark, som i økende grad ble kjønnsmodne etter to vintre i istedenfor etter en (figur 2.1). Det ble også observert en dramatisk økning i aldersbestemt størrelse i smålakselvene i denne regionen, med 24 % vektøkning fra 0 til 50 % genetisk slektskap til oppdrettslaks. Det er viktig å påpeke at de målte endringene i denne studien er underestimat (se Bolstad mfl. 2017 for utdypende forklaring).
2.2.4 - Modellering
Modellering av konsekvenser av innkryssing av rømt oppdrettslaks gir en mulighet til å estimere tidsforløp og omfang av genetisk forandringer i ville laksebestander. Her gjennomgås to publiserte modelleringsarbeid. Det første arbeidet av Hindar mfl. (2006) var basert på best tilgjengelig kunnskap om fitness-forskjeller mellom rømt oppdrettslaks, villaks og deres første- og andregenerasjons avkom midt på 2000-tallet, og estimerte andelen av en villaksbestand som er genetisk innkrysset med rømt oppdrettslaks. Dette ble studert ved forskjellige nivå av rømming. Arbeidet viste at ved 20 % innslag av rømt oppdrettslaks på gyteplassene, fikk en store forskjeller i sammensetningen av bestanden i løpet av 10 laksegenerasjoner (ca. 40 år). Forfatterne viste også at desto mer rømt oppdrettslaks i gytebestanden, desto større andel av den ville bestanden ble genetisk påvirket av rømt oppdrettslaks over tid. Modellen ble senere videreutviklet med en populasjonsdynamisk modell for bestandsstørrelse og en økonomisk modell for verdien av fiske etter laks (Liu mfl. 2013). Modellen hadde ingen fitness-funksjon på den ville bestanden og kunne ikke predikere økologiske konsekvenser av innkryssing annet enn i bestandsstørrelse.
En ny modell (IBSEM – Individual based eco-genetic model) for å studere genetisk innkryssing av rømt oppdrettslaks i villaksbestander er publisert av Castellani mfl. (2015). Modellen ble basert på best tilgjengelig kunnskap om fitnessforskjeller mellom avkom av oppdrettslaks og villaks midt på 2010-tallet. IBSEM er en individbasert modell, og inneholder en realistisk genetisk komponent. Den kan gi et estimat på ev. forandringer i ville bestander over tid for parametere som antall yngel, parr, smolt og voksen laks i bestanden, individuell vekst i de forskjellige livsstadier og kjønnsmodning. Ved bruk av modellen har vi dermed en mulighet til å estimere «biologiske» forandringer i ville bestander over tid som følge av forekomst av rømt oppdrettslaks på gyteplassene.
Analyser utført med IBSEM (Castellani mfl. 2018) viste først og fremst at jo flere rømt oppdrettslaks i gytebestanden, jo større genetisk innkryssing, og jo større genetiske forandringer i den ville bestanden (slik modellen til Hindar mfl. 2006 også viste). Modellen viste imidlertid at ved lav til moderat innslag av rømt oppdrettslaks er forandringen i den ville bestanden relativt liten og dermed vanskelig å kvantifisere på kort sikt. For eksempel, ved 5–10 % innslag av rømt oppdrettslaks på gyteplassene, viste de fleste fenotypiske og livshistorietrekk kun svake forandringer i en villaksbestand etter 50 år med genetisk innkryssing. Kun når innslag av rømt oppdrettslaks på gyteplassene ble økt til 30–50 % ble genetiske forandringer i bestandens fenotypiske og livshistorietrekk tydelige etter 50 år. Forfatterne konkluderte med at dette skyldes at (1) rømt oppdrettslaks har en lavere gytesuksess enn villaks i naturen (Fleming mfl. 1996, 2000), (2) fordi det er en sterk seleksjon mot avkom av oppdrettslaks i naturen (dvs., de har høyere dødelighet enn villaks og påvirker dermed bestandens «karakteristikk» mindre enn dersom de hadde overlevd i større grad), og (3) fordi mange livshistorietrekk i ville bestander er svært plastiske, og tetthetsavhengige. For eksempel, det er godt dokumentert at til tross for at oppdrettslaks vokser langt hurtigere enn vill laks under oppdrettsforhold (Glover mfl. 2017), er det kun funnet svake eller moderate forskjeller i vekst mellom disse gruppene i naturen (McGinnity mfl. 1999; Skaala mfl. 2014; Reed mfl. 2015; Jonsson & Jonsson 2017; Glover mfl. 2018). Det er dermed kanskje ikke overraskende at modellen indikerer at moderat innslag av rømt oppdrettslaks på gyteplassene kun fører til svake eller moderate forandringer i mange fenotypiske trekk.
IBSEM indikerer likevel at den største målbare effekten av innkryssing av rømt oppdrettslaks er at det kommer færre fisk tilbake fra havet (se også Hutchings 1991 og Liu mfl. 2013). En mulig forklaring er tetthetsavhengig dødelighet, og at en del av elvens produksjonskapasitet brukes til å produsere avkom av rømt og genetisk-påvirket laks, med høyere dødelighet i havet, og dermed reduseres antall laks som vandrer tilbake til elven. Det er likevel viktig å påpeke at modellen, under ulike scenarioer, viser at vedvarende innkryssing av rømt oppdrettslaks vil på sikt føre til en svekket bestand med redusert produksjon av laks av vill avstamning (Castellani mfl. 2018; Sylvester mfl. 2019; Bradbury mfl. 2020). Dette er forenlig med all tilgjengelig kunnskap om dette temaet (Glover mfl. 2017).
2.3 - Faktorer inkludert i risikovurderingen
En årlig risikovurdering av miljøproblemer knyttet til norsk fiskeoppdrett har vært gjennomført av Havforskningsinstituttet siden 2011 (Taranger mfl. 2015). Fram til 2018 ble risiko for genetiske endringer i ville laksebestander som følge av innkryssing av rømt oppdrettslaks vurdert basert på observerte innslag av rømt oppdrettslaks i elvene. Informasjon om andel rømt oppdrettslaks i elvene ble hentet fra den årlige rapporten fra det nasjonale overvåkingsprogrammet for rømt oppdrettslaks i vassdrag (se Glover mfl. 2019).
I 2019 ble det opprettet en ny tilnærming til risikovurderingen av norsk fiskeoppdrett (Grefsrud mfl. 2019). For utfordringen knyttet til rømming og genetisk påvirkning, ble den nye risikovurderingen designet for å vurdere den fremtidige risikoen for innkryssing av rømt oppdrettslaks i ville bestander. Ettersom mange ville bestander i Norge allerede er innkrysset med rømt oppdrettslaks (Glover mfl. 2013; Karlsson mfl. 2016; Diserud mfl. 2019b), ble vurderingen av risiko for innkryssing definert som risikoen for ytterligere genetisk endring hos villaks som følge av rømt oppdrettslaks. Risiko er beregnet per produksjonsområde og er basert på følgende faktorer ; andel rømt oppdrettslaks på gyteplassene (rømming, andel rømt oppdrettslaks i elv og utfisking/ fjerning av rømt oppdrettslaks fra elv) og bestandenes robusthet for ny innkryssing (bestandsstatus og genetisk status). H er presenterer vi en utvidet beskrivelse av disse faktoren, samt drøfter ulike biologiske faktorer som på nåværende tidspunkt ikke er inkludert i risikovurderingen. For resultater fra risikovurdering for ytterligere genetisk endring hos villaks som følge av rømt oppdrettslaks, se Grefsrud mfl. 2019 og Glover mfl. 2020.
2.3.1 - Rømt oppdrettslaks på gyteplassene
2.3.1.1 - Rømming
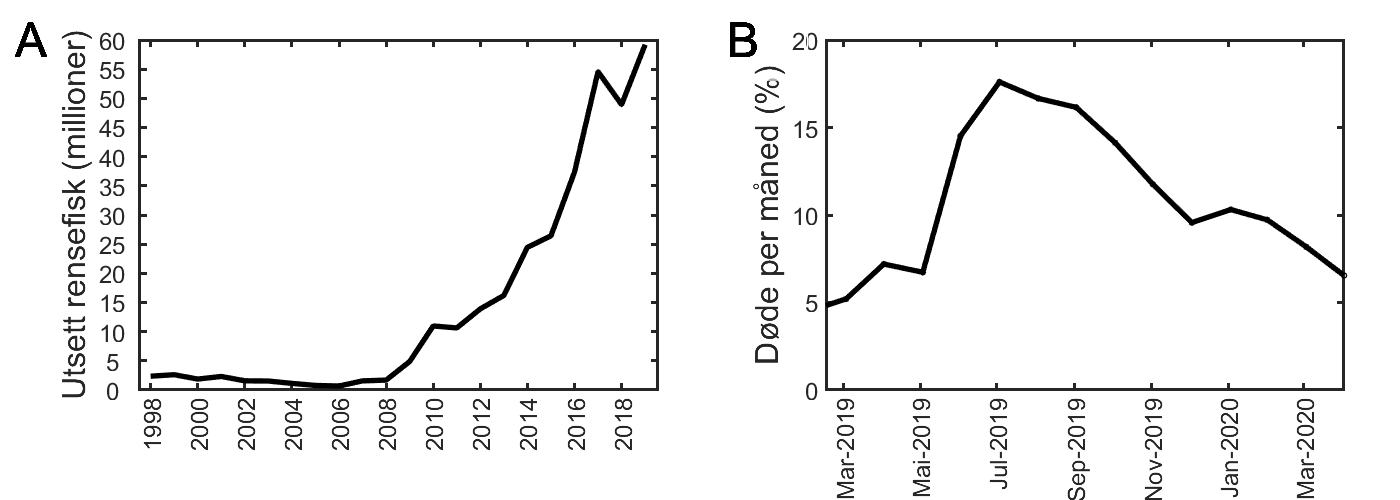
De offisielle innrapporterte rømmingstallene for laks (www.fiskeridir.no) viser at det siden 2001 årlig har rømt flere hundre tusen oppdrettslaks de fleste år (figur 2.2). I 2017 var det kun rapportert 17 000 rømt fisk som er det laveste nivået i hele perioden. Det høyeste rapporterte tallet var i 2006 da 921 000 fisk rømte. Disse tallene er minimumsestimater og de faktiske rømmingstallene er sannsynligvis høyere enn det som rapporteres. Havforskningsinstituttets DNA-identifisering av urapportert rømt oppdrettslaks (Glover 2010; Zhang mfl. 2013), og en større studie med utsetting av merket laks og modellering viste at de faktiske rømmingstallene for perioden 2005–2011 sannsynligvis var 2–4 ganger høyere enn den offisielle statistikken (Skilbrei mfl. 2015a). Det er ikke gjort tilsvarende studier for seinere år. Selv om det er usikkerhet i de offisielle rømmingstallene, er risikovurderingen basert på årlig gjennomsnittlig rapportert rømming per produksjonsområde i perioden 2014–2018 (Appendiks 1, tabell 1.1). I dette tidsrommet ble det meldt om totalt 747 931 * rømte oppdrettslaks.
Figur 2.2 Antall rømt oppdrettslaks rapportert årlig til Fiskeridirektoratet i perioden 2001–2020 og rapportert gjenfangst i perioden 2014–2019 (per 02.05.2020). Kilde: www.fiskeridir.no.
Biologisk status ved rømming påvirker oppdrettslaksens evne til å overleve i naturen, vandre opp i vassdragene og gyte sammen med villaks. Dette omfatter for eksempel livsfase, kjønn, kjønnsmodning, tidspunkt for rømming, lysregime på anlegget før rømming, sykdomsstatus, størrelse, alder og tidsforløp i det fri. Disse faktorene diskuteres her, men er på nåværende tidspunkt ikke inkludert i risikovurderingen da det finnes lite kunnskap om hvordan oppdrettslaksens biologiske status varierer mellom produksjonsområder.
Det er stor variasjon i spredning og overleving hos rømt oppdrettslaks, og rømt oppdrettslaks kan spre seg over store områder (Hansen mfl. 1993; Hansen 2006a, b; Jensen mfl. 2013; Quintela mfl. 2016). Årstid, rømmingslokalitet (eks. fjord vs. kyst), størrelse og alder ved rømming ser ut til å være avgjørende for hvor den rømte fisken svømmer og i hvilken grad de overlever.
Villaks legger ut på lange vandringer, fra elven til storhavet som liten smolt og tilbake som kjønnsmoden laks. Vandringene er synkronisert med årstidene, der smolt vandrer ut til havet i perioden april-juli, mens kjønnsmoden laks vandrer tilbake til elvene fra sent om våren til utpå høsten.
* I selve risikovurderingen står det at 730 179 laks rømte i tidsrommet 2014–2018. Dette skyldes i hovedsak at all data ble hentet ut fra Fiskeridirektoratet sine nettsider i desember 2018, og at rømmingstall for 2018 i etterkant ble noe oppjustert i 2019.
Også laks i oppdrett har disse vandringsinstinktene, men både årstid, størrelse og kjønnsmodningsstatus til fisken når den rømmer, har betydning for hvilken atferd den får i frihet. I noen tilfeller vil rømt oppdrettslaks spre seg hurtig og være lite fangbar, mens den under andre forhold vil holde seg lenge i området og kanskje søke opp i nærliggende elver.
Havforskningsinstituttet har gjennomført en serie med eksperimentelle slipp av oppdrettslaks for å studere ulike atferdsmønstre. Resultatene fra disse forsøkene er satt sammen med kunnskap fra andre kilder som også beskriver slipp av merket oppdrettslaks (Hansen 2006a, b; Chittenden mfl. 2011). Smolt og postsmolt som rømmer den første sommeren etter at de er satt ut i merder i sjøen, vil normalt vandre hurtig mot havet (Skilbrei 2010; Skilbrei mfl. 2015a). Dette innebærer at gjenfangst blir nærmest umulig. Denne fisken vokser opp i de åpne havområdene sammen med villaksen, og en liten andel (0,4 % gjenfangst fra våre utslipp) kommer tilbake til kysten ett til tre år senere som kjønnsmoden fisk. Våre undersøkelser viser at en høy andel av disse vil søke tilbake mot ferskvannskilder i området de rømte fra som smolt, men mange vandrer likevel opp i elver spredd over et område på flere hundre kilometer (Skilbrei mfl. 2015a; figur 2.3).
Vandringsviljen til postsmolt som rømmer om høsten reduseres i takt med at dagene blir kortere. Derfor øker også den lokale gjenfangsten av postsmolt som rømmer utover høsten (13 % gjenfangst fra våre utslipp), blant annet fordi de er blitt store nok til å bli fanget i garn og av sportsfiskere (Skilbrei mfl. 2015a).
Dersom fisk rømmer kort tid etter at de har blitt flyttet fra kar på et settefiskanlegg til merd i sjøen, blir vandringsmotivasjonen påvirket av lysregimet fisken var utsatt for på settefiskanlegget. Kontinuerlig belysning svekker vandringsviljen og kan få stor settefisk til å holde seg i fjorden uvanlig lenge, selv om de rømmer om sommeren (Skilbrei mfl. 2014). I produksjon av høstsmolt kan bruk av kunstig økt daglengde på settefiskanlegget få laksen til å legge ut på vandring mot havet selv om den rømmer fra merden seint om høsten. Sjansen for at disse overlever vinteren i havet er imidlertid lav (Skilbrei 2013), da slipp av høstsmolt ga lavere gjenfangst som voksen (0,004 %) en slipp av vårsmolt (0,17 %). Det er dermed lavere sannsynlighet for at fisk fra denne type rømming overlever fram til kjønnsmodning og vandrer opp i elv.
Gjenfangst av voksen laks avhenger av rømmingsområdet. Slipp av voksen laks fra anlegg på kysten har gitt lavere gjenfangst (4–7 %) enn slipp i fjorder der fisken oppholder seg lenger og er mer eksponert for fiskeredskaper (7–33 %). Slipp fra anlegg helt ute i havgapet har ikke gitt gjenfangster (Skilbrei mfl. 2015a). Flesteparten av gjenfanget voksen laks ble fanget i månedene etter at de ble satt ut, i nærheten av utslippsområdet. Kun 0,09 % ble gjenfanget 1–2 år etter utslippet (Skilbrei mfl. 2015a). En medvirkende årsak til dette er muligens at voksen laks som rømmer har vansker med å lære seg å fange naturlig føde. Undersøkelser av mageinnhold til voksen rømt oppdrettslaks fanget nær norskekysten viser vanligvis at de aller fleste er tomme (Olsen & Skilbrei 2010). Rømt oppdrettslaks fanget under fisket ved Færøyene har derimot samme diett som villaksen der (Jacobsen & Hansen 2001).
Figur 2.3 Gjenfangst av voksen laks i sjø (gule sirkler) og elv (rød) fra slipp av smolt fra forskningsstasjonen Matre (R) i 2005–2010. Fisk gjenfanget < 6 km fra utsettingsstedet (49 % av rapportert gjenfangst) er ikke vist. Størrelsen på sirklene angir antall fisk fra 1 til 5 individer.
Rømminger om høsten, uavhengig av alder på laksen, kjennetegnes ofte av at mange fisk søker mot ferskvannskilder, både elver og kraftverk som slipper ut turbinvann. Dette er som ventet når det gjelder kjønnsmodnende fisk, men det har vist seg at mange umodne laks også søker inn mot elveosene og at noen også går opp i nedre del av elven (Madhun mfl. 2015). Det er kjent at umoden laks kan gå opp i ferskvann (Webb mfl. 2007), men denne atferden er ikke godt kartlagt. Det er mulig at noen av de umodne fiskene svømmer ut av elven igjen etter en stund, mens andre kan stå i elven til de blir kjønnsmodne.
Som nevnt over har merkestudiene vist at smolt og postmolt som rømmer kan komme tilbake som gyteklar laks etter 1–3 år i havet, og at voksen rømt oppdrettslaks som gjenfanges som oftest blir gjenfanget i løpet av det året de rømte (Skilbrei mfl. 2015a). Undersøkelser av et pigment i kjøtt som reflekterer ulik diett mellom oppdrettsmiljøet og naturen, viste tidlig på 90-tallet at om lag halvparten av den undersøkte rømte oppdrettslaksen fanget i elv hadde rømt nylig mens den resterende halvparten hadde tilbrakt mer enn ett år i naturen (Lura & Sægrov 1994). Senere har fettsyreanalyser av voksen rømt oppdrettslaks i elv vist at flertallet nylig hadde rømt, dette basert på at de hadde fettsyreprofiler som var svært lik oppdrettsfôret. En mindre andel på rundt 10–30 % av den voksne rømte oppdrettslaksen hadde fettsyreprofiler som tydet på at den hadde beitet i havet i lang tid og dermed sannsynligvis hadde rømt som smolt eller postsmolt. Det var færre observasjoner av voksen laks som hadde spist begge typer mat; som hadde rømt som relativt stor laks og deretter klart å finne ville byttedyr (Skilbrei mfl. 2015b; Anon. 2018). Dette støtter antakelsen fra merkestudiene om at en stor del av den umodne voksne oppdrettslaksen som rømmer sannsynligvis ikke overlever fram til den blir kjønnsmoden.
Tilgjengelige data tyder på at sannsynligheten for at en rømt oppdrettslaks overlever fram til kjønnsmodning og vandrer opp i elv er størst hvis fisken rømmer som smolt om sommeren, eller det samme året den blir kjønnsmoden. Sannsynligheten er minst for smolt som rømmer om høsten, og umoden laks som ikke blir kjønnsmoden før tidligst neste år.
2.3.1.2 - Andel rømt oppdrettslaks i elv
Siden oppstarten av lakseoppdrett i Norge har flere millioner oppdrettslaks rømt fra en rekke anlegg langs norskekysten. De fleste av disse fiskene «blir borte» i det marine miljøet uten videre spor, men noen vil vandre opp i lakseelver. Det nasjonale overvåkingsprogrammet for rømt oppdrettslaks i vassdrag, som ble utformet og etablert på oppdrag fra Fiskeridirektoratet etter føringer fra Nærings- og fiskeridepartementet i 2014, beregnet innslag av rømt oppdrettslaks i henholdsvis 140 vassdrag i 2014 (Anon. 2015a) og 165 vassdrag i 2015 (Anon. 2016a). Overvåkingen ble videre utvidet til å beregne innslag av rømt oppdrettslaks i 196 vassdrag i 2016 (Anon. 2017a), 197 vassdrag i 2017 (Anon. 2018) og 205 vassdrag i 2018 (Anon. 2019a).
Vassdragene som er overvåket er valgt ut fra en rekke kriterier; god geografisk spredning, inkludering av de nasjonale laksevassdragene, representasjon av vassdrag av ulik størrelse samt å bygge videre på vassdrag med tidsserier og med gode lokale nettverk, som er etablert i ulike prosjekter finansiert av Norges forskningsråd og miljømyndighetene siden 1989 (Fiske mfl. 2006; Diserud mfl. 2019a). Data blir samlet inn fra sportsfiske om sommeren, høstfiske, stamfiske og drivtellinger om høsten. De tre førstnevnte metodene er i hovedsak basert på stangfiske og skiller mellom rømt oppdrettslaks og villaks ved å undersøke fiskens skjell, som gir et bilde av fiskens vekstbetingelser gjennom i livet. Drivtellinger innebærer at snorklere foretar en visuell inspeksjon av fisk i elven, teller opp og karakteriserer vill og rømt oppdrettet laks på basis av utseende og atferd. I mange av elvene som blir undersøkt blir mer enn én metode benyttet. Innsamlete data går gjennom en kvalitetssikringsprosess og blitt enkeltvis vurdert i henhold til en rekke kriterier for å få en total vurdering av dataenes representativitet. Innslaget av rømt oppdrettslaks for hver elv presenteres i en årlig rapport fra overvåkingsprogrammet som prosentandelene registrert ved de ulike metodene, samt som en årsprosent som beregnes fra andel oppdrettslaks i sportsfiske og/eller høstfiske/stamfiske (Fiske mfl. 2006). Årsprosenten tar hensyn til at sportsfisket sannsynligvis gir et for lavt, og høstfisket sannsynligvis et for høyt estimat av innslaget av rømt oppdrettslaks. De ulike metodene som blir benyttet i de forskjellige elvene har sine styrker og svakheter, både i forhold til prøvestørrelsene og sikker identifikasjon av rømt oppdrettslaks. At innslaget av rømt oppdrettslaks i vassdragene kan endre seg i løpet av sesongen og at rømt oppdrettslaks til dels har en annen atferd enn villaks, gjør det krevende både å innhente gode data og å sammenligne data innhentet med de ulike metodene. Ved utregning av et estimat for prosentvis andel oppdrettslaks i elven, kommer det i tillegg en statistisk usikkerhet på anslaget som avhenger av prøvestørrelsen og innslaget av rømt oppdrettslaks. Ulike kilder til usikkerhet i dataene fra overvåkingsprogrammet blir diskutert i rapporten (Anon. 2019a).
Elver blir klassifisert i kategoriene <4, 4–10 og >10 %-, tilsvarende systemet foreslått av Taranger mfl. (2012). Klassifiseringen følger ikke årsprosent slavisk, men er basert på en samlet vurdering av alle datakildene:
Lavt innslag : innslag av rømt oppdrettslaks er estimert til under 4 %.
Moderat innslag : innslag av rømt oppdrettslaks er estimert til mellom 4 % og 10 %.
Høyt innslag : innslag av rømt oppdrettslaks er estimert til over 10 %.
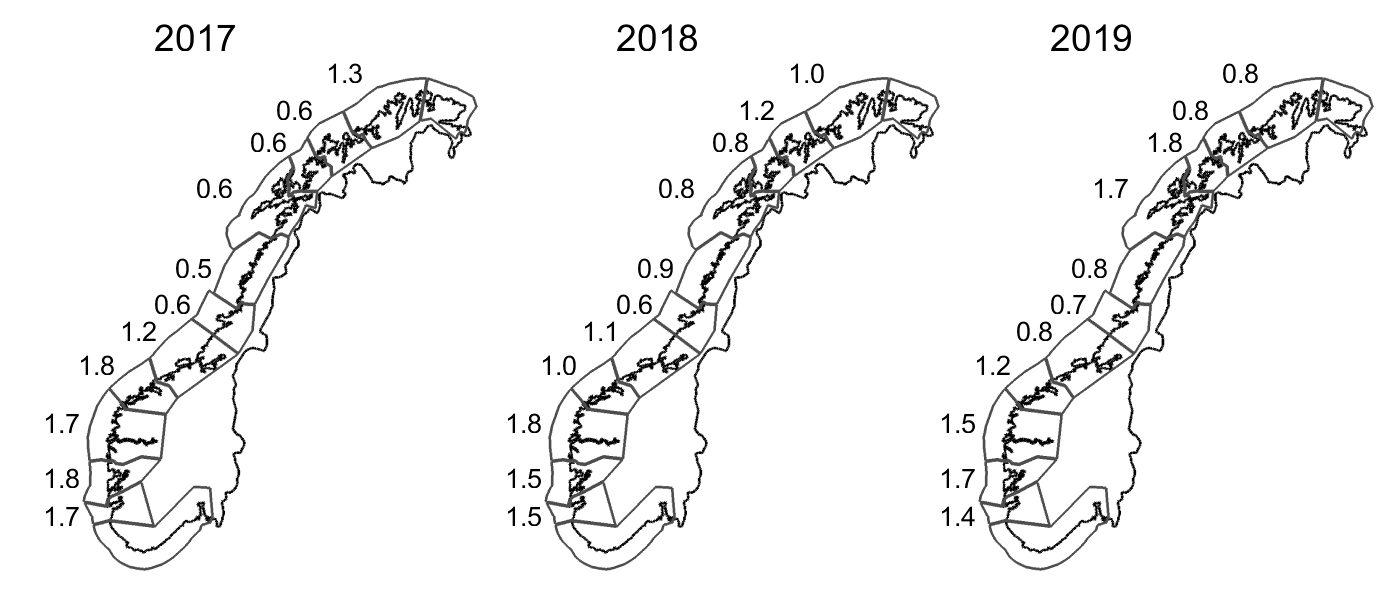
Resultatene fra overvåkingsprogrammet er presentert på to nivåer. Hovedrapporten representerer en oppsummering av hovedresultatene, og viser hvilke metoder som er lagt til grunn (Anon. 2019a) og er publisert elektronisk sammen med vedleggsdokumenter som viser detaljerte resultater for hvert vassdrag. Disse vedleggsdokumentene er organisert fylkesvis og omfatter mer enn 800 sider til sammen ( https://www.hi.no/hi/nettrapporter/fisken-og-havet-2019-4 ). I risikovurderingen har vi brukt klassifiseringene overvåkingsprogrammet har gjort for de enkelte vassdrag i perioden 2014–2017 innenfor hvert av produksjonsområdene og vurdert disse samlet for å kategorisere tilstanden innenfor hvert produksjonsområde (se Appendiks 1, tabell 1.2).
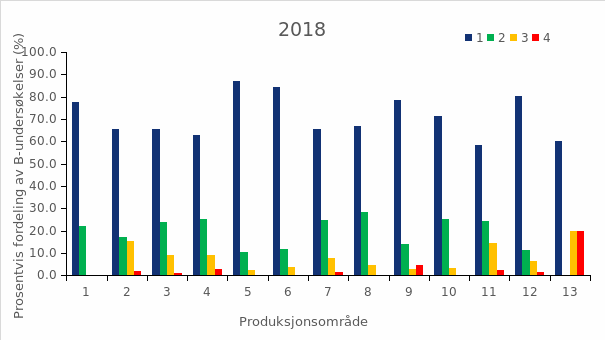
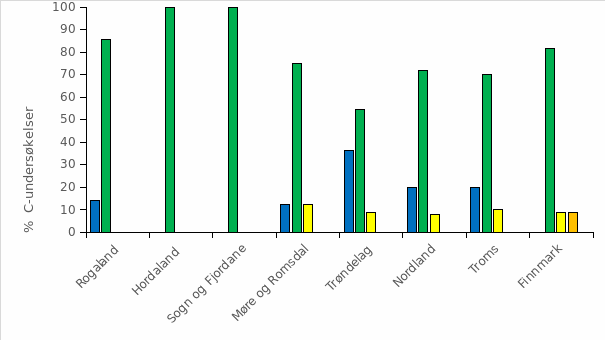
Det ble i 2018 beregnet årsprosent for 123 elver, og det presenteres i rapporten fra overvåkingsprogrammet data fra drivtellinger fra 122 elver. Resultater fra overvåkingsprogrammet for 2018 viste at til sammen 153 elver (75 %) ble vurdert til å ha lavt innslag av rømt oppdrettslaks (mindre enn 4 % innslag), 33 vassdrag (16 %) ble vurdert til å ha moderat innslag (mellom 4 og 10 % innslag), mens 19 (9 %) vassdrag ble vurdert til å ha et høyt innslag av rømt oppdrettslaks (over 10 %). Det var en økning i både antall og andel elver med høyt innslag i 2018 sammenlignet med 2017 (15 vassdrag, 8 %). Innslaget av rømt oppdrettslaks varierte langs norskekysten. I Hardangerfjorden hadde mange av vassdragene høyt innslag av rømt oppdrettslaks, som i tidligere år. Det var også vassdrag med høyt innslag av rømt oppdrettslaks i Møre og Romsdal, Trøndelag, Nordland og Troms. Tilstanden var derimot god, med lave innslag av rømt oppdrettslaks, på hele strekningen fra Østfold til Rogaland (figur 2.4).
Figur 2.4 Lokalisering av elvene der innslaget av rømt oppdrettslaks i 2018 er vurdert av overvåkingsprogrammet til å være lavt (< 4 %, grønne sirkler), moderat (4 – 10 %, gule sirkler), eller høyt (>10 %, røde sirkler) . Se teksten for nærmere forklaring av de tre kategoriene. Fra Anon. (2019a).
Andelen rømt oppdrettslaks i elvene har endret seg mellom år, og det har vært en synkende tendens i registreringene gjennom de siste årene. Gjennomsnittlig innslag av rømt oppdrettslaks for de undersøkte elvene har variert mellom 3 og 16 % (tilsvarer en beregnet årsprosent mellom 3 og 10 %), med en synkende trend som er signifikant over tid (figur 2.5). Nivået av rømt oppdrettslaks nådde i 2018 det laveste nivået siden 2006. Størstedelen av nedgangen har skjedd etter 2013 og er betydelig, med midlere innslag i 2018 på kun en fjerdedel av det som var typisk opp til 2013.
Se Diserud mfl. (2019a) for en oppsummering av resultatene fra overvåkingen før 2014, og Glover mfl. (2019) for en beskrivelse av det nåværende overvåkingsprogrammets aktiviteter og resultater.
Figur 2.5 Gjennomsnittlig % rømt oppdrettslaks (o) i høstundersøkelsene for årene 2006–2018 for totalt 69 elver (Fiske 2013, Fiske mfl. 2014; Anon. 2015a, 2016a, 2017a, 2018, 2019a), vist for hele Norge. Prediksjoner basert på modell er vist med tykk linje (signifikant nedgående trend). Se Anon. (2019a), for mer detaljer.
2.3.1.3 - Utfisking/fjerning av rømt oppdrettslaks fra elv
Data fra det nasjonale overvåkingsprogrammet kommer til direkte anvendelse gjennom utfiskingsforskriften som Nærings- og fiskeridepartementet vedtok i 2015 ( Forskrift om fellesansvar for utfisking mv. av rømt oppdrettsfisk), der oppdrettsnæringens sammenslutning for utfisking av rømt oppdrettsfisk (OURO) er hjemlet. Her er det fastsatt at det skal gjøres tiltak for å redusere mengde rømt oppdrettslaks i elver med mer enn 10 % innslag av oppdrettslaks, dokumentert gjennom overvåkingsprogrammet. Utføring av pålagte oppgaver fra forskriften ble satt i verk av OURO i 2016. Det ble gjennomført tiltak i 37 elver i 2016, 52 elver i 2017, 63 elver i 2018 og 37 elver i 2019. Det gjennomføres således utfisking i flere elver enn de som er vurdert til å ha > 10 % innslag av rømt oppdrettslaks. I tillegg til utfiskingstiltak organisert gjennom OURO, organiserer også Fiskeridirektoratet uttaksfiske som avbøtende tiltak ved akutte rømmingsepisoder og i vassdrag hvor mye oppdrettslaks blir observert. For nærmere informasjon om utfiskingstiltakene og gjennomføring i de ulike vassdragene se også rapporter fra aktører som har deltatt i fisket på OURO sine nettsider (www.utfisking.no).
Antall rømt oppdrettslaks fjernet fra elver av OURO i hvert produksjonsområde i perioden 2016–2017 utgjør datagrunnlaget for vurdering av denne faktoren i risikovurderingen (se Appendiks 1, tabell 1.3).
2.3.2 - Bestandenes robusthet for ny innkryssing
Det er mange biologiske faktorer (bestandens størrelse, tetthet, livshistoriekarakteristikk og timing av kritiske hendelser som gyting) og fysiske faktorer (elvegradient og lengde, temperaturforhold, substrat, vandringshinder) som, i tillegg til forekomst av rømt oppdrettslaks, vil påvirke graden av genetisk innkryssing og konsekvensene for den ville bestanden. Pågående arbeid, blant annet i QuantEscape II (NFR-prosjekt 2016-2020), vil gi en bedre forståelse for risikofaktorer knyttet til sannsynligheten for genetisk innkryssing.
Noen faktorer har allerede vært undersøkt i forbindelse med 5 og 10-årsevalueringen av ordningen med nasjonale laksevassdrag, og -fjorder, hvor en bl.a. har sett på hvilke typer elver som tiltrekker seg rømt fisk (Fiske mfl. 2013; Hindar mfl. 2018). Studiene viser at økt vannføring, større laksebestand og økt oppdrettsintensitet i regionen, øker antallet rømt oppdrettslaks i elven. Andelen rømt oppdrettslaks øker også med økt vannføring og oppdrettsintensitet i regionen, men avtar med økende bestandsstørrelse av villaks. At antall/andel rømt oppdrettslaks i et vassdrag har en sammenheng med nærhet til og omfanget av oppdrett i nærheten sammenfaller med tidligere analyser (Fiske mfl. 2006) og er også dokumentert i andre land (Keyser mfl. 2018). Høy vannføring (og stor laksebestand) ansees å være viktig fordi det tiltrekker rømt oppdrettslaks.
Hvor stor gytesuksess voksen rømt oppdrettslaks har på gyteplassene er avhengig av hvor mange ville konkurrenter de har. Rømt oppdrettslaks har generelt sett en lavere gytesuksess enn villaks (Fleming mfl. 1996, 2000), og et gitt innslag av rømt oppdrettslaks på gyteplassene vil derfor ikke automatisk føre til en tilsvarende prosentvis genetisk innkryssing. Det er derimot grunn til å tro at oppdrettslaksens gytesuksess vil variere i tid og rom, avhengig av blant annet hvor lenge den har vært i havet (Fleming mfl. 1996, 1997) og konkurransen den møter på gyteplassen med vill fisk (Glover mfl. 2012). Derfor vil også genetisk innkryssing kunne variere i tid og mellom bestander i ulike vassdrag. Likevel er det dokumentert en viss sammenheng mellom observert andel rømt oppdrettslaks og beregnet genetisk innkryssing på vassdragsnivå. I et arbeid med 20 vassdrag, fant Glover mfl. (2013) en sammenheng mellom beregnet forekomst av rømt oppdrettslaks over tid og beregnet innkryssing. De fant at 47 % av variansen i genetisk innkryssing ble forklart av andel rømt oppdrettslaks observert (R 2 = 0,47). Dette ble bekreftet i en mer omfattende analyse av det samme datagrunnlaget av Heino mfl. (2015) som også tok hensyn til bestandsstørrelse (R 2 = 0,51).
I 2016 publiserte Karlsson mfl. (2016) et arbeid som viste at forekomst av rømt oppdrettslaks i perioden 1989–2012 (gjennomsnittlig årsprosent, etter Diserud mfl. 2013) forklarte 24 % av variansen i genetisk innkryssing i 77 undersøkte vassdrag. Da forfatterne utførte analysen på et regionalt nivå, økte forklaringsstyrken til 56 %. Oppsummert viser disse analysene at selv om det er en sammenheng mellom andel rømt oppdrettslaks i bestanden og genetisk innkryssing (figur 2.6), så er det også andre faktorer i elven som har betydning.
Figur 2.6 Forholdet mellom gjennomsnittlig årsprosent (andel) av rømt oppdrettslaks i perioden 1989–2012 og beregnet genetisk innkryssing av rømt oppdrettslaks i 77 villaksbestander. Røde diamanter er elver i Sør-Norge (fra Østfold til Sør-Troms), blå er elver i Finnmark, grå er i «overgangssonen» mellom de to regionene, og åpne symboler er regionsvise gjennomsnitt. De stiplete linjene er regresjonslinjen for henholdsvis Sør-Norge (rød) og Finnmark (blå). Figuren er fra Karlsson mfl. (2016).
2.3.2.1 - Villaksens bestandsstatus
Antall gytelaks som trengs for å utnytte elvens produksjonspotensial kalles gytebestandsmål (Hindar mfl. 2007). Bestander som både når gytebestandsmålet og har et høyt produksjonspotensial er trolig mer robuste mot innkryssing av rømt oppdrettslaks enn bestander som har lite produksjonspotensial og/eller som ikke når gytebestandsmålet.
Oppnåelse av gytebestandsmål og høstingspotensial til bestandene beregnes årlig av Vitenskapelig råd for lakseforvaltning (VRL), og brukes her som en indikator for villaksbestandens robusthet for ny innkryssing av rømt oppdrettslaks. I «Kvalitetsnorm for ville bestander av laks ( Salmo salar )», delnorm gytebestandsmål og høstingspotensial , blir bestandene av VRL delt inn i kategoriene: «svært god», «god», «moderat», «dårlig» og «svær dårlig» kvalitet (Anon. 2019b).
Dersom bestandene når sine gytebestandsmål, har de mange nok ville gytelaks på gyteplassene til å kunne utnytte elvas produksjonspotensial. Konkurransen på gyteplassen blir enda sterkere dersom flere villaks enn gytebestandsmålet er til stede på gyteplassen. Bestander med høyt høstingspotensial (vesentlig flere voksenlaks kommer tilbake enn det som er nødvendig for å nå gytebestandsmålet) har også større sannsynlighet for å nå gytebestandsmålet i årene som kommer, selv om overlevelsen i havet skulle bli redusert. Konkurransen mellom ungfiskene i elven vil også være større hvis det er mange fisk som gyter i vassdraget, og avkom av rømt oppdrettslaks vil gjøre det relativt sett dårligere hvis det er mange ville ungfisk å konkurrere med (Skaala et al. 2012). Det antas derfor at bestander som både når gytebestandsmålet og har et høyt produksjonspotensial er mer robuste mot innkryssing av rømt oppdrettslaks enn bestander som har lite produksjonspotensial og/eller som ikke når gytebestandsmålet.
I risikovurderingen er vurderingen av villaksens bestandsstatus i produksjonsområdene basert på beregninger av måloppnåelse for gytebestandsmål og høstingspotensial (Anon. 2014b, 2015b, 2016b, 2017b) for den enkelte villaksbestand i perioden 2014–2017 (se Appendiks, tabell 1.4).
2.3.2.2 - Villaksens genetiske status
Rømt oppdrettslaks og deres avkom har antakeligvis større suksess i konkurranse med innkryssede individer enn med ikke-innkrysset villaks. Det er derfor sannsynlig at høy innkryssing av oppdrettslaks i bestandene vil gjøre dem mindre robuste for innkryssing av rømt oppdrettslaks i framtiden enn bestander som har liten grad av innkryssing.
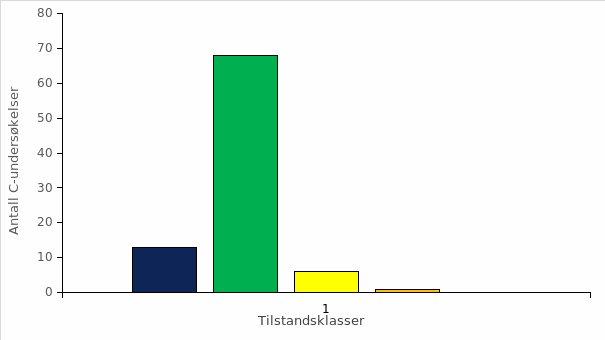
NINA og Havforskningsinstituttet har i samarbeid produsert et omfattende sett med estimater for tidligere genetisk innkryssing av rømt oppdrettslaks i 225 bestander (Diserud mfl. 2019b), som representerer om lag 94 % av villaksressursene i Norge (beregnet som andel av det totale gytebestandsmålet). Resultatene viser at oppdrettslaks er krysset inn i et stort antall elver: store genetiske forandringer er påvist i 67 vassdrag (> 10 % endring), moderate genetiske forandringer er påvist i 16 vassdrag (4-10 % endring), svake genetiske forandringer er indikert i 67 vassdrag (1-4 % endring) og ingen genetiske forandringer er observert i 75 vassdrag (< 1 % endring) (figur 2.7). Dette viser at genetisk innkryssing forekommer i en stor del av elvene i Norge da 2/3 deler av bestandene er plassert i kategoriene svake genetisk forandringer indikert til store genetisk forandringer dokumentert.
Det er utarbeidet kvalitative og kvantitative kriterier for hver av de fire tilstandsklassene, og i alt er det undersøkt om lag 40 000 villaks for å beskrive genetisk innkryssing i ville laksebestander. Beskrivelsen av genetisk status utgjør delnorm genetisk integritet til «Kvalitetsnorm for ville bestander av laks ( Salmo salar )» og første statusrapport ble publisert for 125 elver i 2016 (Diserud mfl. 2016).
Over tid vil innkryssing av rømt oppdrettslaks kunne forandre egenskapene til de ville laksebestandene, redusere antall villaks som produseres og svekke bestandenes evne til å tilpasse seg endringer i miljøet. Innkryssing av rømt oppdrettslaks vil derfor både kunne svekke bestandene, og dermed gjøre dem mindre robuste mot framtidig innkryssing av rømt oppdrettslaks. I risikovurderingen blir den genetiske påvirkningen av rømt oppdrettslaks på ville laksebestander oppsummert per produksjonsområde (Appendiks 1, tabell 1.5).
Figur 2.7 Genetisk status i 225 laksebestander i forhold til kvalitetselementet genetisk integritet. Ingen genetisk endring ble observert i 75 bestander, svake genetiske endringer ble indikert i 67 bestander, moderate genetiske endringer ble påvist i 16 bestander, mens store genetiske endringer ble påvist i 67 bestander. For mer utfyllende forklaring av de fire kategoriene se Diserud mfl. (2019b) der figuren er hentet fra.
Til tross for at det nå er beregnet innkryssingsnivå av rømt oppdrettslaks i 225 vassdrag i Norge, er det i liten grad publisert tilsvarende estimater i andre land med lakseoppdrett. I nyere tid er det derimot publisert flere studier fra Canada (Sylvester 2018, 2019; Wringe 2018) og det pågår arbeid i andre land. Norge fremstår dermed som et globalt kunnskapssenter på denne problemstillingen, da en slik omfattende oversikt ikke foreligger i andre land.
2.4 - Referanser
Abrantes KG, Lyle JM, Nichols PD, Semmens JM. 2011. Do exotic salmonids feed on native fauna after escaping from aquaculture cages in Tasmania, Australia? Canadian Journal of Fisheries and Aquatic Sciences 68, 1539–51.
Anon. 2015a. Rømt oppdrettslaks i vassdrag. Rapport fra det nasjonale overvåkingsprogrammet 2014. Fisken og havet, særnr. 2b–2015.
Anon. 2016a. Rømt oppdrettslaks i vassdrag. Rapport fra det nasjonale overvåkingsprogrammet 2015. Fisken og havet, særnr. 2b–2016.
Anon. 2017a. Rømt oppdrettslaks i vassdrag. Rapport fra det nasjonale overvåkingsprogrammet 2016. Fisken og havet, særnr. 2b–2017.
Anon. 2018. Rømt oppdrettslaks i vassdrag. Rapport fra det nasjonale overvåkingsprogrammet 2017. Fisken og havet, særnr. 2–2018.
Anon. 2019a. Rømt oppdrettslaks i vassdrag. Rapport fra det nasjonale overvåkingsprogrammet 2018. Fisken og havet, nr. 2019–4.
Anon. 2014b. Status for norske laksebestander i 2014. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 6, 225 s.
Anon. 2015b. Status for norske laksebestander i 2015. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 8, 300 s.
Anon. 2016b. Status for norske laksebestander i 2016. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 9, 190 s.
Anon. 2017b. Status for norske laksebestander i 2017. Rapport fra Vitenskapelig råd for lakseforvaltning nr. 10, 152 s.
Araki H, Schmid C. 2010. Is hatchery stocking a help or harm? Evidence, limitations and future directions in ecological and genetic surveys. Aquaculture, 38 (Supp. 1): 2–11.
Besnier F, Glover KA, Skaala Ø. 2011. Investigating genetic changes in wild populations: modelling gene-flow from farm escapees. Aquaculture Environment Interactions 2: 75–86.
Besnier F, Glover KA, Lien S, Kent M, Hansen MM, Shen X, Skaala Ø. 2015. Identification of quantitative genetic components of fitness variation in farmed, hybrid and native salmon in the wild. Heredity 00:1-9.
Besnier F, Solberg MF, Harvey AC, Carvalho GR, Bekkevold D, Taylor MI, Creer S, Nielsen EE, Skaala Ø, Ayllon F, Dahle G & Glover KA (2020) Epistatic regulation of growth in Atlantic salmon revealed: a QTL study performed on the domesticated-wild interface. BMC Genetics21 .
Bicskei B, Bron J, Glover KA, Taggart J B. 2014. A comparison of gene transcription profiles of domesticated and wild Atlantic salmon ( Salmo salar L.) at different life stages, reared under controlled conditions. BMC Genomics 15:884.
Bicskei B, Taggart JB, Glover KA, Bron JE. 2016. Comparing the transcriptomes of embryos from domesticated and wild Atlantic salmon ( Salmo salar L.) stocks and examining factors that influence heritability of gene expression. Genetics Selection Evolution, 48(1):1-16.
Bolstad GH, Hindar K, Robertsen G, Jonsson B, Sægrov H, Diserud OH, Fiske P, Jensen AJ, Urdal K, Næsje TF. 2017. Gene flow from domesticated escapees alters the life history of wild Atlantic salmon. Nature Ecology & Evolution, 1: 0124.
Bourke EA, Coughlan J, Jansson H, Galvin P, Cross TF. 1997. Allozyme variation in populations of Atlantic salmon located throughout Europe: diversity that could be compromised by introductions of reared fish. ICES Journal of Marine Science 54: 974–985.
Bourret V, Kent MP, Primmer CR, Vasemägi A, Karlsson S, Hindar K, McGinnity P, Verspoor E, Bernatchez L, Lien S. 2013. SNP-array reveals genome-wide patterns of geographical and potential adaptive divergence across the natural range of Atlantic salmon ( Salmo salar ). Molecular Ecology 22: 532–551.
Bradbury IR, Duffy S, Lehnert SJ, Johannsson R, Fridriksson JH, Castellani M. Burgetz I, Sylvester E, Messmer A, Layton K, Kelly N, Dempson JB & Fleming I.A. (2020) Model-based evaluation of the genetic impacts of farm-escaped Atlantic salmon on wild populations. Aquaculture Environment Interactions12 , 45–59.
Castellani M, Heino M, Gilbey J, Araki H, Svåsand T, Glover KA. 2015. IBSEM: An individual-based Atlantic salmon population model. PLOS One 10(9): e0138444.
Castellani M, Heino M, Gilbey J, Araki H, Svåsand T & Glover K (2018) Modeling fitness changes in wild Atlantic salmon populations faced by spawning intrusion of domesticated escapees. Evolutionary Applications , 1–16.
Chittenden CM, Rikardsen AH, Skilbrei OT, Davidsen JG, Halttunen E, Skardhamar J, McKinley RS. 2011. An effective method for the recapture of escaped farmed salmon. Aquaculture Environ Interact 2011, 1(3):215-224.
Clifford SL, McGinnity P, Ferguson A. 1998a. Genetic changes in Atlantic salmon ( Salmo salar ) populations of northwest Irish rivers resulting from escapes of adult farm salmon. Canadian Journal of Fisheries and Aquatic Sciences 55: 358–363.
Clifford SL, McGinnity P, Ferguson A. 1998b. Genetic changes in an Atlantic salmon population resulting from escaped juvenile farm salmon. Journal of Fish Biology 52: 118–127.
Crozier WW. 1993. Evidence of genetic interaction between escaped farmed salmon and wild Atlantic salmon ( Salmo salar L) in a Northern Irish river . Aquaculture 113 (1-2):19-29.
Crozier WW. 2000. Escaped farmed salmon, Salmo salar L., in the Glenarm River, Northern Ireland: genetic status of the wild population 7 years on. Fisheries Management and Ecology, 7: 437–446.
Darwish TL, Hutchings JA. 2009. Genetic variability in reaction norms between farmed and wild backcrosses of Atlantic salmon ( Salmo salar ). Canadian Journal of Fisheries and Aquatic Sciences 66: 83–90.
Debes PV, Hutchings JA. 2014. Effects of domestication on parr maturity, growth, and vulnerability to predation in Atlantic salmon. Canadian Journal of Fisheries and Aquatic Sciences 71 , 1371–84.
Diserud OH, Fiske P, Hindar K. 2013. Forslag til kategorisering av laksebestander som er påvirket av rømt oppdrettslaks – Oppdatering for perioden 1989–2012. – NINA Rapport 976. 22 s.
Diserud OH, Hindar K, Karlsson S, Glover K, Skaala Ø. 2016. Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander. Vedlegg Notat NINA/HI, s. 53–85 i Anon. 2016. Klassifisering av 104 laksebestander etter kvalitetsnorm for villaks. Temarapport nr. 4. Vitenskapelig råd for lakseforvaltning.
Diserud, OH, Hindar, K, Karlsson, S, Glover, KA, Skaala, Ø. 2017. Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander – status 2017. NINA Rapport 1337. 55 s.
Diserud OH, Fiske P, Sægrov H, Urdal K, Aronsen T, Lo H, Barlaup BT, Niemela E, Orell P, Erkinaro J, Lund RA, Økland F, Østborg GM, Hansen LP, Hindar K. 2019a Frequency of escapees in Norwegian rivers 1989–2013. Ices Journal of Marine Science 76, 1140–50.
Diserud OH, Hindar K, Karlsson S, Glover KA, Skaala Ø. 2019b. Genetisk påvirkning av rømt oppdrettslaks på ville laksebestander – oppdatert status 2019. NINA Rapport 1659. 72 s.
Einum S, Fleming IA. 1997. Genetic divergence and interactions in the wild among native, farmed and hybrid Atlantic salmon. Journal of Fish Biology 50: 634–651.
Ferguson A, McGinnity P, Baker N, Cotter D, Hynes R, O’Hara B, O’Maoileidigh N, Prodöhl P, Rogan G. 2002A two-generation experiment comparing the fitness and life-history traits of native, ranched, non-native, farmed, and hybrid Atlantic salmon under natural conditions. ICES CM 2002/T:04.
Fiske P, Lund, R A, Hansen, LP. 2006. Relationships between the frequency of farmed Atlantic salmon, Salmo salar L., in wild salmon populations and fish farming activity in Norway 1989–2004. ICES J Mar Sci 63: 1182–1189.
Fiske P, Diserud,OH, Robertsen G, Foldvik A, Skilbrei OT, Heino M, Helland IP, Hindar K. 2013. Midtveisvurdering av nasjonale laksevassdrag og nasjonale laksefjorder. Rømt oppdrettslaks og bestandsstatus. NINA Minirapport, 470: 1–24.
Fiske P. 2013. Overvåking av rømt oppdrettslaks i elv om høsten 2010–2012. NINA Rapport 989.
Fiske P, Aronsen T, Hindar K. 2014. Overvåking av rømt oppdrettslaks i elver om høsten 2013. NINA rapport 1063. 44 s.
Fjelldal PG, Glover KA, Skaala Ø, Imsland A, Hansen TJ. 2009. Vertebral body mineralization and deformities in cultured Atlantic salmon ( Salmo salar L.): Effects of genetics and off-season smolt production. Aquaculture 296: 36–44.
Fleming IA, Jonsson B, Gross MR, Lamberg A. 1996. An experimental study of the reproductive behaviour and success of farmed and wild Atlantic salmon ( Salmo salar ). J Appl Ecol 33:893–905.
Fleming IA, Lamberg A, Jonsson B. 1997. Effects of early experience on the reproductive performance of Atlantic salmon. Behav Ecol 8:470–480.
Fleming IA, Einum S. 1997. Experimental tests of genetic divergence of farmed from wild Atlantic salmon due to domestication. ICES Journal of Marine Science 54: 1051–1063.
Fleming I, Hindar K, Mjølnerød IB, Jonsson B, Balstad T, Lamberg A. 2000. Lifetime success and interactions of farm salmon invading a native population. Proceedings of the Royal Society of London B. 267: 1517–1523.
Fleming IA, Agustsson T, Finstad B, Johnsson JI, Björnsson BTh. 2002. Effects of domestication on growth physiology and endocrinology of Atlantic salmon ( Salmo salar ). Canadian Journal of Fisheries and Aquatic Sciences 59:1323-1330.
Forskrift om fellesansvar for utfisking mv. av rømt oppdrettsfisk. 2015. Lovdata: FOR-2015-02-05-89. Hentet fra: https://lovdata.no/dokument/SF/forskrift/2015-02-05-89.
Fraser DJ, Weir LK, Bernatchez L, Hansen MM, Taylor EB. 2011. Extent and scale of local adaptation in salmonid fishes: review and meta-analysis. Heredity 106: 404–420.
Fraser DJ, Houde ALS, Debes PV, O’Reilly P, Eddington JD, Hutchings JA. 2010. Consequences of farmed-wild hybridization across divergent wild populations and multiple traits in salmon. Ecological Applications 20: 935–953.
Gjedrem T. 2010. The first family-based breeding program in aquaculture. Reviews in Aquaculture 2, 2–15.
Gjøen HM, Bentsen HB. 1997. Past, present, and future of genetic improvement in salmon aquaculture. ICES Journal of Marine Science 54: 1009–1014.
Glover KA, Skaala, Ø. 2006. Temporal stability of sea louse Lepeophtheirus salmonis Krøyer populations on Atlantic salmon Salmo salar L. of wild, farm and hybrid parentage. Journal of Fish Biology 68: 1795–1807.
Glover KA, Skar C, Christie KE, Glette J, Rudra H, Skaala Ø. 2006a. Size-dependent susceptibility to infectious salmon anemia virus (ISAV) in Atlantic salmon ( Salmo salar L.) of farm, hybrid and wild parentage. Aquaculture 254: 82–91.
Glover KA, Bergh Ø, Rudra H, Skaala, Ø. 2006b . Juvenile growth and susceptibility to Aeromonas salmonicida subsp. salmonicida in Atlantic salmon ( Salmo salar L.) of farmed, hybrid, and wild parentage. Aquaculture 254: 72–81.
Glover KA, Otterå H, Olsen RE, Slinde E, Taranger GL, Skaala Ø. 2009. A comparison of farmed, wild and hybrid Atlantic salmon ( Salmo salar L.) reared under farming conditions. Aquaculture 286: 203–210.
Glover KA. 2010. Forensic identification of farmed escapees: a review of the Norwegian experience. Aquaculture Environment Interactions 1: 1–10.
Glover KA, Quintela, M, Wennevik V, Besnier F, Sørvik AGE, Skaala Ø. 2012. Three decades of farmed escapees in the wild: A spatio- temporal analysis of Atlantic salmon population genetic structure throughout Norway. PloS One 7: e43129.
Glover KA, Pertoldi C, Besnier F, Wennevik V, Kent M, Skaala Ø. 2013. Atlantic salmon populations invaded by farmed escapees: quantifying genetic introgression with a Bayesian approach and SNPs. BMC Genetics, 14.
Glover KA, Solberg MF , McGinnity P, Hindar K, Verspoor E, Coulson MW, Hansen MM, Araki H, Skaala Ø, Svåsand T. 2017. Half a century of genetic interaction between farmed and wild Atlantic salmon: Status of knowledge and unanswered questions. Fish and Fisheries 1–38.
Glover KA, Urdal K, Næsje T, Skoglund H, Florø-Larsen B, Otterå H, Fiske P, Heino M, Aronsen T, Sægrov H, Diserud O, Barlaup BT, Hindar K, Bakke G, Solberg I, Lo H, Karlsson S, Skaala Ø, Lamberg A, Kanstad-Hanssen Ø, Muladal R, Skilbrei OT & Wennevik V. 2019. Domesticated escapees on the run: the second-generation monitoring program reports the numbers and proportions of farmed Atlantic salmon in >200 rivers annually Ices Journal of Marine Science 76, 1151–61.
Glover KA, Wennevik V, Hindar K. Skaala Ø, Fiske P, Solberg MF, Diserud OH, Svåsand T, Karlsson S, Andersen L.B, Grefsrud ES. 2020. The future looks like the past: Introgression of domesticated Atlantic salmon escapees in a risk assessment framework. Fish and Fisheries 00:1-15, https://doi.org/10.1111/faf.12478 .
Grefsrud ES, Svåsand T, Glover KA, Husa V, Kupka-Hansen P, Samuelsen OB, Sandlund N, Stien LH. 2019. Risikorapport norsk fiskeoppdrett 2019. Fisken og Havet 2019–5.
Hansen LP, Jacobsen JA, Lund RA. 1993. High numbers of farmed Atlantic salmon, Salmo salar L., observed on oceanic waters north of the Faroe Islands. Aquaculture and Fisheries Management 24: 777–781.
Hansen LP. 2006a. Migration and survival of farmed Atlantic salmon ( Salmo salar L.) released from two Norwegian fish farms. ICES J Mar Sci 63, 1211–1217.
Hansen LP. 2006b. Vandring og spredning av rømt oppdrettslaks. NINA Rapport 162: 1–21.
Harvey A, Glover KA, Taylor MI, Creer S, Carvalho GR. 2016a. A common garden design reveals population-specific variability in potential impacts of hybridization between populations of farmed and wild Atlantic salmon, Salmo salar L. Evolutionary Applications 9, 435–49.
Harvey A, Juleff G, Carvalho G, Taylor M, Solberg MF, Dyrhovden L, Matre IH, Glover KA. 2016b. Does density influence relative growth performance of farm, wild and F1 hybrid Atlantic salmon in semi-natural and hatchery common garden conditions? Royal Society Open Science 3:16152.
Harvey A, Solberg M, Troianou E, Carvalho GR, Taylor M, Creer S, Dyrhovden L, Matre IH, Glover KA. 2016c. Plasticity in growth of farmed and wild Atlantic salmon: is the increased growth rate of farmed salmon caused by evolutionary adaptations to the commercial diet? BMC Evolutionary Biology 16:264.
Heino M, Svåsand T, Wennevik W, Glover KA. 2015. Genetic introgression of farmed salmon in native populations: quantifying the relative influence of population size and frequency of escapees. Aquaculture Environment Interactions 6: 185–190.
Hindar K, Ryman N, Utter F. 1991. Genetic effects of cultured fish on natural fish populations. Canadian Journal of Fisheries and Aquatic Sciences 48: 945–57.
Hindar K, Fleming IA, McGinnity P, Diserud O. 2006. Genetic and ecological effects of salmon farming on wild salmon: modelling from experimental results. ICES J. Marine Sci. 63: 1234–1247.
Hindar K, Diserud OH, Fiske P, Forseth T, Jensen A J, Ugedal O, Jonsson N, Sloreid S.-E, Arnekleiv JV, Saltveit SJ, Sægrov H, Sættem LM. 2007. Gytebestandsmål for laksebestander i Norge. NINA Rapport 226. 78 s.
Hindar K, Diserud OH, Fiske P, Karlsson S, Bolstad GH, Foldvik A, Wennevik V, Bremset G og Rosten C. 2018. Evaluering av nasjonale laksevassdrag og nasjonale laksefjorder: Rømt oppdrettslaks, genetisk innkryssing og bestandsstatus. NINA Rapport 1461. Norsk institutt for naturforskning.
Houde ALS, Fraser DJ, Hutchings JA. 2010. Reduced anti-predator responses in multigenerational hybrids of farmed and wild Atlantic salmon ( Salmo salar L.). Conservation Genetics 11: 785–794.
Houston RD, Haley CS, Hamilton A. 2008 Major quantitative trait loci affect resistance to infectious pancreatic necrosis in Atlantic salmon ( Salmo salar ). Genetics 178: 1109–1115.
Hutchings JA. 1991. The threat of extinction to native populations experiencing spawning intrusions by cultured Atlantic salmon. Aquaculture, 98: 119–132.
ICES. 2016. Report of the Workshop to address the NASCO request for advice on possible effects of salmonid aquaculture on wild Atlantic salmon populations in the North Atlantic (WKCULEF), 1–3 March, Copenhagen, Denmark. ICES CM 2016/ACOM 42: 44 pp.
Jacobsen JA, Hansen LP. 2001. Feeding habits of wild and escaped farmed Atlantic salmon, Salmo salar L., in the Northeast Atlantic. Ices Journal of Marine Science 58, 916–33.
Jensen AJ, Karlsson S, Fiske P, Hansen LP, Hindar K, Østborg G. 2013. Escaped farmed Atlantic salmon in the Arctic Ocean. Aquaculture Environment Interactions 3: 223–229.
Johnsson JI, Björnsson BTh. 1994. Growth hormone increases growth rate, appetite and dominance in juvenile rainbow trout, Oncorhynchus mykiss. Animal Behaviour 48: 177–186.
Johnsson JI, Petersson E, Jönsson E, Björnsson BTh, Järvi T. 1996. Domestication and growth hormone alter antipredator behaviour and growth patterns in juvenile brown trout. Salmo trutta . Canadian Journal of Fisheries and Aquatic Sciences 53: 1546–1554
Johnsson JI, Höjesjö J, Fleming IA. 2001. Behavioural and heart rate response to predation risk in wild and domesticated Atlantic salmon. Canadian Journal of Fisheries and Aquatic Sciences 58: 788–794.
Jonsson N, Jonsson B, Hansen LP. 1998. The relative role of density-dependent and densityindependent survival in the life cycle of Atlantic salmon Salmo salar . Journal of Animal Ecology 67: 751–762.
Jonsson B, Jonsson N, Hansen LP. 2003. Atlantic salmon straying from the River Imsa. Journal of Fish Biology 62: 641–657.
Jonsson B, Jonsson N. 2017. Maternal inheritance influences homing and growth of hybrid offspring between wild and farmed Atlantic salmon. Aquaculture Environment Interactions, 9:231-238.
Jönsson E, Johnsson JI, Björnsson BTh. 1996. Growth hormone increases predation exposure of rainbow trout. Proceedings of the Royal Society of London, Series B, 263: 647–651.
Jönsson E, Johnsson JI, Björnsson BTh. 1998. Growth hormone increases aggressive behavior in juvenile rainbow trout. Hormones and Behaviour 33: 9–15.
Karlsson S, Moen T, Hindar K. 2010. Contrasting patterns of gene diversity between microsatellites and mitochondrial SNPs in farm and wild Atlantic salmon. Conservation Genetics 11: 571–582.
Karlsson S, Moen T, Lien S, Glover KA, Hindar K. 2011. Generic genetic differences between farmed and wild Atlantic salmon identified from a 7K SNP-chip. Molecular Ecology Resources 11: 247–253.
Karlsson S, Diserud OH, Moen T, Hindar K. 2014. A standardized method for quantifying unidirectional genetic introgression. Ecology and Evolution 4, 3256–63.
Karlsson S, Diserud O H, Fiske P, Hindar K. 2016. Widespread genetic introgression of escaped farmed Atlantic salmon in wild salmon populations. Ices Journal of Marine Science, 73: 2488–2498.
Keyser F, Wringe BF, Jeffery NW, Dempson JB, Duffy S & Bradbury IR. 2018. Predicting the impacts of escaped farmed Atlantic salmon on wild salmon populations. Canadian Journal of Fisheries and Aquatic Sciences , 1–7.
Koljonen M-L, Tähtinen J, Säisä M, Koskiniemi J. 2002. Maintenance of genetic diversity of Atlantic salmon ( Salmo salar ) by captive breeding programmes and the geographic distribution of microsatellite variation. Aquaculture 212: 69–9.
Liu YJ, Diserud OH, Hindar K, Skonhoft A. 2013. An ecological-economic model on the effects of interactions between escaped farmed and wild salmon (Salmo salar ). Fish and Fisheries 14, 158–73.
Lura H, Sægrov H. 1991. Documentation of successful spawning of escaped farmed female Atlantic salmon, Salmo salar , in Norwegian rivers. Aquaculture 98: 151–159.
Lura H, Økland F. 1994. Content of synthetic astaxanthin in escaped farmed Atlantic salmon, Salmo salar L., ascending Norwegian rivers. Fisheries Management and Ecology, 1: 205–216.
Madhun AS, Karlsbakk E, Isachsen CH, Omdal LM, Sørvik AGE, Skaala Ø, Wennevik V, Glover KA. 2015. Potential disease interaction reinforced: Double-virus infected escaped farmed Atlantic salmon, Salmo salar L., recaptured in a nearby river. Journal of Fish Diseases 38: 209–219.
Martin-Smith KM, Armstrong JD, Johnsson JI, Björnsson BT. 2004. Growth hormone increases growth and dominance of wild juvenile Atlantic salmon with affecting space use. Journal of Fish Biology 65, Suppl. A: 156–172.
McGinnity P, Stone C, Taggart JB, Cooke DD, Cotter D, Hynes, R, McCamley C, CrossT, Ferguson A. 1997. Genetic impact of escaped farmed Atlantic salmon ( Salmo salar L.) on native populations: use of DNA profiling to assess freshwater performance of wild, farmed, and hybrid progeny in a natural river environment. ICES Journal of Marine Science 54: 998–1008.
McGinnity P, Prodöhl P, Ferguson A, Hynes R, Ó Maoiléidigh N, Baker N, Cotter D, O’Hea B, Cooke D, Rogan G, Taggart J, Cross T. 2003. Fitness reduction and potential extinction of wild populations of Atlantic salmon, Salmo salar , as a result of interactions with escaped farm salmon. Proceedings of the Royal Society, London, Series B, 270: 2443–2450.
Mjølnerød IB, Refseth UH, Karlsen E, Balstad T, Jakobsen KS, Hindar K. 1997. Genetic differences between two wild and one farmed population of Atlantic salmon ( Salmo salar ) revealed by three classes of genetic markers. Hereditas 127: 239–248.
Moen T, Baranski M, Sonesson A K, Kjøglum S. 2009. Confirmation and fine-mapping of a major QTL for resistance to infectious pancreatic necrosis in Atlantic salmon ( Salmo salar ): population-level associations between markers and trait. BMC Genomics 10: 368.
Mork J. 1991. One-generation effects of farmed fish immigration on the genetic differentiation of wild Atlantic salmon in Norway. Aquaculture 98: 267–276.
Mork OI, Bjerkeng B, Rye M. 1999. Aggressive interactions in pure and mixed groups of juvenile farmed and hatchery-reared wild Atlantic salmon Salmo salar L. in relation to tank substrate. Aquaculture Research 30: 571–578.
Normandeau E, Hutchings JA, Fraser DJ, Bernatchez L. 2009. Population-specific gene expression responses to hybridization between farm and wild Atlantic salmon. Evolutionary Applications2 , 489–503.
Olsen RE, Skilbrei OT. 2010. Feeding preference of recaptured Atlantic salmon Salmo salar following simulated escape from fish pens during autumn. Aquaculture Environment Interactions 1, 167–74.
Ozerov M, Vähä J-P, Wennevik V, Svenning M-A, Vasemägi A, Diaz Fernandez R, Unneland L, Haapanen K, Niemelä E, Falkegård M, Prusov S, Lyzhov I, Rysakova K, Kalske T, Christiansen B. 2017. Comprehensive microsatellite baseline for genetic stock identification of Atlantic salmon (Salmo salar L.) in northernmost Europe. ICES Journal of Marine Science, 74(8): 2159–2169.
Perry WB, Solberg MF, Besnier F, Dyrhovden L, Matre HH, Fjelldal PG, Ayllon F, Creer S, Llewellyn M, Taylors MI, Carvalhol G & Glover KA. 2019. Evolutionary drivers of kype size in Atlantic salmon ( Salmo salar ): domestication, age and genetics. Royal Society Open Science6 , 14.
Reed TE, Prodohl P, Hynes R, Cross T, Ferguson A, McGinnity P. 2015. Quantifying heritable variation in fitness-related traits of wild, farmed and hybrid Atlantic salmon families in a wild river environment. Heredity 115, 173–84.
Rengmark AH, Slettan A, Skaala O, Lie O, Lingaas F. 2006. Genetic variability in wild and farmed Atlantic salmon ( Salmo salar ) strains estimated by SNP and microsatellites. Aquaculture, 253: 229–237.
Roberge C, Einum S, Guderley H, Bernatchez L. 2006. Rapid parallell evolutionary changes of gene transcription profiles in farmed Atlantic salmon. Molecular Ecology 15: 9–20.
Roberge C, Normandeau E, Einum S, Guderley H, Bernatchez, L. 2008. Genetic consequences of interbreeding between farmed and wild Atlantic salmon: insights from the transcriptome. Molecular Ecology 17: 314–324.
Robertsen G, Reid D, Einum S, Aronsen T, Fleming I, Sundt-Hansen L, Karlsson S, Kvingedal E, Ugedal O, Hindar K. 2019. Can variation in standard metabolic rate explain contextdependent performance of farmed salmon offspring? Ecology and Evolution9 , 212–22.
Skaala Ø, Høyheim B, Glover KA, Dahle G. 2004. Microsatellite analysis in domesticated and wild Atlantic salmon ( Salmo salar L.): allelic diversity and identification of individuals. Aquaculture 240: 131–143.
Skaala Ø, Taggart JB, Gunnes K. 2005. Genetic differences between five major domesticated strains of Atlantic salmon and wild salmon Journal of Fish Biology 67: 118–128.
Skaala Ø, Wennevik V, Glover KA. 2006. Evidence of temporal genetic change in wild Atlantic salmon ( Salmo salar L) populations affected by farmed escapees. ICES J. Marine Science 63: 1224–1233.
Skaala Ø, Glover KA, Barlaup BT, Svåsand T, Besnier F, Hansen MM, Borgstrøm, R. 2012. Performance of farmed, hybrid and wild Atlantic salmon families in a natural river environment. Canadian Journal of Fisheries and Aquatic Sciences 69: 1994–2006.
Skaala Ø, Glover KA, Barlaup B, Borgstrøm R. 2014. Microsatellite DNA used for parentage identification of partly digested Atlantic salmon ( Salmo salar ) juveniles in a natural rivner environment. Marine Biology Research 10: 323–328.
Skaala Ø, Besnier F., Borgstrom R., Barlaup B., Sorvik A.G., Normann E., Ostebo B.I., Hansen M.M. & Glover K.A. 2019. An extensive common-garden study with domesticated and wild Atlantic salmon in the wild reveals impact on smolt production and shifts in fitness traits. Evolutionary Applications 12 , 1001–16.
Skilbrei OT. 2013. Migratory behaviour and ocean survival of escaped out-of-season smolts of farmed Atlantic salmon Salmo salar. Aquaculture Environment Interactions 3:213-221.
Skilbrei OT, Skulstad OF, Hansen T. 2014. The production regime influences the migratory behavior of escaped farmed Atlantic salmon ( Salmo salar L.). Aquaculture 424–425, 146–150.
Skilbrei OT, Heino M, Svåsand T. 2015a. Using simulated escape events to assess the annual numbers and destinies of escaped farmed Atlantic salmon of different life stages, from farms sites in Norway . ICES Journal of Marine Science, 72 : 670–685.
Skilbrei OT, Normann E, Meier S, Olsen RE. 2015b. Use of fatty acid profiles to monitor the escape history of farmed Atlantic salmon. Aquaculture Environment Interactions 7:1-13.
Solberg MF, Kvamme O, Nilsen F, Glover KA. 2012. Effects of environmental stress on mRNA expression levels of seven genes related to oxidative stress and growth in Atlantic salmon Salmo salar L. of farmed, hybrid and wild origin. BMC Research Notes 5:672.
Solberg MF, Skaala Ø, Nilsen F, Glover KA. 2013a. Does domestication cause changes in growth reaction norms? A study of farmed, wild and hybrid Atlantic salmon families exposed to environmental stress. PLoS ONE 8(1): e54469.
Solberg MF, Zhang Z, Nilsen F, Glover KA. 2013b. Growth reaction norms of domesticated, wild and hybrid Atlantic salmon families in response to differing social and physical environments. BMC Evolutionary Biology 13:234.
Solberg MF, Zhang Z, Glover KA. 2015. Are farmed salmon more prone to risk than wild salmon? Susceptibility of juvenile farm, hybid and wild Atlantic salmon Salmo salar L. to an artificial predator. Applied Animal Behaviour Science 162: 67–80.
Solberg MF, Dyrhovden L, Matre IH, Glover KA. 2016. Thermal plasticity in farmed, wild and hybrid Atlantic salmon: no indication of domestication-driven divergence in lower thermal tolerance during early development. BMC Evolutionary Biology 16:38.
Sylvester EVA, Wringe BF, Duffy SJ, Hamilton LC, Fleming IA, Bradbury IR. 2018. Migration effort and wild population size influence the prevalence of hybridization between escaped farmed and wild Atlantic salmon. Aquaculture Environment Interactions 10, 401–11.
Sylvester EVA, Wringe BF, Duffy SJ, Hamilton LC, Fleming IA, Castellani M, Bentzen P, Bradbury IR. 2019. Estimating the relative fitness of escaped farmed salmon offspring in the wild and modelling the consequences of invasion for wild populations. Evolutionary Applications 12, 705–17.
Sægrov H, Hindar K, Kålås S, Lura H. 1997. Escaped farmed Atlantic salmon replace the original salmon stock in the River Vosso, western Norway. ICES Journal of Marine Science 54: 1166–1172.
Taranger GL, Svåsand T, Bjørn PA, Jansen PA, Heuch PA, Grøntvedt RN, Asplin L. 2012. Forslag til førstegenerasjons målemetode for miljøeffekt (effektindikatorer) med hensyn til genetisk påvirkning fra oppdrettslaks til villaks, og påvirkning av lakselus fra oppdrett på viltlevende laksefisk. Fisken og havet, 13-2010, Havforskningsinstituttet; Veterinærinstituttets rapportserie Nr. 7–2012.
Taranger GL, Karlsen O, Bannister RJ, Glover KA, Husa V, Karlsbakk E, Kvamme BO, Boxaspen KK, Bjørn PA, Finstad B, Madhun AS, Morton HC, Svåsand T. 2015. Risk assessment of the environmental impact of Norwegian Atlantic salmon farming. Ices Journal of Marine Science 72, 997–1021.
Thodesen J, Grisdale-Helland B, Helland SJ, Gjerde B. 1999. Feed intake, growth and feed utilization of offspring from wild and selected Atlantic salmon ( Salmo salar ). Aquaculture 180:237-246.
Verspoor E. 1988. Reduced genetic variability in first-generation hatchery populations of Atlantic salmon ( Salmo salar ). In: Verspoor, E., Stradmeyer, L. & Nielsen J.L. (Eds.) The Atlantic salmon: genetics, conservation and management. Blackwell Publishing, Oxford. Canadian Journal of Fisheries and Aquatic Sciences 45: 1686–1690.
Verspoor E. 1997. Genetic diversity among Atlantic salmon ( Salmo salar L.) populations. ICES Journal of Marine Science 54: 965–973.
Volpe JP, Taylor EB, Rimmer DW, Glickman BW. 2000. Evidence of natural reproduction of aquaculture escaped Atlantic salmon ( Salmo salar ) in a coastal British Columbia river. Conservation Biology 14: 899–903.
Webb JH, McLaren IS, Donaghy MJ, Youngson AF. 1993. Spawning of farmed Atlantic salmon, Salmo salar L., in the second year after their escape. Aquaculture and Fisheries Management 24: 557–561.
Webb JH, Verspoor E, Aubin-Horth N, Romakkaniemi A, Amiro P. 2007. The Atlantic Salmon. Chapter 2. In: The Atlantic salmon: genetics, conservation and management. Verspoor E., Stradmeyer, L.& Nielsen J.L.(Eds). Blackwell Publishing, Oxford, pp. 17–56.
Wennevik V, Quintela M, Skaala Ø, Verspoor E, Prusov S, Glover KA. 2019. Population genetic analysis reveals a geographically limited transition zone between two genetically distinct Atlantic salmon lineages in Norway. Ecology and Evolution
Wringe BF, Jeffery NW, Stanley RRE, Hamilton LC, Anderson EC, Fleming IA, Grant C, Dempson JB, Veinott G, Duffy SJ, Bradbury IR. 2018. Extensive hybridization following a large escape of domesticated Atlantic salmon in the Northwest Atlantic. Communications Biology 1.
Zhang Z, Glover KA, Wennevik V, Svåsand T, Sørvik AGE, Fiske P, Karlsson S, Skaala Ø. 2013. Genetic analysis of Atlantic salmon captured in a netting station reveals multiple escapement events from commercial fish farms. Fish Manage Ecol 20:42-51.
3 - Miljøeffekter som følge av utslipp av løste næringssalter fra fiskeoppdrett
Norsk produksjon av laks og regnbueørret var i 2018 om lag 1 340 000 tonn (uttak til slakt) og innrapportert fôrforbruk var i 2018 på om lag 1 600 000 tonn (kilde: Fiskeridirektoratet). Oppløst nitrogen og fosfor slippes ut fra matfiskanlegg som uorganiske forbindelser dannet ved fiskens metabolisme. Utslippsmengden av løste næringssalter vil være proporsjonal med fiskeproduksjonen. Det er stor usikkerhet rundt mengden av utslipp av løste næringssalter fra matfiskanlegg. Det finnes ulike modeller og massebudsjett for beregning av løste næringssalter (Wang mfl. 2012; Norderhaug mfl. 2016a; Svåsand mfl. 2016; Torrissen mfl. 2016), men ingen av modellene eller beregningsmetodene er godt nok verifiserte. Modellberegninger av utslippene av løst uorganisk nitrogen (DIN) varierer fra 20 kg til 38,4 kilo per tonn produsert fisk, mens utslippene av løst uorganisk fosfor (DIP) varierer fra -1,5 (negativt fosforbudsjett) til 5,1 kilo per tonn produsert fisk (tabell 3.1). Vi har i risikovurderingen derfor valgt å bruke TEOTIL modellen som estimerer de høyeste utslippene av løste næringssalter ( 38,4 kg løst nitrogen og 5,1 kg løst fosfor per tonn laks produsert) , for å ikke underestimere utslippene. Dette vil gi estimerte utslipp på 51 538 tonn løst nitrogen og 6845 tonn løst fosfor i 2018. Til sammenligning slippes det årlig ut om lag 48 000 tonn nitrogen og 2600 tonn fosfor til vann fra jordbruk, avløp og landbasert industri (SSB). Figur 3.1 viser utslippsmengde av løst nitrogen og fosfor fra matfiskproduksjon i produksjonsområdene langs kysten.
DIN-kg/tonn fisk
DIP kg/tonn fisk
Massebalansebudsjett I (Wang mfl. 2012)
36
2,45
Massebalansebudsjett II (Torrissen mfl. 2016)
23
- 1,5
ANCYLUS (Svåsand mfl. 2016)
20
3
TEOTIL (Norderhaug 2016 mfl. 2016a)
38,4
5,1
Tabell 3.1 Utslipp av løst uorganisk nitrogen (DIN) og løst uorganisk fosfor (DIP) (kg) per tonn produsert laks og regnbueørret beregnet med to ulike massebalansebudsjett og to modeller.
Figur 3.1 Årlig (2018) utslippsmengde av løst nitrogen og fosfor fra fiskeoppdrett i produksjonsområdene langs norskekysten.
3.2 - Spredning og fortynning
Når fisk produseres i åpne merdanlegg slippes det ut næringssalter direkte til miljøet. Disse omfatter nitrogen (nitrat, nitritt og ammonium) og fosfor (fosfat) i form av løste uorganiske forbindelser som dannes under fiskens metabolisme og skilles ut via gjeller og nyrer. De løste forbindelsene som slippes ut fra anlegget vil spre seg med overflatestrømmen hovedsakelig i merddyp (0-30 meter). De fortynnes relativt raskt i vannmassene og gjør at det kan være vanskelig å måle forhøyde konsentrasjoner av ammonium ved anlegget, men under fôring av fisken representerer de stadige pulser av lett omsettelige nitrogenforbindelser. En rekke studier har vært gjort av løste næringssalter fra oppdrettsanlegg, og de fleste konkluderer med relativt svake pulser i inntil 1-2 km fra merdene (sammenstilt i Price mfl. 2015). Hvor langt disse pulsene med forhøyde næringssaltverdier strekker seg, vil variere med lokale forhold (vannutskiftning, strømforhold o.a.) og biomassen av fisk i anleggene. Utslippsmengde fra fiskeproduksjonen vil også variere med årstiden siden fisken vokser mest om sommeren, noe som gir høyest utslipp i denne perioden. Figur 3.2 viser et eksempel på fordelingen av ammonium (nitrogen) ved et stort lakseanlegg (5000 tonn) på åpen kyst nær Florø i Sogn og Fjordane ved lav og høy produksjon i anlegget (Jansen mfl. 2018).
Figur 3.2 Ammoniumkonsentrasjoner (dybdeintegrerte middelverdier) målt ved et anlegg på åpen bølgeeksponert kyst ved Florø under lav (april) og høy (september) produksjon av fisk. Figuren viser konsentrasjoner i overflatevannet (0–20 meter) sett ovenfra. Anlegget er markert med svart firkantet boks og prøvetakingspunktene er markert med kryss (verdier i µmol/l, merk ulik fargeskala på figurene) (figur fra Jansen mfl. 2018).
Utslipp fra akvakultur står for det største menneskeskapte bidraget av næringssalter til norsk kystvann på strekningen Rogaland–Finnmark. Utslipp av ekstra næringssalt i områder med dårlig vannutskiftning kan føre til at næringssaltkonsentrasjonen i sjøvannet øker lokalt. Tidligere lå en rekke anlegg delvis innelukkede områder, noe som førte til lokale overgjødslingsprosesser. De fleste norske oppdrettsanlegg ligger i dag lokalisert i områder med god overflatestrøm. Dette er nødvendig for at fisken i anleggene skal trives og få nok oksygen. Samtidig sørger overflatestrømmen for utskifting av overflatevannet og er med på å spre og fortynne de løste næringssaltene.
3.3 - Effekter av utslipp av løste næringssalt
3.3.1 - Endring i planteplanktonproduksjonen (eutrofi)
Effekten av næringssaltutslippene på planteplanktonproduksjonen vil avhenge av sjøareal, oppholdstid og grad av innblanding av andre vannmasser (vannsirkulasjon). Langs norskekysten er uorganisk fosfor sjelden en begrensende faktor for planteplankton, og en ytterligere tilførsel av fosfor vil derfor ikke gi en direkte respons i planteplanktonproduksjonen. Betydelige utslipp av uorganisk nitrogen, som det normalt blir for lite av i sommerhalvåret, kan derimot føre til økt planteplanktonproduksjon, økt nedbrytning av algebiomasse i dypet og oksygenmangel i bunnvannet. Denne tilstanden kaller vi eutrofi.
Overgjødsling (eutrofiering) av de kystvann defineres av OSPAR som ekstra tilførsler av næringssalter som forårsaker en økt vekst av alger eller vannplanter og fører til uønsket forstyrrelse av balansen mellom organismer i vannet og av kvaliteten på vannet de lever i (Anon 2017). Regionale effekter av utslipp av næringssalter er kjent fra flere deler av verden der menneskeskapte utslipp fra jordbruk og kloakk har skapt store problemer.
En rekke studier har undersøkt planteplanktonforekomstene nært oppdrettsanlegg, men har ikke kunnet påvise forhøyede verdier ved anleggene (Taylor mfl. 1992; Pitta mfl. 1999, 2006; Price mfl. 2015). Årsaken til dette er mest sannsynlig at planteplanktonets oppholdstid i området med forhøyede verdier er for kort til at planktonalgene kan respondere med økt produksjon. Noen studier har vist forhøyet planteplanktonproduksjon i oppdrettsområder, men dette har særlig vært i Middelhavet og i Asia (sammenstilt i Price mfl. 2015).
Målinger fra områder med høy tetthet av oppdrettsanlegg i Chile, Skottland, Middelhavet (Gowen & Ezzi 1994; Soto & Norambuena 2004; Pitta mfl. 2006) har ikke vist verdier som tyder på regional overgjødsling av frie vannmasser i områder med god vannutskiftning. Planktonmengde og artssammensetning overvåkes ukentlig langs norskekysten i regi av Mattilsynet gjennom overvåkningsprogrammet for skadelige alger. Det er stor naturlig variasjon i planteplanktonbiomassen og artssammensetningen i løpet av året og mellom årene, og det registreres også betydelige ulikheter innenfor små geografiske områder. En tre-års studie av regionale effekter i oppdrettsintensive Hardangerfjorden viste heller ingen økte næringssalt- eller klorofyllverdier i de åpne vannmassene (Husa mfl. 2014a).
3.3.2 - Teoretisk beregning av respons i planteplanktonproduksjon
Effekten av utslippene på planteplanktonproduksjonen vil avhenge av sjøareal, oppholdstid og grad av innblanding av andre vannmasser (vannsirkulasjon). Det er mulig å gjøre enkle beregninger av responsen i planteplanktonproduksjonen. Midlere planteplanktonproduksjon i norske kyst og fjordområder er ca. 135 gram C/m2 /år (Wassmann 1990 a, b). Hvis en antar at 100 % av det løste nitrogenet som slippes ut fra matfiskanlegg omsettes til planktonproduksjon, ser vi av figur 3.3 at det vil være størst relativ økning i de naturlige nivåene av planteplanktonbiomasse i produksjonsområde 3 Karmøy til Sotra (18,4 %) og minst i produksjonsområde 12 Øst-Finnmark (1,2 %). Med en teoretisk økning på 18,4 % i planteplanktonproduksjonen, slik som i produksjonsområde 3, vil klorofyll-a-verdiene fremdeles ligge innenfor grensen for god vannkvalitet (Svåsand mfl. 2016). Som mål på overgjødsling benyttes her at konsentrasjonen av planteplankton ikke skal være mer enn 50 % over naturlig referansetilstand. Utslippene er beregnet på grunnlag av data fra Fiskeridirektoratets biomassestatistikk for 2018 fordelt på produksjonsområder. Sjøarealene i de ulike produksjonsområdene vil variere, derfor er utslippsmengden vurdert per kystnært sjøareal. Kystnært sjøareal er her definert som arealet innenfor 2 km fra kystlinjen. Dette er gjort fordi samme utslippsmengde vil kunne ha større effekt i et lite sjøareal enn den vil ha i et stort. Beregningene er utført på det totale sjøarealet i hvert produksjonsområde og tar ikke hensyn til lokale forskjeller som områder med mindre vannutskifting og områder med spesielt høy fiskeproduksjon, som kan gi betydelig høyere prosentvis økning enn verdiene vist i figur 3.3.
Figur 3.3 Estimert prosentvis økning i planteplanktonproduksjonen som følge av utslipp av løst nitrogen fra matfiskanlegg i 2018 fordelt på produksjonsområder (basert på 100 prosent utnyttelse av nitrogen til karbonfiksering).
3.3.3 - Endringer i makroalgesamfunn på hardbunn
Makroalgesamfunn kan respondere raskt på ekstra nitrogentilførsel med redusert biodiversitet og økt forekomst av opportunistiske grønnalger på bekostning av flerårige habitatbyggende arter som tang og tare (Klavestad 1967, 1978; Bokn & Lein 1978; Bokn mfl. 1992; Munda 1996; Bartsch & Kuhlenkamp 2000, 2009). Dette fenomenet har vi sett i Oslofjorden i perioden med høye nitrogenverdier på grunn av utslipp fra kloakk, industri og landbruk. Økt forekomst av planteplankton kan redusere tilgangen på lys for makroalger, og kan dermed redusere voksedypet betydelig (Rueness & Fredriksen 1991). Massive oppblomstringer av opportunistiske makroalger i overgjødslede områder er kjent fra flere deler av verden (Liu mfl. 2010; Pang mfl. 2010; Ménesguen 2010). En undersøkelse av makroalger på hardbunn i Hardangerfjorden, der man har hatt lengre tid med høy oppdrettsintensitet, viste ingen overgjødslingseffekter på makroalgesamfunn i fjorden (Husa mfl. 2014b).
3.4 - Overvåkning av miljøtilstand i kystvann
Næringssaltkonsentrasjoner, klorofyll-a verdier (planteplankton) og makroalgesamfunn på hardbunn inngår i overvåkning av miljøkvalitet i kystvann. Som ett ledd i implementeringen av vannforskriften i Norge ble det i 2013 startet opp en trendovervåkning av miljøtilstanden i norske kystområder (ØKOKYST). Denne overvåkningen dekker foreløpig bare noen utvalgte stasjoner i fylkene og er for det meste plassert i upåvirkede områder. Det er planlagt et utvidet nettverk med stasjoner for denne overvåkningen slik at man etter hvert får mer data fra norskekysten.
I Rogaland, Hordaland og i seks fjorder i Nordland startet det i 2012-2013 opp overvåkning av vannkvalitet på ett relativt dekkende stasjonsnett i områder med matfiskproduksjon. Overvåkningsprogrammet følger veiledere gitt etter Vannforskriften (Direktoratsgruppen vanndirektivet, Veileder 02:2018), finansieres av lokale oppdrettere, koordineres av Blue Planet og utføres av akkrediterte, uavhengige firma. Data fra disse tre fylkene gir foreløpig det beste grunnlaget for å si noe om miljøtilstanden i områder med matfiskproduksjon.
3.4. Lokale effekter av utslipp av løste næringssalter
Lokale effekter av utslipp av næringssalt på makroalger på hardbunn eller spesielle grunne naturtyper er ikke vurdert i denne omgang, men dette er problemstillinger som det arbeides med. Slike lokale effekter i grunne områder inngår i dag ikke i noen systematisk overvåkning ved matfiskanlegg.
I matfiskanleggenes påvirkningssone kan det forekomme effekter fra utslipp av næringssalter i strandsonen eller der anlegget er plassert i grunne skjellsandsområder og på tarebanker, slik moderne kystanlegg gjerne ligger i dag. Kontinuerlige pulser av næringssalter kan forårsake lokale overgjødslingseffekter. Effekten av utslippene i grunne områder vil i høy grad bestemmes av faktorer som strømhastighet og bølgepåvirkning. Studier av makroalgesamfunn i overgjødslede områder viser at en etter hvert utvikler et samfunn med redusert biodiversitet og en overvekt av grønnalger i artssamfunnet (Munda 1996). Ammonium som slippes ut fra matfiskanlegg tas lett opp i alger. Studier har vist at økte ammoniumstilførsler stimulerer til økt vekst av hurtigvoksende makroalger med høy volum-/overflateratio, slik som tynne, bladaktige og trådformede arter. Dette kan føre til økte mengder av påvekstalger som kan redusere lystilgangen og konkurrere effektivt om næringssaltene slik at man over tid kan få en reduksjon av flerårige, seintvoksende arter som tang og tare (Worm & Sommer 2000). En studie av makroalgesamfunn ved fiskeoppdrettsanlegg i Tasmania viste høyere biomasse av opportunistiske alger på korallrev i nærheten av fiskeoppdrettsanlegg (100-400 m), men fant ingen reduksjon i de habitatbyggende artene (Oh mfl. 2015). Fiskeoppdrettsanleggene i Tasmania er plassert på svært grunne og innstengte lokaliteter og er ikke direkte sammenlignbare med norske forhold.
Stortareskog er en viktig naturtype på bølgeeksponert kyst og fungerer som levested og oppvekstområde for en rekke andre arter. Økende forekomst av store matfiskanlegg på eksponert kyst kan potensielt påvirke denne naturtypen. Studier av effekten av utslipp av løste næringssalter i stortareskog viser liten respons i assosiert fauna og flora. Tareskogssamfunnene både på blader, stilkene og i hapterene (festerøtter) viser kun små forskjeller mellom anlegg og referansestasjoner og viser at dette samfunnet er relativt robust mot utslipp fra oppdrett (Haugland 2019).
Løsliggende kalkalger (ruglbunn): Utslipp av næringssalter kan føre til at kalkalgene blir overgrodde av ettårige, hurtigvoksende algearter, noe som kan nedsette vekstraten til kalkalgene. Undersøkelser fra Skottland, Spania og Middelhavet har vist at kalkalger generelt er sensitive for økt sedimentering av organiske partikler. Dersom planten blir dekket av et lag med finkornet sediment, vil gassutvekslingen i cellene forhindres, og plantens mulighet til å drive fotosyntese vil svekkes. Sediment som inneholder hydrogensulfid (H2S), som kan dannes i nærheten av oppdrettsanlegg, kan være fatalt for kalkalger, og 14 dagers dekke av slikt sediment førte til 100 % dødelighet (Wilson mfl. 2004). Utslipp av organisk materiale påvirker også dyr som har kalkalgeforekomster som levested. Typisk vil de fleste krepsdyr forsvinne og bli erstattet med opportunistiske børstemark (Sanz-Lazaro mfl. 2011; Hall-Spencer 2006). Det pågår arbeid ved Havforskningsinstituttet for å undersøke effekter av utslipp på denne naturtypen.
Ålegressenger: Hvordan utslipp av næringssalter fra matfiskanlegg påvirker ålegressenger har ikke blitt studert i Norge (Husa mfl. 2016). En rekke studier fra andre områder har imidlertid vist at ålegressenger kan påvirkes ved at det etableres større mengder påvekstalger på ålegresset som skygger for lyset. Dette fører til redusert vekst hos ålegresset og vil på sikt kunne redusere forekomsten av ålegress (Munda 1996; Worm & Sommer 2000; Diaz-Almela mfl. 2008; Duarte mfl. 2008).
3.5 - Referanser
Anon 2017. Eutrophication Status of the OSPAR Maritime Area Third Integrated Report on the Eutrophication Status of the OSPAR Maritime Area. OSPAR Commission, Eutrophication Series 2017.
Bartsch I, Kuhlenkamp R. (2000). The marine macroalgae of Helgoland (North Sea): An annotated list of records between 1845 and 1999. Helgoland Marine Research 54, 160-189.
Bartsch I, Kuhlenkamp R. (2009). Entwicklung der Makrophyten. Vegetation bei Helgoland vor dem Hintergrund der Wasserrahmenrichtlinie. Bundesamt für Seeschiffahrt und Hydrographie (BSH), Hamburg. Meeresumwelt Aktuell: Nord- und Ostsee 1, 1-8.
Bokn T, Lein T.E. (1978). Long-term changes in fucoid assiciation of the inner Oslofjord, Norway. Norwegian Journal of Botany 25, 9-14.
Bokn T, Murray SN, Moy FE, Magnusson JB. (1992). Changes in fucoid distribution and abundances in the inner Oslofjord, Norway: 1974–80 versus 1988–90. Acta Phytogeographica Suecia 78, 117-124.
Diaz-Almela E, Marba N, Alvarez E, Santiago R, Holmer M, Grau A, Mirto S, Danovaro R, Petrou A, Argyro M, Karakassis I, Duarte CM. (2008). Benthic input rates predict seagrass
Duarte CM, Frederiksen M, Grau A, Karakassis L, Marba N, Mirto S, Pérez P, Pusceddu A, Tsapakis M. (2008). Effects of fish farm waste on Posidonia oceanic meadows; Synthesis and provision of monitoring and management tools. Marine Pollution Bulletin 56, 1618-1629.
Gowen RJ, Ezzi IA. (1994). Assessment and prediction of the potential for hypernutrification and eutrophication assosiated with cageculture of salmonids in Scottish waters. Dunstaffnage Marine Laboratory, Oban Scotland, 137 s.
Hall-Spencer J, White N, Gillespie E, Katie G, Foggo A. 2006. Impact of fish farms on maerl beds in strongly tidal areas. Marine Ecology Progress Series 326, 1-9.
Haugland BT. 2019 Effects of fish farm effluents on kelp forest ecosystems: Kelp performance, associated species, and habitats. PhD thesis, Universitetet i Bergen.
Husa V, Kutti T, Grefsrud ES, Agnalt AL, Karlsen Ø, Bannister R, Samuelsen O, Grøsvik BE. (2016). Effekter av utslipp fra akvakultur på spesielle marine naturtyper, rødlista habitat og arter. Rapport fra Havforskningen Nr. 8-2016. 52 s.
Husa V, Kutti T, Ervik A, Sjøtun K, Hansen PK, Aure J. 2014a. Regional impact from finfish farming in an intensive production area (Hardangerfjorden, Norway). Marine Biology Research. 3, 241-252.
Husa V, Steen H, Sjøtun K. 2014b. Historical changes in the macroalgal communities in Hardangerfjorden. Marine Biology Research. 3, 226-240.
Jansen HM, Broch OJ, Bannister R, Cranford P, Handå A, V Husa, Zeng Jie Jiang, Strohmeier T, Strand H. (2018). Spatio-temporal dynamics in the dissolved nutrient waste plume from Norwegian salmon cage aquaculture. Aquaculture Environment Interactions 10:385-399.
Klavestad N. (1967). Undersøkelser over benthos-algevegetasjonen i indre Oslofjord i 1962-1965. Delrapport 9. NIVA. 119 s.
Klavestad N. (1978). The marine algae of the polluted inner part of the Oslofjord. Botanica Marina 21, 71-97.
Liu D, Keesing JK, Dong Z, Zhen Y, Di B, Shi Y, Fearns P, Shi P. (2010). Recurrence of the world’s largest green-tide in 2009 in Yellow Sea, China: Porphyra yezoensis aquaculture rafts confirmed as nursery for macroalgal blooms. Marine Pollution Bulletin 60, 1423-32.
Norderhaug KM, Gundersen H, Høgåsen T, Johnsen TM, Severinsen G, Vedal J, Sørensen K, Walday M. (2016a). Eutrophication status for Norwegian waters. National report for the third application of OSPARs Common Procedure. Rapport frå Miljødirektoratet M- 589.
Ménesguen A, Perrot T, Dussauze M. (2010). Ulva Mass Accumulations on Brittany Beaches: Explanation and Remedies Deduced from Models. Mercator Ocean Quarterly Newsletter, October 2010.
Munda IM. (1996). The northern Adriatic Sea. In Ecological studies Vol 123. Eds. Scramm & Nienhaus. Marine benthic vegetation. Kap 16, 369-402
Oh ES, Edgar GJ, Kirkpatrick JB, Stuart-Smith RD, Barett NS. 2015. Broad -scale impacts of salmon farms on temperate macroalgal assemblages on rocky reefs. Marine Pollution Bulletin 98: 201-209.
Pang SJ, Liu F, ShanTF, Xu N, Zhang ZH, Gao SQ, Chopin T, Sun S. (2010). Tracking the algal origin of the Ulva bloom in the Yellow Sea by a combination of molecular, morphological and physiological analyses. Marine Environmental Research 69, 207-215.
Pitta P, Karakassis I, Tsapakis M, Zivanovic S. (1999). Natural vs. Mariculture derived nutrients and plankton in the Mediterranean Sea. Hydrobiologia 391, 181-194.
Pitta P, Apostolaki ET, Tsagaraki T, Tsapakis M, Karakassis I. (2006). Fish farming effects on the chemical and microbiological variables of the water column: a spatio-temporal study along the Mediterranean Sea. Limn. Hydrobiologia 563, 99-108.
Price C, Black KD, Hargrave BT, Morris JA. (2015). Marine cage culture and the environment: effects on water quality and primary production. Aquaculture Environmental Interactions 6,151-174.
Rueness J. og Fredriksen S. (1991). An assessment of possible pollution effects on the benthic algae of the outer Oslofjord, Norway. Oebalia 17, 223-235.
Sanz-Lazaro C, Belando MD, Marin-Guirao L, Navarrete-Mier F, Marin A. (2011). Relationship between sedimentation rates and benthic impact on Maerl beds derived from fish farming in the Mediterranean. Marine Environmental Research 71, 22-30.
Soto D, Norambuena F. 2004. Evaluation of salmon farming effects on marine systems in the inner seas of southern Chile: a large-scale mensurative experiment. Journal of Applied Ichthyology 20,493-501.
Svåsand T, Karlsen Ø, Kvamme BO, Stien LH, Taranger GL, Boxaspen K. (2016). Risikovurdering av norsk fiskeoppdrett 2016. Fisken og havet, særnummer 2-2016. 192.
Taylor BE, Jamieson G, Carefoot TH. (1992). Mussel culture in British Columbia: the influence of salmon farms on growth of Mytilus edulis . Aquaculture 108, 51-66.
Torrissen O, Hansen PK, Aure J, Husa V, Andersen S, Strohmeier T, Olsen RE. (2016). Næringsutslipp fra havbruk- nasjonale og regionale perspektiv. Rapport fra Havforskningen Nr. 12-2016. 19 s.
Wassmann P. 1990a. Relationship between primary and export production in the boreal coastal zone of the North Atlantic. Limnology and Oceanography 35: 464-471
Wassmann P. 1990b. Calculating the load of organic carbon to the aphotic zone in eutrophicated coastal waters. Marine Pollution Bulletin 21: 183-187.
Wang X, Olsen LM, Reitan KI, Olsen Y. (2012). Discharge of nutrient wastes from salmon farms: environmental effects, and potential for integrated multi-trophic aquaculture. Aquaculture Environment Interactions 2, 267-283.
Wilson S, Blake C, Berges JA, Mags CA. (2004). Environmental tolerances of free-living coralline algae (maerl): implications for European marine conservation. Biological Conservation 120, 283-293.
Worm B, Sommer U. (2000). Rapid direct and indirect effects of a single nutrient pulse in a seaweed-epiphyte grazer system. Marine Ecology Progress Series 2002, 283-288.
4 - Miljøpåvirkning på bunn som følge av partikulære organiske utslipp fra fiskeoppdrett
Forfatter(e):
Pia Kupka Hansen
(HI)
Utforing i merd. Foto: Eivind Senneset/Havforskningsinstituttet
4.1 - Innledning
Oppdrettsfisk i Norge produseres i all hovedsak i åpne merdanlegg og det slippes ut organiske partikler direkte til miljøet i form av fekalier fra fisken og fôr som ikke spises. Partiklene spres over større områder eller akkumuleres på eller i sedimentet og vil kunne påvirke bunnmiljøet i større eller mindre grad rundt oppdrettsanlegget. Mengden organiske partikler som slippes ut fra fiskeoppdrettsanlegg vil være proporsjonalt med produksjonen av fisk, men påvirkningen vil avhenge av de naturlige forholdene på lokaliteten og i området, især strømforholdene.
Fôrpellets og fekalier har ulike fysiske egenskaper. Pelletene er relativt faste og går ikke lett i stykker, men fekalier er ofte skjøre og brekkes lettere opp i mindre deler som synker med ulik hastighet. Synkehastigheten for pellets i størrelsen 4–10 mm ligger i området 6–11 cm/s (Yrong-Song mfl. 1999). Fordeling av synkehastigheten av fekalier fra 1,5 kg stor fisk er vist i figur 4.1 A. Intakte fekalier synker med en fart på 5 til 10 cm/s, og i overkant av 90 % av partiklene synker raskere enn 2,5 cm/s. En liten fraksjon synker langsommere enn 0,1 cm/s, og spres langt videre enn det umiddelbare influensområdet. Fiskestørrelse synes å ha liten betydning for synkehastigheten.
Figur 4.1 A) Fordeling av synkehastighet til fekalier fra 1,5 kg stor laks. B) Spredning av organisk materiale i nær- og fjernsone ved et matfiskanlegg i en fjord på Vestlandet. Simuleringer ble kjørt for 1,5 kg tung fisk over en 14 dagers periode, og er presentert som normalisert partikkeldistribusjon basert på antall partikler som ble sluppet gjennom simuleringsperioden (begge figurer fra Bannister mfl. 2016).
Spredningen av partiklene bestemmes av dyp, vannstrøm, hvor raskt de synker og hvor lett de går i oppløsning. De relativt høye synkehastighetene til spillfôr og intakte fekalier gjør at lokaliteter med lave strømhastigheter (< 5 cm/s) vil få deponert det meste av det organiske materialet under og i den umiddelbare nærhet til anlegget. Lokaliteter med høye strømhastigheter (> 10 cm/s) sprer partiklene over et større område med relativt lite bunnfelling rett under merdene. Ved å kombinere synkehastighetsdata med 3-D hydrodynamiske modeller kan se på spredningen av partikulært organisk materiale (figur 4.1B) (Bannister mfl. 2016).
Strømforholdene er ulike inne i fjordene og ute på kysten. Fjordlokaliteter kan ha god strøm i merddypet, mens det ofte er lite vannbevegelse i dypere vannlag. Dette er i motsetning til anlegg som ligger ute på kysten, som ofte ligger grunnere og har god strøm i hele vannsøylen. Fjordlokaliteter er derfor mer utsatt for overbelastning og har høyere sannsynlighet for å ikke nå miljømålene som er bestemt for virksomheten. Figur 4.2 viser sedimentasjonsrater målt ved to oppdrettsanlegg, ett inne i en fjord og ett ute på kysten, henholdsvis ved anleggene og 800 m fra anleggene (Valdemarsen mfl. 2012; Bannister mfl. 2014). På fjordlokaliteten sedimenterer det meste av det organiske stoffet rett ved anlegget, og tilførslene øker kraftig utover produksjonssyklusen. På kystlokaliteten spres det organiske materialet utover, hvilket resulterer i en langt lavere sedimentasjon, og verdiene er stabile gjennom produksjonsperioden.
Figur 4.2. Sedimentasjonsrater målt over en produksjonsperiode ved to matfiskanlegg (anlegg) og 800 meter fra anleggene (fjern) henholdsvis ute på kysten (a) og inne i en fjord (b). «Tidlig» angir starten av produksjonssyklusen; «Midt» angir midt i produksjonen, mens «Sent» angir måling mot slutten med maksimal biomasse og fôring (Valdemarsen mfl. 2012; Bannister mfl. 2014).
Variasjon i partikkelspredning gjør at størrelsen på påvirkningssonen omkring matfiskanleggene vil variere. Sedimentasjonen vil oftest være størst under og i umiddelbar nærhet av anlegget, og den avtar med økende avstand. De minste fraksjonene av utslippene fra anlegget vil i noen tilfeller kunne spores flere kilometer nedstrøms fra anlegget, men mesteparten av partiklene vil vanligvis bunnfelle mindre enn 500 meter fra merdene. Utviklingen i laksefiskproduksjonen i Norge går mot store anlegg som ligger på bølgeeksponerte, grunne kystlokaliteter og disse ser ut til å ha større bæreevne enn fjordlokaliteter (Keeley mfl. 2019; Keeley mfl. 2010).
4.3 - Lokale effekter av utslipp av partikulært materiale på bløtbunn og hardbunn
Bløtbunn: Nedbrytningen av organisk stoff forbruker oksygen hva enten det foregår ved bakterier eller dyr. Dersom forbruket er større enn tilførslene, oppstår det oksygenmangel i sedimentene og de blir anoksiske. Visse bakterier kan leve uten oksygen og nedbryter det organiske stoffet, men prosessene er langsomme slik at avfallet lettere bygger seg opp og det utvikles giftige gasser som dreper bunndyrene. Det kan også dannes bobler som transporterer partikler og smittestoffer fra bunnen og opp i vannmassene som kan skade fisken i merdene (Hall mfl. 1990; Hansen mfl. 1991; Holmer & Kristensen 1992; Hargrave mfl. 1993; Holmer & Kristensen 1996; Valdemarsen mfl. 2012).
Påvirkning på bunnen vises bla gjennom oksygenforbruket, jo høyere oksygenforbruk jo større er sannsynligheten for anoksisk sediment. Der er stor forskjell på oksygenforbruket på ulike lokaliteter avhengig av hvor mye organisk materiale som er akkumulert. Figur 4.3. viser forskjellen mellom en strømrik lokalitet med lite opphoping av organisk stoff og en strømsvak hvor det organiske stoffet akkumulerte. Oksygenforbruket på den strømrike lokaliteten var relativ lav gjennom produksjonsperioden men økte med økende fiskebiomasse (figur 4.3a). En stasjon 800 meter borte fra anlegget viste også tegn til påvirkning med rikt bunndyrssamfunn og et økende oksygenforbruk på slutten av produksjonsperioden. På den strømsvake stasjonen var oksygenforbruket høyt tidlig i produksjonsperioden men minket ved økende fiskebiomasse, et symptom på at lokaliteten var overbelastet (figur 4.3b). Bunndyrene forsvant, og anoksiske nedbrytningsprosesser tok over. Stasjonen 800 meter lenger vekk var imidlertid lite påvirket, med et relativt fattig bunndyrssamfunn, som er vanlig på dypt vann i fjorder, og et lavt oksygenforbruk gjennom hele produksjonsperioden (Valdemarsen mfl. 2012, 2015; Bannister mfl. 2014).
Figur 4.3. Forbruk av oksygen ved matfiskanlegg (Anlegg) og 800 meter fra anlegg (Fjern) på en strømsterk (a) og på en strømsvak (b) oppdrettslokalitet målt over en produksjonsperiode (tidlig, midt i og sent i produksjonsperioden). Data fra Valdemarsen mfl. (2012) og Bannister mfl. (2014).
Bunndyrsamfunnet er en følsom indikator for organisk påvirkning, og blir mye brukt til overvåking (Black 2001). Som vist i figur 4.4. dannes det ofte en gradient ut fra anleggene. Nær merdene er det først en artsfattig sone med masseforekomst av noen få opportunistiske arter, mens bunndyrsamfunnet i en overgangssone er stimulert med forhøyet antall arter. Lengst ute er dyresamfunnet upåvirket, selv om avfall fra anlegget kan spores med følsomme kjemiske metoder (Kutti mfl. 2007b). Dette samsvarer med det vanlige mønsteret for organisk belastning omkring en punktkilde (Pearson & Rosenberg 1978).
Figur 4.4. Biomasse av bunndyr (B), antall individ (A) og antall arter (S) i ulike avstander fra et oppdrettsanlegg fortøyd i et punkt i en fjord i Hordaland (figur fra Kutti 2008).
Hardbunn: Mange anlegg er i dag plassert i områder hvor man finner hardbunn eller blanding mellom hard og bløtbunn. Man har tidligere antatt at der ville være liten opphoping av organisk materiale fra oppdrett på slike bunner, da der er lite annet materiale som sedimenterer. Mengden organisk materiale som sedimenterer på hardbunn er først og fremst avhengig av strømforholdene, slik det er for bløtbunn. Fauna på hardbunn består av fastsittende organismer som bla svamp og sjøanemoner og bevegelig fauna som bla kråkeboller, sjøpølser og krepsdyr. Organisk materiale som sedimenterer på hardbunn kan enten bli spist av faunaen eller nedbrytes av bakterier slik som på bløtbunn. Vi har imidlertid få undersøkelser som dokumenterer hva som skjer på hardbunn gjennom produksjonsperioden, men der er satt forskning i gang hvor man vil forsøke å definere indikatorer og bestemme grenseverdier for akseptabel påvirkning på hardbunn så det blir mulig å drive overvåking av disse bunntypene slik man gjør på bløtbunn (B og C undersøkelser). Undersøkelse av to dype, hardbunnslokaliteter i en fjord viste at der under produksjonsperioden var akkumulering av organisk stoff og at den opprinnelige faunaen var borte. To typer opportunistiske børstemark dominerte og levde på/av det organiske stoffet. Påvirkningen avtok med økende avstand fra anleggene, og etter om lag 75 meter var de høye tetthetene av børstemark redusert (Hansen mfl. 2011; Eikje 2013). Resultatene svarer til undersøkelser gjort i Canada hvor man også har funnet tap av den opprinnelige faunaen og at opportunistiske børstemark dominerer (Hamoutene mfl. 2016, Salvo mfl. 2017).
4.4 - Regionale effekter av utslipp av organiske partikler
Regionale effekter på bunndyrsamfunn og oksygennivåer i vannsøylen kan oppstå dersom mengden av organisk materiale, f. eks i form av fekalier fra matfiskanlegg som spres utenfor anleggsområdet, blir så stor til at dyresamfunnene på bunn endrer seg. Oksygenforbruket øker og bunndyrssamfunnet endres slik vi kjenner det fra bunnpåvirkning tettere på anleggene. Det økte oksygenforbruket kan gi oksygensvikt i bunnvannet, spesiell der tilførselen av oksygen i vannmassene er dårlig. Effektene av partikulært materiale vil avhenge av størrelsen på utslipp og hydrografiske forhold. Fjordene er mest utsatt for en negativ påvirkning av utslipp fra akvakultur og faktorer som terskeldyp, mengde naturlige tilførsler, vannvolum i dypbassengene og frekvensen av utskiftning av dypvannet vil være avgjørende for hvor mye organisk materiale fjorden tåler (Aure 2013).
4.5 - Rehabilitering av lokaliteter
Ved brakklegging av lokaliteter vil forholdene i sedimentet endre seg mot den tilstand som var før man satte ut fisk. Internasjonal forskning har vist at regenerering av bløtbunn ved permanent brakklegging kan ta fra et par måneder til syv år (Karakassis mfl. 1999; Macleod mfl. 2004; Pereira mfl. 2004). Det er store variasjoner mellom lokaliteter, og regenereringen avhenger blant annet av belastningen på bunnen, bunntypen og strøm. Regenerering av kjemiske forhold i sedimentet kan ta fra en måned til fire år etter endt produksjon, men biologisk regenerering tar vanligvis lengre tid. På hardbunn viser foreløpige resultater at det meste av det organiske materialet og børstemarkene var forsvunnet fra bunnen etter fire måneders brakklegging, men lite av den opprinnelige faunaen var kommet tilbake (Eikje 2013).
4.6 - Sporing av organisk materiale i sedimenter og marine næringskjeder
Oppdrettsanlegg trekker til seg dyr som enten direkte spiser spillfôr eller fekalier, eller bytteetere som spiser disse dyrene (Carss 1990). Sporing av terrestre (landbaserte) fettsyrer som normalt ikke finnes i det marine miljø er en effektiv metode for sporing av utslipp av organisk materiale fra matfiskanlegg til bunnsedimenter og til dyr som spiser mye av oppdrettsavfall (Olsen mfl. 2009, Olsen mfl. 2012, Woodcock mfl. 2017). I tillegg kan undersøkelser av stabile isotoper i fauna og sediment gi utfyllende informasjon. Sporing av DNA fra soyaprodukter i fiskefôr er en ny metode som gir klare signal hos filterfødere (kamskjell) som gjerne får i seg mindre mengder av avfallsprodukter (Woodcock mfl. 2017). Bruk av medisiner med lang halveringstid som tilsettes fôret kan også brukes til sporing av utslipp fra anlegg, men er naturlig nok kun anvendelig ved anlegg som bruker slik medisin (Samuelsen mfl. 2015). Fordelen med å bruke soya- DNA eller medisinrester som sporstoff er at det gir et klart enten eller svar.
Sporingsmetoder er nyttige i områder der undersøkelser viser en dårlig miljøtilstand, og man mistenker at utslipp fra oppdrettsanlegg i nærheten er årsak til den dårlige tilstanden. Man kan også benytte slike sporingsmetoder for å følge utslipp av organisk materiale inn i de marine næringskjedene. Foreløpige studier viser at organisk materiale fra matfiskanlegg går inn i den marine næringskjeden i et vidt spekter av arter. Fisk som sei, torsk, smørflyndre, havmus, brosme og uer spiser av fekalier eller børstemark og andre dyr som lever av fekalier. Vanlig kystreke, sjøkreps og trollhummer i nærheten av anlegg kan også ha fettsyrer i vevet som indikerer at de delvis lever av organiske materiale fra anleggene (Olsen mfl. under arbeid).
Vi har liten kunnskap om hvordan organisk materiale med terrestrisk opprinnelse vil påvirke de marine næringskjeder på sikt. Noen dyr har stor evne til å utnytte dette avfallet og vil dermed ha konkurransefortrinn i forhold til andre arter. Denne matressursen kan også påvirke noen arters funksjonsevne og reproduksjonspotensiale. Nyere studier viser at kråkeboller kan samle seg ved anlegg i fjorder og utnytter avfallet fra oppdrettsanlegg som ekstra matressurs, men når de spiser dette kan både funksjonsevne og reproduksjonssuksess reduseres (White mfl. 2016, White mfl. 2017).
4.7 - Lokale effekter på sårbare bunntyper
Det er ikke gjort en risikovurdering på sårbare bunntyper ennå, men i de neste årene kommer vi til å ha økt fokus på hvordan utslipp av organiske partikler fra matfiskanlegg påvirker sårbare habitater som korallrev, korallskog og svampområder. Dette er naturtyper med en viktig økologisk funksjon langs kysten og i fjordene fordi de har en høy produksjon og er levested for hundretalls andre arter. Disse naturtypene kan ha en forhøyet risiko for påvirkning fordi den enten tåler mindre eller bruker svært lang tid på å reetablere seg. Å etablere et korallrev eller et større kalkalgeområde kan ta tusenvis av år. Kunnskap om dette vil kunne gi en mer optimal plassering av nye anlegg og gi et bedre grunnlag for risikovurdering av effekten av utslipp.
Ut fra den erfaring vi har i dag angående effekter av utslipp på bløtbunn og hardbunn, kan det anslås at utslipp av organiske partikler i umiddelbar nærhet til forekomster av disse naturtypene kan være ødeleggende. Det finnes få nasjonale eller internasjonale vitenskapelige studier om dette og det finnes ingen kunnskap om hvor stor buffersone man bør ha rundt anlegg for å unngå permanent skade.
Koraller: Det er lite kunnskap om hvordan dypvannskorallrev bygget av øyekorall Lophelia pertusa påvirkes av langtidsutslipp fra matfiskproduksjon (Tangen & Fossen 2012). Foreløpige resultater fra et korttidseksperiment har imidlertid vist at organiske utslipp setter ned vekstratene betydelig til Lophelia pertusa nær anleggene (<250 m) og øker erosjonen av det døde kalkskjelettet som revet hviler på (Kutti mfl. 2015). Studier av effekter av utslipp fra akvakultur på korallrev i tropiske og subtropiske områder bekrefter at koraller kan være sensitive overfor utslipp av organiske partikler med negative effekter på både vekst, overlevelse og reproduksjon (Bongiorni mfl. 2003; Villanueva mfl. 2006). I Norge er det begrenset kunnskap om forekomst og utbredelse av korallrev, noe som har bidratt til at en rekke matfiskanlegg i dag ligger plassert slik at de potensielt kan påvirke dypvannskorallrev (Husa mfl. 2016).
Svamp: Nye resultater fra et korttidseksperiment indikerer at forhøyede nivåer av små partikler fra oppdrettsfôr i vannet forårsaker fysiologisk og cellulært stress hos kålrabisvamp Geodia barretti (Kutti mfl. 2016). Hvis dette stemmer kan svampbestander under og nært anlegg forsvinne på sikt, noe som også er indikert fra feltundersøkelser i Hardangerfjorden (Hansen mfl. 2011). En rekke studier fra tempererte og tropiske havområder viser at økt sedimentasjon av uorganiske partikler fører til endringer i struktur og biodiversitet i svampsamfunn og at det også kan føre til sviktende rekruttering hos enkelte svamparter (se for eksempel Fabricius 2005 og Bannister mfl. 2010). Organisk avfall fra oppdrettsanlegg antas å ha en større effekt enn mineralpartikler, siden organisk avfall bruker oksygen når det brytes ned (Weber mfl. 2006, 2012).
4.8 - Overvåking
For å følge med i påvirkningen overvåkes miljøtilstanden i bunnen både rett ved anlegget og i området rundt. I 2000 kom den første standarden for miljøovervåking av bunnpåvirkning fra marine akvakulturanlegg, Norsk Standard NS9410, den ble revidert i 2007 og siste revisjon kom i 2016. I 2005 kom den første forskriften som påla oppdretter å overvåke miljøtilstanden på bunnen ved anleggene. Tidligere hadde det vært opp til myndighetene å bestemme når der skulle utføres miljøundersøkelser.
Overvåkningen av organisk utslipp ved akvakulturanlegg skal følge norsk standard NS9410:2016 (Miljøovervåkning av bunnpåvirkning fra marine akvakulturanlegg). Standarden omfatter to typer undersøkelser, B og C, hvor B-undersøkelsen brukes nær anleggene (anleggssonen) og C-undersøkelsen i området omkring (overgangssonen). Standarden forklarer hvordan undersøkelsene skal utføres, undersøkelsesfrekvens og hvordan resultatene skal vurderes i forhold til definerte miljøstandarder. NS9410:2016 inneholder også en forundersøkelse som skal gjøres på nye lokaliteter før de tas i bruk og ved vesentlig utvidelse av eksisterende lokaliteter. Overvåkingsrapportene leveres til henholdsvis Fiskeridirektoratet (B-undersøkelsen) og Miljødirektoratet (C-undersøkelsen).
4.8.1 - Overvåking i anleggssonen
Miljømål for anleggssonen er at organisk materiale ikke skal akkumulere over tid og påvirkningen skal ikke være større enn at gravende bunndyr kan leve i sedimentet. For anleggssonen gjøres obligatorisk overvåking og der er etablert indikatorer og grenseverdier for påvirkning (NS9410:2016). B-undersøkelsen er en obligatorisk trendovervåkning, og skal brukes nær anlegget der risikoen for påvirkning er størst. Undersøkelsen er en kombinasjon av biologiske, kjemiske og sensoriske variabler og definerer fire miljøtilstander (1-4): Miljøtilstand 1 betyr lite påvirkning, mens tilstand 4 viser stor påvirkning og er definert som overbelastning. Overvåkningen skal utføres ved maksimal belastning og undersøkelsesfrekvensen øker jo dårligere miljøtilstand lokaliteten oppnår. NS9410:2016 inneholder krav om B-undersøkelse også før utsett av ny fisk i anlegget dersom lokaliteten har miljøtilstand 2 eller dårligere. Tilstanden på den enkelte lokaliteten kan endre seg fra produksjonssyklus til produksjonssyklus, men som regel vil man etter noen år kunne justere produksjonen til lokalitetens bæreevne ved hjelp av resultatene fra B-undersøkelsene. Lokalitetens bæreevne er en kombinasjon av de naturgitte forhold som strøm, topografi, bunntype og faunasamfunn og den mengde fisk som oppdrettes, det vil si forholdet mellom lokalitetens sensitivitet og produksjonspresset.
B-undersøkelsen baserer seg på metodikk utviklet for bløtbunn, som er lite egnet til å bestemme påvirkningen på blandingsbunn og hardbunn. NS9410:2016 inneholder veiledning ved undersøkelser på hardbunn men resultatene er usikre. På lokaliteter der det er vanskelig å gjennomføre en B-undersøkelse kan forvaltningen fatte vedtak om alternativ overvåkning. Det satt i gang forskning som skal resultere i overvåking for hardbunn. Resultatene fra B-undersøkelsene rapporteres til Fiskeridirektoratet som kvalitets sikrer og godkjenner undersøkelsene.
Resultatene fra B-undersøkelsene for hele landet gjennom de siste ni årene vises i figur 4.5. Fordelingen av lokaliteter med meget god og god tilstand har vært tilnærmet jevn i perioden, og i 2018 hadde over 90 % av matfiskanleggene meget god eller god tilstand. Antall lokaliteter med dårlig eller meget dårlig tilstand har vært under 10 % i hele tidsperioden.
Figur 4.5. Resultater fra gyldige B-undersøkelser fra hele landet fra 2009 til 2018. Figuren viser hvor stor prosentdel av undersøkelsene som har lokalitetstilstand 1 (meget god, blå), 2 (god, grønn), 3 (dårlig, gul) og 4 (meget dårlig, rød). Kilde: Fiskeridirektoratet.
Antall gjeldende B-undersøkelser i 2018 fordelt på produksjonsområder er vist i tabell 4.1 Lokaliteter som hadde beste lokalitetstilstand i 2017 er med i statistikken, fordi deres siste B-undersøkelse fortsatt var gjeldene ved tidspunktet for gjennomgang av data.
Produksjonsområde
Antall gjeldende B-undersøkelser i 2018
PO1 Svenskegrensen til Jæren
9
PO2 Ryfylke
52
PO3 Karmøy til Sotra
166
PO4 Nordhordland til Stadt
146
PO5 Stadt til Hustadvika
38
PO6 Nordmøre og Sør-Trøndelag
126
PO7 Nord-Trøndelag med Bindal
64
PO8 Helgeland til Bodø
85
PO 9 Vestfjorden og Vesterålen
65
PO10 Andøya til Senja
63
PO11 Kvaløya til Loppa
41
PO12 Vest-Finnmark
61
PO13 Øst-Finnmark
5
Totalt
921
Tabell 4.1. Antall gjeldende B-undersøkelser i 2018 fordelt på fylker. Kilde: Fiskeridirektoratet.
Prosentvis fordelingen av B-undersøkelsene fordelt på lokalitetstilstand og produksjonsområder vises i figur 4.6. For alle produksjonsområdene, bortsett fra Øst-Finnmark, ligger andelen lokaliteter med meget god og god lokalitetstilstand (tilstand 1 og 2) på minst 80%. Total for hele landet lå 14 lokaliteter i tilstand 4 (uakseptabel) og 66 lå i tilstand 3 med risiko for fremtidig overbelastning. Disse lokalitetene følges tett gjennom overvåking for å kunne gripe inn hvis de går i tilstand 4.
Figur 4.6. Prosentvis fordeling av B-undersøkelser i 2018 fordelt på lokalitetstilstand i produksjonssonene. Totalt antall undersøkelser fra hver region finnes i tabell 4.1. Lokalitetstilstanden for den enkelte lokalitet er angitt med fargekode for tilstand 1 (meget god, blå), 2 (god, grønn), 3 (dårlig, gul) og 4 (meget dårlig, rød) Kilde: Fiskeridirektoratet.
4.8.2 - Overvåkning i overgangssonen
Miljømål for overgangssonen er at påvirkningen kun må være liten og på grensen av overgangssonen skal miljøtilstanden være lik området generelt. For overgangssonen gjøres obligatorisk overvåking (C-undersøkelser) og det er etablert indikatorer og grenseverdier for påvirkning (NS9410:2016 og Veileder 02:2013). Her skal man kunne avdekke mindre endringer over tid og undersøkelsen må derfor være mere følsom enn B-undersøkelsen. Hoveddelen er en kvantitativ undersøkelse av bunndyrene, i tillegg kan den omfatte analyser av sediment som kan identifisere utslipp fra oppdrettsanlegg. Prøvetakingsstasjonene er plassert i et område som dekker påvirkningssonen rundt anlegget og den ytterste stasjonen er plassert i en veiledende distanse på 500-600 meter fra anlegget. Utstrekningen av overgangssonen vil være lokalitetsavhengig, og modellering av spredning av organisk avfall samt overvåkning under drift vil bidra til å fastsette yttergrensene for denne. Overvåkningen er risikobasert slik at undersøkelsesfrekvensen øker med økende påvirkning og antall prøvetakingsstasjoner bestemmes av hvor mye fisk som produseres i anlegget. C-undersøkelser har tidligere i hovedsak blitt gjennomført når anleggene har fått pålegg om dette, mens det etter revideringen av standarden i 2016 (NS9410:2016) skal gjennomføres C-undersøkelser ved alle anlegg med en risikobasert frekvens.
Klassifiseringen av faunaprøver gjøres i henhold til veileder: ”Klassifisering av miljøtilstand i vann” (Veileder 02:2013). Det beregnes normaliserte nEQR verdier (normalised ecological quality ratio) for ulike diversitets- og sensitivitetsindekser for faunasammensetningen. Gjennomsnittet av enkeltindeksenes nEQR-verdier brukes til å beregne miljøtilstand på stasjonene i overgangssonen (nEQR) (Veileder 02:2013). En helhetsvurdering av resultatene fra alle stasjoner rundt anlegget gjøres og danner grunnlaget for videre oppfølging av anlegget. Dersom den ytterste stasjonen i overgangssonen har moderat tilstand skal det gjøres ytterligere undersøkelser for å klarlegge om dette gjelder for hele overgangssonen, bakgrunnen for tilstanden og vurderes om det skal settes i gang tiltak.
Lokaliteter der overgangssonen består av hardbunn kan ikke overvåkes med C-undersøkelsen. Forvaltningsmyndighetene har da anledning til å pålegge andre overvåkingsmetoder som for eksempel videofilming. Den pågående forskningen på hardbunns påvirkning vil bli brukt til å definere indikatorer og grenseverdier for påvirkning også i overgangssonen. Miljøovervåkning av overgangssonen har tidligere blitt forvaltet av Fiskeridirektoratet, men fra 2017 er dette ansvaret blitt overført til Miljødirektoratet. Undersøkelsene er ikke kvalitets sikret slik som tidligere, da Miljødirektoratet ikke har fått på plass rutiner for dette. Resultantene brukes derfor med forbehold.
Da stasjonene som inngår i C-undersøkelsen og som ligger nærmere anlegget ikke direkte kan vurderes ut fra miljømål satt i vannforskriften/NS9410-2016 har vi valgt å bruke miljøtilstanden i det ytterste punktet i overgangssonen. Miljøtilstanden i dette punktet skal kunne si noe om hvorvidt anlegget påvirker miljøet i overgangen til regional sone.
C-undersøkelsen utføres ikke hvert år og antall utførte undersøkelser kan derfor variere fra år til år. De fleste C-undersøkelsene utført i 2018 hadde minst god tilstand (tilstand 2) på den ytterste stasjonen i overgangssonen (figur 7).
Figur 4.7. Miljøtilstandsklasse for den ytterste stasjonen i overgangssonen for 88 C-undersøkelser gjennomført i 2018. Miljøtilstandsklasser (gjennomsnittlig indeksverdi, nEQR): 1 (meget god, blå), 2 (god, grønn), 3 (moderat, gul), 4 (dårlig, oransje) og 5 (svært dårlig, rød). Kilder Miljødirektoratet.
Ved innfasingen av den nye veilederen (Veileder 02:2013) ble klassegrensene til enkelte indekser for bløtbunnsfauna endret. Dette var en konsekvens av interkalibrering av indeksene opp mot naboland, som en del av implementeringen av det europeiske vannrammedirektivet. Særlig Shannon Wiener indeksen ble berørt av i denne prosessen, slik at også klassegrensene for nEQR som er en gjennomsnittsverdi av flere indekser endres (se Svåsand mfl. 2016). Som følge av dette er det nå vanskeligere å oppnå meget god tilstand i bunnfaunasamfunn.
Prosentvis fordelingen av C-undersøkelsene fordelt på lokalitetstilstand og fylker vises i figur 4.8. Resultatene vises fylkesvis da de ennå ikke foreligger inndelt i produksjonsområder. I de fleste fylkene lå andelen av lokaliteter med miljøtilstand 2 eller bedre på over 90%.
Figur 4.8. Prosentvis fordeling i miljøtilstandsklasser (nEQR) på den ytterste stasjonen i overgangssonen på 88 matfisklokaliteter i 2018 fordelt på fylker. Tilstandsklasse 1 (meget god), tilstandsklasse 2 (god), tilstandsklasse 3 (moderat), tilstandsklasse 4 (dårlig), tilstandsklasse 5 (svært dårlig). Kilder: Miljødirektoratet.
4.9 - Referanser
Aure J. (2013). Oppdrett og oksygen i Hardangerfjordbassenget. Havforskningsrapporten 2013. 198 s.
Bannister RJ, Battershill CN, de Nys R. (2010). Demographic variability and long-term change in a coral reef sponge along a cross-shelf gradient of the Great Barrier Reef. Marine and Freshwater Research 61, 389-396.
Bannister RJ, Valdemarsen T, Hansen PK, Holmer M, Ervik A. (2014). Changes in benthic sediment conditions under an Atlantic salmon farm at a deep, well flushed coastal site. Aquaculture Environment Interactions 5, 29-47.
Bannister RJ, Askeland Johnsen I, Hansen PK, Kutti T, Asplin L. (2016). Near-and far-field dispersal modelling of organic waste from Atalantic salmon aquaculture in fjord systems. ICES Journal of Marine Science 73, 2408-2419.
Black D. (2001). Environmental Impacts of Aquaculture. Sheffield Academic Press, Sheffield. 212 s.
Bongiorni L, Shafir S, Rinkevich B. (2003). Effects of particulate matter released by a fish farm (Eilat, Red Sea) on survival and growth of Stylophora pistillata coral nubbins. Marine Pollution Bulletin 46, 1120-1124.
Carss DN. (1990) Concentrations of wild and escaped fishes immediately adjacent to fish farm cages. Aquaculture 90, 29-40.
Eikje EM. (2013). Benthic impacts of fish farm waste on hard bottom habitats, the ecology of opportunistic epifauna polychaetes. Master thesis, University of Bergen, Norway. 61 s.
Fabricius KE. (2005). Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Marine Pollution Bulletin 50, 125-146.
Hall POJ, Anderson LG, Holby O, Kollberg S, Samuelsson MO. (1990). Chemical fluxes and mass balances in a marine fish cage farm. I. Carbon. Mar. Ecol. Prog. Ser. 61, 61-73.
Hansen PK, Pittman K, Ervik A. (1991). Organic waste from marine fish farms - effects on the seabed. In: T. Makinen (ed.): Marine aquaculture and environment, Nord 1991:22. s. 105-119.
Hansen PK, Bannister R, Husa V. (2011). Utslipp fra matfiskanlegg. Påvirkning på grunne og dype hardbunnslokaliteter. Rapport fra Havforskningen NR 21-2011. 26 s.
Hamoutene D, Salvo F, Donnet S, Dufour SC. (2016). The usage of visual indicators in regulatory monitoring at hard-bottom finfish aquaculture sites in Newfoundland (Canada). Marine Pollution Bulletin 108, 232-241.
Hargrave BT, Duplisea DE, Pheiffer E, Wildish DJ. (1993). Seasonal changes in benthic fluxes of dissolved oxygen and ammonium associated with marine cultured Atlantic salmon. Mar. Ecol. Prog. Ser. 96, 249-257.
Holmer M, Christensen E. (1992). Impact of marine fish cage farming on metabolism and sulfate reduction of underlying sediments. Mar. Ecol. Prog. Ser. 80, 191-201.
Holmer M, Christensen E. (1996). Seasonality of sulfate reduction and pore water solutes in a marine fish farm sediment: the importance of temperature and sedimentary organic matter. Biogeochem. 32, 15-39.
Karakassis Y, Hatziyanni E, Tsapakis M, Plaiti W. 1999. Benthic recovery following cessation of fish farming: a series of successes and catastrophes. Mar. Ecol. Prog. Ser. 184, 205-218
Keeley N, Valdemarsen, Woodcock S, Holmer M, Husa V, Bannister R. (2019). Resilience of dynamic coastal benthic ecosystems in response to large-scale finfish framing. Aquacult. Envir. Interac.11, 161-179
Keeley NB, Valdemarsen, Strohmeier T, Pochon X, Dahlgren T, Bannister R. (2020). Mixed-habitat assimilation of organic waste in coastal environments – It’s all about synergy. Sci. Tot. Environ. 699, 134281
Kutti T. (2008). Regional impact of organic loading from a salmonid farm – dispersal, sedimentation rates and benthic fauna response. PHD Thesis, University of Bergen. 58 s.
Kutti T, Ervik A, Hansen PK. (2007a). Effects of organic effluents from a salmon farm on a fjord system. I. Vertical export and dispersal processes. Aquaculture 262, 367-381.
Kutti T, Hansen PK, Ervik A, Høisæter T, Johannessen P. (2007b). Effects of organic effluents from a salmon farm on a fjord system. II. Temporal and spatial patterns in infauna community composition. Aquaculture 262(2-4), 355-366.
Kutti T, Nordbø K, Bannister RJ, Husa V. (2015). Oppdrettsanlegg kan true koraller i fjordene. Havforskningsrapporten 2015. Særnummer- Fisken og Havet 1-2015. 220 s.
Kutti T, Krogness C, Husa V. (2016). Effekter av utslipp av oppdrettsfôr på svamp. Rapport fra Havforskningen. Nr. 1-2016. 15 s.
NS 9410:2016. (2016). Norsk Standard: Miljøovervåkning av bunnpåvirkning fra marine akvakulturanlegg). 36 s.
Macleod CK, Crawford CM, Moltschaniwskyj NA. (2004). Assessment of long term change in sediment condition after organic enrichment: defining recovery. Marine Pollution Bulletin 49, 79-88
Olsen SA, Ervik A, Grahl-Nielsen O. (2009). Deep-water shrimp ( Pandalus borealis , Krøyer 1838) as indicator organism for fish-farm waste. J. Exp.Mar.Biol.Ecol. 381, 82-89.
Olsen SA, Ervik A, Grahl-Nielsen O. (2012). Tracing fish farm waste in the northern shrimp Pandalus borealis (Krøyer, 1838) using lipid biomarkers. Aquacult Environ Interact 2, 133–144.
Pearson TH, Rosenberg R. (1978). Macrobenthic successions in relation to organic enrichment and pollution of the marine environment. Oceanogr. Mar. Biol. Ann. Rev. 16, 229-311.
Pereira PMF, Black KD, McLusky DS, Nickell TD. (2004). Recovery of sediments after cessation of marine fish farm production. Aquaculture 235, 315-330.
Salvo F, Mersereau J, Hamoutene D, Belley R, Dufour SC. (2017). Spacial and temporal changes in epibenthic communities at deep, hard bottom aquaculture sites in Newfoundland. Ecological Indicators 76, 207-218.
Samuelsen OB, Lunestad BT, Hannisdal R, Bannister R, Olsen S, Tjensvoll T, Farestveit E, Ervik A. (2015). Distribution and persistence of the anti sea-lice drug teflubenzuron in wild fauna and sediments around a salmon farm, following a standard treatment. Sci. Total Environment 508, 115-121.
Tangen S, Fossen I. (2012). Interaksjoner mellom kaldtvannskoraller og intensivt oppdrett. Kunnskapsstatus og et første skritt mot en konsekvensanalyse. Rapport. Møreforskning Marin. MA 12-10. 43 s.
Valdemarsen T, Bannister RJ, Hansen PK, Holmer M, Ervik A. (2012). Biogeochemical malfunctioning in sediments beneath a Norwegian deep-water fish farm. Environmental Pollution 170, 15-25.
Veileder 02:2013 - revidert 2015. (2015). Klassifisering av miljøtilstand i vann. Økologisk og kjemisk klassifiseringssystem for kystvann, grunnvann, innsjøer og elver. 254 s.
Villanueva RD, Yap HT, Montano MNE. (2006). Intensive fish farming in the Philippines is detrimental to the coral reef-building coral Pocillopora damicornis . Marine Ecology Progress Series 316, 165-174.
White CA, Dworjanyn SA, Nichols PD, Mos B, Dempster T. (2016). Future aquafeeds may compromise reproductive fitness in a marine invertebrate. Marine Environmental Research 122, 67-75.
White CA, Bannister RJ, Dworjanyn SA, Husa V, Nichols PD, Kutti T, Dempster T. (2017) Consumption of aquaculture waste affects fatty acid metabolism of a benthic invertebrate. Science of the Total Environment. 586, 1170-1181.
Woodcock SH, Troedsson C, Strohmeier T, Balseiro P, Sandnes Skaar K, Strand Ø. (2017). Combining biochemical methods to trace organic effluents from fish farms. Aquaculture Environment Interactions. 9, 429-443.
Yrong-Song C, Malcolm CM, Beverage M. (1999). Physical characteristics of commercial pelleted Atlantic salmon feed and considerations of implicates for modeling of waste dispersion through sedimentation. Aquaculture International 7, 89-100.
5 - Miljøeffekter på non-target-arter ved bruk av legemidler
Sjøkreps i forsøk med avlusningsmidler. Foto: Erlend A. Lorentzen/Havforskningsinstituttet
5.1 - Innledning
Her gis en oversikt over legemidler som brukes i norsk akvakultur (t.o.m. 2019) for behandling mot innvollsorm, bakterier (antibakterielle midler) og lakselus (figur 5.1). I tillegg kommer anestesi- og desinfeksjonsmidler. Vi har valgt å fokusere på midler brukt mot lakselus, som gis enten som bademidler eller i fôret, og mulige effekter av disse på andre arter enn lakselus (non-target arter). Kunnskapen i denne rapporten er grunnlaget for risikovurderingen av norsk fiskeoppdrett 2019 (Grefsrud mfl. 2019). Siden juli 2019 har det blitt publisert flere viktige artikler, spesielt på effekter av bademidler på arter relevant for Norge, som vil bli inkludert i kunnskapsstatus for risikovurdering av norsk fiskeoppdrett 2021.
Når det gjelder å sikre at oppdrettsfisk til konsum ikke inneholder rester av legemidler i helseskadelige konsentrasjoner, er det etablert tilbakeholdelsestider, som er tid det skal gå mellom medisinering og slakting av oppdrettsfisk. I tillegg innførte Norge på slutten av 1990-tallet et omfattende kontrollsystem, som følger EUs regelverk (EU 1996). Der analyseres oppdrettsfisk for å sjekke at det ikke er rester av legemidler over den etablerte grenseverdien (Maximum Residue Limit, MRL) (EU 2009). Resultatene for norsk oppdrettslaks publiseres årlig i en egen rapport (Hannisdal mfl. 2018) og vil ikke bli omtalt mer i denne kunnskapsrapporten.
Lakselus tilhører dyregruppen krepsdyr, og medikamenter som dreper disse kan også påvirke andre krepsdyr, alt fra planktonorganismer til krabber og hummer. Spørsmål om negative effekter har særlig vært rettet mot viktige kommersielle arter. I tillegg kommer spørsmål om mulige effekter på planktonorganismer og tidlige livsstadier av fiske- og skalldyrarter. I strandsonen lever skalldyr som kan bli påvirket, men andre organismer som tang og tare vil potensielt også eksponeres. Det gis derfor en oversikt over kunnskapsstatus av effekter av legemidler på non-target arter samt hvordan bademidler og fôrbaserte legemidler spres i miljøet. Det refereres i teksten til behandlingsdose som er den anbefalte doseringen fra produsenten for å behandle laks. Når medikamentet har markedsføringstillatelse, er dose og behandlingsprosedyre godkjent av Statens Legemiddelverk ( www.felleskatalogen.no/medisin-vet ).
Figur 5.1. Skjematisk oversikt over de ulike midlene som går under betegnelsen legemidler i norsk akvakultur. Midlene med heltrukken linje er inkludert i risikovurdering 2019 (Grefsrud mfl. 2019).
5.2 - Desinfeksjonsmidler
Formalin (formaldehyd løst i vann) brukes i fiskeoppdrett både som desinfeksjonsmiddel og mot overflateinfeksjoner på fisk i ferskvann forårsaket av bakterier, parasitter og sopp. Forbruket ble tidligere ikke registrert, men fra 2016 omsettes formalin gjennom godkjente legemiddelgrossister og inngår i registreringene. Ved teknisk bruk kreves ikke forskrivning av autorisert dyrehelsepersonell og bruken reguleres av et annet lovverk en det lovverket som regulerer bruken av legemidler. Forbruket av formalin i 2016 var på 39 815 kg aktiv substans (figur 5.2). Forbruket økte til 46 579 kg i 2017, og i 2019 ble det rapportert forbruk på 41 183 kg. Doser som er beskrevet i litteraturen er 167 og 225 ppm og 0,01% (Treves-Brown 2000, Poppe 1999). Formaldehyd kan produseres naturlig på flere måter og er en del av karbonkretsløpet i naturen. Formaldehyd er godt løselig i vann og metaboliseres via maursyre eller metanol til karbondioksid og vann. Hvor store utslippene av formalin er til miljøet fra oppdrettsvirksomhet er vanskelig å stadfeste.
5.3 - Legemidler mot innvollsorm
Salget av midler mot bendelmark ( Eubothrium sp.) i perioden før 2000 var på mindre enn 240 kg med årlig nedgang fra 2004 til 2009, men har siden hatt en liten årlig økning (figur 5.2). I 2014 ble det benyttet totalt 625 kg, og i 2015 økte forbruket til 942 kg. Forbruket har siden blitt redusert og utgjorde 50 kg i 2019. Det eneste brukte middelet mot innvollsorm i dag er praziquantel. Praziquantel gis til fisken via fôret og kan spres til sedimentet via spillfôr og fekalier. En undersøkelse viste at konsentrasjonen av praziquantel ble halvert i det øverste laget av sedimentet (<2 cm) ila en måned mens i det dypere laget (5-7 cm) var det ingen reduksjon (Hektoen 1995). Effektene av praziquantel på muslinger, snegler, krepsdyr og børstemark var imidlertid små og de vurderte risikoen for påvirkning på miljøet som lav.
5.4 - Antibakterielle midler
Det er få problemer med bakterielle sykdommer i norsk fiskeoppdrett, som har medført lavt forbruk av antibakterielle midler (antibiotika og kjemoterapeutika) over mange år. Denne trenden fortsatte også i 2019. Salget av antibakterielle midler til behandling av oppdrettsfisk, målt som kg aktiv substans, hadde en nedgang fra 511 kg, til 301 kg og 212 kg i perioden 2014-2016, ifølge tall fra Folkehelseinstituttet (figur 5.2). Forbruket var redusert til 213 kg i 2019. I forhold til det totale forbruket (human, veterinær og akvakultur) av antibakterielle midler i Norge, utgjør forbruket i akvakultur rundt 2% av totalen (NORM/NORM-VET 2018). Data fra Veterinært legemiddelregister (VetReg) viser at de fleste resepter er forskrevet til behandling av små rensefisk (tidlige livsstadier) og dermed er hver resept på et lite kvantum sammenlignet med behandling av et stort oppdrettsanlegg. Dette forbruket bidrar derfor lite til det totale forbruket (Hjeltnes mfl. 2019).
Antibakterielle midler gis via fôret og følger for en stor del det organiske avfallet (fekalier og spillfôr) ut i miljøet. Miljøeffektene av en medisinering er derfor i hovedsak begrenset til bunnen i nærområdet rundt anlegg som behandles. Antibakterielle midler er lite toksiske for høyerestående organismer, men kan forårsake utvikling av resistente bakterier. Regelmessig testing av fiskepatogene bakterier isolert fra laksefisk i oppdrett har imidlertid bare avdekket enkelte tilfeller av nedsatt følsomhet for antibakterielle medikamenter (Hjeltnes mfl. 2019). I 2018, som i tidligere år, ble det identifisert nedsatt følsomhet for oksolinsyre i bakterien Flavobacterium psychrophilum isolert fra syk regnbueørret. Mekanismen bak har blitt relatert til kromosomale mutasjoner og derfor vurderes muligheten for overføring av motstandsdyktighet (resistens) til andre bakterier som liten. Det er påvist rester av antibakterielle midler i villfisk og blåskjell fanget nært anlegg under medisinering (Ervik mfl. 1994). På grunn av det lave forbruket av antibakterielle midler og få tilfeller av påvist resistens er det per i dag ikke å anse som noe alvorlig problem.
Figur 5.2. Årlig forbruk (kg) av antibakterielle midler, midler mot innvollsorm, overflateinfeksjon i ferskvann og beroligende og bedøvende midler fra 1990 til 2018 (data fra fhl.no).
5.5 - Legemidler mot lakselus
Basert på hvordan avlusningsmidlene administreres kan en dele medikamentene i to grupper, de som brukes til badebehandling (hydrogenperoksid, azametiphos, cypermetrin, deltametrin) og de som administreres til fisken via fôret (diflubenzuron, teflubenzuron, emamectin-benzoat). Etter et generelt lavt forbruk av midler mot lakselus fra begynnelsen av 2000-tallet økte forbruket årlig fra 2008 og fram til 2015. Fra 2016 har det imidlertid vært en årlig reduksjon i forbruket. Redusert forbruk av legemidler de siste årene er i hovedsak forårsaket av at oppdretterne har tatt i bruk ikke-medikamentelle metoder som varmt vann, ferskvann og mekanisk fjerning av lakselus (se også kunnskapsstatus velferd). Tall for forbruk av alle legemidlene er publisert av Folkehelseinstituttet ( www.fhi.no ).
5.5.1 - Bademidler
5.5.1.1 - Hydrogenperoksid
Hydrogenperoksid er et oksidasjonsmiddel med den kjemiske formelen H2O2 som brukes til avlusning. Når lakselus eksponeres for hydrogenperoksid tas det opp gjennom skallet og oralt. Inni lusa dissosierer hydrogen peroksid til oksygen og vann noe som kan forårsake midlertidig eller permanent skade på innvendige strukturer og som får parasitten til å løsne fra verten og flyte til overflaten (Thomassen 1993, Bruno & Raynard 1994). Virkning av hydrogenperoksid kan også skyldes at substansen har sterk oksiderende effekt på makromolekyler. Det er vist at utvikling av embryo i egg til hunnlusen, samt klekkesuksess påvirkes negativt av hydrogenperoksid (Johnson mfl. 1993, Aaen mfl. 2014). Hydrogenperoksid selges i Norge under navnene Nemona og Paramova. Behandlingsdosen for laks for Nemona varierer fra 1500 til 2100 mg/l, avhengig av temperaturen. Behandlingstiden er anbefalt å vare 20 minutter. Behandlingsdosen ved bruk av Paramova er 1500 mg/l og behandlingstiden er anbefalt å vare 15-20 minutter. For både Nemona og Paramova er det oppgitt (Veterinærkatalogen) at det skal brukes med stor forsiktighet ved sjøtemperaturer mellom 14°C og 18°C. Laks skal ikke behandles hvis temperaturen er over 18°C . Det brukes mot preadult og adult lakselus, når behandling med andre lakselusmidler ikke er hensiktsmessig
5.5.1.1.1 - Forbruk
Hydrogenperoksid brukes både mot lakselus og mot amøbegjellesyke. Statistikken over forbruket viser samlet salg av rent (100 %) hydrogenperoksid. I midten av 1990-tallet begynte næringen å bruke hydrogenperoksid for å bli kvitt lakselus på laksen. Forbruket var veldig lite sammenlignet med dagens nivå (figur 5.3). Fra 1998 til 2009 var stoffet ikke i brukt, men i 2010 og fram til 2013 økte det fra 3 071 til 8 262 tonn. Forbruket var størst i 2015, på 43 246 tonn, men har deretter blitt redusert årlig til 4 523 tonn i 2019.
Figur 5.3. Årlig forbruk av hydrogenperoksid (i tonn) i norsk akvakultur fra 1990 til 2019 (data fra Folkehelseinstituttet).
5.5.1.1.2 - Spredning og fortynning
Hydrogenperoksid spaltes lett til vann og oksygen. I konsentrert form er hydrogenperoksid tyngre enn vann, men på grunn av de små mengdene som tilsettes ved behandling vil tettheten av behandlingsløsningen endres lite i forhold til tettheten av sjøvann. Det kan imidlertid være nok til at behandlingsløsningen kan synke under spesielle forhold. Ved Havforskningsinstituttet arbeides det med å utvikle gode spredningsmodeller for å kunne forutsi hvordan utslipp fra et anlegg vil spres i miljøet. Næringssalter, finpartikulært materiale og badebehandlingsmidler vil i hovedsak følge overflatestrømmen. Strømmene i dypere lag og på bunnen vil bestemme hvor de større organiske partiklene ender opp. Havforskningsinstituttet har gjort noen foreløpige eksperimenter med spredning av en konsentrasjon av et passivt sporstoff basert på numeriske strømmodellresultater fra Hardangerfjorden. Det ble benyttet modellresultater med 200 meters romlige oppløsning. Modellresultatene viste at sporstoffet normalt var fortynnet til under 1% av den opprinnelige konsentrasjonen (behandlingsdosen) etter 24 timer, men at i bakevjer kunne konsentrasjonen fortsatt være opptil 3% (Brokke 2015). Fortynning var høyest i strømrike deler og minst i bakevjer, bukter og andre steder med liten strøm. Liknende resultater er presentert for hydrogenperoksid av Refseth mfl. (2017). I sine simuleringer viste Refseth mfl. (2017) at i enkelte områder kan man finne konsentrasjoner under 10 mg/l hydrogenperoksid etter 4 timer. I strømsvake områder kan slike konsentrasjoner finnes i inntil 10 timer, men utbredelsen vil være mindre i utstrekning. Videre viste simuleringene at konsentrasjoner over 100 mg/l kan man forvente å observere stort sett innenfor en radius på 1-2 km, og 300 mg/l innenfor en radius på 1 km.
5.5.1.1.3 - Nedbrytning
Nedbrytningen (dvs. halveringstid) av hydrogenperoksid i vann er avhengig av flere faktorer som temperatur, pH og tetthet av organiske partikler som hydrogenperoksid kan reagere med (Lyons mfl. 2014). I tillegg kan også tilstedeværelsen av ulike plankton føre til at konsentrasjonen reduseres raskere (Wong mfl. 2003). Bruno & Raynard (1994), i likhet med flere andre studier, fant en halveringstid på omkring 7 dager ved 15°C. Lyons mfl. (2014) beregnet halveringstiden til 28 dager i filtrert sjøvann ved 10°C og en utgangskonsentrasjon på 1386 mg/L. I samme studiet varierte halveringstiden fra 8 til 19 dager ved 10°C når ulike utgangskonsentrasjoner (1800, 1200, 800, 500, 300 mg/L) ble brukt, men resultatene indikerte ingen trend i forhold til utgangskonsentrasjon. Lyons mfl. (2014) fant også at filtrert sjøvann ga raskere degradering enn rå sjøvann med plankton, noe som er meget overraskende. Fagereng (2016) fant derimot at halveringstidene varierte med utgangskonsentrasjon (160, 22, 3 mg/L) og temperatur (8,7 og 12°C). Laveste verdi var 3-4 dager ved 12°C og 28 dager ved 8.7°C. Det er med andre ord stor variasjon mellom de ulike studier og ikke alle oppgir informasjon om vannkvalitet, temperatur eller andre påvirkende faktorer. Det er derfor behov for mer kontrollerte undersøkelser.
5.5.1.1.4 - Tilstedeværelse i miljøet
Tilstedeværelse av hydrogenperoksid har sammenheng med forbruk og hvordan hydrogenperoksid sprer seg, fortynnes og nedbrytes. Det er gjennomført to feltstudier hvor målinger av hydrogenperoksid er gjort i tilknytning til utslipp fra merd. Undersøkelsene er gjort på to oppdrettslokaliteter på Vestlandet en dag tidlig i oktober 2015 (Fagereng & Haavik 2015, Fagereng 2016) og over to dager i midten av april 2016 (Andersen & Hagen 2016, Fagereng 2016). Undersøkelsene i oktober 2015 var ved et anlegg hvor dypet var rundt 190 m, og vannprøver ble tatt i 8 ulike dyp fra 10 til 150 m. 8 ulike tidspunkt ble valgt, fra 2 til 20 minutter etter at hydrogenperoksid ble sluppet. Noen målinger ble også gjort etter 30 og 40 minutter. Prøver ble tatt i tre ulike retninger fra anlegget i to prøveserier. Det var svak strøm på observasjonsdagen. Laksen ble behandlet for AGD med behandlingsdose (1330 mg/l) som er lavere enn ved behandling mot lus. Undersøkelsen i april var gjort ved et anlegg hvor det var ca. 60 m dyp, og det ble gjort prøvetaking ved seks ulike dyp fra 2 til 60 m, på 10 ulike tidspunkt fra 5 minutter før behandling til 25 minutter etter behandling, og ved 4 ulike punkt rundt anlegget hvor den største avstanden var 15 meter fra kanten av anlegget. Det ble gjennomført tre slike serier av undersøkelser. Behandlingsdosen varierte fra 1800 til 2300 mg/l. Sjøtemperaturen var 6°C og strømmen ble vurdert å være svak i området.
I april 2016 ble det målt lave H2O2 konsentrasjoner i majoriteten av målepunktene, men det ble funnet restkonsentrasjoner både i horisontal og vertikal retning fra anlegget, også på siste måletidspunkt dvs. 25 minutter etter slipp ble det målt konsentrasjoner på over 200 mg/l. Totalt var konsentrasjonen i 78% av prøvene under 1 mg/l og i kun 5% av prøvene ble det målt høyere verdier enn 100 mg/l. De høyeste konsentrasjonene ble funnet på 5-20 meters dyp, bortsett fra et målepunkt som ble gjort rett under merden 8 minutter etter slipp. Her hadde hydrogenperoksid sunket til bunnen (60 m). Konsentrasjonen var 723 mg/l, men var redusert til 323 mg/l 7 minutter senere. Totalt sett tyder resultatene på en rasker horisontal enn vertikal spredning (Fagereng 2016).
I oktober 2015 ble det ikke funnet høyere konsentrasjoner enn 5 mg/l i en radius på 70 meter fra anlegget og det ble konkluderte med at hydrogenperoksid ble raskt fortynnet, både i avstand og tid etter behandling (Fagereng & Haavik 2015, Fagereng 2016). Den høyeste konsentrasjonen ble målt til 25 mg/l, målt i én prøve tatt 15 m fra anlegget på 10 m dyp 25 minutter etter utslipp (Fagereng 2016, tabell 4). Det ble ikke observert noe som antydet at hydrogenperoksid sank i dette studiet. Vi vil gjerne presiser at slike feltundersøkelser er meget krevende både med hensyn til fysiske ressurser, mannskap og ikke minst logistikk.
5.5.1.1.5 - Endringer i artssammensetning
Gjennom året vil det naturlig være endringer i artssammensetningen, både i diversitet, mengde og tetthet. For eksempel er det arter som kun finnes i enkelte geografiske områder, og enkelte livsstadier som kun forekommer deler av året. Det kan derfor være utfordrende å skille endringer som følge av eksponering for legemidler og de naturlige endringene. I Skottland ble det i perioden september 1999 til august 2004 undersøkt mulige endringer i flora og fauna ved fire oppdrettslokaliteter som følge av bruk av flere lusemidler (før og etter), deriblant også hydrogenperoksid (SEPA 2005). De fant ingen effekter på dyreplankton, hverken på artsnivå eller samfunnsnivå. Endringer i artssammensetning var ikke forårsaket av lusebehandling, men som for eksempel flekkvis fordeling, sesong og livs-syklus. Videre konkluderte de med at oppblomstringer av planteplankton var også upåvirket av bruken av legemidler. Heller ikke artssammensetningen i noen av de undersøkte områdene var påvirket eller endret som følge av lusebehandlingene. Undersøkelser på bunnslåing av rur viste ingen effekter. Rurlarver er frittsvømmende i vannmassene og kan teoretisk bli påvirket av hydrogenperoksid ved bruk. Det er foreløpig ikke publisert norske studier som viser funn av endringer i artssammensetning eller endringer i en arts mengde som følge av eksponering for hydrogenperoksid.
5.5.1.1.6 - Artenes følsomhet
En arts toleranse for legemiddelet bestemmes på grunnlag av standard toksisitetstester. Mål for overlevelse som benyttes, LC50, er konsentrasjoner som dreper 50% av individene som eksponeres eller utsettes for behandlingen under definerte eksperimentelle betingelser og over et definert tidsrom. Vi har valgt å bruke tilnærmet realistisk eksponeringstid på 1 time i våre studier, supplert med 24-timers eksponering, for å kunne sammenligne med data fra andre publiserte studier.
En times eksponering for hydrogenperoksid hadde liten effekt på testorganismene, bortsett fra hoppekreps (tabell 5.1). Strandreke ( Palaemon elegans ) og tangpungreke ( Praunus flexousus ) eksponert i 1 time med behandlingsløsning på 1700 mg/l gav mindre enn 10% dødelighet (Brokke 2015). Et annet eksperiment undersøkte hvor lang tid det tok før 50% av forsøksdyrene døde ved gitte konsentrasjoner (Brokke 2015). Ved eksponering for 1700 mg/l tok det 1,8 timer å oppnå LC50 for pungreker og 6,3 timer for strandreker. Når konsentrasjonen ble redusert til 425 mg/l, økte tiden til 4,2 timer for pungreker og 23 timer for strandreker (Brokke 2015). I dette forsøket ble rekene eksponert for de ulike konsentrasjonene i 12 timer, og resultatene for strandreke der dødeligheten fortsatte indikerer derfor at en eksponering for hydrogenperoksid har effekt også etter at eksponeringen er over. Når eksponeringstiden ble utvidet til 24 timer var det mulig å beregne LC 50 verdier til 171 mg/l for strandreke og 77 mg/l for pungreke. Torskeegg (Gadus morhua) viste seg imidlertid å være robust ovenfor hydrogenperoksid (Refseth mfl. 2017).
En tilsvarende studie som tidligere beskrevet for andre lusemidler, ble gjennomført med hydrogenperoksid og villfangede hoppekreps (Passamaquoddy Bay, New Brunswick) (Burridge & Van Geest 2014). Hoppekreps som ble eksponert i én time for 1200, 120 og 12 mg/l ble immobilisert i løpet av 15–60 minutter. I to av fire paralleller for de to høyeste konsentrasjonene gav en fargetest indikasjoner på dødelighet (Burridge & Van Geest 2014). Disse resultatene samsvarer godt med lignede studier gjennomført med hoppekreps ( Calanus sp) i Norge (tabell 5.1), med LC 50 35,3 mg/l etter én times eksponering (Escobar Lux 2016). Lengre eksponering (24-96 timer) av hoppekreps for hydrogenperoksid gav betydelig lavere LC 50 verdier (6.0- 2.5 mg/l) (Hansen mfl. 2017). Sukkertare ( Saccharina latissima) er også sensitiv for hydrogenperoksid med registrert dødelighet ved 1 times eksponering for 170 mg/l (10 % av behandlingsløsningen) (Haugland mfl. 2019). Dødelighet ble også observert ved eksponering av Europeisk hummerlarver (Homarus gammarus) i stadium I og II for 10% av behandlingsløsningen i 1 time (Escobar Lux, upublisert). E t utslipp av hydrogenperoksid vil basert på disse toksisitetsstudiene mest sannsynlig ha effekt på ulike planktoniske organismer rundt anlegget, spesielt i de første timene etter utslipp, og ved ugunstige forhold også påvirke organismer i strandsonen som for eksempel sukkertare.
Tabell 5.1. Oppsummering av ulike arters følsomhet for hydrogenperoksid. LC50 (mg/l) er den konsentrasjonen hvor 50% av individene døde. Land er oppgitt fordi ikke alle artene som er oppgitt i tabellen er relevant for norsk fauna. Behandlingsdose med hydrogenperoksid er 1500 til 2100 mg/l.
*Homarus gammarus
** Homarus americanus
5.5.1.2 - Organofosfater
Azametifos hører til gruppen av kjemiske forbindelser som kalles organofosfatene. De virker hemmende på enzymet acetylkolinesterase. Hemming av dette enzymet fører først til overstimulering av musklene, etterfulgt av blokkering som gir lammelse og død. Azametifos selges i Norge under navnet Azasure Vet. Det har effekt på både preadulte og adulte stadier av lakselus. Behandlingsdosen for l aks er 100 μg/l i minst 30 minutter, men ikke lenger enn 60 minutter. Er sjøtemperaturer høyere enn 10°C anbefales det å behandle i maksimum 30 minutter. Neguvon og Nuvan er legemidler hvor virkestoffene er henholdsvis triklorfon og diklorvos. Oppløst i vann spaltes triklorfon til diklorvos, sistnevnte antas å ha sterkest effekt. Triklorfon og diklorvos er organofosfater, med samme virkningsmekanisme som azametifos, men med noe svakere virkning.
5.5.1.2.1 - Forbruk
Forbruket av organofosfater var høyt på begynnelsen av 1980-tallet, med maks forbruk i 1985 på 28 260 kg triklofron (figur 5.4). Fram til 1995 var det triklorfon og diklorvos som hadde størst forbruk, men siden 1997 har azametifos vært enerådende. Fra 2000 til 2007 var ikke azametifos ikke i bruk, mens i 2008 var forbruket 66 kg, for så å øke jevnlig fram til 2014 hvor forbruket var på 4 630 kg og 3904 kg året etter. Siden har forbruket avtatt drastisk, til 154 kg i 2019.
Figur 5.4. Forbruk av organofosfater som legemidler i norsk akvakultur fra 1990 til 2019 (data fra Folkehelseinstituttet). Organofosfatene triklorfon og diklorvos selges ikke lenger i Norge.
5.5.1.2.2 - Spredning og fortynning
Azametifos er svært vannløslig (<1g/L) med en lav oktanol/vann fordelingskoeffisient (log Kow = 1.05). På grunn av den høye vannløseligheten er det veldig liten sannsynlighet for at det binder seg til organisk materiale (SEPA 2005). Spredning etter utslipp vil derfor påvirkes av faktorer som vind og strøm. Modellering gjort på tidlig 1990 tallet i Skottland viste at konsentrasjoner rundt et anlegg vil reduseres til under 0,2 ug/l innen 24 timer for områder uten mye strøm og til 0,02 ug/l i strømrike områder (Turrell 1994 i SEPA 2005). Videre indikerte modellene at azametifos mest sannsynlig vil spres som en sky i de øvre vannlagene, fra 0 til 10 m (SEPA 2005).
5.5.1.2.3 - Nedbrytning
Halveringstid for azametifos er oppgitt å være rundt 9 dager (SEPA 2005). Temperatur var ikke oppgitt, men vi har sett denne halveringstiden både relatert til 8°C og 12°C (ulike kjemikaliekataloger). Det er oppgitt en halveringstid på 10,8 dager ved 20°C og ved pH7 (Worthing& Walker 1987 i Urbina mfl. 2019). Vi har ikke funnet noen av halveringsstudiene publisert i sin helhet. Det er ikke oppgitt om nedbrytning av azametifos påvirkes av sjøtemperaturen.
5.5.1.2.4 - Tilstedeværelse i miljøet
Tilstedeværelse av azametifos har sammenheng med forbruket, hvor mange anlegg som behandles og hvordan azametifos spres, fortynnes og nedbrytes. Etter en behandling ved bruk av presenning slippes behandlingsløsningen direkte i sjøen og kan, avhengig av vær og vind, påvirke strandsonen. Ved bruk av brønnbåt er det mulig å bestemme utslippsområdet og få en raskere fortynning. Det er gjennomført to feltundersøkelser i Canada (nordlige deler av Fundybukta) hvor azametifos ble sluppet sammen med fargestoff for å se spredning etter utslipp (Ernst mfl. 2001, 2014). Den første undersøkelsen ble gjort i oktober 1996, ved tre ulike lokaliteter (uten laks i merd), og viste at det var noe ulik spredning på de tre lokalitetene (Ernst mfl. 2001). Fargestoffet kunne detekteres opp til 5 timer etter utslipp og var synlig i avstand fra 900 m til 3 km. For to av lokalitetene fordelte stoffet seg ned til ca. 10 m, mens for den tredje lokaliteten ble det funnet rester ned til 25 m. De tok vannprøver etter utslipp for følsomhetsundersøkelser på tangloppe, Eohaustorius estuarius, hvorav kun én av 27 vannprøver forårsaket immobilisering etter 48 timers eksponering. Denne prøven ble tatt 30 minutter etter utslipp. Ernst mfl. (2014) gjorde lignede undersøkelse med slipp fra både merd og brønnbåt i september 2010 (med laks i merd og brønnbåt). De fant ved slipp fra merd målbare rester av azametifos (konsentrasjon ikke oppgitt) opp til 1.7 km fra merden og 150 m fra brønnbåt. Høyeste konsentrasjon var på litt over 25 ug/l ca. 1 m fra merdkanten. På grunn av høy mengde organiske partikler i merden gjorde undersøkte de også om azametifos kunne binde seg til disse, men azametifos var i hovedsak oppløst i vann. Også i denne undesøkelsen ble det tatt vannprøver i distanse og tid etter utslipp for å se effekten på tangloppen E. estuarius. Det var kun vannprøver tatt inne i merden før utslipp som forårsaket dødelighet etter 1 times eksponering. De påpekte viktigheten av å gjennomføre 1 times følsomhetstester som er mer realistiske enn for eksempel 48 timers eksponering. Følsomheten var kun 20% eller mindre for vannprøver tatt etter utslipp fra brønnbåt sammenlignet med utslipp fra merd (Ernst mfl. 2014).
Langford mfl. (2015) gjorde undersøkelser ved tre norske anlegg som hadde brukt azametifos og fant konsentrasjoner på opptil 26 ng/l i vannprøver tatt ved ett av anleggene i uken etter behandlingen. Konsentrasjonene ble suksessivt lavere med økende avstand fra anlegget og var 0,5 ng/l én km unna. Det at det ble funnet målbare konsentrasjoner av azametifos så lenge etter en behandling er litt overraskende, men dette ble ikke forklart eller diskutert i rapporten. Denne undersøkelsen indikerer en langsommere fortynningsrate sammenlignet med de utenlandske studiene.
5.5.1.2.5 - Endring i artssammensetning
Gjennom året vil det naturlig være endringer i artssammensetningen, både i diversitet, mengde og tetthet. For eksempel er det arter som kun finnes i enkelte geografiske områder, og enkelte livsstadier som kun forekommer deler av året. Det kan derfor være utfordrende å skille endringer som følge av eksponering for legemidler og de naturlige endringene. Både modeller for spredning av azametifos og målinger gjort i felt etter behandling antyder at individer og arter i de øvre vannlag vil være mest utsatt ved badebehandling.
Burridge & Van Geest (2014) referer til en upublisert studie gjort ved lakseanlegg i New Brunswick Canada august til september 1996, hvor det ble gjort målinger før-under-etter behandlinger med azametifos. De fant ingen endringer i oksygen eller klorofyll-a som indikerer ingen endringer i primær produksjonen. Det er gjort noen få studier hvor individer av ulike arter er plassert i bur i, ved og rundt oppdrettsanlegg som har behandlet med azametifos (Burridge & Van Geest (2014). A merikansk hummer (voksne individer) døde når de var plassert inne i selve behandlingsmerden, men alle overlevde i de burene som var plassert i ulike distanser fra anlegget. Et annet studium i Canada viste lignende resultater for tangloppe Eohaustorius estuarius , hvor kun behandlingsløsningen gav dødelighet etter én times eksponering (Ernst mfl. 2014). Det er foreløpig ikke publisert norske studier som viser funn av endringer i artssammensetning eller endringer i en arts mengde som følge av eksponering for azametifos.
5.5.1.2.6 - Artenes følsomhet
I kontrollerte laboratorieforsøk der ulike non-target arter (strandreke, tangpungreke, hoppekreps, mudderreke Crangon septemspinosa , pungreke Mysid sp.) (se t abell 5.2 ) ble testet viste det seg at effekten var relativt liten ved 1 times eksponering. Unntaket var voksen amerikansk hummer (Homarus americanus) som hadde 50% dødelighet ved en behandlingsløsning på 24,5 μg/l, det vil si en fortynningsfaktor på fire (tabell 5.2). Brokke (2015) fant at etter 24 timers eksponering var LC 50 -verdien høyere enn behandlingsdosen for strandreker, mens tangpungreke var noe mer sensitiv (tabell 5.2). Det er kun publisert et forsøk på gjentatte eksponeringer (Burridge & Van Geest 2014) der amerikansk hummer (voksne) ble eksponert 30 minutter for 1 µg/l azametifos to ganger per dag, i tre dager. I dette forsøket ble det imidlertid ikke registrert dødelighet på hummeren.
Madden mfl. (1991) undersøkte akutt dødelighet på sildelarver. De eksponerte larvene i 96 timer for konsentrasjoner 0-200 ug/l. Ingen dødelighet ble funnet i løpet av denne tiden. Testing ved høyere konsentrasjoner gjorde at LC 50 ble estimert til 250 ug/l ved 96 timers eksponering. De fant at ved behandlingsdosen var 77-78% av AcHE aktviteten redusert i forsøksdyrene. Robertson mfl. 1992 eksponerte europeisk hummerlarver (uten å definere hvilket stadium) i 1 time og fant 100% dødelighet ved eksponering for 100 ug/l. Og 14 av 20 individer (70%) ved 10 ug/l ble immobilisert og døde etter to dager i friskt vann. Når europeisk hummer ble eksponert for diklorvos i en konsentrasjon på 100 μg/l ble det registrert én død hummer etter 4,5 timer og én etter 6 timer (Egidius & Møster 1987). I den samme undersøkelsen ble det vist at både strandkrabbe ( Carsinus maenas ) og taskekrabbe ( Cancer pagurus ) var mindre følsomme enn hummer for eksponering av diklorvos, og overlevde 10 timer i en konsentrasjon på 1000 μg/l.
Tabell 5.2. Oppsummering av ulike arters følsomhet for azametifos. LC50 (mg/l) er den konsentrasjonen hvor 50% av individene døde. NOEC – er den laveste konsentrasjonen som ikke ga noe dødelighet. Land er oppgitt fordi ikke alle artene som er oppgitt i tabellen er relevant for norsk fauna. Behandlingsdose med azametifos er 100 μg/l.
*Homarus gammarus
** Homarus americanus
5.5.1.3 - Pyretroider (deltametrin og cypermetrin)
Deltametrin og cypermetrin tilhører stoffgruppen pyretroider. Disse stoffene påvirker nervecellene ved å forsinke/forhindre at Na + kanalene lukkes etter depolarisering og forstyrrer således en normal repolarisering av cellene. Dermed blokkeres overføring av impulser i parasittenes nervebaner, som igjen medfører eksitasjon, paralyse og død hos lakselusa. Deltametrin selges i Norge under navnet Apha Max. Anbefalte dosering av deltametrin er 2 µg/l (tilsvarende 2000 ng/l), og behandlingstiden er 30 minutter. Det er oppgitt at effekten reduseres ved sjøtemperaturer lavere enn 6°C. Det påpekes at nøtene bør være rene ved behandling for å sikre mest mulig effekt. Deltametrin skal ikke benyttes hvis det oppbevares hummer eller krabbe i umiddelbar nærhet (<200 m).
Cypermetrin var sist solgt i Norge i 2017, og da ble det solgt under navnet Excis vet. og Betamax vet . Cypermetrin er noe mindre potent enn deltametrin, og anbefalte doseringer var 5 µg/l (tilsvarende 5000 ng/l) som Excis vet og 15 µg/l (tilsvarende 15 000 ng/l) som Betamax vet . Pyretrum (solgt under navnet Py-Sal) ble brukt kun i noen få år i Norge, 1993- 1996.
5.5.1.3.1 - Forbruk
Pyretroider ble tatt i bruk i 1994, og da som pyretrum, med et totalt forbruk på 32 kg (figur 5.5). Pyretrum ble noen år seinere erstattet av delta- og cypermetrin. Det totale årlige forbruket fram til 2000 var i snitt 31 kg, men økte så til 96 kg, fordelt på 23 kg deltametrin og 73 kg cypermetrin. I 2009 økte igjen de årlige forbruket til 150 kg, 62 og 88 kg for delta- og cypermetrin. I 2012 til 2014 var forbruket på henholdsvis 353, 347 og 320 kg, og avtok til å utgjøre kun 10 kg deltametrin i 2018 og 2019.
Figur 5.5. Årlig forbruk av pyretroider (i kg) som legemidler i norsk akvakultur fra 1990 til 2019 (data fra Folkehelseinstituttet).
5.5.1.3.2 - Spredning og fortynning
Deltametrin og cypermetrin er lite løselig i vann (5-10 ug/l) og binder seg lett til andre stoffer og partikler (log Kow >4) (SEPA 2005). Etter en behandling ved bruk av presenning slippes behandlingsløsningen direkte i sjøen og kan, avhengig av vær og vind, påvirke strandsonen. Ved bruk av brønnbåt er det mulig å bestemme utslippsområdet og få en raskere fortynning. Ved Havforskningsinstituttet arbeides det med å utvikle gode spredningsmodeller for å kunne forutsi hvordan utslipp fra anlegg og brønnbåt vil spres i miljøet. Næringssalter, finpartikulært materiale og badebehandlingsmidler vil i hovedsak følge overflatestrømmen. Strømmene i dypere lag og på bunnen vil bestemme hvor de større organiske partiklene ender opp. Van Geest mfl. (2014a) fant veldig rask reduksjon av deltametrin i kar som også inneholdt tangloppearten Echinogammarus finmarchicus , 77% reduksjon innen 24 timer. Dette betyr at selv om fortynning er den viktigste årsaken til reduksjon i konsentrasjonen av deltametrin vil andre faktorer som binding/absorpsjon til organisk materiale og annet materiale som plastikk, også bidra. Willis mfl. (2005) simulerte spredning og fortynning av cypermetrin fra et anlegg i Skottland. De høyeste konsentrasjonene i deres modeller var 25 minutter etter behandling, og verdiene varierte fra 195 til 218 ng/l for tre ulike simuleringer (dag 1, 2 og 6 i deres studie). M odelleringsstudiene viste altså at cypermetrin raskt ble fortynnet og spredt etter utslipp. En studie fra Canada viste at konsentrasjoner av cypermetrin på 20-40 ng/l kunne finnes opptil 4 til 5 timer etter utslipp (Ernst mfl. 2001).
5.5.1.3.3 - Nedbrytning
Van Geest mfl. (2014a) fant 77% reduksjon av deltametrin etter 24 timer i kar som også inneholdt tangloppearten Echinogammarus finmarchicus , men det oppgis ikke om denne reduksjonen var som følge av binding til annet materiale eller om det skyldtes ren nedbrytning. To andre studier viser helt motsatte resultater av hverandre. Mens Maguire mfl. (1989) beregnet en halveringstid av deltametrin på mindre enn 3 timer i renset sterilt vann uten tilgang på lys, publiserte Tomlin (1994) halveringstiden for deltametrin løst i vann med temperatur på 25 og pH 9 til å være 2,5 dager. Nye studier bør gjøres for å bestemme halveringstiden i sjøvann.
5.5.1.3.4 - Tilstedeværelse i miljøet
Tilstedeværelse av pyretroidene delta- eller cypermetrin har sammenheng med forbruk og hvordan stoffene sprer seg, fortynnes og nedbrytes. I en feltundersøkelse i Canada hvor behandlingsløsningen med deltametrin var tilsatt fargestoff ble det tatt vannprøver av utslippsvannet i avstander inntil 1500 m fra anlegget (Ernst mfl. 2014). Restkonsentrasjoner ble ikke analysert, men brukte tangloppearten E. estuarius (48 timers LC50 = 1,0-3,6 µg/l) som indikator (Ernst mfl. 2001). Vannprøver tatt inntil 500 m fra anlegget ga noe dødelighet ved 1 times eksponering, men hvis tangloppen ble eksponert for 48 timer ble det observert dødelighet opptil 1000 m fra anlegget. I et feltstudium gjennomført i Norge ble strandreke satt i bur i ulike dyp og avstander fra et oppdrettsanlegg som behandlet laksen med deltametrin. Det ble ikke observert dødelighet i grupper som var plassert lengre enn 50 m fra anlegget (Grøsvik & Andersen 1997) . I en annen norsk undersøkelse fra 2014 ble det ikke funnet pyretroider, verken i vann- eller sedimentprøver, men små rester ble funnet i blåskjell ved anlegget (4,8 ng/g) (Langford mfl. 2015). Prøvene var tatt ved et anlegg en uke etter medisinering med azametifos og muligens også cypermetrin. En lignende undersøkelse i Skottland viste at den høyeste målte konsentrasjonen var 187 ng/l, 25 minutter etter utslipp ca 25 fra anlegget i strømretningen (Hunter & Fraser 1995 i Willis mfl. 2005). Pyretroider er lite vannløselige og disse forbindelsene vil kunne binde seg til ulike substanser , partikler og sediment slik at konsentrasjonen i vannfasen reduseres raskere enn bare ved kjemisk nedbrytning eller fortynning. På den annen side inneholder produktene dispergeringsmidler og stabilisatorer, noe som kan bidra til å holde disse stoffene i løsningen.
5.5.1.3.5 - Endringer i artssammensetning
Gjennom året vil det naturlig være endringer i artssammensetningen, både i diversitet, mengde og tetthet. For eksempel er det arter som kun finnes i enkelte geografiske områder, og enkelte livsstadier som kun forekommer deler av året. Det kan derfor være utfordrende å skille endringer som følge av eksponering for legemidler og de naturlige endringene. I Skottland ble det i perioden september 1999 til august 2004 undersøkt mulige endringer i flora og fauna ved fire oppdrettslokaliteter som følge av bruk av flere lusemidler (før og etter), deriblant også cypermetrin (SEPA 2005, Willis mfl. 2005). De fant ingen effekter på dyreplankton, hverken på artsnivå eller samfunnsnivå. De fant endringer i artssammensetning, men dette ble forklart med flekkvis fordeling, sesong og livs-syklus. Videre konkluderte de med at oppblomstringer av planteplankton også var upåvirket av bruken av legemidler. Undersøkelser på bunnslåing av rur viste ingen effekter. Rurlarver er frittsvømmende i vannmassene og kan teoretisk bli påvirket av badmidler som cypermetrin. Det er foreløpig ikke publisert norske studier som viser funn av endringer i artssammensetning eller endringer i en arts mengde som følge av eksponering for pyretroider.
5.5.1.3.6 - Artenes følsomhet
En arts toleranse for legemiddelet bestemmes på grunnlag av standard toksisitetstester. Mål for overlevelse som benyttes, LC50, er konsentrasjoner som dreper 50% av individene som eksponeres eller utsettes for behandlingen under definerte eksperimentelle betingelser og over et definert tidsrom. Vi har valgt å bruke tilnærmet realistisk eksponeringstid på 1 time i våre studier, supplert med 24-timers eksponering, for å kunne sammenligne med data fra andre publiserte studier.
Eksponeringsforsøk i laboratorier viser at larver av amerikansk hummer er spesielt sensitive (tabell 5.3). Det er foreløpig ikke gjennomført lignende studier med europeisk hummer. Undersøkelser ved Havforskningsinstituttet (Brokke 2015) ga lignende resultater (LC 50 ) for strandreker og tangpungreker som for mudderreke i Canada, både etter 1 og 24 timers eksponering. Innfangete hoppekreps (Passamaquoddy Bay, New Brunswick) eksponert for 200 og 2000 ng/l deltametrin ble ubevegelige (immobile) og sank til bunnen av tanken i løpet av 15–60 minutter (Burridge & Van Geest 2014). Etter fem timer i friskt vann var de fremdeles immobilisert, men ingen dødelighet ble observert.
En studie i Skottland brukte LC 10 for fem ulike arter av krepsdyr (hoppekreps, pungreke, mudderreke, gulfreke, vinkekrabbe) og en fiskeart (tannkarpe) til å beregne sensitivitetskurver med formål å definere grenseverdier for miljøeffekt av deltametrin (Crane mfl. 2011). Analysene viste at en konsentrasjon på 9,3 ng/l var grenseverdien for å beskytte de mest sensitive artene ved en eksponeringstid på 3 timer. Ved 24 timers eksponering ble grenseverdien beregnet til 2,5 ng/l. Noen av artene som er brukt i dette studiet er mindre aktuelle når en snakker om norske forhold. Derfor bør en slik analyse også gjennomføres med relevante norske arter. Eksponering av børstemark ( Nereis virens ) for behandlingsløsning gav ingen effekt, mens eksponering i syv dager for sediment som inneholdt deltametrin (160-400 ng/g) gav både dødelighet, redusert graveevne og redusert bevegelse (Van Geest mfl. 2014a). For tangloppen E. finmarchicus eksponert i 10 dager for sediment tilsatt deltametrin, var LC 50 -verdien 16 ng/g (Van Geest mfl. 2014b). Studiene viser at deltametrin er svært toksisk for enkelte non-target organismer, og et utslipp vil sannsynligvis påvirke planktoniske organismer i nærområdet.
Art
Eksponerings-tid
LC50 (ng/l)
Land
Kilde
Strandreke (Palaemon elegans)
1 time
187.0
Norge
Brokke 2015
Tangpungreke (Praunus flexuosus)
1 time
154.4
Norge
Brokke 2015
Europeisk hummer*, larve (stadium I)
1 time
2.6
Norge
Parsons mfl.; upubliserte data
Europeisk hummer, larve (stadium II)
1 time
2.9
Norge
Parsons mfl.; upubliserte data
Amerikansk hummer**; stadium I
1 time
3.4
Canada
Burridge & Van Geest 2014
Amerikansk hummer; stadium III
1 time
36.5
Canada
Fairchild mfl. 2010
Amerikansk hummer; voksen
1 time
18.8
Canada
Burridge & Van Geest 2014
Tangloppe (Eohaustorius estuarius)
1 time
13.1
Canada
Fairchild mfl. 2010
Tangloppe (Echinogammarus finmarchicus)
1 time
70.0
Canada
Van Geest mfl. 2014b
Pungreker (Mysid sp.)
1 time
13.9
Canada
Burridge & Van Geest 2014
Mudderreke (Crangon septemspinosa)
1 time
142.0
Canada
Fairchild mfl. 2010
Strandreke (Palaemon elegans)
24 timer
12.3
Norge
Brokke 2015
Tangpungreke (Praunus flexuosus)
24 timer
27.8
Norge
Brokke 2015
Amerikansk hummer; stadium I
24 timer
0.8
Canada
Burridge mfl. 2014
Amerikansk hummer; stadium II
24 timer
0.6
Canada
Burridge mfl. 2014
Amerikansk hummer; stadium IV
24 timer
1.7
Canada
Burridge mfl. 2014
Amerikansk hummer; voksen
24 timer
15.0
Canada
Burridge mfl. 2014
Tangloppe (Echinogammarus finmarchicus)
24 timer
9.4
Canada
Van Geest mfl. 2014b
Pungreker (Mysid sp.)
24 timer
1.4
Canada
Burridge mfl. 2014
Mudderreke (Crangon septemspinosa)
24 timer
27.0
Canada
Burridge mfl. 2014
Tabell 5. 3. Oppsummering av ulike arters følsomhet for deltametrin. LC50 (ng/l) er den konsentrasjonen hvor 50% av individene døde. Land er oppgitt fordi ikke alle artene som er oppgitt i tabellen er relevant for norsk fauna. Behandlingsdose med deltametrin er 2000 ng/l.
*Homarus gammarus
** Homarus americanus
5.5.2 - Fôrmidler
5.5.2.1 - Flubenzuroner (diflu- og teflubenzuron)
Flubenzuroner er betegnelsen på en gruppe avlusningsmidler som administreres til fisken via fôret. Stoffene virker ved å hemme syntesen dvs. produksjon av kitin som er et hornaktig stoff som bygger opp hudskjelettet hos blant annet kreps og insekter. I norsk fiskeoppdrett brukes både diflubenzuron og teflubenzuron. Diflubenzuron selges som Reeleze vet i Norge og anbefalt dosering er 3-6 mg/kg laks per dag i 14 påfølgende dager. Teflubenzuron selges som Ektobann og anbefalt dosering er 10 mg/kg laks per dag i syv påfølgende dager. Bør ikke brukes ved sjøtemperaturer <9 ̊C. Legemidlene overføres fra vert (i dette tilfellet laks) til parasitt (lakselus) ved at parasitten spiser slim, hud og blod på fisken. Flubenzuroner er effektive mot alle stadier av parasitten som gjennomgår et skallskifte. Behandling med flubenzuroner bør ikke gjentas før tidligst etter 12 uker på grunn av akkumulering og lang halveringstid i miljøet.
Basert på innspill fra blant annet Havforskningsinstituttet fastsatte Nærings og Fiskeridepartementet i 2017 nye regler som skal bidra til å hindre uakseptable miljøeffekter av legemidler brukt i oppdrettsnæringen, inkludert kitinsyntesehemmere.
Akvakulturdriftsforskriften § 15a. Vilkår for bruk av kitinsyntesehemmere. «Kitinsyntesehemmere gitt gjennom fôret kan ikke brukes på samme lokalitet før det er gått minst 6 måneder fra siste behandling. Dette gjelder uansett hvilken kitinsyntesehemmer som er benyttet. Kitinsyntesehemmere kan ikke brukes på lokaliteter nærmere enn 1000 meter fra rekefelt jf. de felt som til enhver tid vises i Fiskeridirektoratets nettbaserte kartverktøy. Avstanden måles som beskrevet i § 18 andre punktum.»
5.5.2.1.1 - Forbruk
Flubenzuroner ble første gang tatt i bruk 1996, og da var det totale forbruket på 770 kg hvorav 79% var teflubenzuron (figur 5.6). De var i bruk frem til 2001, for så ikke bli brukt igjen før i 2009 og da med et forbruk på 3 441 kg, hvorav 59% var teflubenzuron. Høyeste årlige forbruk ble registrert i 2016 med 4 824 kg diflubenzuron og 4 209 kg teflubenzuron. Siden har det årlige forbruket blitt kraftig redusert, til 1 479 kg i 2019 der diflubenzuron utgjorde 88%.
Figur 5.6. Årlig forbruk av flubenzuroner (i kg) i norsk akvakultur, fra 1990 til 2019 (data fra Folkehelseinstituttet).
5.5.2.1.2 - Spredning og fortynning
Flubenzuroner gis i fôret som fisken spiser, men opptaket fra laksens tarm er imidlertid lavt til moderat (biotilgjengelighet 10–30%). Siden laksen i liten grad omdanner disse stoffene, vil mesteparten bli utskilt i uforandret form via urin eller bundet til fekalier. Medikamentene tilføres dermed miljøet som spillfôr under behandling, og via fekalier og urin i opptil et par uker etter at medisineringen er avsluttet. Løseligheten i vann er derimot liten for både di- og teflubenzuron, og begge stoffene har vannavvisende (hydrofobe) egenskaper.
Spredning og fortynning av flubenzuroner vil altså være både som oppløst i vann og at stoffene binder seg til tilgjengelig organisk materiale. Det at stoffene lett binder seg til organisk materiale vil bidra til å redusere konsentrasjonen i vannfasen. Denne prosessen er beskrevet fra mindre ferskvannsdammer og små estuarier, men ikke i et marint system. Ut lekking av flubenzuroner fra organisk materiale, som pellets og fekalier, til vannfasen vil være liten ( Selvik mfl. 2002, Samuelsen 2016 ). Flubenzuroner vil derfor hovedsakelig spres med det organisk materiale til sedimentet, og vil følge samme spredningsmønster som organisk avfall fra anlegget. Tilførselen til miljøet vil være begrenset til perioden med medisinering og en kort tid etter (Samuelsen mfl. 2015). Kjemisk analyse har vist at fekalier kan ha betydelig høyere konsentrasjoner av medikamentene enn nivået var i den medisinerte pelleten (Samuelsen upubl. data). Medisinerte pellets og intakte fekalier synker raskt, mens små svevepartikler, som kan utgjøre om lag 5–10%, kan spres med strømmen over større områder. Opp til 2,7% av partiklene kan nå lengre enn 2 km fra anlegget (Bannister mfl. 2016), men spredningen er avhengig av strømhastigheten på de ulike lokalitetene. Mengden av flubenzuroner i sedimentet vil derfor være størst nær anlegget.
5.5.2.1.3 - Nedbrytning
Kjemisk nedbrytning av flubenzuroner løst i vann kan forekomme ved spaltning (hydrolyse) og ved påvirkning av sollys (foto-nedbrytning). Oppløst i vann brytes både diflubenzuron og teflubenzuron ned under påvirkning av naturlig lys, men nedbrytningshastigheten påvirkes av vannets pH og temperatur og er beskrevet å være raskere under basiske forhold (Mabury & Crosby 1996, Marsella mfl. 2000). Hydrolyse av diflubenzuron er beskrevet av Ivie mfl. (1980) og Boelhouwers mfl. (1988) med halveringstider på 7 til 32 dager. Andre studier derimot hevder at hydrolyse av flubenzuroner i vann er en svært langsom prosess eller helt fraværende (Marsella mfl. 2000). Betydningen av hydrolyse og foto-nedbrytning av flubenzuroner i sjøvann ved aktuelle temperaturer og lysregimer fra norske farvann er ikke kjent.
To laboratorieundersøkelser viser at flubenzuroner bundet til organisk materiale i marine sedimenter, under både oksygenrike og oksygenfattige forhold, er veldig stabile (Selvik mfl. 2002, Samuelsen 2016). Marint sediment ble anriket med flubenzuroner enten som knuste eller hele medisinerte pellets eller som medisinholdige fekalier og plassert i tanker med rennende vann. Forsøkene gikk over 6 måneder, og resultatene viste ingen signifikant reduksjon i konsentrasjonen av verken diflubenzuron eller teflubenzuron, til tross for stor mikrobiell aktivitet. Dette betyr at mikrobiell nedbrytning og utlekking av flubenzuroner fra organisk materiale til vannfasen er svært liten. Feltundersøkelser konstaterer en gradvis reduksjon av flubenzuroner i sedimentet over tid, og basert på slike undersøkelser er det beregnet halveringstider på 110–170 dager for teflubenzuron (SEPA 1999a, Samuelsen mfl. 2015). For diflubenzuron viste forsøk fra NIVA (1998) at halveringstiden varierte med temperatur, fra 100 dager ved 5°C til 30 dager ved 15°C.
Eliminasjonshastigheten av flubenzuroner i atlantisk laks viser en halveringstid på 2,5 og 6,5 dager avhengig av temperatur (EMEA 1999ab). I yngel av europeisk hummer ble halveringstiden av teflubenzuron beregnet til 3,5 dager ved 14 o C (Samuelsen mfl. 2014). At flubenzuroner skilles ut så raskt fra både laks og hummer indikerer at stoffene ikke vil akkumulere i næringskjeden, i motsetning til hva en ser for en del ikke nedbrytbare (persistente) organiske miljøgifter.
5.5.2.1.4 - Tilstedeværelse i miljøet
Tilstedeværelse av flubenzuroner i miljøet har sammenheng med forbruk og hvordan stoffene sprer seg, fortynnes og nedbrytes. Flubenzuroner oppløst i vannfasen har blitt funnet opptil 1100 m fra oppdrettsanlegg og i en periode på minst to uker etter avsluttet medisinering ( NIVA 1998, Langford mfl. 2011). Den høyeste konsentrasjonen ble målt til 295 ng/l diflubenzuron i en vannprøve tatt 300 meter fra et anlegg som brukte dette medikamentet. Andre undersøkelser har imidlertid påvist bare små konsentrasjoner av flubenzuroner i vannfasen i forbindelse med medisinering (Samuelsen mfl. 2015).
Diflubenzuron og teflubenzuron er funnet i bunnsediment og i organiske svevepartikler samlet inn ved anlegg og i avstand på inntil 1100 m (Langford mfl. 2011, Samuelsen mfl. 2015). Restkonsentrasjonene i prøvene tatt på stasjoner lengst fra anleggene var imidlertid små. Høyeste konsentrasjon av teflubenzuron målt i sediment ved et anlegg var 40,0 mg/kg (våtvekt) (Samuelsen mfl. 2015). Tilsvarende konsentrasjon av diflubenzuron var 49,5 mg/ kg sediment (tørrvekt) (NIVA 1998). Havforskningsinstituttet gjennomførte feltundersøkelser i 2015 og 2016 i områder med anlegg som tidligere hadde brukt både di- og teflubenzuron (Samuelsen upubliserte data). Undersøkelsene avdekket at alle sedimentprøvene samlet inn mellom to anlegg (avstand på ca. 2 km) inneholdt restkonsentrasjoner av di- og teflubenzuron. De høyeste nivåene var 1,23 mg/kg for teflubenzuron og 0,25 mg/kg for diflubenzuron i prøver tatt like ved anleggene. Sedimentprøver fra to andre anlegg viste lignende resultater med høyest konsentrasjoner rett ved anleggene og avtagende konsentrasjoner med økende avstand. I en undersøkelse som inkluderte flere anlegg i Skottland ble de høyeste konsentrasjonene av teflubenzuron funnet like ved anleggene og varierte fra 0,2 mg/kg til 15,0 mg/kg (SARF098 2016). I avstand 100 m fra anleggene varierte konsentrasjonene fra 0 til 2,8 mg/kg. Langford mfl. (2011) konstaterte spredning av flubenzuroner til strandsonen der restkonsentrasjoner ble funnet i en prøve med tanglopper Disse undersøkelsene bekrefter at flubenzuroner kan spres over et stort område, og at de kan detekteres i sedimentet i lang tid etter bruk.
Feltundersøkelser både i Skottland og Norge konstaterte altså en gradvis reduksjon av flubenzuroner i sedimentet over tid (SEPA 1999a, Samuelsen mfl. 2015). Mulige årsaker til en slik reduksjon kan være nedbrytning av organiske materiale, at organismer som børstemark som inneholder medisinrester blir spist av andre organismer, eller at andre mer mobile organismer enn børstemark også konsumerer organisk materiale med medisinrester. Organiske partikler kan også transporteres til andre område via vannstrømmene.
Non target arter som reker, krabber og sjøkreps kan konsumere medisinholdig organisk materiale direkte eller få det via konsum av andre arter (Samuelsen mfl. 2015). Samuelsen mfl. (2015) fant rester av teflubenzuron i 24 av 30 arter (fisk og krepsdyr) som ble undersøkt i tilknytning til et anlegg like etter medisinering. Artene ble fanget inn maksimum 300 m fra anlegget. Konsentrasjonene var generelt lave, men det ble funnet høye verdier i enkeltindivider av for eksempel sei ( Pollachius virens ), noe som kan tyde på konsum av pellet fra oppdrettsanlegget. Høyest var likevel konsentrasjonen i børstemark ( Capitella capitata, Nereis sp.) som lever i sedimentet ved anlegget. Åtte måneder senere var det fremdeles høye konsentrasjoner i børstemark , mens kun små mengder ble funnet i enkelte individer av dypvannsreker (<16 ng/g) og i brunmaten i taskekrabbe (20 ng/g) og sjøkreps (<45 ng/g). Rester av teflubenzuron i prøver av pelagisk eller bunnlevende fisk kunne derimot ikke påvises i prøver samlet inn åtte måneder etter medisinering. Andre feltundersøkelser har påvist flubenzuroner i taskekrabbe, dypvannsreker og amfipoder (NIVA 1998, Langford mfl. 2011). En feltundersøkelse som Havforskningsinstituttet gjennomførte i 2015, i et område med to oppdrettsanlegg som tidligere hadde brukt både di- og teflubenzuron, påviste restkonsentrasjoner i eremittkreps ( Pagurus sp.), kongsnegl ( Buccinum undatum ), svartslangestjerner ( Ophiocomina nigra ), blomsterreker og sjøkreps (Samuelsen upubliserte data). Generelt var konsentrasjonene lave i alle artene, med høyest verdi på 53 ng/g funnet i brunmaten av sjøkreps. I en tidligere feltundersøkelse ble det påvist enkeltprøver av sei som var høyere enn MRL-verdien som er satt til 500 ng/g (Samuelsen mfl. 2015).
5.5.2.1.5 - Endringer i artssammensetning
Gjennom året vil det naturlig være endringer i artssammensetningen, både i diversitet, mengde og tetthet. For eksempel er det arter som kun finnes i enkelte geografiske områder, og enkelte livsstadier som kun forekommer deler av året. Det kan derfor være utfordrende å skille endringer som følge av eksponering for legemidler og de naturlige endringene. Det er foreløpig ikke publisert norske studier som viser funn av endringer i artssammensetning eller endringer i en arts mengde som følge av eksponering for flubenzuroner.
5.5.2.1.6 - Artenes følsomhet
En arts toleranse for legemiddelet bestemmes på grunnlag av standard toksisitetstester. Mål for overlevelse som benyttes, LC50, er konsentrasjoner som dreper 50% av individene som eksponeres eller utsettes for behandlingen under definerte eksperimentelle betingelser og over et definert tidsrom. Siden di- og teflubenzuron påvirker kitin-syntesen vil dyregrupper med kitin i skallet være spesielt sårbare. Dødelighet inntreffer i tilknytning til skallskifte og arter som gjennomgår flere skallskifter årlig og yngre individer med hyppige skallskifter er spesielt sårbare. Det er to kilder til eksponering for stoffene; løst i vann og bundet til organisk materiale.
Effekter av flubenzuroner i vannfasen er undersøkt for tre marine arter, hoppekrepsene Acartia tonsa og Tisbe battaglia og pungreken Mysidopsis baiha , i tillegg til en del ferskvannsarter (Langford mfl. 2011, Macken mfl. 2015). Eksponeringstiden i forsøkene er fra 24 timer til 30 dager. Felles for dem, med ett unntak, er at LC 50 -verdien er høyere enn den høyeste konsentrasjonen på 295 ng/l som har vært målt i vannfraksjonen i forbindelse med bruk av flubenzuroner i akvakultur i Norge (Langford mfl. 2011). Kun en art av vårflue ( Clistoronia magnifica ) hadde LC 50 -verdi under 295 ng/l. Langtidseksponering (7 dager) gav imidlertid NOEC verdier for T. battaglia på 3,2 ng/l for teflubenzuron og tilsvarende 10,0 ng/l for diflubenzuron (Macken mfl. 2015). Disse verdiene er betydelig lavere enn mange av verdier målt i vannfasen men vi vet lite om den reelle eksponeringen av non-target organismer for flubenzuroner løst i vann.
Effekt av flubenzuroner bundet til organisk materiale inkluderer ulike reservoarer for stoffene som fekalier, fôrspill og levende organismer som for eksempel børstemark. Børstemark ( Capitella capitata, Nereis sp.) kan overleve med høye konsentrasjoner av flubenzuroner over lang tid (Mendez 2005, 2006, NIVA 1998, Samuelsen mfl. 2015). Høyeste konsentrasjon i undersøkelsen til Samuelsen mfl. (2015) var 8353 ng/g. I tillegg til dødelighet kan eksponering for et stoff gi andre effekter. Nylige forsøk gjort ved Havforskningsinstituttet viste økende dødelighet og reduksjon i vekst når børstemarken Capitella sp. ble f ô ret med konsentrasjoner av teflubenzuron på 1, 2 og 4 g teflubenzuron per kilo f ô r (Fang, upubliserte data). Samuelsen mfl. (2014) studerte i laboratorieforsøk effekten av oralt administrert teflubenzuron på yngel av europeisk hummer med doser tilsvarende konsentrasjoner i fekalier og medisinert fôr. Eksponeringen varte syv påfølgende dager for å simulere utslipp ved medisinering. Dette gav en dødelighet på 30 til 50% i de medisinerte gruppene sammenlignet med kontrollgruppen. I tillegg ble det registrert misdannelser som deformerte klør, forkalkede gangbein, stive antenner og reduserte gjellelokk hos yngel som overlevde skallskifte. Individer med misdannelser vil ha mindre sjanser for å overleve i naturen. Totaleffekten (dødelighet og misdannelser) var i størrelsesorden 50% i alle grupper. Det laveste nivået målt i hummerne som døde var 12 ng/g, dette er lavere enn det som ble funnet i flere arter i feltundersøkelsen (Samuelsen mfl. 2015). På den annen side var det ett individ som overlevde skallskifte med en konsentrasjon på 351 ng/g. Påfølgende studier med langtidseksponering av hummeryngel for teflubenzuron der doser tilsvarende 20, 10, 5, 2 og 0,4% av konsentrasjonen i medisinert fôr, gav også rundt 50% dødelighet ved høyeste dose, mens de to laveste dosene gav mindre enn 10% dødelighet (Samuelsen upubliserte data). Liknende studier er gjort med voksne blomsterreker ( Pandalus montagui ) og strandreker, og resultater viser at begge artene er mer sensitive for teflubenzuron enn hummeryngel (Samuelsen upubliserte data). Noen av de samme typer av misdannelser som ble funnet på hummeryngel ble også registrert på dypvannsreker. Resultatene tilsier at NOEC (konsentrasjon for ingen observerte effekter) kan estimeres på alle disse tre artene, når de endelige analysene er ferdigstilt . Pelagiske larver av dypvannsreke ( Pandalus borealis ) ble eksponert for små fôrpartikler som inneholdt 0,7 g/kg diflubenzuron (Bechmann mfl. 2018). Partiklene fordelte seg i vann med en snitt konsentrasjon på 1,5 µg/L. Etter 14 dagers eksponering og 20 dager observasjon førte dette til en reduksjon i overlevelsen på 50% sammenlignet med kontroll. En analyse av stadium I larver på dag 8 viste en konsentrasjon på 180 ng/g våtvekt av diflubenzuron. Høyest dødelighet av rekelarvene var observert fra dag 8 til dag 16. Diflubenzuron påvirket reproduksjonen til amfipoden Corophium volutator og NOEC-verdien ble estimert til å være mellom 10 og 100 µg/kg sediment ( NIVA 1998 ).
5.5.2.2 - Emamektin
Emamektin-benzoat, heretter bare kalt emamektin, tilhører stoffgruppen avermektiner og tilsettes fôret til fisken. Det er beskrevet at emamektin påvirker spenningspotensialet i nerveceller (hyperpolarisering) som resulterer først i paralyse og så død, men den eksakte virkningsmekanismen som dreper lakselus er ikke helt klarlagt. Emamektin selges som SLICE i Norge og anbefalt dosering er 50 µg/kg laks/ørret daglig i 7 dager. Det er anbefalt begrensninger i behandlingshyppigheten pga. den lange halveringstiden. Det bør maks. utføres tre behandlinger i løpet av 12 måneder, og maks. fem behandlinger i løpet av en 2 års produksjonssyklus.
5.5.2.2.1 - Forbruk
Emamektin ble først brukt i 1999, 3,5 kg totalt (figur 5.7). Det årlige forbruket var under 100 kg fram til rundt 2011, men økte kraftig i perioden 2014 til 2016 (172, 259 og 232 kg). Det har siden blir redusert noe, og forbruket i 2019 var 114 kg.
Figur 5.7. Årlig forbruk av emamektin (i kg) i norsk akvakultur, fra 1990 til 2019 (data fra Folkehelseinstituttet).
5.5.2.2.2 - Spredning og fortynning
Emamektin har høy affinitet til organisk materiale (log Kow 5,0) og lav vannløselighet (550 ug/l) (SEPA 2005). Det kan spres til miljøet via fôrspill eller som fekalier fra fisk som er behandlet. Spredningen bestemmes av dybde, vannstrøm og hvor raskt partiklene synker og hvor lett de går i oppløsning. Medisinerte pellets og intakte fekalier synker raskt, mens små svevepartikler som kan utgjøre om lag 5–10% (Bannister mfl. 2016), kan spres med strømmen over større områder. Opp til 2,7% av partiklene kan nå lengre enn 2 km fra anlegget. Det er også vist at for norske lokaliteter med lave strømhastigheter <5 cm/s vil det meste av det organiske materiale konsentreres rett under og i nærheten av anlegget (Bannister mfl. 2016). For lokaliteter med høye strømhastigheter >10 cm/s vil partiklene spres over et større område med relativt lave nivåer rett under merdene.
5.5.2.2.3 - Nedbrytning
Undersøkelser i Skottland viser at emamektin er persistent i sediment, og halveringstiden ble beregnet til 164-225 dager (SEPA 1999b). Emamektin oppløst i vann kan brytes ned av lys (fotolyse) med en halveringstid på ca. 1,5 – 21 dager, avhengig av blant annet pH og daglengde (Veterinærkatalogen).
5.5.2.2.4 - Tilstedeværelse i miljøet
Vi vet lite om hva lags konsentrasjoner som finnes i sediment under norske forhold. Det er gjort en del undersøkelser i Skottland, hvor én undersøkelse fant høyeste konsentrasjon av emamektin på 95,7 ng/g sediment under anlegget, mens lengre enn 25 m fra anlegget var høyeste målte konsentrasjon 42,0 ng/g (SARF098 2016). I et annet studium, ble et anlegg med laks behandlet med totalt 33 g emamektin, det ble funnet restkonsentrasjoner i sediment fra 25-100 m fra anlegget 4 måneder etter behandling (Telfer mfl. 2006). Høyeste konsentrasjon var 2,73 ng/g sediment, 10 m fra anlegget. I det samme studiet ble det funnet restkonsentrasjoner i blåskjell opp til 100 m fra anlegget én uke etter behandling (<1 ng/g). I et annet studium ble det funnet restkonsentrasjoner på 3,6 ug/kg sediment i prøver tatt opptil 100 m fra anlegget fire måneder etter behandling (182,5 g ble brukt totalt for anlegget) (SEPA 2005). I en annen skotsk undersøkelse, som omfattet nesten 1000 sedimentprøver, var høyeste konsentrasjon 95,7 ng/g sediment i prøver tatt under anlegget mens lengre enn 25 m fra anlegget var høyeste målte konsentrasjon 42,0 ng/g (SARF098 2016). I en feltundersøkelse ble høyeste konsentrasjon i sedimentet målt til 6,6 ng/g, etter én behandling ( Schlering-Plough 1999). Langford mfl. (2014) fant konsentrasjoner på 2.4 ng/g to måneder etter behandling for et anlegg på Vestlandet i Norge, og 6,5 ng/g ved et annet anlegg. Åtte oppdrettsanlegg på Shetland ble også undersøkt for restkonsentrasjoner av emamektin i sedimentet (Bloodworth mfl. 2019, SEPA 2018). Konsentrasjonene av emamektin var høyest i prøver tatt like ved anleggene, 25,2 ng/g. 17% av prøvene hadde høyere konsentrasjon enn standard «near-field EQS» på 7,6 ng/g våtvekt sediment, mens 100 m fra anleggene var kun 7% av prøvene høyere enn «far-field EQS» 0,76 ng/g sediment (SEPA 2018). Det bør gjøres en vurdering om Norge skal foreslå liknende EQS verdier som Skottland.
5.5.2.2.5 - Endringer i artssammensetning
Gjennom året vil det naturlig være endringer i artssammensetningen, både i diversitet, mengde og tetthet. For eksempel er det arter som kun finnes i enkelte geografiske områder, og enkelte livsstadier som kun forekommer deler av året. Det kan derfor være utfordrende å skille endringer som følge av eksponering for legemidler og de naturlige endringene. I Skottland ble det i perioden september 1999 til august 2004 undersøkt mulige endringer i flora og fauna ved fire oppdrettslokaliteter som følge av bruk av flere lusemidler (før og etter), deriblant også emamektin (SAMS 2005). De fant ingen effekter på dyreplankton, hverken på artsnivå eller samfunnsnivå.
Telfer mfl. (2006) undersøkte fauna før, under og opp til 1 år (september 1997 til september 1998) etter behandling med emamektin på et skotsk oppdrettsanlegg. De vanligste artene rundt anlegget var kongsnegl (Buccinum undatum), eremittkreps (Pagurus spp.), strandkrabbe (Carcinus maenas), svømmekrabbe (Liocarcinus depurator), alle artene er alt-spisende og kunne dermed teoretisk spise fôrrester fra oppdrettsanlegget. De fant at, til tross for at antall av hver art og hvilke arter som ble funnet varierte gjennom året, var det ingen forskjell mellom før- og etter-undersøkelsen. De høyeste restkonsentrasjonene ble funnet i kongsnegl (2,2-5 µg/kg) en uke etter behandling og i eremittkreps (2,5 µg/kg) en måned etter behandling. Det ble ikke funnet målbare restkonsentrasjoner i hverken krepsdyr eller skjell. Artsdiversitet under anlegget var lavere sammenlignet med kontrollområdet like etter behandling, men etter 12 måneder var artsdiversiteten tilsvarende som før behandling. Det er foreløpig ikke publisert norske studier som viser funn av endringer i artssammensetning eller endringer i en arts mengde som følge av eksponering for emamektin , men gitt at forbruket er så høyt bør dette gjennomføres.
5.5.2.2.6 - Artenes følsomhet
Emamektin påvirker nervesystemet og har dermed en direkte effekt på organismen. Det er gjort noen effektstudier med emamektin oppløst i vann, men disse studiene er mindre relevante siden stoffet vil være bundet til organiske partikler. Våre vurderinger er derfor basert på studier hvor non-target arter kan få i seg emamektin via fôrrester og fekalier. Hos voksen amerikansk hummer førte oral administrering av emamektin til fremskyndet skallskifte (Waddy mfl. 2007). Voksen amerikansk hummer var lite følsom, LC 50 >644 µg/g ved eksponering for emamektin gjennom fôret over 7-10 dager (Burridge mfl. 2004). Lignende ble også funnet for voksen sjøkreps, LC 50 >68,2 µg/g (SARF098 2016). Et forsøk over 10 dager, der emamektin var anriket til sediment, gav LC 50 -verdier på henholdsvis 153 ng/g og 1368 ng/g for tangloppe ( Corophium volutator ) og børstemarken Hediste diversicolor (Major mfl. 2008). Lignende verdier ble også rapportert fra Skottland, LC 50 på 193 og 111 ng/g sediment (våtvekt) for mudderreke ( Crangon volutator ) og fjæremark ( Arenicola marina ) (SEPA 1999b). NOEC verdier for sjøkreps eksponert for anriket sediment i 192 timer oppgis til 68,2 μg/g, og 69,3 μg/g for hestereke ( Crangon crangon ). Disse konsentrasjonene er betydelig høyere enn innholdet i kommersielt medisinfôr (10 μg/g) men eksponeringen varte over kun 8 dager (SEPA 1999b). NOEC verdien for sediment anriket med emamektin ble målt til 460 ng/g sediment for børstemarken Capitella capitata (SEPA 1999b). Daoud mfl. (2018) fant LC 50 >250,2 ng/g for hummerlarver (stadium IV-V) når de ble plassert på sediment tilsatt premix av emamektin. Generelt er LC 50 lavere i de eksperimentene hvor bunnlevende dyr eksponeres for sediment (sand) tilsatt emamektin i ren form. Dette gjelder hummeryngel av amerikansk hummer (stadium IV-V), tangloppe, børstemark, mudderreke og fjæremark.
5.6 - Referanser
Aaen SM, Aunsmo A, Horsberg TE. (2014). Impact of hydrogen peroxide on hatching ability of egg strings from salmon lice ( Lepeophtheirus salmonis ) in a field treatment and in a laboratory study with ascending concentrations. Aquaculture 422-423, 167-171.
Andersen PA, Hagen L (2016). Fortynningsstudier – Hydrogenperoksid. Feltstudier. Rapport. Aqua Kompetanse AS, 30 s.
Bannister RJ, Johnsen IA, Hansen PK, Kutti T, Asplin L (2016). Near- and far-field dispersal modelling of organic waste from Atlantic salmon aquaculture in fjord systems. ICES J Mar. Sci. 73 (9), 2408-2419.
Bechmann RK, Lyng E, Westerlund S, Bamber S, Berry M, Arnberg M, Kringstad A, Calosi P, Seear PJ (2018). Early life stages of northern shrimp ( Pandalus borealis ) are sensitive to fish feed containing the anti-parasitic drug diflubenzuron. Aquatic toxicology, 198: 82-91.
Boelhouwers EJ. Joustra KD. Stegman KH (1988). Hydrolysis of 14C labeled diflubenzuron in buffer solutions at pH 5, 7 and 9. UNIROYAL DI-6799. Duphar B.V. The Netherlands, report 56630/08/91.
Brokke KE (2015). Mortality caused by de-licing agents on the non-target organisms chameleon shrimps ( Pranus flexuosus ) and grass prawn ( Palaemon elegans ). Thesis for the degree Master of Science, University of Bergen, Bergen, Norway. 105 s .
Bruno DW & Raynard RS (1994). Studies on the use of hydrogen peroxide as a method for the control of sea lice on Atlantic salmon. Aquaculture International 2, 10-18.
Burridge LE. Van Geest JL (2014). A review of potential environmental risk associated with the use of pesticides to treat Atlantic salmon against infestations of sea lice in Canada. Can. Sci. Adv. Sec. Res Doc 2014/002.
Burridge LE, Hamilton N, Waddy SL, Haya K, Mercer SM, Greenhalgh, R, Tauber R, Radecki SV, Crouch LS, Wislocki PG, Endris RG. (2004). Acute toxicity of emamectin benzoate (SLICE TM) in fish feed to American lobster, Homarus americanus . Aquaculture Research, 35: 713-722.
Crane M, Gross M, Maycock DS, Grant A, Fossum BH (2011). Environmental quality standards for a deltamethrin sea louse treatment in marine finfish aquaculture based on survival time analyses and species sensitivity distributions. Aqua Res. 42, 68-72.
Daoud D, McCarthy A, Dubetz C, Barger DE (2018). The effects of emamectin benzoate or ivermecting spiked sediment on juvenile American lobster ( Homarus americanus ). Ecotoxicology and Environmental Safety, 163: 636-645.
Egidius E, Møster B (1987) Effect of Neguvon and Nuvan Treatment on Crabs ( Cancer pagurus , C. maenas ), Lobster ( Homarus gammarus ) and blue mussel ( Mytilus edulis ). Aquaculture, 60, 165-168.
EMEA (1999a). Committee for veterinary medical products. Diflubenzuron, Summary report (2). European Agency for the Evaluation of Medicinal Products, Veterinary Medicines Evaluation Unit. EMEA/MRL/621/99-Final. 6s.
EMEA (1999b). Committee for veterinary medical products. Teflubenzuron, Summary report (2). European Agency for the Evaluation of Medicinal Products, Veterinary Medicines Evaluation Unit. EMEA/MRL/547/99-Final. 7s.
Ernst W, Doe K; Cook A; Burridge L, Lalonde B, Jackman P, Aube JG, Page F (2014). Dispersion and toxicity to non-target crustaceans of azametiphos and deltamethrin after sea lice treatments on farmed salmon, Salmo salar . Aquaculture 424, 104-112.
Ernst, W, Jackman P, Doe K, Pages F, Julien G, Mackay K & Sutherland T (2001). Dispersion and toxicity to non-target aquatic organisms of pesticides used to treat sea lice on salmon in net pen enclosures. Marine Pollution Bulletin, 42 (6): 433-444.
Ervik A, Thorsen B, Eriksen V, Lunestad BT, Samuelsen OB (1994). Impact of administering antibacterial agents on wild fish and blue mussels Mytilus edulis in the vicinity of fish farms. Dis Aquat Org, 18, 45-51.
Escobar Lux RH (2016). The effects of an anti-sea lice chemotherapeutant, hydrogen peroxide, on mortality, escape response and oxygen consumption on Calanus spp. Masteroppgave (These de master). Peierre et marie Curie University, Paris France. 40 s.
EU (1996) Council Directive 96/23/EC of 29 April 1996 on measures to monitor certain substances and residues thereof in live animals and animal products
EU (2009) Commission Regulation (EU) No 37/2010 of 22 December 2009 on pharmacologically active substances and their classification regarding maximum residue limits in foodstuffs of animal origin
Fagereng M, Haavik S (2015). Monitoring of hydrogen peroxide in surrounding seawater after treatment of farmed fish in netpen. Preliminary report of field experiments in October 2015. Report. University of Bergen. 6 s.
Fagereng M (2016). Bruk av hydrogenperoksid I oppdrettsanlegg; fortynningsstudier og effekter på blomsterreke ( Pandalus motagui ). Masteroppgave i farmasi. Senter for farmasi, Universitetet i Bergen. 104 s.
Fairchild WL, Doe KG, Jackman PM, Arsenault JT, Aubé JG, Losier M, Cook AM (2010). Acute and chronic toxicity of two formulations of the pyrethroid pesticide deltamethrin to an amphipod, sand shrimp and lobster larvae. Can. Tech. Rep. Fish. and Aqua. Sci. 2876, 42 s.
Fang J, Samuelsen OB, Strand Ø, Jansen H (2018). Acute toxic effects of hydrogen peroxide, used for salmon lice treatment, on the survival of polychaetes Capitella sp. And Ophryotrocha spp. Aquaculture Environment Interactions, 10: 363-368.
Gebauer P, Paschke K, Vera C, Toro J E, Pardo M, Urbina M (2017). Lethal and sub-lethal effects of commonly used anti-salmon louse formulations on non-target crab Metacarcinus edwardsii larvae. Chemosphere, 185, 1019-1029.
Grøsvik E, Andersen RE (1997). Investigation of environmental effects in the field. Unpublished report submitted to Apharma, Norway by Rogaland Research. 30 juni 1997. 15 s. ¨
Hannisdal R, Nøstbakken OJ, Lunestad BT, Hove HT, Frøyland L, Madsen L (2018). Monitoring program for pharmaceuticals, illegal substances, and contaminants in farmed fish. ISSN 1893-4536
Hansen BH, Hallmann A, Altin D, Jenssen BM, Ciesielski TM (2017). Acute hydrogen peroxide exposure does not cause oxidative stress in late-copepodite stage of Calanusfinmarchius . J. Toxicol Environ Health, Part A, 80, 820-829.
Haugland BT, Rastrick SPC, Agnalt A-L, Husa V, Kutti T, Samuelsen OB (2019). Mortality and reduced photosynthetic performance in sugar kelp Saccharina latissimi caused by the salmon-lice therapeutant hydrogen peroxide. Aquaculture Environment Interactions, 11: 1-13.
Haya K, Burridge LE, Davies IM, Ervik A. (2005). A review and assessment of environmental risk of chemicals used for the treatment of sea lice infestations of cultured salmon. Environmental effects of marine finfish aquaculture. 1433-6883. Springer-Verlag. Berlin pp. 305-340.
Hektoen H (1995). Miljøvirkninger av havbruk, Sluttrapport til Forskningsrådet 1990-94. Norges Forskningsråd ISBN: 82-12-00558-5. 45 s.
Hjeltnes B, Bornø, G, Jansen, M D, Haukaas,A, Walde, C (red) (2017). Fiskehelserapporten 2016. Veterinærinstituttet rapportserie nr. 4/2017. 124 s.
Ivie GW, Bull DL, Veech JA (1980). Fate of diflubenzuron in water. Journal of Agricultural Food Chemistry 26, 330-337.
Johnson SC, Constible JM, Richard J (1993). Laboratory investigations on the efficacy of hydrogen peroxide against the salmon louse Lepeophtheirus salmonis and its toxicological and histopathological effects on Atlantic salmon Salmo Salar and chinook salmon Oncorhynchus tshawytscha. Disease of Aquatic Organisms. 17: 197-204.
Langford KH, Øxnevad S, Schøyen S, Thomas KV (2011). Environmental screening of veterinary medicines used in aquaculture-diflubenzuron and teflubenzuron. Klima- og forurensningsdirektoratet, Statlig program for forurensningsovervåkning. SPFO-rapport 1086/2011, 46s. Rapporten er tilgjengelig på www.vetinst.no and www.antibiotikaresistens.no
Langford KH, Øxnevad S, Schøyen M Thomas KV (2014). Do antiparasitic medicines used in aquaculture pose a risk to the Norwegian aquatic environment? Environmental Science & Technology 46: 7774-7780.
Langford K, Bæk K, Kringstad A, Rundberget T, Øxnevad S, Thomas KV (2015). Screening of the sea lice medications azamethiphos, deltamethrin and cypermethrin Screening av lusemidlene azametifos, deltametrin og cypermetrin. Miljødirektoratet, Miljøovervåking, M-345. 28 s.
Lyons MC, Wong DKH, Page FH (2014). Degradation of hydrogen peroxide in seawater using the anti-sea louse formulation Interox® Paramove™50. Can. Tech. Rep. Fish. Aquat. Sci. 3080: v + 15p.
Mabury SA, Crosby DG (1996). Fate and disposition of diflubenzuron in rice fields. Environmental Toxicology and Chemistry 15, 11, 1908-1913.
Macken A, Lillicrap A, Langford K (2015). Benzoylurea pesticides used as veterinary medicines in aquaculture: risks and developmental effects on non-target crustaceans. Environmental Toxicology and Chemistry 34 (7), 1533–1542.
Madden S, Callanan K, Roberson N, Moore DC (1991). Determination of acute toxicity (LC50) of CGA18809 to yolk sac and post yolk sac herring larvae ( Clupea harengus L.). Fish. Res. Serv.
Maguire RJ, Carey JH, Hart JH, Tkacz RJ, Lee H-B (1989). Persistence and Fate of Deltamethrin Sprayed on a Pond. J Agrie Food Chem, 37 (4): 1153-1159.
Major DJ, Solan M, Martinez I, Murray L, McMillan H, Paton GI, Killham K (2008). Acute toxicity of some treatments commonly used by the salmonid aquaculture industry to Corophium volutator and Hediste diversicolor : whole sediment bioassay tests. Aquaculture 258, 102-108.
Marsella AM, Jaskolka M, Mabury SA (2000). Aquous solubilities, photolysis rates and partition coefficient of benzoylphenylurea insecticides. Pest Management Science 56, 789-794.
Mendez N (2005). Effects of teflubenzuron on larvae and juveniles of the polychaete Capitella sp. B from Barcelona, Spain. Water, Air and Soil pollution 160, 259-269.
Mendez N (2006.) Effects of teflubenzuron on sediment processing by membres of the Capitella species-complex. Environmental pollution 139, 118-124.
NIVA (1998). Environmental safety evaluation for the use of Lepsidon® vet 0.6g/kg “EWOS” against developing stages of sea lice on fish in aquaculture. Expert report, environmental safety. NIVA Report 3877-98. 45 s.
NORM/NORM-VET (2018). Usage of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Norway. Tromsø/Oslo 2018. ISSN:1502-2307(print)/1890-9965(electronic).
Pahl BC, Opitz HM (1999). The effects of cypermethrin (Excis) and azamethiphos (Salmosan) on lobster Homarus americanus H. Milne Edwards larvae in a laboratory study. Aquaculture Research, 30: 655-665.
Poppe T (1999) redaktør. Fiskehelse og fiskesykdommer. Universitetsforlaget AS.
Refseth GH, Sæther K, Drivdal M, Nøst OA, Augustine S, Camus L, Tassara L, Agnalt AL, Samuelsen OB (2017). Miljørisiko ved bruk av hydrogenperoksid. Økotoksikologiske vurdering og grenseverdi for effekt. Akvaplan-niva AS Rapport 8200 – 1. 55 s.
Robertson NA, Madden S, Moore DC, Davies IM (1992). Repeated short exposure of lobster larvae (Homarus Gammarus L.) to CGA18809. Fisheries Research Services Report No 13/92. mfl. 1992
SAMS (2005). Ecological effects of sea lice medicines in Scottish sea lochs. Final Report 9. Feb. 2005. Scottish Association for Marine Science. 286 s.
Samuelsen OB (2016). Persistence and stability of teflubenzuron and diflubenzuron when associated to organic material in marine sediment. Bulletin of Environmental Contamination and Toxicology, 96, 224-228.
Samuelsen OB, Lunestad BT, Farestveit E, Grefsrud E, Hannisdal R, Holmelid B, Tjensvoll T, Agnalt A-L (2014). Mortality and deformities in European lobster ( Homarus gammarus ) juveniles exposed to the anti-parasitic drug teflubenzuron. Aquatic Toxicology 149, 8-15.
Samuelsen OB, Lunestad BT, Hannisdal R, Bannister R, Olsen S, Tjensvoll T, Farestveit E, Ervik A (2015). Distribution and persistence of the antisea-lice drug teflubenzuron in wild fauna and sediments around a salmon farm, following a standard treatment. Science of the Total Environment 508, 115-121.
SARF098 (2016). Towards understanding of the environmental impact of sea lice medicine-the PAMP Suite. A study commissioned by the Scottish Aquaculture Research Forum (SARF). http://www.sarf.org.uk/
Schering-Plough (1999). Slice. Teknisk monografi. Schering-Plough Animal Health. 40 s.
Selvik A, Hanssen PK, Ervik A, Samuelsen OB (2002). The stability and persistence of Diflubenzuron in marine sediments studied under laboratory conditions and the dispersion to the sediment under a fish farm following medication. Science of the Total Environment 285, 237-245.
SEPA (1999a). Calcide (Teflubenzuron) - Authorisation for use as an in-feed sea lice treatment in marine cage salmon farms. Risk assessment, EQS and recommendations. Scottish Environmental Protection Agency, Policy no 29. 15 s.
SEPA (1999b). Emamectin Benzoate. An environmental Risk Assessment. Scottish Environmental Protection Agency. 23 s.
SEPA (2005). Ecological effects of sea lice medicines in Scottish sea lochs. Final report 9 February 2005. 286 s.
SEPA (2018). Evaluation of a new seabed monitoring approach to investigate the impacts of marine cage fish farms. Scottish Environmental Protection Agency, Report 39 s. http://contactscotland-bsl.org/
Telfer TC, Baird DJ, McHenery JG, Stone J, Sutherland I, Wislocki P (2006). Environmental effects of the anti-sea lice (Copepoda: Caligidae) therapeutant emamectin benzoate under commercial use conditions in the marine environment. Aquaculture, 260 (1-4): 163-180.
Thomassen, JJVI. (1993) A new method for control of salmon lice. In: Fish Farming Technology (eds H Reinertsen, LA Dahle, L Jørgensen and K Tvinnereim) Balkema: Rotterdam. pp. 233-236.
Tomlin C (red) (1994). The pesticide manual: A world compendium. Incorporating the agrochemicals Handbook. 10 th ed. British Crop Protection Council and Royal Society and chemistry. Thornton health. UK.
Treves-Brown KM (2000). Applied Fish Pharmacology. Kluwer Academic Publisher, London
Urbina MA, Cumillaf JP, Paschke K, Gebauer P (2019). Effects of pharmaceuticals used to treat salmon lice on non-target species: Evidence from a systematic review. Science of The Total Environment, 649: 1124-1136.
Van Geest JL, Burridge LE, Kidd KA (2014a). Toxicity of two pyrethroids-based anti-sea lice pesticides, AlphaMax and Excis to a marine amphipod in aqueous and sediment exposure. Aquaculture 434, 233-240.
Van Geest JL, Burridge LE, Kidd KA (2014b). The toxicity of the anti sea-lice pesticide AlphaMax to the polychaete worm Nereis virens . Aquaculture 430, 98-106.
Waddy SL, Merritt VA, Hamilton-Bibson MN, Aiken DE, Burridge LE (2007). Relationship between doses of emamectin benzoate and moulting respons of ovigerous American lobsters ( Homarus americanus ). Ecotoxicology and Environmental Safety 67, 95-99.
Willis, KJ, Gillibrand PA, Cromey CJ, Black KD (2005). Sea lice treatments on salmon farms have no adverse effects on zooplankton communities: a case study. Marine Pollution Bulleting, 50: 806-816.
Wong GTF, Dunstan WM, Kim D-B (2003). The decomposition of hydrogen peroxide by marine phytoplankton. La décomposition du peroxyde d’hydrogène par le phytoplancton marin. Oceanologica Acta 26: 191–198.
6 - Miljøeffekter ved bruk av villfanget leppefisk i fiskeoppdrett
Grønngylt hann (venstre) og hunn (høyre). Foto Erlend A. Lorentzen/Havforskningsinstituttet
6.1 - Innledning
Bruk av rensefisk som avlusningsmetode mot lakselus i norsk havbruk, har økt jevnt de siste årene. I 2019 passerte for første gang totalt utsett av rensefisk 60 millioner individer til en samlet verdi på over 1,3 mrd NOK (https://www.fiskeridir.no/Akvakultur/Tall-og-analyse/Akvakulturstatistikk-tidsserier/Rensefisk). Både leppefisk og rognkjeks benyttes som rensefisk og metoden er regnet som et miljøvennlig alternativ til bruk av legemidler. Rensefisk påfører også oppdrettsfisken mindre stress enn avlusningsmetoder som krever håndtering. Rensefisk som settes ut i norske oppdrettsanlegg er definert som akvakulturdyr, og dermed underlagt det samme regelverket som andre oppdrettsorganismer. Anlegg med rensefisk er dermed flerartskulturer (polykulturer). All rognkjeks som brukes som rensefisk kommer fra oppdrett. Av leppefisk er noe av berggylten fra oppdrett, men storparten av forbruket kommer fra villfanget leppefisk, og de vanligste artene som høstes er grønngylt, bergnebb, berggylt og gressgylt.
Flere problemstillinger har reist seg i kjølvannet av bruken av rensefisk. De berører både fiskeriet av leppefisk, den storstilte flyttingen mellom områder, smittespredning og genetiske endringer. Disse temaene vil bli omtalt i denne kunnskapsstatusen som også er grunnlaget for risikovurdering av miljøeffekter ved bruk av villfanget leppefisk i oppdrett.
Fisket etter leppefisk er et blandingsfiske hvor fangstene består av ulike arter salgbar leppefisk, samt en bifangst av undermåls leppefisk og andre arter. Fisket etter leppefisk er regulert ved redskapsbegrensning, seleksjonsinnretninger i redskap som brukes, minstemål, båtkvoter og fangstperiode og håndtering av bifangst. Informasjon om reguleringen finnes på http://www.fiskeridir.no/Yrkesfiske/Tema/Fiske-etter-leppefisk. Det fiskes i hovedsak med spesialbygde teiner, mens et mindretall av fiskerne benytter også ruser. Selv om det forekommer variasjoner, gir teiner generelt sett lavere bifangst av andre arter sammenliknet med ruser (Halvorsen mfl., 2017b, 2017a). Ruser er i mindre grad selektive og har et økt innslag av arter som torsk, krabber og ål i forhold til teiner. Det må søkes om tillatelse for å bruke ruser i fisket, og i 2020 er det bare yrkesfiskere med merkeregistrerte fartøy som får benytte disse. Fra 2021 vil det være forbudt å bruke ruser i fiske etter leppefisk. Fra 2015 måtte både teiner og ruser ha fluktåpninger for å selektere ut den minste fisken. Dessuten ble det påbud om inngangssperre for å hindre oter, stor fisk og hummer å gå inn i fiskeredskapene. Fra 2021 må alle redskap som benyttes i fisket etter leppefisk ha sirkulære åpninger i kalven med maks diameter på 6 cm. Dette vil redusere fangst av stor berggylt (> 28 cm), samt antageligvis også bifangst av andre større arter, som torsk, krabbe og hummer (Halvorsen mfl., 2019).
I de fleste områder foregår fisket etter leppefisk relativt lokalt. Fisken holdes i en eller flere oppbevaringstanker om bord, og leveres direkte til fiskeoppdrettsanleggene, eller har en kort mellomlagring før levering. Er det større avstander mellom fiskeområder og oppdrett (som er tilfelle fra Sørlandet og Sverige), samles leppefisken opp i tanker eller samlemerder og fraktes videre med tankbiler.
Det er viktig at fangstmetodene er skånsomme og at fisken er lytefri ved levering. Hvis fisken står lenge i redskapen vil den lett få skader, og det er derfor innført regler for ståtid (maks ett døgn utenom helligdager). Det er ikke funnet forskjeller i kvalitet og overlevelse av fisk som er fisket med henholdsvis ruser eller teiner (Skiftesvik mfl., 2014b). Skadd fisk og bifangst blir sortert ut. Hvis fisken sorteres mens fiskerne forflytter seg fra en lokalitet til en annen og blir sluppet ut i åpen sjø, vil den trolig ha redusert overlevelse. Reguleringene har de siste årene vært at «All bifangst skal straks tilbake i sjøen på ein slik måte at den ikkje vert skadd og at den finn tilbake til sitt naturlige miljø. Denne ordninga gjeld også leppefisk fanga i strid med gjeldande minstemål eller som av andre grunnar må setjast tilbake i sjøen.» og at «Det er ikkje tillate å behalde bifangst i teiner eller ruser som er sett ut til fangst av leppefisk. Dette gjelder også bifangst av krabbe.» https://fiskeridir.no/Yrkesfiske/Tema/Leppefisk/Leppefisk-reguleringa-2020 Mange fiskere har montert rør for å slippe ut fisken under vannflaten. Når den slippes ut igjen på fangststedet (nært land) øker dette muligheten for at den kommer trygt til bunns og i skjul. Forhold rundt fangst, transport og bruk av rensefisk er beskrevet i veilederne, på: http://lusedata.no/for-naeringen/veiledere-leppefisk/ .
Uttak av leppefisk har variert mellom år. I 2017 var det registrerte uttaket i underkant av 28 millioner ville leppefisk på landsbasis. Dette var nesten 10 millioner mer enn anbefalt uttak. Fisket ble kvoteregulert fra 2018, og uttaket av leppefisk har nå stabilisert seg rundt anbefalt uttak. I 2019 var uttaket av leppefisk for de ulike geografiske områdene (avrundet til nærmeste 100 000): 4,1 millioner for Sørlandet (fra grensa mot Sverige til Varnes fyr på Lista), 11,5 millioner for Vestlandet (fra og med Varnes fyr på Lista til 62°N), og 3,4 millioner nord for 62°N. Totalt 19 millioner leppefisk på landsbasis (oppdatert 27.04.2019). Det fiskes mest av bergnebb (45%) og grønngylt (42%) (https://www.fiskeridir.no/Yrkesfiske/Tall-og-analyse/Fangst-og-kvoter/Fangst-av-leppefisk).
Fisketrykket varierer betydelig mellom ulike geografiske områder, samtidig som det også kan være store lokale og regionale variasjoner i naturlige forekomsten av de ulike leppefiskartene (Skiftesvik mfl., 2015, Halvorsen mfl., 2016). Dette skyldes at leppefisk er svært stedbundne og har spesifikke habitatpreferanser, slik at tettheten påvirkes i stor grad av romlig variasjon i miljøbetingelser. Fiskeriet vil ha ulik påvirkning i ulike områder, siden det er påvist høy geografisk variasjon i vekst og kjønnsmodning (Halvorsen mfl., 2016, Olsen mfl., 2018).
Fisket etter leppefisk vil påvirker bestandene til de ulike artene forskjellig. Fangbarheten forventes å være artsspesifikk, noe som i stor grad avhenger av artenes habitat-overlapp med fiskeriet, som for eksempel dybde. Grønngylt og berggylt har høyest forekomst på 0-6 meter, som også er måldybde for dette fiskeriet (Halvorsen mfl., 2020). Bergnebb, grasgylt og Rødnebb/blåstål finnes også like stor eller økende tetthet ned mot 20 meters dyp. Sannsynligheten for at disse artene kommer i kontakt med teiner og blir fanget er derfor lavere. Merkeforsøk viser at grønngylt og bergnebb har høyest fangbarhet i teiner, mens den er tilsynelatende lav for berggylt (Halvorsen mfl., 2020, Ruud 2020). De underliggende årsakene for disse forskjellene er ukjente, men antas at de skyldes ulik grad av tiltrekking til agn, samt motivasjon og evne til å ta seg inn og ut av teinene (Ruud 2020).
Leppefiskartene med kortest generasjonstid vil kunne restitueres raskere etter et overfiske enn arter med lengre generasjonstid. I tillegg er de artsspesifikke minstemålene i ulik grad er tilpasset størrelse ved kjønnsmodning. Berggylt står her i særklasse, med et minstemål på 14 cm, mens den kjønnsmodnes som hunn rundt 22 cm, og skifter kjønn til hann ved 34-40 cm (Muncaster mfl., 2013). Med andre ord, et høyt uttak av bestanden over 14 cm kan ha en sterk påvirkning på kjønnsfordeling og produktiviteten hos berggylt. Generasjonstiden er lengst for berggylt og bergnebb, og vi forventer derfor at disse artene vil bli mest negativt påvirket av et intensivt fiske, og da spesielt berggylt da den, som før nevnt, ikke er kjønnsmoden ved minstemålet som er innført. Bergnebb kjønnsmodner når den er 6-7 cm, godt før oppnådd fangststørrelse, men det er små eggmengder det er snakk om her da fisken er liten. Grønngylt har kort generasjonstid på Sørlandet, og vil derfor ha potensial til å hente seg inn raskere enn bergnebb og berggylt.
Fisket skjer oftest ved at det fiskes intensivt på lokalitet etter lokalitet, slik at en lokalitet «fiskes ned» for salgbar fisk, hvorpå fiskeren forflytter seg til den neste lokalitet, som så «fiskes ned», osv. Selv om fangst per redskapsenhet viser ingen eller moderat reduksjon, vil det kunne være en dramatisk effekt på små, lokale og gjerne geografisk isolerte bestander. På bakgrunn av både referansefiske, våre egne undersøkelser og innkommet informasjon ser vi en fare for at det intensive fisket vil kunne føre en endring i arts-, størrelses - og kjønnsfordeling i leppefisksamfunnene langs kysten (Darwall mfl., 1992, Skiftesvik mfl., 2014a, Halvorsen 2017b).
Å gi en helhetlig vurdering av graden fiskeripåvirkning på de fem artene leppefisk over større områder er derfor utfordrende. Merkestudier gjennomført på Vestlandet har vist at fiskeriet tar ut minst 40 % av grønngyltbestanden i løpet av to måneders fiske (Halvorsen mfl., 2017c). På Skagerrakkysten er det påvist høyere tetthet av bergnebb og grønngylt i verneområder sammenliknet med områder åpent for fiske (Halvorsen mfl., 2017b). Disse enkeltstudiene viser at fiskeriet kan ha en tydelig påvirkning på leppefiskbestandene, men det er fortsatt store kunnskapshull på fiskedødelighet i andre områder og særlig for berggylt. I en nylig avlagt masteroppgave ble det ikke funnet noen tydelige forskjeller i fangstrater eller størrelsesfordeling for berggylt, grønngylt og bergnebbb mellom bevaringsområder i Tvedestrand og nærliggende områder åpne for fiske etter leppefisk i perioden 2010-2019 (Reamon 2020). Det er ukjent hvor hardt disse områdene har blitt fisket i denne perioden da fiskere ikke har vært pålagt å rapportere hvor de fisker. Påbud om AIS-system i dette fisket vil gi verdifull informasjon om fordeling av fisketrykk i tid og rom, og gi mulighet for å bedre studier av generelle effekter av fiskeriet på bestandene i de kommende årene.
Strandnotundersøkelsen for Skagerrakkysten har registrert antall leppefisk på artsnivå siden 1989 (Fig 6.1). Undersøkelsen gir en god indeks på rekruttering, da det først og fremst er 0-gruppe leppefisk som fanges. Lav indeks på høsten tilsier at det er en svak årsklasse som kommer inn i fiskeriet påfølgende år. Strandnotundersøkelsen dekker kun Skagerrak, men siden fiskeriet utgjør en relativ stor del av totalfangst (22 % av den nasjonale kvoten kan tas her), så kan serien gi en indikasjon om i hvilken grad et relativt intensivt fiskeri har påvirket bestandsutviklingen. Det er ingen tydelig negativ trend for berggylt, bergnebb og grønngylt (målartene) for Agder, som er regionen som er mest intensivt fisket (~ 80 % av fangsten på Sørlandet tas i Agder). Øst for Agder har berggylt og bergnebb hatt en tydelig negativ utvikling siden 2010, men siden fiskepresset er lavere her er det sannsynlig at andre årsaker enn fiskeriet etter leppefisk spiller inn. Dette underbygges av trenden til grasgylt øst for Agder, som ikke er en målart i disse områdene, har hatt en tilsvarende negativ utvikling som berggylt og bergnebb.
Figur 6.1 Gjennomsnittlig antall leppefisk i strandnotundersøkelsene Skagerrak 1989-2018, delt inn i underområder. Stiplet linje indikerer middelverdi for hele perioden i alle områdene sett under ett.
6.2.2 - Kunnskapsstyrke
Kunnskapen om leppefiskbestandene er mangelfull når det kommer til forståelse av hvilke mekanismer som regulerer rekruttering og variasjoner i bestandene (f.eks sommer- og vintertemperatur, tetthetseffekter), og i hvilken grad fiskeriet har en regulerende innvirkning på bestandsstørrelsene hos de ulike artene. Pågående forskning ved Havforskningsinstituttets forskningsstasjon på Austevoll undersøker dette, og resultater er ventet å bli presentert i 2021 og 2022. Datagrunnlaget er for bestandsutviklingen er god på Sørlandet gjennom strandnotundersøkelsen og årlige rusetokt. Det er tilsynelatende ikke noen klar nedgang i bestandene etter fisket ble intensivert fra 2010. For resten av landet er det samlet inn fangst-per-enhet-innsats data fra referansefiskere siden 2011. Det har vært utfordrende å konkludere med bestandsendringer basert på disse dataene, grunnet stor variasjon i fangstrater innad og mellom år, og mellom fiskere. Det er imidlertid registrert en urovekkende nedgang i fangstrater av berggylt og bergnebb i noen områder frem til 2015. Nedgangen, særlig på Vestlandet, samtidig som innsatsen i fisket økte, førte til at Havforskningsinstituttet for 2016 anbefalte en «frysing» av leppefiskuttaket på Sørlandet og nord for Stad på samme nivå som 2015 uttaket, og en reduksjon på 15-20% på Vestlandet. Med bakgrunn i dette innførte Fiskeridirektoratet kvoter fra og med 2016. Tanken er så å bruke fangstrater fra tokt og fiskeri for å se om disse øker eller minker ved gjeldende kvotenivå, og da ta stilling til om kvotenivået bør/kan justeres opp eller ned i samsvar med dette. Fra 2019 ble det gjennomført endringer i denne datainnsamlingen som vil redusere variasjon og feilkilder. Forsøksfiske på Austevoll tyder på relativt robuste bestander på tross av høyt fiskepress i dette området (Kunnskapstøtte til Fiskdir 2019, Halvorsen mfl., 2020).
Det er også manglende kunnskap om dynamikken i rekrutteringen og restituering av leppefiskbestandene. Spesielt er det behov for å undersøke konsekvensene av selektivt uttak av artene med kjønnsdimorfisme, hvor hannene står for yngelpleie og større hanner bygger større og flere reir (Kindsvater mfl. 2020). Undersøkelser av dette vil være vil være viktig for å skaffe mer kunnskap om hvilke konsekvenser fisket etter leppefisk vil ha både på kort og lengre sikt.
6.2.3 - Bekymringsmeldinger
Observasjoner fra både forskere, dykkere, fiskere, fritidsfiskere, hytteeiere og fastboende gjør at det til tider kommer inn bekymringsmeldinger. Det rapporteres om at leppefisken i mange områder forsvinner. Mange av bekymringsmeldingene går på at berggylt er forsvunnet fra områder der de har vært vanlige tidligere. En del av fiskerne rapporterer også om svært intensiv fisking som resulterer i at de må forflytte seg til stadig nye områder for å få fisk. I tillegg kommer det en del bekymrings-meldinger om bruk og «svartsalg» av hummer fra fisket etter leppefisk.
6.2.4 - Sortering/gjenutsetting av leppefisk og fiskeripåvirkning på andre arter
Arter kan påvirkes indirekte (negativt eller positivt) ved en endring i artssammensetning som en følge av uttak fra bestandene. Det er en varierende grad av bifangst i fisket, som også varierer med sesong og område. Regelverket er tydelig på at all bifangst (undermåls leppefisk og andre arter) skal gjenutsettes på grunt vann. Havforskningsinstituttet har ikke kjennskap til at kontrollmyndighetene har avdekket systematiske brudd på regelverket rundt gjenutsetting eller rapporter om ulovlig beholdt bifangst. De fleste bifangstarter klarer seg bra dersom de settes ut igjen der de er fisket, slik som krabbe, hummer, ål, torsk og undermåls leppefisk, mens spesielt sypike som mister mye fiskeskjell under fangst, og noe sei og lyr kan ha økt dødelighet. Vi antar at skadeomfanget på sårbare arter vil øke i forhold til hvor mange ganger de fiskes og settes ut igjen. Ved et intensivt fiske etter leppefisk er det derfor fare for uønskede effekter på andre arter som lever i de samme biotopene som leppefisken. Mange fiskere har montert rør for å slippe ut fisken under vannflaten. Når den slippes ut igjen på fangststedet (nært land) øker dette muligheten for at den kommer trygt til bunns og i skjul. Det rapporteres om at dette ikke alltid respekteres.
Havforskningsinstituttet har siden 2011 hatt avtaler med et nettverk av 10-15 leppefisk-fiskere langs norskekysten mellom Hvaler og Flatanger. Disse fiskerne rapporterer både beholdt og utsatt leppefisk, samt bifangst av andre arter. Basert på deres rapporter etter at de tok i bruk inngangsbegrensning og fluktåpninger i henhold til dagens regelverk, viser vi i figur 6.2 hvor stor andel av leppefiskartene som settes ut igjen på grunn av størrelse og/eller liten kjøperinteresse, og hvilke andre arter som dominerer bifangsten. Hvor stort det totale omfanget kan være hvis man skalerer gjennomsnittet for referansefiskerne opp til alle båter som er registrert som leppefisk fiskere er vist i tabell 1 og 2 (Appendiks).
Figur 6.2 viser at i perioden 2015-2018 ble nesten all fanget berggylt beholdt (bare 14% satt ut). For bergnebb og grønngylt beholdes omtrent halvparten, mens lite av grasgylt og rødnebb/blåstål blir beholdt. Utsatt leppefisk som prosent av beholdt leppefisk var om lag 26%, 127%, 121%, 430% og 179% for hhv berggylt, bergnebb, grønngylt, grasgylt og rødnebb/blåstål.
Figur 6.3 viser fangst av alle arter/artsgrupper rapportert av referansefiskerne i 2015-2018 for henholdsvis teiner (N=1115 fangstrapporter) og ruser (N=229 fangstrapporter). Som prosent av beholdt antall leppefisk utgjorde bifangsten av torsk, lyr, ål, hummer og taskekrabbe hhv 2,8, 1,4, 3,5, 0,4 og 16,8% i antall. Som prosent av total fangst av både beholdt og utsatt fangst av alle arter utgjorde bifangsten av torsk, lyr, ål, hummer og taskekrabbe hhv 1,2, 0,5, 1,3, 0,2 og 2,6 % i antall (Figur 6.3).
Figur 6.2 Andeler av leppefisk fisket med teiner satt ut igjen på grunn av størrelse eller liten kjøperinteresse. Basert på jevnlig rapportering fra instituttets referansefiskere, og presentert som gjennomsnittlig andel utsatt inkl. standardfeil over perioden 2015-2018 (N=1115 fangstrapporter).
Figur 6.3 Relativ artssammensetning av all fangst (i antall) i teine- (N=1115 fangstrapporter) og ruse-fisket (N=229 fangstrapporter) etter leppefisk, basert på jevnlig rapportering fra instituttets referansefiskere i perioden 2015-2018.
I noen områder (varierer mye) er det et betydelig innslag av hummer i leppefisk fisket (Appendiks tabellene 1-2). Norsk sluttseddelstatistikk for 2018 viser en total norsk rapportert fangst av hummer på ca. 50 tonn. Dersom nedenfor bifangsttall per fisker skaleres opp til alle registrerte leppefisk fiskere, så gir det en bifangst av hummer på i størrelsesorden 20 tonn om man regner med en gjennomsnitts-vekt på 500 g per bifanget hummer. Fiskerne rapporterer at disse settes ut igjen, og da kan det også hende at samme hummer blir fanget igjen flere ganger. I Havforskningsinstituttets omfattende forskningsfiske i Austevoll og Flødevigen med ruser og teiner fanges det av og til hummer, og per 2020 er ikke registrert skadet eller død hummer i disse studiene. Havforskningsinstituttet har også omfattende fangst-merke-gjenfangst tokt på hummer i Skagerrak, og samlet tilsier disse erfaringene at hummer ikke tar skade av å bli fanget og sluppet ut igjen (Fernandes-Chacon mfl. 2020). Ved fangst av flere hummere i samme hummerteine er det rapportert om klo-tap og død hummer, mest sannsynlig som følge av aggressive interaksjoner i redskapen (Sørdalen mfl. 2020). Det er betydelig lavere fangstrater av hummer i leppefiskredskap sammenliknet med hummerteiner, og det er svært sjelden blitt fanget mer en hummer per teine/ruse i Havforskningsinstituttets prøvefiske etter leppefisk.
Zimmermann mfl. (2020) beregnet den totale bifangsten av taskekrabbe i leppefisk fisket til gjennomsnittlig 235 krabber per fartøy per år for fartøy som fisker med ruser, mens for fartøy som fisker med teiner er gjennomsnittet 2911 krabber per år. Utkastraten av taskekrabbe er høy og ligger i gjennomsnitt på 72% over alle år og redskapstyper (varierer mellom 44 og 100%), men vi kan anta at de fleste krabber som gjenutsettes overlever. Vi antar derfor at gjennomsnittlig mer enn en fjerdedel av krabbene som tas som bifangst, landes og brukes til agn. Ekstrapolert til hele leppefiskkvoten kan dette bety at omtrent 150 000 krabber (rundt 60 tonn) brukes til agn per år, men en stor usikkerhet er knyttet til dette estimatet.
Under hele referanseperioden 2015-2018, dvs. etter at inngangssperre ble innført, er det kun rapportert 5 skarver og 1 oter som fanget av referansefiskerne. Ingen i teiner, bare i ruser.
En stor del av bifangsten er undermåls leppefisk, som i hovedsak slippes fri (Halvorsen mfl., 2016b). I 2015 ble det påbudt med kryss eller not i ytterste kalv på ruser og maksstørrelse på inngangskalv i teiner brukt til fiske etter leppefisk. Dette har redusert bifangsten av oter, hummer, krabbe og stor fisk. Det er også påbud om seleksjonsinnretning (12 mm spalter) for å redusere fangsten av undermåls fisk. Dette har vist seg effektivt i å redusere innslaget av undermåls bergnebb og delvis undermåls grønngylt (Jørgensen mfl., 2017). Størrelsene på åpningen er tilpasset med en målsetning om at undermåls bergnebb skal sorteres ut på fiskedypet. Høyere minstemål og tykkere kroppsfasong hos grønngylt og berggylt betyr at en større andel av undermåls fisk av disse artene ikke kan rømme ut av fluktåpningene. Fluktåpningene er mer effektive i teiner enn i ruser (Halvorsen mfl., 2017a, Jørgensen mfl., 2017).
Overholdes regelverket vil overlevelsen til både fangst og bifangstarter være høy grunnet kort ståtid og ved at fisket kun foregår på grunt vann. Derfor vurderes dette fiskeriet å ha en begrenset påvirkning utover målartene. Usikkerheten rundt bifangst er først og fremst knyttet til om dette pålegget overholdes, noe som i liten grad er kjent. Hvis det forekommer avvik ved at bifangst beholdes eller settes ut på dypere vann, så er det derimot sannsynlig at fisket i en eller annen grad påvirker bestandene av bifangstarter.
6.2.5 - Indirekte økosystemeffekter
Leppefisk har en sentral rolle i kystøkosystemet. De er opportunistiske beitere på en rekke bunnlevende og fastsittende dyr som en rekke arter virvelløse dyr, blant annet krepsdyr, snegler, muslinger, børstemark (Alvsvåg, 1993, Sayer mfl., 1995; Deady and Fives, 1995). Videre er de byttedyr for større fiskearter og sjøfugl (Steven 1933, Nedreaas mfl., 2008, Olsen mfl., 2018). Leppefisk er også føde for større fisk, som kysttorsk, lyr, lange og ål, og i noen områder sjøfugl (skarv). Hvis lokale bestander av leppefisk fiskes ned, vil dette kunne resultere i et redusert fødetilbud for andre arter (Bergström et al 2016).
En nedfisking av lokale leppefiskbestander vil kunne ha en effekt på mengde og tetthet av disse organismene og således på bunnfauna og flora (begroing). En studie fra Sverige har påpekt at et fiske av leppefisk kan i så måte ha en positiv effekt på tareskogen, ved at en for høy tetthet av leppefisk beiter ned snegler og krepsdyr som igjen beiter på trådalger (Östman mfl., 2016).
Noe motstridende funn ble rapportert i et eksperimentelt studie nylig publisert, som viste at høy tetthet av bergnebb kan ha en positiv effekt på flerårlige makroalger og forfatterne advarer mot at en reduserte bestander av leppefisk kan påvirke bunnsamfunn i negativ retning dominert av trådformede grønnalger (Kraufvelin mfl., 2020). Samme studiet viser at blåskjell er en viktig del av dietten til leppefisk - og det er blitt fremsatt en hypotese om at redusert forekomst av blåskjell kan ha en sammenheng med økt tetthet av leppefisk. De økologiske interaksjonene er likevel for lite undersøkt til at man kan si noe håndfast om økologiske effekter av uttak av leppefisk, spesielt siden fødevalg og habitatvalg er ulikt både mellom arter og størrelsesklasser. Det er ingen kjente studier av effekter av fiskeriet på økosystemet som helhet, enten som en direkte påvirkning gjennom bifangst, eller indirekte som en følge av endret artssammensetning, størrelsesfordeling og tetthet av leppefisk. Det er derfor et klart behov for målrettet forskning for å bedre forstå de økologiske sammenhengene mellom leppefisk og andre arter, og hvordan disse påvirkes av endringer i arts og størrelsessammensetning av leppefisk som en følge av fiskeriet.
6.3 - Biologi, velferd og svinn
Leppefiskartene har ulik livshistorie. Bergnebb og berggylt kan bli 25 år, mens grønngylt kan bli opp mot 10 år, som regel ikke mer enn 7–8 år (bare 3 år på Sørlandet) (Darwall mfl., 1992, Muncaster mfl., 2013, Halvorsen mfl., 2016). Rognkjeksyngel lever 1-2 år i tareskogen, før de blir pelagiske ved 5-6 cm lengde (Myrseth, 1971, Davenport, 1985, Holst 1993, Wienerroither mfl., 2011). De lever pelagisk til de blir kjønnsmodne, hanner når de er 4-5 år og 23-26 cm, hunner når de er 5-6 år og 32-39 cm (Myrseth, 1971, Thorsteinsson, 1981, Davenport, 1985, Albert mfl., 2000).
Det er et betydelig svinn av rensefisk i merdene (Nilsen mfl., 2014). Siden mesteparten av rensefisken som holdes i merdene ikke overlever en produksjonssyklus betyr det at fisken får et kort liv. Svinnet kompenseres med "etterfylling” av rensefisk gjennom hele sesongen, og svinnet er derfor også en direkte drivende faktor i fisket etter leppefisk. Svinnet utgjøres i hovedsak av rømming, predasjon og dødelighet forårsaket av skader, sykdom eller alder. Vi kjenner ikke forholdet mellom disse faktorene, og forholdet vil i stor grad variere gjennom sesongen og mellom oppdrettsanlegg. Grønngylt blir som oftest ikke eldre enn tre år på Sørlandet (Halvorsen mfl., 2016). Fisk fra Sørlandet kan således godt være i sitt tredje leveår, og det kan derfor ikke forventes at den skal leve lenger enn ut sesongen. Praksis har i mange år vært at det aller meste av leppefisken blir brukt i én sesong, men med tilrettelegging av skjul og fôring er det i mange anlegg noe leppefisk som overlever i merdene til våren.
Fisk som er skadet ved fangst eller transport får ofte skjelltap og infeksjoner og dør i løpet av noen uker. Fisk kan også skades ved spyling av nøter, i dødfisksamlere, ved lusebehandling og andre driftsrutiner, og stress og skader gjør at rensefisken er utsatt for ulike sykdommer. Ved dårlige rutiner hos fisker eller på anleggene kan mye av fisken forsvinne i løpet av uker eller måneder etter utsett. All leppefisk vil i naturen oppholde seg nær bunn, berg eller tang og tare der de kan skjule seg for predatorer og finne føde. Rognkjeksyngel vil ofte sitte fastsugd til alger. I en oppdrettsmerd blir det lagt til rette for dette med mange skjul, ofte i form av plasttare og hengende plastplater til rognkjeksen. Mangel på skjul og hvileflater (til rognkjeks) vil sannsynligvis føre til et økt stressnivå hos rensefisken. Bare å beite på lakselus gir ikke nok føde til rensefisken og ikke all rensefisk fungerer som lusespisere. For at fisken skal ha motivasjon til å beite lus av laksen holdes nøtene reine for å unngå at rensefisken spiser seg mett på begroingsorganismer. I forsøk har vi konstatert at leppefisk som har beitet ned lusen på laksen og går i reine nøter, raskt får redusert kondisjon – de sulter (Skiftesvik mfl., 2013). Det foregår uttesting av ulike fôringsmetoder og fôrtyper for leppefisk og rognkjeks. Mange oppdrettere bruker ressurser på å forbedre miljøet for rensefisken, gi den bedre fôr, og håndtere den mer skånsomt. Det pågår også mye FoU-aktivitet for å forbedre velferden til fisken. Innen forskningen arbeides det også med å øke kunnskapsbasen for de ulike artenes atferd og deres velferdsbehov i merdene. Selv det gradvis kommer på plass bedre metoder og kunnskapen øker gjør den økte bruken av rensefisk at problemet med dårlig fiskevelferd hos rensefisk øker i omfang.
6.4 - Smitte og innførsel av uønskede organismer via transport
Under transport av fisk blir også andre organismer, som for eksempel sykdomsfremkallende organismer (agens), flyttet fra opprinnelseslokalitet til utsettingslokalitet, både via transportvannet, transportenheten og via fisken. Gjentatte transporter til samme område vil kunne øke sannsynligheten for spredning og etablering av uønskede organismer (Peeler & Feist, 2011). Når transportvannet slippes ut eller skiftes, vil overlevelsen i stor grad være bestemt av forholdene på utslippsstedet. Dette kan være fysiske forhold som tid på året, temperatur, strømforhold osv. Etablering av agens krever i tillegg mottakelig(e) vert(er). De fleste agens er knyttet til én eller noen få, nært beslektede vertsorganismer. Noen sykdomsfremkallende organismer har imidlertid et bredere vertsspekter og kan smitte mellom arter. Det er også forskjeller i hvilken evne agens har til å tilpasse seg nye verter.
Villfanget leppefisk har, etter at kvotene kom på plass, stått for mellom 30 – 40 % av det totale forbruket av rensefisk i norske oppdrettsmerder (drøye 38% i 2018 og 31% i 2019). I tillegg fiskes og transporteres det omtrent 1 million villfangede leppefisk fra Sverige til Norge. Den omfattende transporten av leppefisk som forekommer langs norskekysten, skjer uten at vi kjenner helsestatus til denne fisken. Forsendelsene av leppefisk skjer i sommerhalvåret, som er den perioden hvor det er størst sannsynlighet for at det kan forekomme levende organismer som er skjult i forsendelsene. Det finnes ingen god nok oversikt over geografiske forskjeller på utbredelsen av sykdommer og parasitter hos leppefisk i norske og svenske farvann, men det foreligger data som tyder på at det er geografiske forskjeller på enkelte gjellepatogener hos bergnebb og grønngylt og på nodavirus-isolater fra bergnebb, grønngylt og berggylt (Korsnes mfl., 2017). Sannsynligvis er det også geografiske forskjeller på enkelte bakterier som kan forårsake sykdom.
Selv om man i dag ikke kjenner til konkrete eksempler på spredning av sykdom via villfanget leppefisk, finnes det mange eksempler på at sykdomsfremkallende organismer er flyttet og introdusert til nye områder som følge av transport av levende fisk (Gozlan mfl., 2006, Peeler mfl., 2011, Peeler & Feist 2011). I Norge er kanskje de mest kjente eksemplene innførsel av parasitten Gyrodactylus salaris fra Sverige på 70-tallet (Mo 1994) og furunkulosebakterien, Aeromonas salmonicida som kom med laksesmolt fra Skotland på midten av 1980-tallet (Egidius 1987).
Eksempler på kjente agens som er påvist hos leppefisk, og som kan tenkes å følge partier av rensefisk er Viral hemmoragisk septikemi virus (VHSV), viral nervevevsnekrose eller nodavirus (VNN) og bakterier som Aeromonas salmonicida, Pasteurella spp. og parasitter som Paramoeba perurans. Det har også være en del fokus på Salmonid alfavirus SAV (som forårsaker pankreassykdom, forkortet PD), Infeksiøs pankreasnekrose virus (IPNV), og ulike typer av Vibrio spp. bakterier.
6.5 - Sykdommer og sykdomsutbrudd i merd
Sykdom og sykdomsutbrudd står for en betydelig andel av svinnet av rensefisk i merd. Dødelighetsmønsteret er ulikt hos de ulike artene. Dette er ikke kartlagt i detalj, men det ser ut til at bergnebb og berggylt er de mest robuste artene, mens det er betydelige problemer med dødelighet hos grønngylt, grasgylt og rognkjeks. En kartlegging av dødelighet og dødelighetsårsaker for rensefisk i merd er beskrevet av Nilsen mfl. (2014) og i rensefiskkampanjen som Mattilsynet gjennomførte i 2019 i samarbeid med NTNU samfunnsforskning og HI (https://www.mattilsynet.no/fisk_og_akvakultur/akvakultur/rensefisk/mattilsynet_sluttrapport_rensefiskkampanje_2018_2019.37769/binary/Mattilsynet%20sluttrapport%20rensefiskkampanje%202018%202019). I tillegg finnes det data fra fisk som er innsendt via fiskehelsetjenesten og analysert ved Veterinærinstituttet (Sommerset mfl., 2020), data fra Havforskningsinstituttets laboratorieforsøk og fangstforsøk (Harkestad 2011, Skiftesvik mfl., 2014), samt informasjon fra fiskere og oppdrettsbedrifter.
Under sykdomsutbrudd hos rensefisken vil det kunne oppstå et økt smittepress ved at agens oppformeres i anleggene og spres derfra. Oppdaterte oversikter over sykdommer hos rognkjeks og leppefisk er gitt i Fiskehelserapporten ugitt av Veterinærinstituttet (Sommerset mfl., 2020) og VKM-rapporten «Risk assessment of fish health associated with the use of cleaner fish in aquaculture» fra 2017.
Videre følger en omtale om de «viktigste agens» relatert til rensefisk.
6.5.1 - Virus
Virusinfeksjoner i rensefisk er lite studert, og det er derfor relativt få virus beskrevet fra rensefisk. Det er sannsynlig at det allerede er blitt spredd fisk smittet med ukjente virus, og de kan også ha bidratt til sykdom og dødelighet blant rensefisk i merder.
6.5.1.1 - Pankreas sykdom (PD)
Pankreas sykdom (PD) forårsakes av salmonid alfavirus (SAV) og per dags dato den virussykdommen som står for de største utfordringene og tapene i norsk oppdrettsnæring. På grunn av de alvorlige konsekvensene knyttet til denne sykdommen på laks, har det vært utredet faren for spredning av PD hos laks via transport av rensefisk. Faren for en slik overføring ble vurdert i en risikovurdering utført ved Veterinærinstituttet, og ble hovedsakelig knyttet til transportvannet (Olsen mfl., 2011). Rensefisk er sett på som lite mottakelig for SAV og det er hittil ikke påvist overføring av SAV mellom lakse- og rensefisk. Men det kan ikke utelukkes at rensefisk kan fungere som en mekanisk vektor (VKM 2017).
6.5.1.2 - Infeksiøs pankreasnekrose (IPN)
Rensefisk ser ikke ut til å utvikle infeksiøs pankreasnekrose (IPN), men kan være bærere av IPN-virus (IPNV) (Gibson & Sommerville, 1996, Gibson mfl., 2002). IPNV har et bredt vertsspekter og det vil derfor være en viss fare for innførsel av IPNV via rensefisk.
Nodavirus, forårsaker viral nervevevsnekrose (VNN) og viral encephalopati og retinopati (VER)
Nodavirus er påvist hos vill berggylt, bergnebb og grønngylt (Korsnes mfl., 2017). Nodavirus har et bredt vertsregister og er kjent fra sykdomsutbrudd hos marin fisk som kveite og torsk i oppdrettssituasjoner (Patel mfl., 2007; Grotmol, 1996).
Viral hemorrhagisk septikemivirus (VHSV) har en stor evne til å tilpasse seg nye verter og habitater. I Norge er VHS en meldepliktig sykdom (http://web.oie.int/eng/maladies/en_classification2010.htm) og vi har fristatus for viruset i oppdrett. Det er derfor viktig å se på faren for innførsel av VHSV til Norge. Viruset finnes i fire ulike genotyper (I-IV) og er vist å kunne infisere mer enn 80 arter i både fersk- og saltvann (OIE). VHSV er påvist i flere arter av villfisk langs norskekysten og sild er særlig mottakelig (Johansen mfl., 2013, Sandlund mfl., 2014). Rensefisk er også mottakelig for VHSV. Viruset er påvist hos leppefisk på Shetland (Munro mfl., 2015) og på rognkjeks på Island (Guðmundsdóttir mfl., 2019). Smitte via føde er en kjent smittevei for VHSV (Ahne 1980, Schönherz mfl., 2012 ) . Det er kjent at liten leppefisk blir spist av laks og regnbueørret, særlig i sulteperioden før slakting. Det er derfor mulig at laksefisk kan bli eksponert for VHSV fra infisert leppefisk, for eksempel ved at leppefisken blir spist. VHS-virusets overlevelse i vann er rapportert som dager (Hawley & Garver, 2008) og uker (Brun & Lillehaug, 2010). Viruset kan også overleve i frossen fisk og vil kunne smitte videre om frossen fisk benytte som fôr (Gudding & Lillehaug, 2018). Nødvendig smittedose er ikke godt kjent, men vil sannsynligvis avhenge av fiskens størrelse/alder, allmenntilstand, temperatur, omgivelsene generelt og virusisolat.
6.5.1.4 - «Nye virus» hos rensefisk
De senere år er nye virus isolert og beskrevet fra rognkjeks. Betydningen av disse virusene, både i forhold til oppdrett men også villfisk, er fortsatt lite kjent. Men det viser igjen at man må forvente at med nye arter i oppdrett vil også nye og hittil ukjente sykdommer dukke opp.
Viruset er assosiert med betennelse og nekroser i lever hos rognkjeks (Skoge mfl., 2018). I tillegg er også bleke gjeller og anemi observert, men det er usikkert hvorvidt denne klinikken er direkte knyttet til virusinfeksjonen (Johansen mfl., 2019). Etter at dette viruset ble identifisert i slutten av 2016, har kartlegging og sykdomsovervåking vist at det er til stede i alle ledd av produksjonen og med relativt høy prevalens. Det har, tilsynelatende, ingen geografisk begrensning, og mye tyder på at viruset både kan overføres horisontalt og vertikalt.
I starten av 2019, ble ytterligere to virus identifisert og karakterisert fra flere tilfeller med forøkt dødelighet på rognkjeksyngel. I enkelte tilfeller var dødeligheten høy. Virusene fikk tentative navn, basert på slektskap, Cyclopterus lumpus Coronavirus (CLuCV) og Cyclopterus lumpus Totivirus (CLuTV). Kliniske tegn og patologi CLuCV er væskefylt tarm (diaré) og irritert tarmepitel. Ingen spesiell klinikk er assosiert med CLuTV (Stian Nylund pers komm).
6.5.1.4.2 - Ranavirus
Ranavirus hos rognkjeks ble påvist første gang på Færøyene i 2014 og siden på Island i 2015 og i Skotland og Irland i 2016. Genetiske analyser tyder på at variantene isolert fra rognkjeks er en ny art av ranavirus (Stagg mfl., 2020). Viruset er foreløpig ikke påvist på rensefisk i Norge (Fiskehelserapporten 2020).
6.5.2 - Bakterier
Det er identifisert en del sykdomsfremkallende bakterier hos både leppefisk og rognkjeks. Noen av disse kan potensielt også forårsake sykdom hos laksefisk. Det er uklart om det er beslektede stammer som er isolert, eller i hvilken grad bakteriene fra leppefisk over tid kan endre sine egenskaper slik at de kan infisere nye vertsarter. Eksempler på bakterier som forårsaker sykdom hos leppefisk og rognkjeks er ulike stammer av Vibrio anguillarum, Aliivibrio ordali, Pasteurella spp., atypisk A. salmonicida, Pseudomonas anguillisepticaog Vibrio splendidus (se f.eks. Harkestad 2011, Johansen 2013, Treasurer 2012, Sommerset mfl., 2020). Infeksjoner med ulike varianter av Pasteurella spp. har økt i forekomst hos oppdrettsfisk, både rensefisk og laks, i de siste årene (Sommerset mfl., 2020). I hvilken grad smitten overføres mellom rensefisk og oppdrettsfisk er usikkert (Poppe mfl., 2012, Alarcón mfl., 2015a). Bakterier som Tenacibaculum spp. og Moritella viscosa er assosiert med sårdannelse hos mange fiskeslag, og særlig laks. Men disse bakteriene er blitt isolert fra sår hos både vill og oppdrettet rognkjeks, og fra oppdrettet berggylt. For de ulike Tenacibaculum artene, T. finnmarkense, T. dicentrarchi og T. maritimum er også direkte smitte mellom rensefisk og laks i produksjon en mulighet, da like varianter av disse er påvist både på laks og rognkjeks (Frisch mfl., 2018; Olsen mfl., 2011; Småge mfl., 2016).
Ut fra den informasjonen som er tilgjengelig i dag, vil rensefisk kunne være bærere av V. anguillarum, Aliivibrio ordali, Pasteurella spp., atypisk A. salmonicida, Pseudomonas anguilliseptica, Aliivibrio spp., V. splendidus, M. viscosa og Tenacibaculum spp. Laks i oppdrett er vaksinert mot V. anguillarum, Aliivibrio salmonicida, M. viscosa og typisk A. salmonicida.
6.5.3 - Parasitter
I 2013 og senere har det vært betydelige problemer med amøbeindusert gjellesykdom (AGD) hos laks i Sør-Norge, forårsaket av amøben Paramoeba perurans. Det er uklart hva som er de viktigste, naturlige reservoarene for P. perurans, men amøben er blitt påvist hos villfanget berggylt og blåstål (VKM, 2014). Infeksjoner og AGD er også observert hos oppdrettet berggylt (Karlsbakk mfl., 2013) og rognkjeks (Bornø & Gulla, 2016), og i grønngylt, berggylt og rognkjeks holdt som rensefisk i laksemerder (Nilsen mfl., 2014, Karlsbakk, 2015). Laks kan smittes med P. perurans som er isolert fra berggylt (Mo mfl., 2014a, Dahle mfl., 2015), og berggylt av amøber fra laks (Dahle mfl., 2015). Dette tyder på at amøben har lav artsspesifisitet, og vi må derfor anta at flytting av fisk med P. perurans representerer en fare for overføring av denne til fisk på mottaksstedet.
Leppefiskartene kan være parasittert av kveis. Norske og skotske studier antyder at infeksjoner med kvalorm (Anisakis simplex) forekommer, men er uvanlige i leppefisk. Det er sannsynligvis lokal variasjon i forekomsten av denne parasitten i leppefiskene. Infeksjoner kan oppstå ved at leppefisken spiser utkast (slo) fra annen fisk, for eksempel i havner. Laks i merd kan bli infisert med A. simplex (Mo mfl., 2014b), men dette er svært uvanlig (Levsen & Maage, 2015). Oppdrettslaksen smittes muligens ved å spise viltfanget infisert leppefisk brukt som rensefisk. En annen kveistype, selorm (Pseudoterranova spp.), kan forekomme i muskulaturen hos leppefisk tatt nær selkolonier. Denne forekomsten er lite undersøkt, følgelig vet vi ikke om P. decipiens og P. krabbei-infeksjoner i leppefisk representerer en trussel med hensyn til kveisinfeksjon i oppdrettslaks. Omfattende screening av norsk oppdrettslaks og regnbueørret har ikke avdekket kveisinfeksjoner i slaktefisk (Levsen & Maage, 2015). Små mengder kveis kan derimot forekomme i den utsorterte taperfisken (Mo mfl., 2014, Levsen & Maage, 2015, Roiha mfl., 2017). Om dette i noen tilfeller har sammenheng med bruk av rensefisk er uvisst.
Skottelus (Caligus elongatus) er en parasitt som lever på ulike fiskearter. Den ligner lakselus, men er mindre. Samtidig med den økende bruken av rognkjeks som rensefisk, har også utfordringene med skottelus-infeksjoner på rognkjeks økt. Rapporter om behov for avlusing av rognkjeks forekommer. Selve innrapporteringen av bruken av avlusningsmetoder og avlusningsmedikamenter gjør at man ikke har oversikt over hvor stor andel som benyttes for avlusning av laks vs rensefisk (Sommerset mfl., 2020). Skottelus fra rognkjeks kan smitte over på annen fisk, men det er trolig uvanlig da rognkjeks synes å være en foretrukket vert (Øines mfl., 2006).
6.5.4 - 6.5.4 Gjenbruk av rensefisk
Oppdrettslokaliteter skal generelt brakklegges etter en produksjonssyklus for å bryte smittesykli. Samtidig er det et behov for å øke overlevelsen av leppefisk, og det diskuteres derfor om gjenbruk av overlevende leppefisk kan være aktuelt. Faren er at noen fisk kan være bærere av agens som så kan spres ved gjenbruk i neste produksjonssyklussyklus. Agens kan i noen tilfeller også endre virulens og tilpasse seg nye verter. Det eksisterer således en fare for at smitteoverføring mellom arter øker (se f.eks. Kennedy mfl., 2015, Sundberg mfl., 2016, Pulkinen mfl., 2010). Det finnes lite relevant informasjon om virulensutvikling som kan relateres til norske forhold, og dette er derfor ikke risikovurdert i denne sammenhengen.
6.6 - Genetiske påvirkninger
Leppefisk lever i geografisk atskilte bestander med begrenset grad av utveksling. Leppefisk som flyttes over lange avstander og deretter rømmer, kan blande seg med lokale bestander og på denne måten påvirke den bestandsgenetiske strukturen på mottaksstedet. Leppefiskene er små, og selv små hull i nøtene representerer en rømningsvei. Dette gjelder særlig bergnebb, som er slankere enn de andre artene (Woll mfl., 2013). Når oppdretterne skifter til nøter med større maskevidde, kan dette resultere i at små leppefisk forsvinner ut av noten. All håndtering av laks medfører en fare for rømming eller dødelighet for rensefisken. Studier av bergnebb og grønngylt tyder på at en innblanding av importert fisk i lokale bestander kan ha funnet sted i et importområde i Trøndelag (Jansson mfl., 2017, Faust mfl., 2018). Et nylig gjennomført studie på grønngylt, som nå er under bearbeiding, viser indikasjoner på at det hittil er de nordligste områdene som er mest påvirket. En teori er at det er enklere for rømt fisk å få rotfeste i områder med nylig etablerte populasjoner, har lav populasjonstetthet og /eller høyt fiskepress på lokale populasjoner (Faust mfl., In prep). Transport representerer derfor en fare for en permanent genetisk påvirkning av lokale leppefiskbestander. Faktorer som spiller inn vil variere mellom arter av leppefisk. Arter med lav overlevelse etter utsett, gjør faren for innblanding i ville bestander er liten.
Berggylt vil være følsom for genetisk endring som følge av å ha lav populasjons tetthet og høy skjevfordeling mellom kjønn. Det er også fare knyttet til bruk av oppdrettet berggylt, som kommer fra få stamfisk, men blir spredd ut til mange oppdrettslokaliteter. Villfanget berggylt har en høyere lovlig minstestørrelse ved fangst enn de andre leppefiskartene, så sannsynligheten for at denne rømmer er lav. Imidlertid kan oppdrettet berggylt settes ut ved mindre størrelser og disse kan ha større sannsynlighet for å rømme gjennom notmaskene enn villfanget fisk. Oppdrettet berggylt er ofte ikke stedegen. Selv om det er større sannsynlighet at liten berggylt rømmer, vil den lange tiden det tar før de rømte små berggyltene kjønnsmodner (ved 22-23 cm). Nylige gjennomførte studier gir indikasjoner på at inndelingen av populasjonsstrukturen til berggylt ligner grønngylt med en nordvestlig og sørøstlig inndeling. Skillet går ved Jæren hvor sandstrendene utgjør en naturlig barriere for genetisk utveksling (Seljestad mfl., 2020). Studiet konkluderer med at transport av berggylt fra Sverige og Sørlandet til Vestlandet og Midt-Norge, vil kunne føre til en miksing av og endring av genetisk forskjellige populasjoner.
Arten som sannsynligvis påvirkes minst er bergnebb. Med sine pelagiske egg vil frekvensen av genutveksling naturlig sett være høyere sammenlignet med andre leppefisk. I tillegg har denne arten høyest populasjonstetthet og størst utbredelse. Men bergnebb er også den vanligste arten i transportene, har best overlevelse etter transport og rømmer mest (Woll mfl., 2013). Bergnebb er småvokst og kan lett unnslippe gjennom maskene. Den er sannsynligvis også mer robust. Ved en undersøkelse av bergnebb samlet inn langs en syd-nord-gradient ble det vist en genetisk variasjon som fulgte gradienten relatert til avstand (Jansson mfl., 2017). Fisken i det eneste "importområdet" i studien avvek imidlertid fra det generelle mønsteret, og var mer lik fisk fra eksportområdet enn fra de nærmeste prøvetakingslokalitetene. Dette kan ha sammenheng med innblanding av importert fisk; det gjøres nå oppfølgende studier i området.
Grønngylt vil i mindre grad rømme (Woll mfl., 2013). Det er rapportert at noen oppdrettere tidligere hadde en praksis hvor leppefisk ble satt fri etter bruk.
Endringer i den bestandsgenetiske strukturen vil også kunne skje i lokale populasjoner ved overfiske, som følge av tap av genetisk variasjon. Dette er en ofte ignorert, men like vel viktig faktor. Leppefiskene lever i mer eller mindre isolerte, lokale populasjoner. Bruk av leppefisk som er fraktet nordover (eksempelvis fra Sverige til Nordland) kan representere akvakultur med arter som ikke forekommer naturlig i området (grønngylt i nordlige områder) og/eller bruk av ikke-stedegen fisk.
Rognkjeks som benyttes som rensefisk kommer fra oppdrett. Det er dels omfattende flytting av befruktet rogn og settefisk, og rognkjeks som settes ut har i dag ofte ikke lokalt eller regionalt genetisk opphav. Studier av genetikken til rognkjeks langs norskekysten viser tvetydige resultater i forhold til grad av genetiske forskjeller. Jónsdóttir mfl. (2018) fant liten eller ingen forskjell i den genetiske strukturen, men undersøkelser gjort av Whittaker mfl. (2018) viste det motsatte. I det sistnevnte arbeidet vises det til funn av distinkt forskjellige genetiske populasjoner, som i tillegg er små. Forfatteren peker på at dette gjør rognkjekspopulasjoner sårbare i forhold til genetisk påvirkning og endring. Disse ulike funnene viser behovet for å undersøke dette nærmere.
6.7 - Referanser
Ahne W (1980). Experimental infection of pike ( Esox lucius ) with Egtvedvirus. Tierarztl Umsch 35: 225-229.
Alarcón M, Gulla S, Røsæg MV, Rønnseth A, Wergeland H, Poppe TT, Nilsen H, Colquhoun DJ (2015). Pasteurellosis in lumpsucker Cyclopterus lumpus , farmed in Norway. J Fish Dis (doi:10.1111/jfd.12366).
Albert OT, Bertelsen B, Jonsson ST, Pettersen IH, Torstensen E (2000). Lumpsucker ( Cyclopterus lumpus ) otoliths: dissection, mounting and age-reading. Fiskeriforskning, Rapport 2000, 1-22.
Alvsvåg J (1993). Fødeval, vekst og energiallokering hos gras gylt (Centrolabrus exoletus L.) og grønngylt (Symphodus melops L.) (Pisces: Labridae) på vestkysten av Noreg, og diettoverlapping med O og l-gruppe torsk (Gadus morhua L.). University of Bergen.
Bergström, L., Karlsson M, Bergström U, Pihl L, Kraufvelin P (2016) Distribution of mesopredatory fish determined by habitat variables in a predator-depleted coastal system. Marine Biology 163(10): p. 201.
Bornø G, Gulla S (2016). Helsesituasjonen hos rensefisk. S. 70-73 i (Hjeltnes m.fl. red). Fiskehelserapporten 2015. Oslo: Veterinærinstituttet.
Brun E, Lillehaug A (2010). Risikoprofil for sykdommer i norsk fiskeoppdrett. Norge, Veterinærinstituttet.
Dahle OM, Nylund A, Karlsbakk E, Andersen L, Blindheim S, Glosvik H, Breck O (2015). Smitteoverføring av Paramoeba perurans mellom laks og berggylt - bruk av to klonale isolat fra henholdsvis laks og berggylt. Abstract, Frisk Fisk, Tromsø.
Darwall WRT, Costello MJ, Donnelly R, Lysaght S (1992). Implications of life-history strategies for a new wrasse fishery. Journal of Fish Biology 41(sb):111–123.
Davenport J (1985). Synopsis of biological data on the lumpsucker Cyclopterus lumpus (Linnaeus, 1758). FAO Fisheries Synopsis. No. 147, 1-31.
Deady S, Fives JM (1995). Diet of ballan wrasse, Labrus bergylta, and some comparisons with the diet of corkwing wrasse, Crenilabrus melops. Journal of the Marine Biological Association of the United Kingdom 75(03):651–665. Cambridge University Press.
Egidius E (1987). Import of furunculosis to Norway with Atlantic salmon smolts from Scotland. ICES C.M., 1987/F:8, 7 p.
Fernández‐Chacón, A., D. Villegas‐Ríos, E. Moland, M. L. Baskett, E. M. Olsen, S. M. Carlson. 2020. Protected areas buffer against harvest selection and rebuild phenotypic complexity. Ecological Applications:eap.2108. Ecological Society of America.
Faust E, Halvorsen KT, Andersen P, Knutsen H, André C (2018). Cleaner fish escape salmon farms and hybridize with local wrasse populations. Royal Society Open Science 5(3):171752.
Faust E*, Jansson E *, André C, Halvorsen K, Seljestad GW & Glover KA (* with equal contribution). In preparation. “Large scale survey of escape and hybridisation of cleaner fish in aquaculture
Frisch K, Småge SB, Brevik J, Duesund H, Nylund A (2018). Genotyping of Tenacibaculum maritimum isolates from farmed Atlantic salmon in Western Canada. J. Fish Dis. 41, 131–137. https://doi.org/10.1111/jfd.12687
Gibson DR, Sommerville C (1996). The potential for viral problems related to the use of wrasse in the farming of Atlantic Salmon. s 240-246 I: Sayer MDJ, Treasurer JW, Costello MJ (red.): Wrasse biology and use in aquaculture. Fishing news books. Blackwell, Oxford.
Gibson DR, Smail DA, Sommerville C (2002). Infectious pancreatic necrosis virus: experimental infection of goldsinny wrasse, Ctenolabrus rupestris L. (Labridae). J Fish Dis 21, 399-406.
Gozlan RE, Peeler EJ, Longshaw M, St-Hilaire S, Feist SW (2006). Effect of microbial pathogens on the diversity of aquatic populations, notably in Europe. Microbes and Infection, 8, 1358-1364.
Grotmol S, Totland GK, Thorud K, Hjeltnes BK (1997). Vacuolating encephalopathy and retinopathy associated with a nodavirus-like agent: a probable cause of mass mortality of cultured larval and juvenile Atlantic halibut Hippoglossus hippoglossus . Dis Aquat Org 29, 85-97.
Gudding R, Lillehaug A (red.) (2018). Smitte mellom oppdrettsfisk og villfisk: Kunnskapsstatus og risikovurdering. Rapport 12, Veterinærinstituttet
Guðmundsdóttir S, Vendramin N, Cuenca A, Sigurðarsdóttir, Kristmundsson A, Iburb TM, Olesen NJ (2019). Outbreak of viral haemorrhagic septicaemia (VHS) in lumpfish ( Cyclopterus lumpus ) in Iceland caused by VHS virus genotype IV. J Fish Dis 42(1): 47-62.
Halvorsen KT, Sørdalen TK, Durif C, Knutsen H, Olsen EM, Skiftesvik AB, Rustand TE, Bjelland RM, Vøllestad LA (2016). Male-biased sexual size dimorphism in the nest building corkwing wrasse ( Symphodus melops ): implications for a size regulated fishery. ICES Journal of Marine Science 73(10):2586–2594.
Halvorsen KT, Bjelland R, Jørgensen T, Skiftesvik A B (2017a). Forsøksfiske for selektiv fangst av berggylt. Fisken og Havet (8).
Halvorsen KT, Larsen T, Sørdalen TK, Vøllestad LA, Knutsen H, Olsen EM (2017b). Impact of harvesting cleaner fish for salmonid aquaculture assessed from replicated coastal marine protected areas. Marine Biology Research 13(4):359–369.
Halvorsen KT, Sørdalen TK, Vøllestad LA, Skiftesvik AB, Espeland SH, Olsen EM (2017c). Sex- and size-selective harvesting of corkwing wrasse (Symphodus melops)—a cleaner fish used in salmonid aquaculture. ICES Journal of Marine Science 74(3):660–669.
Halvorsen K, Skiftesvik AB, Jørgensen T (2019). Kunnskapsstøtte – anbefaling om redusert inngangsstørrelse i teiner i fisket etter leppefisk.
Halvorsen K, Sørdalen TK, Larsen T, Rafoss T, Skiftesvik AB (2020). Kunnskapsbasert innovasjon for optimal ressursutnyttelse i leppefiskeriet. Rapport fra Havforskningen.
Harkestad LS (2011). Eksperimentell smitte av grønngylt, Symphodus melops, med V.tapetis-isolatene CECT 4600, LP2 og NRP45. Mastergradsoppgave i havbruksbiologi, Universitetet i Bergen 2011, 136 s.
Hawley LM, Garver KA (2008). Stability of viral hemorrhagic septicemia virus (VHSV) in freshwater and seawater at various temperatures. Dis Aquat Org 82, 171-178.
Holst JC (1993). Observations on the distribution of lumpsucker ( Cyclopterus lumpus , L.) in the Norwegian Sea. Fish Res 17, 369-372.
Jansson E, Quintela M, Dahle G, Albretsen J, Knutsen H, André C, Strand Å, Mortensen S, Taggart JB, Karlsbakk E, Kvamme BO, Glover KA (2017). Genetic analysis of goldsinny wrasse reveals evolutionary insights into population connectivity and potential evidence of inadvertent translocations via aquaculture. ICES Journal of Marine Science. doi:10.1093/icesjms/fsx046.
Johansen R (red.) (2013). Fiskehelserapporten 2012. Oslo, Veterinærinstituttet.
Johansen Johansen R, Tønnessen Ø, Nylund S (2019) Hvordan få kontroll på helseutfordringer på rensefisk. Foredrag Rensefiskkonferansen 2019
Jónsdóttir ÓDB, Schregel J, Hagen SB, Tobiassen C, Aarnes SG, Imsland AKD (2018). Population genetic structure of lumpfish along the Norwegian coast: aquaculture implications. Aquacult Int 26, 49–60.
Jørgensen T, Bjelland R, Halvorsen K, Durif C, Shema S, Skiftesvik AB (2017). Seleksjon i leppefiskredskap. Fisken og Havet (2).
Karlsbakk E (2015). Amøbisk gjellesykdom (AGD) – litt om den nye plagen. Havforskningsrapporten 2015. Fisken og havet, særnr. 1- 2015: 33-35.
Karlsbakk E, Olsen AB, Einen A-CB, Mo TA, Fiksdal IU, Aase H, Kalgraff C, Skår S-Å, Hansen H (2013). Amoebic gill disease due to Paramoeba perurans in ballan wrasse ( Labrus bergylta ). Aquaculture 412-413, 41-44.
Kennedy DA, Kurath G, Brito IL, Purcell MK, Read AF, Winton JR, Wargo AR (2015). Potential drivers of virulence evolution in aquaculture. Evolutionary Applications 9, 344-354.
Korsnes K, Karlsbakk E, Skaar CK, Sælemyr L, Nylund A, Kvamme BO, Mortensen S (2017). High nervous necrosis virus (NNV) diversity in wild wrasse (Labridae) in Norway. Diseases of Aquatic Organisms 126:43-50.
Kraufvelin P, Christie PH, Gitmark JK (2020) Top-down release of mesopredatory fish is a weaker structuring driver of temperate rocky shore communities than bottom-up nutrient enrichment. Marine Biology 167(4):1–20. Springer Berlin Heidelberg.
Levsen A, Maage A (2015). Nasjonal undersøkelse av forekomst av Anisakis i norsk oppdrettslaks (pp. 10). Bergen: Nasjonalt institutt for ernærings- og sjømatforskning (NIFES).
Mo TA, Emilsen V, Hansen H, Poppe TT (2014a). Smitteforsøk med isolat av Paramoeba perurans fra berggylt og laks for å studere mottakelighet og infeksjonsutvikling hos laks. Rapport, Veterinærinstituttet 31 Jan. 2014. 13 s.
Mo TA, Gahr A, Hansen H, Hoel E, Oaland Ø, Poppe TT (2014b). Presence of Anisakis simplex (Rudolphi, 1809 det. Krabbe, 1878) and Hysterothylacium aduncum (Rudolphi, 1802) (Nematoda; Anisakidae) in runts of farmed Atlantic salmon, Salmo salar L. J Fish Dis 37 , 135-140.
Mo TA (1994) Status of Gyrodactylus salaris problems and research in Norway. In: Parasitic Diseases of Fish (Ed. J.W. Lewis), pp. 43–48. Samara Publishing, Tresaith, Dyfed, Wales.
Munro ES, McIntosh RE, Weir SJ, Noguera PA, Sandilands JM, Matejusova I, Mayes AS, Smith R (2015). A mortality event in wrasse species (Labridae) associated with the presence of viral haemorrhagic septicaemia virus. J Fish Dis 38, 335-341.
Muncaster, S., B. Norberg, and E. Andersson. 2013. Natural sex change in the temperate protogynous Ballan wrasse Labrus bergylta. Journal of Fish Biology 82(6):1858–1870. Blackwell Publishing Ltd.
Myrseth B (1971). Fekunditet, vekst, levevis og ernæring hos Cyclopterus lumpus L. Hovedoppgave, Univ. Bergen/HI 1971. 113 s.
Nedreaas K, Aglen A, Gjøsæter J, Jørstad K, Knutsen H, Smedstad O, Svåsand T, Ågotnes P (2008). Kysttorskforvaltning på Vestlandet og langs Skagerrakkysten. Vurdering av status for kysttorsk på strekningen svenskegrensen–Stad med forslag om forvaltningstiltak. Fisken og Havet, 5-2008. 106 pp.
Nilsen A, Viljugrein H, Røsæg MV, Colquhoun D (2014). Rensefiskhelse – kartlegging av dødelighet og dødelighetsårsaker, Veterinærinstituttets rapportserie nr 12, 2014.
Olsen AB, Jensen BB, Nilsen H, Grøntvedt RN, Gjerset B, Taksdal T, Høgåsen HR (2011). Risikovurdering for spredning av pancreas disease virus (PD-virus) ved bruk av leppefisk i norsk lakseoppdrett. Veterinærinstituttets rapportserie 7-2011. Oslo: Veterinærinstituttet.
Olsen AB, Nilsen H, Sandlund N, Mikkelsen H, Sørum H, Colquhoun DJ (2011) Tenacibaculum sp. associated with winter ulcers in sea-reared Atlantic salmon Salmo salar . Diseases of Aquatic Organisms 94 (3): 189-199.
Olsen EM, Halvorsen KT, Larsen T, Kuparinen A (2018). Potential for managing life history diversity in a commercially exploited intermediate predator, the goldsinny wrasse ( Ctenolabrus rupestris ). ICES Journal of Marine Science 76(1):357–357.
Patel S, Korsnes K, Bergh Ø, Vik-Mo F, Pedersen J, Nerland AH (2007) Nodavirus in farmed Atlantic cod Gadus morhua in Norway. Dis Aquat Org 77: 169-173
Peeler EJ, Oidtmann BC, Midtlyng PJ, Miossec L & Gozlan R (2011) Non-native aquatic animals introductions have driven disease emergence in Europe. Biological Invasions, 13, 1291-1303.
Peeler, EJ & Feist, SW (2011) Human intervention in freshwater ecosystems drives disease emergence. Freshwater Biology.
Poppe TT, Taksdal T, Skjelstad H, Sviland C, Vågnes Ø, Colquhoun DJ (2012). Nye arter – nye diagnostiske utfordringer. Norsk Veterinærtidsskrift 124(1): 19-21.
Pulkinen K, Suomalainen L-R, Read AF, Ebert D, Rintamäki, Valtonen ET (2015). Intensive fish farming and the evolution of pathogen virulence: the case of columnaris disease in Finland. Proc Royal Soc B 277, 593-600.
Reamon, M (2020). Assessing the impacts of marine protected areas on wrasse populations in Norway. Master thesis. University of Agder.
Ruud, M (2020). Catchability and selectivity in a multispecies wrasse fishery in Western Norway. Master thesis. University of Agder.
Roiha IS, Maage A, Levsen A (2017). Nasjonal undersøking av førekomst av Anisakis simplex i norsk oppdrettsaure (Onchorhynchus mykiss). Rapport, NIFES 26.01.2017; 12 p.
Sandlund N, Gjerset B, Bergh Ø, Modahl I, Olesen NJ, Johansen R (2014) Screening for viral hemorrhagic septicemia virus in marine fish along the Norwegian coastal line." PLoS ONE9 (9): e108529.
Sayer MDJ, Gibson RN, Atkinson RJA (1995). Growth, diet and condition of goldsinny on the west coast of Scotland. Journal of Fish Biology 46(June 1993):317–340.
Schönherz AA, Hansen MHH, Jorgensen HBH, Berg P, Lorenzen N, Einer-Jensen K (2012). Oral transmission as a route of infection for viral haemorrhagic septicaemia virus in rainbow trout, Oncorhynchus mykiss (Walbaum). J Fish Dis 35, 395-406.
Seljestad GW, Quintela M, Faust E, Halvorsen K, Besnier F, Jansson E, Dahle G, Knutsen H, André C, Folkvord A, Glover KA (2020) “A cleaner-break”: Genetic divergence between geographic groups and sympatric phenotypes revealed in ballan wrasse ( Labrus bergylta ). Ecology and Evolution. Crossref DOI link: https://doi.org/10.1002/ECE3.6404
Skiftesvik AB, Bjelland RM, Durif CMFI, Johansen IS, Browman HI (2013). Delousing of Atlantic salmon ( Salmo salar ) by cultured vs. wild ballan wrasse ( Labrus bergylta ). Aquaculture 402-403, 113-118.
Skiftesvik AB, Blom G, Agnalt A-L, Durif CMF, Browman HI, Bjelland RM, Harkestad LH, Farestveit E, Paulsen OI, Fauske M, Havelin T, Johnsen K, Mortensen S. (2014a). Wrasse (Labridae) as cleaner fish in salmonid aquaculture – The Hardangerfjord as a case study. Mar Biol Res 10, 289-300.
Skiftesvik AB, Durif D, Bjelland R, Browman H, Holm E, Jørgensen T, Løkkeborg S, Utne Palm AC, Skar Tysseland AB, Hoddevik Ulvestad B, Aasen A, Axelsen BE, Mortensen S, Sælemyr L, Skår CK, Karlsbakk E, Einen ACB, Nordbø J, Omdal LM, Isachsen CH, Espeland SH, Semb Johannesen I, Olausson S og Elen E ( 2014b). Bestander og fangstkvaliteter av leppefisk. Rapport fra Havforskningen nr. 3-2014. Sluttrapport FHF prosjekt 900609.
Skiftesvik AB, Durif CMF, Bjelland RM, Browman HI (2015). Distribution and habitat preferences of five species of wrasse (Family Labridae) in a Norwegian fjord. ICES Journal of Marine Science 72(3):890–899.
Skoge RH, Brattespe J, Økland AL, Plarre H, Nylund A, (2018). New virus of the family Flaviviridae detected in lumpfish ( Cyclopterus lumpus ). Arch. Virol. 163, 679–685. https://doi.org/10.1007/s00705-017-3643-3
Småge SB, Brevik ØJ, Duesund H, Ottem KF, Watanabe K, Nylund A, (2016). Tenacibaculum finnmarkense sp. nov., a fish pathogenic bacterium of the family Flavobacteriaceae isolated from Atlantic salmon. Antonie van Leeuwenhoek, Int. J. Gen. Mol. Microbiol. 109, 273–285.
Sommerset I, Walde C S, Bang Jensen B, Bornø B, Haukaas A og Brun E (red). (2020) Fiskehelserapporten 2019, Oslo, Veterinærinstituttet
Stagg HEB, Guðmundsdóttir S, Vendramin N, Ruane NM, Sigurðardóttir H, Christiansen DH, Cuenca A, Petersen PE, Munro ES, Pobov VL, Subramaniam K, Imnoi K, Waltzek TB, Olesen NJ (2020). Characterization of ranaviruses isolated from lumpfish Cyclopterus lumpus L. in the North Atlantic area: proposal for a new ranavirus species (European North Atlantic Ranavirus). Journal of General Virology 101(2): 198-207.
Steven GA (1933). The food consumed by shags and cormorants around the shores of Cornwall (England). Journal of the Marine Biological Association of the United Kingdom 19(01):277. Cambridge University Press.
Sundberg L-R, Ketola T, Laanto E, Kinnula H, Bamford JKH, Penttinen R, Mappes J (2016). Intensive aquaculture selects for increased virulence and interference competition in bacteria. Proc Royal Soc B 283, 20153069. DOI: 10.1098/rspb.2015.3069.
Sørdalen, TK, Halvorsen K T, Vøllestad L.A, Moland E, Olsen EM (2020). Marine protected areas rescue a sexually selected trait in European lobster. Evolutionary Applications:1–12.
Thorsteinsson V (1981). The ageing validation of the lumpsucker ( Cyclopterus lumpus ) and the age composition of the lumpsucker in Icelandic lumpsucker fisheries. ICES CM.1981/G58 Demersal Fish Committee, 26 p.
Treasurer JW (2012). Diseases of north European wrasse (Labridae) and possible interactions with cohabited farmed salmon, Salmo salar L. Review article. J Fish Dis 35, 555-562.
VKM (2014). Risk assessment of amoebic gill disease, Panel on Animal Health and Welfare; VKM Report 2014: 11, Norwegian Scientific Committee for Food Safety (VKM), Oslo, Norway.
VKM (2017). Risk assessment of fish health associated with the use of cleaner fish in aquaculture. Opinion of the Panel on Animal Health and Welfare of the Norwegian Scientific Committee for Food and Environment. VKM report 2017:32, Norwegian Scientific Committee for Food and Environment (VKM), Oslo, Norway (https://vkm.no/risikovurderinger/allevurderinger/rensefiskogrisikoforoverforingavsmittetiloppdrettslaks.4.d44969415d027c43cf6562a.html)
Wienerroither R, Johannesen E, Dolgov A, Byrkjedal I, Bjelland O, Drevetnyak K, Eriksen K, Høines Å, Langhelle G, Langøy H, Prokhorova T, Prozorkevich D, Wenneck T (2011). Atlas of the Barents Sea Fishes. IMR/PINRO Joint Report Series (ISSN 1502-8828). 1, 272 s.
Whittaker BA, Consuegra S, Garcia de Leaniz C. (2018) Genetic and phenotypic differentiation of lumpfish (Cyclopterus lumpus) across the North Atlantic: implications for conservation and aquaculture. Peer J 6:e5974. doi:10.7717/peerj.5974
Woll A, Solevåg SE, Hansen Aas G, Bakke S, Skiftesvik AB, Bjelland R (2013). Velferd leppefisk i merd. Rapport nr MA 13-07, Møreforskning.
Øines Ø, Simonsen JH, Knutsen JA, Heuch PA (2006). Host preference of adult Caligus elongatus Nordmann in the laboratory and its implications for Atlantic cod aquaculture. J Fish Dis. 29,167-74.
Östman Ö, Eklöf J, Eriksson BK, Olsson J, Moksnes PO, Bergström U (2016). Top-down control as important as nutrient enrichment for eutrophication effects in North Atlantic coastal ecosystems. Journal of Applied Ecology 53(4):1138–1147.
6.8 - Appendiks
Region
Antall referanse-fiskere
Gj.snitt antall redskaper per fangst-rapport
Torsk – Antall per fisker per år - variasjon
Torsk – Gj.snitt antall per fisker per år
Lyr – Antall per fisker per år - variasjon
Lyr – Gj.snitt antall per fisker per år
Ål – Antall per fisker per år - variasjon
Ål – Gj.snitt antall per fisker per år
Hummer. Antall per fisker per år - variasjon
Hummer. Gj.snitt antall per fisker per år
Taske-krabbe. Antall per fisker per år - variasjon
Taske-krabbe. Gj.snitt antall per fisker per år
1
5
35
7-2990
474
0-307
67
0-1680
222
0-160
18
0-679
168
2
4
191
0-2450
395
0-2740
482
30-5070
1420
10-680
129
60-3420
819
3
5
200
8-4508
903
0-1506
215
0-5574
682
0-1152
100
0-30517
2986
Tabell 1 Bifangst av torsk, lyr, ål, hummer og taskekrabbe i det norske leppefisk fisket med teiner basert på rapporter fra Havforskningsinstituttets nettverk av referansefiskere i 2014-2018 (N=1399 fangstrapporter). Bare teiner med inngangssperre og seleksjonsinnretning er tatt med, men for 2014 er også teiner med 10 mm fluktåpning inkludert. Legg merke til den store variasjonen i bifangsten per fisker per år. Region 1 = Sørlandet, region 2= Vestlandet og Region 3 = kysten nord for Stad.
Region
Antall fiskere1)
Torsk – Totalt antall per år
Lyr – Totalt antall per år
Ål – Totalt antall per år
Hummer – Totalt antall per år
Taskekrabbe – Totalt antall per år
1
136
64513
9083
30153
2477
22906
2
212
83675
102102
301122
27364
173563
3
124
112011
26668
84522
12369
370318
SUM
472
260198
137853
415796
42210
566787
Tabell 2 Gjennomsnittlig bifangst per fisker per år i Tabell 1 (Appendiks) oppskalert med gjennomsnittlig (2014-2018) antall fiskere i hver region for å gi et grovt estimat av den totale bifangsten av torsk, lyr, ål, hummer og taskekrabbe i det norske leppefisk fisket med teiner. Region 1 = Sørlandet, region 2= Vestlandet og Region 3 = kysten nord for Stad.
1)Gjennomsnitt for 2014-2017. Kilde: Fiskeridirektoratet
Laks i merd. Foto: Frode Oppedal/Havforskningsinstituttet
7.1 - Innledning
Fisk er beskyttet av Lov om Dyrevelferd på samme måte som andre produksjonsdyr, og loven sier bl.a. at: «Dyr har egenverdi uavhengig av den nytteverdien de måtte ha for mennesker. Dyr skal behandles godt og beskyttes mot fare for unødige påkjenninger og belastninger (§3)» og at «Dyreholder skal påse at driftsformer, metoder, utstyr og tekniske løsninger som brukes til dyr er egnet til å ivareta hensynet til dyrenes velferd” (§8). Dette er klare krav, men hva som menes med «unødige påkjenninger og belastninger» og hvor grensene går er uklart. Hva som egentlig menes med «dyrenes velferd» er heller ikke definert, og har store rom for fortolkning. I de nylig publiserte kunnskapssoppdateringen om velferd og velferdsindikatorer hos laks og regnbueørret (Noble mfl. 2018, 2020) blir fiskevelferd definert som «livskvalitet som oppfattet av dyret selv». Denne definisjonen samsvarer godt med hvordan folk flest oppfatter begrepet dyrevelferd, der særlig dyrenes (selvopplevde) mulige lidelser er hovedgrunnen til at de bekymrer seg for dyrenes velferd.
For at fisken skal oppleve god velferd må fisken tilbys miljøforhold og omgivelser den er i stand til å tilpasse seg og mestre. Indikatorer på god mestring er at de er friske og sunne, vokser normalt og har normal atferd. Ved håndtering av fisken må det også brukes metoder som ikke skader fisken, eller at den utsettes for miljøforhold som påfører dem høyt stressnivå eller smerte. Indikatorer på dårlig velferd er sykdom, parasitter og skader, dårlig vekst og avmagring, og avvikende atferd, som f.eks. at de står og sturer langs nota og viser liten respons på fôring (Noble mfl. 2018, 2020). At noen fisk dør er også selvfølgelig en indikator på dårlig velferd, da vi må anta at fisk som har dødd har opplevd kortere eller lengre perioder med dårlig velferd og at det er flere syke eller skadete fisk i oppdrettsenheten.
Selv om dødelighet ikke er et direkte mål på hvor god livskvaliteten er, så er dette den eneste velferdsindikatoren som oppdretterne rapporterer regelmessig til Fiskeridirektoratet. Antall fisk i begynnelsen av måneden og antall døde i måneden før rapporteres innen 7. i hver måned for hver enkelt oppdrettsmerd som er i drift. Hvis fisk blir flyttet fra en merd til en annen (f.eks. i forbindelse med en avlusing) blir det svært vanskelig å følge en gruppe fisk gjennom en hel produksjon og beregne dødeligheten for hele produksjonen. For å omgå dette problemet har vi derfor valgt å benytte kun månedlig dødelighet som velferdsindikator i dataanalysen under. Denne analysen kunne ha blitt gjort betydelig mer nøyaktig og gitt bedre informasjon hvis det ble rapportert en fiskegruppe-ID sammen med de månedlige produksjonsdataene som blir rapportert til Fiskeridirektoratet og de ukentlige luse- og avlusingsdataene som blir rapportert til Mattilsynet.
Nedenfor vil vi denne gangen bare sette søkelys på velferd hos oppdrettslaks og rensefisk, og ikke vurdere regnbueørret, kveite og andre arter i oppdrett. Alle antall- og dødelighetsberegninger er basert på månedlig innrapportert data til Fiskeridirektoratet og Mattilsynet fra alle norske oppdrettslokaliteter som var i drift i perioden 2004-2020.
Atlantisk laks (Salmo salar L.) har som de fleste fiskearter et rikt sanseapparat og en avansert hjerne som gjør de i stand til å oppleve sine omgivelser, fange byttedyr og unngå farer. Lakseyngel og parr lever de første 2-5 årene i elvene som bunnfisk og er tilpasset et liv i sterk vannstrøm, varierende miljøforhold, relativt små vannvolumer, og høy risiko for å bli spist av predatorer. Etter smoltifisering, når de er rundt 20 g, vandrer de relativt få som har overlevd ned elva og videre ut i Norskehavet for å gjøre seg store og feite (Aas mfl. 2010). Også der er de utsatt for predatorer, sykdom og parasitter, og som oftest er det langt mindre enn 10% som 1-3 år senere og 2-20 kg større vandrer tilbake for å gyte, som oftest i elva de kom fra (Aas mfl. 2011). Laksen er derfor en robust fisk tilpasset store miljøforskjeller og lange vandringer, og derfor også en fisk som er godt egnet til et liv i oppdrett. Oppdrettslaksen er i tillegg tilpasset livet i oppdrett gjennom mange generasjoner av selektivt utvalg av familiene med de beste egenskapene for å mestre livet i oppdrett, og vokser nå langt bedre under oppdrettsbetingelser enn avkom fra vill laks (Solberg mfl. 2013). Til forskjell fra villaksen er oppdrettslaksen fanget i et begrenset vannvolum der den er i tett kontakt med tusener av andre laks, er utsatt for et høyere smittepress fra virus, bakterier og parasitter, og får tildelt et næringsrikt, men ensformig og dødt fôr med høyt tørrstoff- og planteinnhold. I tillegg er oppdrettslaksen utsatt for håndteringsprosedyrer som sortering, vaksinering, trenging og avlusing som utsetter de for stress og skader.
7.2.1 - Velferd og dødelighet i ferskvannsfasen
Figur 7.1 Produksjon av lakseyngel og laksesmolt i årene 2004-2019. Stolpediagrammet viser totalt antall klekt yngel per år (blå stolper), antall rapportert som død (røde stolper), antall rapportert som destruert (oransje stolper) og antall rapportert som rømt og annet svinn (lilla stolper). Linjen indikerer antall laksesmolt solgt for utsett i sjø det angitte året. Kilde: Fiskeridirektoratet, tall for 2019 er oppgitt som foreløpige.
Antall solgte laksesmolt økte fra 148 millioner i 2004 til 373 millioner i 2019 (Figur 7.1). For å produsere laksesmolten ble det siden 2010 årlig klekket over 400 millioner lakseegg, hvor 75-80 % vokste opp til salgbar smolt. Dødeligheten i klekkeriene varierte i perioden 2004-2019 fra 27 til 65 millioner per år. I tillegg ble omtrent like mye av settefisken destruert, både av kvalitetsårsaker (størrelse, deformiteter og skader), sykdom og trolig også på grunn av markedsmessige årsaker og produksjonsbegrensninger i matfiskfasen. Mellom 3 og 13 millioner lakseyngel og parr er også oppgitt som rømt og annet svinn ulike år. Totalt sett betyr det at dødeligheten i settefiskanleggene er høyere enn i sjøfasen, noe som indikerer betydelige velferdsproblemer også der. I settefiskanleggene dør mest fisk under 3 g (Tørud mfl. 2019). I en spørreundersøkelse blir det svart at også vaksinering, sortering og transport medfører høy risiko for dødelighet (Tørud mfl. 2019)
7.2.2 - Velferd og dødelighet i sjøfasen
Antall laks i sjø, dvs. antall laks som til enhver tid er i sjømerder, økte raskt fra år til år frem til 2012 hvor en for første gang passerte over 400 millioner laks i sjøen deler av året (Figur 7.2A). Fra 2013 til nå har veksten vært mer moderat. Antall laks i sjøen varierer gjennom året avhengig av når ny fisk blir satt ut og når stor fisk blir slaktet.
Figur 7.2
Laks i sjø og prosent døde 2005-2019. A) Antall fisk registrert i sjø per måned. B) Prosent døde oppdrettslaks per måned. Sirkler angir månedsverdiene, mens de heltrukne svarte linjene angir snitt månedsverdiene per år (for 2019 er maiverdien ikke inkludert i årssnittet). Data: Fiskeridirektoratet.
I perioden 2005-12 lå gjennomsnittlig månedlig innrapportert dødelighet for alle størrelser rundt 1.0 %, men med betydelig variasjon fra måned til måned mellom 0,5-1% (Figur 7.2B). I 2012 ble det registrert en tydelig nedgang i dødeligheten, men dødeligheten økte i årene senere til omtrent samme nivå, men med noe mindre variasjon enn før 2012 (Fig. 7.2B). Som vist nedenfor skyldes dette i hovedsak høyere dødelighet etter lusebehandling av større laks (se 20.2.4). Den svært høye dødeligheten registrert i mai 2019 (Figur 7.2B) er en utligger som skyldes oppblomstringen av den giftige algen Chrysochromulina leadbeateri i deler av Ofoten og Lofoten som tok livet av over 7 millioner laks.
Hvor godt laksesmolten tåler overgangen fra settefiskanlegg til sjø er en indikator på hvor robust fisken er for livet i merdene. Denne overgangen medfører nødvendigvis risiko for stress og skader i forbindelse med overføring til bil eller brønnbåt, transporttid, vær og sjø -og veiforhold under transport, vannkvalitet i transporttanker, utpumping til merdene. Overføring til en merd medfører også et helt nytt miljø hva gjelder lysforhold, temperatur, vannkvalitet, patogener og fysiske forhold som laksen må både fysiologisk og atferdsmessig tilpasse seg. Laks som er svekket av tidligere sykdom eller som har opplevd sub-optimale oppdrettsbetingelser i ferskvannsfasen har dårligere forutsetninger for å mestre summen av de overnevnte stressfaktorer. Den største risikofaktoren er imidlertid at ikke alle individene er skikkelig smoltifisert, som vil si at de er fysiologisk tilpasset sjøvann og kan regulere salt- og vannbalansen i cellene. Utsett med laks der deler av populasjonen er ufullstendig smoltifisert, eller er forbi «smolt-vinduet» og har resmoltifisert, vil få langvarig osmotisk ubalanse ved overføring til sjøvann og risiko for høy dødelighet, særlig ved lav sjøvannstemperatur (Sigholt og Finstad, 1990). Dette samsvarer med at rapportert dødelighet i utsettingsmåned er høyest ved 0-4°C (Figur 7.3AB.)
Dataene viser også at dødelighet ved utsett var generelt høyere før 2012 enn etter, og kanskje særlig for høstutsett. I tillegg viser analysen at dødelighet i forbindelse med høy temperatur ved utsett etter 2012 har vært på samme nivå som de moderate temperaturene. I 2019 lå dødeligheten i forbindelse med utsett på samme nivå som årene før på våren, men tydelig lavere dødelighet for høstutsettene (Figur7.3F). Basert på dette kan det se ut som at særlig høstsmolten nå jevnt over er mer robust enn før. Vi har ikke data på om bedringen skyldes bedre smoltkvalitet, eller om transport og håndteringsmetodene ved utsett har blitt mer skånsom. Smoltstørrelsen (som rapportert ved slutt på kalendermåneden med utsett) økte gradvis fra under 110 g i gjennomsnitt før 2012 til 160 g i 2019, noe som kan ha gjort fisken mer robust. En annen medvirkende forklaring på denne nedgangen kan også være introduksjon av lakseegg fra stamfisk selektert for IPN-resistens fra 2010-11, som har ført til at forekomst av denne sykdommen har blitt redusert med mer enn 90%. I 2011-12 gikk rapporterte lokaliteter med IPN kraftig ned, noe som fortsatte ytterligere i 2013, og IPN har ligget på et relativt lavt nivå siden (Sommerset m.fl. 2020; Fig 7.6).
Figur 7.3
Dødelighet samme kalendermåned som utsett ved temperatur mellom 0 og 4 °C, 4 og 8 °C, 8 og 12 °C, 12 og 16 °C og 16 og 20 °C for laks satt ut i første halvdel av året («vårutsett»), andre halvdel av året («høstutsett») for årene 2005-2011, 2013-18 og 2019.
7.2.3 - Dødelighet, stress og skader under avlusning
Selv om dødeligheten første kalendermåned etter utsett har gått ned (Fig 7.3), har dødeligheten på større og eldre fisk dessverre vært økende. Selv om det rapporteres tilfeller der stormer, predatorer, forstyrrelser fra røkting og utstyr har stresset fisken og ført til skader, antar vi at dette har relativt lite avtrykk på fiskevelferden for oppdrettsfisken som helhet. Analyse av data fra databasen til Fiskeridirektoratet sammenstilt med innrapporterte avlusingsoperasjoner til Mattilsynet viser tydelig at det er stor risiko for økt dødelighet under avlusingsoperasjoner (Fig 7.4). Etter 2012 har metodene for lusebehandling endret seg fra badebehandling med pyretroider eller organfosfater, via hydrogenperoksid til hovedsakelig ikke-medikamentell-behandling (IMM) (Figur 7.4A). Figuren viser også en økning i antall innrapporterte behandlinger fra 2016. Dette samvarierer med endring i avlusingsmetode til IMM, men også med at Mattilsynet desember 2015 kom med en presisering om at en kan behandle enkeltmerder i anlegget med mye lus for å holde smittepresset nede og ha en mer målrettet tilnærming til lusebehandling. Før denne presiseringen var mer enn 50 % av innrapporteringene for hele anlegget, mens har siden ligget på 30 - 40%.
Figur 7.4 A) Avlusingsoperasjoner rapportert til Mattilsynet 2012-2019. Rød = termisk, blå = mekanisk, grønn = hydrogenperoksid bad, grå = andre bademidler. B) Innrapportert månedlig dødelighet generelt (grå åpne punkt) vs. innrapportert månedlig dødelighet for lokalitet som har rapportert avlusing (svarte punkt). Heltrukken linje = generell årlig trend for alle lokaliteter (grå linje) og lokaliteter som har gjennomført avlusing gitte måned (svart linje).
I 2012 var 95 % av alle innrapporterte avlusingsoperasjoner kjemisk badebehandling, 5 % badebehandling med hydrogenperoksid og i tillegg noen få mekaniske avlusinger, mens i 2019 var nær 60 % termisk, 26 % mekanisk, og kun 12 % kjemisk badebehandling og 3 % behandling med hydrogenperoksid. Når vi sammenligner dødelighet i merder med og uten lusebehandling ser vi en tydelig økning i forbindelse med innføring av ikke-medikamentelle-metoder (IMM) fra 2016 (Fig 7.4B). Rapportert dødelighet i avlusingsmåned gikk noe ned fra 2018, som trolig skyldes forbedring av teknologi og mer erfaring med bruk av IMM.
I 2019 var nær 75 % av alle innrapporterte avlusingsoperasjoner i produksjonsområde 2-5 termiske, mens det var en mer jevn fordeling mellom mekanisk og termisk fra produksjonsområde 6 og nordover. Fra produksjonsområde 9 var det også et vesentlig innslag av hydrogenperoksid. Ved å sammenligne den månedlige dødeligheten for lokaliteter uten innrapportert avlusing med lokaliteter med gjennomført avlusing får vi et bilde av hvor risikofylte de ulike avlusingsmetodene er for fisken (Figur 7.5). Kjemisk badebehandling gav ofte økt dødelighet, men det er også mange måneder med tilsvarende dødelighet som normalen. Avlusing med hydrogenperoksid gav tilsvarende dødelighet som ved kjemisk badebehandling, men dødeligheten har til tider vært mer variabel og svært høy og denne metoden må karakteriseres som mer risikofylt. Etter at mekanisk behandling ble tatt i utstrakt bruk (fra og med 2016) ligger nesten alle punktene over normalen. Tilsvarende ser vi for termisk avlusing. Legg imidlertid merke til at for mekanisk og termisk avlusing så har dødeligheten gått ned i 2019. Dette kan være et resultat av økt oppmerksomhet rundt dødelighet i forbindelse med avlusing, optimalisering av utstyr og prosedyrer, og presiseringen fra Mattilsynet om at fiskehelsepersonell skal vurdere forsvarligheten ved avlusingen. En medvirkende årsak til nedgangen i dødelighet kan også være bløggebåter som ligger standby og bistår med å fjerne døende fisk etter avlusing.
Figur 7.5
Sammenligning av dødelighet per måneder for lokaliteter som ikke har rapportert en avlusingsoperasjon vs. dødelighet for lokaliteter som har rapportert kjemisk badebehandling, avlusing med hydrogenperoksid, mekanisk avlusing eller termisk avlusing. Analysen er gjennomført basert på produksjonsområde 6-12 for laks over 1 kg. Produksjonsområder 2-5 er utelatt fordi disse er sterkt dominert av termisk avlusing. Filter: Laks over 1 kg, produksjonsområde 6-12.
Effekt på fiskevelferd ved termisk avlusing ble vurdert av uavhengige forskningsinstitutt under utviklingen av begge de to meste brukte systemene i næringen (Grøntvedt mfl. 2015, Roth 2016). Til tross for dette er det tydelig at termisk avlusing i mange tilfeller kan gi høy dødelighet (Overton mfl. 2018), og det har blitt rapportert om blødninger og sårskader (Poppe mfl. 2018) og fisk som kolliderer i vegger i behandlingskammeret (Hjeltnes mfl. 2019). På oppdrag fra Mattilsynet har Havforskningsinstituttet sammen med Veterinærinstituttet avdekket at laks som blir eksponert for temperaturer over 28 °C viser tydelig flukt- og smerteatferd (Nilsson mfl. 2019). For mekanisk avlusing blir det rapportert om at både spylingen og børsting kan påføre fisken skjelltap og blødninger (Holan mfl. 2017, Erikson mfl. 2019, Sommerset mfl. 2020). Effekt av skjelltap, og når fisken er mest sårbar for dette er et område hvor det mangler mye viktig grunninformasjon og hvor det bør fremskaffes ny kunnskap (Noble mfl. 2018). Generelt synes det som om det arbeides fortløpende med å forbedre de ulike teknologiene for avlusing, og dataene fra Figur 7.5 tyder på at det har vært en vesentlig forbedring siden de først ble introdusert, men at det fortsatt er forbundet med økt fiskedødelighet å gjennomføre en avlusing.
7.2.4 - Sykdom og parasitter
Fravær av alvorlig sykdom og skadelige mengder parasitter er en forutsetning for god velferd. Særlig infeksjoner som er smertefulle og langvarige og fører til nedsatt organfunksjon medfører sterkt redusert velferd. Det rapporteres inn en rekke sykdommer for laksefisk i oppdrett, og med unntak av IPN er det lite som tyder på at sykdomsbildet har bedret seg de siste årene. Blant virussykdommene er Pankreassykdom (PD), hjerte og skjelettmuskelbetennelse (HSMB) og hjertesprekk (CMS) de hyppigst rapporterte (Figur 7.6). Symptomer på PD inkluderer tap av appetitt, muskelskader (inkludert spiserørmuskulatur), celledød i bukspyttkjertelen (pankreas) og redusert produksjon av fordøyelsesenzymer, endret adferd (fisken stiller seg i overflaten) og økt dødelighet. Særlig PD forårsaket av SAV3, som er vanligst på Vestlandet, er forbundet med økt dødelighet. Fisk med SAV3 nord for produksjonsområde 5 skal destrueres, eller flyttes sørover (Forskrift om tiltak for å forebygge, begrense og bekjempe PD hos akvakulturdyr ). For produksjonsområde 6-7 er det i hovedsak SAV2. I 2019 ble det ikke registrert noen forekomster av SAV3 i produksjonsområde 3, men det ble registrert enkelttilfeller av SAV2 (Sommerset mfl. 2020). Ifølge Fiskehelserapporten (Sommerset mfl. 2020) rapporteres HSMB fra hele landet, med de siste årene med en overvekt i produksjonsområde 9 -11 . Både PD, HSMB og CMS gir varierende dødelighet, og dødelighet forårsakes gjerne av stress i forbindelse med håndtering.
Figur 7.6 Registrert forekomst av ulike virussykdommer hos laks i oppdrett i perioden 2005 – 2019 diagnostisert av Veterinærinstituttet (Sommerset mfl. 2020). I tillegg er det diagnostisert sykdommer i private laboratorier, og en må anta at mange forekomster ikke er registrert i denne statistikken.
Siden PD er en meldepliktig sykdom, har vi gode data for denne, og en relativ stor andel av anleggene sør for PO8 rapporterer om PD infeksjoner (Figur 7.7A). Ved å plotte dødelighet for anlegg i produksjonsområder fri for PD, for anlegg uten påvist PD i produksjonsområder med PD og for anlegg med påvist PD kommer risikoen for økt dødelighet ved pankreassykdom tydelig frem (Figur 7.7B). Blant bakteriesykdommene er spesielt vintersår et stort velferdsproblem. Vintersår forekommer langs hele kysten, men særlig i Nord-Norge. En grundig gjennomgang av status for de ulike fiskesykdommene finnes i Fiskehelserapporten 2019 (Sommerset mfl. 2020).
Figur 7.7 A) Prosentandel aktive lokaliteter med PD registrert i 2017, 2018 og 2019 per produksjonsområde. I produksjonsområde 2-5 dominerer SAV3 (røde stolper), mens i produksjonsområde 6-7 destrueres eller flyttes lokaliteter med SAV3. Her vil det derfor være i hovedsak SAV2 som blir stående i sjø (hvite stolper). B) Snitt månedlig dødelighet for produksjonsområde 6-12 i måneder uten rapportert avlusingsoperasjon (åpne sirkler), snitt månedlig dødelighet i måneder uten avlusingsoperasjon for lokaliteter i produksjonsområde 2-5 uten påvist PD (svarte sirkler) og med påvist PD (røde sirkler) i gitt kalenderår. Filter: Laks > 500 g.
7.2.5 - Skadelig vannmiljø
Vannet flyter fritt gjennom merdene og utskiftingsgraden varierer med vannstrøm på ulike dyp, maskevidde og begroing på merdveggene. En har derfor ingen kontroll eller rensing av vannet som fisken blir eksponert for. Vanntemperaturen varierer sterkt med sesong, men i perioder, spesielt på vår og sommer, kan det være store temperaturforskjeller mellom ulike dyp. Det kan oppstå høye fisketettheter i de foretrukne temperaturområdene (Oppedal mfl. 2011). Svært lave eller høye temperaturer er særlig problematisk når fisken er stresset, og i tiden etter sjøutsett. Lave temperaturer øker også risikoen for visse sårsykdommer som vintersår (forårsaket av Moritella viscosa) og sår forårsaket av Tenacibaculum spp. I figur 7.8 vises antall lokaliteter som har registrert svært høye (>18°C) eller svært lave temperaturer (<4°C). Høye temperaturer forekommer relativt sjeldent og bare fra produksjonsområde 6 og sørover, mens svært kalde perioder forekommer naturlig nok mest i nord, men i 2019 var det også svært kalde perioder på Vestlandet (særlig PO2).
Vannmiljøet blir også påvirket av fiskens oksygenforbruk og utslipp av avfallsstoffer og ekskrementer. Mengden tilgjengelig oksygen i merdene blir ikke rapportert, men studier viser at det kan oppstå perioder med lave oksygenverdider i merder med mye fisk i perioder med lav vannstrøm. Risikoperioder er her perioder hvor tidevannet snur, og områder med tidevannsdrevet strøm er mest utsatt. De siste årene har en tatt i bruk såkalte «luseskjørt», som er lusetette presenninger som dekker merdene i de øverste 4-10 meterne for å beskytte mot lakselus. Disse reduserer også vannutskiftingen og det kan oppstå lave oksygenverdier hvis mye fisk står innenfor skjørtene (Stien mfl. 2012, 2018). Opphopning av avfallsstoffer fra fisken, som ammoniakk og CO2, kan føre til skadelige konsentrasjoner i kar, men i åpne merder blir ikke konsentrasjonene høye nok til å bli et problem.
Figur 7.8 Andel lokaliteter med rapportert månedstemperatur under 4 °C A) eller over 18 °C B). Hvert produksjonsområde har en søyle for henholdsvis 2017, 2018 og 2019.
Merdene holdes rene ved å bruke spylemaskiner som fjerner alger og organismer som vokser på notveggene. Omfanget av dette har økt for å gjøre rensefisken mer effektive ved at de har mindre alternativ mat tilgjengelig. Ulempen er at algefragmenter og særlig nesleceller fra hydroider kommer ut i merdene og kan skade gjellene til fisken.
Vi vet lite om hvilke organismer som flyter gjennom merdene og hvilke effekter disse har på fisken. Vi vet imidlertid at det er en svært høy forekomst av gjelleskader. Fiskehelsetjenestene melder om utfordringer med algeoppblomstringer på seinsommer og tidlig høst. Når det er høye algebiomasser kan algene skade gjellene direkte ved mekanisk påvirkning, og noen algetyper er direkte giftige og kan føre til fiskedød også ved lave tettheter. Disse algegruppene har skjøre celler, som kan løse seg opp når de treffer fiskegjellene og slippe ut skadelige stoffer. Eksempler på slike arter i våre farvann er Chrysochromulina leadbeateri som drepte mye laks i Nordland i 2019, Prymnesium parvum, Pseudochatonellafarcimen. Andre mindre studerte arter er alger fra slektene Dictyocha and Vicicitus. Alle disse artene skader gjellene ved at en får nekrotiske endringer i gjellefilamentene. Den mekanistiske forklaringen bak fiskedød på grunn av alger er lite utforsket, men det eksisterer mange teorier. Disse inkluderer frie fettsyrer, reaktive oksygentyper og spesielle algetoksiner. Virkelige algegifter er bare kjent fra noen få arter som f.eks. prymnesiner fra P. parvum eller brevetoxiner fra Karenia brevis.
7.2.6 - Dødelighet per produksjonsområde
Gjennomsnittlig månedlig dødeligheten er relativt stabil fra år til år for de ulike produksjonsområdene (Figur 7.9). Dødeligheten kan variere noe fra år til år på grunn av variasjoner mellom generasjoner og variasjon i hvilken størrelsesfase størsteparten av fisken i det aktuelle produksjonsområdet er. Typisk så er dødeligheten høyest på Vestlandet og lavest i Nord-Trøndelag og Helgeland, hvor gjennomsnittlig månedlig dødeligheten kan være opp til 3 ganger høyere på Vestlandet. Dette skyldes til dels høyere forekomster av sykdom (særlig PD) og mer lakselusbehandling i sør, som igjen kan ha sammenheng med høyere temperaturer og at anleggene ligger tettere. Fjoråret 2019 skiller seg ut med uvanlig høye verdier for PO9 og 10, som skyldes at dødeligheten for disse produksjonsområdene var over 12 % i mai i forbindelse med oppblomstringen av giftalger. Andre produksjonsområder med nedadgående dødelighetstrend inkluderer PO2 (Boknafjorden) og PO12 (Vestfinnmark). Dette kan tyde på at oppdretterne i disse PO-ene har klart å forbedre velferden til fisken i merdene. En medvirkende faktor kan være at i forhold til mange andre PO-er er disse PO-ene relativt lite påvirket av tilstøtende PO-er.
Figur 7.9
Gjennomsnittlig prosent døde oppdrettslaks per måned for årene 2017, 2018 og 2019 for produksjonsområde 2 til 12. PO1 (Sørlandet) og 13 (Østinnmark) er ikke inkludert siden det her er svært få lokaliteter å basere beregningene på.
7.2.7 - Preventive metoder mot lakselus og nye driftsformer
Norsk oppdrettsnæring er under stadig utvikling. Særlig lakselus er en driver av innovasjon og det testes i dag ut flere ulike system for å unngå påslag av lakselus. Dette inkluderer produksjon av post-smolt i landbaserte resirkuleringsanlegg, semi-lukkede merder i sjø, snorkelmerder, merder med skjørt og nedsenkede merder. Alle disse nye driftsformene medfører nye livsbetingelser for fisken som kan ha konsekvenser for dyrevelferden.
Luseskjørt: Dette er et relativt enkelt tiltak hvor en lusetett presenning blir plassert rundt de øverste meterne av merden for å sperre luselarvene ute. Luseskjørt brukes på mange oppdrettsanlegg og det finnes flere vitenskapelige publikasjoner på bruk av luseskjørt og effekt fra luseskjørt på vannkvalitet og fiskevelferd (Frank mfl. 2015, Lien mfl. 2014, Grøntvedt mfl. 2018, Stien mfl. 2012, 2018). Studiene viser at luseskjørt har en reduserende effekt på lusepåslag, men at dette varierer mellom lokaliteter og reduseres ved høy strøm. Studiene viser også at luseskjørt ikke trenger å gi negative effekter på fisken, men at det kan oppstå lave oksygenforhold og dårlig vannkvalitet i skjørtvolumet (Stien mfl. 2012). I Veterinærinstituttet sin spørreundersøkelse til fiskehelsepersonell for 2019 var det flere som rapporterte at de hadde observert utfordringer med luseskjørt knyttet til lave oksygenverdier og negativ påvirkning på gjellehelse (Sommerset mfl. 2020). Det advares også om at lav vannutskifting kan føre til at fisken utsettes for høye konsentrasjoner av gift fra not-impregnering (Sommerset mfl. 2020).
Nedsenket merd: Det har vært forsket på å benytte nedsenkede merder for å oppdrette laks i flere tiår og det finnes mange vitenskapelige publikasjoner på dette (f.eks. Dempster mfl. 2009, Korsøen mfl. 2012, Glaropoulos mfl. 2019, Oppedal mfl. 2020). Å senke ned laksen har imidlertid flere utfordringer, både rent praktisk og for laksen selv. Laksefisk har en åpen (fysostom) svømmeblære og må gå til overflaten for å svelge luft for å fylle svømmeblæren nok til å opprettholde nøytral oppdrift der den svømmer. For å løse dette problemet har det blitt forsket på undervannsmerder med luftlommer i taket, regelmessig løfting av undervannsmerden til overflaten og bruk av luftbobler. Selv om det nå er flere som tester ut ulike prinsipper for å sikre at laksen klare å fylle svømmeblæren i dypet må denne teknologien fortsatt sies å være på utprøvingsstadiet.
Snorkelmerd: Snorkelmerd, eller tubenot, er i prinsippet en nedsenket merd, men hvor der er en lusetett tunell («snorkel») fra merdvolumet til overflaten hvor laksen kan svømme opp for å fylle svømmeblæren i overflaten. Det er vist at laksen mestrer å svømme opp snorkelen og fylle luft i overflaten for deretter å svømme ned igjen i merdvolumet (Stien mfl. 2016, Oppedal mfl. 2017). Effekten snorkelen har mot lusepåslag avhenger av hvor dyp snorkelen er, dyp av eventuelt brakkvanslag og strømforhold (Samsing mfl. 2016, Oppedal mfl. 2019). Det har også vært tilfeller med økt innslag av AGD (Neoparamoeba perurans) hos fisk i snorkelmerd, men det har blitt vist at dette kan behandles ved å tilsette ferskvann i de øverste meterne av snorkelen. Snorkelmerd er mer krevende å drifte og er fortsatt bare i begrenset bruk i næringen til tross for til dels svært god effekt mot lusepåslag (Geitung mfl. 2019).
Semi-lukket anlegg: Dette er enheter i sjøen med en tett barriere mellom vannvolumet med fisken og omgivelsene. Det kalles semi-lukket fordi det fortsatt er kontakt med det omliggende vannet via vannet som pumpes inn fra dypet, og vannet som pumpes ut. Slike enheter gir en effektiv beskyttelse mot lakselus (Nilsen mfl. 2017), men det er også krevende teknologi der alt må fungere som det skal til enhver tid for at fiskevelferd skal være sikret (Holan mfl. 2017, Noble mfl. 2018). På grunn av høye kostnader blir fisken også holdt i høyere tettheter i semi-lukkede anlegg kontra åpne sjømerder (Nilsen 2019) og en må derfor hele tiden overvåke og tilsette oksygen (Holan mfl. 2017). En rekke ulike former for semi-lukkede anlegg er under uttesting, men bortsatt fra noen få enkeltstudier vet vi lite om risiko for teknisk eller menneskelig svikt når de kommer i mer allmenn bruk.
Stor post-smolt: Produksjon av post-smolt i semi-lukkede anlegg i sjø (opptil 1 kg), gjennomstrømmingsanlegg eller i resirkuleringsanlegg på land er en måte å redusere tiden laksen er i sjøen, og dermed redusere faren for lusepåslag og påfølgende avlusing. Alle disse produksjonsformene er sårbare for uregelmessigheter i driften og oppblomstringer av patogener som kan være katastrofale for laksen, i tillegg til redusert vannkvalitet og velferdsproblemer knyttet til dette (Holan mfl. 2017, Noble mfl. 2018, Sommerset mfl. 2020). Det kan også være velferdsutfordringer i forbindelse med transport av fisken til sjømerdene og i overgangen fra et kontrollert karmiljø til et mer eksponert miljø i åpne merder.
Eksponert oppdrett: Ved å flytte merdene til havs kan det bli lenger mellom lokalitetene og mindre lusesmitte. Sterk strøm og høye bølger kan imidlertid skape problemer for laksen (Hvas og Oppedal 2017), og i enda større grad for rensefisk (Hvas mfl. 2018, Yuen mfl. 2019, Jónsdóttir mfl. 2019). Det er også flere logistiske utfordringer, og lange perioder med dårlig vær kan hindre oppdretter adgang til anlegget. Det utvikles nå systemer for oppdrett til havs, men disse er fortsatt under utprøving og det er foreløpig lite erfaringsdata tilgjengelig.
7.2.8 - Triploid laks
Triploid laks er steril og har blitt tatt i bruk som et miljøtiltak i forbindelse med «grønne tillatelser» for å hindre genetisk påvirkning av ville laksestammer ved en eventuell rømning. Triploid laks har økt forekomst av deformiteter og katarakt, men dette kan motvirkes ved å inkubere eggene ved lave temperatur og ved å tilsette mer fosfor og histidin til fôret. Havforskningsinstituttet har fulgt uttesting av triploid laks i flere samarbeidsprosjekt med næringen (Kjøglum mfl. 2016, Stien mfl. 2020). Resultatene fra disse oppfølgingene viser at triploid laks er mer sårbar for infeksjoner som resulterer i sår og høy dødelighet, men det har også vært flere utsett der den triploide laksen har oppnådd tilsvarende eller bedre fiskevelferd som fisken i referansemerder med diploid («normal») laks. Typisk for disse utsettene er at laksen har blitt satt ut i sjø om våren, mens for triploid laks satt ut om sommeren har det vært høyt innslag av parvikapsulose, og triploid laks satt ut om høsten har vært plaget av Tenacibaculum («munnråte») og vintersår.
7.3 - Velferd hos rensefisk
Rognkjeks og flere arter leppefisk blir benyttet som rensefisk for å holde nede tallet på lakse- og skottelus på oppdrettslaks og -ørret (Imsland mfl. 2014). Bruken av rensefisk har steget kraftig siden 2008 og antall satt ut i merdene var kommet opp i nærmere 60 millioner i 2019 (Figur 7.10A). Rognkjeks og leppefisk står langt fra laks og ørret i både miljøbehov, svømmeevne og biologi, og siden oppdrettsmerdenes design og lokaliseringen er tilpasset laksefisk er de mindre egnet til å dekke rensefiskenes miljøbehov. Leppefiskene er tilpasset et relativt rolig liv mellom klipper, steiner og alger nært land, og miljøpreferanser og -tålegrenser for leppefisk varierer med både art og størrelse.. Rognkjeksen vandrer også ut i havet, men voksne individ finnes vanligvis dypere 50 m og foretrekker kaldt vann (Powell mfl. 2018). Når det gjelder svømmekapasitet skiller rensefiskartene seg mye fra laks og ørret. Både rognkjeks og leppefiskartene har dårlig kapasitet for å motstå sterk strøm over tid, mens laks kan tåle betydelig høyere hastigheter (Remen mfl. 2016, Hvas mfl. 2018, Yuen mfl. 2019). Da rensefisken vanligvis er av betydelig mindre størrelse enn laksen gjør den relative strømhastigheten (kroppslengde/s) forskjellen i toleranse for vannstrøm enda større. Selv om rognkjeks til en viss grad kan beskytte seg mot sterk strøm ved å bruke «sugekoppen» under buken hvis den har substrat til dette, gjelder dette bare opp til et visst nivå (Hvas mfl. 2018). Vannmiljøet i en laksemerd er i mange tilfeller ikke innenfor toleranseområdet til rensefisk. Det vil være sesongmessige variasjoner som i perioder gir for lav temperatur for leppefisk eller for høy temperatur for rognkjeks. På eksponerte lokaliteter kan vannstrømmen være for sterk for rensefisk, og hvis den ikke kommer seg i ly for strømmen vil den ende opp i notveggen.
Figur 7.10
A) Utsett av rensefisk rapportert til Fiskeridirektoratet. B) Prosent månedlig dødelighet hos rensefisk rapportert til Fiskeridirektoratet sin database i 2019.
Det er hovedsakelig villfanget bergnebb, grønngylt og berggylt, samt oppdrettet berggylt og rognkjeks som brukes som rensefisk. Den villfangede leppefisken blir utsatt for stress og skader under fiske og transport, så overlevelse ved utsett er avhengig av i hvilken kondisjon fisken er i når den kommer fram til anlegget. Noe fisk blir fanget lokalt og blir overført til merdene samme dag som de fiskes, mens andre blir lagret i oppsamlingstanker og kan bli transportert i bil over lange avstander, for eksempel fra Sørlandet til Trøndelag. Den oppdrettede rensefisken er utsatt for sykdommer i klekkeriene og kan være svekket før transport. Ifølge oppdretterne selv dør mer enn 10% av villfanget leppefisk første måned i merdene. Særlig grønngylt har høy utsettdødelighet, mens utsettdødeligheten er lavere for oppdrettet berggylt og oppdrettet rognkjeks (Stien mfl. 2020). Dataene fra Fiskeridirektoratets database for 2019 viser de samme tendensene (Figur 7.11). Dessverre har denne databasen hatt en samlekategori ‘Rensefisk oppdrett’ som gjør det vanskelig å skille mellom artene, men der art er oppgitt er det en tydelig forskjell i dødelighet første kalendermåned i sjø mellom leppefisk-artene og rognkjeks (Figur 7.11BC).
Figur 7.11 Dødelighet samme kalendermåned som utsett i 2019 ved temperaturer mellom 0 og 4 °C, 4 og 8 °C, 8 og 12 °C, 12 og 16 °C og 16 og 20 °C der art er oppgitt som A) ‘Rensefisk (oppdrett)’, B) en leppefisk art B) og C) som rognkjeks.
Mattilsynet gjennomførte en nasjonal tilsynskampanje med helse og velferd hos rensefisk 2018-2019 (Stien mfl. 2020). Som del av denne kampanjen utarbeidet Mattilsynet en spørreundersøkelse om fiskevelferd og dødelighet hos rensefisk. Denne spørreundersøkelsen viste at 42 % av rensefisken som ble satt ut i merdene ble registrert som døde i merden. Forfatterne påpeker imidlertid at den reelle dødeligheten sannsynligvis er vesentlig høyere siden død rensefisk ofte ikke faller ned i dødfiskhåven, men blir fanget i notveggen, hvor de råtner, går i oppløsning, eller blir spist av predatorer på utsiden. Rensefiskene er utsatt for en rekke sykdommer og parasitter, samtidig som stress, uegnede miljøforhold, håndtering og skader er årsaker til høy dødelighet (Stien mfl. 2020). Siden våren 2019 har oppdretterne rapportert inn antall rensefisk i merdene og antall døde rensefisk siste måned på samme måte som for oppdrettslaks. Disse dataene viser at mellom 5 og 18 % av rensefisken som står i merdene blir rapportert som dødfisk hver måned (Figur 7.10), noe som er rundt 10 ganger høyere enn tilsvarende gjennomsnittstall for laks (Figur 7.2). Dette viser at på tross av at oppdretterne gjør en stor innsats for å tilpasse miljøet i merdene med skjulesteder og spesialfôr til rensefisken, så er det tydelig at disse artene ikke mestrer livet i en laksemerd.
7.3.1 - Håndteringstress og dødelighet hos rensefisk
Under lusebehandling, notskift og andre operasjoner må fisken som regel tvinges mot eller over overflaten, hvilket medfører trykkreduksjon. Flytting med vakuumpumpe medfører ytterligere trykkreduksjon. Når trykket minker øker volumet på gassen i svømmeblæren. Laksen sin åpne svømmeblære gjør at den tåler rask reduksjon i trykk, da overflødig gass vil lekke ut. Leppefisk har lukket svømmeblære, og reduksjon av gassmengden må skje fysiologisk (via blodbanen) over tid. Ved stort trykkfall på kort tid risikerer derfor svømmeblæren å sprekke, eller fisken kan bli flyte opp og miste oppdriftskontrollen, dvs. uten mulighet for å komme seg ned på dypere vann for å utjevne trykket. Rognkjeks har ikke svømmeblære og vil derfor være mindre påvirket av selve trykkreduksjonen.
Ifølge Akvakulturdriftsforskriften §28 skal rensefisk sorteres ut før det utføres operasjoner som kan føre til belastning, skade og unødvendig påkjenning på rensefisken. På bakgrunn av dette har Mattilsynet presisert at rensefisken må utsorteres før dagens metoder for mekanisk og termisk avlusing. Unntaket er hvis rensefisken selv behøver avlusing. Mange lokaliteter, særlig i Finnmark, har fortalt om til tider store mengder skottelus på rognkjeks og at dette har ført til dødelighet. Noen av de ikke-medikamentelle metodene hevdes å være skånsom for rognkjeks, men det finnes lite dokumentasjonen av velferdseffekter av ikke-medikamentell avlusing av rensefisk. Foreløpig er det også et alternativ å i stedet behandle mot skottelus via fôret eller ved kjemisk badebehandling siden det foreløpig ikke har vært den samme resistensutviklingen for skottelus. Å fiske ut rensefisken fra merden, før laksen skal trenges og gjennomgå avlusing, er krevende. Noen mekaniske systemer har derfor inkludert avsilere for å sortere ut rensefisen, men det er lite dokumentasjon på hvor godt disse virker i praksis og hvor stressende dette er for fisken. Selv om noen oppdrettere forteller at de lykkes godt med utfiskingen anser vi det som sannsynlig at mye av rensefisken i praksis gjennomgår avlusing sammen med laksen. Rognkjeks er ansett som en relativt hardfør fisk, men leppefisk er betydelig mer sårbar. Samtidig kan det være lettere å utsortere den lille smale leppefisken før den går inn i avlusingssystemet enn den mer tykke rognkjeksen.
7.4 - Fiskeernæring og velferd
For at fisk i oppdrett skal ha god velferd må de ha tilgang til næringsrik og sunn mat som tilfredsstiller deres fysiologiske behov gjennom hele produksjonen. Dette betyr at de må få mat av en størrelse den kan spise, riktig sammensetning av næringsstoffer, og i tilstrekkelige mengder og riktig fordeling til at all fisk i oppdrettsmerden får tilgang til mat. Tidligere ble fôret i hovedsak produsert av marine råvarer og bestod essensielt av det samme som oppdrettsfisken spiser i vill tilstand. Det siste tiåret har det imidlertid på grunn av råvaremangel vært en overgang til mer bruk av planteproteiner og –oljer (Aas mfl. 2019). Nye råvarer har annen sammensetning av næringsstoffer enn de marine råvarene, både i mengde og kjemisk form. Det er derfor viktig at behovet for de enkelte næringsstoffer oppfylles gjennom riktig sammensetning av råstoff og eventuelle tilsetninger av spesielle næringsstoff som det lett kan blir for lite av ved bruk av planter eller andre fôrråstoff (www.arraina.eu). Planteråstoff har også andre risiki enn marine ressurser med tanke på uønskete stoffer, for eksempel plantevernmidler, soppgifter og naturlige antinæringsstoff som kan gjøre enkelte næringsstoff lite tilgjengelig (Glencross mfl. 2020). Rent historisk har de fleste store endringer i fôrmidler til oppdrettsfisk medført uønskete innslag av velferdslidelser som beindeformiteter, katarakt og redusert motstand mot stress og infeksiøse sykdommer (Waagbø 2008). Dette er ofte knyttet opp mot interaksjoner mellom ernæring og miljøbetingelser i et intensivt oppdrett. Ved bruk av planteråstoff har man sett økte innslag av beindeformiteter og endret motstand mot stress og infeksiøse sykdommer (Waagbø 2006; 2008; Bæverfjord mfl. 2019). Dette har vært knyttet opp mot redusert mengde og tilgjengelighet av fosfor og andre mineraler, samt endret balanse av essensielle fettsyrer med relativt mer omega-6 fettsyrer og mindre omega-3 fettsyrer i fôret. Det er usikkert hvor stor betydning de reelle endringene i uønskete stoffer har å si for fiskens velferd. Her vil store variasjoner i innhold kunne øke usikkerheten.
Perioder med sulting forekommer i løpet av produksjonen for å ivareta fiskens velferd under ulik håndtering, vaksinasjon, behandlinger og transport. Dette er knyttet til utfordringer med vannkvalitet under transport og fiskens respons til fysisk stress. For eksempel skal sulting før transport og slakt ivareta vannkvaliteten under lukket transport (mindre ammoniumutskilling, oksygenforbruk, akkumulert karbondioksid), øke fisken kapasitet for å håndtere trenging og stress, og ivareta hygieniske forhold med tom tarm ved slakt.
For lavt innhold og tilgjengelighet av beinmineralet fosfor i fôret fører til beindeformiteter, spesielt hos rasktvoksende laks og triploid laks. Fosfor kan tilsettes fôret, men man må samtidig ha kontroll over de lokale miljøeffektene som gir seg utslag i eutrofiering av høyt fosfor i fôret. Mange mineraler har etter Matloven og Forskrift om fôrvarer en øvre grenseverdi satt av nasjonale og internasjonale myndigheter (Mattilsynet og EFSA). For mineralet sink ble denne grenseverdien nylig redusert fra 200 til 180 mg/kg for laksefisk, hovedsakelig med bakgrunn i miljøhensyn. Nyere forskning har på den annen side vist et økt behov for sink opp mot grenseverdien i stor laks gitt plantebaserte fôr i sjøvann. Dette illustrerer et dilemma mellom å dekke fiskens behov for sink og samtidig beskytte det lokale miljøet mot unødig utslipp. En ideell mer bærekraftig løsning ligger i å gjøre fôrets og fôringrediensenes naturlige innhold av sink mer tilgjengelig eller benytte mer tilgjengelige former for sink i fôret. Et studium med økende menger av mineralet mangan, tilsatt i uorganisk eller organisk form til plantebaserte fôr viste at organisk mangan hadde dobbelt så god tilgjengelig som uorganisk mangan (Prabhu mfl. 2019). Med tanke på fôrmengdene som benyttes i norsk lakseoppdrett per år (ca 1,8 millioner tonn) og gjennomsnittlig innhold av mangan i norske fôr (47 mg/kg; Sele mfl. 2019), er potensialet en reduksjon i utslipp i norske fjorder fra ca. 80 til 32 tonn per år. Unødig høye nivå av mineraler i fôret har potensielt velferdsmessige utfordringer knyttet til interaksjoner på opptak og status med andre mineraler hos fisken, noe som kan føre til suboptimal ernæringsstatus.
Mangel på aminosyren histidin er satt i sammenheng med utvikling av katarakt (grå stær) hos oppdrettslaks (Waagbø og Remø 2020). Dette er knyttet opp til lave nivå av en histidinforbindelse (NAH) i laksens linse. I praktisk oppdrett vil laksesmolt gitt kommersielle fôr uten tilsetning av histidin ha økt risiko for utvikling av katarakt når temperaturen stiger, selv om behovet for vekst er dekket (Remø mfl. 2014). Regnbueørret gitt samme fôr er ikke tilsvarende sensitiv, mens triploid laks er mer sensitiv enn vanlig laks (Sambraus mfl. 2018). Det er vist at stor laks i sitt andre år i sjø også utvikler katarakt når temperaturen stiger dersom de ikke får tilskudd av histidin i fôret. Med alvorlig kataraktutvikling reduseres fôrinntaket og man risikerer sekundære lidelser og sykdommer. Ettersom tilsetning av histidin fordyrer fôret, blir det en avveiing for oppdretter om når og hvor lenge man bør gi histidinrikt fôr for å redusere kataraktutviklingen.
For å ha kunnskap om trygge øvre grenser av uønskede stoffer i dagens og fremtidens fiskefôr blir uønskede stoffer som kan påvirke fiskehelse og mattrygghet overvåket og studert. Nyere fokus har vært på pesticider og mykotoksiner fra planteråvarer som har blitt produsert og lagret under ugunstige forhold. Stoffene blir undersøkt både med tanke på direkte negative helseeffekter og samspillseffekter med andre gifter eller næringsstoff. Overvåkningen av fiskefôr i 2018 som Havforskningsinstituttet gjør for Mattilsynet viste ingen overskridelser av øvre grenseverdier for uønskete stoffer i fôr eller fôrmidler, verken for organiske eller uorganiske (Sele mfl. 2019). Noen av de undersøkte mykotoksinene har imidlertid ikke etablerte grenseverdier ennå. Risikoen for uønskete hendelser med kontaminanter og giftstoffer i fôr og fôrmidler ansees i dag som liten. Et økende innslag at soppgifter i fôr til landdyr kan endre denne statusen i fremtiden.
7.4.1 - Ernæring hos rensefisk
Av rensefiskene våre er det kun de artene det drives oppdrett av, rognkjeks og berggylt, hvor det forskes på hvilke ernæringskrav som oppfyller god velferd. Rognkjeks og leppefisk har behov for en annen sammensetning av fôret enn laks, og for disse finnes det egne kommersielle spesialfôr. Det er betydelig forskningsaktivitet for å optimalisere sammensetningen av fôret mot bedre velferd for rensefisk, tilpasset deres anatomi, fysiologi og helse.
Oppdrett av berggylt startet før rognkjeks derfor har en mer kunnskap om denne arten. Anatomi og fysiologi er svært ulik hos disse artene, og det er dermed vurdert at de har ulike ernæringsbehov. Rognkjeksen gyter store egg som gir godt utviklede larver. Disse er store nok til å startfôres på formulert fôr. Den har god appetitt og vokser raskt. Den gode appetitten gjør at den kan være utfordrerne å få den til å kun spise sitt fôr i laksemerden. Berggylten gyter små egg som gir små (4 mm) larver, disse trenger levendefôr i oppimot 2 mnd. Denne arten er vanskelig å få til å spise, så smakelighet på fôret et svært viktig. Dette gjør fôret svært dyrt og gir begrensinger med tanke på sammensetning.
Berggylt er en saktevoksende art (Dipper mfl. 1977) som i naturen beiter på gastropoder, dekapoder, echinodermer, bivalver og lignende (Figueiredo mfl. 2005). Den har velutviklede tenner i forkjeven, som gjør den velegnet til å skrape byttet av overflater. Deretter følger kraftige svelgbeinsplater med tenner for å knuse byttet før det svelges direkte ned i tarmen, da denne arten mangler mage. Tarmen er kraftig og veldig kort, kun 2/3 av fiskens totale lengde (Le mfl. 2019b). Selv om tarmen er kort fordøyes formulerte pellets av høy kvalitet godt, 74 % av protein og 50 % av fettet er absorbert i første ¼ av tarmen etter 4 timer. Innen fôret et passert gjennom tarmen etter 10 til 14 timer er henholdsvis 90 og 80 % protein og fett absorbert (Le mfl. 2019b). Fremre del av tarmen har en videre diameter enn midt og baktarm, ofte kalt bulbus. Bulbusen har vært referert til som pseudogaster, altså et reservoar for spist mat som kan bearbeides og fordøyes, men uten en ekte mages lave pH og pepsinogen drevne proteinfordøyelse. Det finnes ikke observasjoner som tilsier at bulbusen har en slik funksjon (Le mfl. 2019b, Lie mfl. 2018) Det berggylten har er et intrikat peristaltisk system som aktivt bringer fordøyelige deler av maten tilbake til første del av tarmen for optimal fordøyelse, mens ikke-fordøyelige komponenter evakueres raskere (Le mfl. 2019a). Med bakgrunn i kunnskap om adferd, fordøyelsessystemets fysiologi og anatomi er det anbefalt å fôre berggylten hyppig i alle livsstadier.
Vi vet lite om ernæringskrav til berggylt under larvestadiene. Praksis har vært å tilby anrikete rotatorier fram til 30 etter klekking dek, for deretter å tilby Artemia med noen dagers overlapp med rotatorier. Larvene tilvennes så et formulert fôr etter 50 dager ette klekking eller senere. Tidligere har en tilvendt larvene tørrfôr rett etter rotatoriefôring (Hansen mfl. 2013), men denne praksisen har en gått bort fra, da en periode med Artemia ser ut til å redusere deformiteter. Det har vært spekulert i om levendefôr var bedre for larvene da fôrdyrets egne fordøyelsesenzymer kunne bidra til å tilgjengelig gjøre næringsstoffer fra seg selv. Denne hypotesen er imidlertid forkastet. Det er derfor næringssammensetningen i fôrdyret som er overlegen det formulerte fôret (Hansen mfl. 2013). Grunnen til at formulert fôr har utilstrekkelig næringssammensetning er at overflate til volum ratio i så små partikler gjør at vannløselige næringsstoffer lekker. Det finnes nå bedre alternativer til rotatorier og Artemia på marked nå. Både nauplier av copepoden Arcatia tonsa og av rur har gitt positiv utvikling i vekst, overlevelse og deformiteter hos berggyltlarver (Øie mfl. 2015) (pers com Espen Grøtan ved MOWI). Dette er et felt det jobbes med. Det er utfordrende å få denne arten over på formulert fôr. Så langt er det nødvendig med rekemel i fôret for å få larvene til å vise interesse (Kousoulaki mfl. 2015). Så vidt vi vet i dag handler dette om smakelighet og ikke ernæringskrav.
Også større fisk trenger rekemel i fôret for å vise interesse. Det er generelt et problem med å opprettholde appetitten hos denne arten. Dette er et komplekst felt der en fortsatt har mye upløyd mark. Vi vet at evolusjonært tap av mage i denne arten har ført til tap av enkelte appetitt stimulerende gener. Dette har sannsynligvis ført til at andre stimuli, som lukt/smak er desto viktigere for appetitt (Lie mfl. 2018).
Berggyltens behov for protein er ca. 65 %, for lipid ca. 12 %, og den tåler karbohydratinnhold på 16 % av fôret (Hamre mfl. 2013). Behov for vitaminer og mineraler samt sammensetning av lipider undersøkes i det pågående prosjektet CleanFeed. Vi har også pågående forskning på bedre fôr til stamfisk.
Utfordringen i merd er å kunne presentere et smakelig fôr til fisken i skjulet. Her har det heldigvis kommet løsninger med fôrblokker som fisken liker å spise av, samtidig som disse fôrblokkene er utilgjengelig for laksen. De representerer derfor en mulighet for selektiv medisinering av leppefisk uten at laksen får i seg dette fôret.
Rognkjeks er en forholdsvis hurtigvoksende art. En årsak til den raske veksten er nok det lave tørrstoffinnholdet i fisken, hvor hele 87,1 ± 0,2 % av juvenil fisk er vann (Sæle mfl. upubliserte data). Dette er en strategi den har for å erstatte svømmeblæren (Davenport og Kjørsvik 1986). Den legger store egg som gir forholdsvis velutviklede larver (Brooker mfl. 2018). Disse kan startfôres med tørrfôr, men vanlig praksis i dag er å starte med Artemia de 20 til 30 første dagene etter klekking før tilvenning med tørrfôr. Dette er en art som er enkel å få til å spise. Når de når en vekt på 1,5 til 2,0 g har de en funksjonell mage og den utvikler et fordøyelsessystem med mage, pylorussekker og en lang tarm.
Rognkjeksens behov for makronæringsstoffer, vitaminer og mineraler undersøkes i det pågående prosjektet CleanFeed. Her legger vi fiskens helse til grunn for anbefalte næringsnivå. Preliminære undersøkelser tilsier at denne arten ikke skal ha like energitett fôr som det laks får. Erfaring tilsier at den er meget opportunistisk og vil ta til seg den føden som er lettest tilgjengelig. Den vil derfor like gjerne spise laksefôr som et spesialtilpasset fôr, om pelletstørrelsen tillater det. Dette er ikke bare uheldig for fiskens helse, men også for fiskens funksjon som rensefisk. Det ser ut som om rognkjeks beiter mindre på lus når den kan spise laksefôr. For best mulig velferd for fisken, er det anbefalt å fôre fisken med tilpasset fôr i eller ved skjul. Praksis der man bruker samme utfôringspunkt som til laks kan derfor være stressende for rognkjeksen, spesielt i lokaliteter med mye strøm.
7.5 - Referanser
Aas, Ø, Einum, S., Klemetsen, A, Skurdal, J. 2010 Atlantic salmon ecology. Blackwell Publishing DOI:10.1002/9781444327755
Aas TS, Ytrestøyl T, Åsgård T 2019. Utilisation of feed resources in the production of Atlantic salmon ( Salmo salar ) in Norway: An update for 2016. Aquaculture 15, 100216, 1-10.
Brooker AJ, Papadopoulou A, Gutierrez C, Rey S, Davie A, Migaud H 2018. Sustainable production and use of cleaner fish for the biological control of sea lice: recent advances and current challenges. Vet Rec 183, 383.
B æverfjord G, Antony JPP, Fjelldal PG, Albrektsen S, Hatlen B, Denstadli V, Ytteborg E, Takle H, Lock E, Berntssen MH, Lundebye A, Åsgård T. Waagbø R 2019. Mineral nutrition and bone health in salmonids. Rev Aquacult. 11, 740-765.
Davenport J, Kjørsvik E 1986. Buoyancy in the lumpsucker Cyclopterus lumpus . Journal of the Marine Biological Association of the United Kingdom 66, 159–174.
Dempster T, Korsøen Ø, Folkedal O, Juell J-E, Oppedal F 2009. Submergence of Atlantic salmon ( Salmo salar L.) in commercial scale sea-cages: A potential short-term solution to poor surface conditions. Aquaculture 288, 254-263.
Dipper FA, Bridges CR, Menz A 1977. Age, growth and feeding in the ballan wrasse Labrus bergylta Ascanius 1767. Journal of Fish Biology.
Erikson U, Solvang T, Schei M, Ag S, Strand A, Aalberg K 2019. Hydrolicer ‐ Utredning av system, stress og velferd ved avlusing. SINTEF, rapport nr 2018:01081
Frank K, Gansel LC, Lien AM, Birkevold J 2015. Effects of a Shielding Skirt for Prevention of Sea Lice on the Flow Past Stocked Salmon Fish Cages. Journal of Offshore Mechanics and Arctic Engineering 137, 011201.
Figueiredo M, Morato T, Barreiros JP, Afonso P, Santos RS 2005. Feeding ecology of the white seabream, Diplodus sargus , and the ballan wrasse, Labrus bergylta , in the Azores. Fisheries Research 75, 107–119.
Geitung L, Oppedal F, Stien LH, Dempster T, Karlsbakk E, Nola V, Wright DW 2019. Snorkel sea-cage technology decreases salmon louse infestation by 75% in a full-cycle commercial test. International Journal for Parasitology 49, 843-846.
Glaropoulos A, Stien LH, Folkedal O, Dempster T, Oppedal F 2019. Welfare, behaviour and feasibility of farming Atlantic salmon in submerged cages with weekly surface access to refill their swim bladders. Aquaculture 502, 332-337.
Glencross BD, Baily J, Berntssen MHG., Hardy R, MacKenzie S, Tocher DR 2020. Risk assessment of the use of alternative animal and plant raw material resources in aquaculture feeds. Reviews in Aquaculture, 12, 703–758.
Grøntvedt RN, Nerbøvik I-KG, Viljugrein H, Lillehaug A, Nilsen H, Gjerve A-G. 2015. Termisk avlusing av laksefisk – dokumentasjon av fiskevelferd og effekt, In: Veterinærinstituttets rapportserie nr. 13/2015. Veterinærinstituttet, Oslo.
Grøntvedt RN, Kristoffersen AB, Jansen PA 2018. Reduced exposure of farmed salmon to salmon louse ( Lepeophtheirus salmonis L.) infestation by use of plankton nets: Estimating the shielding effect. Aquaculture 495, 865-872.
Hamre K, Nordgreen A, Grotan E, Breck O 2013. A holistic approach to development of diets for Ballan wrasse ( Labrus berggylta ) - a new species in aquaculture. PeerJ 1, e99.
Hansen TW, Folkvord A, Grotan E, Sæle Ø 2013. Genetic ontogeny of pancreatic enzymes in Labrus bergylta larvae and the effect of feed type on enzyme activities and gene expression. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology 164, 176–184.
Hjeltnes B, Bang Jensen B, Bornø G, Haukaas A, Walde C S (red) 2019. Fiskehelserapporten 2018, utgitt av Veterinærinstituttet 2019
Holan AB, Roth B, Breiland MSW, Kolarevic J, Hansen ØJ, Iversen A, Hermansen Ø, Gjerde B, Hatlen G, Mortensen A, Lein I, Johansen L-H, Noble C, Gismervik K, Espmark AM 2017. Beste praksis for medikamentfrie metoder for lakseluskontroll (MEDFRI). Nofima, Rapport 10/2017
Hvas M, Oppedal F 2017. Sustained swimming capacity of Atlantic salmon. Aquaculture Environment Interactions 9, 361-369. https://doi.org/10.3354/aei00239
Hvas M, Folkedal O, Imsland A, Oppedal F 2018. Metabolic rates, swimming capabilities, thermal niche and stress response of the lumpfish, Cyclopterus lumpus . Biology open 7, bio036079
Imsland, AK., Reynolds P, Eliassen G, Hangstad TA, Foss A, Vikingstad E, Elvegård TA 2014. The use of lumpfish ( Cyclopteruslumpus L.) to control sea lice ( Lepeophtheirus salmonis Krøyer) infestations in intensively farmed Atlantic salmon ( Salmo salar L.). Aquaculture 425-426,18–23.
Jónsdóttir KE, Hvas M, Alfredsen JA, Føre M, Alver MO, Bjelland HV, Oppedal F 2019. Fish welfare based classification method of ocean current speeds at aquaculture sites. Aquaculture Environment Interactions 11, 249-261.
Kjøglum S, Mommens M, Santi N, Hultgren H 2016. Milepælsrapport – Storskal produksjon av triploid laks under kommersielle forhold. Aquagen AS.
Korsøen ØJ, Fosseidengen JE, Kristiansen TS, Oppedal F, Bui S, Dempster T 2012. Atlantic salmon ( Salmo salar L.) in a submerged sea-cage adapt rapidly to re-fill their swim bladders in an underwater air filled dome. Aquacultural Engineering 51, 1-6.
Kousoulaki K, Bogevik AS, Skiftesvik AB, Jensen PA, Opstad I 2015. Marine raw material choice, quality and weaning performance of Ballan wrasse ( Labrus bergylta ) larvae. Aquaculture Nutrition 21, 644–654.
Le HTMD, Lie KK, Etayo A, Rønnestad I, Sæle Ø 2019a. Nutrient sensing; transcriptomic response and regulation of gut motility in an agastric vertebrate. Biorxiv 827659.
Le HTMD, Shao X, Krogdahl Å, Kortner TM, Lein I, Kousoulaki K, Lie KK, Sæle Ø 2019b. Intestinal Function of the Stomachless Fish, Ballan Wrasse ( Labrus bergylta ). Frontiers Mar Sci 6, 140.
Lie KK, Tørresen OK, Solbakken MH, Rønnestad I, Tooming-Klunderud, A., Nederbragt, A.J., Jentoft, S., Sæle, Ø., 2018. Loss of stomach, loss of appetite? Sequencing of the ballan wrasse ( Labrus bergylta ) genome and intestinal transcriptomic profiling illuminate the evolution of loss of stomach function in fish. Bmc Genomics 19, 186.
Lien AM, Volent Z, Jensen Ø, Lader P, Sunde LM 2014. Shielding skirt for prevention of salmon lice ( Lepeophtheirus salmonis ) infestation on Atlantic salmon ( Salmo salar L.) in cages – A scaled model experimental study on net and skirt deformation, total mooring load, and currents. Aquacultural Engineering 58, 1-10.
Nilsen A, Nielsen KV, Biering E, Bergheim A 2017. Effective protection against sea lice during the production of Atlantic salmon in floating enclosures. Aquaculture 466, 41-50.
Nilsen A, 2019. Production of Atlantic salmon ( Salmo salar ) in closed confinement systems (CCS) - salmon lice, growth rates, mortality and fish welfare. PhD thesis, Department of Food Safety and Infection Biology, Norwegian University of Life Sciences.
Nilsen A, Nielsen KV, Biering E, Bergheim A 2017. Effective protection against sea lice during the production of Atlantic salmon in floating enclosures. Aquaculture 466, 41–50.
Nilsson J, Moltumyr L, Madaro A, Kristiansen TS, Gåsnes SK, Mejdell CM, Gismervik K, Stien LH 2019. Sudden exposure to warm water causes instant behavioural responses indicative of nociception or pain in Atlantic salmon. Veterinary and Animal Science 8, 100076.
Noble C, Nilsson J, Stien LH, Iversen MH, Kolarevic J, Gismervik K 2018. Velferdsindikatorer for oppdrettslaks: Hvordan vurdere og dokumentere fiskevelferd. 328pp. ISBN 978-82-8296-531-6.
Noble C, Gismervik K, Iversen MH, Kolarevic J, Nilsson J, Stien LH, Turnbull JF 2020. Velferdsindikatorer for regnbueørret i oppdrett: Hvordan vurdere og dokumentere fiskevelferd 309 pp.
ISBN 978-82-8296-620-7
Oppedal F, Dempster T, Stien LH 2011. Environmental drivers of Atlantic salmon behaviour in sea-cages: a review. Aquaculture 311, 1-18.
Oppedal F, Bui S, Stien LH, Overton K, Dempster T 2019. Snorkel technology to reduce sea lice infestations: efficacy depends on salinity at the farm site, but snorkels have minimal effects on salmon production and welfare. Aquaculture Environment Interactions 11, 445-457.
Oppedal F, Folkedal O, Stien LH, Vågseth T, Fosse JO, Dempster T, Warren-Myers F 2020. Atlantic salmon cope in submerged cages when given access to an air dome that enables fish to maintain neutral buoyancy. Aquaculture 525, 735286.
Overton K, Dempster T, Oppedal F, Kristiansen TS, Gismervik K, Stien LH 2018. Salmon lice treatments and salmon mortality in Norwegian aquaculture: A review. Reviews in Aquaculture, 1–20.
Poppe TT, Dalum AS, Røislien E, Nordgreen J, Helgesen KO 2018. Termisk behandling av laks. Norsk veterinærtidsskrift 3, 148–156.
Powell A, Treasurer JW, Pooley CL, Keay AJ, Lloyd R, Imsland AK, Garcia de Leaniz C 2017. Use of lumpfish for sea‐lice control in salmon farming: challenges and opportunities. Reviews in Aquaculture 10, 683-702.
Prabhu PAJ, Silva MS, Kröeckel S, Holme M-H, Ørnsrud R, Amlund H, Lock E-J, Waagbø R 2019. Effect of levels and sources of dietary manganese on growth and mineral composition of post-smolt Atlantic salmon fed low fish meal, plant-based ingredients diets. Aquaculture 512, 734287, 1-9.
Remen M, Solstorm F, Bui S, Klebert P, Vågset T, Solstorm D, Hvas M, Oppedal F 2016. Critical swimming speed in groups of Atlantic salmon Salmo salar. Aquaculture Environment Interactions 8, 659-664.
Remø SC, Hevrøy EM, Olsvik PA, Fontanillas R, Breck O, Waagbø R 2014. Dietary histidine requirement to reduce the risk and severity of cataracts is higher than the requirement for growth in Atlantic salmon smolt, independently of the dietary lipid source. Brit. J. Nutr., 111, 1759-1772.
Roth B 2016. Avlusing av laksefisk med Optilice: Effekt på avlusing og fiskevelferd. Nofima, Inc., Tromsø, Norway.
Sambraus F, Remen M, Olsen RE, Hansen TJ, Waagbø R., Torgersen T., Lock E-J, Imsland A, Fraser TWK, & Fjelldal PG (2018) Changes in water temperature and oxygen: the effect of triploidy on performance and metabolism in large farmed Atlantic salmon. Aquacult Environ Interact 10,157-172.
Samsing F, Johnsen I, Stien LH, Oppedal F, Albretsen J, Asplin L, Dempster T 2016. Predicting the effectiveness of depth-based technologies to prevent salmon lice infection using a dispersal model. Preventive Veterinary Medicine 129, 48-57
Sele V, Sanden M, Berntssen MHG, Storesund J, Lie KK, Espe M, Lundebye A-K, Hemre G-I, Waagbø R, Ørnsrud R 2019. Program for overvåking av fiskefôr – Årsrapport for prøver innsamlet i 2018. Mattilsynets overvåkningsprogram. Rapport fra Havforskningen 30-2019, ISSN 1893-4536 (online), 56 pp.
Sigholdt F, Finstad B, Effect of low temperature on seawater tolerance in Atlantic Salmon (Salmo salar) Smolts. Aquaculture 84, 167-172.
Solberg MF, Skaala Ø, Nilsen F, Glover KA 2013. Does Domestication Cause Changes in Growth Reaction Norms? A Study of Farmed, Wild and Hybrid Atlantic Salmon Families Exposed to Environmental Stress. PLoS ONE 8(1): e54469.
Sommerset I, Walde CS, Bang Jensen B, Bornø B, Haukaas A, Brun E 2020. Fiskehelserapporten 2019, utgitt av Veterinærinstituttet 2020.
Stien LH, Nilsson J, Hevrøy EM, Oppedal F, Kristiansen TS, Lien AM, Folkedal O 2012. Skirt around a salmon sea cage to reduce infestation of salmon lice resulted in low oxygen levels. Aquacultural Engineering 51, 21-25.
Stien LH, Dempster T, Bui S, Glaropoulod A, Fosseidengen JE, Wright DW, Oppedal F 2016. ‘Snorkel’ sea lice barrier technology reduces sea lice loads on harvest-sized Atlantic salmon with minimal welfare impacts. Aquaculture 458, 29-37.
Stien LH, Lind MB, Oppedal F, Wright DW, Seternes T 2018. Skirts on salmon production cages reduced salmon lice infestations without affecting fish welfare. Aquacultlure 490, 281-287.
Stien LH, Størkersen KV, Gåsnes SK, 2020. Analyse av dødelighetsdata fra spørreundersøkelse om velferd hos rensefisk. Rapport fra Havforskningen NR. 2020-6.
Stien LH, Sæther PA, Kristiansen TS, Fjelldal PG og Sambraus F. 2019. Første samlerapport: Velferd for triploid laks i Nord-Norge — Fra utsett til slakt, utsett 2014-2017. Rapport fra Havforskningen 2019-47 ISSN:1893-4536
Tørud, B, Bang Jensen B, Gåsnes S, Grønbech S, Gismervik K. m.fl. 2019: Dyrevelferd i settefiskproduksjonen – SMÅFISKVEL. VI Rapport 14-2019
Waagbø R 2006. Chap 13 Feeding and disease resistance in fish, In Mosenthin, R., Zentek, J. & Zebrowska, T. (eds.) Biology of growing animal : Elsevier Limited, pp. 387-415.
Waagbø R 2008. Reducing production related diseases in farmed fish. In: Lie, Ø. (ed.) Improving farmed fish quality and safety : VS Woodhead Publishing, pp. 363-398.
Waagbø R, Remø SC 2020. Chap 7. Functional diets in fish health management. In Aquaculture Health Management (edited by Fred Kibenge and Mark Powell), Elsevier (in press)
Yuen JW, Dempster T, Oppedal F, Hvas M 2019. Physiological performance of ballan wrasse ( Labrus bergylta ) at different temperatures and its implication for cleaner fish usage in salmon aquaculture. Biological Control 135, 117-123.
Øie G, Galloway T, Sørøy, M., Hansen, M.H., Norheim, I.A., Halseth, C.K., Almli, M., Berg, M., Gagnat, M.R., Wold, P.-A., Attramadal, K., Hagemann, A., Evjemo, J.O., Kjørsvik, E., 2015. Effect of cultivated copepods ( Acartia tonsa ) in first-feeding of Atlantic cod ( Gadus morhua ) and ballan wrasse ( Labrus bergylta ) larvae