Denne rapporten er en leveranse til bestillingen «Grunnlagsstudie: Pelagisk økosystem» fra Oljedirektoratet til Havforskningsinstituttet. Studien er bestilt i forbindelse med at regjeringen har igangsatt en prosess for åpning av norsk sokkel for havbunnsmineralvirksomhet. Rapporten sammenstiller vår nåværende kunnskap om de pelagiske økosystemenes struktur og funksjon og belyser viktige kunnskapshull. Det er lagt særlig vekt på å beskrive økosystemkomponenter og prosesser som er essensielle for arbeidet med planprogram for konsekvensutredning av havbunnsmineralvirksomhet i norske havområder, og fokus vil derfor tidvis være litt annerledes enn det man vil finne i en generell beskrivelse av Norskehavet, Islandshavet og Grønlandshavet. Vi har lagt mer vekt på eksisterende kunnskapshull, og fokusert mer på utredningsområdet og på økologi og prosesser i pelagisk sone i dyphavet, enn det som kan forventes i for eksempel en lærebok om plankton-, fisk eller sjøpattedyrøkologi. Selv om formålet dermed har farget fokuset for beskrivelsene, beskriver kapitlene generell fordeling og økologi i de pelagiske systemene i Norskehavet, Islandshavet og Grønlandshavet.
Tina Kutti har koordinert arbeidet med rapporten som er inndelt i forskjellige fagkapittel med følgende forskere som ansvarshavende: 1) fysisk oseanografi - Kjell Arne Mork, 2) kjemisk oseanografi - Melissa Chierici, 3) biogeokjemi/marinkjemi – Knut Yngve Børsheim, 4) mikrobiologi - Knut Yngve Børsheim, 5) planteplankton - Knut Yngve Børsheim, 6) mesozooplankton - Espen Bagøien, Tor Knutsen, Cecilie Thorsen Broms, 7) makroplankton og mesopelagisk fisk - Thor Klevjer, Tor Knutsen, Espen Strand, Harald Gjøsæter, 8) dyreplankton i relasjon til hydrotermale kilder – Tor Knutsen, 9) pelagisk fisk - Erling Kåre Stenevik, Åge Høines, 10) fisk knyttet til kontinentalskråningene rundt Norskehavsbassenget - Kristin Windsland, 11) bunnfisk på dypet - Harald Gjøsæter, 12) sjøpattedyr - Anne Kirstine Frie, 13) bentisk-pelagisk kobling - Tina Kutti. Arbeidet er utført i forsknings- og rådgivningsprogram Marine prosesser og menneskelig påvirkning, ledet av Frode Vikebø.
Sammendrag
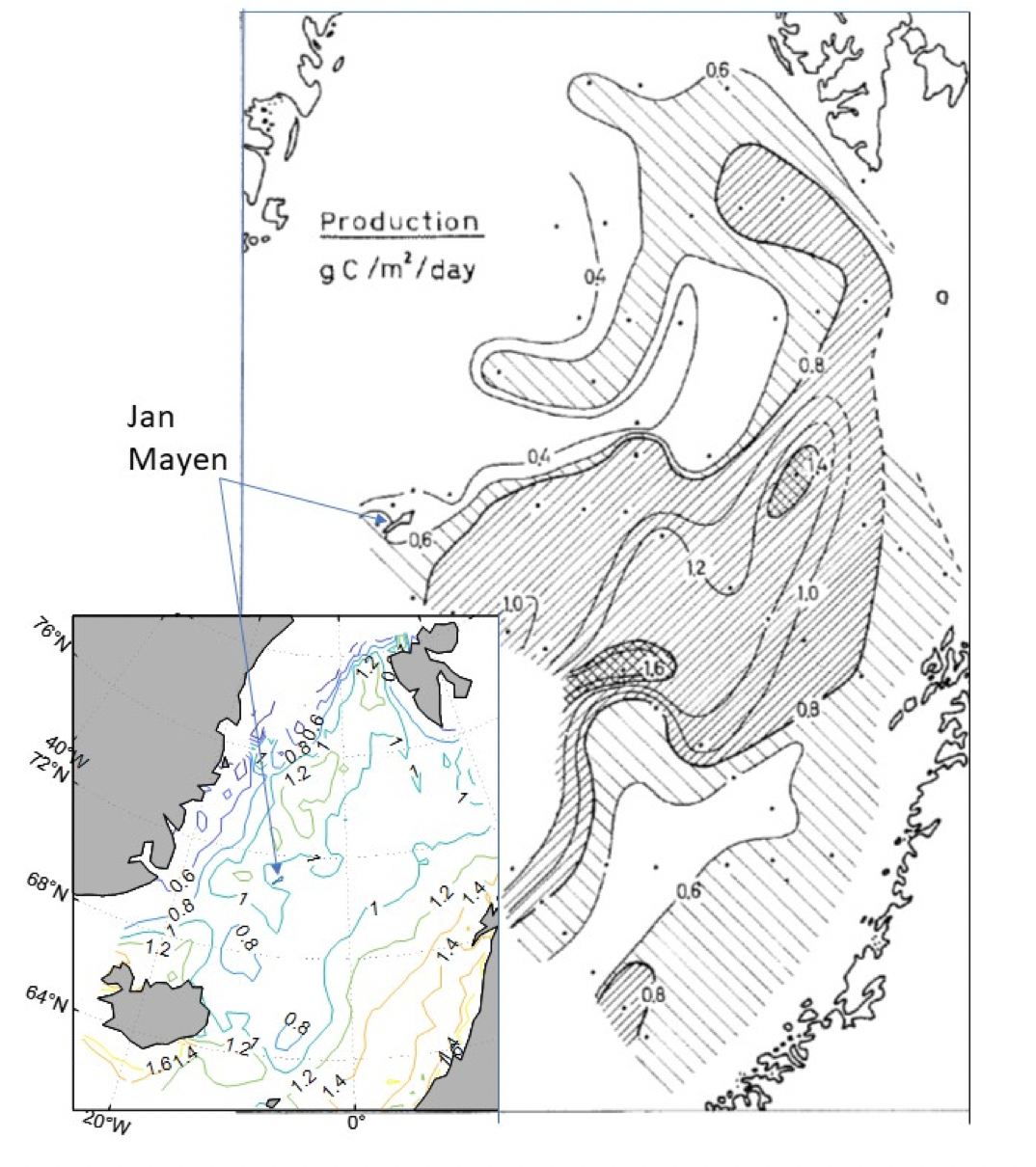
Denne rapporten er utarbeidet på oppdrag fra Oljedirektoratet (OD) og utgjør et faglig grunnlag til myndighetenes konsekvensutredning om åpning av deler av norsk sokkel for havbunnsmineralvirksomhet. Rapporten sammenstiller eksisterende kunnskap om fysiske og kjemiske forhold, og den romlige fordeling av arter og samfunn over dyp og mellom forskjellige geografiske områder, i havområdene vest for Norge. Rapporten tar for seg alle de trofiske nivåene i det pelagiske økosystemet, fra mikroorganismer til sjøpattedyr, og de nøkkelprosesser som knytter forskjellige økosystemkomponenter sammen. Hovedfokus er på det geografiske området hvor regjeringen har igangsatt åpningsprosessen for havbunnsmineralvirksomhet. Dette utredningsområdet strekker seg over deler av De nordiske hav. Det dekker 26 % av norske havområder, er halvannen gang større enn Norges samlede landareal og har et bunndyp som varierer mellom 100 og 4000 meter.
De nordiske hav er en fellesbetegnelse for Norskehavet, Islandshavet og Grønlandshavet. De brer seg nordover fra Grønland-Skottlandryggen, opp mot Framstredet og Spitsbergen og begrenses videre av Norge i øst og Grønland i vest. Havområdet karakteriseres av en sterk nordgående strøm av varmt vann på østsiden, langs norskekysten, og en sydgående strøm av kaldt vann på vestsiden. Langs midthavsryggene Jan Mayenryggen, Mohnsryggen og Knipovichryggen dannes en relativt skarp front mellom de varme og kalde vannmassene, som blir kalt Den arktiske front. Hvordan de to vannmassene er fordelt over havområdet og i dypet, og blandingen av vannmasser med ulik temperatur og saltinnhold, er svært viktig fordi det styrer utbredelsen av arter, populasjoner og samfunn av både plankton, fisk og sjøpattedyr.
Med sin geografiske plassering spiller De nordiske hav en viktig klimatisk rolle. Tyngre vannmasser blir dannet her ved at varmt og salt atlanterhavsvann som strømmer inn i området, mister mye av sin varme til atmosfæren. Når dette skjer blir vannet tyngre og synker ned i dypet og strømmer senere ut av De nordiske hav og inn i sørligere deler av Atlanterhavet, og bidrar slik til omveltnings-sirkulasjonen i det nordlige Atlanterhavet. Denne transport av vann fra overflaten til dypet er av stor betydning for den globale karbonsyklusen, og overfører CO2 som overflatevannet har tatt opp fra atmosfæren ned i dypet av havet. Beregninger viser at en stor del av de menneskeskapte karbondioksidutslippene til atmosfæren er blitt absorbert nettopp i dette geografiske området. Denne prosessen påvirker karbonkjemien slik at både volumet og arealet av vann hvor kalkskjell eller andre kalkformasjoner begynner å gå i oppløsning, har økt tydelig de siste 20 årene. Samtidig er det detektert en trend med synkende pH gjennom hele vannsøylen og med en raskere pH-reduksjon i dypbassengene i Norskehavet enn i de øvrige delene av dette havområdet.
Man har i dag god oversikt over de generelle hydrografiske forhold og havstrømmer i De nordiske hav, men det mangler detaljert kunnskap (av høy romlig oppløsning) i topografiske komplekse områder som rundt midthavsryggene. Å etablere slik kunnskap vil kreve flere målestasjoner for innsamling av fysiske og kjemiske oseanografiske data og utplassering av rigger med måleinstrumenter. Havforskningsinstituttets faste snitt strekker seg ikke inn i utredningsområdet, med unntak av de forlengede Gimsøy-NV og Bjørnøya-V snittene som tas én gang i året.
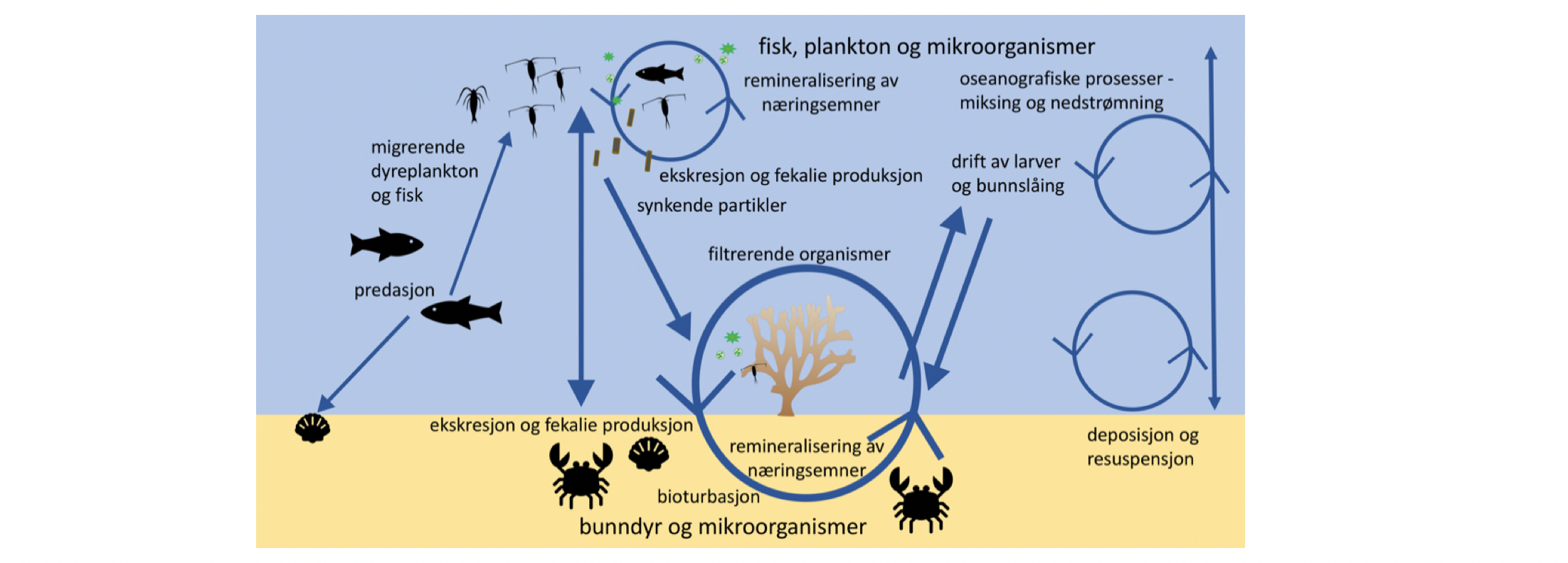
Plankton forekommer fra bunn til overflate gjennom hele utredningsområdet, men mengden og artssammensetningen varierer geografisk og med miljøforholdene i ulike typer vannmasser. Mengden av en gitt planktonart i et bestemt område og dyp vil videre endre seg gjennom sesongsyklusen. Bakteriene er de mest tallrike organismene, og deres funksjon i det pelagiske økosystemet er i hovedsak knyttet til nedbryting av organisk materiale. Planteplanktonet omdanner uorganisk karbon til biologisk bundet organisk karbon ved fotosyntese. Dyreplanktonet spiller en viktig rolle i det pelagiske systemet ved å beite på planteplanktonet, mens de selv spises av organismer på høyere trofiske nivå. Slik kanaliseres energien fra primærprodusentene oppover i næringsnettet. De viktigste dyreplanktongruppene i De nordiske hav er mikrodyreplankton, hoppekreps, muslingkreps, pilormer, krill, amfipoder, pelagiske reker, geléplankton, pelagiske snegler, dessuten fiskeegg og -larver og mesopelagisk fisk. Fysiske og biologiske prosesser styrer energiflyten mellom øvre vannlag med primærproduksjon og dypereliggende økosystemer. Mange arter vandrer daglig fra mesopelagiske dyp til vannmasser nær overflaten. Her beiter de på mindre planktonarter, før de vandrer ned i dypet igjen. På denne måten bidrar de til transport av organisk karbon til dypet.
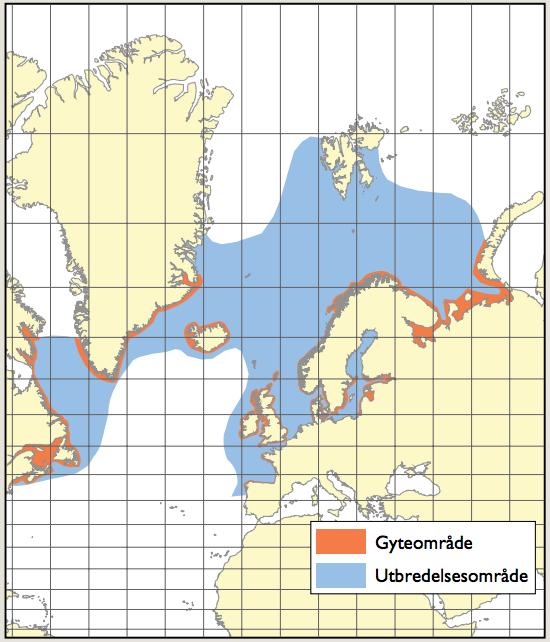
Hoppekrepsen Calanus finmarchicus (raudåte) vurderes å være arten som bidrar mest til biomassen av mellomstore dyreplankton i Nord-Atlanteren, og spiller en økologisk nøkkelrolle i Norskehavet. Unge individer av C. finmarchics er hovedføden for larvene til en rekke fiskebestander av kommersiell og økologisk betydning, mens eldre individer representerer den viktigste føden for planktonspisende fiskebestander som sild og makrell. Norskehavs-, Lofoten- og Islands-bassengene er overvintringsområder for C. finmarchicus. Bassenget i Grønlandshavet er kjerneområde for raudåtens større, arktiske slektning Calanus hyperboreus. Krill og amfipoder er to svært viktige grupper av det større dyreplanktonet i De nordiske hav, både mht. biomasse og økologi. Dyreplankton kan gjennomføre betydelige vertikale vandringer og har blitt observert på store dyp, også i nærheten av områder med aktive varme havkilder.
Mens undersøkelser av virus fra utredningsområdet ikke er kjent i tilgjengelig litteratur, er det publisert noen få studier på bakterier, hovedsakelig i tilknytning til karbonsyklus i vann nær overflaten, men også fra bunnvann nær varme havkilder. Planteplankton i utredningsområdet har vært karakterisert i enkelte studier som også omfatter årstidsvariasjon. Prøvetakingen av dyreplanktonet innen utredningsområdet fokuserer i hovedsak på den øvre delen av vannsøylen, og går kun unntaksvis dypere enn ca. 1000 m. Slike dype prøver er meget spredt geografisk og mht. sesong. Mens kunnskap om dyreplanktonartenes mengder, fordelinger, produksjon og trofiske interaksjoner selv i øvre vannlag er begrenset, er den meget mangelfull på større dyp. Det er et stort behov for denne typen kunnskap generelt og ved Den arktiske front spesielt.
Det pelagiske fiskesamfunnet i Norskehavet er dominert av store bestander av atlantisk sild, makrell og kolmule. I tillegg finnes det store mengder av lodde, og mindre mengder atlantisk laks, rognkjeks og makrellstørje. Norskehavet er først og fremst et viktig beiteområde for de arter som vandrer inn i Norskehavet, bl.a. for å beite på dyreplankton om våren og sommeren. Sild, makrell, lodde, laks, rognkjeks og makrellstørje holder seg fremfor alt i de øvre vannlag mens kolmulen ofte finnes dypere, mellom 200 og 500 m. Kontinentalskråningen er påvirket av en uttalt hydrografisk front mellom varme atlantiske vannmasser og kalde vannmasser. Her forekommer en rekke arter, herunder flere kommersielt viktige arter som blåkveite, snabeluer og vanlig uer.
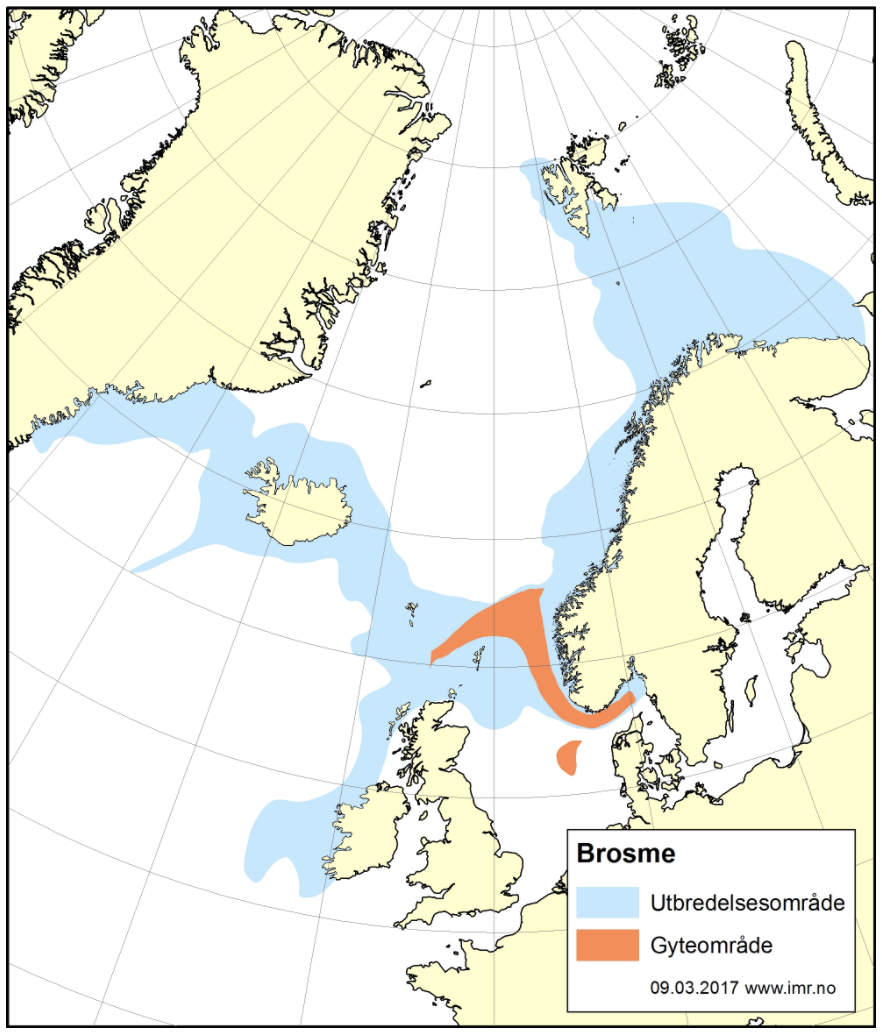
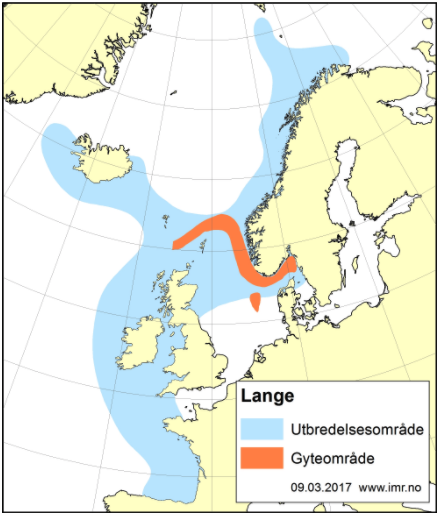
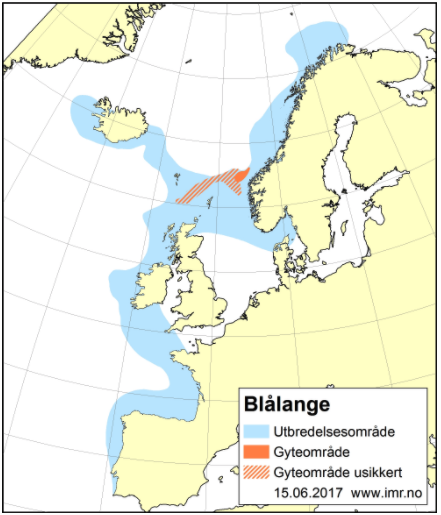
Det brukes betydelige ressurser på å overvåke tilstanden til de kommersielt viktige fiskeartene, og lange tidsserier med data fra fiskeri og forskningstokt gir oss et godt kunnskapsgrunnlag for å forvalte bestandene. For de fleste bestander er status god og de høstes bærekraftig med gytebestander over føre-var-nivå. Noen arter er listet som sårbare, sterkt truet eller kritisk truet på Norsk rødliste for arter, herunder vanlig uer, blålange og en rekke bruskfiskarter. For ikke-kommersielle arter mangler ofte informasjon om bestandsstørrelse, reproduksjons- og oppvekstområder, samt kunnskap om vandringsmønster. For områder dypere enn 1000 m er kunnskapsmangelen betydelig. Det finnes kun sporadiske undersøkelser på større dyp, hvor mange ikke-kommersielle arter lever. Behovet for mer kunnskap om disse ikke-kommersielle artene er stort, da disse kan ha en viktig funksjon i økosystemet, som bentisk-pelagisk kobling i næringsnettet. Tråling på slike dyp er utfordrende, men ikke umulig.
Tolv sjøpattedyrarter blir jevnlig observert i utredningsområdet. Vågehval, finnhval og knølhval er de mest tallrike bardehvalene. De observeres fremfor alt om sommeren, rundt hele Den arktiske front og til dels også i mer sørlige områder av Norskehavet. Hanner av tannhvalarten spermhval beiter også over det meste av utredningsområdet om sommeren. De nevnte storhvalene antas generelt å dra til sørlige farvann i løpet av vinteren for å pare seg eller kalve. Tannhvalartene spekkhogger, nebbhval og kvitnosdelfin blir stort sett i disse nordlige farvann gjennom hele året. Både for de stasjonære og de migrerende artene er beitingen i og rundt utredningsområdet antakelig av stor betydning for overlevelse og reproduksjonsevne. Bardehvalartene beiter i stor grad på dyreplankton og mindre pelagisk fisk, mens tannhvalene fortrinnsvis spiser blekksprut og fisk. Selarten klappmyss beiter over store deler av utredningsområdet, fremfor alt på fisk og blekksprut. Sjøpattedyrenes beitesuksess påvirkes av naturlige og menneskeskapte svingninger i forekomsten av byttedyr, men kan også påvirkes av direkte forstyrrelser fra andre menneskelige aktiviteter enn fiskerier.
I De nordiske hav ble bestandene av alle storhvalarter sterkt redusert av hvalfangst. Etter fredningen, i løpet av 1900-tallet, har finnhval og knølhval kommet tilbake på et livskraftig nivå, mens blåhval, seihval og nordkaper fortsatt er svært sjeldne. Vågehvalbestanden, som ble mindre hardt beskattet enn de større bardehvalartene, har holdt seg på et stabilt høyt nivå de siste tiårene. Bestandene av spekkhogger og kvitnosdelfin er også klassifisert som livskraftige. Den nordøstatlantiske bestanden av klappmyss ble desimert i de første to årene etter 2. verdenskrig, grunnet overfangsting. En manglende gjenoppretting av bestanden etter fredning i 2006 viser at det også er andre begrensende faktorer for bestanden. Fremtidig status for sjøpattedyrene i utredningsområdet vil variere betydelig mellom arter på grunn av forskjeller i biologi, økologi og nåværende utgangspunkt.
Det eksisterer god kunnskap over de generelle hydrografiske forhold og havstrømmer i De nordiske hav. Det er imidlertid mangler med hensyn til høyoppløst informasjon om bunnstrømmer i topografiske komplekse områder som rundt midthavsryggene. Det meste av vår eksisterende kunnskap om plankton og fisk i De nordiske hav er basert på undersøkelser fra de øvre vannlag (0-1000 m) og vår kunnskap om de dypere vannlag er svært begrenset. Sjøpattedyr observeres først og fremst på overflaten, og det finnes meget begrenset kunnskap om deres aktivitet på dypet. De nordiske hav dekker et meget stort og heterogent areal, slik at vår kunnskap, også om de best undersøkte områdene, er meget fragmentert. Fremover må det gjøres en innsats for å tette disse kunnskapshullene.
1 - Innledning
1.1 - Havbunnsmineralvirksomhet på norsk sokkel
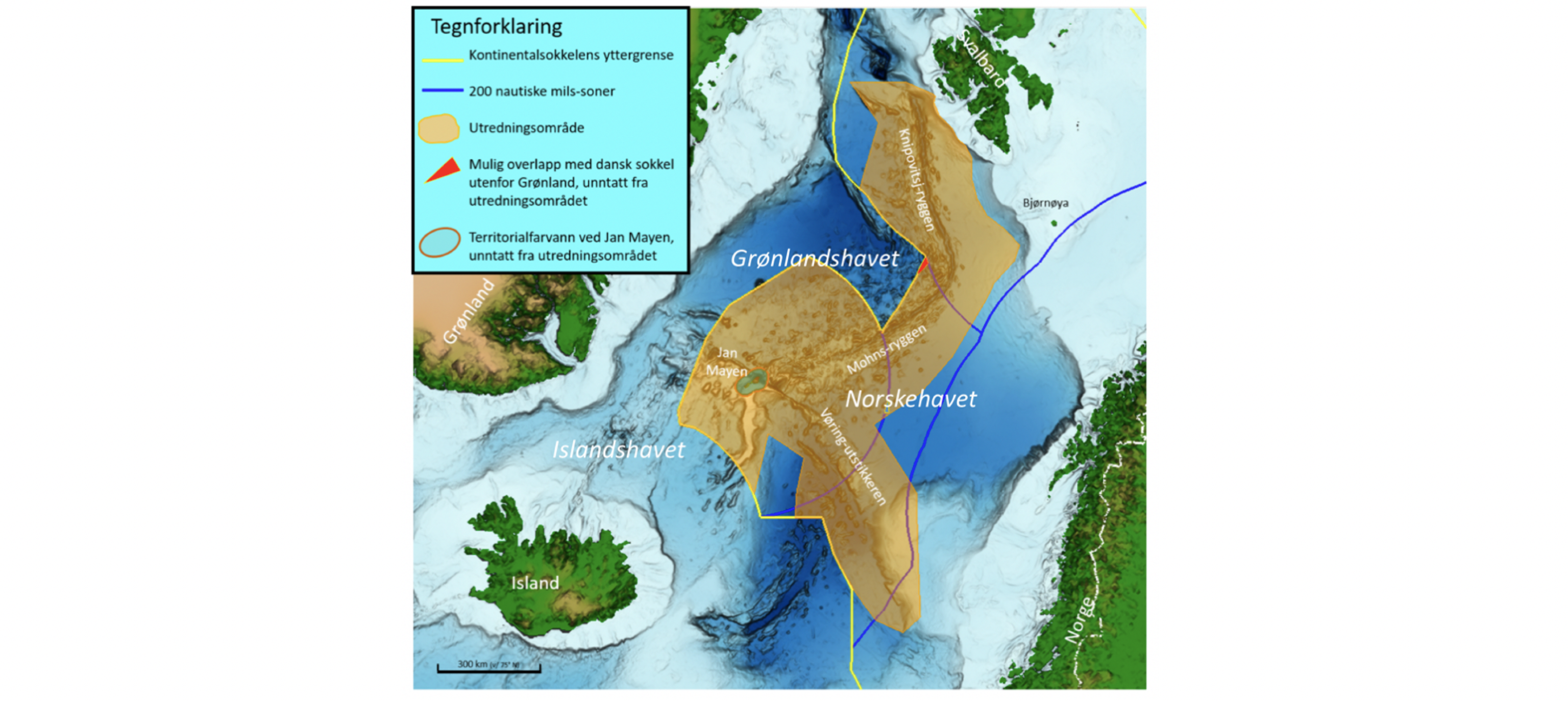
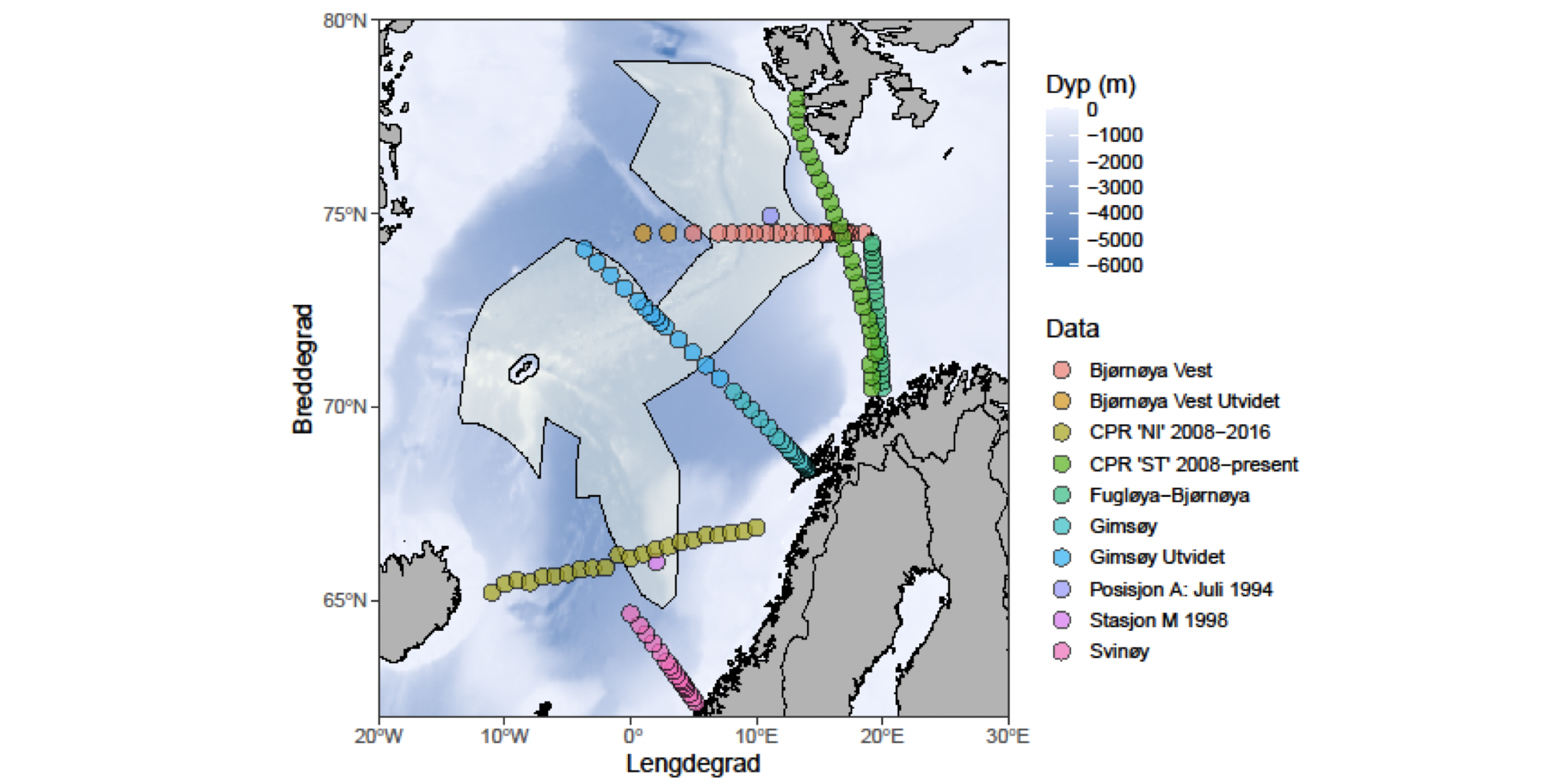
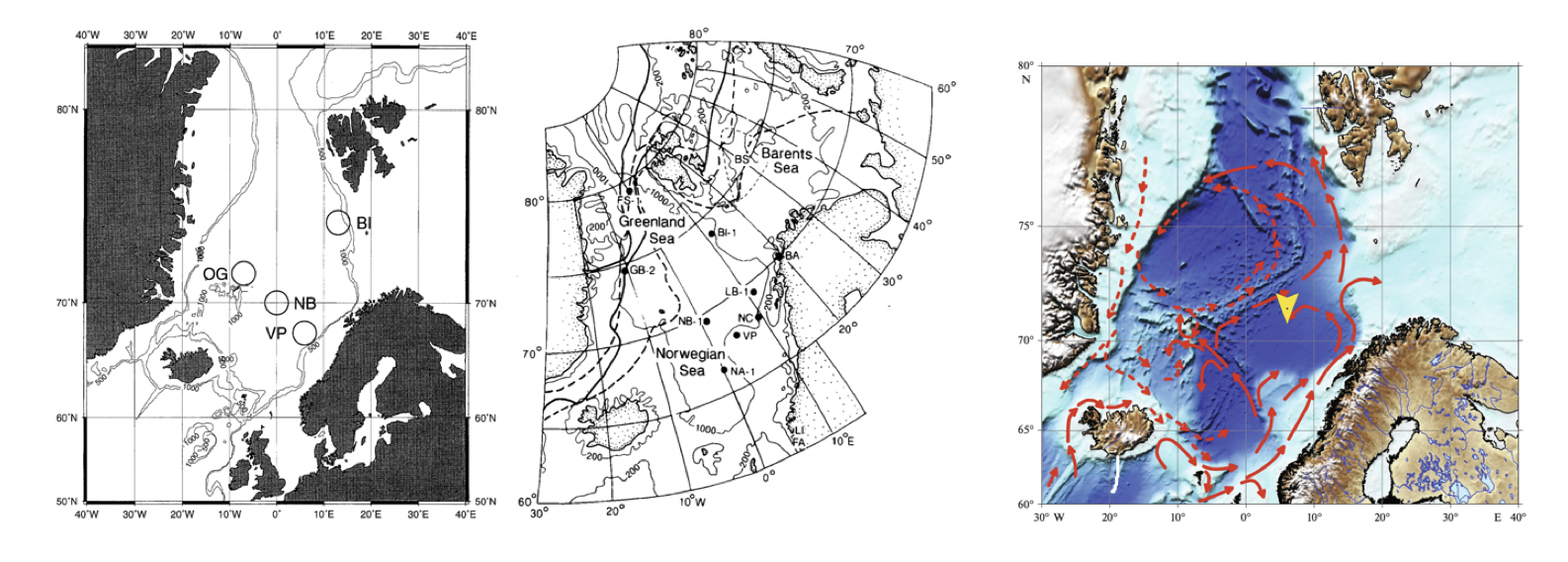
Regjeringen har igangsatt en prosess for å åpne norsk sokkel for havbunnsmineralvirksomhet. Som et av de første steg i den prosessen ønsker man å få en oversikt av tilgjengelig kunnskap om det fysiske, kjemiske og biologiske miljø i de områdene hvor det kan bli aktuelt å utvinne havbunnsmineraler. Et utredningsområde er blitt definert (se Figur 1.1), som omfatter et areal på 592 000 km2 og hvor midhavsryggene Jan Mayenryggen, Vøringutstikkeren, Mohnsryggen og Knipovichryggen står i senter. Utredningsområdet strekker seg over store deler av De nordiske hav, en fellesbetegnelse for Norskehavet, Islandshavet og Grønlandshavet. Det dekker 26 % av norske havområder, er halvannen gang større enn Norges samlede landareal og har et bunndyp som varierer mellom 100 og 4000 meter. Denne rapporten sammenstiller nåværende kunnskap om pelagiske økosystemkomponenter i dette havområdet.
Figur 1.1 Kart over De nordiske hav (Norskehavet, Islandshavet og Grønlandshavet) med området som utredes for åpning for havbunnsmineralvirksomhet, utredningsområdet, markert i brunt (kart fra OD).
1.2 - De nordiske hav: geografi og kunnskapsstatus
Området nord for Grønland-Skottland-ryggen (som strekker seg fra Skottland over Færøyene mot Island og Grønland), sør for Framstredet/Spitsbergen og vest for kontinentalskråningen mot Barentshavet benevnes Det nordiske hav. De nordiske hav har et totalt areal på 2.6 millioner km2, knapt 1% av klodens totale havareal. Gjennomsnittsdyp er 1600 m og havområdet har et totalt volum på 4.1 millioner km3. Området er svært heterogent. Bunnen er topografisk sett meget kompleks og består av grunn kontinentalhylle og skråning, dype bassenger (Boreasbassenget, Grønlandsbassenget, Lofotenbassenget og Norskehavsbassenget), midthavsrygger og undersjøiske fjell, platåer, samt utallige undervannscanyoner og raviner.
Vannmassene i De nordiske hav karakteriseres av en sterk nordgående strøm av varmt vann på østsiden og en sydgående strøm av kaldt vann på vestsiden. Transporten av varmt og salt atlanterhavsvann inn i Norskehavet fra sørlige deler av det nordlige atlanterhav har blitt beregnet til omtrent 8 millioner tonn per sekund, noe som er betydelig. Langs midthavsryggene Jan Mayenryggen, Mohnsryggen og Knipovichryggen dannes en relativt skarp front mellom de varme og kalde vannmassene, som blir kalt Den arktiske front. Hvordan de to vannmassene er fordelt over De nordiske hav, horisontalt og i dypet, og blanding av vannmasser med forskjellig temperatur og saltholdighet, er svært viktig fordi det styrer utbredelsen av organismer på alle de trofiske nivåene, fra mikroorganismer og dyreplankton til fisk og sjøpattedyr. Mens noen arter holder seg fortrinnsvis til varmere vannmasser er andre sterkt knyttet til kalde vannmasser. Noen arter forekommer kun i varme vannmasser, andre kun i kalde vannmasser. Fordelingen av vannmasser i området styrer på denne måten utbredelsen av arter, så vel som størrelsen av forskjellige populasjoner og strukturen (det vil si artssammensetningen) av de pelagiske samfunn vi finner i de forskjellige delene av De nordiske hav. Området karakteriseres ikke av sin artsrikdom men heller av at finnes svært høye biomasser av mange av de vanligste artene og av at havområdet i sin helhet er svært produktivt.
I over tusen år har man i Norge hatt kystnære fiskerier av betydelig art på fiskebestandene av den nordøstarktiske torsken og norsk vårgytende sild. Hvor torsken og silden befant seg når den ikke var ved kysten visste man ikke før på slutten av 1800-tallet. En av hypotesene var at den holdt seg lenger ute til havs, men det fantes ikke noen vitenskapelige observasjoner som kunne støtte dette. Havområdene utenfor Norge ble vitenskapelig og grundig studert for første gang i 1876-1878, under Den norske nordhavsekspedisjonen. Ekspedisjonen dekket Norskehavsområdet fra Færøyene til Svalbard og samlet inn en mengde vitenskapelig informasjon om blant annet vannmassenes fysiske egenskaper, plankton, fisk og bunndyr. Etter ekspedisjonen utviklet man det første strømkartet over De nordiske hav, man beskrev torskens vandring og at den torsken som gyter ved norskekysten om vinteren/våren tilhørte samme bestand som om sommeren oppholder seg sør for Svalbard i Barentshavet. Dette var starten på den norske havforskningen. Siden den tid er det blitt lagt ned en betydelig innsats for å beskrive dette havområdets struktur (det vil si hvilke arter som forekommer hvor) og for å forstå hvordan de forskjellige artene, populasjonene og samfunnene som man finner her er avhengige av hverandre og det fysiske og kjemiske miljøet i havet. Aller størst kunnskap har vi om økosystemene i Norskehavet og dette er veldig godt sammenfattet i boken The Norwegian Sea Ecosystem hvor HR Skjoldal var redaktør (2004). Om den norske havforskningens historie kan man lese i Historier om og fra havforskningen av E Moksness og O Nakken (https://www.hi.no/resources/Historier-om-og-fra-havforskningen.pdf).
Figur 1.2 Forskningsskipet Michael Sars I, brukt mellom 1900 og 1914 (fotograf ukjent) og måling av saltholdighet og temperatur ved hjelp av vannhenter (fotograf: Alf Dannevig).
I dag drifter Havforskningsinstituttets rederi 6 fartøy som brukes til forskningstokt i norske havområder av Havforskningsinstituttet, Universitetet i Bergen, Universitetet i Tromsø og Norsk Polarinstitutt. Årlig står disse båtene for 1300-1500 effektive toktdøgn. I tillegg bruker Havforskningsinstituttet leiefartøy med omtrent det samme antall toktdøgn årlig og en referanseflåte på omtrent 20 havgående fiskefartøy for å samle inn vitenskapelig informasjon fra norske havområder. Kunnskapen om det marine miljø i disse havområdene er derfor betydelig og stadig voksende.
1.3 - Rapportens formål og inndeling
Denne rapporten sammenstiller nåværende kunnskap om de pelagiske økosystemenes struktur og funksjon i De nordiske hav. Kunnskapen er i hovedsak hentet fra publisert engelskspråklig og norsk litteratur, men også noe upubliserte data fra Havforskningsinstituttets databaser er benyttet. Rapporten er delt inn i forskjellige underkapitler som omhandler de forskjellige fagfeltene: a) fysisk oseanografi, b) kjemisk oseanografi, c) plankton (i.e. biogeokjemi, mikrobiologi, planteplankton og dyreplankton), d) fisk (i.e. pelagisk fisk, fisk knyttet til kontinentalskråningene rundt Norskehavsbassengen og bunnfisk på dypet, e) sjøpattedyr og f) bentisk-pelagisk kobling. Ettersom det ikke er tydelige skillelinjer mellom disse fagfeltene vil noe kunnskap bli repetert i to eller flere kapitel, slik at alle underkapitler vil kunne leses som selvstendige enheter. Etter hvert underkapittel følger en konklusjon hvor de viktigste kunnskapshullene i fagfeltet blir trukket frem.
2 - Fysisk oseanografi
2.1 - Innledning
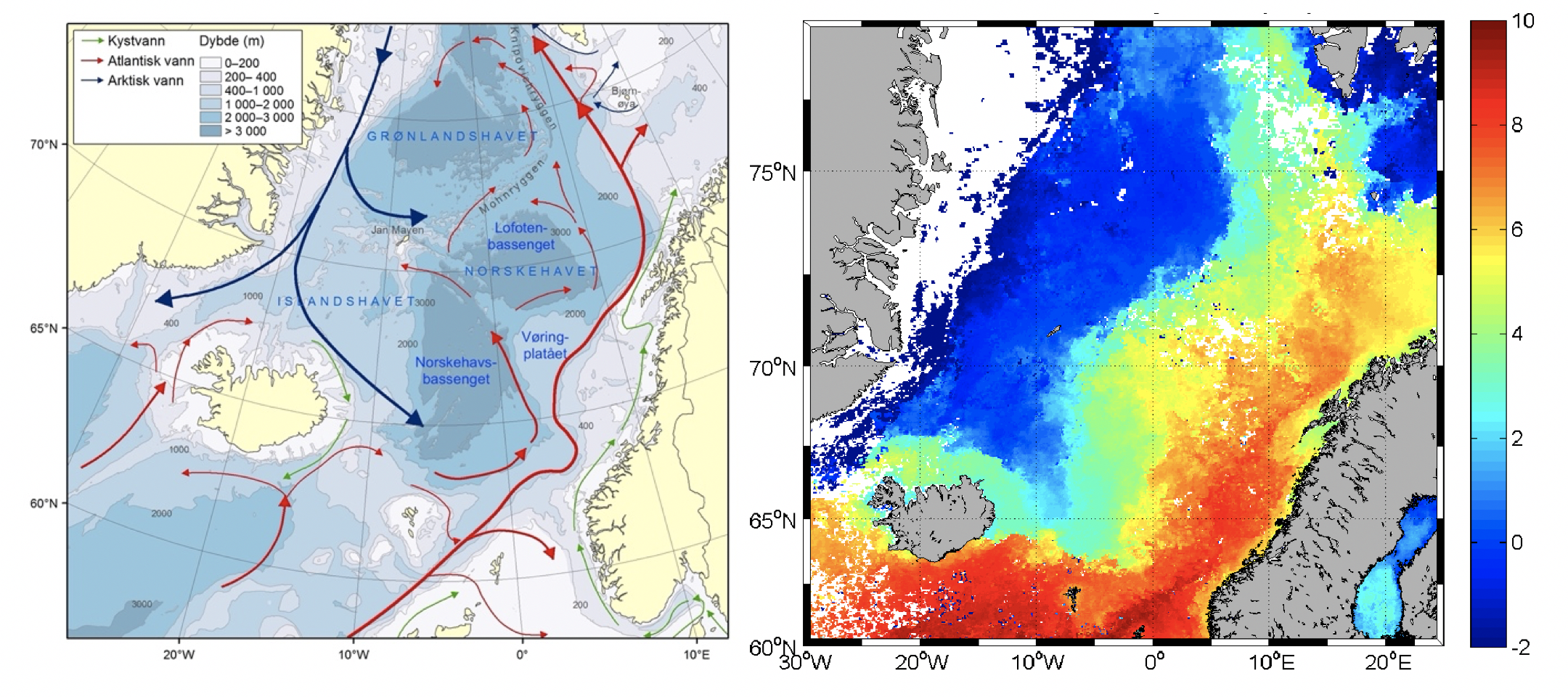
Med sin plassering spiller De nordiske hav (fellesbetegnelse for Norskehavet, Grønlandshavet og Islandshavet) en viktig klimatisk rolle. Tyngre vannmasser blir her dannet ved at varmt, salt atlanterhavsvann som strømmer inn i området mister mye av sin varme til atmosfæren. De tyngre vannmassene synker ned i dypet og senere ut av De nordiske hav og inn i Atlanterhavet, og bidrar til omveltningssirkulasjonen i det nordlige Atlanterhavet. Kaldt, ferskere vann fra Polhavet vil også passere De nordiske hav på sin ferd sørover mot Atlanterhavet langs Øst-Grønland. Noe av dette vannet blir transportert inn i sentrale områder via flere sidegrener (Figur 2.1).
Figur 2.1 a) Kart over Norskehavet med bunntopografi og de viktigste havstrømmer, b) overflatetemperatur fra satellitt data (eksempel fra februar 2008).
Sirkulasjonen og fordelingen av de forskjellige vannmassene er i stor grad bestemt av bunntopografien. Først og fremst spiller Grønland-Skottland-ryggen en stor rolle fordi den isolerer de dype vannmassene i Atlanterhavet fra de dypere lag i De nordiske hav. Derfor er dypvannet overalt i De nordiske hav karakterisert av arktiske vannmasser med temperaturer under 0°C. Med unntak av overgangslaget under det vindblandede laget i overflaten, er lagdelingen i vannsøylen liten, og uten store vertikale sprang i tettheten. Dette er med på å forsterke den topografiske styringen av strømmene i området. Selv dype topografiske strukturer kan spille en viktig rolle, som for eksempel Mohns- og Knipovich-ryggene nord for Jan Mayen og Jan Mayen-ryggen sør for Jan Mayen. Ved disse ryggene møtes varme og kalde vannmassene som skaper fronter med store horisontale gradienter i temperatur og saltholdighet. Strømmene vil her i hovedsak ha retning langs med ryggene og den arktiske fronten følger også ryggene. I hovedtrekk er strømforholdene i De nordiske hav, slik de er skissert i figur 2.1, dominert av en nordgående strøm av varmt vann på østsiden, Den norske atlanterhavsstrømmen, og en kald sydgående strøm på vestsiden, Øst-Grønlandsstrømmen, med en forholdsvis skarp arktisk front i skillet mellom de kalde og de varme vannmassene (Figur 2.1). Vann fra Atlanterhavet kommer hovedsakelig inn i De nordiske hav gjennom Færøy-Shetland-kanalen og over ryggen mellom Island og Færøyene. Øst-Grønlandsstrømmen transporterer overflatevann med lav saltholdighet og is fra Polhavet. Under det kalde overflatevannet, i et varmere intermediært lag (ca. 100–200 m dyp), er det vann av atlantisk opprinnelse som kommer sørover igjen fra Vest-Spitsbergenstrømmen som er den nordligste grenen i Den norske atlanterhavsstrømmen (Figur 2.1). I dypere lag fører Øst-Grønlandsstrømmen dypvann fra Polhavet som er en viktig komponent av dypvannet i De nordiske hav. Øst-Grønlandsstrømmen har to viktige grener som går inn i De nordiske hav. Den sørligste grenen går sørøstover inn i den sørlige delen av Islandshavet og er kalt Øst-Islandsstrømmen. Øst-Islandsstrømmen har stor betydning for forholdene ved Nord-Island og sørlige Norskehavet. Den nordligste grenen fra Øst-Grønlandsstrømmen har større betydning i området rundt Jan Mayen. Grenen tar av østover der Øst-Grønlandsstrømmen møter Islandsplatået på sørsiden av det dype Grønlandsbassenget. Denne grenen bærer navnet Jan Mayenstrømmen og fører kaldt polarvann østover på nordsiden av Jan Mayen og videre nordøstover langs Mohnsryggen. Her går den parallelt med det varmere atlantiske vannet fra den vestligste grenen av Den norske atlanterhavsstrømmen som medfører at den arktiske fronten er skarp i dette området. Variasjonene i utbredelse og egenskapene til atlantiske og arktiske vannmasser sammen med endringer i hav-atmosfære flukser bidrar hovedsaklig til de klimatiske endringene som er observert i De nordiske hav, og som igjen har stor betydning for det marine økosystemet.
2.2 - Data og metoder
Altimetri (havnivå) fra satellitt for perioden 2010-2020 er brukt for å estimere midlere (geostrofisk) overflatestrøm for sommer og vinter. Dataene er produkter fra AVISO (http://www.aviso.ocean- obs.com/duacs) og har en oppløsning på 1/3 grad i lengde og bredderetning. De samme dataene er også brukt for å beregne mesoskala (mindre enn 100 km utstrekning) virvel aktivitet, men pga. den horisontale oppløsningen til dataene vil ikke virvler med diameter mindre enn ca. 20-30 km oppløses.
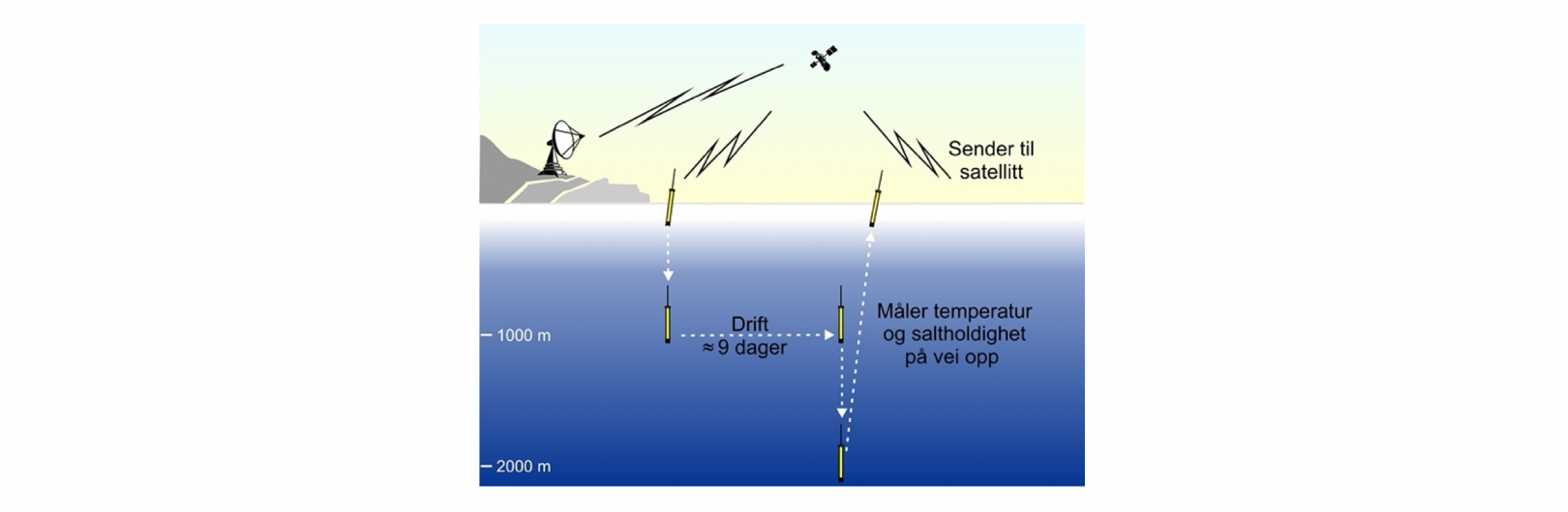
Hydrografiske målinger med CTD («Conductivity, Temperature, Depth»)-sonde har vært vanlig siden 1970-tallet. I tillegg er mange CTD-stasjoner de siste 20 årene tatt med profilerende autonome Argo bøyer (se avsnittet under om Argo bøyer for beskrivelse av Argo). Alle disse dataene gir en god oversikt over de hydrografiske forholdene i Norskehavet, men pga. den komplekse bunntopografien over ryggene oppløses ikke romlige variasjoner der. De fleste stasjonene fra fartøy er dessuten tatt ned til 1000 m dyp mens med Argo bøyer er de fleste tatt ned til 2000 m dyp. Det er flere faste hydrografiske snitt som gjentas flere ganger i året der målingene tas ned 1500 m dyp eller til bunn. Disse snittene vil også brukes for tidsvariasjoner av temperatur og saltholdighet. Siden 1995 har det i mai måned blitt gjennomført et økosystemtokt i samarbeid med andre land (WGINOR) der hele Norskehavet dekkes. Disse dataene sammen med andre CTD-data (fra fartøy og Argo) er griddet og brukt til å lage horisontale kart for temperatur og saltholdighet. Figur 2.2 viser eksempel på stasjoner med CTD-data for mai 2019 der noen faste hydrografiske snitt inngår (røde punkter).
Figur 2.2 Kart viser CTD-stasjoner tatt mai 2019. WGINOR-tokt: blå punkter, andre tokt: røde punkter, Argo data: gule punkter. Bilder: Argo bøye på overflaten før den synker ned i dypet og CTD-sonde som senkes fra fartøy. Foto: Tor de Lange (UiB) og Cecilie Broms (HI).
Argo bøyer er fritt drivende autonome profilerende bøyer som måler hydrografiske data i de øvre 2000 m (Figur 2.2). Mesteparten av levetiden driver de i 1000 m dyp og drivbanene derfra er brukt til å estimere havstrømmene i dypet. Drivbaner for ca. 300 Argo bøyer i perioden 2001-2020 inngår i dette datasettet.
Resultater fra publiserte arbeider basert på enkelte strømmålinger i utredningsområdet blir presentert. Blant annet inkluderer dette strømmålinger fra to rigger sør for Jan Mayen.
Sjøisutbredelse målt med satellitt vil bli presentert for middel og maksimum sjøisutbredelse de siste 20 årene (2001-2020). Data er hentet fra «Climate Data Store» hos Copernicus (https://cds.climate.copernicus.eu/#!/home) og består av to datasett: «European Organisation for the Exploitation of Meteorological Satellites (EUMETSAT) Ocean and Sea Ice Satellite Application Facility (OSI SAF)» og «European Space Agency Climate Change Initiative Phase 2 project (ESA CCI)».
2.3 - Strømforhold
Overflatestrøm
Den norske Atlanterhavsstrømmen består av to grener, en indre gren langs med eggakanten, og en ytre gren, ofte kalt polar- eller frontstrømmen siden den er assosiert med Den arktiske fronten (Figur 2.1 og 2.3). Den indre grenen følger eggakanten (over 500-800 m dyp) nordover som en sterk, ensrettet og smal Atlanterhavsstrøm (Orvik og Niiler 2002). Ved inngangen til Barentshavet går en gren inn i Barentshavet og en annen gren nordover som Vest-Spitbergenstrømmen inn i Polhavet.
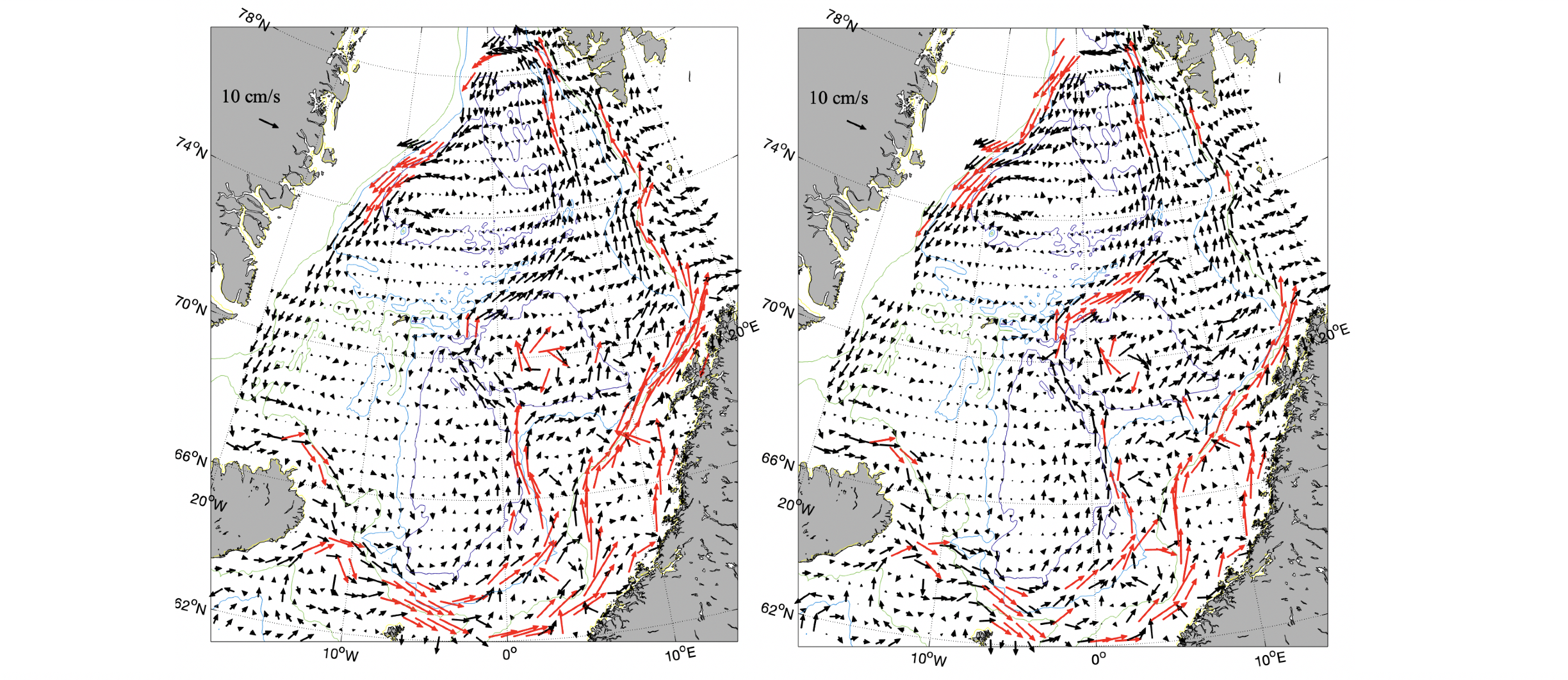
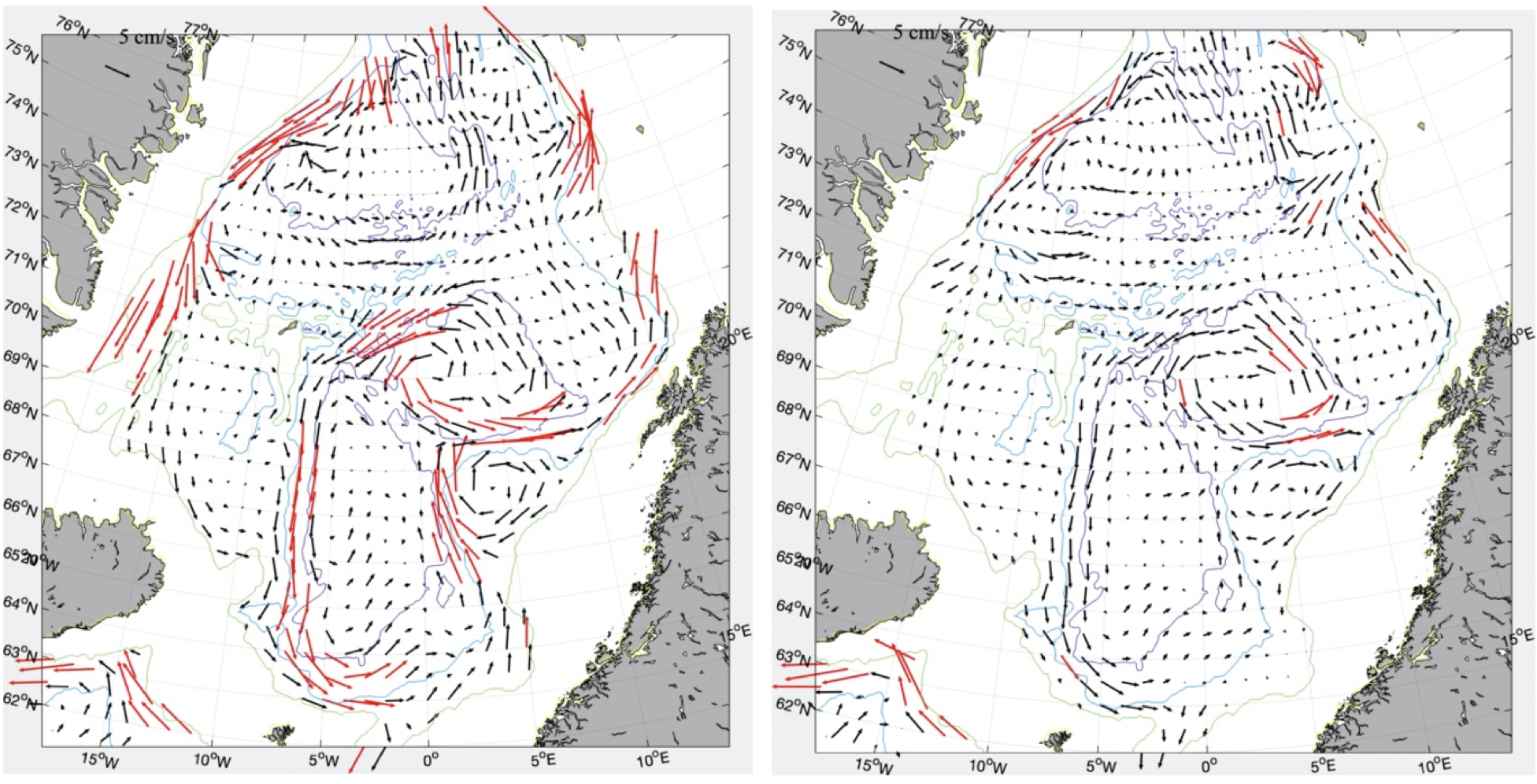
Figur 2.3 Middel overflatestrøm for vinter (desember-april, venstre figur) og sommer (juni-oktober, høyre figur) fra satellittdata (havnivå) 2010-2020. Røde vektorer er hastighet større enn 10 cm/s.
Den ytre grenen følger ca. 2000 m dybdekonturen i sørlige Norskehavet og på vestsiden av Vøringplatået. Nord for Vøringplatået deler den seg i to grener der den ene grenen går østover nord for Vøringplatået mens den andre grenen har en mer vestlig retning mot Jan Mayen. Øst for Jan Mayen strømmer mesteparten av dette vannet nordøstover på østsiden av Mohns-ryggen og følger da Mohns- og Knipovichryggen nordover. Vest for Spitsbergen vil noe av dette vannet resirkuleres og gå inn i Grønlandshavet mens resten fortsetter sammen med den indre grenen inn i Polhavet. Typiske middelhastigheter er 10-30 cm/s (Figur 2.3) med noe sterkere strøm om vinteren. Tilsvarende strømforhold er observert med overflatedriftere i 15 m dyp, men med hastigheter større enn 30 cm/s på vestsiden av Vøringplatået (Orvik og Niiler 2002, Koszalka mfl. 2011). Sterkest strøm ble målt i Lofotenbassenget utenfor Lofoten der farten oversteg 100 cm/s (Anderson mfl. 2011).
Dypstrøm
Dypstrømmene i De nordiske hav ble estimert med drivbanene (se Figur 2.4 og 2.5) til 61 Argo bøyer av Voet mfl. (2010). Datagrunnlaget er nå større og inneholder drivbaner, i 1000 m dyp, til ca. 300 Argo bøyer. Dataene er fra 2001-2020 og strømvektorene er griddet for vinter (desember-april) og sommer (juni-oktober, Figur 2.5). I de dype bassengene i Norskehavet, det sørlige Norskehavsbassenget og det nordlige Lofotenbassenget har strømmen en tydelig syklonisk sirkulasjon (retning mot klokken). Strømmene er svakere i de flate og indre områdene av bassengene sammenlignet med ytterkantene av bassengene (Figur 2.5). I ytterkant av bassengene er hastighetene 5-10 cm/s vinterstid og <=5 cm/s sommerstid. Variasjoner i vindstressvirvlingen (antall/styrken på lavtrykk) påvirker sirkulasjonen i dypet der økt lavtrykksaktivitet gir sterkere sirkulasjon (Voet mfl. 2010, Mork mfl. 2014). Det er verdt å merke seg at på østsiden av Mohnsryggen går strømmene i dypet motsatt vei (sørlig retning) enn de gjør i overflaten (nordlig retning, sammenlign Figur 2.3 og 2.5).
Figur 2.4 En syklus til en Argo bøye. Driften i dypet kan brukes til å estimere strømmen der. En Argo bøye kan utføre ca. 150 sykluserFigur 2.5 Vinter (venstre) og sommer (høyre) strømmer i dyphavet (1000 m dyp). Røde piler er hastighet større enn 5 cm/s.
Virvler
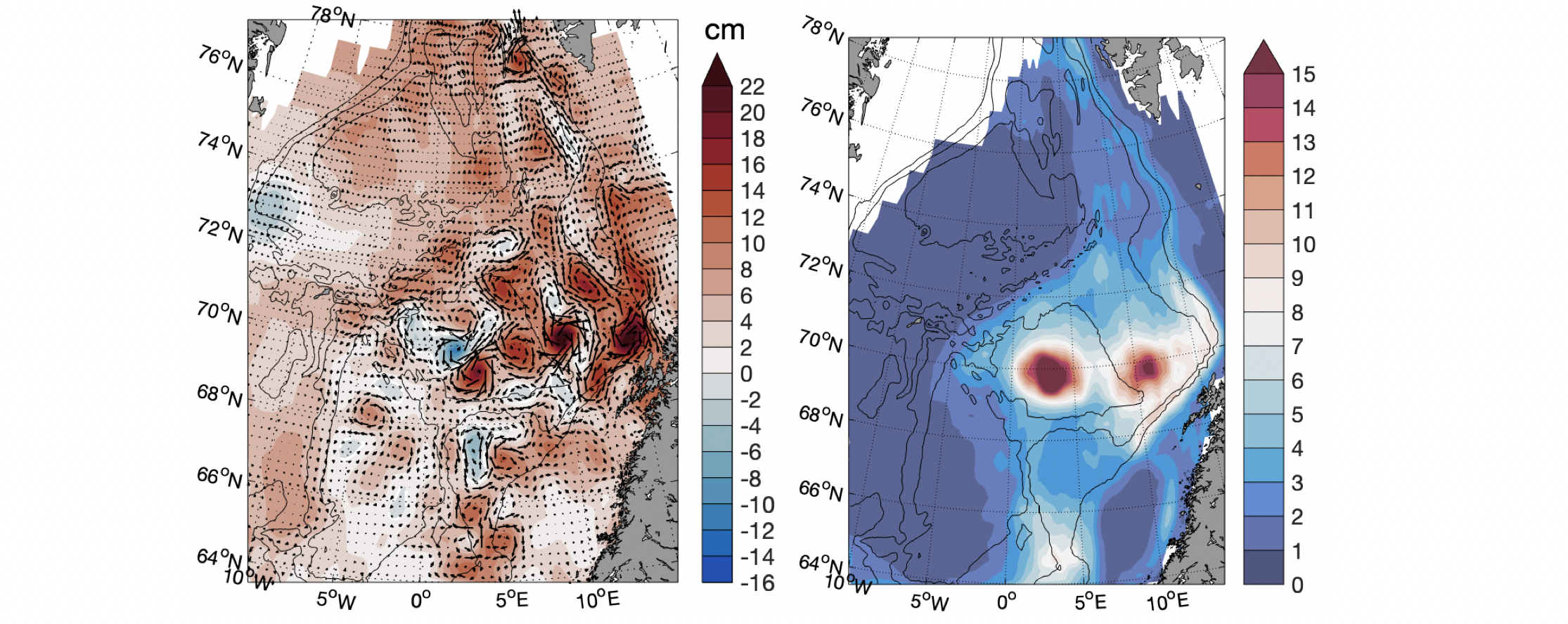
I havstrømmer kan det oppstå virvler på grunn av strømskjær (strømhastigheten endrer seg på tvers av strømretningen), for eksempel ved at strømmen blir påvirket av topografi som kyst eller undersjøiske fjell. Noen virvler kan vare i flere måneder før de blir oppløst. Virvler kan derfor forflytte seg over lengre distanser og inneholde vannmasser med andre egenskaper, fra der de ble generert, enn det utenforliggende vannet. Det er flere virvler i Lofotenbassenget enn i de andre havområdene i De nordiske hav (Koszalka mfl. 2011). En grunn til dette er at virvler genereres når den indre grenen av Den norske atlanterhavsstrømmen passerer langs den bratte skråningen utenfor Lofoten, og disse virvlene driver vestover inn i Lofotenbassenget. Det er både sykloniske (retning mot klokken) og antisykloniske (retning med klokken) virvler i Norskehavet (Raj mfl. 2016). En spesiell virvel er Lofoten virvelen som er en permanent antisyklonisk virvel i sentrum av bassenget (ca. 70N, 3E) som har atlantisk vann ned til 1000-1200 m dyp (Søiland og Rossby 2013). Den har en diameter på 30-40 km og maksimum orbital hastighet på 0.8 m/s. Eksempler på virvler i Norskehavet for et tidspunkt er vist i Figur 2.6a. Figur 2.6b viser høyere virvelaktivitet i Lofotenbassenget enn i de andre havområdene i De nordiske hav. Over undersjøiske fjell og banker kan det dannes topografiske strømvirvler slik at vannmassene oppholder seg der over lengre tid. Dette er tilfellet for Schulzbanken (07°29´E, 73°51´N), der toppen er på 560 m dyp, i overgangen mellom Mohns- og Knipovichryggen, hvor det er observert retensjon av partikler over banken (Roberts mfl. 2018).
Figur 2.6 a) Anomali av havnivå med strømvektorer 2. Mars 2020. Virvler med høyt (lavt) havnivå har antisyklonisk (syklonisk) rotasjon. b) Middel virvel («eddy») kinetisk energi (10-3 m2 s-2) fra altimetri data (2010-2020).
Strømmålinger fra rigger
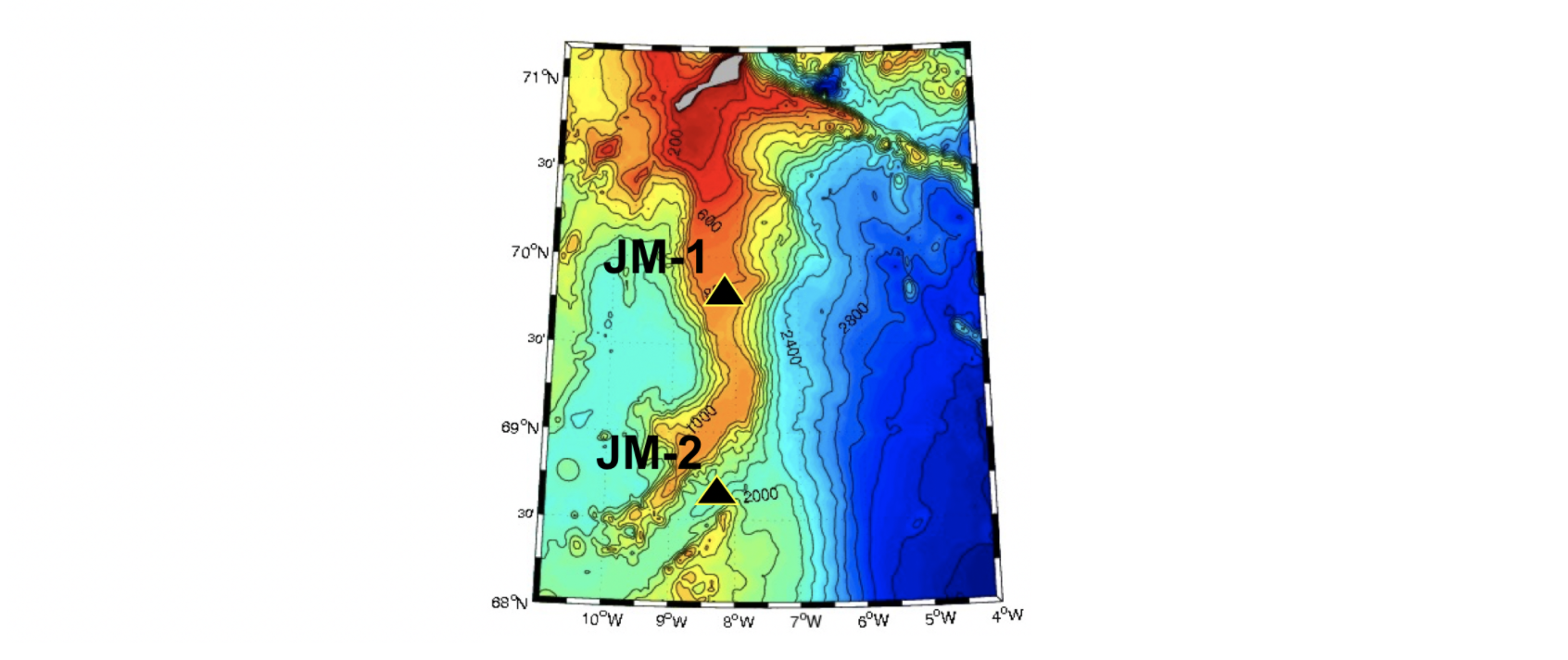
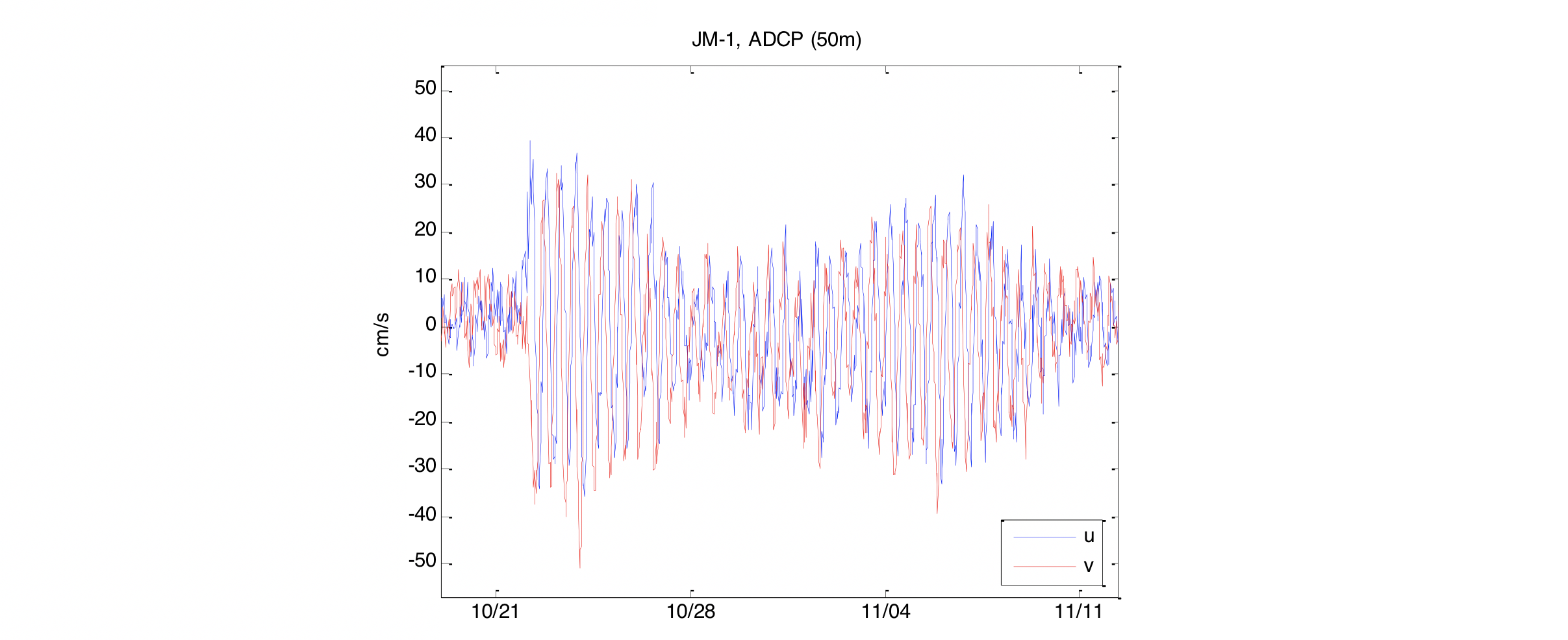
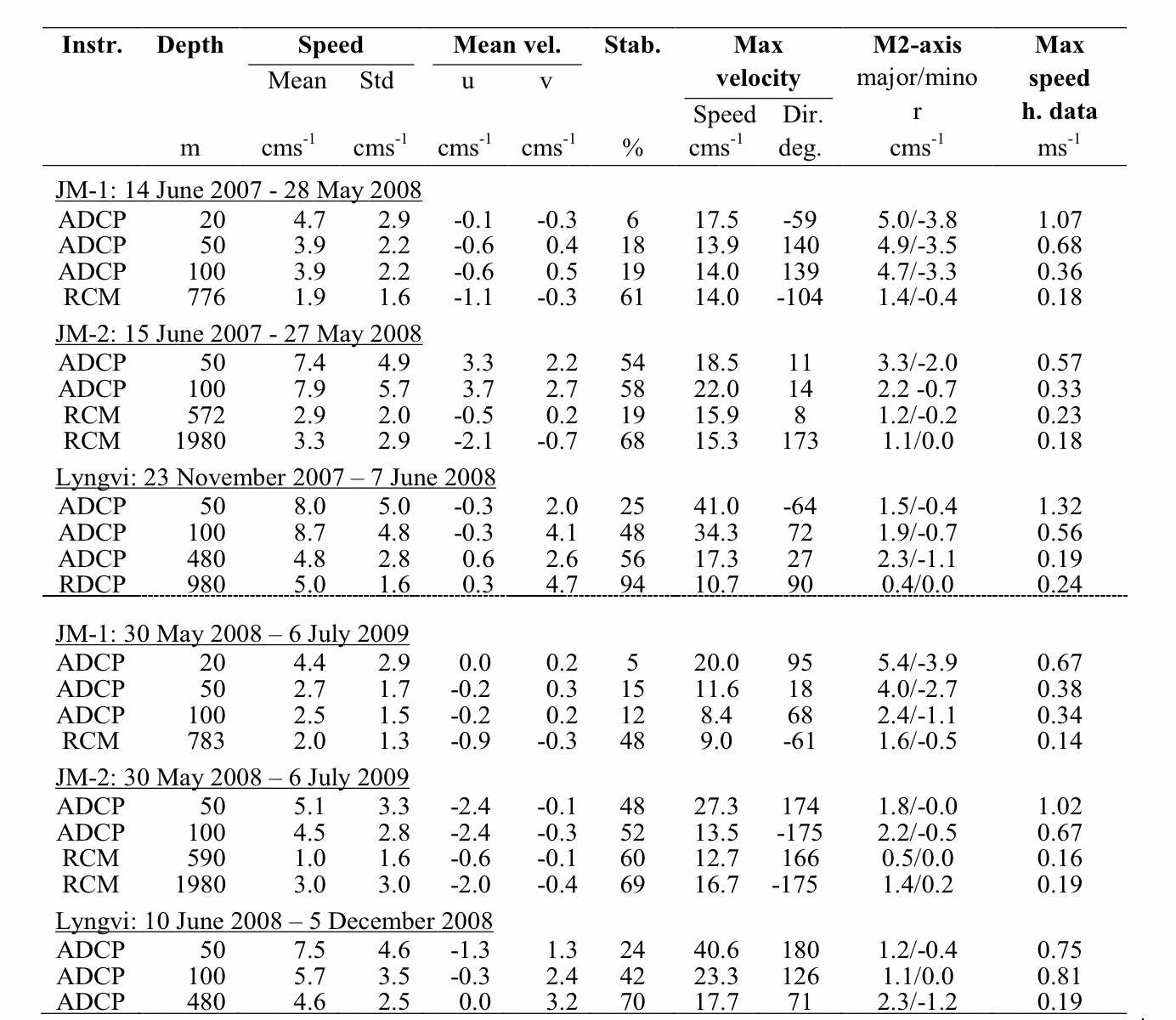
I forbindelse med det internasjonale polaråret (IPY) 2007-2008 ble det satt ut to strømrigger på Jan Mayen ryggen (Figur 2.7), en nordlig rigg på 800 m dyp (JM-1) og en sørlig rigg i en 2000 m dyp kanal (JM-2). Statistiske størrelser (middel og maks strøm, tidevann, etc.) fra strømmålingene er gitt i tabell 1 i Appendiks. Resultatene fra disse målingene har blitt presentert i Jan Mayen rapporten (Fossum mfl. 2012) og i Mork mfl. (2014) og bare hovedresultatene gjengis her. Tidevannskomponenten M2 (halvdaglig periode) er den dominerende tidevannskomponten i dette området (Padman og Erofeeva 2004). Amplituden til M2 varierte fra 5 cm/s i øvre lag til ca 1 cm/s ved bunn (appendiks tabell 1). Strømmene over Jan Mayen ryggen var relative svake og strømmen skiftet ofte retning der retningen i de øvre 50 m var sterkt påvirket av de lokale vindene. Selv om middelstrømmen var svak kunne det i perioder registreres strøm på over 1 m/s i øvre lag forårsaket av atmosfæriske forhold. Lavtrykksepisoder forårsaket at strømhastighetene kom i svingninger med periode på en halv dag. Dette er vist i Figur 2.8 der et lavtrykk passerte over området 21-22 oktober 2007.
Undervannsbilde av sandbunn på 1000 m dyp med riflet mønster etter sterk strøm. Foto: Mareano
Figur 2.7 Plassering av Jan Mayen riggene for strømmåling i 2007-2008.
Figur 2.8 Hastighet i 50 m dyp ved JM-1 for oktober-november 2007. (u, v) = (øst, nord).
Turbulens/vertikal miksing
Naveira Garabato mfl. (2004) estimerte den turbulente miksingen i De nordiske hav. Miksingen var sterkt påvirket av lagdelingen der svakere lagdeling ga sterkere miksing. Den var sterkere under 2000 m der lagdelingen er mindre enn i 500-1500 m dyp. I 500-1500 m dyp var miksingen i Norskehavet som regel liten (~10-5 m2 s-1) mens den var en størrelsesorden høyere i Grønlandshavet (~10-4 m2 s-1). Under 2000 m dyp var den vertikale miksingen sterkere (3×10-4 - 10-2 m2 s-1) og den var spesielt stor over Mohnsryggen (~5×10-3 m2 s-1).
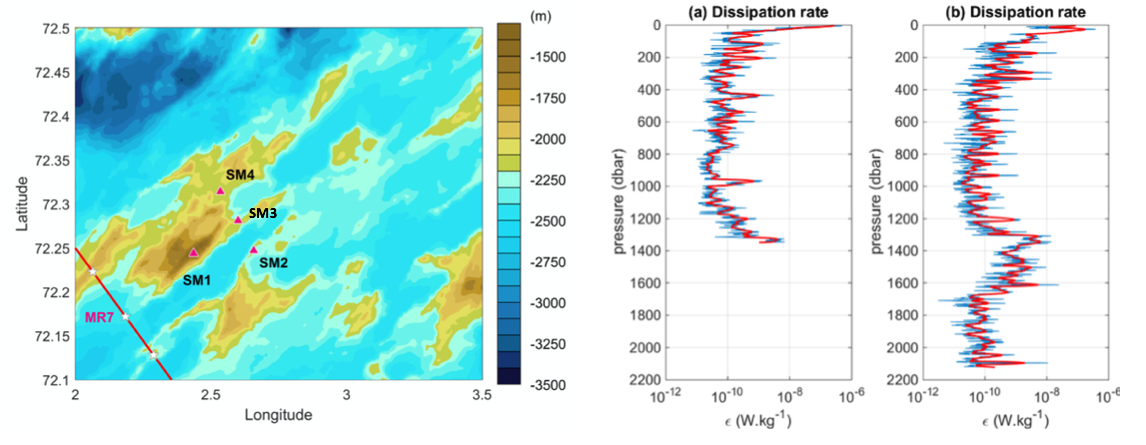
Flere mikrostrukturprofiler ble tatt i Norskehavet på et tokt med F/F Håkon Mosby (26 mai-15 juni, 2016) for målinger av dissipasjon (energitap pga. turbulens). Målingene ble gjennomført blant annet over Mohnsryggen (se Figur 2.9 for posisjonene). Resultatene viser at det undersjøiske fjellet forsterker turbulensen på det samme dypet som den høyeste fjelltoppen i området, som ved SM3 der dissipasjonen viser høyere verdier flere 100 m over bunn (Figur 2.9, høyre figur (b), SM3). Strøm og hydrografiske målinger ved Schulzbanken viste også økt turbulens og vertikal miksing som følge av brytning av indre tidevannsbølger (Roberts mfl. 2018, Hanz mfl. 2021). Observasjoner av høye konsentrasjoner 3He, fra hydrotermale kilder, i de øvre 500 m over Mohns- og Knipovichryggen indikerer også økt vertikal miksing eller strømninger der (Stensland mfl. 2019).
Figur 2.9 Venstre: posisjonene til mikrostrukturprofiler over Mohnsryggen. Høyre: profiler av dissipasjon ved (a) SM1 og (b) SM3.
Disse studiene viser at den komplekse bunntopografien langs med Mohns- og Knipovichryggen sammen med sterk frontstrøm langs med ryggen bidrar til økt turbulens og miksing i dypet. Kunnskapen om hvordan strømforholdene, turbulens og miksing påvirkes av frontstrømmen og/eller indre tidevannsbølger i utredningsområdet er derimot begrenset.
2.4 - Vannmassefordeling
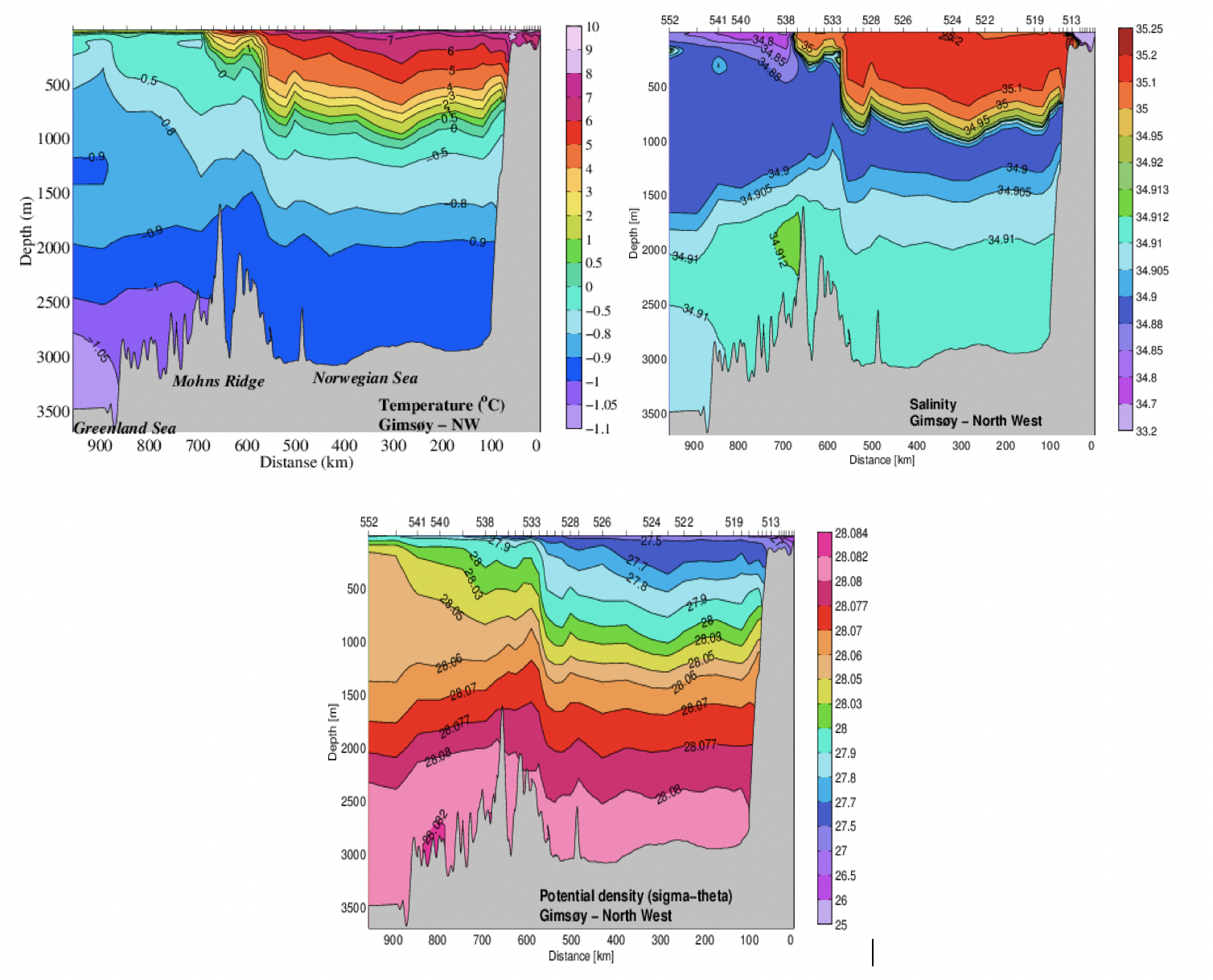
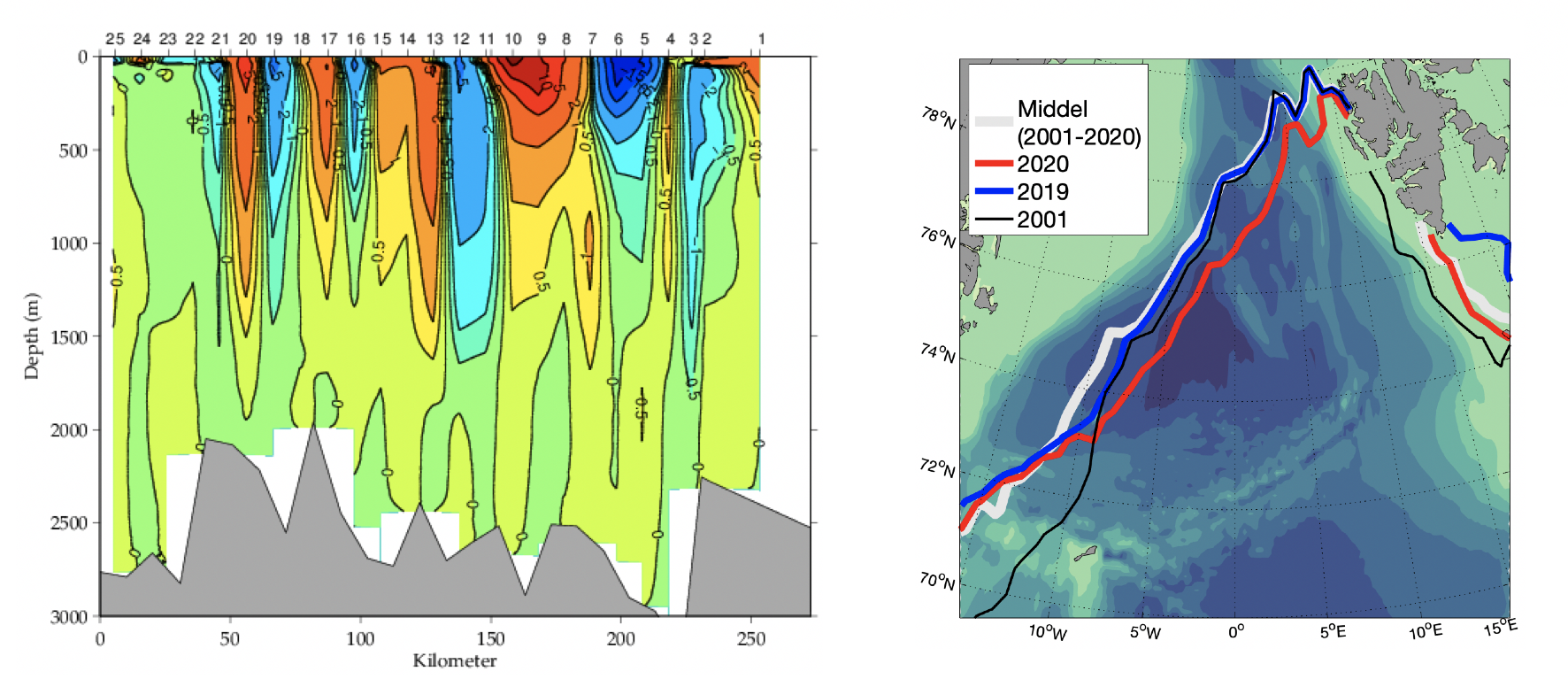
Hvordan det varme atlantiske vannet spres inn i Norskehavet kan sees i Figur 2.1. I sør er det atlantiske vannet ca. 10°C og det avkjøles nordover og vestover ved at det mister varme til atmosfæren og innblanding av kaldere vannmasser. Fordelingen av de forskjellige vannmassene i Norskehavet, vertikalt og over bassengene, kan sees i et hydrografisk snitt som går nordvest fra Lofoten, over Lofotenbassenget og Mohnsryggen og inn i Grønlandshavet (Figur 2.10, se Figur 2.2, røde punkter, for lokalisering av snittet). I Lofotenbassenget ligger det atlantiske vannet, saltholdighet større enn 35, ned mot maks 800 m dyp. Under det atlantiske laget ligger det arktisk intermediære laget med temperaturer rundt nullgraden og som best kan observeres ved et minimum i saltholdighet i ca. 900-1300 m dyp. Over tid har saltholdigheten i det intermediære laget endret seg slik at det enkelte år ikke har et klart saltholdighetsminimum i Norskehavet. Under dette laget ligger dypvannet der temperaturen er under null grader. På vestsiden av Mohnsryggen i Grønlandshavet er temperaturene i hele vannkolonnen, med unntak av et overflatelag, under null grader. Vannet her har også lavere saltholdighet enn i Norskehavet. Påvirkningen av Mohnsryggen på de hydrografiske forhold kan sees i hele vannkolonnen med en skarp arktisk front over ryggen. Her endres temperaturen med flere °C over ca. 20 km (avstand mellom to CTD-stasjoner).
Figur 2.10 Temperatur, saltholdighet og sigma-theta i forlenget Gimsøy-NV snitt mai/juni 2002.
Dybden på det atlantiske vannet varierer fra område til område. Fra de griddete CTD-data for mai måned er dybden av det atlantiske laget beregnet (Figur 2.11a). Det Atlantiske laget er dypest i sentrum av Lofotenbassenget (ca. 800 m) og blir gradvis grunnere mot Mohnsryggen og sørover mot Norskehavsbassenget. Laget har laveste tykkelse i de sørvestlige områdene av Norskehavsbassenget. Det skyldes Øst-Islandsstrømmen som medfører at Arktisk vann er mer fremtredende der.
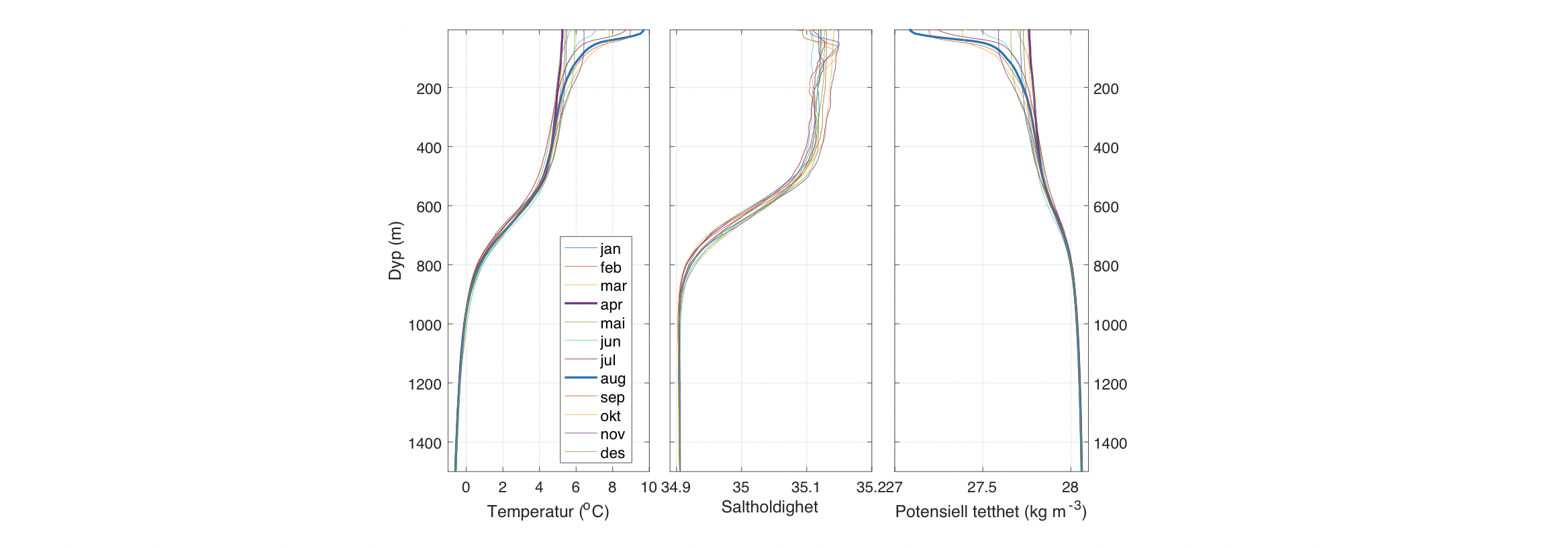
Figur 2.11. a) Midlere dybde på det atlantiske laget (saltholdighet>35). Middel b) temperatur og c) saltholdighet i 300 m dyp. Dataene er middel for mai måned (1995-2019).2.12 Månedlig vertikalprofiler for temperatur, saltholdighet og tetthet i vestlig del av Lofotenbassenget langs med Mohnsryggen.
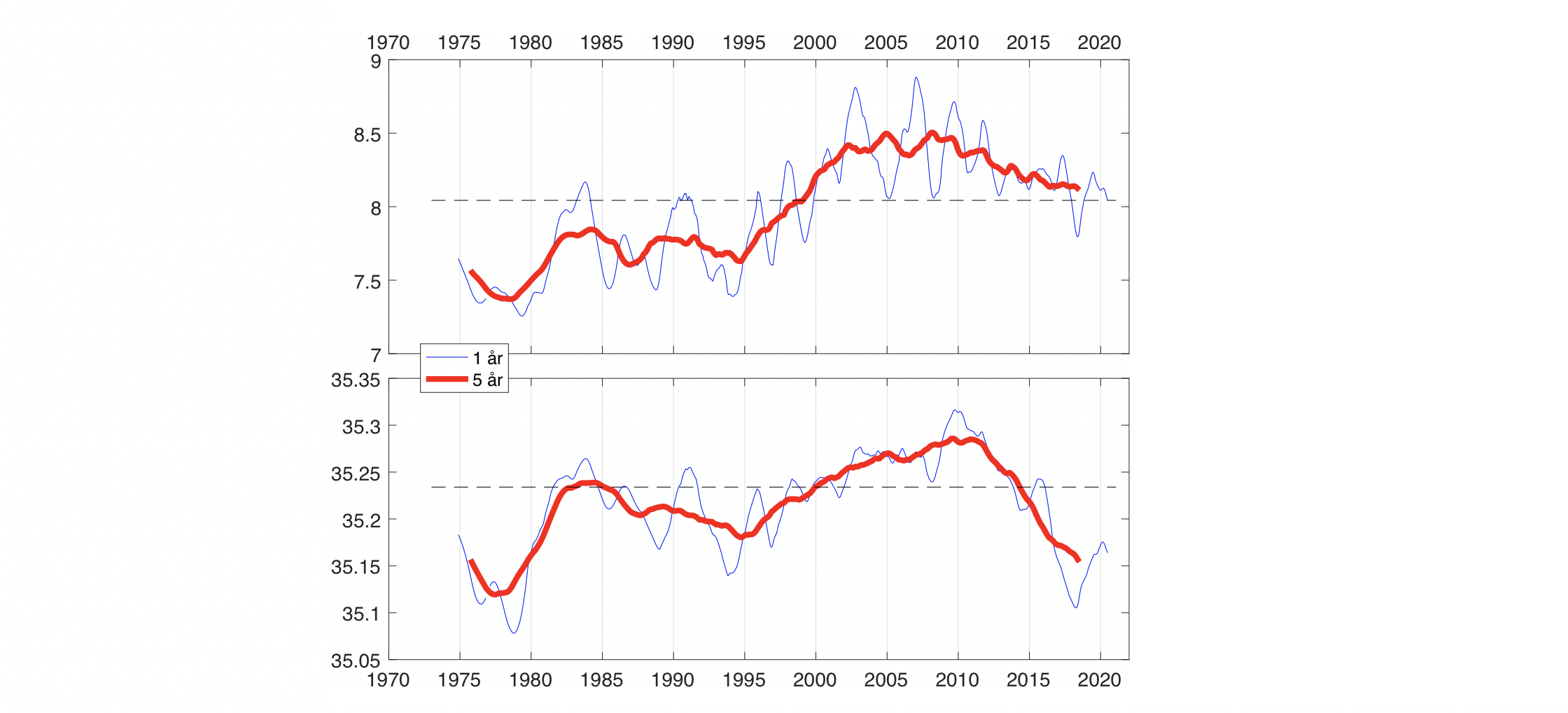
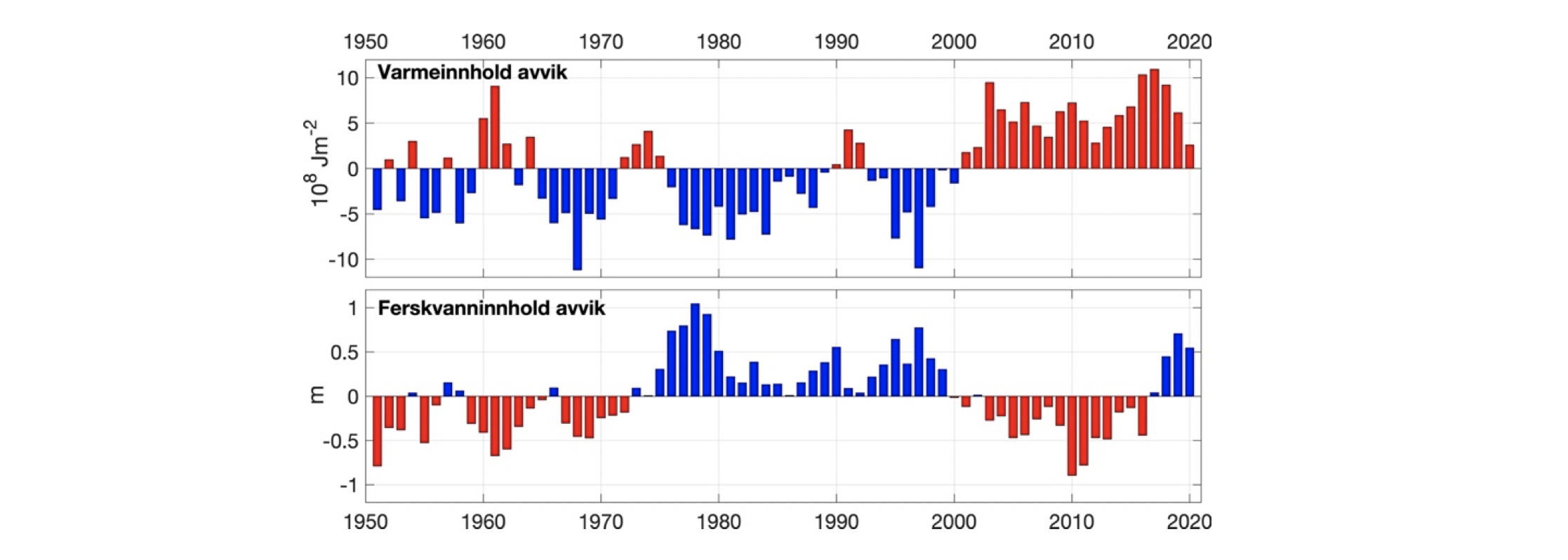
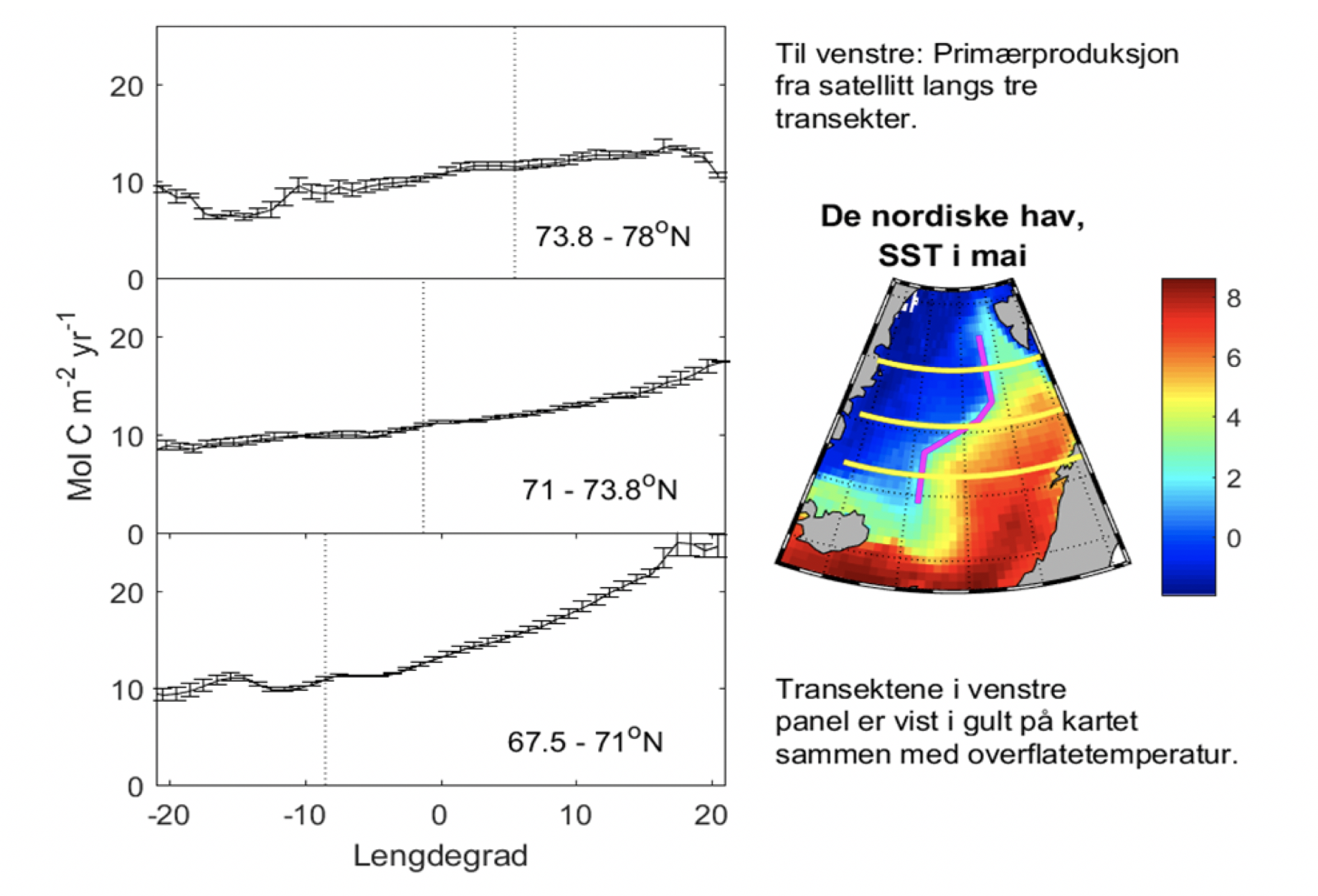
Den horisontale fordeling av atlantisk og arktiske vannmasser kan sees i Figur 2.11b,c der temperatur og saltholdighet i 300 m dyp er vist. Det er varmere og saltere vann over Vøringplatået og vannet blir raskt kaldere og ferskere vestover. Det er store gradienter på vestsiden av Vøringplatået og over Mohns og Knipovichryggen mens det er mindre gradienter ved Jan Mayen Ryggen. I Grønlandshavet og Islandshavet er det vann med temperaturer under 0°C (stiplet hvit linje). Om vinteren blir det øvre laget utsatt for sterk avkjøling og kraftige vinder som medfører at vinterblandingen rekker ned til ca. 300 m og 600 m dyp for henholdsvis Norskehavs- og Lofotenbassenget (Nilsen og Falck 2006). Månedlig variasjon av temperatur, saltholdighet og tetthet som funksjon av dyp er beregnet fra Argo data (2001-2020) i den vestlige delen av Lofotenbassenget, langs østsiden av Mohnsryggen. Figur 2.12 viser at vinterstid er vannmassene i de øvre 500 m gjennomblandet med temperaturer på ca. 5°C. I løpet av sommeren dannes et tynt varmt overflatelag (~50 m dyp) med temperaturer på ca. 10°C. Målinger viser at det innstrømmende atlanterhavsvannet langs med eggakanten har blitt varmere siden 1978. På Svinøysnittet (ca. 63° N, 3° Øst, se også Figur 2.2 for lokalisering av snittet) har temperaturen økt med omkring 0,8°C. 2007 var det varmeste året noensinne siden målingene startet (Figur 2.13). Særlig etter 2000 og frem til omtrent 2012 har atlanterhavsvannet vært bemerkelsesverdig varmt og salt. Økningen i temperatur og saltholdighet begynte i midten av 1990-årene og skyldes hovedsakelig storskala endringer i havsirkulasjonen i det nordlige Atlanterhavet. De siste årene har temperaturen hatt en nedadgående trend, og i 2020 var temperaturen nær langtidsmiddelet. Atlanterhavsvannet ble fra midten av 1990-tallet betydelig saltere frem til 2010-2011 men saltholdigheten har siden avtatt. De siste årene det vært oppsiktsvekkende lave saltholdighetsverdier der man må tilbake til slutten av 1970-årene for å finne tilsvarende lave verdier. Basert på de griddete data for mai måned sammen med andre historiske norske og russiske data er varmeinnholdet og ferskvanninnholdet i atlantisk vann beregnet for Norskehavet fra 1951 til 2021. Det har vært både varme og kalde perioder med flere års varighet (Figur 2.14). Siden 2000 har varmeinnholdet vært høyere enn langtidsmiddel (dvs. varmere enn normalt) men har avtatt de siste årene og er nå nær langtidsmiddelet. Ferskvanninnholdet var på det laveste i 2010-2011 (dvs., havet var saltere enn normalt) men har økt siden. Endringene i varme- og ferskvanninnhold skyldes i hovedsak endringer i det innstrømmende atlanterhavsvannet inn i Norskehavet og i hav-atmosfære flukser over Norskehavet.
Figur 2.13 Temperatur (°C) og saltholdighet i kjernen av det innstrømmende Atlanterhavsvannet på Svinøysnittet (nordvest fra Stad, ca. 63°N, 3°Ø, se også figur 3.2 for lokalisering av snittet). 1 og 5 årsmiddel er vist.Figur 2.14 Varmeinnhold og ferskvanninnhold i et område som dekker atlantisk vann i Norskehavet. Oppdatert fra Mork mfl. (2019).
2.5 - Den arktiske front
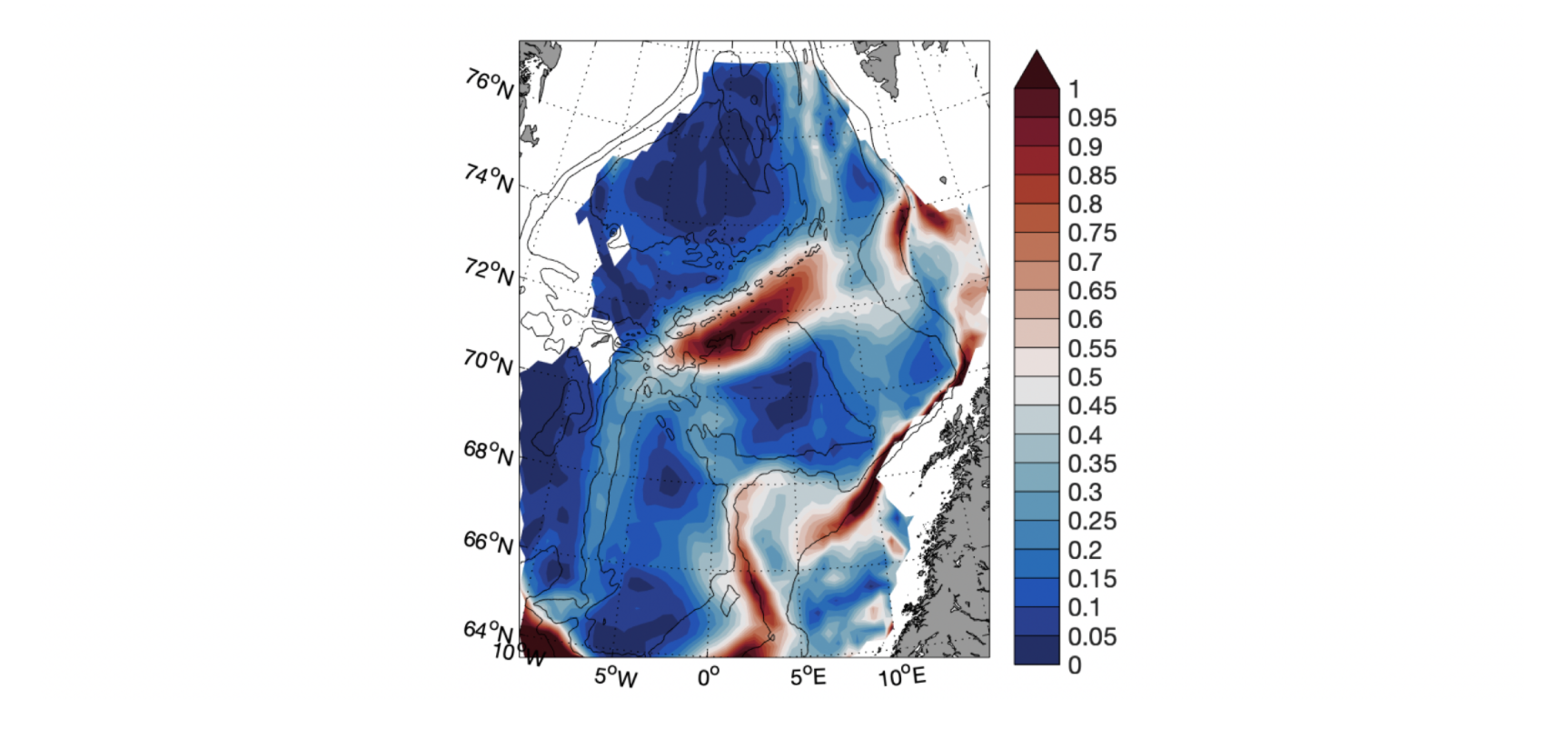
Den arktiske front er definert som grensen mellom varmt Atlanterhavsvann og kaldt Arktisk vann. I Figur 2.15 vises frontområdene i 300 m dyp og hvor tetthetsgradienten er størst (som et mål på hvor skarp fronten er).
Figur 2.15 Tetthetsgradient i 300 m dyp. Større gradient er skarpere front.
I det sørlige Norskehavsbassenget følger fronten 2000 m dybdekoten før den bøyer av mot Jan Mayen etter den passerer vestsiden av Vøringplatået. Nordøst for Jan Mayen sammenfaller fronten med Mohnsryggen og videre nordover med Knipovichryggen. Fronten er sterkest over Mohnsryggen og vest for Vøringplatået. Et frontområde kan også sees vest for Bjørnøya der arktisk vann fra Barentshavet møter den indre grenen av den norske Atlanterhavsstrømmen.
Den Arktiske fronten har en relativ fast lokalisering over ryggene i Norskehavet. De store temperatur- og saltholdighetsendringer på tvers av fronten gir relative sterke strømmer langs med fronten. I kjernen av fronten er strømhastighetene rundt 50 cm/s (Bosse og Fer 2019). Sør for Jan Mayen vil derimot temperatur- og saltholdighetsendringene kompensere hverandre slik at tetthetsendringene her blir mindre (Figur 2.15). Det medfører at strømmene langs med Jan Mayen ryggen er svakere enn strømmen langs med Mohnsryggen (Fossum mfl. 2012, se også Figur 2.3). Siden den arktiske fronten som regel ligger litt øst for Jan Mayen er området rundt Jan Mayen som regel dominert av arktiske vannmasser (Gabrielsen mfl. 1997). Om det er de kalde arktiske eller varmere atlantiske vannmassene som dominerer ved Jan Mayen kan derimot variere i både tid og rom. Denne dynamikken gjenspeiles også til en viss grad i det biologiske mangfoldet der ulike arktiske og nordlige arter befinner seg i området avhengig av hvilke vannmasser som dominerer (Fossum mfl. 2012).
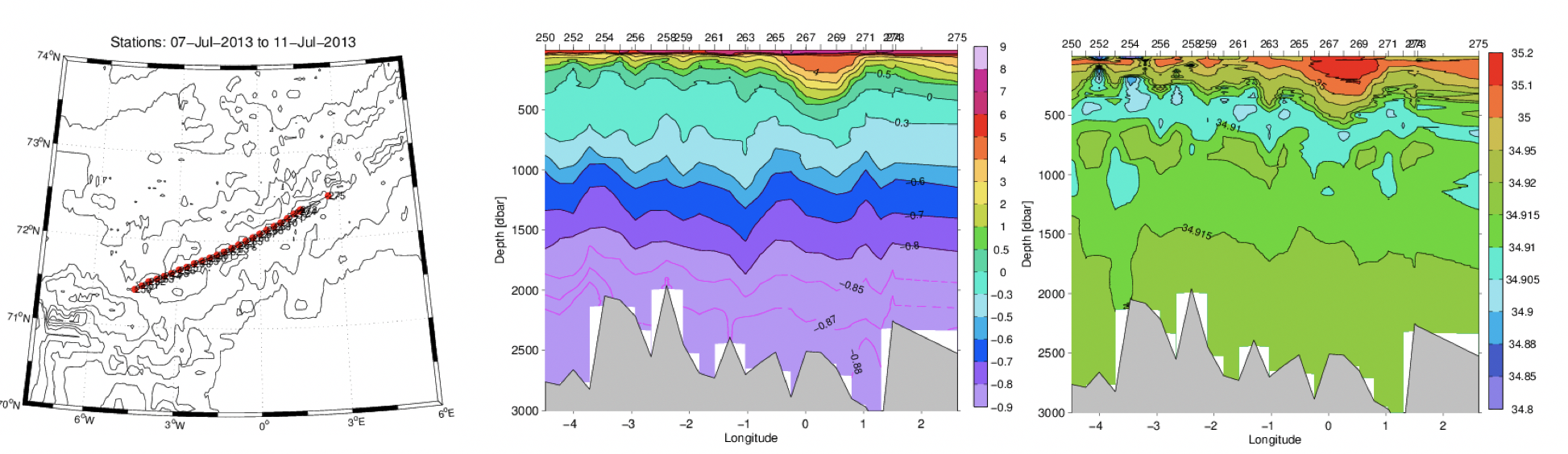
Figur 2.16. CTD-stasjoner, temperatur (°C) og saltholdighet i et hydrografisk snitt over Mohnsryggen juli 2013.
I juli 2013 ble det tatt et hydrografisk snitt over Mohnsryggen der avstanden mellom CTD-stasjonene var relativ liten (ca. 10 km, Figur 2.16). Atlantisk vann kan sees i overflaten, men temperaturen avtar raskt med dypet og under 500 m er vannet kaldere enn 0°C. Spesielt interessant er laget med minimum saltholdighet der saltholdigheten er mindre enn 34.91 mellom ~300-600 m dyp. Dette er Arktisk vann som strømmer fra Grønlandshavet inn i Norskehavet hvor det legger seg som et intermediært lag under det atlantiske laget. Fra de hydrografiske dataene over Mohnsryggen ble geostrofiske hastigheter på tvers av ryggen beregnet. Hastighetene er relativ til bunnhastighetene på tvers av ryggen som antas å være små. Dataene viser et vertikal skjær i strømmen der strømmen under 1500 m dyp er liten og nærmest konstant med dypet (Figur 2.17a). Største hastigheter er i overflaten der hastigheten kan være over 15 cm/s. Det er flere mesoskala virvler i snittet, både sykloniske og antisykloniske virvler som har 20-50 km utstrekning. Til sammenligning estimerte Van Aken mfl. (1995) horisontal utstrekning av virvler til 25-45 km i samme området basert på strømmålinger og CTD-stasjoner.
Figur 2.17 A. Geostrofisk hastigheter (cm/s) over Mohnsryggen. Positive verdier (rød farge) er strøm mot vest (inn i Grønlandshavet) mens negative verdier er strøm mot øst (blå farge). Fargeskalaen er ikke lineær. B. Utbredelse av sjøis for April (15% sjøiskonsentrasjonen er brukt for beregning av utbredelse).
2.6 - Sjøis
Drivisbeltet som dannes utenfor Grønlands østkyst blir kalt «Vesterisen». Området er hovedsakelig påvirket av den kalde sørgående Øst-Grønlandsstrømmen som transporterer is og kalde vannmasser sørover fra Polhavet langs med Grønlands østkyst (Figur 2.1, Helland-Hansen og Nansen 1909). Selv om noe av sjøisen i Grønlandshavet blir dannet lokalt, kommer mesteparten av sjøisen fra Polhavet med Øst-Grønlandsstrømmen. Tidligere kunne isen strekke seg helt til Jan Mayen om våren, men de siste årene har iskanten vært mye lengre vest. De siste tiårene har vært en nedadgående trend i utbredelse av sjøis (Onarheim mfl. 2018). I gjennomsnitt er det maksimum isdekke i april og minimum isdekke i september (Kvingedal 2005). Figur 2.17b viser utbredelse av sjøis for april måned for 2001-2020 middel og årene 2019-2020. Året 2001 er også vist siden dette året har størst østlig utbredelse ved Jan Mayen de siste 20 årene. I 2020 lå iskanten lengre øst i Grønlandshavet enn middelet for de siste 20 årene. Det kan være store endringer fra år til år (f.eks. for 2019 og 2020). Mellomårlige variasjoner i isutbredelse påvirkes i stor grad av endringer i det atmosfæriske trykkfeltet (Deser mfl. 2000).
2.7 - Fremtidsutsikter
Det er forventet at temperaturen vil øke i utredningsområdet som følge av klimaendringer fra økt drivhuseffekt, men naturlige variasjoner vil i stor grad dominere over klimasignalet de neste 10-20 årene (Hanssen-Bauer mfl. 2015). Den globale oppvarming vil sannsynligvis føre til at Atlanterhavsstrømmen inn i Norskehavet svekkes, men observasjonene så langt viser ikke noen tegn på en reduksjon av strømmen (Hanssen-Bauer mfl. 2015). De siste tiårene har det vært en nedadgående trend i utbredelsen av sjøis (Onarheim mfl. 2018) og det er stor sannsynlighet at denne trenden vil fortsette i fremtiden (Portner mfl. 2019).
2.8 - Oppsummering og kunnskapsbehov
Eksisterende hydrografiske data fra fartøy, rigger og bøyer, satellitt data, drivbøyer og strømmålinger gir en god oversikt over hydrografiske forhold og havstrømmene i de pelagiske områdene (åpne vannmassene) av Norskehavet. Observasjonene viser at utbredelse av de forskjellige vannmassene og strømforhold er stort sett bestemt av topografiske forhold. Sterkest strøm er det ved ytterkantene av havbassengene hvor det er størst gradient i bunntopografien. Det mangler derimot detaljerte kunnskaper om hydrografi og spesielt strømforhold nær bunn i de mer komplekse bunntopografiene som Mohns- og Knipovichryggen. Utredningsområdet dekker flere forskjellige vannmasser og i skjæringspunktene mellom disse, ofte over undersjøiske rygger, vil andre fysiske prosesser være mer fremtredende enn i de mer åpne vannmassene. For eksempel vil sterk strøm langs med ryggene og/eller tidevann sammen med grov topografi gi kompliserte strømforhold. Observasjonene viser også at vertikal blanding og strømninger forsterkes over undersjøiske rygger. En mer detaljert romlig miljøbeskrivelse i utvalgte områder vil kreve flere målestasjoner/rigger for innsamling av oseanografiske data og etablering av en havmodell med tilstrekkelig romlig oppløsning i disse områdene.
3 - Kjemisk oseanografi
3.1 - Innledning
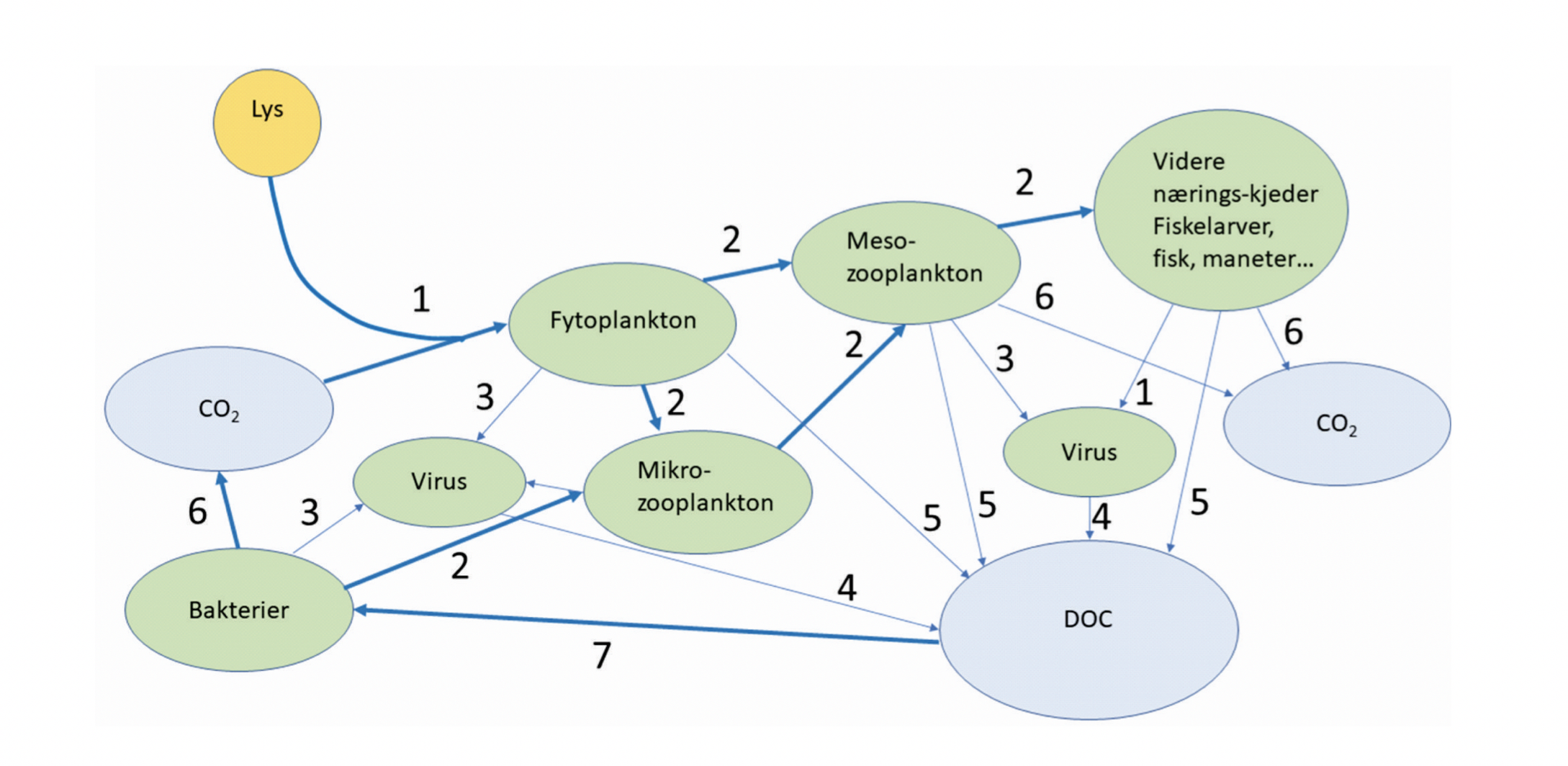
Karbon er naturlig tilstede i atmosfæren som CO2 (karbondioksid). Det finnes også bundet i organisk materiale f.eks. i skoger og jordsmonn. I havet finner man karbon i flere forskjellige uorganiske forbindelser f.eks. kalk, karbondioksid og bikarbonat og som organisk materiale i marine planter og dyr (se den biologiske karbonpumpen, Figur 4.4 i kapittel om plankton). I utgangspunktet eksisterer det en balanse mellom de ulike karbonlagrene. Ligning 1 (nedenfor) viser det marine CO2 systemet og dens kobling til atmosfæren, et syre-base system med målsetting om å holde havets pH basisk, ca. 8 i de øvre vannmassene. Når atmosfærisk CO2 løses i overflatevannet blir det en del av havets CO2 system og danner raskt karbonsyre (H2CO3) og frigjør hydrogenioner (H+). Ettersom havet er basisk frigjøres ytterligere H+ og det dannes bikarbonat (HCO3-) og karbonationer (CO32-). Dette systemet påvirkes av alle naturlige prosesser som endrer dens komponenter, slik som fysiske prosesser (omblanding, sirkulasjon og ferskvannstilførsel), temperatur, biologiske prosesser (produksjon og nedbrytning), og utveksling mellom hav-luft. I De nordiske hav spiller de biologiske prosessene en stor rolle. Det er observert store sesongvariasjoner med lav pH om vinteren og høy pH om sommeren grunnet CO2 opptak i planteplanktonproduksjon. I isdekkede områder, slik som ved Øst-Grønland og i den vestre delen av Fram Stredet, påvirkes karbonatkjemien i tillegg av sjøisprosesser så som smelting og isdannelse.
Havets karbonsyklus og dens rolle i opptak, transport og blanding av karbondioksid (CO2) fra forbrenning av fossilt brensel, sementproduksjon og endring i bruk av land, er et sentralt tema. Siden år 1780 har havene absorbert omtrent en tredjedel av de menneskeskapte karbondioksidutslippene til atmosfæren. En stor del av dette er blitt absorbert i Nord-Atlanteren (Sabine mfl. 2004, Olsen mfl. 2010). Det er grunnet avkjøling av den varme norske atlanterhavsstrømen på sin vei nordover. Ettersom kaldt vann absorberer CO2 fra atmosfæren mer effektivt, vises den høyeste andelen av menneskelig CO2 innehold i De nordiske hav, spesielt i Norskehavet. Det er flere grunner til dette, inkludert kjøling av overflatevannet med samtidig økning i løseligheten av karbondioksid, at vindhastigheten generelt er høy, spesielt i løpet av vintermånedene, og at dypvann dannes ved hjelp av dyp blanding og konveksjon. Det viktigste dypvannsproduksjonsområdet på den nordlige halvkulen er sannsynligvis Grønlandshavet. Når overflatevannet avkjøles og tettheten i Grønlandshavet øker, synker det og transporterer dermed nylig absorbert menneskeskapt CO2 til dype og intermediære vannlag. Disse vannmassene som blir produsert nord for den arktiske delen av midthavsryggene, er en av komponentene i bunnvannet som strømmer sørover inn i Nord-Atlanteren og transporterer CO2 videre til verdenshavene. De arktiske vannmassene som transporteres i Øst-Grønlandsstrømmen har lavere CO2 innhold enn atmosfæren (ca 410 µatm) gjennom hele året, altså det er pCO2 undermettet, hvilket innebærer at det vil ta opp store mengder CO2 fra atmosfæren (Chierici mfl. 2019, Olafsson mfl. 2021). Derfor har Grønlandshavet og Norskehavet en betydning i den globale syklusen av karbon og det videre CO2 opptak. En del av vannet resirkuleres og returnerer til Norskekysten som et dypere vann med høy CO2 innehold noe som gir lav pH og kalkmetning. Hovedopptaket av karbon er synlig i overflaten, i de dype lagene i Framstredet, og i overflatevannet med direkte atlantisk opprinnelse. Hovedutstrømningen av karbon skjer i overflatelaget over Jan Mayen og Knipovich-ryggen, og gir transport av karbon inn i Atlanterhavet via henholdsvis Danmarks-stredet og mot Arktis via Norskehavet. Studier viser en nettotransport av karbon ut av Grønlandshavet om vinteren (e.g. Chierici mfl. 1999) og at det arktiske vannet forsterker CO2 opptaket i nordiske havområder (Olafsson mfl. 2021).
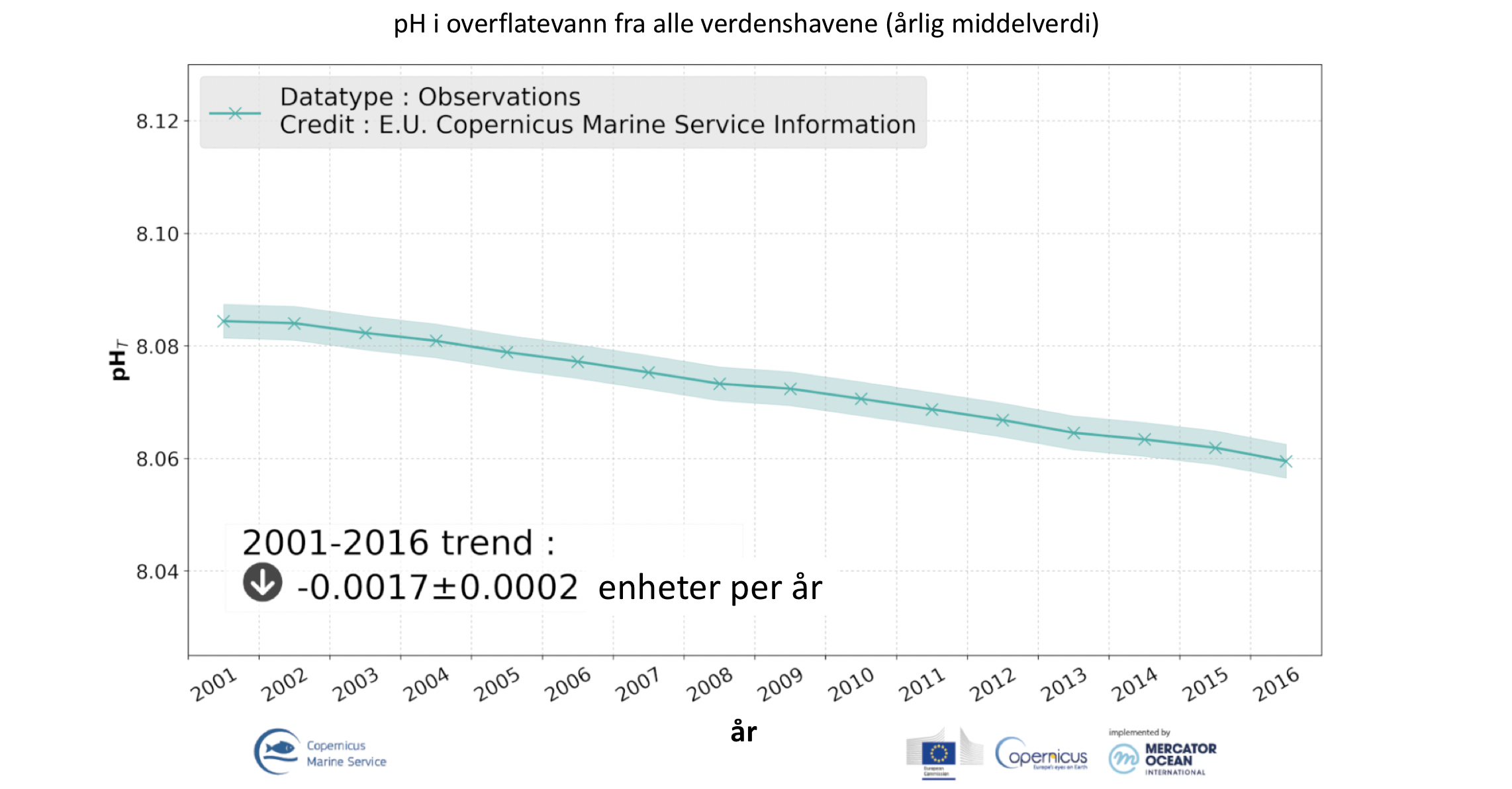
Havet har en pH mellom 7,5 og 8,5 og er basisk. Mengden CO2 i atmosfæren og i havet har økt svært raskt det siste århundret med den konsekvens at den mest basiske komponenten, karbonationer, i havet har minket og havet er dermed blitt «surere» (egentlig, mindre basisk). Dette fenomen refereres til som havforsuring (eng. ocean acidification). Observasjoner ved ulike havstasjoner viser en midlere pH-reduksjon på ca 0,002 per år siden 1985 (Figur 3.1, Copernicus Marine Service), hvilket er den forventede pH-reduksjon basert på CO2 økningen i atmosfæren på ca 2 µatm hvert år. Ettersom økt CO2 i havet gir avtagende pH og redusert antall karbonationer, medfører det en økende kalk-oppløselighet (Ω). Dessuten fører dette til at det dypet hvor kalk løses opp, såkalt kalkmetningsdyp (Ω), stiger i vannkolonnen. Havforsuring vil derfor først og fremst påvike de artene som bygger skjelett eller skall av kalk. Marine arter i faresonen i norske områder vil for eksempel være dypvannskoraller, som vokser langs store deler av norskekysten, og vingesnegl, som blant annet lever i Norskehavet og Grønlandshavet. I tillegg påvirker havforsuring tilgjengelighet og toksisitet av metaller, på grunn av endring i pH og redoks-potensial. Ettersom havforsuringen er et resultat av at havet har tatt og fortsatt tar opp CO2 fra atmosfæren, vil forsuring av havet pågå i lang tid, selv om CO2 utslippene skulle avta (Portner mfl. 2019). Chierici mfl. (2019) viste at det fortsatt er stort opptak av CO2 fra atmosfæren i nord og at det er viktig å følge med på hvor fort denne utviklingen går. De ulike egenskapene til vannmassene i området, med kaldt arktiskt og varmt atlanterhavsvann med en forholdsvis skarp arktisk front i skillet mellom de kalde og de varme vannmassene, vil ha stor betydning for havforsuringstilstanden og kalkoppløsingspotensiale. Arktisk vann er eldre og har allerede høyt CO2 (lav pH og aragonittmettning) (Chierici mfl. 2016), hvilket betyr at økt transport av arktisk vann inn i Norskehavet og langs kysten sannsynligvis vil bidra til lavere metning med kalkmetningsdyp stigende oppover i vannkolonnen over tid.
Figur 3.1 Årlig middelverdier på pH fra 1985 til 2019 i overflaten fra alle verdens hav viser en årlig pH reduksjon med trend på ca 0,0016. EU Copernicus Marine Services
3.2 - Data og metoder
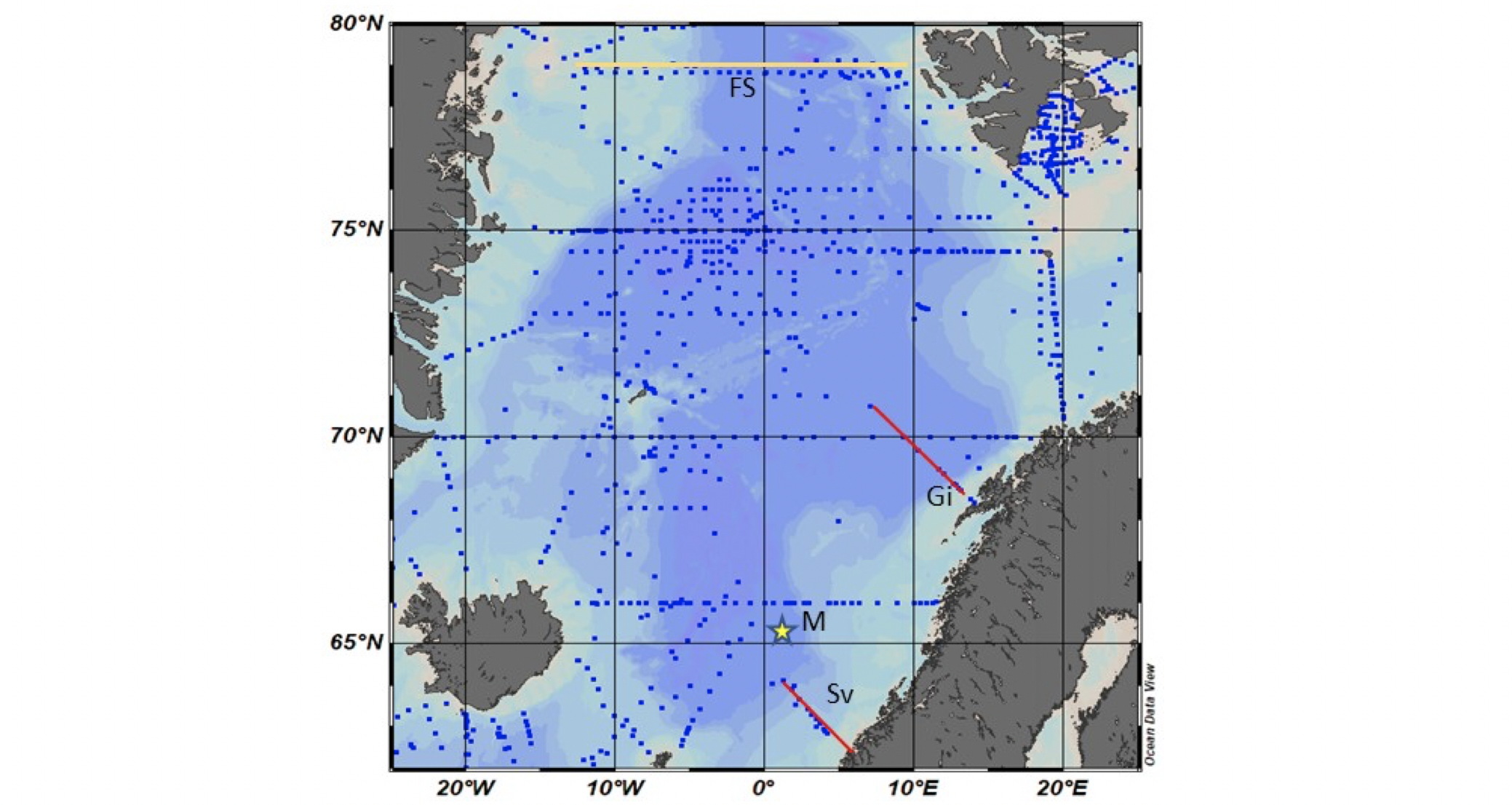
Antall målinger fra Norskehavet er økt siden programmet «Overvåking av havforsuring i norske farvann» ble startet opp i 2012, der undersøkelser av havforsuring blir utført av HI, NIVA, NORCE og UiB på oppdrag fra Miljødirektoratet og presenteres i årlige rapporter på www.miljodirektoratet.no (f.eks. Chierici mfl. 2014, 2020, Jones mfl. 2018, 2020). Overvåkningen er i hovedsak basert på informasjon fra tokt med fartøy der vannsøylen prøvetas en gang i året, i hovedsak vinterstid, langs de faste snittene Svinøy-NV, Gimsøy-NV av HI (Figur 3.2). NORCE utfører vannsøylemålinger ved Stasjon M i sørlige delene av Norskehavet. Gjennom et samarbeid mellom HI og NPI i Framsenterets Flaggskip «Havforsuring og effekter i nordlige hav» samles det inn data for havforsuringsovervåkning i et transekt over Framstredet i Arktis. I Grønlandshavet er det også blitt samlet inn data på enkelte forskningstokt, lagret i databasen GlodapV2, som kan brukes for å beskrive hele karbonatkjemien og alle etterspurte havforsuringsindikatorer som pH og aragonittmetning (Figur 3.2).
Figur 3.2 Kart over stasjoner (blå prikker) presentert i GLODAPv2 (Olsen mfl. 2016), de snitt som prøvetas av HI (røde strekene): Svinøy-NV (Sv) og Gimsøy-NV (Gi) og Stasjon M (M, stjerne, NORCE) som del av havforsuringsovervåkingen, og snittet i Framstredet mellom Grønland og Svalbard som prøvetas av HI og NPI som del av Flaggskipet havforsuring i FRAM (oransje strekk).
Fartøy på faste snitt: Fra CTD-sonde utstyrt med mellom 12-24 Niskin-flasker tas vannprøver fra ulike dyp i vannsøylen sammen med målinger på salt, temperatur og trykk. Analyser av total alkalinitet (AT), løst uorganisk karbon (CT, summen av alle komponentene i marine CO2 systemet, Lign. 1, to av fire mulige parametere) utføres på HI i Tromsø etter internasjonal standard (Dickson mfl. 2007). Data for AT og CT brukes sammen med temperatur, dybde (trykk), saltholdighet, fosfat og kiselsyre i en kjemisk spesieringsmodell, CO2SYS (Pierrot mfl. 2006) for å beregne de andre CO2-systemkomponentene som in situ pH og metningsgrad for kalsitt og aragonitt (ΩCa og ΩAr), og for å estimere kalkoppløsingsdypet (Ω<1).
Autonome sensorer for pH og pCO2: Det er en sterk teknisk utvikling for å forbedre sensorer for å måle pH og pCO2 på bøyer, rigger og andre autonome plattformer. Det finnes flere pH og pCO2 sensorer og Argo programmet har satt inn noen få «biogeokjemiske» sensorer som måler pH i studieområdet. Disse sensorene kan gi informasjon som er av god oppløsing i tid, men dessverre er nøyaktigheten ikke tilstrekkelig for å brukes til trendanalyse. Utvikling pågår og forhåpentligvis vil det finnes sensorer med tilstrekkelig god kvalitet innen noen år. Det samme gjelder pCO2 sensorer. Det pågår flere prosjekt for å utføre ulike tester og internasjonale interkalibreringer for å forbedre og utvikle disse sensorene.
3.3 - pH og kalkmetning i De nordiske hav
Overvåking i Norskehavet viser en klar variasjon i pH-verdiene gjennom året i de øverste 100 meterne. Denne variasjonen er naturlig og henger i stor grad sammen med den biologiske aktiviteten i havet, som er høyest om våren og sommeren. Når den biologiske aktiviteten er høy blir det mindre CO2 i vannet, fordi CO2 tas opp og bindes gjennom fotosyntesen. Dette gjør at pH-verdien stiger og vannet blir mer basisk (mindre surt). Oppløst CO2 og pH varierer ikke bare gjennom året, det er også en geografisk variasjon i disse parameterne. Ut til ca. 50 kilometer fra land er overflaten påvirket av fortynning med ferskere vann fra kysten. Kystvannet inneholder generelt mindre karbonationer. Lenger ut i havet observeres en tydelig innflytelse av atlanterhavsvann. Dette vannet har generelt et høyere CO2-innhold enn de mer kystnære vannmassene. Ettersom dette vannet også inneholder mer karbonationer (høyere AT), som er basisk, er likevel pH-verdiene høyere enn i kystvannet. Områdene lengst vest i Norskehavet er påvirket av innstrømming av arktiske vannmasser fra Grønlandshavet. Det arktiske vannet har lavere pH og aragonittmetning enn atlantervannet. Generelt er det store geografiske og sesongssmessige variasjoner i pH, CO2 og karbonat i Norskehavet.
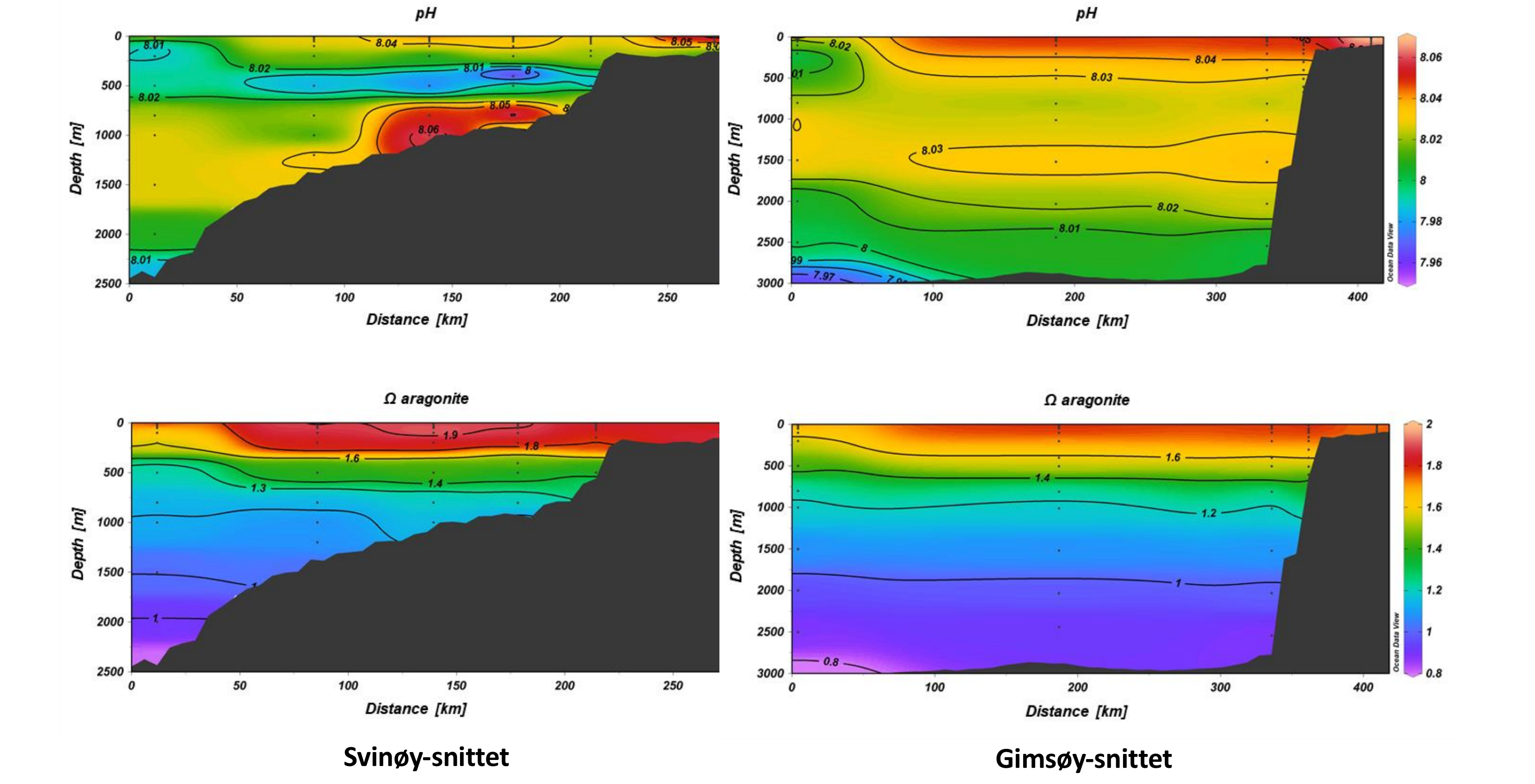
Figur 3.3 Variasjon av pH og aragonitt metning i Norskehavet ved Svinøy-snittet og Gimsøysnittet, fra Norske kysten til Norske bassenget. Figur fra Jones mfl. 2019.
Fra Svinøy-snittet (Figur 3.3) ser vi tydelig påvirkning av kyststrømmen på karbonatkjemien med lave AT og CT verdier ut til ca 50 km fra kysten. I det arktiske vannet er pH og aragonittmetning lavere. Med en økende inblandning av atlanterhavsvann øker pH og aragonittmetningen. CT øker gradvis til høye verdier i atlanterhavsvannet i nord og utover i Norskehavsbassenget. De laveste pH verdiene (ca 8,01) finner man i det dypeste vannet. Aragonittmetning viser relativt homogene verdier i overflaten på ca 1,8 og 2,0. Metning synker gradvis vertikalt og undermetning av aragonitt vises ved 2000 meter ved den nordligste stasjonen. Middelverdi for aragonittmetning langs hele snittet viser 1,7±0,3. I Grønlandshavet er det observert variasjon i parameterne dels fra påvirkning av ulike vannmasser med forskjellig hydrografisk signatur og dels som et resultat av biologisk aktivitet om våren/sommeren. I overflatevannet er CT-verdiene relativt lave og pH høy grunnet biologisk aktivitet. Vannmasser dypere enn ca 500 m viser små variasjoner både for CT og AT. Lenger nord i Norskehavet viser data fra Gimsøy-snittet lignende variasjoner og drivere (i.e. fysiske og biologiske prosesser som påvirker karbonkjemien) som lenger sør. Her ser vi imidlertid større påvirkning fra det arktiske vannet med lavere pH og aragonittmetning i dypvannet og med undermetning på 1900 m dyp, ca 100 m lengre oppe i vannsøylen sammenlignet med langs Svinøy-snittet (Figur 3.3). Denne regionen har mange kaldtvannskorallrev som potensielt er spesielt følsomme for vann med lav pH og lite metning av kalsiumkarbonatmineraler.
Undervannsbilde av vingesnegl (Limacina helicina) (A) og kaldtvannskorall (Lophelia pertusa) (B). Foto: Erling Svensen og Jan Helge Fosså
3.4 - Fremtidsutsikter
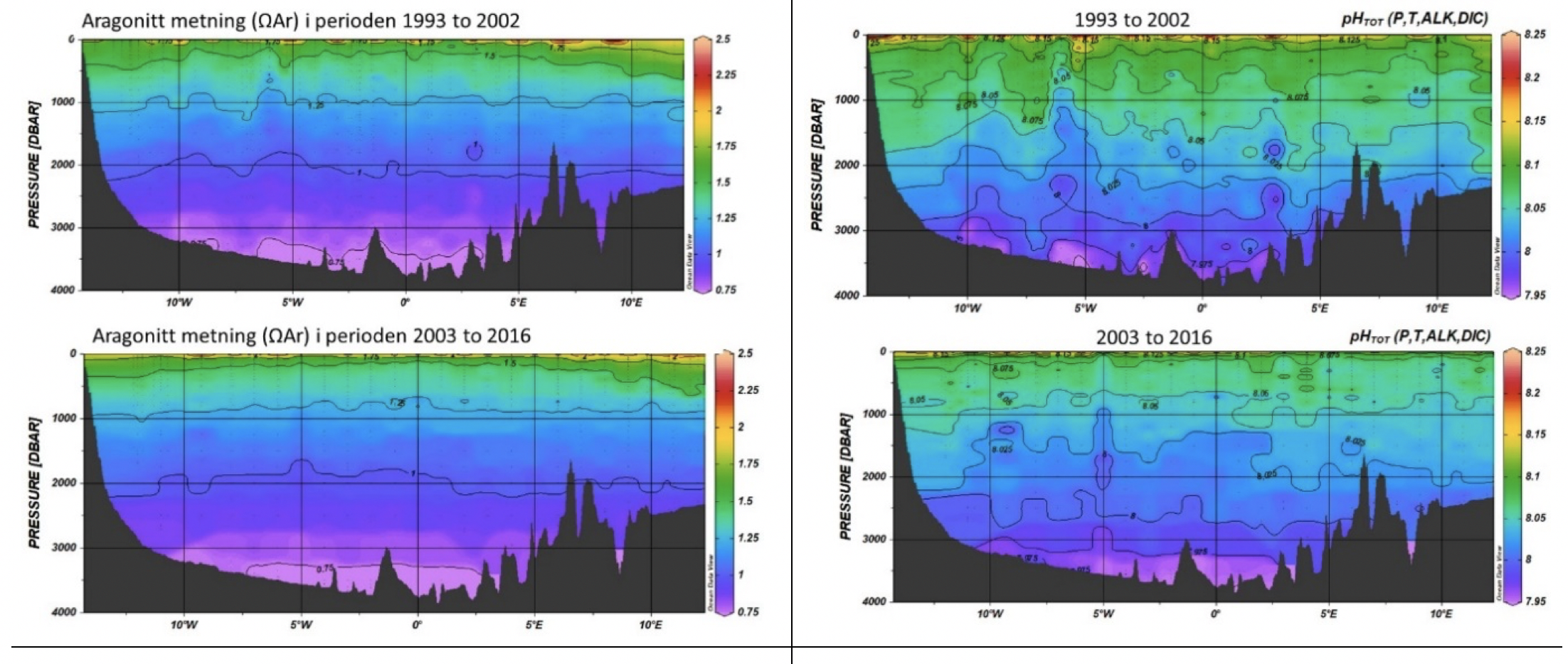
3.4.1 - Grønlandshavet
Observasjoner langs snittet 75°N (basert på data i GLODAPv2) viser at metningshorisonten for aragonitt i Grønlandshavet er lik den i Norskehavet. Undermetning ligger omtrent ved 2000 m dyp (Figur 3.4), men noe høyere opp i vannsøylen på vestsiden enn i øst. Forskjellen er sannsynligvis drevet av en større innblanding av arktisk vann med lavere aragonittmetning og resirkulert atlantervann med lav pH og høyt CO2 innhold i vest (Figur 3.4). Integrert over to ulike perioder, 1993 til 2002 og 2003 til 2016, ser vi at aragonitt metningshorisonten (Ω=1) er på ca 1800-2100 m i vest og ca 2100-2200 m i øst. Dessuten ser vi at Ω=1 har flyttet seg oppover i vannkolonnen med ca 100-200 m mellom disse periodene. Det betyr at metningshorisonten stiger i vannsøylen med ca 70 m per tiår. I tillegg er det tydelig at volumet og arealene med vann med kalkoppløsning (Ω<1) har økt mellom 1993 og 2016. Data for pH-verdier mellom de to periodene viser tydelig minkende pH i hele vannsøylen (Figur 3.4).
Figur 3.4 Utvikling av aragonitt metning (venstre) og pH (høyre) i Grønlandshavet langs 75°N i perioden fra 1993 til 2002 (øvre) og i 2003 til 2016 (nedre) basert på GlodapV2 data langs 75°N. Kilde: Chierici/HI
3.4.2 - Norskehavet
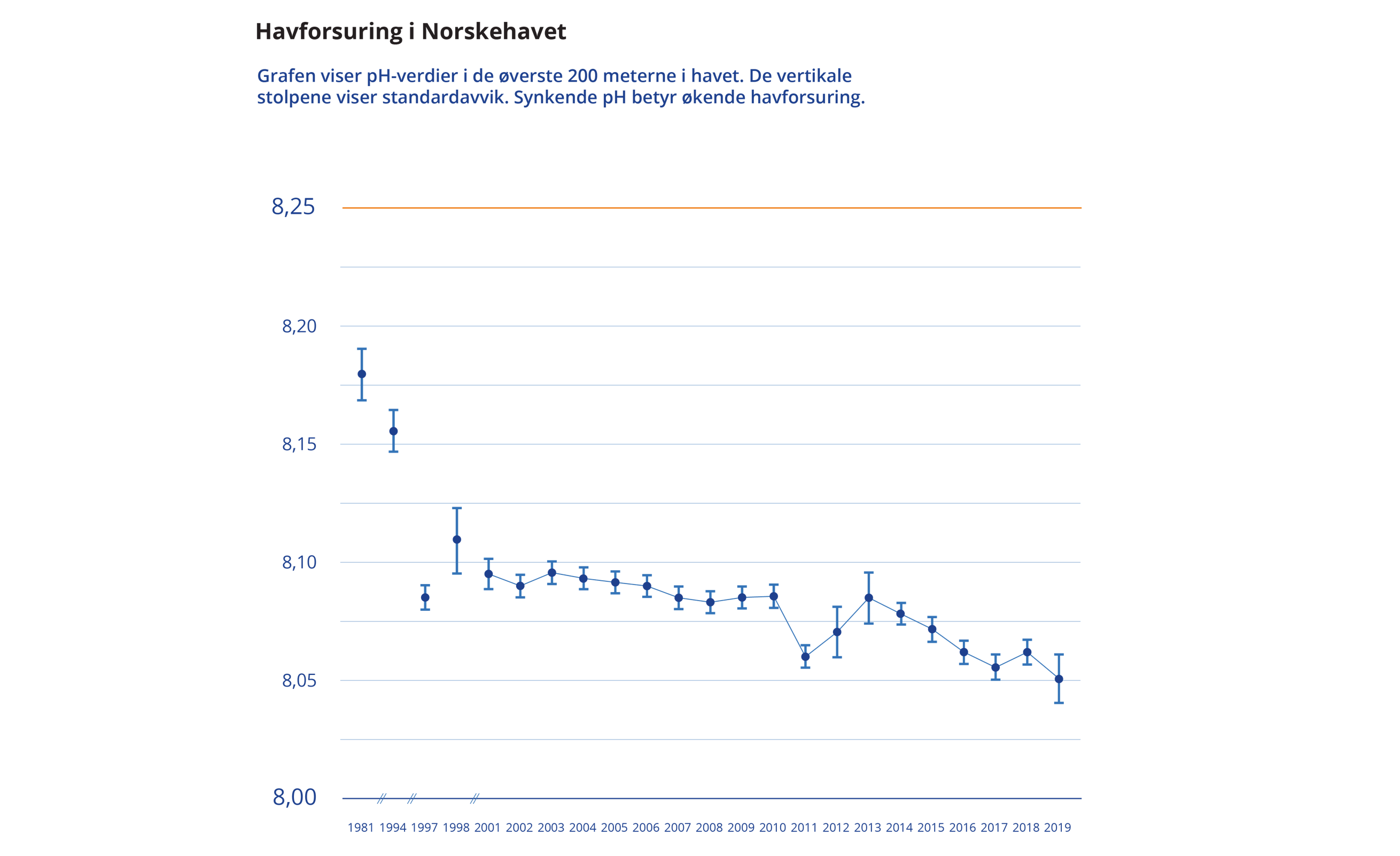
Data fra Havforsuringsovervåkingsprogrammet (www.miljodirektoratet.no) og GLODAPv2 viser at pH-verdien i de øvre 200 m i Norskehavet har sunket med 0,12 pH-enheter de siste 40 årene (Figur 3.5, NORCE/Miljøstatus).
Til sammenligning har pH ved stasjon M i Norskehavet sunket med 0,003 per år og kalkmetningen med 0,014 per år kun de siste 10 årene, i.e i perioden 2011-2019 (Jones mfl. 2020, HI). Det viser at dypbassengene i Norskehavet opplever en raskere pH-reduksjon (-0,003 per år) enn andre deler av Norskehavet (Skjelvan mfl. 2014, Jones mfl. 2018) og det globale gjennomsnittet på -0,002 per år (Figur 3.5, Copernicus Marine Data). I det nordlige Norskehavet er laveste Ω aragonitt funnet på 3120 m dyp. Metningshorisonten for aragonitt ligger her på mellom 1500 m og 2000 m dybde. Dette er mye grunnere enn metningshorisontdybden ved stasjon M og ved Svinøy-snittet i sørlige deler av Norskehavet hvor metningshorisonten ligger på 2000 m.
Figur 3.5 Utvikling av middel år verdier av pH i de øvre 200 m mellom 1985 til 2019 og viser en tydelig synkende pH trend. Kilde: NORCE/Miljøstatus
3.5 - Oppsummering og kunnskapsbehov
Havforsuringsovervåkningen i Norskehavet har vist at både volumet og arealet av vann hvor kalkskjell eller andre kalkformasjoner begynner å gå i oppløsning har økt tydelig de siste 20 årene. Samtidig er det detektert en trend med synkende pH gjennom hele vannsøylen. Reduksjonen er raskere i de øvre vannlag enn i de dype bassengene men også i dypet skjer det en endring. Den dokumenterte pH endringen i Norskehavet er raskere enn i andre åpne havområder. I Grønlandshavet, og også fra sentrale deler av utredningsområdet, mangler man standardiserte målinger av karbonatkjemi fra faste stasjoner, nedover i dypet, tatt på den samme tidsperiode over flere år. Slike lange tidsserier er nødvendig for å påvise trender og for å dokumentere hvor raskt pH, innholdet av karbonationer og aragonittmetningshorisonten forandres med god presisjon. Det råder også store usikkerheter knyttet til de fremtidige effektene av havforsuring på de organismer som lever i havet, både på de organismer som vil bli påvirket direkte ved at det for eksempel vil kreve mer energi å bygge skall av kalk og de som vil bli påvirket indirekte gjennom endringer i næringsnettet.
4 - Plankton
4.1 - Innledning
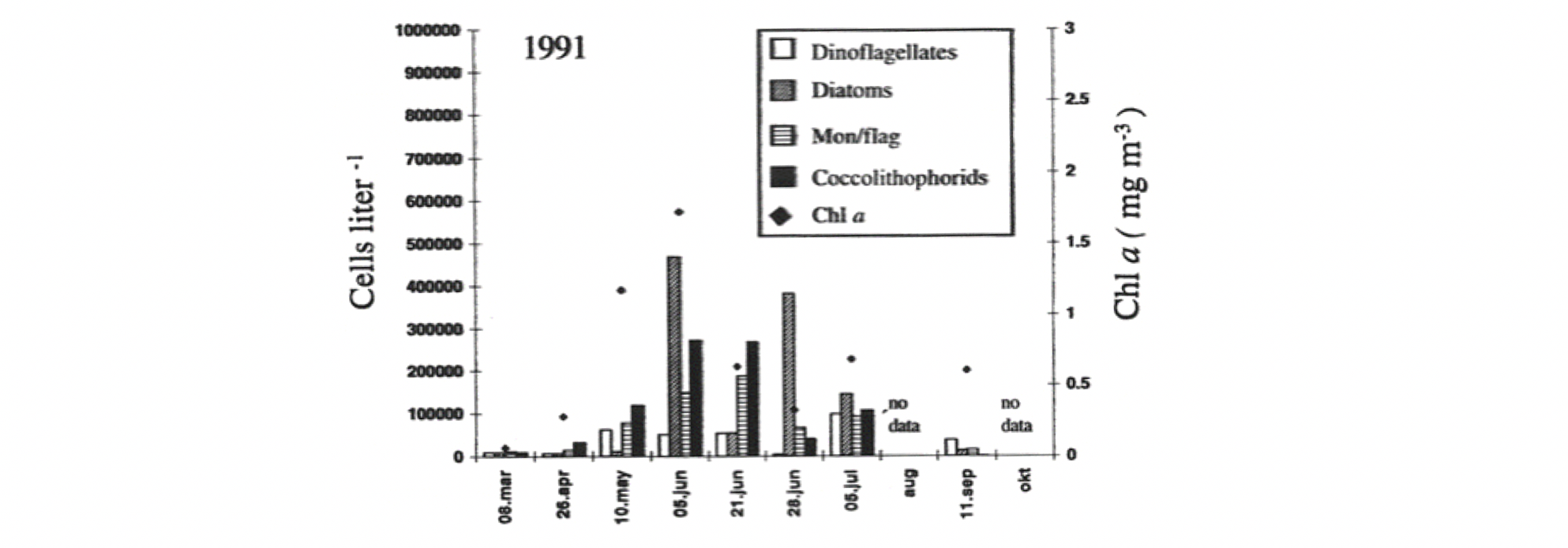
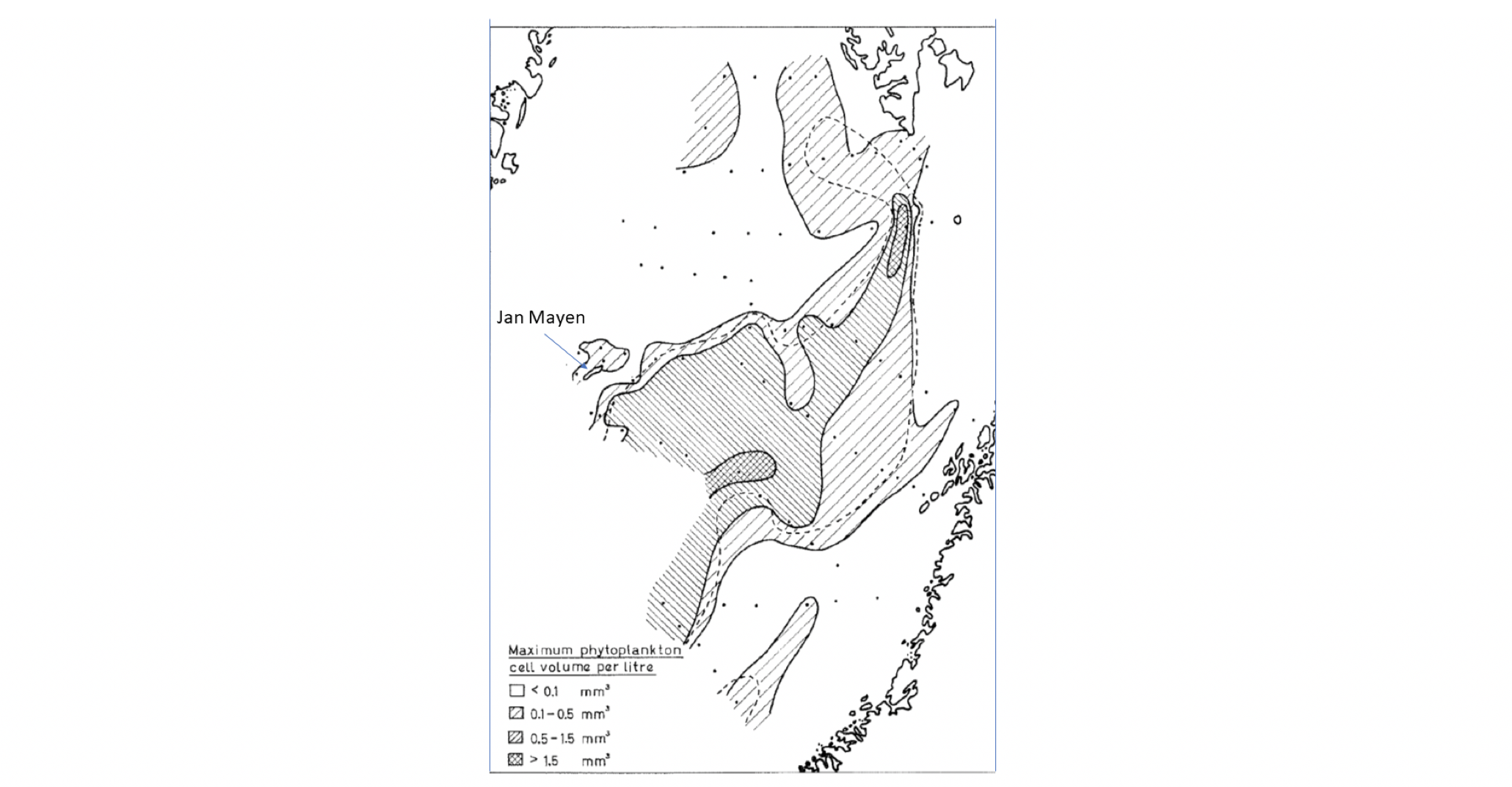
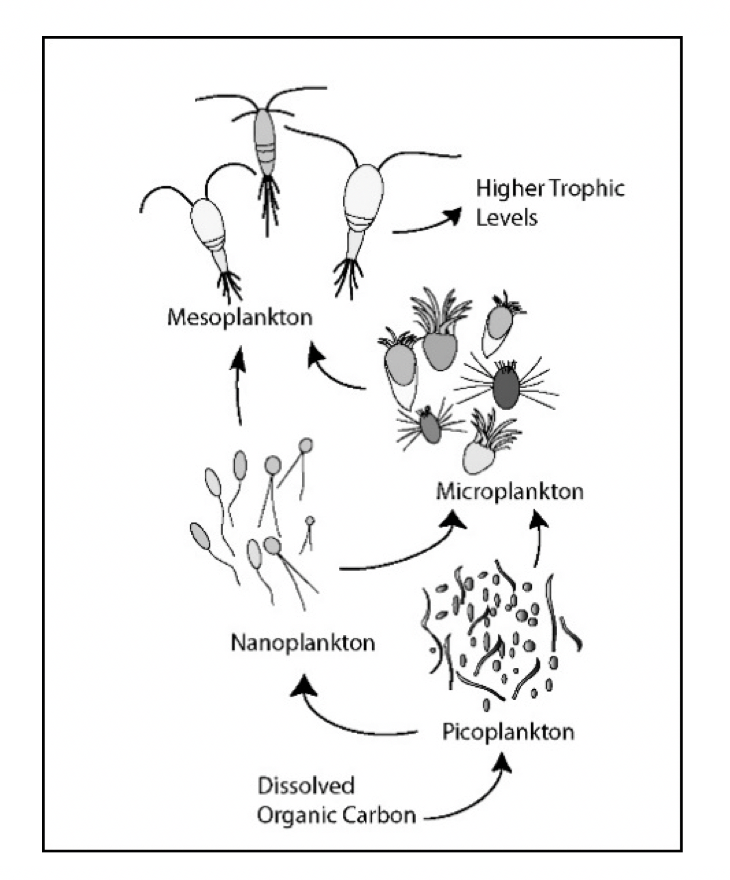
Livsformer i havet som driver med strømmene kalles plankton (Hensen 1887). Mange arter starter sitt liv som planktoniske egg og larver, men blir i stand til å forsere havstrømmene når de vokser opp (meroplankton). Andre forblir prisgitt strømmenes bevegelse gjennom hele sin livssyklus (holoplankton), enten på grunn av størrelsen, eller fordi de har liten evne til egenbevegelse, som for eksempel manetene. Planktonet omfatter både planter og dyr, encellede så vel som flercellede, og gruppen dekker et bredt størrelsesspekter. Plankton kan typisk påvirke sin vertikale posisjon i vannsøylen ved å svømme eller regulere oppdriften, mens horisontal forflytning først og fremst er knyttet til vannmassenes bevegelser. Romlige fordelinger av planktonpopulasjoner er dermed sterkt påvirket av havstrømmer og sirkulasjonsmønstre (Melle mfl. 2004). Plankton i en eller annen form finnes nær sagt overalt i verdenshavene, og forekommer fra bunn til overflate gjennom hele utredningsområdet. Mengden og sammensetningen av ulike arter vil variere geografisk og med miljøforholdene, og dermed også mellom habitater karakterisert ved ulike typer vannmasser, dyp, fysiske faktorer inkludert lysforhold og temperatur, predatorregimer, etc. Mengden av en planktonart i et bestemt område og dyp vil typisk endre seg gjennom sesongsyklusen, bl.a. grunnet synkronisering med tilgang til næring, fysiske forhold, reproduksjonssyklus, og for visse arter også overvintring i dypt, kaldt vann versus opphold i overflatelagene om sommeren når føde er tilgengelig og høyere temperatur gir økte vekstrater.
Bakteriene i sjøvann er de mest tallrike organismene, og deres funksjon i økosystemet er i hovedsak knyttet til nedbryting av organisk materiale og annen resirkulering. Virus er parasittiske biologiske enheter som er enda mer tallrike enn bakterier (Bergh mfl. 1989). Alle organismer blir parasittert av artsspesifikke virus, og virus fremkaller en viktig del av dynamikken i mangfoldet av organismer i alle miljø. En art som vokser seg spesielt tallrik og dominerende vil ha stor sannsynlighet for å bli desimert av virusepidemier, og høy mortalitet i en art åpner for andre arter (Suttle 2005).
Planteplankton omdanner uorganisk karbon i havet til biologisk bundet organisk karbon ved den biokjemiske prosessen fotosyntese, hvor lys benyttes som energikilde (Williams mfl. 2002). Dyreplankton spiller rollen som bindeledd mellom planteplankton og organismer høyere i næringsnettet. Dette fungerer ved at såkalt herbivort dyreplankton som beiter på planteplankton overfører energi fra primærprodusentene til sin egen biomasse, og selv blir spist av annet dyreplankton, fisk, sjøfugl og hval.
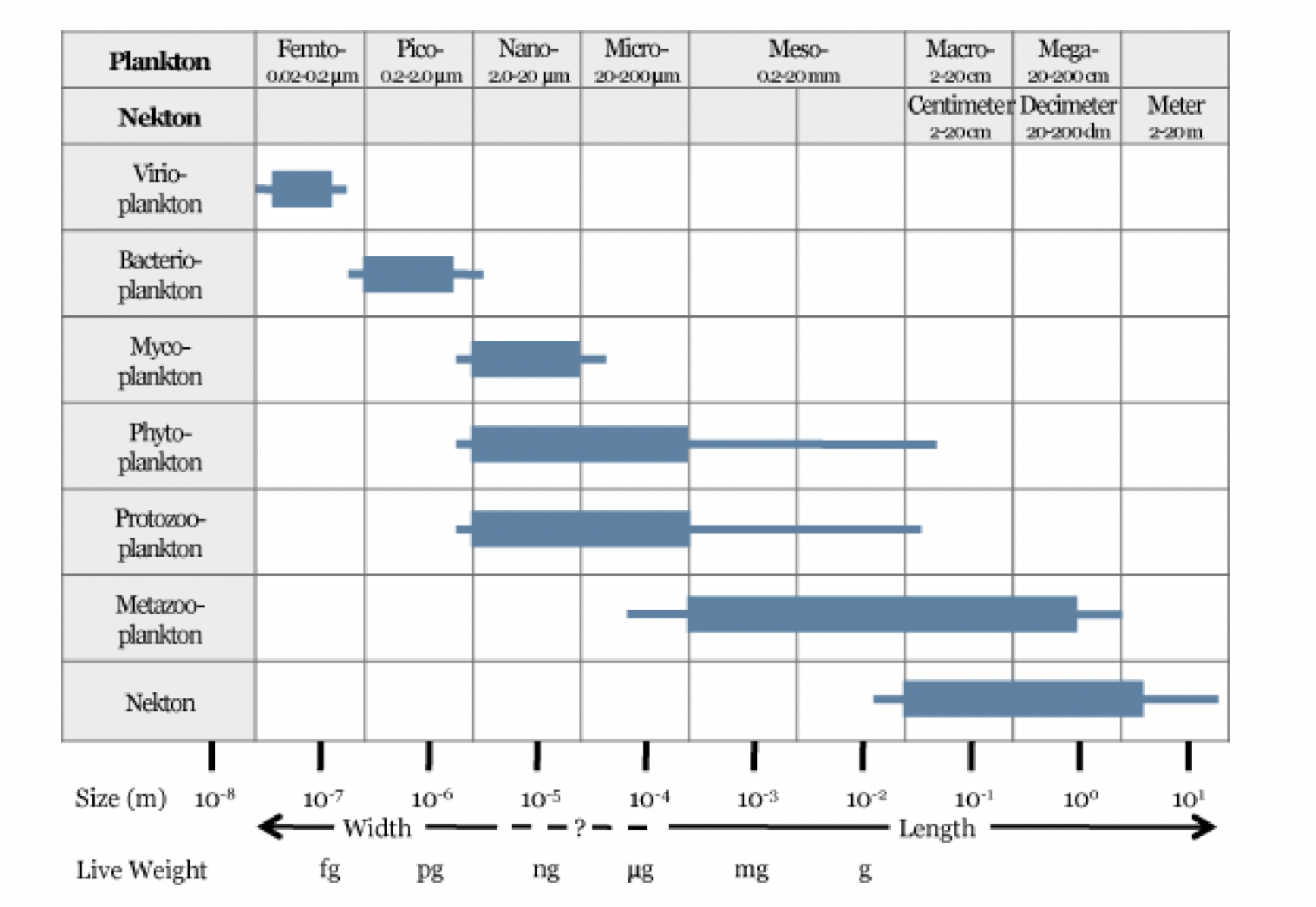
Noen typer dyreplankton har mange generasjoner per år, mens andre trenger flere år før de er klare til å reprodusere. Viktige dyreplanktongrupper i Norskehavet er mikrodyreplankton, kopepoder (også kalt hoppekreps), ostrakoder (muslingkreps), pilormer, krill, amfipoder, pelagiske reker, geléplankton (ulike typer maneter), pelagiske gastropoder, fiskeegg og larver (Melle mfl. 2004). Kopepodene er små krepsdyr, typisk 1–10 mm lange, og representerer den antallsmessig dominerende gruppen av flercellede dyr i havet (Melle mfl. 2004, Kiørboe 2011 med referanser). Det mellomstore dyreplanktonet kalles også for mesodyreplankton, og er representert av organismer med kroppsstørrelse mellom ~ 0.2-20 mm (Figur 4.1). Mens dyreplankton er definert ved å ha sin utbredelse styrt av horisontale strømmer, er nekton dyr med nok egenbevegelse til å selv styre sin utbredelse. Makroplankton og mikronekton er ofte brukte som samlebetegnelser på grupper av dyr med størrelser fra ~ 2 til ~ 20 cm. I praksis er det lite forskjell mellom disse to gruppene, så i denne rapporten slår vi dem sammen, og definerer makroplankton som alle organismer større enn ~ 2 cm som ikke foretar store horisontale vandringer. Denne gruppen vil inneholde både fisk og evertebrater; viktige grupper av makroplankton i utredningsområdet er amfipoder, krill, pelagiske reker, maneter (inkludert ribbemaneter), pilormer og mesopelagisk fisk.
Figur 4.1 Størrelsen av pelagiske organismer relatert til mye brukte begreper for klassifisering av bl.a. virus, bakterier, planteplankton, dyreplankton og nekton. Merk at de ulike organismegruppene er allokert til sine mest korrekte størrelsesspekter. Tilpasset fra Sieburth mfl. (1978).
Generelt sett har makroplankton flerårige livssykluser, og for mange av artene er vertikalfordeling ontogenetisk styrt, dvs. at forskjellige alders- eller størrelsesgrupper typisk finnes på ulike dyp. Mens enkelte arter av mesodyreplankton i området demonstrerer svært klare ontogenetiske vertikale sesongmigrasjoner (for eksempel raudåte, Calanus finmarchicus), er ikke tilsvarende mønstre for makroplankton allment akseptert. Imidlertid er det godt kjent at vertikal fordeling av makroplankton varierer med miljøforhold og vannmasser (f.eks. Dale mfl. 1999a, Norheim mfl. 2016), og vertikal fordeling kan forventes å variere med sesong. I tillegg til makroplankton er pelagiske blekksprut antatt å være en gruppe av betydning i området (Olsen mfl. 2006), men her mangler i stor grad datagrunnlag for å kunne vurdere hvor viktig denne gruppen er.
Økologiske studier av de frie vannmassene (altså de deler av vannsøylen som ikke er i direkte kontakt med havbunnen), oftest kalt pelagialen, deler gjerne vannsøylen inn i vertikale soner. Den øverste kalles epipelagisk sone og er vanligvis definert som den delen av vannsøylen der det er nok lys på dagtid til fotosyntetisk aktivitet. Sonen kalles derfor også fotisk eller eufotisk sone. I praksis foregår så og si all primærproduksjon i denne sonen, som også har de største tetthetene av konsumenter, for eksempel kopepoder og fisk som makrell og sild. Under den epipelagiske sonen kommer den mesopelagiske, som oftest blir definert som den delen av vannsøylen der lys fra overflaten er til stede i tilstrekkelige grad til at organismer kan detektere det, men i for liten mengde til å tillate fotosyntese. Som tommelfingerregel kaller vi dyp fra ~200 til 1000 m for mesopelagiske. Fysiske og biologiske prosesser i øvre vannlag (e.g. epipelagisk og mesopelagisk sone) styrer energiflyten mellom primærproduksjonen i epipelagisk sone og dypereliggende økosystemer (Turner 2015), både bathypelagisk sone (frie vannmasser der overflatelys ikke lengre er detekterbart for organismer) og dyptliggende bentiske systemer.
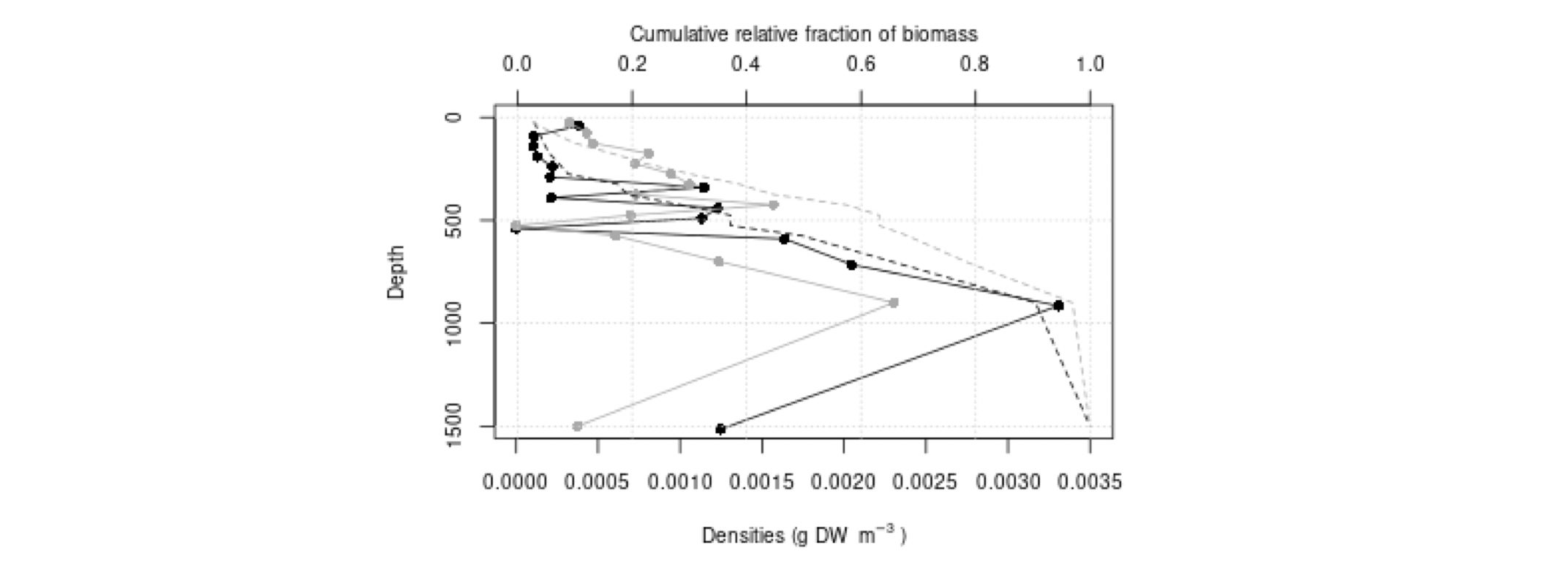
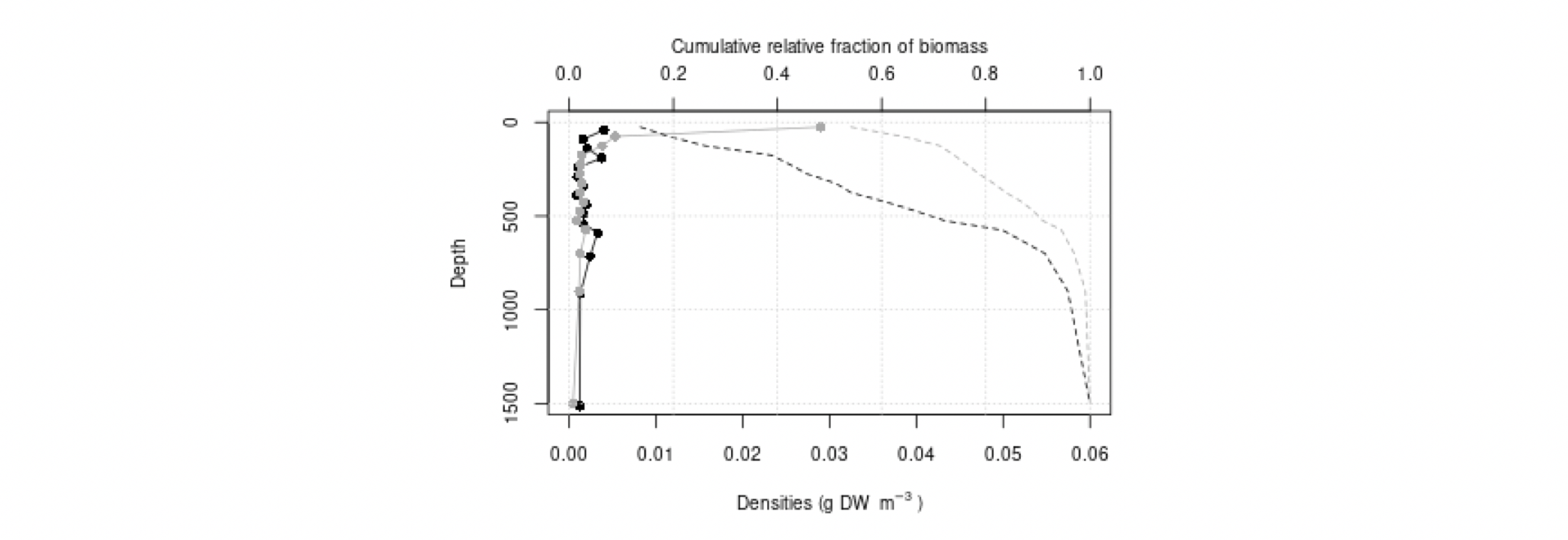
I tillegg til at meso- og makroplankton er essensielle komponenter i de pelagiske næringskjedene, er de sannsynligvis viktige for transport av næring til økosystemene i dyphavet. Svært mange av artene i Norskehavet vandrer daglig fra mesopelagiske dyp til vannmasser nær overflaten: her beiter de på mindre planktonarter, før de vandrer ned i dypet igjen, og på denne måten transporter de effektivt karbon til dypet. Dette kalles aktiv karbontransport, i motsetning til passiv karbontransport, som er betegnelsen på partikler som synker ut fra øvre vannlag. Dyreplankton er også viktige for den passive transporten, det antas at en stor andel av det utsynkende, partikulære karbonet finnes i avføring fra dyreplankton (Turner 2015). Studier fra andre havområder har antydet at mer enn halvparten av karbonet eksportert dypere enn 1000 m dyp kan gå via makroplankton (Davison mfl. 2013), men for norske farvann er det store hull i kunnskapen om hvor mye av den totale karboneksporten som går via plankton på veien ned gjennom vannsøylen.
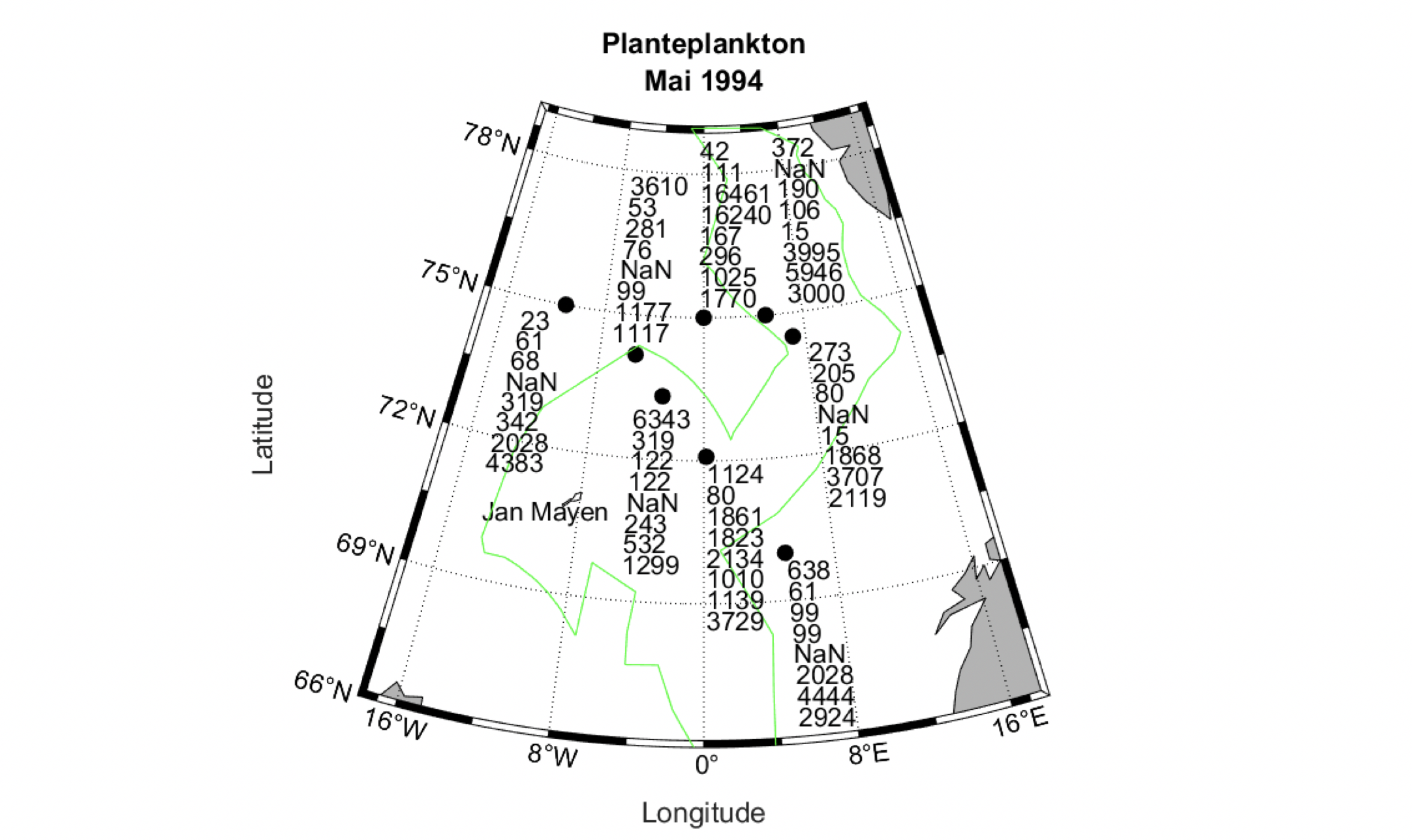
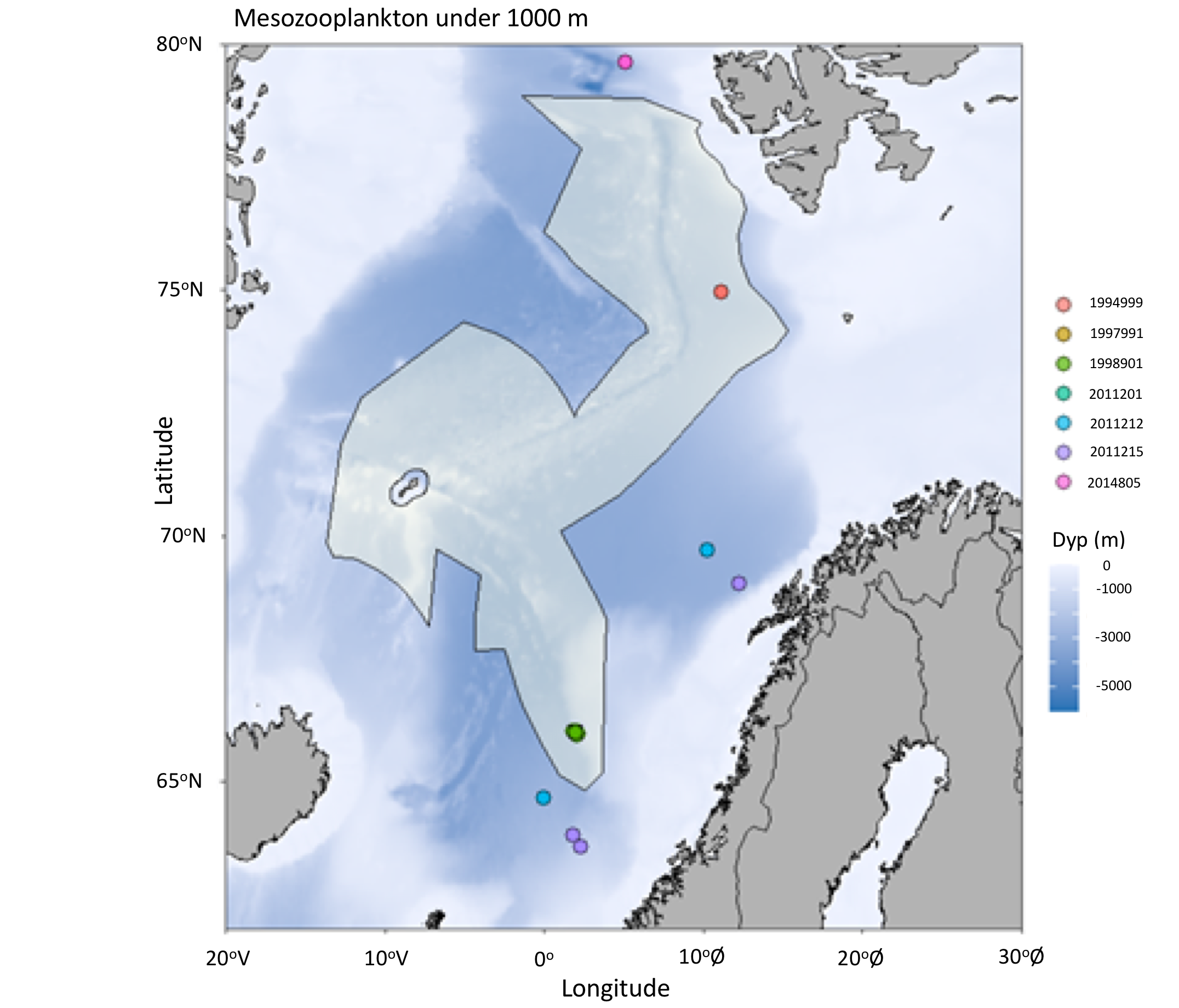
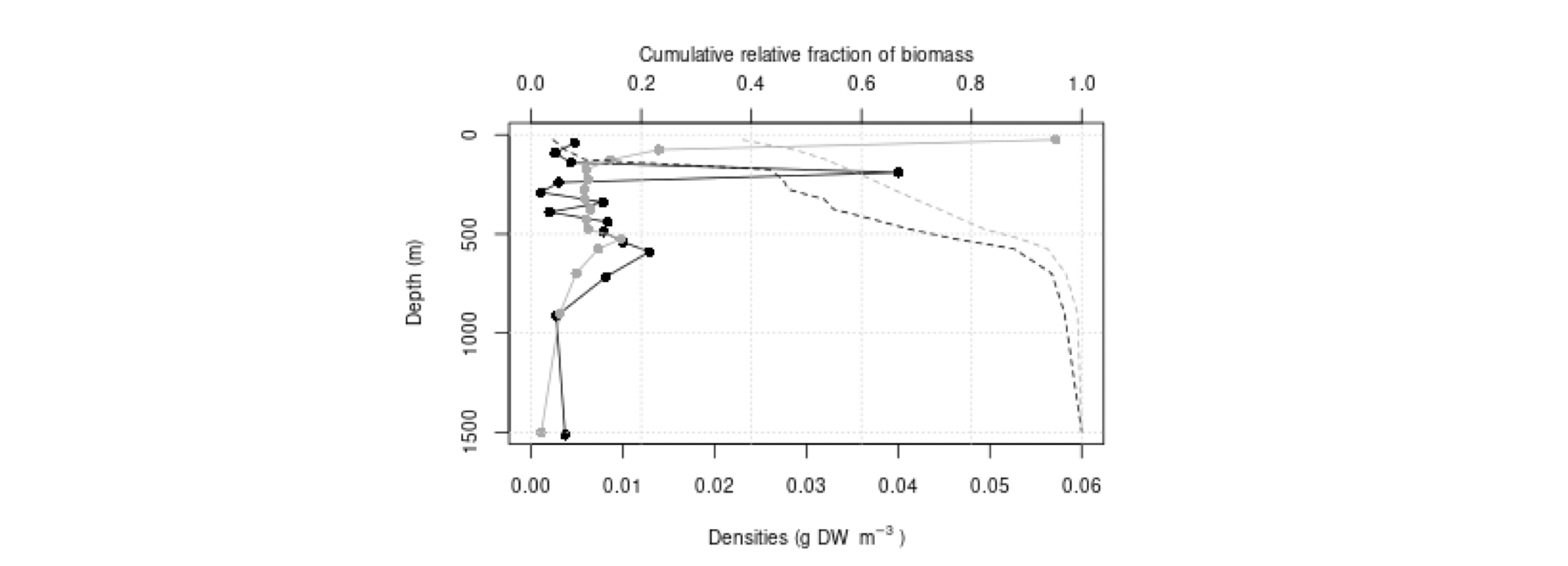
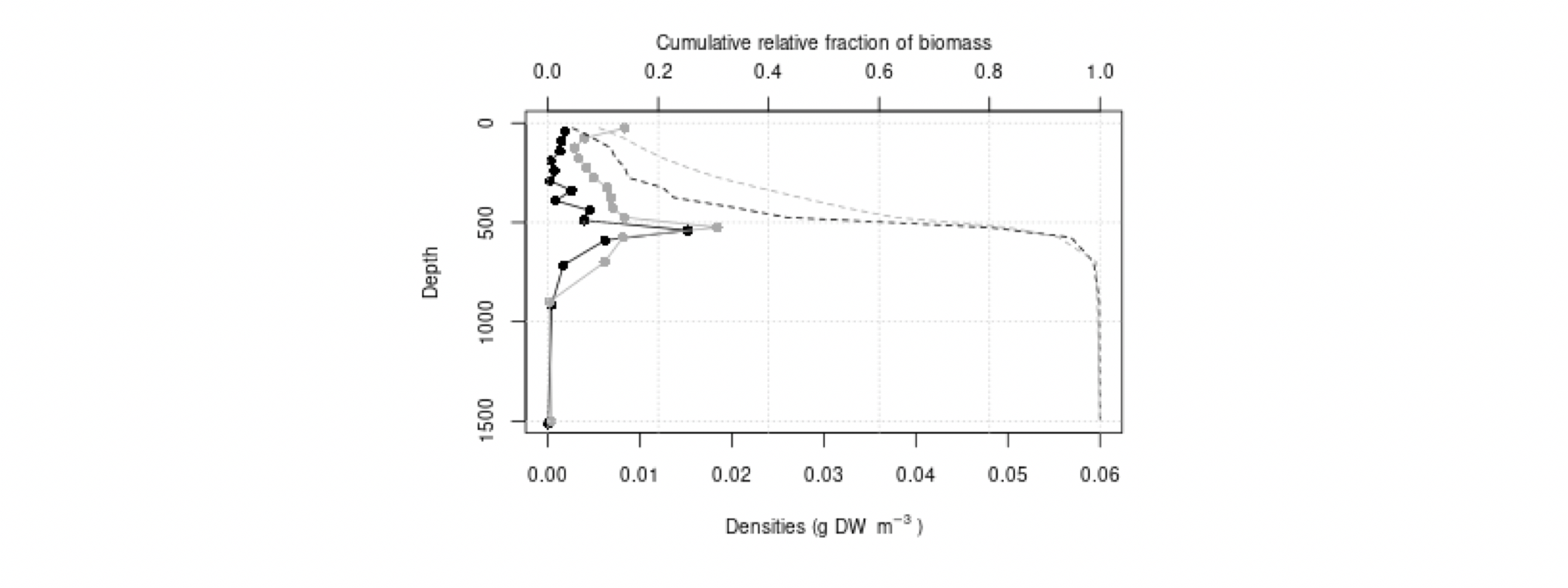
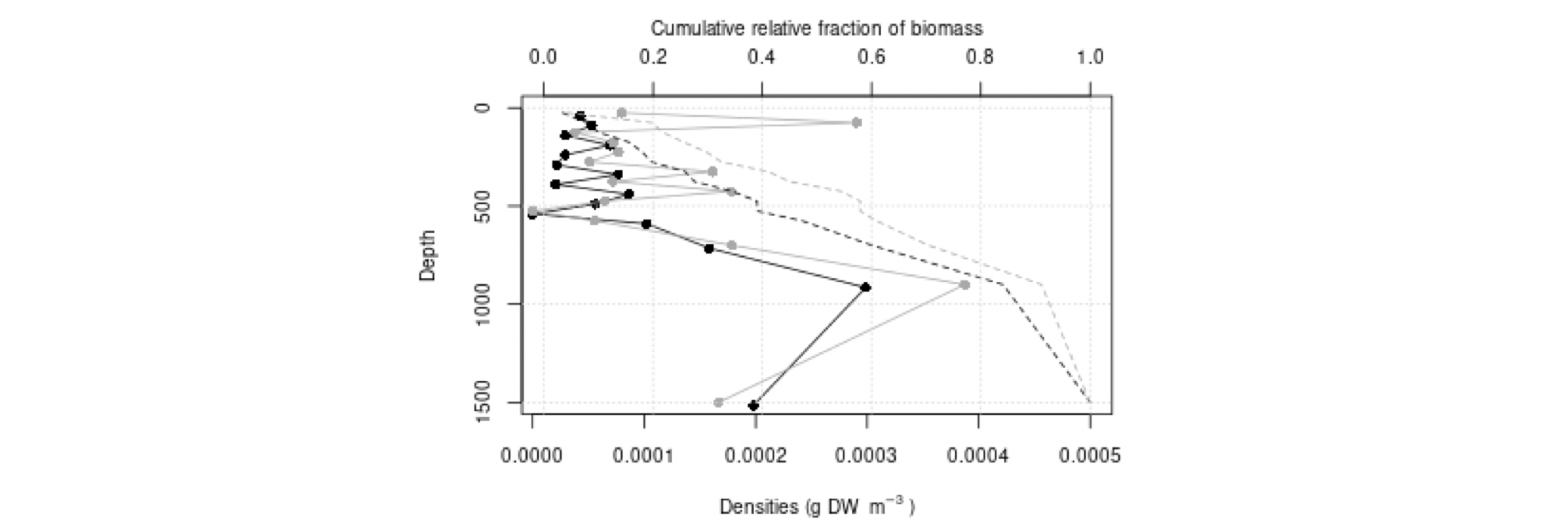
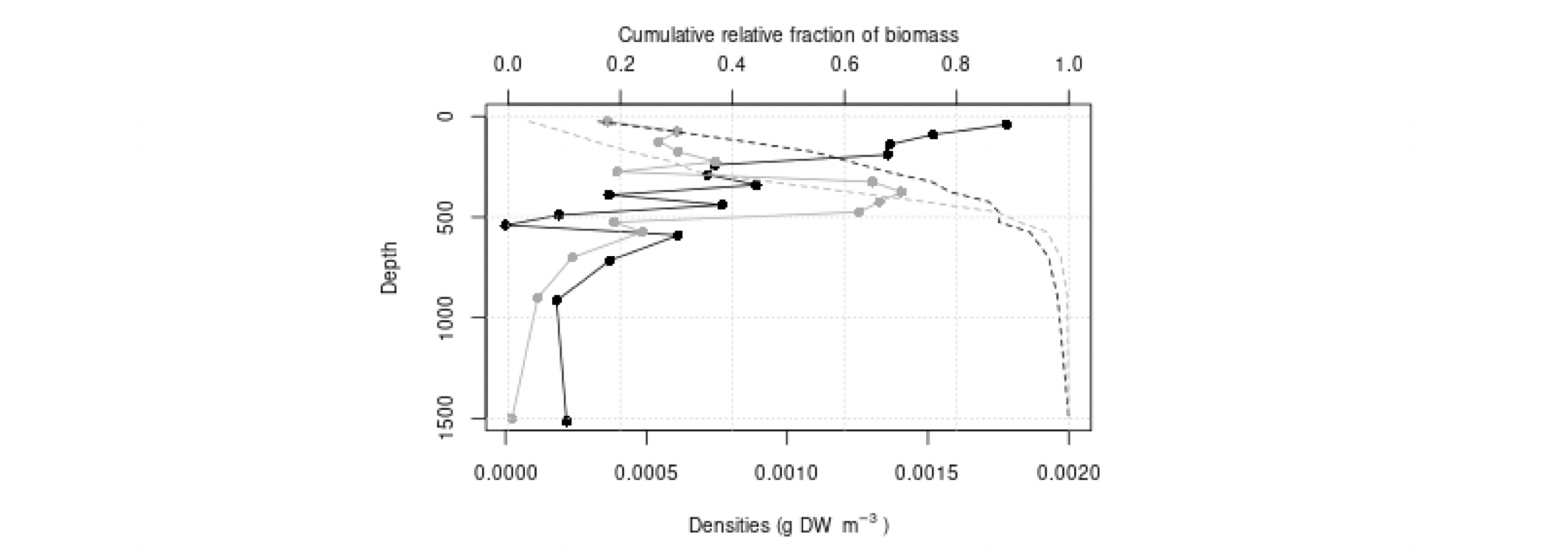
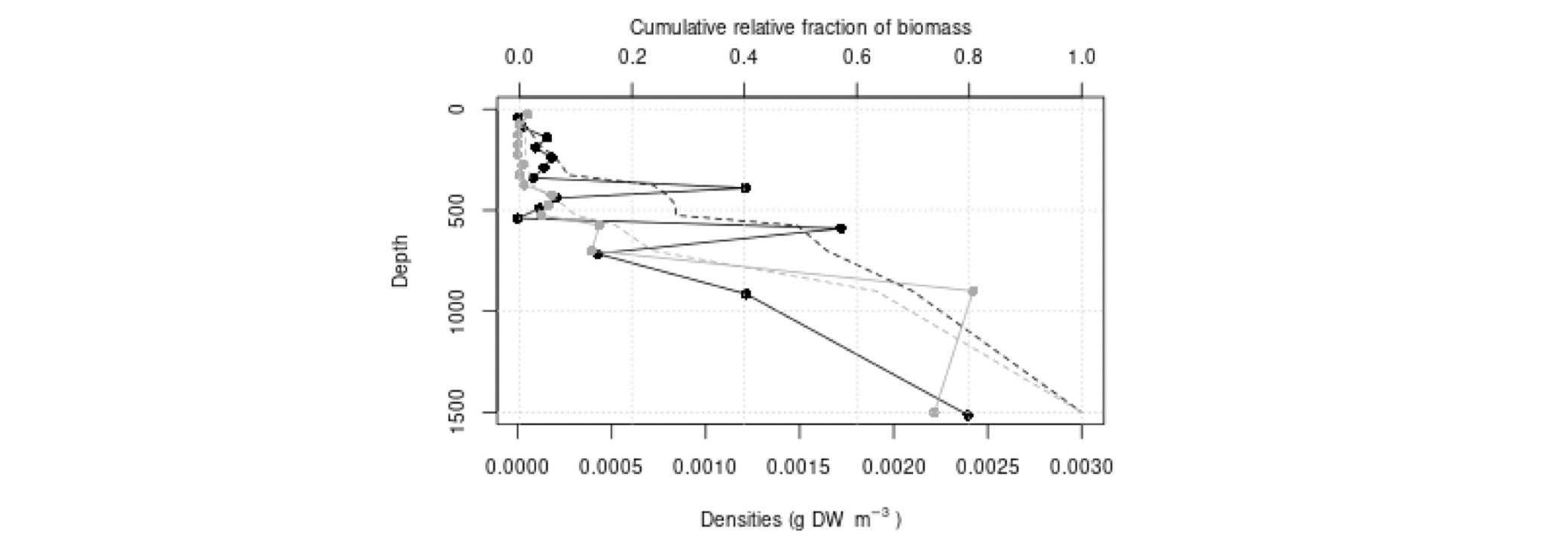
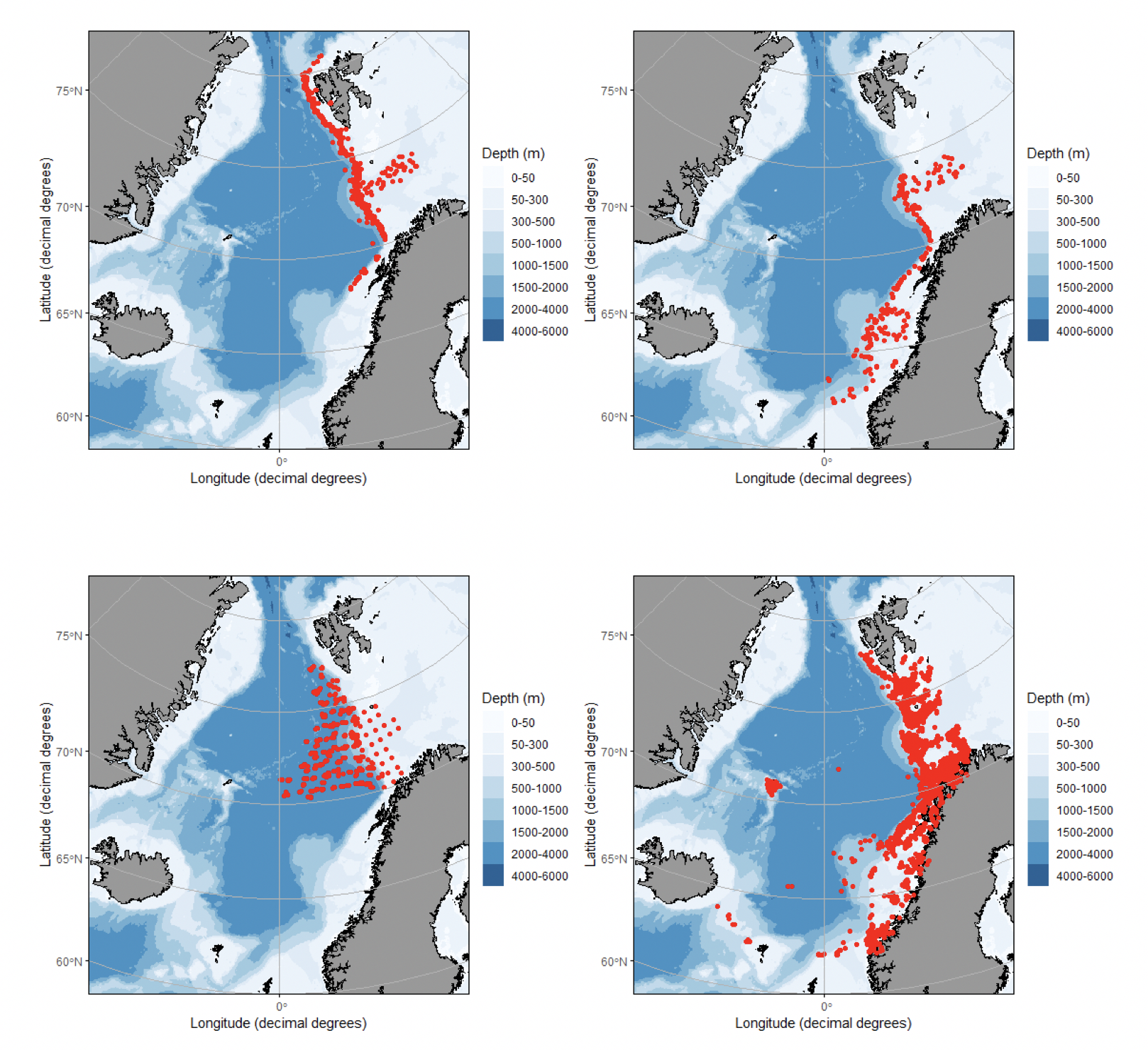
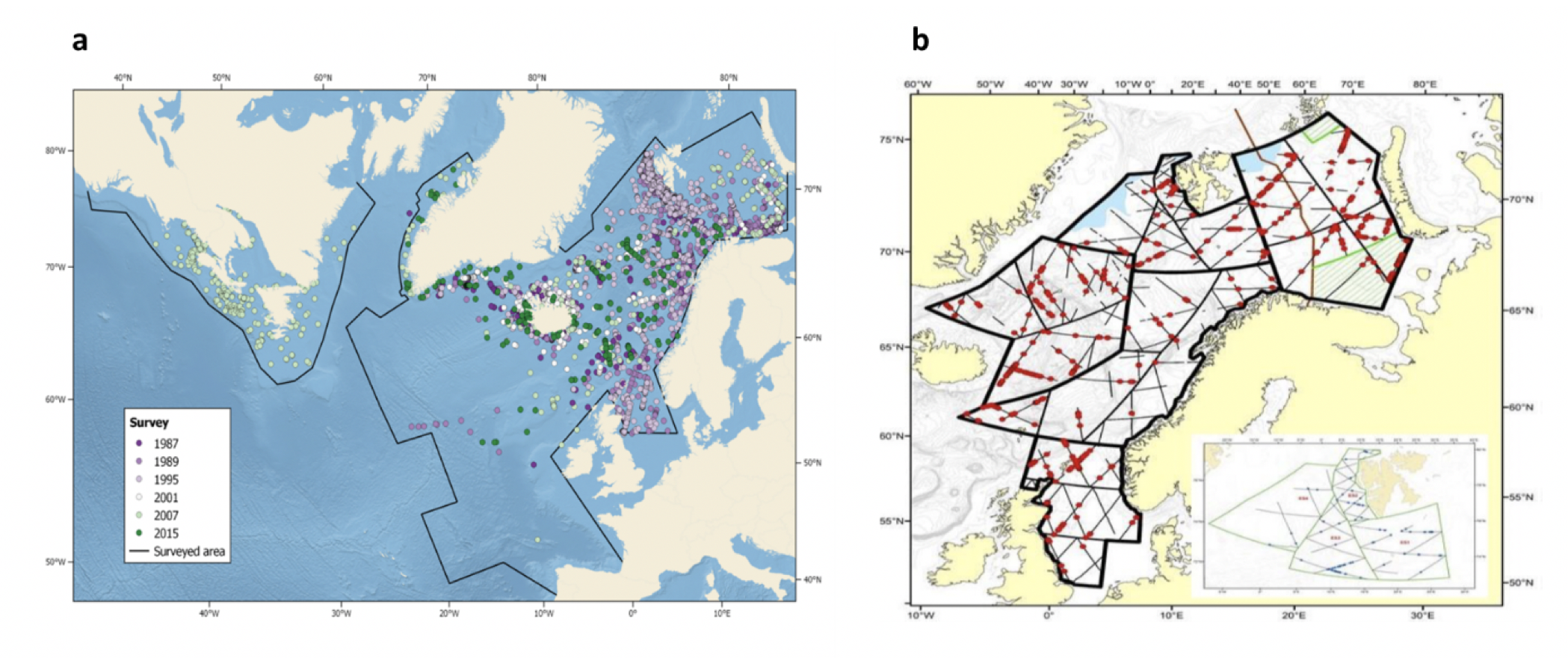
Det definerte utredningsområdet (Figur 1.1) representerer ingen naturlig biologisk inndeling (i det minste for pelagisk sone), fordelinger av dyr og biologiske prosesser vil derfor spenne ut over områdets grenser. De pelagiske økosystemene innenfor utredningsområdet vil heller ikke være homogene, og man kan forvente forskjeller mht. planktonets artssammensetning, biomassenivåer og “timing” av årssyklus innen området. Ettersom planktonets horisontale utbredelse i stor grad styres av havstrømmer, er kunnskap om havstrømmer og sirkulasjonsmønstre for området (se oseanografi-kapitlet i rapporten) nødvendig for å forstå områdets pelagiske økologi. For eksempel vil utredningsområdet ha en viss romlig overlapp med Norskehavs-, Lofoten- og Islands-bassengene, og dermed også viktige overvintringsområder for hoppekrepsen Calanus finmarchicus (raudåte) (Melle mfl. 2014). Utredningsområdet overlapper også noe med bassenget i Grønlandshavet, som er kjerneområde for raudåtens større, arktiske slektning Calanus hyperboreus (Hirche 1997). Siden horisontale strømmer kan føre plankton inn og ut av utredningsområdet, samt fordi det finnes få planktonprøver fra dyp større enn ca. 700 m innenfor utredningsområdet, suppleres det i denne rapporten med informasjon basert på prøver hentet i utredningsområdets nærhet (Figurer 4.2 og 4.3).
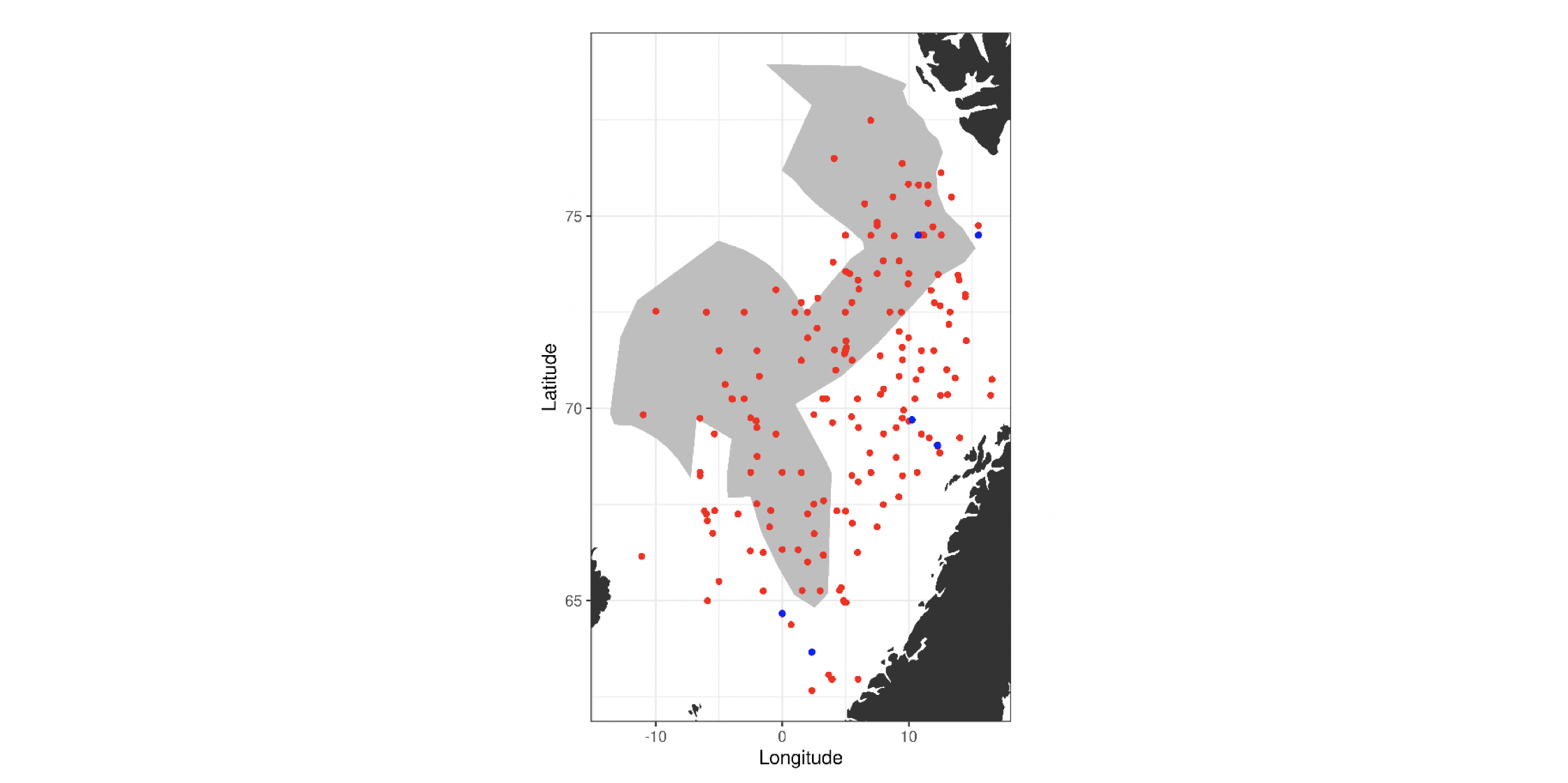
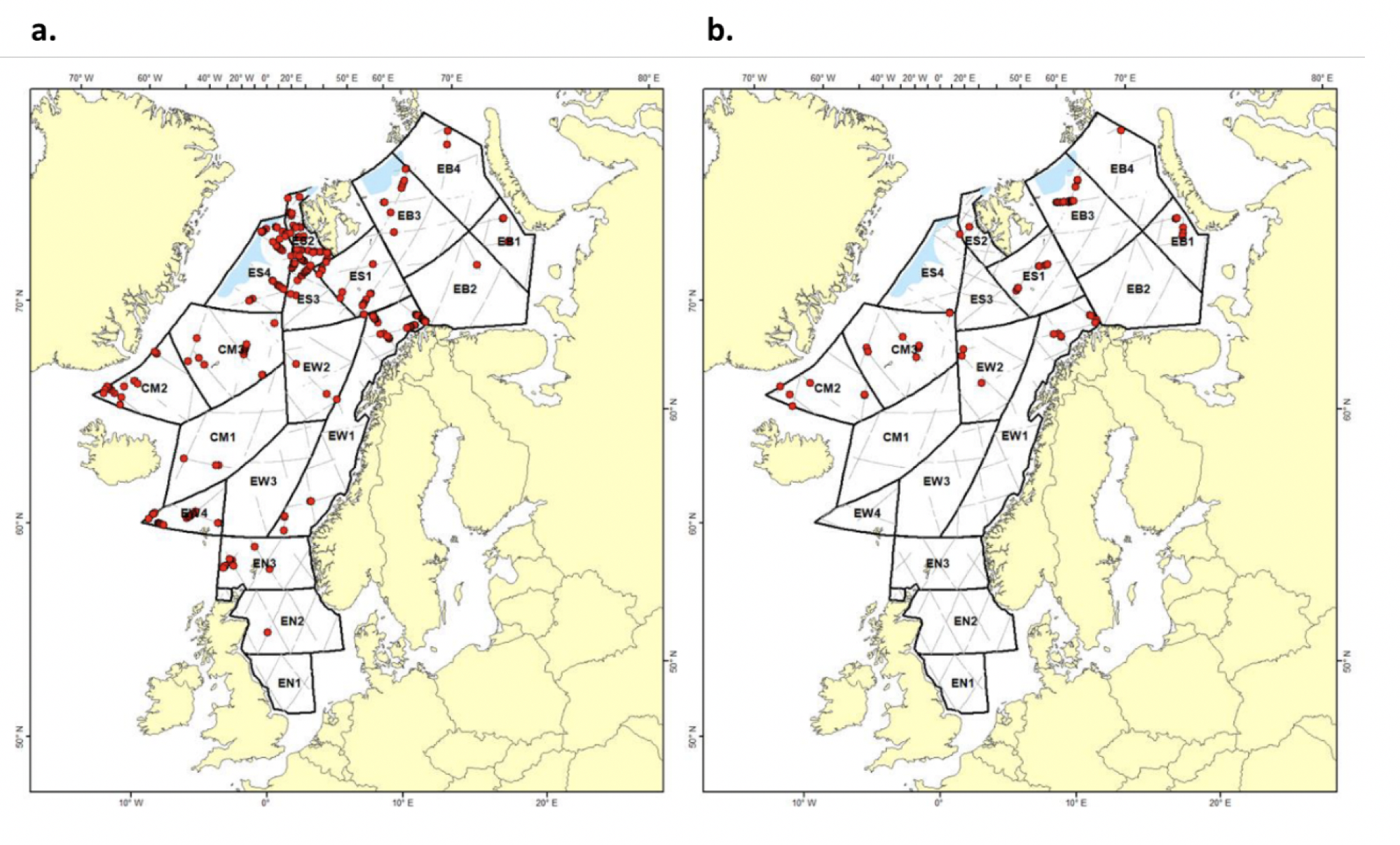
4.2 Prøvetagningsposisjoner for dybdestratifiserte dyreplankton prøver samlet inn med redskapet MOCNESS t.o.m. 2016. Røde punkter viser stasjoner med dybdestratifiserte prøver ned til minst 500 m, mens blå punkter viser stasjoner med prøvetagning dypere enn 1000 m. Grått felt angir utredningsområdet. Som vist ved de blå punktene har Havforskningsinstituttet svært få virkelig dype (> 1000 m), dybdestratifiserte prøver. I tillegg har HI tilgang til en del ikke-dybdeoppløste prøver, for eksempel dyreplankton fra bunn eller 2000m til overflaten, tatt med WP2-håv (disse stasjonene er ikke vist i figuren).
4.2 - Planktonovervåkning og tidsserier
Havforskningsinstituttets regulære overvåking i Nord-Atlanteren består i hovedsak av årlige storskaladekninger av følgende havområder; Nordsjøen, Norskehavet og Barentshavet, kombinert med et sett av faste snitt som dekkes 4-6 ganger per år (noen snitt er geografisk forlenget én gang per år). Formålet med snitt som dekkes gjentatte ganger årlig er å fange opp sesongutvikling hos planteplanktonet og sekundærprodusentene, biodiversitet og klimarelaterte effekter på økosystemene – for eksempel ved forflytning/transport av sørlige arter inn i våre nordlige produksjonsområder, samt å kunne estimere mellomårstrender i sekundærprodusentenes antall og biomasse.
Når det gjelder den dypere delen av vannsøylen, er Havforskningsinstituttets og andre institusjoner/nasjoner sin overvåking mangelfull. I Norskehavet og dets randsone foregår den storskala områdedekningen to ganger i året, der første dekning er i mai og andre dekning er i juli og august. Prøvetakingen av dyreplankton utføres her med WP2 håv (Unesco 1968) med maskevidde 180 µm fra 200 m dyp til overflaten, samt med jevne mellomrom også dybdestratifiserte MOCNESS / Multinet Mammoth (Wiebe m.fl. 1976, 1985, Hydrobios) fra 700 m til overflaten. I tillegg tas WP2 prøver fra snittene Svinøy-NordVest (5x/år) og Gimsøy-NordVest (4-5x/år) og Bjørnøya-Vest (4-5x/år) (se Hassel mfl. 2020) (plassering av snittene er vist i Figur 4.3). Her tas prøver fra 200 m til overflaten (evt. bunn til overflaten i grunnere områder), samt noen dypere prøver og enkelte dybdestratifiserte MOCNESS/Multinet Mammoth. I tillegg dekkes snittet Fugløya-Bjørnøya, som krysser den sørvestlige inngangen til Barentshavet, og hvor det tas WP2-prøver fra bunn til overflate (Hassel mfl. 2020) (4-6 x/år).
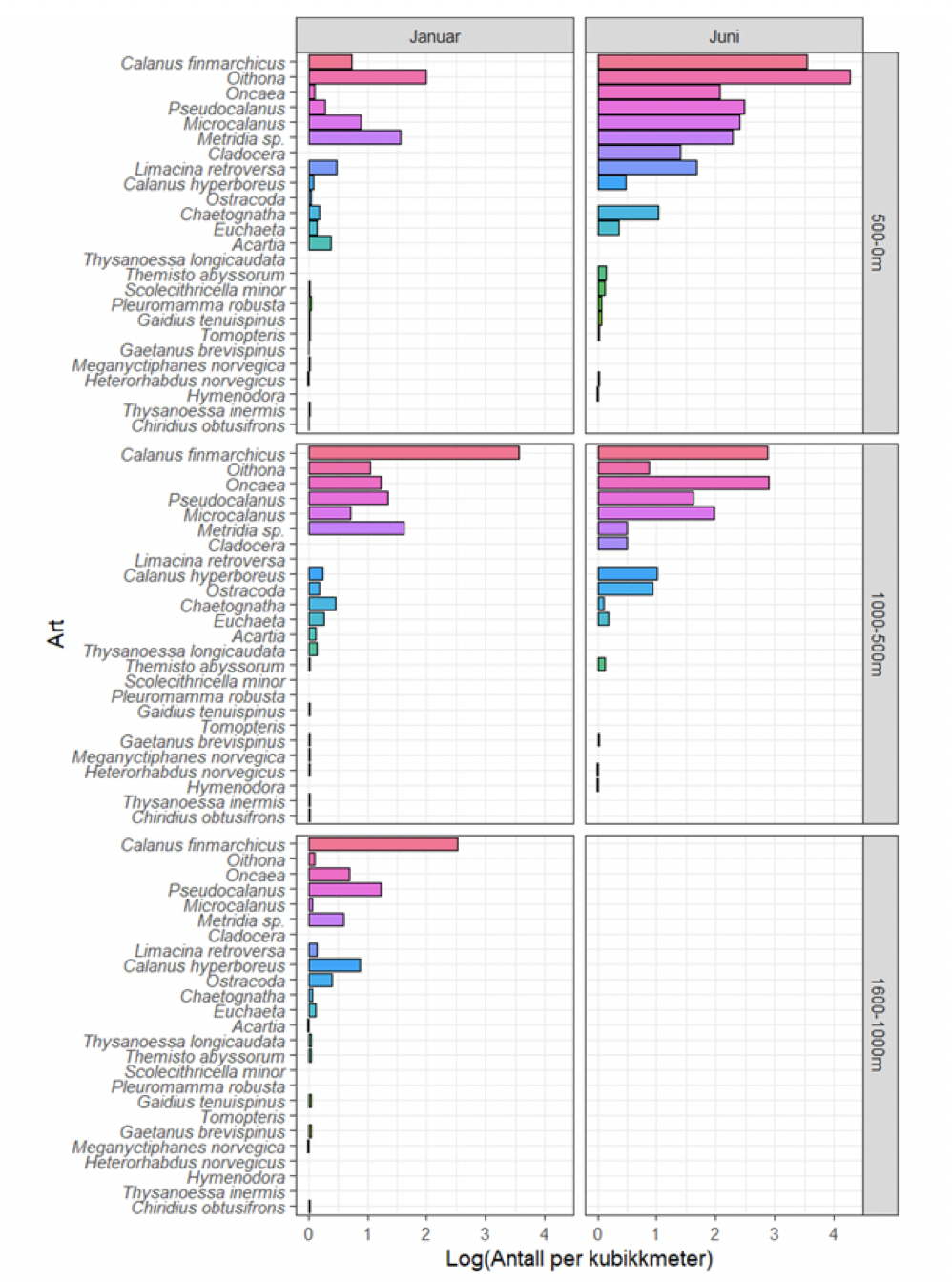
Mens de storskala områdedekningene i Norskehavet overlapper med utredningsområdet, går kun snittet Bjørnøya-vest inn i utredningsområdet (samt det forlengede Gimsøy-snittet en gang per år – se Figur 4.3). De andre snittene er likevel relevante siden plankton kan drive inn og ut av utredningsområdet med havstrømmene, og snittene dermed kan representere planktonet oppstrøms og nedstrøms og dermed ha overføringsverdi til utredningsområdet. Vertikalstratifisert prøvetaking av mellomstort dyreplankton går noen ganger til ca. 2000 m dyp. I Norskehavet gjelder dette svært få prøver (se Figurer 4.2 og 4.3). Videre foretas artsopparbeiding kun av et fåtall utvalgte prøver da det er svært arbeidsintensivt og kostbart å utføre taksonomiske analyser. Dette betyr igjen at av de i utgangspunktet få eksisterende vertikaloppløste planktonprøvene fra Norskehavet med øvre lukkedyp dypere enn 1000 m, er kun et lite antall analysert mht. artssammensetning. Imidlertid er rutinen at for alle dyreplanktonprøver veies størrelsesfraksjonert biomasse, som dermed gir data på dybdestratifisert biomasse for ulike størrelsesgrupper. Dette er nyttig for å forstå hvor store mengder dyreplankton som befinner seg på forskjellig dyp i vannsøylen i forskjellige områder og til ulike tider på året, gitt begrensingene i prøvetakingen nevnt over. Per i dag er det klare begrensninger mht. hva man kan sammenfatte om ulike typer planktonbestander dypere enn ca. 700 m.
I tillegg til prøvetakingen nevnt ovenfor, samarbeider Havforskningsinstituttet med The Marine Biological Association (www.mba.ac.uk), som driver “the Continuous Plankton Recorder Survey” (CPR) (Batten mfl. 2003). Prinsippet for innsamling med CPR og den standardiserte taksonomiske opparbeidingen er beskrevet i Richardson mfl. (2006). Dermed finnes artsinformasjon for plankton samlet inn ved ca. 7 m dyp langs to ruter; hvor den ene gjennom årene 2008-2016 gikk mer eller mindre månedlig mellom Nord-Norge og Island, mens den andre ruten gikk omtrent månedlig mellom Nord-Norge og Longyearbyen (Spitsbergen). Sistnevnt rute har av logistiske årsaker mindre vinterdata. CPR data er basert på høyhastighetssamlere som taues etter fartøy (“ships of opportunity”). Fig. 4.3 viser hovedlinjene for de to beskrevne rutene – merk at plasseringen av disse kan variere litt fra dekning til dekning.
Prøvetakingen skissert ovenfor gjelder altså i hovedsak mellomstort dyreplankton (mesodyreplankton) og i mindre grad større makroplankton og mikronekton (se Figur 4.1). Skal man bedre få grep på fordeling, mengde og artssammensetning av makroplankton og mikronekton må man ta i bruk bedre egnede redskaper, fordi flere av disse organismene har betydelig svømmekapasitet og lettere unnviker de noe mindre og finmaskede redskapene beskrevet ovenfor som benyttes til prøvetaking av mellomstort dyreplankton. For å forbedre prøvetakingen av makroplankton og mikronekton, har man gjennom noen år benyttet en nyere type pelagisk trål som har fått navnet Makroplankton trål (36 m2 åpning, 3x3 mm maskevidde). Med dette redskapet foretar vi, bl.a. på overvåkingstoktene i Norskehavet, V-hal mellom ~1000 m og overflaten som gir integrerte prøver av det som står i vannsøylen av organismer som nevnt over. Å tråle med denne type utstyr på slike dyp er tidkrevende. I forbindelse med spesialundersøkelser kan man tråle dypere, men det igjen vil kreve særlige forberedelser og sannsynligvis spesialutrustning (jfr. MARECO – Wenneck mfl. 2008).
Kvantitative undersøkelser av dyreplankton gjennom hele vannsøylen ned mot 4000 m i Norskehavet er ikke realistisk i konvensjonelle overvåkingsprogrammer. Konsekvensen er at de dypeste områdene er ekstremt dårlig kartlagt mht. planktonforekomster. Man har etter hvert akkumulert en grov forståelse og kjennskap til makroplankton organismene i de øvre 1000 m, men kunnskapen om de dypere lagene er svært mangelfull. Når det gjelder mellomstort dyreplankton som samles inn med andre typer redskaper, og som i tillegg kan ta dybdestratifiserte prøver, finnes i alle fall noe kunnskap om planktonorganismene som står ned til ca. 2000 meters dyp. Denne informasjonen er dog meget spredt geografisk og mht. sesongvariasjon, og er dessuten mer mangelfull i Grønlandshavet og i nærheten av den midtatlantiske ryggen enn sentralt og øst i Norskehavet.
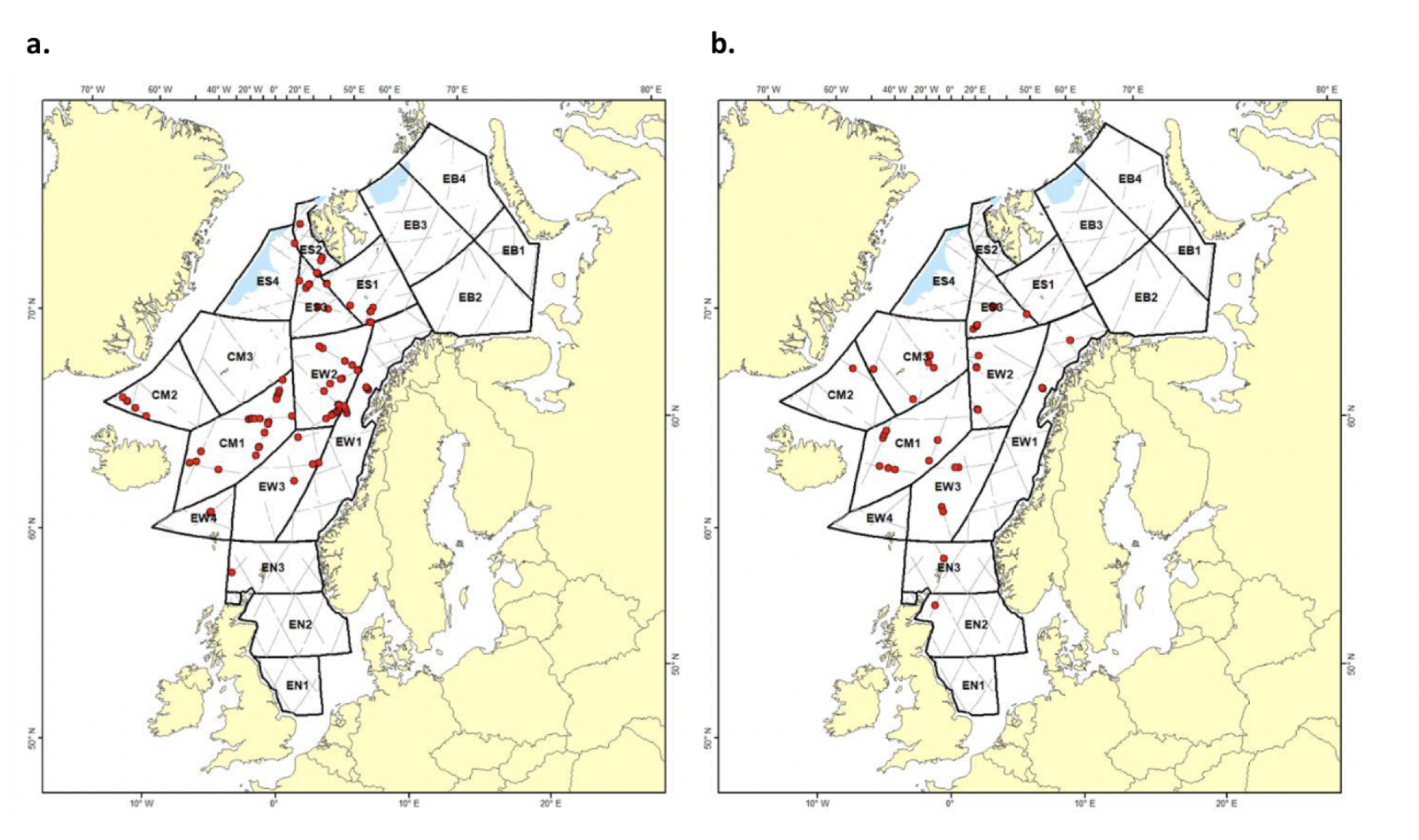
Figur 4.3 Utvalgte planktonovervåkningssnitt og stasjoner. Grått felt angir utredningsområdet. Se teksten for beskrivelse av ulike prøvetyper, innsamlingsredskap og dyp, samt hvorvidt det eksisterer taksonomisk informasjon og/eller biomassedata.
4.3 - Mikrobiologi
Biogeokjemi omhandler biologiske prosesser som omformer biokjemiske eller mineralske forbindelser. Økologisk betydning av slike prosesser kan illustreres ved å følge transformasjonene i sykliske diagrammer hvor karbonsyklus er et eksempel (Figur 4.4). I alle biogeokjemiske sykluser er mikroorganismer essensielle aktører, men i karbonsyklus er også fisk, fugler og større predatorer spektakulære og interessante aktører. Mikroskopiske encellede organismer står for primærproduksjonen, og andre mikrober står for mineralisering tilbake til karbondioksid. Flere viktige grupper av beitere er også encellede mikroskopiske organismer, særlig ciliater og flagellater, og disse er samlet under fellesbetegnelsen mikrodyreplankton (Figur 4.4). Parasittisme er en faktor for mortalitet hos alle livsformer, og i sjøvann er virus, men også spesielle bakterier, sterkt til stede. Lengre oppe i næringskjeden finnes også en rikelig fauna av flercellede parasitter.
Figur 4.4 Karbonsyklus i øvre vannmasser i havet. Blå bokser inneholder oppløste forbindelser. Prosesser er markert med tall. 1: Primærproduksjon. 2: Beiting eller predasjon. 3: Parasittisme. 4: Lysis. 5: Ekskresjon eller defekasjon. 6: Respirasjon. 7: Bakterieproduksjon. Karbondioksid er plassert to steder å forenkle grafikken.
Lignende diagrammer kan brukes for å illustrere omsetning av næringssalter som inneholder elementer som nitrogen, fosfor og svovel, og noen av disse kretsløpene involverer svært spesialiserte grupper av bakterier. Mikronæringstoffer er grupper av elementer som cellene trenger i svært små mengder, for eksempel kobber og litium. Disse vil bli omsatt av spesialiserte eller mindre spesialiserte mikroorganismer. Videre er det mange elementer som ikke inngår i cellenes byggestoffer, for eksempel kvikksølv og andre tungmetaller, som også omsettes av mikroorganismer både aktivt ved enzymatisk opptak, og passivt ved at de adsorberes til overflater (Gadd 2010). Omsetning av metaller er viktige prosesser fordi de i visse former og konsentrasjoner er særdeles giftige. I utredningsområdet finnes rimelig gode observasjoner av karbonsyklus og omsetning av næringssalter. De mer eksotiske biogeokjemiske syklusene som omfatter metaller har vært forsket på i mindre grad, og hovedsakelig i forbindelse med undersøkelse rundt geokjemiske lekkasjer fra jordens indre langs det midtatlantiske sprekkområdet.
4.3.1 - Planteplankton
I havet er primærprodusentene hovedsakelig mikroskopiske encellede planktoniske organismer, med samlebetegnelse planteplankton eller fytoplankton. Felles for disse er at de inneholder klorofyll som gjør dem i stand til å drive fotosyntese, og disse fargestoffene gjør at planteplankton kan kvantifiseres med optisk metodikk. Fargen på havet observert fra satellitt gir informasjon med høy geografisk oppløsning om mengde planteplankton i sjøen.
Figur 4.5 Konsentrasjon av klorofyll på tidspunktet våroppblomstringen kollapset. Undersøkelsesområdet er vist i sort, den arktiske fronten er vist i magenta. Skala for klorofyll er i briggske logaritmer: log10 (mg Chla m-3).
Figur 4.5 viser konsentrasjonen av klorofyll på det tidspunkt konsentrasjonen av planteplankton når sitt maksimum. Dette tidspunktet markerer slutten på våroppblomstringen, og etterfølges av en sommerperiode med lavere algebiomasse. Oversikten fra satellitt viser at utredningsområdet er mangfoldig når det gjelder den årvisse oppvekst av planteplankton. I det arktiske domenet er den maksimale konsentrasjonen høyere enn i det atlantiske domenet, men slutten av våroppblomstringen er tidligere i det atlantiske domenet (Børsheim mfl. 2014). Den årvisse våroppblomstringen sees i området rundt Jan Mayen fra tidlig mars til midt i juni (Figur 4.5), og den mellomårlige variasjonen er høy i området. Dette går fram av standardavviket for gjennomsnittet (Figur 4.6). Observasjonene i figuren er fra øst for den arktiske fronten, det vil si atlantiske domene. I det arktiske domene er den mellomårlige variasjonen, målt som standardavviket på årsvariasjonen, dobbelt så høy som på østsiden av den arktiske fronten.
Figur 4.6 Årsvariasjon av klorofyll øst for Jan Mayen, som ukentlige gjennomsnitt for tidsperioden 2003 til 2019. Vertikale markeringer viser standardavvik. Data fra NASA sin satellitt MODISaqua.
Utredningsområdet er ikke blant Havforskingsinstituttets mest undersøkte områder (jfr. kapittel 4.2), med interessante unntak. I mange år har det gjennomførts ekspedisjoner langs 75ºN, og disse har vanligvis også tatt prøver langs Gimsøysnittet og videre på kursen fra Lofoten retning nordvest til Grønland, slik at prøvetakingen krysset utredningsområdet to ganger. Disse ekspedisjonene er ikke hovedsakelig regelmessige overvåkningstokt, men har primært vært plattform for dedikerte forskningsprosjekter.
Figur 4.7 Planteplankton nord for Jan Mayen i mai, celler per milliliter (NaN står for ikke registrert). Grønt polygon viser utredningsområdet, og svarte sirkler viser posisjoner hvor det ble tatt prøver av planteplankton. Tallkollonnene viser telletall av hovedgrupper. Fra øverst: 1. Bacillariophyceae (Kiselalger); 2 Dinophyceae (dinoflagellater). 3 Prymniophycetes (svepeflagellater). 4. Phaeocystis pouchetii (en koloniformende svepeflagellat). 5 Monader. 6 Chryptophyceae (svelgflagellater). 7 Salpingoeca spp. (diverse heterotrofe krageflagellater). 8 Uidentifiserte flagellater.
I 1994 ble det tatt vannprøver for visuell inspeksjon og telling av planteplankton (Rey mfl. 2000). Resultatene viser at man i våroppblomstringen finner en karakteristisk fordeling av hovedgruppene av planteplankton (Figur 4.7). Klassisk metodikk for algetelling har mange begrensninger, og den kan ikke registrere det som nå kalles picoplankton. Dette er former for planteplankton som er mindre enn 2 µm i diameter, en størrelse som overlapper med heterotrofe bakterier. Det meste av picoplankton er det vi på norsk kaller blågrønnalger, og de er bakterier. I sjøvann er disse blågrønnalgene til stede i alle farvann. De er primærprodusenter, mange fikserer nitrogen, og som er viktig føde for beitere (Poulson mfl. 2017). Det er også foretatt tellinger av planteplankton fra sør i utredningsområdet (Tabell 4.1). Det generelle bildet fra artssammensetningen er ikke dramatisk forskjellig fra Grønlandshavet (kiselalger, dinoflagellater), men Naustvoll mfl. (2020) rapporterte også tellinger av ciliater, som er viktige dyreplankton og beitere. I utredningsområdet vil både biomasse og artssammensetning variere sterkt i løpet av året. Sesongvariasjon i planteplanktonet er beskrevet på værstasjon M øst i Norskehavet (Halldal 1953, Dale mfl. 1999b). Den geografiske plasseringen av stasjon M er indikert i Figur 4.3. Planteplanktonets sammensetning gjennom i 1991 er vist i Figur 4.8. Paasche (1960) viste for øvrig at våropplomstringen var dominert av små kiselalger. Fra stasjon M finnes det også en grundig studie av planteplankton fra oktober 1948 til november 1949 med ukentlig prøvetaking i hele perioden (Halldal 1953). Tabeller i denne artikkelen angir artssammensetning og telletall fra fem dyp gjennom hele året. I juni 1954 ble det samlet prøver fra De nordiske hav som ble analysert for innhold av planteplankton (Paasche 1960). Denne artikkelen gir detaljert informasjon om artssammensetningen fra tiden på sommeren da biomassen er på sitt høyeste. Artikkelen bekrefter også høye tall for heterotrofe flagellater og ciliater. Størrelsen av algene ble estimert med lysmikroskopi, dette ga grunnlag for å estimere total biomasse som volum (Figur 4.9).
Tabell 4.1. Planteplankton sør for Jan Mayen våren 2013 (Naustvoll mfl. 2020).
Celler/ml
Primo mai
Primo juni
Norskehavet
Islandshavet
Norskehavet
Islandshavet
Flagellater
1500
1500
2000
3000
Dinoflagellater
80
90
180
160
Diatomeer
300
500
400
Ciliater
0,15
0,2
0,4
Figur 4.8 Fytoplankton biomasse ved 0.5 m dyp på Stasjon M (66ºN, 2ºØ) i Norskehavet i 1991. Fra Dale mfl. 1999b.
Figur 4.9 Distribusjon av biomasse som mm3liter-1 i juni 1954. Skalaen 0,1; 0,5; 1,5 mm3 liter-1 tilsvarer 0,9; 2,6; 5,3 mg Chl m-3 (i figur 4.5 brukes logaritmer av klorofyllverdien, da er tilsvarende skala -0,05; 0,41; 0,72). Figuren er hentet fra Paasche (1960).
4.3.2 - Primærproduksjon
Observasjoner fra satellitter kombinert med empirisk basert modellverktøy har gitt estimater av primærproduksjon med samme detaljerte oppløsning som for klorofyll (Behrenfeld og Falkowski 1997). Produksjonsforholdene langs den arktiske fronten ble undersøkt på flere ekspedisjoner knyttet til polaråret 2007-2008 (Børsheim mfl. 2014).
Figur 4.10 Årlig netto primærproduksjon langs tre transekter som krysser utredningssområdet. Modifisert fra Børsheim mfl. 2014. Overflatetemperatur (Sea Surface Temperature, SST) i mai fra satellittdata som ºC.
Primærproduksjonen i det arktiske domenet var rundt 10-11 mol C m-2 år-1, men var høyere i det atlantiske domenet og har en økende gradient mot norskekysten. I det atlantiske domenet minker den årlige produksjonen nordover i området (Figur 4.10). De høyeste verdiene av klorofyll finner vi i det arktiske domenet (Figur 4.5), men de høyeste beregningene av produksjon forekommer i det atlantiske domenet. Det er fremdeles ganske åpent hvilke fysiske eller biologiske faktorer som styrer disse forholdene.
Standard metode for måling av primærproduksjon i vannprøver har benyttet radioaktivt C14-CO2 og in situ inkubasjon. En serie av slike inkuberinger ble utført i juni 1954 (Berge 1958). En sammenstilling av disse målingene ble vist i Paasche (1960) og i figur 4.11 er de sammenlignet med gjennomsnittlig primærproduksjon i juni for tidsrommet som satellitten MODIS (NASA) har levert data. Satellittdataene er justert for sammenligning med karbon-14 data på våre breddegrader (Børsheim 2017).
Figur 4.11 Primærproduksjon i Norskehavet fra juni. Til høyre karbon-14 målinger fra 1954 Berge (1958) reanalysert av Paasche (1960). Nede til venstre gjennomsnitt for perioden 2003-2020 (bare juni) med samme isolinjer som til høyre.
4.3.3 - Næringssalter
I noen deler av utredningsområdet er det god dekning av data for mengde næringssalter fra profiler tatt gjennom hele vannsøylen. Tabell 4.2 viser konsentrasjon av nitrat, fosfat og silikat langs breddegrad 75°N fra iskanten øst av Grønland til Bjørnøya. Det kan påvises forskjeller i profilene på hver side av den Arktiske fronten, men disse forskjellene er ikke mer enn på et par prosent (Børsheim 2017).
Tabell 4.2. Næringssalter langs 75ºN sensommeren 2006 som µmol kg-1 (gjennomsnitt, standardavvik, og antall prøver analysert). Data fra Børsheim (2017).
Nitrat
Fosfat
Silikat
Dybde (m)
Arktisk
Atlantisk
Arktisk
Atlantisk
Arktisk
Atlantisk
0-20
0.58±0.79(29)
2.46±0.98(28)
0.17±0.05(29)
0.25±0.06(28)
0.83±0.52(29)
1.45±0.53(28)
20-50
6.02±3.17(54)
6.08±2.95(45)
0.56±0.18(54)
0.50±0.16(45)
3.29±1.36(54)
2.97±1.05(45)
50-100
10.06±1.69(53)
9.85±2.53(39)
0.75±0.05(53)
0.71±0.09(39)
4.86±0.56(53)
4.46±0.47(39)
100-500
12.47±0.48(134)
12.20±1.11(89)
0.84±0.03(133)
0.82±0.05(88)
6.02±0.38(134)
5.39±0.59(89)
500-2000
13.62±0.70(135)
14.34±0.54(79)
0.91±0.05(131)
0.96±0.04(78)
8.35±1.78(135)
9.71±1.85(79)
>2000
14.83± 0.20(60)
15.04±0.13(17)
0.98±0.02(58)
1.00±0.02(17)
12.03±0.31(60)
12.49±0.30(17)
4.3.4 - Planteplanktondynamikk og metodebetraktninger
Havforskingsinstituttets fartøyer tar rutinemessig vannprøver fra hele vannsøylen for en rekke hydrografiske og biologiske variabler, deriblant næringssalter og klorofyll i den fotiske sonen. I tillegg settes det ut bøyer med sensorer for relevante variabler. Datadekningen fra disse målingene vil variere med prosjekter og oppdrag. For å analysere mellomårlig variasjon i våroppblomstring og andre dynamiske forhold knyttet til planteplankton, er observasjoner fra satellitt et viktig tillegg til målinger fra skip og autonome bøyer i sjøen.
Figur 4.12 Dagnummer for etablering av våroppblomstringen i de nordiske hav (dagen klorofyll overstiger 0.6 mg m-3, Siegel mfl. 2002). Basert på tidsserier av gjennomsnittlig konsentrasjon av klorofyll fra satellitt i tidsrommet 2003-2019.
Figur 4.12 viser tidspunkt for etablering av våroppblomstringen i de nordiske hav, basert på ukentlige gjennomsnitt i et kontinuerlig nettverk av ca 50x50 km store gridruter. Denne oppløsningen i tid og rom gir et bilde av hvordan våroppblomstringen langs norskekysten forflytter seg ikke bare nordover, men også ut fra kysten i retning nordvest. Mønsteret er påvirket ikke bare av lyset, men også av havstrømmene som beveger seg nordover langs norskekysten, men sørover langs Grønland. Slik detaljrikdom er et viktig supplement til data fra tokt. Satellittdata er imidlertid kun estimater avledet fra optiske indikatorer («proxies») og kan ikke erstatte in situ observasjoner, og gir bare informasjon om det øvre vannlag. Kombinert med toktdata er de et viktig verktøy for økosystemanalyse.
Utredningsområdet er et lite besøkt område i forhold til andre deler av de nordiske hav. Tettest prøvetaking foregår nærmere norskekysten, og vest for Jan Mayen er vi sjelden til stede. I de siste tiår har det blitt analysert klorofyll i profiler fra ca. 3000 stasjoner årlig i De nordiske hav, som tilsvarer havområdet som er vist i Figur 4.12. Stasjonene fra disse observasjonene ligger i hovedsak øst for utredningsområdet, og gjennomsnittlig bare 180 årlige stasjoner (minimum 48, maksimum 311) har vært analysert fra posisjoner innenfor utredningssområdet de siste ti år. Det lar seg derfor ikke gjøre å etablere baselinjeanalyser for viktige biologiske variabler basert på historiske data fra utredningssområdet.
4.3.5 - Mikrodyreplankton
Mikrodyreplankton består i hovedsak av encellede dyr fra gruppene heterotrofe mikroflagellater og ciliatene. Disse er særdeles aktive beitere i de pelagiske økosystemene. Møller mfl. (2006) fant at 60% av beitetrykket på planteplankton i Grønlandshavet i mai var fra ciliatene, og bare 30% fra kopepoder. Rapporter fra andre havområder har vist at ciliatene har tilsvarende høy andel av beitingen på fytoplankton, og at de også spiser bakterier (Schmoker mfl. 2013). Imidlertid er heterotrofe flagellater de mest aktive beiterne på bakterier, og flagellatene beites også av ciliatene (Figur 4.13). Dynamikken i våroppblomstringen er sterkt påvirket av beiting og fordelingen mikrodyreplankton blant beiterne i De nordiske hav (Børsheim mfl. 2014). For utredningsområdet har vi ikke funnet relevante prosesstudier om disse organismegruppene, men det eksisterer telletall for både ciliater og flagellater (Halldal 1953, Paasche 1960, Naustvoll mfl. 2020).
Figur 4.13 Den mikrobielle sløyfen (Wikipedia)
4.3.6 - Bakterier
Omsetning av løst organisk karbon gir produksjon av mikroorganismer, og også de minste nedbryterne bli beitet (Figur 4.13). Først de siste 40 år er dette tilfanget til næringskjedene blitt tatt hensyn til, gjerne under betegnelsen den mikrobielle sløyfen (Azam mfl. 1983, Herndl mfl. 2008). Encellede organismer uten cellekjerne utgjør to hovedgrener, eubacteria og archaebacteria. Moderne genetikk har påvist at begge grupper spiller viktige roller, både med felles prosesser og spesialiserte tilpasninger. De vil i det følgende bli omtalt samlet som bakterier. I karbonsyklusen har heterotrofe bakterier en hovedrolle som konsument av løst organisk karbon, som de delvis bruker som næring for vekst, og ellers respirerer til CO2. Organisk karbon frigjøres fra alle organismer, i hovedsak som fekalier og i mindre grad i form av kadavre. Den del av avfallet som er i oppløst form, vil i sjøvann hovedsakelig bli omsatt av bakterier. Sopp, gjær og sopplignende former som chytrider er viktige nedbrytere i terrestriske miljøer, men disse er bagatellmessige aktører i sjøvann. Ved mineralisering vil cirka 1/3 av det organiske materialet bakteriene konsumerer brukes til produksjon av bakterieceller, resten respireres til CO2. Ved å måle bakterieproduksjon får man derfor også et mål for total mengde nedbrutt organisk materiale. I havområdet over Mohnsryggen var bakterieproduksjon i de øvre 50 meter på sensommeren 1996 mellom 7 og 11 mmol C m-2 d-1, dette tilsvarer 21-33 mmol organisk C m-2 d-1 (Børsheim 2000). Figur 4.14 viser posisjoner som ble undersøkt i 1996, med integrerte verdier for bakterieproduksjon og dybdefordeling av biomasse og vekstrater.
Gjennomsnittlige verdier (snitt ± standardavvik) for biomasse hos heterotrofe bakterier, målt i august. I parantes er angitt antall minimum og maksimum for bakterieproduksjon.
Dybdeintervall
(m)
Bakterieproduksjon
(µmol C l-1 d-1)
Totall bakterier
(celler ml-1x10-5)
Vekstrate
(d-1)
0-50
0,26± 0,23 (0,02-1)
11± 7
0,7± 0,8
51-100
0,04± 0,02 (0,02-0,08)
6± 2
0,1± 0,08
101-200
0,02± 0,01(0,0002-0,05)
4± 8
0,1± 0,1
201-500
0,006± 0,004 (0,001-0,01)
2± 1
0,07± 0,06
Figur 4.14 Bakterielle variabler fra de nordiske hav på slutten av vekstsesongen (Børsheim 2000).
Nedbryting av organisk materiale omfatter langt flere elementer enn karbon, og bakteriene er sentrale i omsetning av de fleste elementene. Nitrogensyklus er et eksempel der noen biogeokjemiske transformasjoner foregår ved spesialiserte bakteriegrupper. Dyr produserer urea og ammonium, og fra dødt organisk materiale vil aminosyrer og ammonium bli avspaltet ved hjelp av bakterielle enzymer. Ammonium er i nærvær av oksygen en energikilde for spesialiserte bakterier som omdanner ammonium, via nitritt, til nitrat, og denne oksidasjonen er disse bakteriegruppenes kilde til energi for vekst. Til tross for at dette har vært kjent fra lærebøker lenge, er forståelsen av de viktige aspektene omkring disse bakteriegruppene kun i sin begynnelse, og helt nord i utredningsområdet er det påvist en rekke hydrografiske påvirkningsfaktorer (Müller mfl. 2018). Forskingsgruppen fra Universitetet i Bergen som sto bak denne genetisk avanserte studien har også vært involvert i mikrobiologiske undersøkelser ved lokaliteter i utredningsområdet, ved havbunn som er direkte påvirket av vulkanske lekkasjer (Urich mfl. 2014). Den herværende rapporten fokuserer på de øvre vannmasser, men det er viktig å påpeke at det eksisterer unik mikrobiologisk innsikt omkring områdene langs den arktiske del av den midt atlantiske rygg som er direkte påvirket av geokjemiske lekkasjer. Ekspedisjoner som har tatt prøver fra for eksempel Trollveggen nord for Jan Mayen har påvist en rekke mikrobiologiske samfunn som har helt spesiell taksonomisk sammensetning og som ivaretar helt spesielle biologiske prosesser knyttet til omsetning av de kjemiske komponentene som slippes ut fra den vulkanske aktiviteten (Storesund og Øvreås 2013, Jørgensen mfl. 2012, Lanzén mfl. 2011). Det er mikrobiologiske prosesser som er ansvarlig for hvordan utslipp av metaller og andre grunnstoffer fra undersjøiske vulkaner blir avsatt på havbunnen.
4.3.7 - Kunnskapshull, mikrobiologi
Økende mengde drivhusgasser i atmosfæren vil fortsette å gi høyere temperaturer i havet og fortsatt havforsuring. Havforsuring er diskutert i eget kapittel, men alt plankton vil påvirkes fysiologisk med høyere respirasjon ved øket temperatur. De siste tiår har temperaturen i atlantisk vann i de øvre 100 meter steget med omtrent 1ºC, og ventes å stige med 2 til 3ºC mot slutten av dette århundre (Bindoff mfl. 2019). Den fysiologiske responsen alene vil gi lavere total pelagisk biomasse (Lotze mfl. 2019). Global oppvarming påvirker også artssammensetning regionalt ved at arter generelt flytter mot høyere breddegrader (Molinos mfl. 2016, Poloczanska mfl. 2016). Denne forflytningen er fullt merkbar allerede, og betyr at vi har en kontinuerlig forandring i både artssammensetning og diversitet i vente de neste tiår.
Utredningsområdet har vært gjenstand kun for sporadiske undersøkelser av mikrobiologiske variable, og det hovedsakelig i forbindelse med prosjekter av kort varighet omkring karbonsyklus eller fiskeriene. Planteplankton har vært karakterisert i et fåtall studier. Undersøkelser av virus fra utredningssområdet er ikke kjent i tilgjengelig litteratur. Bakterier har vært undersøkt i noen få publiserte studier, hovedsakelig omkring karbonsyklus i vann nær overflaten, og lokalt i områder påvirket av vulkanske utslipp. Det finnes undersøkelser av årstidsvariasjon for planteplankton i utredningsområdet, men ikke for hverken bakterier eller virus. Kunnskap om heterotrofe mikrobielle samfunn gjennom hele vannsøylen mangler. Det er derfor ikke mulig å etablere basislinjer for sentrale biologiske prosesser i noen del av utredningsområdet.
Det er klart påvist at i sprekkområdene langs den Midtatlantiske ryggen finnes store forekomster av spesielle mikrobiologiske samfunn som er knyttet til omsetning av stofflekkasjer fra jordens indre når disse frigjøres i sjøvann. Et fåtall ekspedisjoner for mer enn ti år siden foretok undersøkelser av slike samfunn ved Mohnsryggen, og påviste spektakulære resultater, men kunnskapen er langt fra fullstendig. Et klart kunnskapshull er derfor at vi vet for lite om mikrobiologisk omsetning av kjemikalier som vil frigjøres til sjøvann dersom avsetningene langs den midtatlantiske ryggen blir gjenstand for gruvedrift.
4.4 - Mesodyreplankton
4.4.1 - Innledning
Kopepoder. Mesodyreplanktonet i Norskehavet er representert ved en rekke forskjellige grupper og arter, hvor kopepodene dominerer både antallet og biomassen (Wiborg 1954, Melle mfl. 2004). Herbivore kopepoder beiter på fotosyntetiserende planteplankton, og er selv viktig føde for karnivort dyreplankton, fisk, sjøfugl og marine pattedyr i Norskehavet (Dalpadado mfl. 2000, Melle mfl. 2004, 2014, Skjoldal mfl. 2004, Langøy mfl. 2012). Mange kopepoder regnes også som omnivore, ved at de kan ernære seg på både primærprodusenter og dyr som for eksempel mikrodyreplankton. Kopepoder kan registrere lys, men mangler i all hovedsak reelt syn. For å sanse og identifisere mulig føde, predatorer og partnere i sine omgivelser, er kopepodene avhengige av å detektere og tolke hydrodynamiske og kjemiske signaler (se Kiørboe 2011 med referanser). Fødeopptak kan foregå ved at kopepoden forholder seg stasjonær i vannet og fanger svømmende eller synkende bytter (for eksempel mikrodyreplankton) i nærheten som detekteres ved hydrodynamiske signaler, og/eller ved at fødeobjekter (som planteplanktonceller) i kopepodens egengenererte fødestrømmer detekteres og fanges individuelt (Paffenhöfer mfl. 1982, Strickler 1985, se detaljerte beskrivelser og referanser i Kiørboe 2011). Slike fødeobjekter detekteres gjerne ved lekkasjer av kjemiske stoffer fra planteplanktonceller. Beiting ved filtrering av mindre fødepartikler kan også forekomme hos små kopepoder, men denne mekanismen er ikke godt forstått ihht. Kiørboe (2011). Kopepoder er i stand til å tolke detaljert hydrodynamisk informasjon (Fields mfl. 2002). Både fjerndeteksjon av bytte ved hydrodynamiske signaler og plukking av fødeobjekter fra fødestrømmer øker dyrenes søkevolum og dermed fødeopptaksrater. Fjerndeteksjon av partner kan skje både ved hydrodynamiske og kjemiske signaler, og hvilken type signal som benyttes kan variere mellom arter. Effekten av begge er økt søksvolum, og dermed større sannsynlighet for reproduksjon i et miljø hvor avstand mellom individer er en utfordring (se for eksempel Tsuda og Miller 1998, Doall mfl. 1998, Kiørboe og Bagøien 2005 med referanser).
I Norskehavet forekommer fire Calanus-arter, hvorav Calanus finmarchicus og C. hyperboreus finnes i store mengder (Conover 1988, Melle mfl. 2004). C. finmarchicus, også kalt raudåte, er omtrent 3-4 mm lang, og vurderes å være arten som bidrar mest til mesodyreplankton biomassen i hele Nord-Atlanteren (Melle mfl. 2014), og mer spesifikt å være den viktigste kopepoden i Norskehavet på grunnlag av sin biomasse så vel som økologiske rolle (Melle mfl. 2004). I Norskehavet er C. finmarchicus særlig tallrik i den Atlantiske vannmassen øst for den Arktiske fronten, selv om den er meget vanlig også i Arktisk vann vest for fronten (Broms mfl. 2009, Melle mfl. 2014, Strand mfl. 2020 A). C. finmarchicus er essensiell mat for en rekke organismer på høyere trofiske nivå (Skjoldal mfl. 2004 med referanser). Unge individer av C. finmarchicus er hovedføden for larvene til de fleste kommersielle fiskebestander i De nordiske hav og omkringliggende områder, mens eldre individer representerer den viktigste føden for planktivore fiskebestander som sild, lodde og makrell (se Melle mfl. 2004 – Faktaboks 6.8, Melle mfl. 2014 med referanser). Den mye større slektningen, C. hyperboreus, som er 6–9 mm lang, forekommer først og fremst i kalde Arktiske vannmasser, og har sannsynligvis de dype bassengene i Grønlandshavet og Baffin Bay som sine kjerneområder (Conover 1988, Hirche 1997). Med havstrømmene transporteres denne arten sørover i Norskehavet, hvor den er mest vanlig vest for den Arktiske fronten (Melle mfl. 2004, Broms og Melle 2007, Broms mfl. 2009, Strand mfl. 2020 A). En tredje slektning, C. glacialis er knyttet til Arktiske vannmasser og særlig assosiert med kontinentalsokkelområdene i nord (Conover 1988). Den siste arten, C. helgolandicus, er en mer sørlig art som i Nordøst-Atlanteren har sin hovedutbredelse mellom Middelhavet og nordlig del av Nordsjøen (Bonnet mfl. 2005 med referanser), men som også observeres i sør i Norskehavet, og nordover i områdene påvirket av den Norske kyststrømmen.
Calanus finmarchicus gjennomfører store sesongmessige vertikalvandringer. Den tilbringer sommeren nær overflaten, og overvintrer generelt som subadult på store dyp i Norskehavet (se Melle mfl. 2014 med referanser) i en dvalelignende tilstand hvor den ikke tar til seg føde, men tærer på akkumulerte lipidreserver (Hirche 1996). Artens utbredelsessentre vurderes å være den sørlige Norskehavsgyren over Norskehavsbassenget samt den subpolare gyren i Irmingerhavet (Aksnes og Blindheim 1996, Jaschnov 1970, Conover 1988, Sundby 2000). I Norskehavet betraktes det dype Norskehavsbassenget som det viktigste overvintringsområdet (Melle mfl. 2014), men også Lofotenbassenget lenger nord vurderes å være et viktig overvintringshabitat (Halvorsen mfl. 2003). I en tidsserie-undersøkelse på Stasjon M i 1948-1949 begynte individene å akkumuleres i dypet allerede på sensommeren. Om vinteren ble mesteparten av populasjonen funnet dypere enn 600 m, og betydelige antall også dypere enn 1000 m (Østvedt 1955). Mot slutten av vinteren og tidlig om våren avtok mengdene i dypet samtidig som antallet adulte individer økte i øvre vannlag (Østvedt 1955). Om våren bruker kopepodene som har vandret opp etter overvintringen gjenværende lipidreserver kombinert med energi fra beiting på planteplanktonet til å produsere egg. Den nye generasjonen gjennomgår så 11 sekvensielle utviklingsstadier. Hovedperioden for eggproduksjon og utvikling av unge individer sammenfaller med utviklingen av planteplanktonets våroppblomstring (Diel og Tande 1992, Niehoff mfl. 1999, Niehoff og Hirche 2000, Broms og Melle 2007, Stenevik mfl. 2007). Mens flesteparten av de eldre, subadulte individene tilhørende årets generasjon vandrer ned på dypet for å overvintre, vil noen individer kunne utvikle seg til adulte og starte årets andre generasjon (Melle mfl. 2014 med referanser, Strand mfl. 2020 A).
Den herbivore, arktiske slektningen C. hyperboreus har en liknende strategi som inkluderer flere overvintringer i dypet, og det har blitt foreslått at den kan bruke inntil 3-4 år på å gjennomføre sin livssyklus i Grønlandshavet (Hirche 1997). Dette er en tilpasning til planteplanktonets korte produksjonssesong i det nordlige området. I Grønlandsgyren og Vest-Spitsbergen strømmen overvintrer C. hyperboreus generelt dypere enn hhv. 500 m og 1000 m (Hirche 1997). Det er vist at individene kan oppholde seg på dyp helt ned til 3000 m under overvintring (Hirche 1997). Oppvandringen starter i april, og nedvandringen i juli måned. Arten har en tilsvarende rolle, betydning og trofisk posisjon i Grønlandshavet som dens mindre slektning raudåte har i Norskehavet.
Figur 4.15 Stasjoner i Norskehavet med artsbestemte dyreplankton prøver med øvre lukkedyp på 1000 m eller dypere som er registrert i Havforskningsinstituttet sine databaser. Kun 2 slik stasjoner er registrert innenfor utredningsområdet. Sirkelen farget med grønt lengst sør i utredningsområdet viser Stasjon M, og det finnes noen flere prøver for denne stasjonen enn de benyttet i den påfølgende Figur 4.16.
Den årlige produksjonen til C. finmarchicus i Norskehavet er anslått å være på noen hundre millioner tonn våtvekt (ca. 190 millioner tonn våtvekt estimert av Skjoldal mfl. 2004, og ca. 290 millioner tonn våtvekt estimert av Hjøllo mfl. 2012), og den sesongmessige forflytningen av biomasse mellom overflaten og dype vannmasser antas å være stor nok til å ha betydning for den globale karbonsyklusen (Jonasdottir mfl. 2015, Boyd mfl. 2019). Selv om Calanus-artene regnes som de absolutt viktigste, finnes det i Norskehavet også en rekke andre kopepodslekter, med annen økologi og andre livshistorier. Disse kopepodene varierer i størrelse fra vel under en millimeter til opp mot en centimeter, og kan være detritusspisere, herbivore, omnivore eller karnivore, og inkluderer slektene/artene; Oncaea sp., Oithona similis, Microcalanus pusillus, Microcalanus pygmaeus, Pseudocalanus minutus, Paracalanus sp., Metridia longa, Metridia lucens, Acartia clausi, Centropages typicus, Centropages hamatus, Temora longicornis, Paraeuchaeta spp. (Wiborg 1955, Østvedt 1955, Melle mfl. 2004, Strand mfl. 2020 A og B). De numerisk dominerende taksa hos mesodyreplanktonet i Norskehavsbassenget i mai og juni 2013 (Strand mfl. 2020 B) var Oithona sp., C. finmarchicus, Pseudocalanus spp., Microcalanus pusillus, Metridia sp., larvaceer, cirripedier og ostrakoder. På Stasjon M (lokalisert langt sørøst i utredningsområdet se Figur 4.15) i 1948-1949, visualiserte Østvedt (1955) i sin Figur 6 artssammensetningen i ulike dybdeintervaller gjennom årssyklusen, og viste at den taksonomiske sammensetningen varierer gjennom sesongen både på grunt og dypt vann, og er forskjellig i overflatelagene versus på dypt vann.
Figur 4.16 Planktonets arts/slekts-sammensetning i tre dybdestrata (500-0, 1000-500, 1600-1000m) ved “Stasjon M” (se geografisk posisjon i forgående figur) eksemplifisert ved data fra januar og juni 1998. Januar dataene er basert på prøver fra 2 dybdeprofiler med Multinet, mens juni dataene er basert på kun én prøve. Merk at det ikke ble tatt prøver dypere enn 1000 m i juni – denne del-figuren må derfor ikke oppfattes som en presentasjon av manglende artsforekomster. Artsantallene er gitt på loge-skala.
I Figur 4.16 vises som et eksempel de viktigste artene som forekom i ulike dybdestrata på Stasjon M i januar og juni i 1998. Taksonomiske data fra prøver dypere enn 1000 m foreligger kun for svært få stasjoner i Norskehavet, og i dette konkrete eksemplet bare for januar-prøvene. Figuren inkluderer makroplankton som krill, amfipoder og reker. Mange av de samme slektene/artene som ovenfor ble beskrevet som typiske i Norskehavet er inkludert i prøvene fra Stasjon M i januar og juni 1998, og i tillegg vises gastropoden Limacina retroversa, ostracoder, cladocerer og pilormer. Merk at disse resultatene kun er basert på totalt tre dybdeprofiler innsamlet med Multinet, og dermed representerer resultatene enkeltanledninger fremfor generelle mønstre.
Pilormer. Pilormer (rekke Chaetognatha) er gjennomsiktige dyr med pilfasong som varierer i lengde mellom 2 mm og 10 cm. De vanligste slektene i De nordiske hav er Sagitta og Eukrohnia. Pilormer er pelagiske rovdyr som i stor grad lever av forskjellige typer mindre krepsdyr, og vurderes å ha potensiale til å kunne utøve et signifikant beitetrykk på sine byttedyrpopulasjoner. Gruppen kan forekomme i dietten til fisk, og dens betydning i det pelagiske økosystemet er trolig undervurdert. Den samlede biomassen av pilormer i Norskehavet er stor, og våre data viser at svært mye av denne biomassen befinner seg i dyphavet (Figur 4.17), med de høyeste tetthetene funnet under 500 m.
Figur 4.17 Vertikal fordeling av biomasse (tørrvekt, g m-3) av pilormer fra Norskehavet, punkter angir tørrvekt per kubikkmeter (svart er data fra natt, grå er data fra dagtid), stiplede linjer angir kumulativ andel av populasjon mot dyp. Figuren er basert på gjennomsnittlige biomassetettheter per dybdeintervall i tilgjengelige prøver tatt med 1 m2 MOCNESS planktontrål.
Geléplankton. Populærbetegnelse for organismer som tilhører to ulike grupper, hhv. nesledyr (maneter, rekke Cnidaria) og ribbe/kammaneter (rekke Ctenophora). Geléplanktonet er en mangfoldig gruppe som består av medlemmer med ulik taksonomisk tilhørighet, men som kjennetegnes ved likhetstrekk som en myk og transparent kropp (Melle mfl. 2004). Også chaetognathene som er nevnt ovenfor inkluderes gjerne i geléplankton-gruppen. Geléplanktonet kan variere voldsomt i størrelse, livnærer seg av annet dyreplankton, og kan utøve et sterkt beitepress på andre organismer i systemet. Cnidariene benytter nesleceller og tentakler til å fange bytte, og inkluderer en rekke undergrupper, med individer som kan variere i størrelse fra millimeter- til meterskala (maneter derfor er også nevnt i kapitlet om makrodyreplankton nedenfor). Eksempler er hydrozoer som typisk har kroppsstørrelse på milli- eller centimeterskala, inkludert kolonidannende siphonophorer, så vel som store scyphozoer som inkluderer kjente arter som Aurelia aurita (glassmanet), Cyanea capillata (rød brennmanet), C. lamarckii (blå brennmanet) og Periphylla periphylla (kronemanet). C. capillata kan ha skivediameter på opptil flere meter. Ctenophorene kjennetegnes av 8 kammer av cilier (flimmerhår), som brukes til forflytning. Denne gruppen mangler nesleceller, men fanger i stedet sitt bytte vha. tentakler med klebeceller (Melle mfl. 2004 med referanser). Vanlige arter i Norskehavet er Pleurobrachia pileus (sjøstikkelsbær), Beröe cucumis og Bolinopsis infundibulum (Melle mfl. 2004). Generelt er kunnskapen om geleplankton i De nordiske hav mangelfull eller begrenset. Habitatdybde, individ tettheter, og sesongutvikling vil variere for de ulike gruppene av geléplankton.
Gastropoder. En annen vanlig komponent i planktonet er pelagiske snegler (klasse Gastropoda). Av artene med skall betraktes Limacina helicina som en arktisk art, mens L. retroversa er mer typisk i de atlantiske vannmassene i Norskehavet. L. retroversa spises av fisk som sild (Dommasnes mfl. 2004), mens nakensneglen Clione limacina blir spist av predatorer som fisk, sjøfugl og hval (Melle mfl. 2004).
Figur 4.18 Vertikalfordeling av planktonbiomasse (gjennomsnittlig tørrvekt i g per m3) for størrelsesfraksjonen mellom 0.18 og 1 mm. Sorte punkter og heltrukne linjer er data for perioden mellom høst- og vår-jevndøgn (vinterdata), grå punkter og linje er data fra perioden mellom vår- og høstjevndøgn (sommerdata). Stiplede linjer viser kumulativ andel av total. For denne størrelsesfraksjonen finnes for eksempel mer enn ~50 % av total biomasse i øvre 25 m om sommer, mens mer enn 50% av biomasse finnes i øvre ~400 m om vinter. Figuren er basert på gjennomsnittlige biomassetettheter per dybdeintervall i tilgjengelige prøver tatt med 1 m2 MOCNESS planktontrål i Norskehavet.Figur 4.19 Vertikalfordeling av planktonbiomasse (gjennomsnittlig tørrvekt i g per m3) for størrelsesfraksjonen mellom 1 og 2 mm, sorte punkter og heltrukne linjer er vinterdata, grå punkter og linje er sommerdata. Stiplede linjer viser kumulativ andel av total. Figuren er basert på gjennomsnittlige biomassetettheter per dybdeintervall i tilgjengelige prøver fra 1 m2 tatt med MOCNESS planktontrål i NorskehavetFigur 4.20 Vertikalfordeling av biomasse av plankton, for plankton større enn 2 mm. Figuren viser vertikalfordeling av biomasse (gjennomsnittlig tørrvekt i g per m3) av plankton større enn 2 mm, sorte punkter og heltrukne linjer er vinterdata, grå punkter og linje er sommerdata. Stiplede linjer viser kumulativ andel av total, for denne størrelsesfraksjonen finnes for eksempel ~40 % av total biomasse fra 500 m og dypere, uansett sesong. Basert på gjennomsnittlige biomassetettheter per dybdeintervall i prøver fra 1 m2 MOCNESS planktontrål i Norskehavet.
4.4.2 - Mønstre i tid og rom
Hvert år i mai gjennomføres en internasjonal innsats for å overvåke nivået av dyreplanktonbiomasse over hele Norskehavet med tilstøtende områder (ICES 2020). Både Atlantiske vannmasser i øst, og Arktiske vannmasser i vest blir dekket, og overvåkningsområdet overlapper med mye av utredningsområdet. I den aktuelle overvåkningen måles tørrvekten til dyreplanktonbiomassen i de øverste 200 meterne av vannsøylen. Figur 2.3 i ICES rapporten viser geografiske fordelinger av biomassen aggregert over ulike tidsperioder (1995-2015, 2016-2018, og 2019). Hovedtrekket for mai-dataene i perioden 1995-2015 er at biomassene tenderte til å være høyest i den vestlige delen av innsamlingsområdet, mens det for de aller siste årene ble indikert at tyngdepunktet lå lenger øst mot Norge. Basert på disse dataene er det videre beregnet en biomasse-indeks for mai-tidsserien for årene 1995-2019. Indeksen beregnes også for hvert av fire delområder (ICES 2020, Figur 2.4). De fire subområdene er definert som “Det sørlige Norskehavet inkl. Norskehavsbassenget”, “Det nordlige Norskehavet inkludert Lofotenbassenget”, “Jan Mayen Arktiske front”, og “Området øst for Island” (se ICES 2020, Figur 2.3 for subområdenes geografiske inndeling på kart). For overvåkningsområdet i sin helhet, lå indeksen relativt høyt i årene 1995-2002, med en gjennomsnittsverdi på 11.5 g tørrvekt per m2 overflate. I perioden 2003-2006 falt gjennomsnittet, og har siden ligget lavt, selv om et visst oppsving indikeres fra omtrent 2010 (ICES 2020, Fig. 2.4). Gjennomsnittet for årene 2003-2019 var 7.9 g tørrvekt per m2 overflate for hele området. Trendene for de 4 ulike subområdene i Norskehavet for 1995-2019 følger mer eller mindre trenden for totalområdet. Årsaken(e) til den beskrevne biomassetrenden gjennom 1995-2019 er uavklart, men det er fremsatt hypoteser om at dette kan være relatert til bl.a. variasjon i bestandsstørrelsene til planktonspisende pelagisk fisk og større dyreplankton som krill og amfipoder, hvor godt vårutviklingen i dyreplanktonpopulasjonene er synkronisert med planteplanktonets våroppblomstring, samt fysiske forhold (se ICES 2020). ICES (2020) presenterer også tilsvarende data for juli-august perioden. Også her vises geografiske fordelinger av biomassen (ICES 2020, Figur 2.5). Juli/august tidsserien er kortere, 2010-2019, og indekser er beregnet for hele området og de samme fire subområdene som beskrevet ovenfor (ICES 2020, Figur 2.6). I ICES (2020) vurderer forfatterne at tidsserien er for kort til å trekke noen solid konklusjon om mulige mellomårstrender.
Havområdet rundt Jan Mayen er omgitt av Norskehavet, Grønlandshavet og Islandshavet, og inkluderer både arktiske og boreale arter av dyreplankton (Skreslet 2004). Se Fossum mfl. (2012) for detaljer om dyreplankton biomasser, arter og vertikale fordelinger i området rundt Jan Mayen. Selv om mengden dyreplankton i dette området om høsten har blitt antydet å være sammenlignbar med mengdene ved Norskekysten og i det østlige Norskehavet (se Fossum mfl. 2012), er det viktig å presisere at dyreplanktonet ved Jan Mayen ikke overvåkes like systematisk som andre områder i Norskehavet. Dette begrenser muligheten til å vurdere lokale mengder og produksjon, samt sesongutvikling og mellomårlige variasjon. Flere grupper dyreplankton kan være tallrike i områdene ved Jan Mayen (se beskrivelser og figurer 6.15 – C. hyperboreus, 6.22 – amfipoder og 6.25 – krill i Melle mfl. 2004). Den arktiske front, hvor atlantiske og arktiske vannmasser møtes, har en relativt fast lokalisering over den midtatlantiske ryggen. Frontsystemer kan være mer produktive enn omgivende vannmasser, men det er generelt sett ikke observert økt dyreplankton biomasse i pelagialen og øvre vannlag i den Arktiske fronten om våren (mai) eller sensommeren (juli/august), som er tidspunktene for de storskala overvåkningstidsseriene. Det kan derimot indikeres at enkeltområder langs fronten kan ha høyere biomasse, i hvert fall periodevis (ICES 2020). En enkeltundersøkelse av den arktiske fronten sør for Jan Mayen, viste høye konsentrasjoner av mesodyreplankton nær fronten og knyttet dette til virvel-aktivitet (C. Broms, under utarbeidelse). Vi kan derfor ikke se bort fra at dette området kan bidra til økt produksjon av plankton f.eks. ved meso-skala fysiske prosesser. Da atlantiske og arktiske vannmasser dominerer hhv. øst og vest for ryggen, blir de pelagiske vannmassene over ryggen et møtested for ulike habitater med forskjellige dyreplanktonsamfunn. Det vil derfor kunne være høyere diversitet i området. Både C. finmarchicus og C. hyperboreus finnes i store mengder i området (Hirche 1991, Melle mfl. 2004, Broms mfl. 2009). Sesongutviklingen av C. finmarchicus er forsinket på den arktiske siden av fronten sammenliknet med den atlantiske siden, noe som fører til ulik sammensetning av stadier på tvers av fronten. Det finnes generelt lite informasjon angående planktonproduksjon og variasjon i mengden av ulike arter langs frontområdet gjennom sesongen og mellom år.
4.4.3 - Vertikale mønstre
Havforskningsinstituttet sin overvåkningsinnsats fokuserer i hovedsak på de øvre 200 m av vannsøylen. Dette er begrunnet i en implisitt antagelse om at det er her hoveddelen av biomassen av mesodyreplankton befinner seg, og at de viktigste trofiske interaksjoner som involverer mesodyreplankton forekommer i dette dybdeområdet (f.eks. fødeinntak hos mesodyreplankton og kommersielle bestander av fisk). Dette er en sannhet med modifikasjoner, og hvor korrekt dette paradigmet er avhenger sannsynligvis av årstid og hvilke grupper av mesodyreplankton man fokuserer på. For eksempel er biomassefordeling av mesodyreplankton i størrelsesfraksjonen mindre enn 2 mm i sommerhalvåret helt i overensstemmelse med disse antagelsene (Figurer 4.18 og 4.19), mens dataene viser at for vinterhalvåret, samt for større komponenter (Figur 4.20) finnes mesteparten av biomassen dypere enn 200 m. Våre data viser at gjennomsnittlig biomasse av potensielt viktige predatorer som pilormer, amfipoder og reker øker mot dypet, noe som antyder at prosesser dypere enn 200 m er viktig for den overordnede pelagiske økologien i området, men det er store hull i vår kunnskap om energiflyt og interaksjoner i dyphavet.
4.5 - Makrodyreplankton og mikronekton
4.5.1 - Innledning
Gitt korrekt metodikk lar det seg fint gjøre å kvantifisere mengden av makroplankton innen et definert område, men gruppen er i et størrelsesspenn der kunnskapsgrunnlaget per i dag er mindre enn for mange andre grupper: disse organismene er for små til å måles kvantitativt med utstyret vi bruker for å fange pelagisk fisk (e.g. typiske fisketråler, mesteparten av makroplanktonet forsvinner gjennom maskene på trålen), men er for store til at det vanlige planktonutstyret (e.g. håver) gir oss gode estimater, bl.a. på grunn av aktiv unnvikelse. Flere av de viktigste artene kan også danne tette svermer eller stimer, noe som igjen fordrer større innsats fra vår side for å oppnå gode estimater. Grunnet utstrakt bruk av innsamlingsutstyr som ikke er egnet for kvantitativ innsamling av makroplankton, prøvetagning som dekker bare deler av den vertikale utbredelsen til makroplanktonet og generell lav innsamlingsinnsats rettet direkte mot makroplankton, bør selv fagfellevurdert tallmateriale på biomasser og tettheter i utgangspunktet ansees som usikre tall.
Den mest komplette oversikten over makroplankton økologi og ressurser i Norskehavet som helhet finnes i Skjoldal mfl. (2004). Utover dette har HI samlet en del materiale med spesialiserte tråler (“Makroplanktontrål”), spesielt i det siste tiåret. Disse dataene er i stor grad upublisert (men se for eksempel Klevjer mfl. 2019). Det er imidlertid allokert relativt liten innsats til dette arbeidet, og selv om grov romlig fordeling kan estimeres, vil det sannsynligvis ikke være nok data til å kunne vurdere for eksempel sesongvariasjoner i dette materialet. Disse tråldataene er heller ikke dybdestratifiserte, og det er kun i de senere år innsamlingen har foregått så dypt som 1000 m, mange av trålene har kun gått ned til 500 m dyp. Havforskningsinstituttet har nesten ikke data fra kvantitative tråler dypere enn 1000 m i Norskehavet, og biomassetettheter av makroplankton i bathypelagiske dyp er derfor høyst usikre. Innsamlingen med tråler er heller ikke egnet til å estimere populasjonsnivåer for makroplanktonarter som danner tette svermer, som for eksempel noen av krill- og amfipode- artene. For de fleste av artene har man derfor ikke tilfredsstillende oversikt over eventuelle populasjonstrender over tid i Norskehavet.
Makroplankton utgjør en viktig komponent i fødeutvalget til fisk og hval, og dyr fra denne gruppen er også viktige som beitere på planteplankton (krill) og som predatorer på mindre dyreplankton som kopepoder. Makroplankton er sannsynligvis også viktige i overføringen av karbon til dyphavet, både som transportører (e.g. aktiv transport gjennom vertikal døgnvandring hos for eksempel krill og mesopelagisk fisk), “feller” (dyptlevende makroplankton spiser på vertikalvandrende plankton), og i “tapsleddet” av vertikal-transporten (e.g. dyptlevende makroplankton spiser enten direkte på utsynkende materiale, eller på organismer som spiser på dette materialet). Økologiske interaksjoner mellom makroplankton og lavere og høyere trofiske nivåer er strukturert vertikalt, det meste av vår kunnskap er om prosesser som foregår i de øvre vannlag (hovedsakelig epipelagisk), og det er store hull i vår kunnskap om pelagiske biomasser og prosesser i dyphavet. Under følger en kort beskrivelse av de forskjellige gruppene av makroplankton, inkludert en oppsummering av antatt vertikal fordeling.
4.5.2 - Amfipoder
Pelagiske amfipoder (Hyperiidae) er en svært viktig økosystemkomponent i store deler av utredningsområdet. Særlig viktig er slekten Themisto, som inkluderer artene Themisto abyssorum og T. libellula som er spesielt tallrike i henholdsvis Atlantiske og Arktiske vannmasser. Begge artene er predatorer på annet dyreplankton. Særlig hoppekreps (f.eks. Calanus finmarchicus) men også andre typer dyreplankton, eksempelvis pilormer står på menyen. T. libellula er tradisjonelt antatt å være en organisme som er viktig i de øverste vannlag fra omtrent 300 m til overflaten. Den er viktig føde for blant annet Grønlandssel, torsk og polartorsk i områder i nærheten av iskanten og i Arktiske vannmasser i det nordlige Atlanterhavet. Svermer av T. libellula er imidlertid også observert på betydelige dyp i Norskehavet (Aarbakke 2007) og i Grønlandshavet (~1270 m, Vinogradov 1999). Lignende observasjoner av T. abyssorum er gjort på Vøringplatået i ca. 800 meters dyp like over bunn (Vinogradov 1999), assosiert med typiske Atlantiske vannmasser i de øvre vannlag, men hvor det over bunn er langt kaldere vannmasser som dominerer.
Betydelig usikkerhet er knyttet til livshistorien til begge disse amfipodene, og det finnes begrenset kunnskap om hvordan disse gjennom livsløpet utnytter vannsøylen med hensyn til reproduksjon, fødeopptak, mulig overvintring, osv. Grove anslag av Dalpadado mfl. (1998) og Olsen mfl. (2006) har antydet at stående biomasse av amfipoder i De nordiske hav kan være i størrelsesorden 40-200 millioner tonn i årets tredje kvartal. Amfipod-biomassen, vurdert på grunnlag av Havforskningsinstituttets trålhal i årene 1990–1999, ble for Norskehavet, Grønlandshavet og Islandshavet funnet å være aller høyest i området vest og nord for Jan Mayen (Melle mfl. 2004). T. libellula, den største arten i denne slekten, er tallrik i det kalde vannet i vestlige deler av Norskehavet (Dalpadado mfl. 1998, Skjoldal mfl. 2004). Mens denne arten typisk opptrer i de øvre vannlag, og ofte i form av svermer, viser HI sine fangster (MOCNESS) at mye av biomassen av gruppen amfipoder også finnes i dyphavet (Figur 4.21), med ~50% biomassen av gruppen estimert til å leve dypere enn 500 m.
Figur 4.21 Vertikal fordeling av amfipod-biomasse (tørrvekt, g m-3) i MOCNESS prøver fra Norskehavet. Data basert på aritmetisk gjennomsnitt. Svarte punkter og linjer er data fra natt, grå er data fra dagtid, stiplede linjer angir kumulativ andel av populasjon mot dyp.
4.5.3 - Krill
Krill (Euphausiidae) er en annen sentral økosystemkomponent i utredningsområdet hvor man kan observere 3-6 arter, og der Thysanoessa inermis, Meganyctiphanes norvegica og T. longicaudata er de antalls- og biomassemessig viktigste. Storkrillen M. norvegica, som når en totallengde på ca. 4 cm, er den største og sannsynligvis biomassemessig viktigste krillarten. Den er svært tallrik langs kontinental-skråningene, særlig i øst, og følger Atlanterhavsstrømmen vest og nord for Svalbard. Den finnes også i de sentrale deler av Norskehavsbassengene, og i området rundt Jan Mayen. Arten M. norvegica er lyssensitiv, og foretar døgnlige vertikalvandringer. Den befinner seg i overflatelagene om natten, og vandrer til dypere vann, gjerne ned til 500 m eller dypere på dagtid (Knutsen og Serigstad 2001).
En annen viktig krillart er T. inermis, hvis voksne individer når en totallengde på 25-32 mm. Den er særlig knyttet til kontinentalsokkel-områdene rundt Svalbard og området rundt Jan Mayen, men er også regelmessig til stede i norske kystfarvann. Voksne individer er registrert så dypt som ~ 700 m over kontinentalskråningen i det østlige Norskehavet. Også denne arten er lyssensitiv og foretar døgnlige vertikalvandringer, men dens vertikale fordeling i vannsøylen avhenger også av årstid og generelle fysiske forhold, og kan variere for området over kontinentalsokkelen versus dyphavet. Den tredje hyppig forekommende krillarten i området er T. longicaudata. Den er langt mindre i størrelse enn artene nevnt over og dens totale biomasse mest sannsynlig mye lavere. I hovedsak er arten knyttet til Atlantisk-påvirkede vannmasser, og den har en 1-2 årig livssyklus. Overvintring skjer på dypt vann i Norskehavet og den har en sesongmessig oppvandring til overflatelagene om våren.
Det er velkjent at en rekke krillarter kan samles i veldefinerte lag vertikalt i vannsøylen, men også tidvis i større og ofte dynamiske stimer. Denne type ansamlinger og adferd kan være knyttet til predatortetthet eller et resultat av parrings-/gyteadferd da hunner og hanner må være i direkte kontakt med hverandre. Grove anslag indikerer at stående krillbiomasse i De nordiske hav kan være i størrelsesorden 30 – 160 millioner tonn i årets tredje kvartal (Dalpadado mfl. 1998, Melle mfl. 2004, Olsen mfl. 2006).
Høy krillbiomasse (alle arter sett under ett) er indikert nær Jan Mayen, basert på horisontal interpolering av data (se Melle mfl. 2004, Figur 6.22). Nyere estimater basert på kvantitative tråler anslår biomassen i Norskehavet til ~1-2 g våtvekt per m2 (Klevjer mfl. 2019), men dette er konservative tall, som ikke inkluderer biomasse tilhørende svermer, for både krill og amfipoder antar vi at mye av biomassen vil finnes i slike tette svermer.
T. inermis vurderes å være en sokkel-art (Skjoldal mfl. 2004), og fordelingsmønsteret kan indikere at den hører hjemme i kaldt vann (Dalpadado mfl. 1998). Krillen T. longicaudata betraktes å være en basseng-art, og data viser at denne arten forekommer i store deler av De nordiske hav. Krillen Meganyctiphanes norvegica er utbredt over store deler av Nord-Atlanteren, tidligere ble denne arten vurdert å være knyttet til sokkel-skråninger (Skjoldal mfl. 2004), og arten er vanlig i det varmere atlantiske vannet lenger øst i Norskehavet (Dalpadado mfl. 1998). Nettfangster antyder at mye av biomassen av krill finnes på mesopelagiske dyp på dagtid, e.g. Figur 4.22, men at det kun finnes relativt lave biomasser under ~500 m i det sentrale Norskehavet.
Figur 4.22 Vertikal fordeling av krill-biomasse (tørrvekt, g m-3) i MOCNESS prøver fra Norskehavet. Punkter angir tørrvekt per kubikkmeter (svart er data fra natt, grå er data fra dagtid), stiplede linjer angir kumulativ andel av populasjon mot dyp.
4.5.4 - Pelagiske reker
Pelagiske reker representert ved slektene Sergestes, Hymenodora, og Pasiphaea kan bli relativt store, med totallengder på ca. 5-20 cm. De er tallrike og har sitt kjerneområde dypt i pelagialen i Norskehavet. Dog kan larver og yngre individer av enkelte arter være tallrike høyere opp i vannsøylen. Flere av artene foretar døgnlige vertikalvandringer. Dybdeintervallet de vandrer over er trolig noe mindre enn beskrevet for storkrill (M. norvegica), men voksne individer befinner seg sannsynligvis på et noe større gjennomsnittlig dagdyp enn storkrillen. Disse rekene representerer en betydelig komponent i det dyp-pelagiske økosystemet i utredningsområdet, er predatorer som beiter på annet dyreplankton og er viktige aktører i økosystemet, bl.a. ved å bringe organisk materiale fra de øvre til dypere deler av vannsøylen. De er mest sannsynlig også viktige fødeorganismer for fisk, blekksprut og maneter.
Når det gjelder mengden av disse pelagiske rekene, eksisterer få estimater av tetthet og biomasse (men se f.eks. Dalpadado mfl. 1998). Tidligere estimater er mest sannsynlig veldig usikre, som de også er for krill og amfipoder (se anslag over), særlig fordi disse var basert på trålfangster som sannsynligvis ikke dekket disse organismenes vertikale utbredelse. I data fra kvantitative tråler kan denne gruppen utgjøre omtrent samme biomasse som summen av krill, amfipoder og mesopelagisk fisk (Klevjer mfl. 2020), men det er som sagt grunn til å anta at disse tråldataene ikke er kvantitative for stimende krill og amfipoder. Havforskningsinstituttets data basert på dybdeoppløste MOCNESS prøver antyder at mye av biomassen av pelagiske reker øker med dypet (Figur 4.23), og det er derfor svært sannsynlig at våre trålestimater (som bare går ned til ca. 1000 m) er underestimater av total biomasse av pelagiske reker i Norskehavet.
Figur 4.23 Vertikal fordeling av biomasse (tørrvekt, g m-3) for pelagiske reker, basert på aritmetisk gjennomsnitt av MOCNESS-prøver tatt i Norskehavet.
4.5.5 - Maneter
Maneter er en fellesbenevnelse for en heterogen gruppe organismer som har sin opprinnelse i to ulike phyla: Cnidaria (nesledyr) og Ctenophora (kammaneter). Ulike typer av nesledyr og kammaneter finnes fra overflaten til de største havdyp. Mange grupper/arter er flerårige og de kan være tallrike i mesopelagialen (200-1000 m). Kronemaneten Periphylla periphylla er en slik nøkkelorganisme i det mesopelagiske samfunnet og Dalpadado mfl. (1998) beregnet den stående biomassen av denne arten til 11 millioner tonn i Norskehavet. De totale biomassene til disse gruppene er veldig usikre.
4.5.6 - Mesopelagisk fisk
Disse regnes oftest som mikronekton, men det finnes foreløpig lite vitenskapelig støtte for at disse fiskene vandrer horisontalt, så de kan regnes som funksjonelt plankton (Kaartvedt mfl. 2008). Viktige arter i Norskehavet er Nordlig lysprikkfisk (Benthosema glaciale), laksesild (Maurolicus muelleri) og liten laksetobis (Arctozenus risso) (Dalpadado mfl. 1998) (Tabell 4.3). Disse er alle flerårige. Laksesild er knyttet til sokkelkantene (nerittisk art) i Atlantisk vann, og forekommer derfor i relativt liten grad sentralt i bassengene (f.eks. Dalpadado mfl. 1998, Figur 6). Fangster antyder også at den lille laksetobisen hovedsakelig er fanget i Atlantisk vann (e.g. Dalpadado mfl. 1998, Figur 7).
Tabell 4.3 Mesopelagiske/bathypelagisk arter påvist i Norskehavet kategorisert med økotype i hht. Mecklenburg mfl. (2018).
Familie
Art
Økotype
Gonostomatidae
Lysfisker
Cyclothone microdon
Svart lysfisk
Mesopelagisk/ Bathypelagisk
Sternoptychidae
Laksesilder
Argyropelecus hemigymnus
Flekket perlemorsfisk
Mesopelagisk
Maurolicus muelleri
Laksesild
Mesopelagisk
Stomiidae
Storkjefter
Chauliodus sloani
Hoggormfisk
Mesopelagisk/ Bathypelagisk
Paralepididae
Laksetobiser
Arctozenus risso
Liten laksetobis
Pelagisk/Meso-pelagisk
Myctophidae
Lysprikkfisker
Benthosema glaciale
Nordlig lysbrikkfisk
Mesopelagisk
Lampanyctus macdonaldi
Brun lysprikkfisk
Mesopelagisk
Myctophum punctatum
Liten lysprikkfisk
Mesopelagisk
Notoscopelus kroyeri
Stor lysprikkfisk
Mesopelagisk
Trachipteridae
Båndfisker
Trachipterus arcticus
Sølvkveite
Pelagisk/Meso-pelagisk
4.5.7 - Blekksprut
En rekke pelagiske blekksprutarter forekommer i utredningsområdet. I hovedsak er det imidlertid én art, Gonatus fabricii som dominerer i området, og som det finnes data for. Arten er vidt utbredt og tallrik i Nord-Atlanteren fra Nordpolen til de sørlige områdene av Norskehavet (Arkhipkin og Bjørke, 1999). Den er en viktig fødeorganisme for blant annet fisk, flere hvalarter og sel. De yngste og minste individene (0.3-4 cm PL, pennlengde) er hovedsakelig lokalisert i de øvre 80 m av vannsøylen (Bjørke og Gjøsæter, 1998). Arten viser en ontogenetisk vandring til dypere vann ved økende størrelse, og individer større enn 200 mm PL (pennlengde) har blitt fanget på dyp fra 400 til 1200 m, som er maksimumdypet det ble fisket på av Bjørke og Gjøsæter (1998). Det foreligger tall på stående biomasse og produksjon, henholdsvis 8.2 og 20 mill. tonn i det som kan defineres som De nordiske hav (se Bjørke og Gjøsæter 1998, Olsen mfl. 2006).
4.5.8 - Dyreplankton ved varme havkilder
Innledningsvis har vi beskrevet hvordan Havforskningsinstituttets overvåking av dyreplankton i de Norske havområdene er lagt opp, hvor hyppig dette foregår og hvilken metodikk som anvendes (kapittel 4.2). Den kanskje viktigste måten vi får informasjon fra områder og dyp hvor ellers få prøver er tatt, er i forbindelse med spesialundersøkelser, f.eks. knyttet til studier av biologi og miljøforhold i nærheten av undersjøiske skorsteiner på den midtatlantiske ryggen (se Aarbakke 2007, Schander mfl. 2010, Olsen mfl. 2014, 2015, Falkenhaug mfl. 2016, Falkenhaug og Hobæk 2017, 2021). Schander mfl. (2010) studerte faunaen rundt hydrotermale kilder på Mohnsryggen. På de undersøkte lokalitetene var bunndypet rundt 500-700 m og altså betydelig grunnere enn gjennomsnittsdypet i Norskehavet og Grønlandshavet på hver sin side av ryggen. Det beskrives at svermer av planktoniske organismer ble observert over disse kildene og det vises til at lignende skyer er observert i andre tilsvarende områder (Wiebe mfl. 1988, Thomson mfl. 1992, Burd og Thomson 1994, 1995, Burd mfl. 2002), men det understrekes at lite er kjent mht. trofiske relasjoner. Det er også klart at der ved slike undersøkelser ikke alltid foreligger data på mengde eller tetthet av organismer.
I løpet av BioDeep2006 toktet ble det gjennomført foreløpige studier og prøvetaking av de hydrotermiske «planktonsvermene» som først ble sett under BioDeep2005, i et område lokalisert ca. 50 km nord for Jan Mayen på 71°N (jfr. Pedersen mfl. 2005). Dette ble gjort ved bruk av akustisk utstyr og Mocness-trål. Fra dette materialet ble det konkludert at planktonskyene hovedsakelig besto av krepsdyr, og 18 arter/grupper ble identifisert (se Aarbakke 2007, Table II). Der foreligger også en del kvantitative data fra disse preliminære undersøkelsene (se Aarbakke 2007, Table V-VI). Imidlertid er det lite i dette materialet (Aarbakke 2007), som indikerer at tettheten av mellomstort dyreplankton («mesodyre-plankton», jfr. Tabell 4.4) er veldig forskjellig i disse hydrotermiske områdene sammenlignet med det man har registrert i tilsvarende dyp, men over de abyssale slettene øst og vest for den midtatlantiske ryggen.
Tettheten av individer som befinner seg i området nær disse hydrotermale feltene (jfr. Tabell 4), er ikke vesentlig forskjellig fra det en finner på tilsvarende dyp andre steder i Nord-Atlanteren selv om sammenligningen kan være vanskelig gitt tidspunkt for data som sammenlignes, også i forhold til sesongmessig utvikling i dyreplanktonet i de områdene prøvene ble tatt. Det kan nevnes at like vest og nord av Svalbard i 2014 (st 551, 80º49.5’N, 15º33’Ø) og (st 547, 79º38.13’N, 5º08.33’Ø), den siste lokaliteten ikke langt fra Molloy dypet, så var tettheten av mellomstore hoppekreps (av størrelse Calanus og Metridia) henholdsvis ~100 ind./m3 og ~375 ind./m3 mellom 250-500 m dyp (SI_Arctic, Ingvaldsen mfl. 2016). Dette er klart mer enn det som ble observert av Aarbakke (2007), ~4-12 ind./m3 (jfr. Tabell 4.4).
Det gjøres oppmerksom på at prøvene i Tabell 4.4 (jfr. Aarbakke 2007) ble tatt i juni 2006, mens prøvene fra SI_Arctic (se Ingvaldsen mfl. 2016) er tatt noe lengre nord og seinere i sesongen, 23.-25. august, delvis utenfor utredningsområdet. Også tidspunktet spiller inn og Aarbakkes data viser at hovedtyngden av den viktigste hoppekrepsen, Calanus sp., i hovedsak står i de øvre 100 m i juni måned, og tilsynelatende ikke har vandret ned til overvintringsdyp enda.
Aarbakke (2007) benyttet også en Makroplankton trål (jfr. Wenneck mfl. 2008), til å ta prøver av makroplankton og mikronekton. Prøve ble tatt ved å trekke trålen horisontalt ca. 450 m i 80 minutter, i en ekkoregistrering observert i omtrent det samme dypet. De fleste av artene som ble identifisert fra trålprøvene i denne undersøkelsen, er kjente og rutinemessig funnet i tilstøtende hav (Dalpadado mfl. 1998, Melle mfl. 2004). Den gammaride amfipoden Eusirus holmii regnes som arktisk og er kjent fra de omkringliggende hav; Barentshavet, Karahavet, Laptevhavet og Grønlandshavet (Macnaughton mfl. 2007). Arten har tidligere blitt funnet nord / nord-vest og sør-øst for Island (Weisshappel 2000) og under den vestlige iskanten av Spitsbergen (Macnaughton mfl. 2007). Arten synes å være assosiert med havisen i Arktis, men er også funnet nær bunnen på 1200 m dyp (se Macnaughton mfl. 2007). Denne siste observasjonen er interessant siden den speiler opplysninger om de hyperiidae artene i slekten Themisto som omtales under.
Noen av planktonsvermene som er omtalt av Aarbakke (2007) og Schander mfl. (2010) er trolig en beskrivelse som har sitt utgangspunkt i hva som ble sett på video-opptak fra ROV’en som ble benyttet i det samme området under BioDeep2005 (se Aarbakke 2007, Figur 18-19). Amfipodene på disse bildene ble bestemt til Themisto libellula. Flere forfattere har beskrevet at de to amfipode-artene T. abyssorum og T. libellula tidvis observeres nær bunn på veldig store dyp: Svermer av T. libellula er observert i Grønlandshavet (~1270 m, Vinogradov 1999). Observasjoner av T. abyssorum (Atlantisk boreal art) er gjort på Vøringplatået i ca. 800 meters dyp like over bunn (Vinogradov 1999). Interessant nok i et område assosiert med typiske Atlantiske vannmasser i de øvre vannlag, men hvor det over bunn er langt kaldere vannmasser som dominerer (jfr. Knutsen og Serigstad 2001, Figur 2).
I forbindelse med oppfølgende geologiske undersøkelser av hydrotermale felt i Norskehavet, har det også blitt gjennomført noen delundersøkelser med fokus på planktoniske organismer. Olsen mfl. (2014, 2015) var spesielt ute etter å studere amfipoden T. abyssorum fordi preliminære undersøkelser hadde identifisert denne boreale pelagiske amfipoden som en potensielt viktig predator i dette kjemosyntetiserende habitatet. Prøver ble samlet på to vitenskapelige tokt med FF G. O. Sars i 2009 og 2011 (Olsen mfl. 2014), henholdsvis på Loki’s Castle (Lokeslottet/LOKI, Pedersen mfl. 2010a), Hakon Mosby Muddervulkan (HMMV) og på det hydrotermale feltet nord-øst av Jan Mayen (se også Aarbakke, 2007). LOKI (2350 meters dybde) er et svartrøykerfelt («black smoker field» med væsketemperatur på 310–320°C som er lokalisert i overgangen mellom Mohnsryggen og Knipovichryggen vest for Bjørnøya (73°30′N og 8°E). For ytterligere detaljer ang. plassering og biologi, vises det til Pedersen mfl. (2010a) og Tandberg mfl. (2011). Håkon Mosby Muddervulkan (på ca. 1300 m dybde), er en såkalt «cold seep» muddervulkan som ligger i overgangen mellom Barentshavet og Norskehavet (Niemann mfl. 2006). Det hydrotermiske feltet nord-øst for Jan Mayen ligger betydelig grunnere (550-750 m dybde), og består av flere undersjøiske «vulkaner», såkalte «white smokers» hvor den utstrømmende væsken har en temperatur opp mot 270°C. Feltet ligger på Mohn-ryggen nord-øst for Jan Mayen (71°18′N og 5°46′W) (Pedersen mfl. 2005, 2010b, Schander mfl. 2010, Sweetman mfl. 2013). Undersøkelsene i 2009 og 2011 hadde ikke primært fokus på å bestemme mengden av dyreplankton i områdene ved disse vulkanene. Likevel ble det benyttet en liten kvantitativ flerpose planktontrål, en 1 m2 Mocness (Wiebe mfl. 1976, 1985) på Lokeslottet og ved Håkon Mosby Muddervulkan. Dette er et redskap som gjør det mulig å ta en prøve innenfor et ønsket dybdeintervall og opptil 8 nett kan åpnes og lukkes manuelt fra forskningsfartøyet. Innsamlingen foregikk ved at redskapet ble trukket horisontalt så nær utslippet fra vulkanene som mulig, og flere nett åpnet og lukket i omtrent samme dyp. På Jan Mayen feltet ble det brukt et vertikalt operert Multinet Midi (Hydrobios, 300-µm maskevidde, 0.25 m2 åpning), for å samle inn materiale over den hydrotermale kilden Trollveggen. Imidlertid ble det funnet veldig få individer av disse amfipodene, selv om flere prøver ble tatt. Det ble nødvendig å supplere materiale ved også å ta i bruk en sugepumpe montert på ROV’en som ble benyttet, men det ble uansett innsamlingsredskap kun hentet ut og arbeidet videre med planktonmateriale av amfipodeslekten Themisto. Gitt at det var vanskelig å få nok individer av Themisto med de tradisjonelle innsamlingsmetodene, indikerer dette at lokaliteten som ble samplet neppe kan ha hatt veldig høye tettheter av disse amfipodene. Imidlertid er det kjent at dyreplankton kan være veldig flekkvis fordelt i tid og rom. Hydrotermale felt er irregulært fordelte og ansamling av bakterier eller andre typer organismer, fastsittende eller hyperbenthiske, kan derfor bidra til å strukturere det biologiske miljøet som igjen kan utnyttes av organismer som vi vet er i stand til å foreta betydelige vertikale vandringer (se Mauchline og Fisher 1969, Tarling mfl. 1998, Vinogradov 1999, Hirai og Jones 2012). Men samtidig kan det være store avstander mellom slike hydrotermale kilder, hvor tettheten av dyreplankton ikke er vesensforskjellig fra de åpne vannmassene lengre ut fra den midt-Atlantiske ryggen.
Basert på DNA fra mageinnhold hos Themisto fant Olsen mfl. (2014) et bredt spekter av byttedyr inkludert mikro-eukaryoter og flercellede organismer, samt detritus. Themisto abyssorum fanget ved Lokeslottet hadde det høyeste mangfoldet av byttedyr. Dette antyder at T. abyssorum kan ta til seg føde fra mere enn ett trofisk nivå og at arten derfor bør betraktes som en omnivor («alteter») og ikke en streng carnivor (rovdyr) slik mesteparten av litteraturen har beskrevet arten tidligere. En del av dyreplankton prøvematerialet tatt med Mocness som er fiksert på absolutt alkohol fra BioDeep2009, er fortsatt tilgjengelig ved Havforskningsinstituttet, mens en del annet materiale er lagret ved Universitet i Bergen. En oversikt over utplukket hoppekrepsmateriale (Copepoda) fra prosjektene BioDeep2009 og SI_Arctic stasjoner som ligger i eller nær utredningssområdet, er gitt i Tabell 4.5. Disse artene ble funnet i prøvene, men det sier ingenting om hvilke andre organismer som var til stede, eller var dominante i prøvene.
4.5.9 - Hva gjør Themisto ved bunnen på store dyp?
Observasjoner av større ansamlinger av amfipoder nær havbunnen (på dybder rundt 1000 – 3000 m), er rapportert for de nordlige Themisto-artene, T. abyssorum og T. libellula (Havermans mfl. 2019). Slike ansamleringer ble forklart med at dyrene beitet på detritus eller planteplankton (Vinogradov 1999 med referanser). I Svalbard-farvann ser T. libellula ut til å være den viktigste fødeorganismen til vortekjeks («Atlantic spiny lumpsucker», Eumicrotremus spinosus). Dette er en bunnfisk det antas ikke er blant de hurtigste svømmerne og som man neppe ville tro er en aktuell predator på raskt svømmende og vandrende amfipoder i vannsøylen (Berge og Nahrgang 2013). Tilsynelatende migrerer T. libellula til bunnområdene i løpet av dagen hvor den samler seg, en adferd som er observert ved hjelp av et nedsenkbart kamerasystem (Vinogradov 1999) og artene utgjør derfor en midlertidig, men viktig komponent av hyperbenthos – en gruppe organismer som er tidvis svært tallrike i området like over bunn. Det samme kan også være tilfelle for T. gaudichaudii i Antarktis, siden individer av denne arten har blitt funnet i epibenthiske sledefangster på dybder rundt 3000 m (jfr. Havermans mfl. 2019, Havermans C. upubliserte data). På kontinentalsokkelen rundt Prince Edward-øyene i Antarktis har T. gaudichaudii ofte blitt fanget med trålredskap nær bunnen (Pakhomov og Froneman 1999), mens voksne T. japonica er registrert på 3000 m dyp (Semenova 1974). Fra en rekke kilder ser vi nå at slekten Themisto som har representanter i viktige områder i verdenshavene, tidvis befinner seg i større ansamlinger nær bunn, hvor de kan utgjøre en viktig føderessurs for bunnlevende fisk eller evertebrater og flere av disse Themisto-artene er dokumentert å gjennomføre betydelige vertikale vandringer. Hvorvidt dette representerer tradisjonelle døgnlige vertikalvandringer som er velkjent hos krill, er fremdeles ikke avklart (Vinogradov 1999). Disse organismene har sannsynligvis en svært viktig funksjon når det gjelder karbonsyklusen. Derfor bør en bl.a. avdekke hvordan dietten hos juvenile og voksne individer av disse artene er sammensatt, det gjelder i forhold til planteplankton i vannsøylen og mulig fødeopptak på/i sedimentene på havbunnen. Slik kunnskap vil ha stor betydning for vår forståelse av konsekvensene for pelagisk-benthiske koblingsprosesser og den biologiske karbonpumpen. Vertikale migrasjoner til havbunnen for disse amfipodene kan også ha å gjøre med skallskifte, reproduktiv adferd og muligens frislipp av juvenile fra hunnenes ynglekammer (Havermans mfl. 2019). Men også kunnskapen om disse forholdene er svært mangelfull per i dag, inkludert for Themisto-artene i Nord-Atlanteren.
4.5.10 - Krillansamlinger nær bunn
Flere av krillartene som forekommer i Nord-Atlanteren gjennomfører betydelige døgnlige vertikale vandringer. En av disse artene er Meganyctiphanes norvegica – også kalt storkrill og endatil fra gammelt av «raudnakkåte». Den er kanskje den viktigste krillarten i Nord-Atlanteren, og finnes sentralt over dypbassengene, men er kanskje særlig knyttet til kontinentalskråningene langs Nord-Atlanterens randsoner (jfr. beskrivelsene for Makroplankton og Mikronekton, se kapittel 4.4.2). Denne arten og andre er studert inngående over mange år, både i felt og i eksperimentelle settinger (se Mauchline og Fisher 1969, Mauchline 1980, Schmidt 2010). Et sentralt aspekt her er artens fødepreferanser og dens betydelige vertikale døgnlige vandringer. M. norvegica er ofte observert ved bunn på dypt vann i forbindelse med ROV operasjoner og er fanget på videoopptak ved en rekke anledninger (se Hirai og Jones 2012). Om arten har fulgt utsetting av ROV’ene og blitt med ned i dypet, fordi den potensielt kan tiltrekkes av lyskasterne på slike farkoster, vites ikke sikkert. Det kan være en mulig forklaring, men med stor sannsynlighet tiltrekkes dyr som allerede er ved bunn disse lyskildene og konsentreres i området rundt farkosten når lys slåes på.
Fra eldre undersøkelser vet vi at storkrillen i enkelte områder oppholder seg svært nær bunn, f.eks. i Loch Fyne, Skottland (MacDonald 1927). Selv om dette ikke er av de dypeste områdene hvor arten finnes, er føden i stor grad vegetarisk detritus fra land og kystområdet rundt. Spesielt syntes dette å være en viktig fødekilde for noe større individer med totallengde ca. 21-29 mm. M. norvegica er en viktig aktør i det planktoniske samfunnet i Clyde-området. Den er tidvis tallrik, og i området hvor maksimalt bunndyp er rundt 110-150 m, står den ofte mellom 18 og 36 m over en gjørmete sedimentoverflate. Senere studier bekrefter disse observasjonene i Firth of Clyde området og viser at den mest vanlige bestanddelen i mageinnholdet til krillartene Thysanoessa raschii og M. norvegica var suspendert materiale og rester av sedimenter (Mauchline 1960, Mauchline og Fisher 1969). I St. Lawrence gulfen fant Berkes (1973) mudder i magene til krillartene T. inermis, T. raschii og M. norvegica, men bare i januar måned, mens undersøkelser av Sameoto (1980) i det samme området dokumenterer at M. norvegica aktivt beitet på kopepoder (hoppekreps) i de dypere vannlagene på dagtid, mens føden bestod av en blanding av fytoplankton og kopepoder grunnere enn 75 m på dagtid. Dette vitner om en omnivor fødestrategi, hvor artens fødetilgang kommer fra minst to trofiske nivå. Nyere undersøkelser støtter undersøkelsen gjort av Sameoto (1980) og gir et nyansert bilde av føden hos denne arten, også i Clyde-området (se Lass mfl. 2001).
De noe eldre observasjonene som er omtalt over viser at M. norvegica er i stand til å utnytte detritus og suspendert materiale, og har betydelig fleksibilitet i sitt fødeopptak (Lass mfl. 2001). Artens ustrakte vertikale vandringer gjennom døgnet (Mauchline og Fisher 1969, Tarling mfl. 1998), og at den er observert på store havdyp i utredningsområdet (jfr. Figur 4.16), støttes av nyere undersøkelser der M. norvegica er observert med ROV på betydelige dyp i Faroe-Shetland kanalen (Hirai og Jones 2012). Høyeste tettheter av M. norvegica ble der funnet nær bunn i dybdeområdet 400-600 m (596 ± 261 ind./m3), men ingen observasjoner ble gjort mellom 600-1500 m.
4.5.11 - Noen felles betraktninger
Det observeres at krillens vertikalfordeling (Figur 4.22) er noe annerledes enn den som er gitt for amfipoder (Figur 4.21), men krillens biomasse i nedre del av vannsøylen rundt 1500 m er på omtrent samme nivå som i ca. 500 m dyp. Imidlertid har vi ikke en direkte kobling til krillartene som utgjør biomassen oppgitt i disse figurene. Sammenlignet med biomassen for amfipoder (Figur 4.21) er krillens biomasse på dagtid i 1500 m dyp omtrent på samme nivå. De høyeste amfipodbiomassene (g DW m-3) vises for den dypeste delen av vannsøylen (ca. 1000-1500 m), men disse tallene sier ikke noe om artssammensetningen. Selv om Themisto med stor sannsynlighet er representert i dette materialet, kan ikke forekomst av andre amfipodarter utelukkes. Det kan heller ikke utelukkes sesongmessige variasjoner i antall og biomasse. Prøvematerialet og derigjennom tallmaterialet for tetthetene (g DW m-3) beregnet for krill og amfipoder på de største dypene (Figur 4.22 og 4.21) er betydelig mindre enn prøvegrunnlaget høyere oppe i vannsøylen. Derfor er det mulig at tetthetene vist for større dyp ikke er representative for disse gruppene gjennom hele sesongen, og gitt områdets heterogene karakter, kan det også være betydelige forskjeller over området på samme tidspunkt i sesongen.
I Figur 4.16 er vist en oversikt over arter/slekter av dyreplankton integrert over dybde-intervallene 0-500 m, 500-1000 m og 1000-1600 m. I de to nederste dybdeintervallene finner vi amfipoden T. abyssorum, storkrillen M. norvegica og den langt mindre T. longicaudata, som skulle bety at artene utvilsomt kan gå dypere enn 1000 m, og dypere enn der mange av de hydrotermale kildene på den midt-Atlantiske ryggen er registrert. Imidlertid er tettheten av organismer her bare ~0.01-0.03 ind./m3, som er mye lavere enn det som ble maksimalt registrert mot bunn for M. norvegica i ~480 m dyp på 596 ind./m3 (Hirai og Jones 2012). Men de lave estimerte håv-verdiene gitt over, må også sees i lys av at innsamlingsredskapet (Multinet, 0.25 m2 åpning), ikke er godt egnet for å ta prøver av mange makroplankton arter som i stor grad har evne til å unnvike redskapet grunnet hurtige reaksjonsmønstre og gode svømmeegenskaper. På stasjon M er avstanden til Arktiske overflatevannmasser og polarfronten betydelig og det er derfor naturlig at den Arktiske amfipoden Themisto libellula ikke er registrert på denne lokaliteten da kun et begrenset datamateriale har vært tilgengelig for denne rapporten.
4.5.12 - Kunnskapshull, varme havkilder og andre bunnære områder
Det er hittil bare et fåtall prøver som er tatt ned mot de hydrotermale feltene knyttet til den midt-Atlantiske fjellkjeden nord for ca. 62ºN, selv om ryggen mange steder ikke er så veldig dyp. Det samme gjelder prøvetaking ned mot bunndyp som er innenfor det som betegnes som bathypelagialen, altså dybdeområdet fra 1000-4000 m. Komplisert topografi i nærheten av den midt-Atlantiske fjellkjeden gjør det her vanskelig å benytte innsamlingsredskaper som man tradisjonelt bruker for overvåkingsformål. Å foreta representativ prøvetaking i et så potensielt heterogent habitat er derfor svært vanskelig. Kanskje må ny teknologi utvikles og tas i bruk for å kunne dokumentere en sannsynlig og betydelig flekkvis forekomst av pelagiske og hyperbenthiske organismer rundt hydrotermale kilder. Her kan både bruk av akustiske og optiske teknikker/metoder sammen med eDNA kanskje bidra til å skape et mer sannferdig bilde av type og mengde organismer som kan utgjøre en betydningsfull andel av dyreplankton biomassen i de hydrotermale nærområdene.
Det er også svært mangelfull kunnskap om hvordan amfipoder og kanskje også krill kan dra fordel av kjemoautotrof produksjon og/eller sedimentert organisk materiale i områder hvor hydrotermale kilder er en viktig del av det undersjøiske landskapet. På den annen side er det også på dypt vann utenfor disse områdene (f.eks. på kontinentalskråningene) vesentlig å forstå hva disse gruppene foretar seg på eller like over bunn. Kan deres tilstedeværelse knyttes til reproduktiv adferd, fødeopptak, overvintring, predatorunnvikelse eller ha innslag av ontogenetisk migrasjon? Både hydrotermale habitater, og andre områder hvor bunnen er innenfor rekkevidde til vertikalmigrerende amfipoder og krill, kan altså ha betydning på populasjonsnivå for disse makroplanktoniske krepsdyrene som igjen er svært viktige fødekomponenter for en rekke fiskearter, fugl og marine pattedyr. Her er det stort behov for ny kunnskap både om biologi og økosystemfunksjon.
Meganyctiphanes norvegica er kjent for å ha en tradisjonell DVM (døgnlig vertikal migrasjon), og vandrer opp mot overflaten om natten mens den beiter, noe omtrent 65% av alle individene gjennomfører, mens resten av bestanden blir værende på dypt vann (Tarling mfl. 1998). Hvorvidt dette er en observasjon som gjelder for både grunne og veldig dype lokaliteter, er usikkert. Det viser imidlertid at en betydelig andel av bestanden på en gitt lokalitet kan unnlate å vertikalvandre og heller blir værende nær bunn. Sannsynligvis er årsaken til dette at det på en gitt lokalitet kan være fordelaktig å oppholde seg for denne bestandskomponenten. Mye er fortsatt uklart mht. de vertikale migrasjonene hos Themisto-artene og M. norvegica, både gjennom døgn og over årssyklus. Det må være en prioritert oppgave å tette slike kunnskapshull om viktige dyreplanktonbestander.
4.6 - Fremtidsutsikter
Global oppvarming påvirker artssammensetning regionalt (Molinos et al. 2016, Poloczanska et al. 2016), typisk ved at sørlige former forflyttes mot høyere breddegrader. Denne spredningen er fullt merkbar allerede, og betyr at vi har en kontinuerlig forandring i både artssammensetning og diversitet i vente de neste tiårene. Mengden Calanus finmarchicus er antatt å øke noe generelt i Norskehavet i nær fremtid (Kjesbu mfl. Innsendt). I det sørlige Norskehavet derimot, lever C. finmarchicus og C. helgolandicus i utkanten av sine utbredelsesområder, og økt havtemperatur vil kunne gi redusert mengde C. finmarchicus og økt mengde C. helgolandicus, tilsvarende situasjonen i Nordsjøen (Beaugrand mfl. 2002), noe som vil påvirke produksjonen og sesongmønsteret i området. I utredningsområdet finnes både Atlantiske (“varmtvanns”) og Arktiske (“kaldtvanns”) systemer, side om side på samme breddegrad og med tilsvarende lysregimer, med strømsystemer som transporterer vann i til dels motsatte retninger, grunne områder så vel som dype bassenger med gyrer. I De nordiske hav foregår kortsiktige og langsiktige endringer i disse systemene, men sannsynligvis ikke likt over hele utredningsområdet. Hvordan dette vil påvirke biologiske enkeltkomponenter eller økosystemene vet vi lite om. Her ligger det en betydelig usikkerhet som bør legges til grunn, også ved en fremtidig konsekvensutredning og forvaltning av disse økosystemene.
4.7 - Oppsummering og kunnskapsbehov
Mikrobiologi
Utredningsområdet har vært gjenstand kun for sporadiske undersøkelser av mikrobiologiske variable, og det hovedsakelig i forbindelse med prosjekter av kort varighet omkring karbonsyklus eller fiskeriene.
Planteplankton i utredningsområdet har vært karakterisert i et fåtall studier, som også inkluderer undersøkelser av årstidsvariasjon.
Undersøkelser av virus fra utredningsområdet er ikke kjent i tilgjengelig litteratur. Bakterier har vært undersøkt i noen få publiserte studier, hovedsakelig omkring karbonsyklus i vann nær overflaten, og lokalt omkring områder påvirket av vulkanske utslipp.
Undersøkelser av årstidsvariasjon hos bakterier og virus i utredningsområdet er ikke kjent, og kunnskap om heterotrofe mikrobielle samfunn gjennom hele vannsøylen mangler.
Det er påvist at i sprekkområdene langs den Midtatlantiske ryggen finnes store forekomster av spesielle mikrobiologiske samfunn som er knyttet til omsetning av stofflekkasjer fra jordens indre når disse frigjøres i sjøvann. Man vet for lite om disse mikrobesamfunnene og mikrobiologisk omsetning av kjemikalier som frigjøres til sjøvann fra den vulkanske aktiviteten langs den Midtatlantiske ryggen.
Dyreplankton
Planteplankton omdanner uorganisk karbon i havet til biologisk bundet organisk karbon ved fotosyntese. Herbivort dyreplankton beiter på planteplanktonet, bygger opp sin egen biomasse, og blir selv spist av karnivort dyreplankton, fisk, sjøfugl og hval. Slik flyter energien fra primærprodusenter til dyr på høyere trofiske nivå i næringsnettet. Produksjonen på lavere trofiske nivå er høy, og representerer viktig føde for store pelagiske fiskebestander, samt sjøfugl og sjøpattedyr.
Sesongsyklusen i lys- og fysiske forhold i De nordiske hav fører til kraftig sesongdynamikk hos planktoniske primær- og sekundærprodusenter. Produksjonen er høyest i vår-sommer perioden, og fraværende eller lav om vinteren. Dette reflekteres videre i tidspunkt for viktige livsbegivenheter og sesongutvikling hos mange av de øvrige organismene i økosystemet.
Mesodyreplanktonet i Norskehavet er representert ved en rekke forskjellige grupper og arter, hvor kopepodene dominerer både antall og biomasse. Vurdert ut fra biomasse og økologisk rolle er Calanus finmarchicus (raudåte) den viktigste kopepodarten i Norskehavet og omkringliggende områder.
Unge individer av C. finmarchicus er hovedføden for larvene til en rekke fiskebestander av kommersiell betydning i De nordiske hav, mens eldre individer representerer den viktigste føden for planktivore fiskebestander som sild og makrell.
C. finmarchicus utfører sesongmessige vertikale migrasjoner. Mens den produktive vår- og sommerperioden tilbringes i øvre vannlag, overvintrer den i en dvalelignende tilstand på store dyp i Norskehavet.
Det er behov for bedre kunnskap om planktonet knyttet til den Arktiske fronten over den Midtatlantiske ryggen; det gjelder bl.a. variasjoner i mengde og produksjon over tid, artssammensetning og vertikalfordeling.
Vertikalvandring og utsynking av plankton og nekton, samt deres fekalier, er viktig for vertikal transport av karbon, og som fødegrunnlag for den betydelige biomassen av mesopelagiske organismer.
Spesielt viktige makroplanktongrupper er krill og amfipoder.
Fødeopptak i dyreplanktonet er ikke begrenset til epipelagialen, men finner også sted på dypt vann. Planktonet kan dermed utnytte både den øvre og dypere delen av vannsøylen, inklusive områder med hydrotermale kilder.
Det er store hull i kunnskapen om biologi, energiflyt og trofiske interaksjoner i dyphavet. Det er i tillegg et stort behov for bedre forståelse av bentisk-pelagiske koblinger generelt, og spesielt i områdene på og over den Midtatlantiske ryggen.
Det er «underskudd» på kvantitative data om artsmengder, biomasse og fordeling av makroplankton i utredningsområdet, spesielt gjelder dette dybdestratifierte data fra meso- og bathypelagisk sone.
5 - Fisk
5.1 - Innledning
Fisk i Norskehavet handler først og fremst om de store, vandrende bestandene av pelagisk fisk, som makrell, sild og kolmule. Langs kontinentalskråningene ned mot de dype bassengene i Norskehavet lever også mer bunntilknyttede arter, som blåkveite, lange og brosme. Nede på bunnen i de dype bassengene, og på de undersjøiske ryggene hvor det for det meste er dypere enn 1000 m, lever trolig ingen arter som utgjør noen særlig stor biomasse, og i alle fall ingen som har noen kommersiell verdi i tradisjonelt fiskeri. Når vi skriver trolig, er det fordi både artssammensetning og mengder er svært dårlig kartlagt. Den vanligste måten å få kunnskap om artssammensetning på bunnen er ved hjelp av trål, og på så dypt vann er det vanskelig å tråle på bunnen. Det er derfor en kjensgjerning at mens vi har svært god kunnskap om hva som lever i de øverste par hundre meterne av Norskehavet, og hva som lever på skråningene rundt dette havet ned til ca. 1000 meter, er med få unntak de dypere delene og i særdeleshet bunnen av havet foreløpig et “Mare incognitum”.
Vi deler gjerne de frie vannmassene i et hav inn etter dypet, hvor de øvre 200 meterne kalles epipelagialen, området fra 200 til 1000 m kalles mesopelagialen, og de dypere delene bathypelagialen. Epipelagialen er der hvor primærproduksjonen, dvs. karbonproduksjonen basert på sollyset, foregår og omsettes i et næringsnett som består av dyreplankton som spiser planteplankton, pelagisk fisk som spiser dyreplankton, og store rovfisker og sjøpattedyr som spiser pelagisk fisk. Dypere nede, i mesopelagialen, er det også en relativt rikholdig sammensetning av organismer, som består av mesopelagisk fisk, krepsdyr, bløtdyr og nesledyr. Mange av disse mesopelagiske organismene vandrer opp i epipelagialen om natten for å beite, og ned igjen til mesopelagialen om dagen. Dette er en tilpasning som gir dem tilgang på mat i de øvre vannlagene, men samtidig skjermer dem fra å bli spist av predatorer som benytter synet ved jakt. Mesopelagiske organismer som ikke døgnvandrer på denne måten er avhengig av mat som “drysser ned” fra de øvre vannlag. Enda dypere nede, i bathypelagialen, er det langt mindre liv, siden tilgangen på føde er begrenset til det som måtte dale ned fra oven. Nede på bunnen er det igjen et noe rikere liv, for all biomasse som drysser ned fra produksjonslagene øverst i vannmassene og som ikke blir spist på veg ned i dypet, vil akkumuleres på bunnen, og her lever noen bunntilknyttede fiskearter, krepsdyr og pigghuder som kan nyttiggjøre seg disse næringsstoffene. Når det en sjelden gang dumper et hvalkadaver eller fiskekadaver ned på bunnen vil dette i lang tid gi opphav til et eget lite samfunn av åtseletere som lever av denne overfloden av mat så lenge den varer. Der det er vulkansk aktivitet på havbunnen, og hvor varmt vann rikt på kjemiske stoffer kommer opp fra havbunnen, dannes det særegne økosystemer hvor primærproduksjonen ikke er basert på fotosyntese, men på kjemosyntese. Disse økosystemene har mange organismer som ellers er ukjente, men trekker trolig til seg også fisk og andre bunnlevende organismer som finner næring rundt disse vulkanene.
Organismene som lever i de frie vannmassene, deler vi gjerne inn i gruppene plankton og nekton. Mens plankton er definert som dyr som har sin utbredelse styrt av horisontale strømmer, er nekton dyr med nok egenbevegelse til å selv styre sin utbredelse. Nekton større enn ca. 20 cm består hovedsakelig av fisk, som blir omtalt i dette kapitlet. Men særlig i mesopelagialen er det mange organismer som vi kan kalle makroplankton og mikronekton, med størrelse fra ca. 2 til 20 cm. I praksis er det små forskjeller mellom disse to gruppene, så vi slår sammen og definerer som makroplankton alle organismer over ~2 cm som ikke har store horisontale vandringer. Denne gruppen blir omtalt i planktonkapittel Makrodyreplankton og mikronekton. Gruppen inneholder både fisk og evertebrater, viktige grupper av makroplankton i utredningsområdet er amfipoder, krill, pelagiske reker, maneter (inkludert ribbemaneter), pilormer og mesopelagisk fisk. I tillegg til makroplankton er pelagiske blekksprut antatt å være en viktig gruppe i området (Skjoldal mfl. 2004), men her mangler vi i stor grad underlagsmateriale for å si noe om hvor viktig denne gruppen er: vi har ikke utstyr til å måle denne gruppen kvantitativt. Også denne gruppen er videre omtalt i kapitlet om makroplankton. Når det gjelder de bunntilknyttede ressursene på dyphavet dekkes disse av en rapport som utarbeides av Universitetet i Bergen, og vi vil derfor ikke omtale disse i denne rapporten.
5.2 - Pelagisk fisk i de øvre delene av pelagialen
Det pelagiske fiskesamfunnet i Norskehavet er dominert av store bestander av artene atlantisk sild (Clupea harengus), atlantisk makrell (Scomber scombrus) og kolmule (Micromesistius poutassou). I tillegg finnes andre pelagiske arter som lodde (Mallotus villosus), atlantisk laks (Salmo salar), rognkjeks (Cyclopterus lumpus), og makrellstørje (Thunnus thynnus). Norskehavet er først og fremst et viktig beiteområde for disse artene som vandrer inn i Norskehavet for å beite på blant annet dyreplankton om våren og sommeren. Det er sannsynligvis et komplekst sett av faktorer som påvirker vandringsmønsteret som varierer over tid, men det er funnet at størrelsen på bestanden og alderssammensetningen er viktige faktorer, spesielt hos norsk vårgytende (nvg) sild. Stor (gammel) fisk har et større vandringspotensiale enn mindre (yngre) fisk og vil ofte vandre lengre vestover i beiteperioden. I tillegg vil miljøfaktorer som temperaturforhold og mattilbud være viktig. Gytefeltene til de pelagiske bestandene finnes ofte i tilstøtende hav og kystområder. De store pelagiske bestandene i Norskehavet er svært tallrike og viktige kommersielle bestander med stor betydning for fiskeriene, og de blir utsatt for varierende grad av fiskepress. Derfor har det over lang tid blitt brukt betydelige ressurser på å overvåke tilstanden til disse bestandene og kunnskapsgrunnlaget er derfor godt og oppdatert. Man har lange tidsserier med data både fra fiskerier og fra forskningstokt. Data fra fiskeriene strekker seg tilbake til tidlig 1900-tallet (nvg-sild) mens datainnsamling av pelagisk fisk fra forskningstokt i Norskehavet startet på 1980-tallet. I beiteperioden i Norskehavet blir det årlig utført to internasjonale tokt for å overvåke de pelagiske bestandene; ett tokt i mai og ett i juli/august. Kunnskapsgrunnlaget er derfor godt i denne perioden. Selv om forskningstoktene ofte har de kommersielt viktigste artene som målarter for toktene blir også annen pelagisk fisk registrert slik at data fra arter som laks og rognkjeks også samles inn.
Overvåkningen inkluderer også innsamling av mager til diettanalyser. Resultater her har vist at de pelagiske artene har betydelig evne til å tilpasse seg ulike miljøforhold og matkilder. Det er stor grad av overlapp i diett mellom arter som makrell og nvg-sild (Bachiller mfl. 2016). Føden til disse er dominert av hoppekreps (kopepoder) og spesielt raudåte (Calanus finmarchicus) er viktig, men også krill og larvesekkedyr (appendicularier) er viktige byttedyr i noen år/årstider. Kopepoder som raudåte og feitåte (C. hyperboreus) er også viktige byttedyr for lodde (Astthorsson og Gislason 1997), mens kolmuledietten er dominert av større byttedyr som krill og amfipoder og det er lite overlapp mellom dietten til kolmule og dietten til sild/makrell (Bachiller mfl. 2016). Hovedføden til rognkjeks er ulike arter av maneter (Bjelland og Holst 2004) mens laksen spiser et vidt spekter av arter inkludert krill, amfipoder, blekksprut og ulike fiskearter (Holm mfl. 2004). Makrellstørjen som vandrer inn i Norskehavet er ofte stor og spiser hovedsakelig fisk, men også blekksprut og noen krepsdyr som krill og reker (Olafsdottir mfl. 2016). Nedenfor følger en kort oppsummering av kunnskap om de viktigste artene/bestandene av pelagisk fisk i Norskehavet.
5.2.1 - Norske vårgytende sild
Det er en kompleks bestandsstruktur av sild i Norskehavet, men nvg-sild er den desidert mest tallrike og kommersielt viktigste sildebestanden med et fiske på i gjennomsnitt ca. 900 000 tonn de siste 15 år (Figur 5.1). Den umodne del av bestanden har oppvekstområder i Barentshavet og vil derfor stort sett være fordelt utenfor utredningsområdet. Den modne silda vil etter gyting på kystbanker nær Norskekysten i februar/mars vandre vestover og inn i Norskehavet. I perioden etter gyting og fram til høsten fordeler nvg-silda seg utover store deler av Norskehavet inkludert havområdene som ligger innenfor utredningsområdet. Om høsten avtar beitingen og nvg-silda vandrer i hovedsak østover for å overvintre i norske kyst- og havområder. Nvg-sild vil derfor i større eller mindre grad befinne seg innenfor utredningsområdet i perioden mars – desember. Toktresultater viser betydelig mellomårlig variasjon både i horisontal- og vertikalfordeling. Silda fordeler seg stort sett i øvre vannlag, men i enkelte år kan silda i mai beite i kalde vannmasser på dyp ned mot 400 m, spesielt i vestlige deler av Norskehavet.
Figur 5.1 Beiteområde, gyteområde og oppvekstområde til norsk vårgytende sild.
5.2.2 - Makrell
Makrell er også en kommersielt svært viktig art i Norskehavet med et årlig fiske på i overkant av 800 000 tonn de siste 15 år (Figur 5.2). Makrellbestanden gyter fra vest av de Britiske øyer og sørover til Biscayabukten i perioden februar-juli (tidligst i sør). Hovedgytingen er i april/mai. Etter gyting vandrer store deler av makrellen inn til beiteområdene i Norskehavet der den blir fram til august/september da den vandrer sørover der den oppholder seg fram til gytevandringen starter. Mye av fiskeriet skjer i beiteperioden i Norskehavet. Makrell-bestanden vil i beiteperioden befinne seg i stor grad innenfor utredningsområdet. Siden makrell er en varmekjær fisk vil den i motsetning til sild i svært liten grad vandre inn i kalde vannmasser nord for Island, selv om sommeren. Det har imidlertid vært en tendens til økt nordlig utbredelse av makrellen og den registreres regelmessig nord til sørspissen av Spitsbergen i juli. I noen år har også makrellen vandret vest til Grønlandske farvann i de relativt varme vannmassene i vest og sør for Island. Makrellen er stort sett knyttet til de varme vannmassene i øverste 50 m i beiteperioden i Norskehavet.
Figur 5.2 Gyteområde og oppvekstområde til makrell.
5.2.3 - Laks
Atlantisk laks er en anadrom fiskeart som gyter i elver, men som bruker blant annet Norskehavet som beiteområdet i den oseaniske fasen av livet som varer 1-4 år og der mesteparten av fiskens individvekst skjer (Figur 5.3). Laksen er oppdelt i enkeltbestander og man regner med at det er en eller flere bestander i hvert vassdrag. Laks som overlever gyting og har flere beiteperioder i havet kan bli svært store, opptil 40 kg. Men, i motsetning til makrell og sild er laksen lite tallrik i Norskehavet. Laks fiskes både i havet og når den er på gytevandring i elvene. Fisket er imidlertid relativt lavt og har hatt en nedadgående trend siden 1970-tallet, med et årlig uttak på ca. 1000 tonn de siste årene. Systematisk innsamling av laks i pelagiske forskningstokt i Norskehavet startet på 1990-tallet og tidsseriene her er derfor kortere enn for de store pelagiske bestandene. Laks har i forskningstoktene stort sett blitt fanget i de øverste vannlagene, noe som indikerer at laksen stort sett beiter nær overflaten i Norskehavet.
Figur 5.3 Utbredelsesområde til Atlantisk laks.
5.2.4 - Rognkjeks
Rognkjeks finnes over store deler av nordøstlige Atlanterhavet, inkludert Norskehavet (Figur 5.4). Den gyter langs kysten om våren, men er i beiteperioden fordelt pelagisk i de øverste 50-60 m over store deler av Norskehavet som også er oppvekstområder for rognkjeksen. Den tåler lave temperaturer godt. Rognkjeks er den arten som fanges hyppigst i trålen på forskningstoktene (spesielt i toktet i juli/august) og kunnskapen om fordeling i Norskehavet i beiteperioden er derfor godt. Men kunnskapen om bestandsstruktur og vandringsmønster mellom gyte- og vekstområdene er svært begrenset. Rognkjeksen vil i beiteperioden befinne seg i stor grad innenfor utredningsområdet. Det er et begrenset fiskeri av kjønnsmoden rognkjeks for rogn, om våren langs kysten i nord. Det norske fisket har variert mellom 14 og 690 tonn rogn de siste 15 år.
Figur 5.4 Gyteområde og oppvekstområde til rognkjeks
5.2.5 - Makrellstørje
Det er to bestandskomponenter av makrellstørje i det nordlige Atlanterhavet (Figur 5.5). Den vestlige makrellstørjen gyter i området ved Mexico-gulfen. Den østlige makrellstørjen gyter om våren i mai–juni, i Middelhavet. Det er hovedsakelig den østlige makrellstørjen som vandrer inn og beiter i Norskehavet om sommeren/høsten. Makrellstørje kan bli opp mot 750 kg og er en svært ettertraktet matfisk. Makrellstørje fanges ikke opp i de ordinære forskningstoktene fordi den er for raskt svømmende til å fanges i pelagiske tråler. Dokumentasjon av utbredelse kommer derfor fra fangster, visuelle observasjoner og i senere tid fra satellittmerking. Disse viser en økt tilstedeværelse av makrellstørje i Norskehavet og langs norskekysten i senere tid, og den norske fangsten har siden gjenåpningen av et ordinært fiske i 2017, økt til 315 tonn i 2021. Data fra satellittmerket makrellstørje viser at den ikke bare er knyttet til overflaten, og dykk ned til 500 m er registrert.
Figur 5.5 Gyteområde og oppvekstområde til makrellstørje
5.2.6 - Lodde
I havområdene mellom Island, Grønland og Jan Mayen finnes en bestand av lodde (Figur 5.6). Denne bestanden gyter langs sør- og vestkysten av Island i februar-mars, og beiter nord for Island om sommeren, inkludert i varierende grad innenfor utredningsområdet vest og sørvest av Jan Mayen. Fisket på denne bestanden har variert fra 0 til ca. 400 000 tonn siste 15 år med et snitt på ca. 275 000 tonn. Bestanden overvåkes årlig med to tokt av det islandske havforskningsinstituttet; ett tokt om høsten og ett like før gyting om vinteren.
Figur 5.6 Beiteområde, gyteområde og oppvekstområde til lodde.
5.3 - Pelagisk fisk dypere nede i pelagialen
5.3.1 - Kolmule
Kolmule er en svært tallrik art i Norskehavet (Figur 5.7) og årlig fangst har i gjennomsnitt vært i underkant av 1,2 millioner tonn de siste 15 år. Kolmule gyter hovedsakelig vest for de Britiske øyer om våren (hovedsakelig mars-april), der også det viktigste fiskeriet foregår. Den voksne delen av bestanden vandrer deretter inn i Norskehavet der den beiter om sommeren og utover høsten før den på nytt starter vandringen mot gyteområdene. Kolmule er noe mer varmekjær enn for eksempel sild, men om sommeren er den fordelt over store deler av Norskehavet inkludert deler av utredningsområdet, men sjelden i det kalde vannet vest for polarfronten. Voksen kolmule finnes ofte i dypet mellom 200 og 500 m, men spesielt ung kolmule kan finnes høyere oppe i vannmassene. Voksen kolmule kan også finnes høyt oppe i vannmassene, spesielt i vestlige deler av Norskehavet der det er kalde vannmasser i dypere vannlag. Umoden kolmule kan være fordelt over store områder i Nordsjøen, Norskehavet og Barentshavet, men de viktigste oppvekstområdene er sørlige og østlige Norskehavet.
Figur 5.7 Utbredelsesområde, ungfisk og gyteområde til kolmule
5.4 - Fisk knyttet til kontinentalskråningen rundt Norskehavsbassenget
5.4.1 - Innledning
Kontinentalskråningen i Norskebassenget er påvirket av en uttalt hydrografisk front mellom varme atlantiske vannmasser og kalde vannmasser i Norskehavet. Hallfreðsson og Primicerio (2017) identifiserte totalt 78 arter/taxa langs kontinentalskråningen og Bjørnøyrenna, hvilket er lavt sammenlignet med de 296 artene observert langs kontinentalsokkelen i det nordatlantiske bassenget (Haedrich og Merrett 1988). De fleste artene vi finner tilhører typiske arktiske grupper i stedet for dypvannsfiskfamiliene som vi finner andre steder. Bergstad mfl. (1999) registrerte 34 arter fordelt på ulike dyp. De grunneste stasjonene (<600 meter) hadde størst diversitet, og var dominert av snabeluer (Sebastes mentella), lusuer (Sebastes viviparus), blåkveite (Reinhardtius hippoglossoides) og kolmule (Micromesistius poutassou). Mellomdype stasjoner (ca. 500-1000 m) hadde lavere biodiversitet og tetthet og var dominert av blåkveite og isgalt (Macrourus berglax). På de dypeste stasjonene (1500-2000 m) var både diversitet og tettheter veldig lave. Kun 7 arter ble observert, sterkt dominert av arktisk ålebrosme (Lycodes frigidus) og svart ringbuk (Paraliparis bathybius). I tillegg til artene nevnt er det observert store arter i torskefamilien som brosme (Brosme brosme), lange (Molva molva) og blålange ((Molva dipterygia), vassild (Argentina silus) og en rekke bruskfiskarter med mer eller mindre kjent utbredelse og populasjonsstørrelse.
Fisk som lever langs kontinentalskråningen finner føde både på bunnen og høyere opp i vannsøylen. Føde lengre opp i vannsøylen blir tilgjengelig gjennom fødevandringer, adveksjon eller synking (Bjelland mfl. 2004). De dominerende artene har en variert diett med bytte fra flere dyregrupper. Blåkveite, og de store torskefiskene brosme og lange, har potensielt en viktig økologisk rolle langs skråningen som predator på hvitting, sild og lodde (Michalsen og Nedreaas 1998, Bjelland mfl. 2000). Blåkveite tar i tillegg en del reker og blekksprut. Dietten til vassild består av maneter, mindre fisk, krepsdyr og blekksprut. Uerartene lever hovedsakelig av dyreplankton de første leveårene, men går så gradvis over til en diett som domineres av fisk, herunder lodde, sild, små torskefisk og store dyreplankton. Som byttedyr er juvenile uer viktig føde for torskefisk og kveite. I tidligere år, da rekrutteringa av snabelueryngel var god og stabil, utgjorde snabeluer under 25 cm rundt 10 prosent av dietten til nordøstarktisk torsk. Larver og små yngel av uer har dessuten blitt observert i sildemager. Blåkveite er funnet i magen til håkjerring, torsk og annen blåkveite. Sjøpattedyr kan være viktige predatorer, men det er lite som tyder på at blåkveite er utsatt for høyt beitepress.
Mange av artene som lever langs kontinentalskråningen når en høy levealder. Brosme blir omtrent 20 år mens blåkveite, lange og blålange blir over 30 år. Vassild og isgalt kan bli 40 år mens vanlig uer og snabeluer kan bli over 60 år. Den lengstlevende arten er likevel håkjerring som kan bli over 270 år (Nielsen mfl. 2016). Langtlevende arter deler ofte flere utviklingstrekk som for eksempel langsom vekst, sen modning og lav fruktbarhet. Slike bestander vil være ekstra utsatt for forstyrrelser fordi det tar lengre tid for populasjonene å gjenopprettes etter en forstyrrelse (Pianka 1970). De fleste av disse artene lever på store dyp der lysmengden er svært begrenset. Mange av artene er derfor avhengige av lyd for grunnleggende livsfunksjoner som for eksempel kommunikasjon, navigasjon og fødesøk. Lange, blålange og brosme har alle lydproduserende strukturer som brukes i forbindelse med reproduksjon (Hawkins og Picciulin 2019).
Havforskningsinstituttet har tre toktserier som dekker disse artenes utbredelse langs kontinental skråningen (Eggakanten) og i Norskehavet (Figur 5.8). EggaSør toktet går om våren (mars-april) og dekker Eggakanten fra 62-75⁰N. Toktet er spesielt viktig for vassild, fordi det samler inn de eneste fiskeri-uavhengige dataene for arten. Toktet dekker også de viktigste gyteområdene for snabeluer, samt deler av utbredelsesområdet for vanlig uer og voksen blåkveite. EggaNord toktet går om høsten (august-oktober) og dekker Eggakanten i området fra 68-80oN og er hovedtoktet for voksen blåkveite. I tillegg dekker toktet deler av utbredelsen til vanlig uer. Snabeluer dekkes også selv om bare en liten del av bestanden regnes å være innfor toktet dekningsområde. Pelagisk dyphavstokt i Norskehavet går sommer/høst (august) og dekker nordlige del av Norskehavet i området fra 70-77⁰N og østover mot Eggakanten. Toktets målart er voksen snabeluer i Norskehavet. Eggatoktene går annethvert år (alternerende), mens dyphavstoktet går hvert tredje år. I tillegg foregår en innsamling av prøver fra fiskeriene som gir informasjon om lengdefordelinger, begrenset info om utbredelse, kjønnsfordelinger osv. for en rekke arter (Figur 5.8). Flere av disse bestandene har også utbredelse i Barentshavet som dekkes av det årlige økosystemtoktet.
Bruskfisk er av liten kommersiell betydning og kunnskapsgrunnlaget er begrenset ettersom det brukes lite ressurser på å overvåke tilstanden til disse bestandene. Bestandsgrenser er ikke kjent for artene i dette området. Det samme gjelder deres bevegelser mellom kystområder og offshoreområder. Ytterligere undersøkelser er nødvendige for å bestemme potensielle vandringer eller interaksjoner mellom bruskfiskpopulasjoner til og fra tilstøtende områder. Mangel på informasjon om demografi og populasjonstrender gjør det vanskelig å vurdere sårbarhet for menneskelig aktivitet. Nedenfor følger en kort oppsummering av kunnskap om de viktigste artene/bestandene av fisk tilknyttet kontinentalskråningen i Norskehavet. Følgende og ytterligere informasjon er tilgjengelige på https://www.hi.no/hi/temasider/arter/.
Figur 5.8 Stasjoner som inngår i forskningstokt for området (ikke alle stasjoner brukes hvert år): EggaNor (øverst til venstre), EggaSør (øverst til høyre) og Dyphavstoktet (nederst til venstre), samt Prøver fra 2020 (Referanseflåten, Kommersielle prøver, Fiskerisampling, Kystvakt, Møreforskning og Prøvebåt) (nederst til høyre).
5.4.2 - Blåkveite
Kjønnsmoden blåkveite finnes hovedsakelig langs Eggakanten mellom fastlands-Norge og Svalbard, med en mer begrenset forekomst i Barentshavet (Figur 5.9). Juvenile fisk finnes hovedsakelig nord og øst for Svalbard til Kvitøya og Frans Josefs land der bestanden har sine oppvekstområder. Blåkveite er en bunnlevende art som hovedsakelig finnes på dypt vann, men foretar pelagiske fødevandringer. Blåkveita gyter langs Eggakanten mellom 70 og 75°N. Gytingen foregår om høsten/vinteren med en topp i desember, på dypt vann (500–800 m) (Albert mfl. 2001). Egg og larver driver med strømmen avhengig av hvor gytingen foregår (Ådlandsvik mfl. 2004). Blåkveite er en kommersiell art i Norskehavet med et årlig fiske på rundt 25 000 tonn i året de siste årene. Mesteparten av fangsten av tas langs Eggakanten, hovedsakelig av Norge og Russland. Den fangstbare mengden blåkveite, definert som fisk som er 45 cm eller lenger, økte etter at strengere regulering bli innført i 2009 og frem til 2012, men viser nedadgående trend etter 2014. Blåkveitas videre bestandsutvikling avhenger blant annet av rekrutteringen som er variabel, med topper i enkelte år. Siste registrerte gode årsklasse er fra 2010. En god årsklasse vil ha stor effekt på blåkveitas bestandsutvikling.
Figur 5.9 Utbredelsesområde og gyteområde til blåkveite.
5.4.3 - Snabeluer
Snabelueren lever i sentrale deler av Norskehavet og langs kontinentalskråningen mot Norskehavet, fra 400 til 600 meters dyp fra Shetland og nordover til Andøya (Figur 5.10). Her finnes det lite snabeluer mindre enn 28–30 cm. Nord for Andøya finnes snabeluer også grunnere. Barentshavet og Svalbard (også nord for Spitsbergen) er oppvekstområdet for arten. Snabelueren beiter i sentrale deler av Norskehavet, men yngler langs Eggakanten, fra Shetland til Tromsøflaket. I Barentshavet er det påvist gytevandring av hunnfisk mot dette området. Snabeluer føder levende 4–6 mm yngel i mars–april. Snabeluer har vært gjenstand for et målrettet pelagisk fiske i Norskehavet utenfor norsk økonomisk sone siden 2004. Direkte fiske har vært tillatt innenfor norsk økonomisk sone siden 2014, med bunntrål så vel som med pelagisk trål. De norske og russiske fangstene (54 000 tonn i 2020) er hovedsakelig fra norsk økonomisk sone eller tas som bifangst i Svalbard-sonen. Siden de forskjellige artene av uer er vanskelig å skille fra hverandre, spesielt når de er unge, gjelder alle bifangstbestemmelser for både snabeluer og vanlig uer.
Figur 5.10 utbredelsesområde og yngleområder til snabeluer.
5.4.4 - Vanlig uer
Vanlig uer lever på 100–500 meters dyp på Eggakanten, langs kysten og enkelte steder inne i fjordene (Figur 5.11). Utbredelsen trekker seg til nordvest for Spitsbergen, men den finnes sjelden i fiskbare mengder nord for Tromsøflaket/Bjørnøya. Yngleområdet strekker seg langs Eggakanten og kontinentalsokkelen fra Shetland og nordover til Andøya, med Storegga, Haltenbanken og Vesterålen som de viktigste områdene. Vanlig uer føder levende yngel i april–mai. Paringa foregår om høsten, og i yngleområdet om våren kan det derfor være rene hunnfisk-konsentrasjoner. Vanlig uer er registrert som truet (EN) på norsk rødliste og direkte fiske er forbudt, med unntak av fiske med håndline fra båter under 15 m i lengde. Disse fiskeriene utgjør 1% av den totale fangsten. Reglene for bifangst er de samme som for snabeluer på grunn av den morfologiske likheten mellom de to artene. Til tross for råd om nullfangst har fangstene de siste årene ligget på over 5000 tonn.
Figur 5.11 utbredelsesområde og yngleområder til vanlig uer
5.4.5 - Brosme
Brosme finnes hovedsakelig på steinbunn på kontinentalsokkelen og -skråningen fra 100 til 1000 meter (Figur 5.12). Voksne individer lever i relativt dype områder, men ungfisk kan påtreffes ganske grunt. Arten gyter langs kysten av Sør- og Midt-Norge, samt sør- og sørvest for Færøyene og Island. Gytingen foregår på 100-400 meters dyp i perioden april-juni. Brosme er primært bifangst i lange- og torskefisket. Fangstene tas langs Eggakanten og i kystnære områder, i tillegg til mindre mengder i Barentshavet. Det landes i overkant av 10 000 tonn brosme, hovedsakelig av Norge. Fisket etter brosme er demersalt.
Undervannsbilder av brosme, lange og blålange. Foto: Erling Svensen og Maeano
Figur 5.12 Utbredelsesområde og gyteområde til brosme.
5.4.6 - Lange
Lange finnes primært på hardbunn eller sandbunn med store steiner i dype områder på kontinentalsokkelen (Figur 5.13). Den finnes vanligvis på 300–400 meters dyp, men kan påtreffes mellom 60 og 1000 meter. Ungfiskene er utbredt i relativt grunne, kystnære områder og på bankene, inkludert den nordlige delen av Nordsjøen. I våre farvann foregår gytingen i april–juni, på 100–300 meters dyp. Den har trolig en alders- eller størrelsesavhengig utvandring til dypere områder og til gyteområdene i Nordsjøen, på Storegga, ved Færøyene, bankene vest for De britiske øyer og sørvest for Island. Fisket etter lange er demersalt. Det fanges rundt 11 000 tonn i ICES statistikkområde 1 og 2, i all hovedsak av Norge. Fangstene av lange tas både i et direkte fiskeri og som bifangst i andre fiskerier. Fangstene tas langs Norskekysten, langs Eggakanten og rundt Shetland.
Figur 5.13 Utbredelsesområde og gyteområde til lange.
5.4.7 - Blålange
Blålange er utbredt fra Marokko til Island, i Nordsjøen og Skagerrak, og i det sørvestlige Barentshavet (Figur 5.14). Den er mest tallrik i varme, dype sokkelområder, på Eggakanten og i fjordene. Blålangen er vanligst på 350–500 meters dyp, men kan finnes mellom 200–1500 meter. Kjente hovedgyteområder er Reykjanesryggen sør av Island, ved Færøyene, vest for Hebridene og langs Storegga, men tallrikheten i disse områdene er usikker. Gytedyp og -tid varierer mellom gyteområdene, og skjer på 500–2000 meters dyp (februar–april) ved Skottland og Færøyene, på 500–1000 meters dyp ved Island (midten av mars–midten av april), mens langs Storegga skjer gytingen fra 350–700 meters dyp (mai–juni). Gyting er dessuten observert ved Øst-Grønland sent på sommeren. Modnende blålange er også observert i Skagerrak. Det er ikke kjent om forekomsten av blålange i de norske fjordene er egne bestander med lokale gyteområder. Til forskjell fra lange og brosme opptrer blålange spesielt konsentrert i gyteperioden. Siden 2006 har blålange stått på den norske rødlisten som truet (EN). For å sikre gjenoppbygging av bestanden i norske områder, ble det i 2009 innført forbud mot direkte fiske og en bifangstgrense på 10 % innblanding av blålange.
Figur 5.14 Utbredelsesområde og gyteområde til blålange.
5.4.8 - Vassild
I norske farvann er vassild utbredt i dype sokkelområder og fjorder fra Skagerrak og Norskerenna til sør og vest i Barentshavet . Særlig store konsentrasjoner finnes på Eggakanten og i dyp på sokkelen utenfor Midt-Norge. Vassilda gyter i dypere deler av utbredelsesområdet, primært på Eggakanten. Gyting skjer hovedsakelig om våren, men gyting er påvist over en lang periode. Ungfisken har vid utbredelse i grunnere områder, 100-200 m, i Nordsjøen kan de forveksles med strømsild. Målrettede fiskerier etter vassild foregår hovedsakelig i sokkeldyp fra Stad til Vestfjorden. I senere år er betydelig økt kvantum av vassild tatt som bifangst i Norskerenna i Nordsjøen. De siste årene har totalfangst på denne bestanden ligget mellom 18 000 og 23 000 tonn.
5.4.9 - Isgalt
Isgalt har utbredelse på begge sider av Nord Atlanteren; i norske farvann er den funnet langs Eggakanten fra 65°N til Spitsbergen på dyp rundt 700 m. Tidligere norske og russiske studier har definert området utenfor Nord Norge som gyteområde; gytetidspunkt er hovedsakelig om vinteren. Isgalt fanges kun som bifangst i det norske utbredelsesområdet og da særlig i fiskeriet etter blåkveite med bunntrål og line. Det er ingen reguleringer for isgalt i dette området.
5.4.10 - Bruskfisk
De fleste bruskfisk befinner seg svært høyt i næringsnettet og spiller derfor en viktig rolle i marine økosystemer. Spesielt haier spiller en viktig rolle i å opprettholde balansen i økosystemer ved å regulere byttedyrpopulasjoner. Forekomsten av bruskfisk-arter i det arktiske hav og omkringliggende havområder ble gjennomgått av Lynghammar et al. (2013) og Williams (2007). 30 arter av bruskfisk ble rapportert i Norskehavet, herav 6 arter kun som enkeltobservasjoner. Dette antallet er høyt i forhold til mange andre arktiske havområder. Den høye artsrikheten skyldes antageligvis innflytelsen av vannmasser fra Atlanterhavet. Kystnære strøk domineres av kloskate (Amblyraja radiata) (Figur 5.15), men piggskate (Raja clava), hvitskate (Rajella lintea), sandskate (Leucoraja circularis), nebbskate (Leucoraja fullonica), rundskate (Rajella fyllae), isskate (Amblyraja hyperborea) og gråskate (Bathyraja spinicauda) er også vanlige. Spisskate (Dipturus oxyrinchus) finner man frem for alt langs de sørlige delene av den norske kystlinjen, syd for 65oN. I dypere områder av Norskehavet er kloskate og isskate de mest tallrike skateartene. Gråskate og rundskate forekommer også regelmessig, særlig nord for 70°N (Skjaeraasen og Bergstad, 2001; ICES, 2018). Vanlige haiarter i Norskehavet inkluderer pigghå (Squalus acanthias), svarthå (Etmopterus spinax), håbrann (Lamna nasus), brugde (Cetorhinus maximus), håkjerring (Somniosus microcephalus), hågjel (Galeus melastomus) og småflekket rødhai (Scyliorhinus canicula). I tillegg finnes en art av helhodefisker, havmus (Chimaera monstrosa) (Figur 5.15).
De fleste bruskfisk har lav produktivitet sammenlignet med benfisk, hvilket gjør dem mer sårbare for ytre påvirkninger. Et typisk bruskfisk-fiskeri er sjelden bærekraftig over tid, og det er mange eksempler på fiskeri-indusert kollaps for både skater og haier (McPhie og Campana 2009, Campana mfl. 2008, Fordham mfl. 2006). Selv om ingen fiskerier i Norskehavet retter seg mot bruskfisk ender de ofte opp som bifangst i andre fiskerier. Rundt 50% av global registrert fangst av bruskfisk tas som bifangst. I tillegg kommer utkast av individer med liten kommersiell verdi. Det finnes lite informasjon om hvor mye som kastes ut, men det er anslått at omtrent halvparten av fangsten av bruskfisk, på verdensbasis, ender opp som utkast (Stevens mfl. 2000). I Norskehavet er bifangstproblematikken størst for store individer av spisskate, gråskate og piggskate. Håbrann er listet som sårbar (VU), brugde og pigghå er listet som Sterk truet (EN) og storskate er listet som kritisk truet (CR).
Som nevnt innledningsvis er kunnskapen om hvilke fisker som lever i tilknytning til bunnen på store dyp svært mangelfull. Med unntak av sporadiske undersøkelser med undervannsfarkoster hvor video benyttes til å få et bilde av hva som måtte finnes av organismer, eller der hvor slike undersøkelser har vært gjennomført systematisk i forbindelse med utforsking av vulkansk aktivitet, er det lite kjent hva som befinner seg ved bunnen på så store dyp. Videopptak vil måtte medføre bruk av kunstig lys, siden det ellers er stummende mørkt der nede, og det er usikkert hvordan kunstig lys vil påvirke de fiskene som måtte være der. Det er tenkelig at lys kan tiltrekke seg fisk, og at en derfor får et feilaktig positivt bilde av hvor tett det er med fisk der, eller at lyset skremmer fisken bort, og at en derfor får et feilaktig negativt bilde av fisketettheten.
Som nevnt er bunntråling på dypere vann enn ca. 1000-1200 m vanskelig, og vi har ikke funnet noen slike dype stasjoner fra selve Norskehavsbassengene i Havforskningsinstituttets databaser. Noe over 1000 stasjoner dypere enn 1000 m er registrert, men alle er tatt langs Eggakanten, og de fleste av disse er knyttet til undersøkelser av blåkveite og andre bunnfisk på dypt vann. Det er ikke helt umulig å tråle dypere om en utruster fartøyet med dette for øyet; i 2004 ble det som et ledd i det store internasjonale prosjektet «Census of Marine Life» gjennomført et tokt til den Midt-Atlantiske ryggen mellom Island og Asorene kalt «Mar-Eco», hvor det ble trålt både i pelagialen og på bunnen. På bunnen ble det tatt 22 hal med en standard samplingtrål (hvor kulene som holder trålen åpen var byttet ut med kuler som tåler slike ekstreme trykk) på dyp fra 800 til 3500 m. Det ble også brukt liner og feller med agn og videokamera for å fange fisk på dyp ned til ca. 4000 m (Porteiro mfl. 2017). Omtrent 300 arter tilhørende 82 familier ble identifisert, hvor av omtrent halvparten var knyttet til bunnen. Dette området ligger så langt fra utredningsområdet at artsammensetningen trolig er helt forskjellig, og det er usikkert om artsmangfoldet er like stort i samme dyp i Norskehavet og i Atlanterhavet.
Noen få undersøkelser av fisk på dypt vann i Norskehavet er verdt å nevne. I 1975 var det en svensk/fransk ekspedisjon til de dype bassengene av Norskehavet, som primært undersøkte den bentiske faunaen i disse bassengene. Blant redskapene de brukte var også en bomtrål, som ikke er designet for å fange fisk, men hvor en ofte fanger noen små bentiske arter (Dahl mfl. 1976). Kun tre arter av fisk ble fanget i bomtrålen; to fra ringbukfamilien og en fra ålekvabbefamilien. I 1995 og 1996 ble det gjort undersøkelser av fiskesamfunnet på kontinentalskråningen i det østlige Norskehavet (Bergstad mfl. 1999). Det ble brukt bunntrål på dyp fra 300 til 2050 m. De fant at biomassen avtok bratt på dypere vann enn ca. 720 m, hvor en krysset 0°C-isotermen. De fanget total 34 ulike arter. Biomassen av fisk i minusvannet var bare 11% av biomassen funnet på Eggakanten og i den øvre delen av kontinentalskråningen. Mens fiskesamfunnet grunnere enn 720 m var dominert av arter som blåkveite, vanlig uer, snabeluer, skolest og isskate, var samfunnet på dypere vann dominert av arktiske arter som arktisk ålebrosme, svart ringbuk og kongeringbuk. I 2011 ble det gjennomført undersøkelser på sokkelen sør for Jan Mayen i forbindelse med et utredningsprogram for olje- og gassvirksomhet i Jan Mayen-sonen (Fossum mfl 2012). Rapporten sammenfattet både det dedikerte toktet til området og litteratur om tidligere undersøkelser. Syttien fiskerarter har vært kjent fra Jan Mayen-sonen fra tidligere, og under toktet i 2011 ble det funnet ytterligere tre arter som ikke var kjent fra området fra før, inkludert en ny art for norske farvann. På toktet i 2011 ble det tatt ni bunntrålhal, på dyp fra 173 m til 924 m. Det ble også tatt tre hal med bomtrål. De hyppigst forekommende artene var nordlig knurrulke (Triglops murrayi), tverrhalet langebarn (Leptoclinus maculatus), langhalet langebarn (Lumpenus lampretaeformis), tornulke (Icelus bicornis), halvnaken ålebrosme (Lycodes seminudus), blek ålebrosme (Lycodes pallidus) og silkeålebrosme (Lycodes adolfi).
Som vi ser, er det ingen undersøkelser av bunnfisk med egnet redskap fra de dypere delene av Norskehavet. I 2018 ble det publisert et atlas over fisk i Arktiske områder (Mecklenburg mfl. 2018a og b) som også kan sies å inkludere store deler av Norskehavet, ettersom negative vanntemperaturer ved bunnen innebærer arktiske forhold. Selv om dette altså i liten grad bygger på observasjoner fra Norskehavet, viser det hvilke bunnfiskarter som med stor sannsynlighet finnes der. Atlaset lister 229 arter, hvorav ca. 85% er knyttet til bunnen. Dette atlaset skiller mellom bentiske arter, som lever på bunnen og inkluderer arter som graver i substratet på bunnen, som ulker, ålekvabber og flyndrer, og demersale (eller bentopelagiske) arter, som lever nær bunnen men også svømmer aktivt over bunnen. Eksempler på demersale arter er torskefisker, skolestfisker, og steinbiter. De ulike økotypene deles igjen inn etter hvilke dyp de typisk lever på, med forstavelsene epi- (0-200 m), meso- (200-1500 m), bathy- (1500-3000 m), og abysso- (dypere enn 3000 m). Tabell 5.1 viser hvilke bunnfiskarter som ifølge atlasets kart og beskrivelser sannsynligvis har en utbredelse i Norskehavet, hvilke dyp de typisk finnes på, og hvilket levesett de har.
Tabell 5.1 Bunnfiskarter med sannsynlig utbredelse i Norskehavet.
Familie
Art
Biotype
Somniosidae
Haier
Somniosus microcephalus
Håkjerring
Meso-demersal
Rajidae
Skater
Amblyraja hyperborea
Isskate
Meso/bathy-bentisk
Amblyraja radiata
Kloskate
Epi/meso-bentisk
Bathyraja spinicauda
Gråskate
Meso-bentisk
Rajella phyllae
Rundskate
Meso-bentisk
Macrouridae
Skolestfamilien
Coelorinchus labiatus
Piggskjellet skolest
Meso/bathy-denersal
Coryphaenoides rupestris
Skolest
Meso/bathy-demersal
Macrourus berglax
Isgalt
Meso-demersal
Lotidae
Brosmefamilien
Brosme brosme
Brosme
Epi/meso-demersal
Gaidropsarus argentatus
Sølvtangbrosme
Meso-demersal
Molva dipterygia
Blålange
Meso-demersal
Molva molva
Lange
Epi/meso-demersal
Sebastidae
Uerfamilien
Sebastes mentella
Snabeluer
Meso-demersal
Sebastes norwegicus
Vanlig uer
Epi/meso-demersal
Cottidae
Ulker
Artediellus atlanticus
Krokulke
Meso-bentisk
Psycrolutidae
Paddeulker
Cottunculus microps
Paddeulke
Meso-bentisk
Agonidae
Panserulker
Leptagonus decagonus
Tiskjegg
Epi/meso-bentisk
Liparidae
Ringbuker
Careproctus micropus
Liten ringbuk
Meso-bentisk
Careproctus reinhardti
Nordlig ringbuk
Meso-bentisk
Paraliparis bathybius
Svart ringbuk
Bathy-demersal
Rhodichtys regina
Kongeringbuk
Meso/bathy-demersal
Zoarcidae
Ålekvabber
Gymnelus retrodorsalis
Spitsbergenålebrosme
Epi-bentisk
Lysenchelys kolthoffi
Marmorert ålebrosme
Meso-bentisk
Lysenchelys muraena
Havålesbrosme
Meso-bentisk
Lysenchelys platyrhina
Flatsnuteålebrosme
Bathy-bentisk
Lycodes adolfi
Silkeålebrosme
Meso-bentisk
Lycodes esmarkii
Ulvefisk
Meso-bentisk
Lycodes eudipleurostictus
Båndålebrosme
Meso-bentisk
Lycodes frigidus
Arktisk ålebrosme
Bathy-bentisk
Lycodes luetkenii
Rosa ålebrosme
Meso-bentisk
Lycodes paamiuti
Paamiutålebrosme
Meso-bentisk
Lycodes pallidus
le kålebrosme
Epi/meso/bathy-bentisk
Lycodes seminudus
Halvnaken ålebrosme
Meso-bentisk
Lycodes squamiventer
Skjellålebrosme
Bathy-bentisk
Lycodonus flagellicauda
Spisshalet ålebrosme
Meso/bathy-bentisk
Stichaeidae
Hornkvabber
Leptoclinus maculatus
Tverrhalet langebarn
Epi-bentisk
Lumpenus lampretaeformis
Langhalet langebarn
Epi-entisk
Anarhichadidae
Steinbiter
Anarhichas denticulatus
Blåsteinbit
Epi/meso/bathy-demersal
Anarhichas lupus
Gråsteinbit
Epi/meso-demersal
Anarhichas minor
Flekksteinbit
Epi/meso-demersal
Pleuronectidae
Flyndrer
Hippoglossoides platessoides
Gapeflyndre
Epi/meso/bathy-bentisk
Reinhardtius hippoglossoides
Blåkveite
Meso-bentisk/pelagisk
5.6 - Oppsummering og kunnskapsbehov
Norskehavet domineres av de store vandrende bestandene av pelagisk fisk som makrell, sild og kolmule. Langs kontinentalskråningen finnes videre en rekke arter, herunder flere kommersielt viktige arter som blåkveite, snabeluer og vanlig uer. For de fleste bestandene der vi har tilstrekkelig kunnskapsgrunnlag til å si noe om bestandsstatus, er status god og bestandene høstes bærekraftig med gytebestander over føre-var-nivå. Noen arter derimot er listet som sårbare, sterkt truet eller kitisk truet på norsk rødliste (Henriksen og Hilmo 2015), herunder vanlig uer, blålange og en rekke bruskfiskarter.
Det brukes betydelige ressurser på å overvåke tilstanden til de kommersielt viktige artene og lange tidserier med data fra fiskeri og forskningstokt gir oss et godt kunnskapsgrunnlag. Bestandsberegningene for sild, makrell, kolmule, blåkveite, vanlig uer, snabeluer baseres på lange tidserier fra forskningstokt, i tillegg til data fra fiskeriene. I bestandsberegningene av blåkveitebestanden brukes en lengdebasert modell, da aldersestimering på denne arten er utfordrende. I tillegg brukes fangstbar biomasse til å beskrive utviklingen i bestanden istedenfor gytebestand. Det arbeides med å utvikle en modell som inkorporerer aldersdata og som vil beregne gytemasse. Dette ventes å påvirke fremtidig rådgivning. Bestandsberegningen for snabeluer og vanlig uer har en noe redusert kvalitet på grunn av manglende dekning av utbredelsesområde. Bestandene av lange, brosme, blålange og vassild dekkes ikke av tradisjonelle undersøkelser og bestandsberegningene er basert på data fra fiskeriene. Slike data er ikke representative og gir kun et indirekte mål på bestandsstørrelsen. For å få et direkte mål på bestandsstørrelse trengs fiskeri-uavhengige data.
Kunnskapsbehovet er større for ikke-kommersielle arter, der det ofte mangler bestandsestimater, informasjon om bestandsstruktur, reproduksjon- og oppvekstområder og vandringer. For en del ikke-kommersielle arter finnes nyttige data fra tidsserier av forskningstokt som har kommersielle arter som målart. Grundig analyse av allerede innsamlet materiale og fremtidig innsamlet materiale vil kunne forbedre kunnskapsgrunnlaget. Merkeforsøk vil kunne gi oss informasjon om vandringer mellom havområder.
Det største kunnskapsbehovet er imidlertid på områder dypere enn 1000-1200 meter, hvor kunnskapsmangelen er betydelig. Det finnes bare sporadiske undersøkelser på større dyp, hvor mange ikke-kommersielle arter lever. Behovet for mer kunnskap om disse ikke-kommersielle artene er til stede da disse kan ha en viktig funksjon i økosystemet som bentisk-pelagisk kobling i næringsnettet. Tråling på slike dyp er utfordrende, men ikke umulig; det stiller spesielle krav både til redskapene som skal brukes og til fartøyenes maskinkraft og dekksutstyr.
Det må videre gjøres en innsats for å sikre mer kunnskap om effekter av klimaendringer, fordi det påvirker utbredelsesområdet, mattilgang og dermed produksjonen i de enkelte bestandene.
6 - Sjøpattedyr
6.1 - Innledning
I løpet av de siste 30 årene med systematisk overvåking har omkring 12 arter av sjøpattedyr opptrådt jevnlig innenfor utredningsområdet. Listen inkluderer 2 selarter (grønlandssel (Pagophilus groenlandicus) og klappmyss (Cystophora cristata)), 5 tannhvalarter (nebbhval (Hyperoodon ampullatus), spermhval (Physeter macrocephalus), spekkhogger (Orcinus orca), narhval (Monodon monoceros) og kvitnosdelfin (Lagenorhyncus albirostris) samt 5 bardehvalarter (vågehval (Balaenoptera acutorostrata), knølhval (Megaptera novaeanglia), finnhval (Balaenoptera physalus), blåhval (Balaenoptera musculus) og grønlandshval (Balaena mysticetus). Flere andre sjøpattedyrarter vites eller antas å forekomme mer eller mindre sporadisk i utredningsområdet, f.eks. ringsel (Pusa hispida), storkobbe (Erignatus barbatus), steinkobbe (Phoca vitulina), seihval (Balaenoptera borealis), grindhval (Globicephala melas), nise (Phocoena phocoena), hvithval (Delphinapterus leucas), kvitskjeving (Lagenorhyncus obliquidens) og nordspisshval (Mesoplodon bidens) samt nordatlantisk retthval også kalt nordkaper (Eubalaena glacialis). Historisk sett har sistnevnte vært vanlig, men de siste 100 årene har man bare kjennskap til en nordkaper, som med stor sannsynlighet har krysset utredningsområdet. Navnet nordkaper er opprinnelig gitt av nederlandske hvalfangere og kommer av at arten kunne fangstes på så langt nord som til Nordkapp (Rice 1998).
Systematisk overvåking av sjøpattedyr i utredningsområdet blir foretatt under Havforskningsinstituttets tokt for telling av vågehval, som har foregått siden 1987. Fra og med 2002, har disse tellinger vært gjennomført som såkalte «mosaikksurveys» med tellinger i forskjellige områder i løpet av en 6 års syklus, som deretter danner grunnlag for et bestandsestimat. Disse toktene har også dannet grunnlag for bestandsestimater og bedre forståelse av forekomstmønster for mange andre hvalbestander. Observasjoner fra Havforskningsinstituttets økosystemtokter samt satellittmerkinger og akustisk overvåking foretatt av flere forskjellige forskningsinstitusjoner har også gitt viktig informasjon om sjøpattedyrenes bruk av Norskehavsområdet.
Alle de nevnte selartene unntatt steinkobbe, yngler på is. Disse arter er også i større eller mindre grad knyttet til is resten av året, særlig under hårfellingsperioden, som foregår 1-3 måneder etter yngleperioden. Isbjørn (Ursus maritimus) jakter av og til helt ute i drivisen (McKinney mfl. 2013), hvor grønlandssel og klappmyss har sine yngleområder. Siden 1990 tallet har isutbredelsen imidlertid blitt såpass redusert at yngleområdene for grønlandssel og klappmyss har ligget utenfor utredningsområdet. Hvalross (Odobenus rosmarus) er knyttet til grunne områder og forekommer kun unntaksvis i drivisen over dyphavsområdene omfattet av denne utredningen.
Blant hvalene i og nært utredningsområdet er det bare grønlandshval, hvithval og narhval som er knyttet til havis året rundt. Hvithvaler forekommer imidlertid kun sjeldent utenfor kontinentalsokkelen i Nordøst-Atlanteren og vil derfor ikke bli ytterligere omtalt i denne rapporten. Narhval er heller ikke direkte observert i utredningsområdet, men data fra lyttebøyer viser at de er til stede i den vestlige delen av Framstredet året rundt (Ahonen mfl. 2019). Noe overlapp med utredningsområdet synes derfor sannsynlig for denne dypdykkende arten. Dessuten er narhval kjent for å være spesielt sensitiv overfor støy og vil dermed kunne påvirkes av aktiviteter inne i utredningsområdet. Sårbarhet overfor menneskeskapt støy er imidlertid ikke unikt for narhval, men gjelder i større eller mindre grad for de fleste sjøpattedyrarter (Kvadsheim mfl. 2020). Nedenfor oppsummeres kunnskapen om bardehvaler, tannhvaler og seler i separate avsnitt. Kunnskap om bruk av lyd og sårbarhet overfor menneskeskapt støy og andre stressfaktorer gjennomgås også.
6.2 - Bardehvaler
6.2.1 - Systematikk og generellt kjennetegn
Bardehvaler (Mysticeti) er en betegnelse for en underorden av hvalene (Cetacea), som kjennetegnes ved ikke å ha frembrutte tenner. Istedenfor henger store keratinplater (barder) ned fra overkjeven (se Figur 6.1). Disse brukes til å tilbakeholde byttedyr i havvann som hvalene tar inn i munnhulen under beiting. Teknikken muliggjør effektiv utnyttelse av store mengder av dyreplankton og små stimefisk og har sannsynligvis vært avgjørende for at de største dyr på kloden har tilhørt denne gruppen (Goldbogen mfl. 2019). De fleste og største bardehvaler tilhører finnhvalfamilien (Balaenopteridae). Dette er strømlinjeformede hurtige hvaler som vågehval, finnhval, seihval, blåhval og knølhval (se Figur 6.2). Den største arten er blåhvalen, som kan bli rundt 30 meter lang og veie opp til 150 ton. Den minste arten er vågehvalen, som kan bli opp mot 10 m lang og veie rundt 9 ton. Grønlandshvalen og nordkaperen tilhører retthvalfamilien (Balaenidae) og kan veie opp mot henholdsvis 100 og 70 tonn. I motsetning til hvaler i finnhvalfamilien, har retthvalene ingen ryggfinne. De er også langt mindre strømlinjeformede enn finnhvalene og svømmer mye langsommere.
Figur 6.1 Nordatlantisk retthval (nordkaper) undersøkes av havforskningspioner Johan Hjort. Bildet menes tatt i Mehamn i 1901 og viser sannsynligvis en av de siste nordkaperne, som ble tatt av norske hvalfangere (privat fotografi vist med tillatelse av Nils Lid Hjort). Hvalen ligger på ryggen og de svarte «børstene» som delvis stikker ut av munnen er bardene. Hos retthvalartene nordkaper og grønlandshval kan bardene bli rundt henholdsvis 3 og 4 meter lange. Hos andre bardehvaler varier den maksimale bardelengde fra rundt 20 cm hos vågehval til rundt 90 cm hos blåhval.
Figur 6.2 Representanter for noen av bardehvalarterne med nåværende eller historisk forekomst i Norskehavet: Øverst t.v. typisk observasjon av vågehval. I motsetning til de større bardehvalene viser vågehvalen sjeldent tydelig blåst (foto: Kjell-Arne Fagerheim, Havforskningsinstituttet); øverst t.h. blåhval i Norskehavet i startfasen av et blåst. Ved full «utblåsing» kan blåhvalblåsten bli rundt 10 meter høy og vil da typisk ha form som en enkel slank søyle, selv om den kommer fra to tettsittende «nesebor» kalt blåsthull. (foto: Kjell-Arne Fagerheim, Havforskningsinstituttet), nederst t.v. beitende knølhvaler med halsfolder, som er typiske for hvaler i finnhvalfamilien (foto: Kjell-Arne Fagerheim, Havforskningsinstituttet), nederst t.h. nordkaperen «Porter» fotografert i Kvænangen etter vandring fra østsiden av USA. Den tydelig todelte blåsten er typisk for retthvalene (foto: Karl-Otto Jacobsen).
Aldersbestemmelse av bardehvaler kan foretas ved hjelp av års-soner i ørevoks og kjemiske analyser av øyelinser eller DNA. Det er usikkerheter knyttet til alle disse metoder, men basert på flere tekniker ser det ut til at vågehvalene kan bli rundt 40-50 år gamle (Hauksson mfl. 2011), knølhvaler opp mot 70 år (Riekkola mfl. 2018) og finnhvaler opp mot 90 år gamle (Konradsson og Gunnlaugsson 1990) og muligvis eldre (Hoffmann 2011). Analyser av øyelinser har vist at grønlandshvaler kan bli over 200 år gamle og først blir kjønnsmodne rundt 20 års alderen (George mfl. 1999, Rosa mfl. 2013). Livslengden hos den nært beslektede nordkaperen, menes å være nærmere 70 år.
Bardehvaler av finnhvalfamilien har folder i underhakepartiet, som setter dem i stand til å utvide munnhulen til en stor pose under beiting (se Figur 6.2). Hos blåhvalen kan en fylt «pose» utgjøre opp mot 2/3 av totalvekten. Det kreves imidlertid mye energi for å fylle og tømme posen, særlig for de største artene, som dermed er særlig avhengige av høy byttedyrtetthet (Goldbogen mfl. 2011, 2012). Dette er antakelig grunnen til at både finnhval og blåhval ofte beiter på 200-300 meters dybde, hvor tettheten av dyreplankton ofte er størst (Friedlaender mfl. 2020).
Grønlandshvaler og nordkapere har ikke samme fleksibilitet i munnhulen som finnhvalene. På grunn av sin eksepsjonelt store munnhule og lange barder (opptil 4 meter), kan de likevel filtrere store mengder vann. Satellittmerking av grønlandshval i Vest-Grønland har vist at de kan beite ned til 582 m og kan være neddykket i opptil 48 minutter (Heide-Jørgensen mfl. 2021). De fleste dykk er imidlertid grunnere enn 300 meter. Under beiting svømmer grønlandshvalen ofte svært langsomt for å redusere vannmotstanden (Simon mfl. 2009). Satellittmerking av nordkapere i Nordvest-Atlanteren har vist beitedykk ned til 175 m og dykketider på opptil 20 minutter, men langt de fleste dykk er mindre enn 30 meter (Baumgartner og Mate 2003).
Et interessant spørsmål er hvordan bardehvalene finner frem til områder med høy byttedyrtetthet. I motsetning til tannhvaler kan de ikke bruke biosonar og synet er antakelig også av begrenset verdi på større vanndyp. Sannsynligvis foretar de ofte prøvedykk eller «prøvefiltreringer», for å vurdere kvaliteten av et beiteområde (Simon mfl. 2009, Friedlaender mfl. 2020). Det ser også ut som om de kan orientere seg ut fra signaler fra andre arter. F.eks. kan sildespisende knølhval bli tiltrukket av lyder fra sildespisende spekkhoggere (Benti mfl. 2021).
6.2.2 - Utbredelse og bestandshistorikk
Sommertellinger av hvaler over perioden 1987-2019 har vist høy forekomst av vågehval (Figur 6.3), finnhval (Figur 6.4a) og knølhval (Figur 6.4b) rundt hele den arktiske fronten og til dels også de mere atlantisk dominerte deler av utredningsområdet (Øien 2009, Leonard og Øien 2020a, b, NAMMCO 2021) I perioden 2014-2018 ble det også gjort flere observasjoner av blåhval i dette området (Leonard og Øien 2020b). Vågehval og finnhval er også observert i relativt store tettheter i områdene rundt Knipovichryggen (Øien 2009, Leonard og Øien 2020 a,b, NAMMCO 2021). Mohnryggen forbinder områder med høy tetthet av bardehval i Danmarksstredet, vestkysten av Spitsbergen og områder lenger øst og nord.
Knølhval er mest observert i ytterpunktene av aksen fra Danmarksstredet til Vest-Spitsbergen, bortsett fra perioden 2014-2018, hvor observasjoner også ble gjort rundt Mohnryggen (Leonard og Øien 2020a og 2020b). Observasjoner av både finn- og knølhval i perioden 1995-2001 antyder imidlertid også en alternativ vandringsrute fra området rundt Jan Mayen til eggakanten inn mot Barentshavet via Vøringutstikkeren og den nordlige kanten av Vøringplatået (Øien 2009). Det er bare gjort noen få observasjoner av seihval på sommertelletoktene for vågehval i Norskehavsområdet og Barentshavet (Leonard og Øien 2020 a, b) og på opportunistiske observasjonsplattformer rundt Svalbard (Storrie mfl. 2018). Dette står i sterk kontrast til store seihvalfangster som ble rapportert fra kysten av Møre og Finnmark på slutten av 1800 tallet (Christensen mfl. 1992). Skift i fangstfordelingen mellom år er blitt tilskrevet fluktuasjoner i fordelingen av Calanus finnmarchicus, som var det vanligst observerte innhold i seihvalmager på hvalfangststasjonene (Ingebrigtsen 1929). I de seneste årtier er de største Nordatlantiske forekomstene av seihval observert i Irmingerhavet og områder rundt den midtatlantiske ryggen (Prieto mfl. 2014, Houghton mfl. 2020), men det er usikkert om disse hvalene tilhører samme bestand, som ble fangstet på i Norskehavet og Barentshavområdet (Cooke 2018a, Huijser mfl. 2018).
Figur 6.3 Observasjoner av vågehval i Nord-Atlanteren basert på a) alle tilgjengelige telletoktdata for perioden 1987-2015 ifølge NAMMCO (2021) og b) Havforskningsinstituttets sommertelletokter i perioden 2014-2019 (Havforskningsinstituttet, upubliserte data)
Figur 6.4 Fordeling av a) finnhvaler og b) knølhvaler under Havforskningsinstituttets sommertelletokter i perioden 2014–2018. Blå områder viser havis. Fra Leonard og Øien 2020b
Grønlandshvaler i Nordøst-Atlanteren ser i hovedsak ut til å følge iskanten året rundt (Kovacs mfl. 2020, De Boer mfl. 2019) og er ikke blitt direkte observert i utredningsområdet under vågehvaltoktene (Leonard og Øien 2020 a, b). Satellittmerkingsdata antyder imidlertid noe overlapp om høsten/vinteren i den nordligste delen (Hamilton mfl. 2021). Det er også gjort flere observasjoner utenfor arktiske farvann, blant annet tre observasjoner utenfor kysten av Finnmark i 1980-1981 (Wiig mfl. 2010) og 3-4 observasjoner i farvannet rundt Irland, Sør-England og Frankrike i 2016 (De Boer mfl. 2017). De nyeste observasjonene menes å være av unge hvaler. I Nordvest-Atlanteren er en ung grønlandshval blitt observert i Gulf of Maine over tre somrer i perioden 2012-2017, mer enn 10 breddegrader sør for den normale sommerutbredelsen for nordvestatlantiske grønlandshvaler (Accardo mfl. 2018). Her ble hvalen observert beitende sammen med nordkapere og seihvaler. Det er ikke observert noen nordkapere i Norskehavsområdet fra vågehvaltoktene, men 5 mulige observasjoner er gjort i farvannene rundt Norge, Færøyene og Island i perioden 1999-2003 (Monsarrat mfl. 2015). Av disse ble 2 observasjoner med sikkerhet identifisert som nordkapere med tidligere tilhold ved vestkysten av USA (Monsarrat mfl. 2015). Alle bardehvalene i Nord-Atlanteren har vært utsatt for et betydelig kommersielt fangsttrykk i løpet av de seneste århundrer. Kystnære konsentrasjoner av nordkapere i nordøst Atlanteren ble fangstet ned allerede på 1500- og 1600-tallet. Fangsten ble gradvis utvidet til det åpne havet og fra midten av 1800-tallet anses nordkaperen som tilnærmet utryddet i norske farvann (Cooke 2020). Arten ble imidlertid fortsatt fangstet rundt Færøyene, Skottland og Shetland tidlig på 1900 tallet (Monsarrat mfl. 2015). En enkelt nordkaper ble også tatt ut for Vest-Norge i 1926 (Jonsgård 1977). I dag antas det at stort sett alle nordkapere tilhører en bestand på rundt 400 individer som holder til på østkysten av Nord-Amerika (Pettis mfl. 2020). Det nyeste bestandsestimatet tyder på en tilbakegang fra det høyest registrerte nivået etter hvalfangstperioden på 476 i 2010 (Waring mfl. 2016). Arten er nå klassifisert som kritisk truet (Cooke 2020). Modellering av fangstdata kombinert med økologiske data fra en nært beslektet retthvalart i det Nordlige Stillehavet (Eubalaena japonica) har gitt estimater for bestandsstørrelsen før hvalfangstperioden på henholdsvis ~5000-12000 for Nordøst-Atlanteren og ~3000-9000 for Nordvest-Atlanteren (Monsarrat mfl. 2016). Genetiske undersøkelser har imidlertid sådd sterk tvil om hvorvidt retthvalene på hver sin side av Nord-Atlanteren faktisk utgjorde separate bestander (Foote mfl. 2013).
Grønlandshvalbestanden i Nordøst-Atlanteren var også nesten utryddet allerede på 1800-tallet (Allen og Keay 2006). Fra et estimert bestandsnivå på om lag 50000 rundt år 1600 (Allen og Keay 2006), teller bestanden i dag bare noen få hundre individer og er klassifisert som kritisk truet i Norsk rødliste for arter (Henriksen og Hilmo 2015). Dette er motsatt til utviklingen i Stillehavsområdet og Nordvestatlanten, hvor bestandene har tatt seg kraftig opp og i dag er estimert til henholdsvis ~17000 og ~7000 individer (Givens mfl. 2016, Doniol-Valcroze mfl. 2020). I de seneste årtier har antallet observasjoner av grønlandshval også økt i Nordøst-Attlanteren, men det er usikkert om dette skyldes økt oppmerksomhet rundt arten, innvandring fra andre bestander eller en reell bestandsøking (Wiig mfl. 2010, Vacquie-Garcia mfl. 2017a, De Boer mfl. 2019, Boertmann mfl. 2020). Det er funnet forskjeller i forekomstratene av forskjellige genotyper mellom Nordøst-Atlanteren og alle andre havområder (Bachmann mfl. 2020). Dette viser, at hvalene ikke fordeler seg tilfeldig, men utelukker likevel ikke at det kan foregå en vis utveksling mellom områder. Eventuell utveksling vil være mest sannsynlig i form av innvandring fra de store bestandene rundt Beringhavet og i Nordvest-Atlanteren til den historisk lave bestanden i Nordøst-Atlanteren. Selv relativt få innvandrede individer vil kunne få betydning for bestandsutviklingen i dette området. Flere genetiske analyser (Foote mfl. 2013, Alter mfl. 2012) samt dateringer av subfossil benmateriale (Dykes og Savelle 2001) viser at det sannsynligvis har vært betydelig utveksling mellom grønlandshvalbestandene rundt polhavet i tidligere perioder med lite is. Under fangstperioden ble det også funnet flere harpunspisser av antatt nordatlantisk opprinnelse i grønlandshvaler tatt i stillehavsområdet (Bockstoce og Burns 1993). Det kjennes ikke til sikre nåtidige utvekslinger av grønlandshvaler mellom de 4 definerte bestander i verden (Baird og Bickham 2021), men flere observasjoner utenfor vanlige utbredelsesområder viser at særlig unge grønlandshvaler kan utforske nye områder (Heide-Jørgensen mfl. 2012, De Boer mfl. 2017). Fysisk mulighet for nåtidige vandringer gjennom polhavsområdet støttes av at atlantiske arter som klappmyss og narhval er blitt observert i det nordlige Stillehavet (Burns og Gavin 1980, COSEWIC 2004). Omvendt er det i perioden 2010-2013 påvist innvandring av 2 gråhvaler fra Stillehavet til Atlanterhavet, hvor denne kystnære benthos-spisende arten ellers har vært utdød i mere enn 200 år (Scheinin mfl. 2011, Alter mfl. 2015, Hoelzel mfl. 2021). Et annet tegn på utveksling av sjøpattedyr mellom disse to havområder er at selsykdommen PDV, populært kallet «selpest», har spredt seg fra Atlanterhavet til Stillehavet (Van Wormer mfl. 2019).
Nye fangsttekniker på slutten av 1800 tallet muliggjorde effektiv fangst av de hurtigsvømmende bardehvalene i finnhvalfamilien. De største artene som blåhval, finnhval, knølhval og seihval ble deretter fort drevet til ekstremt lave nivåer i Nord-Atlanteren (Christensen mfl. 1992, Vikingsson mfl. 2015). Etter fredningstiltak på 1950 tallet har totalbestandene av finnhval og knølhval imidlertid tatt seg kraftig opp i Nordøst-Atlanteren og er nå vurdert som livskraftige i den norske rødlisten (Henriksen og Hilmo 2015). Sommerforekomstene av finnhval og knølhval i områdene dekket av Havforskningsinstituttets vågehvaltellinger ble estimert til henholdsvis 11387 (CI: 8072–16063) og 10708 (CI: 4906–23370) under seneste tellesyklus i perioden 2014-2018 (Leonard og Øien, 2020b). Disse estimater skiller seg ikke vesentlig fra tidligere tellinger i periodene 2002-2007 og 2008-2013 (Leonard og Øien 2020a). Antallet av blåhvalobservasjoner var for lavt til å gi grunnlag for estimater. Også ellers i Nord-Atlanteren ser denne arten ut til å ha hatt vanskeligheter med å ta seg opp efter overfangstingen. Estimatet for blåhval i hele Nordøst-Atlanten var bare ~1000 dyr i perioden 1987-2001 (Pike mfl. 2009). Senere tellinger rundt Island og Grønland har imidlertid gitt et estimat for dette område på 3000 (CI 1377–6534) (Pike mfl. 2009). Seihvaler har opptrådt for sjeldent i Norskehavsområdet til at antallet kan estimeres, men Storrie mfl. (2018) rapporterte en del observasjoner rundt Svalbard. Dette er i den nordligste delen av artens utbredelse. Under storhvalfangsten, var finnmarkskysten ett av de viktigste fangstområdene for alle store finnhvalarter (Christensen mfl. 1992). Dette gjaldt også blåhvaler og seihvaler, som imidlertid ikke har blitt observert der i nyere tid (Leonard og Øien 2020a,b). Det tyder på at bestandskomponentene som beitet i disse områdene ble mer eller mindre utryddet, og at disse artene enda ikke har blitt tallrike nok til å rekolonisere hele sitt tidligere utbredelsesområde.
Vågehvalen ble først utsatt for betydelig fangst fra 1930 tallet (Skaug mfl. 2004). I de tidlige årene kan denne fangsten ha redusert antallet av vågehvaler noe, men etter en 5 års pause i vågehvalfangsten fra 1988-1993, har den nordøstatlantiske bestanden av vågehval holdt seg på et stabilt nivå på rundt 100000 (Solvang mfl. 2015, IWC 2018). Vågehvaler rundt Jan Mayen anses å tilhøre en annen bestand (den sentralatlantiske vågehvalbestanden), som har vist betydelig variasjon i forekomst over perioden 1987 til 2013 (IWC 2018). Årsaken til denne dynamikken er ikke kjent, men endringer i fordelingsmønster kan sannsynligvis spille inn. Rundt Jan Mayen ble den estimerte forekomsten av vågehval redusert fra rundt 25000 ved tellinger i 1997 og 2005 til 11000 (CI 4498-35911) i perioden (2008-2013) (NAMMCO 2021). Usikkerheten rundt det seneste estimatet er vel å merke stor. Etter gjenopptakelsen av vågehvalfangsten i 1993 har den norske fangsten ligget på under 1% av bestandsestimatet på de berørte bestandene (Solvang mfl. 2015) og antas ikke å kunne føre til bestandsnedgang.
6.2.3 - Vandringsmønster, habitatbruk og diett
De fleste bardehvaler i finnhvalfamilien foretar beitevandringer mot polene om våren fra parings og kalvingsområder i tropiske og subtropiske områder. Det er imidlertid betydelig variasjon i dette mønsteret både innenfor og mellom arter. Det er også fortsatt stor usikkerhet om hvor store deler av populasjonene som foretar disse vandringene hvert år, og når på året de vandrer. Opptak fra stasjonære og mobile lyttebøyer i Nordøst-Atlanteren viser at noen finnhvaler og knølhvaler oppholder seg i de nordiske hav utover vinteren og for begge arter er det registrert signaler, som kan tyde på parringsatferd eller «øving» på parringsatferd (Simon mfl. 2010, Ahonen mfl. 2017, Magnusdottir og Lim 2019, Aniceto mfl. 2020). Så langt ser det ut til at det bare er hanner som produserer disse lydene men tilstedeværelsen av knølhvalhunner er også dokumentert basert på DNA analyser av biopsier tatt om vinteren (A. Rikardsen, Norges Arktiske Universitet, upubliserte data). Under perioden for storhvalfangst i Nord-Norge frem til tidlig på 1900 tallet ble det observert både veldig unge fostre og nesten ferdigutviklede fostre hos knølhval (Ingebrigtsen 1929). Basert på dette har det blitt foreslått, at både paring og kalving kan foregå i nordlige farvann, men dette er ikke bevist. Nyere studier av individuelt gjenkjennbare mønstre på halefinner av knølhvaler har derimot vist at knølhvaler som beiter utenfor Nord-Norge og ellers i Nord-Atlanteren ofte vandrer til kjente parringsområder i Karibien og utenfor Vest-Afrika (Stevick mfl. 2003, Wenzel mfl. 2020). Satellittmerking av 25 finnhvaler ved Svalbard i september 2015-2019, viste at over halvparten vandret sørover i løpet av 1-2 måneder (Lydersen mfl. 2020). En god del av disse viste tegn på beiteatferd i området rundt Mohnryggen og i noen grad også Knipovichryggen. Resten ble igjen i området rundt Svalbard og beitet særlig langs kontinentalskråningen vest og nord for Spitsbergen (Lydersen mfl. 2020). Av de finnhvalene som vandret sørover, ble noen sporet helt til kysten av Portugal og Marokko, hvor atferden tydet på paring eller kalving (Lydersen mfl. 2020). Satellittmerking av blåhvaler og finnhvaler ved Azorene om våren har vist at de fleste vandret mot Island og Øst-Grønland i løpet av noen uker (Silva mfl. 2013). De viste imidlertid også beiteatferd rundt Azorerne. Om vinteren er blåhvaler også observert i høyproduktive oppstrømningsområder utenfor Mauritania, hvor beiteforholdene antas å være gode (Baines og Reichelt 2014). Ingen av disse har imidlertid blitt satellittmerket. Veldig lite er kjent om overvintringsområdene for nordøstatlantiske vågehvaler (Cooke 2018b). Sommerbeitingen i nordlige farvann er antakelig avgjørende for de migrerende bardehvalenes overlevelse og reproduksjonsevne, selv om det også foregår noe beiting andre steder. Beregninger viser at forstyrrelser av beiteatferd eller reduksjon i byttedyrtetthet fort kan få betydelig effekt på energibalansen til f.eks. blåhval (Guilpin mfl. 2020). Blåhvalen og grønlandshvalen beiter nesten utelukkende på dyreplankton (Cooke 2018c, Cooke og Reeves 2018), mens de andre nevnte artene har en mer opportunistisk diett med større innslag av småfisk som lodde og sild (Nøttestad mfl. 2015, Vikingsson mfl. 2015, Windsland mfl. 2007, Tartu mfl. 2020).
Selv om grønlandshvalene i Nordøst-Atlanteren stort sett holder seg til drivisen året rundt, kan de likevel foreta betydelige vandringer. En hunnhval merket i Framstredet våren 2010, migrerte om lag 950 km sørover langs iskanten i løpet av 14 dager (Lydersen mfl. 2012). Den tok imidlertid også en del avstikkere ut i åpent hav utenfor iskanten, hvor den kunne tilbringe flere dager i et begrenset område, antakelig på grunn av beiting. Det meste av tiden tilbrakte den over kontinentalskråningen mot Øst-Grønland eller langs med undervannsrygger i nærheten. Ytterligere 13 grønlandshvaler ble merket mai-juni 2017 (Kovacs mfl. 2020) og viste generelt høy preferanse for havisområder, særlig om vinteren. De fleste oppholdt seg da i den vestlige delen av Framstredet i iskonsentrasjoner opp mot 100%. Det finnes ikke noen analyser av diett for Nordøstatlantiske grønlandshvaler, men fra Vest-Grønland/Øst-Canada og Stillehavsområdet er det kjent at de kan beite både på subarktiske og arktiske arter av hoppekreps (Calanus finnmarchicus, C. glacialis, C. hyperboreus) samt diverse arter av krill (Laidre mfl. 2007, Pomerlau mfl. 2011). Det er også vist at grønlandshvalene beiter året rundt (Fortune mfl. 2020). Om vinteren beiter de antakelig på overvintrende stadier av hoppekreps og krill, som står dypt i vannsøylen eller helt på bunnen (Fortune mfl. 2020). Dette i kombinasjon med ekstrem fettlagringskapasitet gir sannsynligvis arten økt fleksibilitet og mulighet for å tåle faste over lengre perioder enn andre hvalarter (Fortune mfl. 2020). Satellittmerkingsstudier tyder på at grønlandshvaler foretrekker havområder med en overflatetemperatur på mindre enn 2°C (Chambault mfl. 2018, Kovacs mfl. 2020). Dette er blitt tolket som tegn på problemer med regulering av kroppstemperaturen ved høyere temperaturer (Chambault mfl. 2018). Detaljerte studier av grønlandshvalenes anatomi og fysiologi indikerer imidlertid at arten har gode forutsetninger for å kunne regulere sin kroppstemperatur også i varmere vann (George mfl. 2021). Generell preferanse for kalt vann med et vis isdekke er sannsynligvis styrt av flere faktorer i grønlandshvalens livsstrategi. Studier i kanadiske og vestgrønlandske farvann tyder på at grønlandshvaler bl.a. søker disse områder for å unngå konkurranse fra andre bardehvaler (Laidre mfl. 2012) samt predasjon fra spekkhoggere (Matthews mfl. 2020a). Det siste er spesielt viktig for at grønlandshvalene skal kunne gjennomføre sin ekstremt langsomme livssyklus. I kanadiske farvann ser man imidlertid at grønlandshvalene oftere befinner seg i åpent vann når det ikke er spekkhoggere i nærheten (Matthews mfl. 2020a). Spekkhoggerpredasjon på store bardehvaler er ikke observert i like stor grad i Nordøst-Atlanteren som i Nordvest-Atlanteren (McCordick mfl. 2014).
Flere forskere foreslår at en tilsynelatende ekstra sterk ispreferanse hos nordøstatlantiske grønlandshvaler kan skyldes at kun de mest istilknyttede individene overlevde fangstperioden her og har «opplært» sine etterkommere i samme habitatbruk (Boertmann mfl. 2015, Kovacs mfl. 2020). I de seneste år er det imidlertid observert enkelte yngre grønlandshvaler langt sør for isutbredelsesgrensene i både Nordvest-Atlanteren (Accardo mfl. 2018) og Nordøst-Atlanteren (De Boer mfl. 2017), herunder Nordsjøområdet. Konkrete årsaker til denne habitatbruken er ukjent, men sannsynligheten for atypisk atferd hos enkeltindivider stiger med økende bestandsstørrelse etter fredning, som med stor sannsynlighet er påvist hos den nordvestatlantiske bestanden (Doniol-Valcroze mfl. 2020). De sørlige observasjonene illustrerer, at yngre grønlandshvaler kan oppsøke andre habitattyper enn de som er dokumentert for overleverne av hvalfangsten og deres etterkommere. Fysiologisk sett skulle det heller ikke være umulig for grønlandshvaler å beite i en del av de samme områdene som de nært beslektede nordkapere. Dette støttes av at samme grønlandshval er observert beitende med nordkapere og seihvaler over flere år i Gulf of Maine (Accardo mfl. 2018). På grunn av farge, størrelse og kraftigere lydproduksjon er grønlandshvaler imidlertid lettere å oppdage i åpent vann enn nordkapere og vil derfor antakelig være mere utsatt for spekkhoggerpredasjon. Yngre og mindre grønlandshvaler vil derfor sannsynligvis ha størst suksess med denne habitatbruken. Det kan være nok en grunn til at det nettopp er slike hvaler, som er observert langt utenfor artens typiske habitat.
Nordkaperne i Nordvest-Atlanteren vandrer mellom kalvingsområder i Florida og beite- og paringsområder i grenseområdene mellom USA og Kanada (Gowan mfl. 2019). De er spesielt knyttet til områder med høye konsentrasjoner av kopepodittstadie 5 (C5) av Calanus finnmarchicus (Baumgartner mfl. 2003). Det er imidlertid også gjort en del registreringer i det dype Irmingerhavet både visuelt og ved hjelp av lyttebøyer (Mellinger mfl. 2011, Monsarrat mfl. 2015). Hovedparten av registreringene er gjort i juli-august og overlapper både i tid og rom med høye konsentrasjoner av C5stadiet av C. finnmarchicus i de øvre vannlag (Heath mfl. 2008). Lignende beiteforhold finnes i store deler av Norskehavet (Broms mfl. 2009) og habitatmodelleringsstudier identifiserer Norskehavet som et potensielt meget gunstig habitat for arten (Monsarrat mfl. 2016). Manglende observasjoner utelukker ikke at arten kan forekomme i området. Beitende nordkapere er spesielt vanskelige å få øye på, da de er mørke i fargen, mangler ryggfinne og ofte opptrer alene. Det har heller ikke vært foretatt målrettede akustiske undersøkelser av nordkaperforekomsten i Norskehavet. Observasjonen av nordkaperen «Porter» i Kvænangsfjorden september 1999 (Jakobsen mfl. 2003) viser imidlertid at en vis forekomst av individer på beitevandring er sannsynlig. Basert på kjennetegn på rygg og hode, ble det fastslått at samme hval var registrert med navn i en amerikansk hvalfotokatalog. Den var sist observert ved Cape Cod 117 dage tidligere og hadde altså svømt minst 5700 km i den mellomliggende periode. «Porter» ble observert beitende en ukes tid i Kvænangen og ble senere igjen observert ved Cape Cod (Jacobsen mfl. 2003, se også Figur 6.2).
6.2.4 - Bruk av lyd og sårbarhet ovenfor undervannsstøy
Bardehvaler kommuniserer ved hjelp av mellom- og lavfrekvente lyder som i stor grad overlapper med frekvensområdet for skipsstøy, seismikk og militære sonarer og derfor kan bli helt eller delvis overdøvet eller «maskert» av dem. Hvalene må derfor ofte modifisere enten frekvens, hyppighet eller styrke på sine lydsignaler for å kunne gjennomføre sin kommunikasjon (Kvadsheim mfl. 2020). Det kan være ekstra energikrevende og kvaliteten på kommunikasjonen blir muligvis ikke like god. Blåhvalen er den eneste bardehvalen, som jevnlig lager lyd om sommeren under sitt beiteopphold i Norskehavet (Ahonen mfl. 2017). Lydopptak fra Framstredet har vist veldig aktiv lydproduksjon av grønlandshval om vinteren, særlig i den vestlige delen (Ahonen mfl. 2017). Signalene hadde karakter av kompliserte «sanger», som tolkes som tegn på paringsatferd. Nordkaperen har generelt lavere kildestyrke på sine lyder enn andre bardehvaler i Nord-Atlanteren og anses også for å ha lite mulighet for å kompensere for støy ved å øke lydstyrken (Hatch mfl. 2012).
Forskning på effekter av militære sonarer på sjøpattedyr i Norskehavsområdet har vist, at vågehval er en særlig sårbar bardehvalart, som kan unngå det eksponerte området også etter at aktiviteten er opphørt (Kvadsheim mfl. 2017). Knølhval viser mindre tydelige reaksjoner (Sivle mfl. 2015). Det er ikke utført studier av sjøpattedyrs reaksjoner på seismikkskyting i Norskehavsområdet (Kvadsheim mfl. 2020). Det er uheldig, da reaksjoner på forstyrrelser ofte vil variere både mellom arter, bestander og områder. Eksempelvis ser man at reaksjonene på forstyrrelser kan avhenge av hvilken atferd sjøpattedyrene normalt har i eksponeringsområdet. Grønlandshvaler ser f.eks. ut til å fortsette å beite i produktive områder under seismikkskyting, men reduserer sin tid i overflaten (Robertson mfl. 2016). Grønlandshvaler på gjennomreise vil derimot ofte være mer tilbøyelige til å øke avstanden til seismikkeksponerte områder (Koski mfl. 2009). Selv om hvalene ikke nødvendigvis fjerner seg fra en støykilde kan de likevel ha en fysiologisk stressreaksjon. Dette er bl.a. dokumentert ved forandringer i hormonnivåer hos nordkapere ved østkysten av USA på dager med sterkt nedsatt trafikk etter terrorangrepet 11. september 2001 (Rolland mfl. 2012).
Det kan oppstå midlertidige eller permanente skader på hørselsorganer ved eksponering veldig nært ved støykilder som seismikk-kanoner og sonarer (Kvadsheim mfl. 2020). Vurderinger av potensialet for dette hos bardehvaler er imidlertid usikre da flere elementer i bardehvalenes hørselsfysiologi fortsatt er dårlig kjent (Southall mfl. 2019, Kvadsheim mfl. 2020). På grunn av åpenlyse problemer med å utføre sansefysiologiske eksperimenter på så store dyr er det også begrenset kunnskap om bardehvalenes optimale hørselsområder og grenseverdier for hørbare frekvenser og lydstyrker. Dette øker usikkerheten på angivelse av sikkerhetsavstander for f.eks. seismikkskyting og sonarbruk. Det antas imidlertid at permanente fysiske skader generelt kun vil forekomme innen en avstand av omkring 100 meter for alle sjøpattedyr (Kvadsheim mfl. 2020). Unntaket er diverse undervanns-detonasjoner, som kan ha skadeeffekt på mye lengre avstand. Forstyrrelser av viktig atferd som beiting, parring, ivaretakelse av unger etc. kan over tid føre til nedsatt helsetilstand og bestandsstørrelser i sjøpattedyrbestander, men det er vanskelig å sette grenseverdier for når disse effektene kan forventes.
6.2.5 - Andre menneskelige påvirkninger av bardehval
Bardehvaler er generelt lavt plassert i fødekjeden og akkumulerer derfor sjeldent så store mengder av miljøgifter at det kan anses å utgjøre en helserisiko for dem (Thomas mfl. 2016, Tartu mfl. 2020). De er antakelig mer sårbare overfor fysiske påvirkninger som innvikling i fiskeredskap (også drivende «spøkelsesgarn») og kollisjoner med fartøy. Det er imidlertid lite kunnskap om omfanget av dette i Norskehavsområdet. Effekter av klimaendringer på bardehvalene vil antakelig primært avhenge av hvordan disse påvirker forekomsten av byttedyr og fødekonkurrenter. Flere studier tyder bl.a på, at en viss økning av havtemperaturene sammenlignet med nivåene mellom 1950 og 1970 så langt kan ha hatt en positiv effekt på fødetilgangen for grønlandshvalene i Nordøst-Atlanteren (Falk-Petersen mfl. 2009) og andre steder (George mfl. 2015). Derimot er økende havtemperaturer i de nordvestatlantiske beiteområdene for nordkaper ledsaget av en reduksjon i kroppskondisjon (Record mfl. 2019), som settes i forbindelse med endret byttedyrfordeling (Davis mfl. 2017, Record mfl. 2019). Også nordkaperne selv har endret fordeling og er dermed antakelig blitt mer utsatt for skipskollisjoner og innvikling i fiskeredskaper (Thomas mfl. 2016). I en periode med generelt stigende temperaturer fra 1993 til 2013 er det også observert en reduksjon i kroppskondisjonen til nordøstatlantiske vågehvaler (Solvang mfl. 2017). Dette antas bl.a. å skyldes økt konkurranse mellom vågehval og torsk, som over den angitte perioden både opplevde mer gunstige miljøbetingelser og redusert fangsttrykk. Stigende temperaturer har også blitt knyttet til økende forekomster av giftige alger i flere havområder, inkludert nordlige og arktiske farvann (Gobler mfl. 2017, McKenzie mfl. 2021). Det er registrert forekomster av algegifter i mange sjøpattedyrarter i disse områder, men foreløpig er det ikke registrert sikre tilfeller av forgiftingssymptomer i nordlige farvann (Lefebvre mfl. 2016, Hendrix mfl. 2021). Mer subtile effekter på helse og reproduksjon kan imidlertid ikke utelukkes. Slike effekter anses bl.a. for mulige medvirkende årsaker til den dårlige bestandssituasjonen hos nordkaperen (Doucette mfl. 2012) og visse steinkobbebestander rundt de britiske øyer (Jensen mfl. 2015). Giftige algeoppblomstringer rapporteres primært i kystnære områder, men noen potensielt giftige alger er rapportert fra Norskehavsområdet (Okolodkov 2005). Det er ikke foretatt undersøkelser av algegiftforekomster hos sjøpattedyr i norske farvann eller øvrige deler av utredningsområdet.
6.3 - Tannhvler
6.3.1 - Systematikk og generelle kjennetegn
Som navnet sier er hvalene i tannhvalunderordenen (Odontoceti) kjennetegnet ved å ha frembrutte tenner. Disse hvaler beiter i all hovedsak på fisk og blekksprut og tennene brukes til å gripe tak i byttedyrene. Tannhvalene har også spesialiserte organer, som setter dem i stand til å bruke ekkolokalisering for å finne frem til sitt bytte (Foskolos mfl. 2019). Størrelsesspekteret for tannhvaler spenner fra niser på rundt 1,5 meter til spermhvaler på opptil 16 meter. Kvitnosdelfiner og narhvaler blir henholdsvis opptil 3 og 6 meter lange, mens spekkhoggere og nebbhvaler kan bli om lag 10 m lange. Spermhvaler og spekkhoggere kan bli opp mot henholdsvis ~70 og ~90 år gamle (Olesiuk mfl. 1990, Evans mfl. 2002), mens narhvaler kan bli helt opp til 115 år gamle (Garde mfl. 2007). Til sammenlikning var de eldst kjente nebbhvaler fra hvalfangsttiden blott 37 år gamle (Christensen 1973). Liknende livslengde rapporteres for kvitnosdelfin, men det er veldig lite data på denne arten.
Figur 6.5 Representanter for tannhvalartene i Norskehavsområdet: Øverst t.v. nebbhval med sitt typiske runde hode og markante nebblignende kjeveparti. Som de fleste tannhvaler har nebbhvalene ett enkelt blåsthull på på toppen av hodet. Det er imidlertid 2 neseganger som leder inn til det ene hullet (Kjell-Arne Fagerheim, Havforskningsinstituttet); øverst. t.h.: spermhval som ligger og puster i overflaten. Den skrå vinkel på blåsten er typisk for spermhvaler og skyldes at blåsthullet sitter på venstre siden av hodet hos spermhval. Ulikt andre tannhvaler har spermhvaler 2 helt oppdelte neseganger, hvorav kun det venstre har forbindelse til et blåsthull (Kjell-Arne Fagerheim, Havforskningsinstituttet); nederst t.v. spekkhogger med unge (Arve Kristiansen, Havforskningsinstituttet); nederst t.h. kvitnosdelfin (Kjell-Arne Fagerheim, Havforskningsinstituttet)
6.3.2 - Utbredelse og bestandshistorikk
De mest utbredte tannhvalarter innen utredningsområdet er spermhval, nebbhval, kvitnosdelfin og spekkhoggere (Øien 2009, Leonard og Øien 2020 a, b, se også Figur 6.5). De to første av disse er utpreget dypdykkere med kapasitet for lange dykk, som reduserer muligheten for å observere og telle dem. Spermhvalen observeres imidlertid relativt ofte fra telletoktene for vågehval (Figur 6.6), særlig i de sørlige og østlige deler av utredningsområdet langs Vøringsutstikkeren, sørlige kanten av Vøringplatået og rundt Mohn og Knipovich ryggene samt eggakanten inn mot Barentshavet (Øien 2009, Leonard og Øien 2020 a, b). Noen observasjoner er også gjort rundt Jan Mayen. Spermhvalen ble et ettertraktet fangstobjekt på 1800 tallet og intensiv jakt over hele Nord-Atlanteren har med all sannsynlighet redusert forekomsten i Norskehavet til et historisk lavt nivå frem til fredning i 1973 (Christensen mfl. 1992). Forekomsten av spermhval i Norskehavet har holdt seg relativt stabil over perioden 1995-2018, men det estimerte nivået rundt 5000 dyr er sannsynligvis for lavt, da det ikke er korrigert for redusert synlighet på grunn av lange dykketider (Øien 2009, Leonard og Øien 2020 a,b). Spermhval yngler ikke i norske farvann og finnes derfor ikke på den norske rødliste for arter. På europeisk nivå er arten klassifisert som sårbar, da den fortsatt menes å være under 50% av bestandsstørrelsen før fangsten begynte (Taylor mfl. 2008a).
Figur 6.6 Fordeling av a) spermhvaler og b) spekkhoggere under telletokter i perioden 2014–2018. Blå områder viser havis. Fra Leonard og Øien (2020b)
Nebbhval er i nyere tid primært blitt observert rundt Jan Mayen og langs Vøringutstikkeren (Leonard and Øien 2020 a og 2020b). Fangstposisjoner fra perioden 1938-71 viser imidlertid også høy forekomst ved eggakanten utenfor Svalbard og på Knipovichryggen (Øien og Hartvedt 2011). Det siste støttes også av observasjoner rapportert av Storrie mfl. (2018). Nebbhvalen kalver antakelig i de nordiske hav, men det er ikke kjent om det er spesielle kalvingsområder. Bestanden av nebbhval i Norskehavet anses som livskraftig i norsk rødliste for arter ut fra en antatt bestand over 2000 dyr (Henriksen og Hilmo, 2015). Dette er basert på et estimat på 2700 individer (95 % CI 900-8000) rapportert for området rundt Jan Mayen før 2011 (Fossum mfl. 2012). Vurderingen for hele arten er imidlertid, at det er for lite data til å avgjøre status (Taylor mfl. 2008b). Bestanden av nebbhvaler i Norskehavet antas å ha blitt betydelig redusert under hvalfangstperioden frem til 1973 (Øien og Hartvedt 2011). Antallet nebbhvalobservasjoner under telletokter for vågehval har vært for lave for å gi grunnlag for pålitelige bestandsestimater, men det ble gjort flere observasjoner i perioden 2014-18 enn tidligere (Leonard og Øien 2020 b).
Det er gjort 2 observasjoner av Nordspisshval i Norskehavsområdet (Carlstrøm mfl. 1997). Denne art er imidlertid særdeles vanskelig å få øye på, og undersøkelser fra Nordvest-Atlanteren viser at data fra lyttebøyer antakelig gir et bedre inntrykk av forekomsten av disse og andre nebbhvalarter (Stanistreet mfl. 2017). Det er også rapportert flere strandete kadavr av nordspisshval i kystområdene rundt utredningsområdet.
Narhval er aldri blitt registrert innenfor utredningsområdet, hverken på sommertelletokt eller ved hjelp av satellittmerking. Det har imidlertid vært merket noen få narhvaler og lyttebøyer i Framstredet registrerer narhval året rundt (Ahonen mfl. 2019). Det bør derfor tas høyde for at denne arten kan forekomme i de nordligste delene av utredningsområdet. Genetiske undersøkelser har vist, at narhvalene i nordøst-Atlanteren er genetisk forskjellige fra langt større narhvalbestander i Nordvest-Atlanteren (Louis mfl. 2020). Det er også funnet signifikante genetiske forskjeller mellom narhvaler i Nordøst-Grønland og ved Svalbard (Louis mfl. 2020). Tellinger av narhval i Øst-Grønland tyder på en markant nedgang eller omfordeling av arten i dette område (NAMMCO 2019a, NAMMCO 2019b), mens tellinger nord for Svalbard har vist større forekomst enn forventet (Vacquie-Garcia mfl. 2017a). Totalt sett har de siste tellinger gitt estimater på i underkant av 3000 narhvaler i Nordøst-Atlanteren, men begge tellinger har sannsynligvis vært ufullstendige.
Sommertelletoktene for vågehval viser relativ høy forekomst av spekkhogger i den sørvestlige delen av utredningsområdet rundt Jan Mayen, langs Vøringutstikkeren og i området rundt Mohnryggen (Leonard og Øien 2020a og 2020b, se Figur 6.6). I tillegg rapporterer Storrie mfl. (2018) om en del observasjoner ut for eggakanten vest for Spitsbergen. Spekkhogger oppholder seg antakelig i store deler av utredningsområdet året rundt, men de fleste observasjoner stammer fra sommerperioden (Jourdain mfl. 2019, Leonard og Øien 2020 a og 2020b). Mellom 1938 og 1981 ble det skutt rundt 3000 spekkhoggere i Nord-Atlanteren av norske hvalfangere og den amerikanske marinen (Naval Aviation News 1956, Øien 1988). Dette påvirket antakelig bestandsnivået, men det finnes ingen telledata fra denne perioden. Basert på data fra vågehvaltellingene har forekomsten av spekkhoggere i Norskehavet variert fra 18821 (11529-30753) i perioden 2002-2007 til 9563 (CI:4713-19403) i perioden 2008-2013 (Leonard og Øien 2020a,b) . Det seneste estimatet, for perioden 2014-2018, er på 15056 (CI: 8423-26914), så det har ikke vært noen klar trend. Spekkhoggerbestanden er klassifisert som livskraftig i norsk rødliste for arter (Henriksen og Hilmo 2015).
Delfinarten kvitnos registreres ofte utenfor kontinentalskråningen vest av Svalbard og Bjørnøya (Leonard og Øien 2020 a,b). Disse dyr er antakelig del av en større bestand, som har sin hovedutbredelse i Barentshavet og teller rundt 190000 dyr. Arten kan forveksles med arten kvitskjeving (Lagenorhyncus obliquidens), som det imidlertid finnes meget få sikre observasjoner av i området (Leonard og Øien 2020 a og 20202b). Springere er en fellesbetegnelse for disse to artene. Ved siste telling i perioden 2014-2018, var det få tvetydige artsbestemmelser og den estimerte totalbestanden av springere på ~193000 var antakelig nesten utelukkende kvitnos-delfiner (Leonard og Øien 2020b). Bare rundt 20000 av disse ble imidlertid estimert å forekomme innen utredningsområdet under tellesyklusen fra 2014-2018, særlig rundt Knipovich-ryggen (Leonard og Øien 2020b). Kvitnos er klassifisert som livskraftig i norsk rødliste for arter (Henriksen og Hilmo 2015).
6.3.3 - Vandringsmønster, habitatbruk og diett
Det er bare kjønnsmodne hanner av spermhval, som forekommer i norske farvann (Fossum mfl. 2012). Hunndyrene og de yngste hannene oppholder seg i subtropiske og tropiske farvann. Spermhvalen er kjent for å kunne dykke dypere enn 2000 meter (Watkins mfl.1993). Dykkedata fra Norskehavsområdet viser, at spermhvalene her beite både i det benthopelagiske laget under 700 meter og på dyp mellom 1000 og 2000 meter (Sarvas 1999, Isojunno og Miller 2018). Diettdata for spermhval i Nordøst-Atlanteren finnes bare fra hvalfangsten (Martin og Clarke 1986, Christensen mfl. 1992) og fra strandete dyr (f.eks Santos mfl. 1999, Santos mfl. 2002). Blekkspruten Gonatus fabricii dominerer mageinnholdet i de fleste rapporter, men spermhvaler som ble tatt ut for Øst-Grønland under islandsk hvalfangst inneholdt også en del fisk, primært rognkjeks (Cyclopterus lumpus) (Martin og Clarke 1986). Disse data er fra 1977-81 og ingen nyere fangstbaserte data eksisterer, da det ble helt slutt på spermhvalfangsten i hele Nord-Atlanteren kort tid etter disse undersøkelsene.
Studier av et mindre antall strandete eksemplarer av nebbhval i Nordøstatlanten indikerer at denne hvalen også langt overveiende spiser G. fabricii, gjerne i livsfaser som opptrer på store dyp (f. eks. Fernandez mfl. 2014). Data fra fangstperioden frem til 1973 viser, at nebbhval også kan spise fisk som uer (Sebastes spp.), blåkveite (Rheinhardtius hippoglossoides), havmus (Chimaera monstrosa), lange (Molva molva) og pigghå (Squalus acanthias) (Benjaminsen og Christensen 1979). Innslaget av fisk i nebbhvaldietten var imidlertid langt større i Labradorhavet enn i Norskehavet og Grønlandshavet. Det er også eksempler på at nebbhval kan spise bentiske organismer som sjøpølser, sjøstjerner og reker (Reeves mfl. 1993, Taylor mfl. 2008b). Dykkedata fra uforstyrrete nebbhvaler rundt Jan Mayen viser hyppige dykk ned til 500-700 meters dyp og noe sjeldnere dykk til 1600 m (Wensween mfl. 2019).
For narhvaler er kunnskapen om habitatbruk i Nordøst-Atlanteren primært basert på hvaler, som er satellittmerket under sommeropphold i Scoresbysundfjorden i Øst-Grønland. Disse hvaler viser sterk preferanse for vannmasser med temperaturer mellom 0.5 og 2.0°C og beveger seg ikke lengre enn ca. 100 km ut fra fjordmunningen under beitesesongen på vinteren (Heide-Jørgensen mfl. 2020). Det finnes imidlertid en større gruppering av narhvaler, som holder til nord for Svalbard om sommeren (Vacquie-Garcia mfl. 2017a) og det er ukjent hvor denne gruppen har sine vinterbeiteområder. Det samme kan sies om narhvaler som er observert i en bukt lenger nord om sommeren (NAMMCO 2019a). Predasjon fra spekkhogger menes å være en viktig kontrollerende faktor for narhvalbestandene i Nordvest-Atlanteren (Lefort mfl. 2020), men det finnes ikke like god oversikt over dette i Nordøst-Atlanteren.
Spekkhoggerne i Norskehavet beiter hovedsakelig på pelagisk fisk som sild og makrell, men noen grupper kan også angripe og spise andre sjøpattedyr (Jourdain mfl. 2019). Den totale hyppigheten av dette i norskehavsområdet er imidlertid vanskelig å bedømme da eksisterende data primært er innsamlet i kystnære områder (Foote mfl. 2009, Jourdain mfl. 2017). Lite er kjent om dietten til kvitnos, men i andre områder beiter de primært på pelagisk fisk og noe blekksprut (Craddock mfl. 2009, Fall mfl. 2014).
6.3.4 - Bruk av lyd og sårbarhet for undervannsstøy
Tannhvaler produserer lyd både i forbindelse med romlig orientering og kommunikasjon. Deres lydrepertoar har et mer begrenset frekvensoverlapp med de fleste menneskeskapte støykilder enn for bardehvalene, og de antas derfor å være mindre utsatt for «maskering» (Kvadsheim mfl. 2020). Det er imidlertid dokumentert at skipsstøy kan føre til maskering av visse typer av spekkhoggerlyder til kommunikasjon og at spekkhoggerne øker styrken på disse signaler for å kompensere for støyen (Holt mfl. 2011). Spermhvaler bruker et spesielt bredt frekvensspekter av klikklyder til ekkolokalisering og en studie fra Mexikogulfen viser en reduksjon i spermhvalklikk i forbindelse med eksponering for skipsstøy (Azzara mfl. 2013). Den nøyaktige årsak til dette er imidlertid ikke klar. I norskehavsområdet har det vært fokusert spesielt på effekter av militære sonarer på sjøpattedyr (Kvadsheim mfl. 2020). Eksperimentelle studier har vist at nebbhvaler reagerer særlig sterkt på støy fra mellomfrekvente militære sonarer (Miller mfl. 2015, Wensveen mfl. 2019). Reaksjonene inkluderte opphør av beiting samt unnvikelse av det eksponerte området i opptil 24 timer etter eksponeringens opphør. Noen nebbhvaler utførte også abrupte dypdykk, som innebærer en risiko for «dykkersyke» (Kvadsheim mfl. 2020). Flere massestrandinger av forskjellige nebbhvalarter har sammenfalt i tid og rom med militær sonarbruk og det anses for sannsynlig, at det er en form for årsakssammenheng (Kvadsheim mfl. 2020). Funn av gassbobler og blødninger i vevet hos noen av disse hvaler kan tyde på at dykkersyke som følge av forstyrrelser i dykkeatferden kan være en mulig årsak (Fernández mfl. 2005). I motsetning til nebbhvalene ser spekkhoggere, spermhvaler og grindhvaler ut til å reagere på sonarstøy ved å redusere antall dype dykk (Sivle 2012, Kvadsheim mfl. 2020) og responsen skjer ved høyere eksponeringsstyrke enn hos nebbhval. På tross av hyppig bruk av seismikk ved olje- og gassleting i norske farvann er det overraskende lite informasjon om reaksjoner på seismikkstøy hos sjøpattedyr i Norskehavsområdet. Studier av narhval i Scoresbysundområdet viser, at disse oppholder seg nærmere overflaten ved eksponering for seismikkstøy (Boertmann mfl. 2020). I andre farvann har tannhvaler typisk reagert på seismikk ved å øke sine dykketider og forflytte seg horisontalt fra lydkilden (Kavanagh 2019, Heide-Jørgensen mfl. 2013, Harwood og Kingsley 2013, Ahonen mfl. 2019, Kyhn mfl. 2019).
6.3.5 - Andre menneskelige påvirkninger av tannhvaler
Tannhvaler har særlig høy sannsynlighet for opphopning av miljøgifter i kroppen. Det skyldes at de er høyt plassert i fødekjeden, kan leve lenge og er dårligere til å nedbryte miljøgifter enn f.eks. seler (Boon mfl. 1997). Til tross for at miljøgiften PCB har vært forbudt siden 1980 tallet er det likevel funnet nivåer over grenseverdien for helseskadelige effekter hos flere tannhvalarter i Nordatlanteren (f.eks. Jepson mfl. 2016, Desforges mfl. 2018, Dietz mfl. 2019, Andvik mfl. 2020). De fleste spekkhoggere i Norskehavsområdet ser ut til å beite primært på sild og makrell, men noen grupper tar også sel og har vesentlig høyere miljøgiftnivåer enn de som bare spiser fisk (Andvik mfl. 2020). Teoretiske modelleringsstudier har antydet, at miljøgiftnivåene i pattedyrspisende spekkhoggere kan føre til utrydding av disse bestander innen 2100 (Desforges mfl. 2018). Der er imidlertid knyttet betydelig usikkerhet til disse prognoser (Desforges mfl. 2019, Witting 2019). Undersøkelser av nyere miljøgifter som perfluorerte forbindelser har vist stigende forekomst i tannhvaler rundt Færøerne (Rotander mfl. 2012). Giftstoffer produsert av mikroalger er påvist i flere tannhvalarter (e.g. Lefebvre mfl. 2016, Davis mfl. 2019), men så langt ikke i Nord-Atlanteren. Fra sørlige farvann er det også observert sykdommer hos delfiner knyttet til flere forskjellige algegifter (f.eks. Davis mfl. 2019, Fire mfl. 2020).
6.4 - Seler
6.4.1 - Systematikk og generelle kjennetegn
Alle selarter med tilknytning til utredningsområdet tilhører familien ekte seler (Phocidae) i underordenen Pinnipedia, som også omfatter sjøløver, pelsseler og hvalrosser. De ekte selene har ikke noe ytre øre og har et hårdekke uten underull. Varmeisolasjon oppnås i stedet nesten utelukkende ved hjelp av spekklaget. Ekte seler er også kjennetegnet ved å ha en mye kortere dieperiode enn andre sjøpattedyr. Den varierer i lengde fra 4 dager hos klappmyss til rundt 6 uker hos ringsel (Pomeroy 2011). Mødrene har generelt redusert fødeinntak under dieperioden og må derfor «spare opp» energi under resten av året. Parring foregår umiddelbart etter at hunnen har sluttet med å die årets unge. Fosteret setter seg imidlertid først fast (implanteres) og begynner å vokse i livmoderen etter 3-4 måneder. Før dette har selene gjennomgått en hårfelningsperiode på et par uker, hvor fødeinntaket også er sterkt begrenset. Klappmyss og storkobbe er de største selartene i Nord-Atlanteren og begge kan bli over 2,5 meter lange og veie rundt 400 kg. Ringselen er den minste selarten og kan bli opptil 1,6 meter lang. Seler orienterer seg i vann ved hjelp av et fremadrettet og høyt spesialisert syn (Debey og Pyenson 2013), god hørsel (Reichmut mfl. 2013) og registrering av vibrasjoner med værhårene (Dehnhart mfl. 2001, Mariott mfl. 2013). Det har av og til vært hevdet, at noen selarter kan ekkolokalisere, men uten spesialiserte fysiologiske tilpasninger er det lite trolig at selenes bruk av lyd kan sammenliknes med tannhvalenes biosonar (Schustermann mfl. 2000). Nylig har man imidlertid funnet at Weddellsel (en dypdykkende sel fra Antarktisområdet) lager ultrasoniske lyder, som potensielt kan brukes til primitiv ekkolokalisering på nært hold (Cziko mfl. 2020). Selenes gode evne til retningsbestemmelse av lyder er også svært nyttig for orientering og beiting.
Figur 6.7 Klappmyss ved Østgrønland kommer til overflaten etter å ha fanget en uer på dypet. (Foto: Aqqalu Rosing-Asvid, Grønlands Natursinstitutt)
6.4.2 - Utbredelse og bestandshistorikk
Klappmyss tilbringer det meste av beitesesongen langs undervannsrygger og kontinentalskråninger i De nordiske hav og er den mest utbredte selarten i utredningsområdet (Folkow mfl. 1996, Folkow mfl. 2010, Vacquie-Garcia mfl. 2017b, Hamilton mfl. 2021, se Figur 6.7 og 6.8). Modellering av klappmyssbestanden viser en dramatisk bestandsnedgang fra rundt 1,4 millioner (CI:671147-2049240) i 1946 til rundt 200000 i 1980 (CI:110983-300624) og en ytterligere nedgang til ~80-90000 (CI:61845-109857) over perioden 2005-2018 (Se Figur 6.9) (ICES 2019). På grunn av dette er klappmyssbestanden i Nordøst-Atlanteren klassifisert som sterkt truet i Norsk rødliste for arter (Henriksen og Hilmo 2015). For hardt fangsttrykk menes å være hovedårsaken til nedgangen frem til 1980 og kan fortsatt ha spillet en vis rolle frem til 2006, da kommersiell fangst av denne arten ble helt stoppet (Øigård mfl. 2014). Den manglende respons på innstilling av fangsten tyder imidlertid på, at klappmyss også er påvirket av andre faktorer som f.eks. klimaendringer og evt. andre miljøendringer. Både klimaendringer og fiskerier kan i tillegg ha ført til endringer i byttedyrtetthet og sammensetting, som kan ha redusert næringsgrunnlaget. I takt med at yngle - og hårfellingsområdene er kommet nærmere land ser det også ut til at klappmyssen utsettes for hardere predasjonspress fra isbjørn (McKinney mfl. 2012). Rester av klappmyss er også funnet i en av 7 undersøkte spekkhoggermager med innhold rapportert av Bourque mfl. (2018). Alle disse spekkhoggerne var tatt på sensommeren ved Sørøst-Grønland og er derfor mest sannsynlig nordvestatlantisk klappmyss på vei til eller fra hårfellingsområder på pakkisen i Danmarksstredet (Andersen mfl. 2013). Det er også gjort observasjoner av spekkhoggere lenger mot nord (De Boer mfl. 2019), som sammenfaller noenlunde i tid og breddegrad med hårfellingen hos nordøstatlantiske klappmysser (Folkow mfl. 1996). Spekkhoggerne er imidlertid kun observert i åpent vann og ser også generelt ut til å unngå havis ellers i Nord-Atlanten (e.g. Jourdain mfl. 2019, Matthews mfl. 2020a). Utenom yngle- og hårfellingsperiodene vil de nordøstatlantiske klappmyssene imidlertid også være vanskelige å jakte på da de generelt opptrer ganske spredt, dykker dypt og ikke bruker lyd under beiting, som tannhvalene gjør. De seneste årtiers utvikling mot mindre og mer ustabil is i Grønlandshavet kan imidlertid ha gjort de yngste klappmyssene mer sårbare for spekkhoggerpredasjon, da det gir mindre adgang til en trygg hvileplattform etter den ultrakorte dieperiode på 4-7 dager. Både klimaendringer og fiskerier kan i tillegg ha ført til endringer i byttedyrtetthet og sammensetting, som kan ha redusert næringsgrunnlaget sammenliknet med f.eks. 1940 og 50 tallet, hvor nedgangen ser ut til å ha begynt. På dette tidspunktet skjedde det bl.a. en voldsom økning i de kommersielle fiskerier på grunn av nye fiskeletings-teknologier mm. (Gullestad mfl. 2014).
I motsetning til klappmyss, viser den modellerte bestandsutviklingen for grønlandssel en stigende tendens fra et lavpunkt på rundt 300000 dyr (CI:267708-364791) i 1970 til rundt 400000 dyr (CI:313400-532107) (ICES 2019). Det er imidlertid betydelig usikkerhet omkring denne utvikling, særlig på grunn av et observert fall i ungeproduksjonen i 2018 sammenlignet med tidligere flytellinger. Denne arten har som helhet lite overlapp med utredningsområdet, men kan likevel av og til forekomme i betydelige antall på grunn av stor bestandsstørrelse (Hamilton mfl. 2021). Det kan også forekomme noen grønlandsseler fra Barentshavsbestanden i området rundt Framstredet (Nordøy mfl. 2008).
Det finnes ingen fullstendige estimater av antallet av ringsel eller storkobber i bestander relevante for utredningsområdet. Bestanden av ringsel på vestsiden av Spitsbergen kan imidlertid ha gått tilbake på grunn av særdeles dårlige yngleforhold de seneste to årtier (Descamps mfl. 2017). Dette skyldes redusert forekomst av snødekket fjord-is, som er det optimale ynglehabitatet for ringsel. Storkobber kan yngle på flak av bre-is og klarer seg derfor sannsynligvis foreløpig bedre. De opptrer imidlertid svært spredt og er vanskelig å telle. Steinkobber forekommer i en liten bestand på Vestsiden av Spitsbergen hvor de yngler på land om sommeren. Denne bestanden ble estimert til 1888 (95% CI: 1660–3023) dyr i 2009-2010 (Merkel mfl. 2013) og er klassifisert som sårbar i den norske rødliste for Svalbard (Henriksen og Hilmo 2015).
Figur 6.8 Prosentvis årlig habitatbruk for klappmyss basert på a) individer og b) lokaliteter. Modifisert fra Hamilton mfl. (2021). Prikkete områder angir det totale utbredelsesområdet.
6.4.3 - Vandringsmønster, habitatbruk og diett
Satellittmerkingsstudier har vist, at klappmyssene tilbringer ca 60% av tiden sin på lange beitevandringer utenfor drivisområdene (Folkow mfl. 1996). Arten er den eneste spesialiserte dypdykker blant de nordatlantiske selene, og kan dykke over 1000 m (Folkow og Blix 1999). Områdene rundt Jan Mayen ryggen, Mohnryggen, Knipovichryggen og kontinentakryggen inn mot Barentshavet er blant de områder, som brukes mest (se Figur 6.8 a og b).
Det er ikke kjent hvilke byttedyr klappmyssene beiter på her, men data fra andre områder viser, at klappmyss kan beite på flere fiskearter, som er vanlige i utredningsområdet. Dette gjelder bl.a. lodde, vassild, uer, blåkveite, sild og torsk (Hammill og Stenson 2000). Klappmyssdietten i drivisen langs Øst-Grønland er dominert av arktiske arter som blekksprutten G. fabricii og polartorsk (Haug mfl. 2004, 2007), som også kan finnes i deler av utredningsområdet. Basert på klappmyssens fordeling og dykkemønster, har også kolmule vært foreslått som en mulig byttedyrart (Folkow og Blix 1999), men dette er ikke verifisert av konkrete funn. Så langt har datainnsamling og analysemetoder ikke vært egnet til å bestemme artsspesifikk diett utenfor drivisområdene. Vesterisbestanden av grønlandssel beiter langs iskanten i hele området fra Øst-Grønland og Jan Mayen området til det østlige Barentshavet (Folkow mfl. 2004). Satellittmerkingsdata fra 1990 tallet viser, at voksne grønlandsseler krysser den nordlige delen av utredningsområdet på vegen til og fra Barentshavet. Grønlandsselunger merket i 2017 vandrer imidlertid primært nord for Svalbard (Hamilton mfl. 2021). Også grønlandssel fra Barentshavet forekommer av og til i den nordlige delen av utredningsområdet (Nordøy mfl. 2008). Grønlandssel beiter primært på krill og amfipoder fra vår til høst og på småfisk som lodde og polartorsk om vinteren (Haug mfl. 2004, Enoksen mfl. 2016). Denne bestanden føder sine unger på isen i Kvitsjøen ca. en måned tidligere enn i Vesterisen.
Steinkobber fra en liten bestand ved Kong Karls forland beiter av og til ved eggakanten vest for Spitsbergen (Blanchet mfl. 2014). De opptrer imidlertid hovedsakelig mer kystnært i likhet med de arktiske selartene ringsel og storkobbe (Hamilton mfl. 2021).
6.4.4 - Bruk av lyd og sårbarhet for undervannsstøy
Alle arter av sel i Nord-Atlanten bruker lyd i forbindelse med parringslek eller markering av revir under vann (Stirling 1973, Perry og Terhune 1999, Van Parijs mfl. 2001, Van Parijs mfl. 2003). En del av selenes lydrepertoar overlapper med menneskeskapt støy fra f.eks. skipsfart, men det er gjort få studier på hvordan selenes atferd påvirkes av dette. En studie fra Alaska viste imidlertid, at steinkobber reduserte sin vokalisering under påvirkning av overlappende skipsstøy (Matthews mfl. 2020b). Det finnes noen få studier av selers reaksjoner på lyd fra seismikk-kanoner eller sonarer (Harris mfl. 2001, Kvadsheim mfl. 2010). En typisk reaksjon er at selene holder hodet over vannet. Dette vil potensielt kunne redusere beiteaktiviteten. Hørselsområdet for seler overlapper med de fleste lyder produsert av hvaler (Duarte mfl. 2021). Det kan dermed ikke utelukkes at seler kan bruke lydsignaler fra f.eks. ekkolokaliserende tannhvaler som hjelp til lokalisering av byttedyrkonsentrasjoner.
6.4.5 - Andre menneskelige påvirkninger av sel
Som fiskespiser befinner klappmyssen seg på et relativt høyt nivå i fødekjeden og har derfor ikke uventet noe høyere verdier av miljøgifter enn andre arter, som spiser mere dyreplankton og/eller fisk eller blekksprut av mindre størrelse (Dietz mfl. 2019). De målte miljøgiftnivåene i de senere år overstiger imidlertid ikke grenseverdier for antatte effekter på reproduksjonsevnen hverken hos klappmyss eller andre selarter relevante for utredningsområdet. Det kjennes til flere tilfeller av dødelige epidemier hos seler, men dette har så vidt man vet ikke forekommet hos selarter i områdene omfattet av dette studie. Økt spredning av sykdomsorganismer sørfra som følge av høyere temperaturer kan imidlertid innebære en større risiko for sykdomsforekomst hos arktiske bestander (Burek mfl. 2008). Giftstoffer produsert av mikroalger er påvist i en lang rekke selarter (Jensen mfl. 2015, Lefebvre mfl. 2016), men det eksisterer ingen data for bestandene som opptrer innen utredningsområdet.
6.5 - Fremtidsutsikter
Fremtidig status for sjøpattedyrene i utredningsområdet vil variere betydelig mellom arter på grunn av forskjeller i biologi, økologi og nåværende utgangspunkt. Både menneskeskapte og naturlige miljøendringer vil være vesentlige drivere av utviklingen. Da de fleste bestander har betydelig vandringskapasitet, vil dessuten habitatkvaliteten i andre områder kunne påvirke forekomsten i utredningsområdet. Knølhval og finnhval i Norskehavsområdet har økt markant etter fredning og vågehvalbestanden har vært stabilt høy over lang tid. Fremtidig utvikling vil avhenge av habitatkvaliteten i det totale området som er tilgjengelig for disse meget mobile arter. Det ser blant annet ut som om de i økende grad søker inn i tidligere isbelagte farvann rundt Svalbard og Øst-Grønland, delvis på bekostning av oppholdstiden i Norskehavsområdet. De mer spesialiserte plankton-etere har respondert langsommere på fredning, men blåhval og grønlandshval viser nå tegn til økning rundt utredningsområdet. Basert på observerte habitatpreferanser virker det sannsynlig at forekomsten av blåhval og seihval fremover kan øke i deler av utredningsområdet, mens fraværet av is gjør det lite sannsynlig at grønlandshval vil forekomme systematisk i området. Observasjoner av enkelte unge grønlandshvaler langt sør for arktiske farvann de senere år reiser likevel spørsmålet om hvorvidt denne arten kan være i stand til å utnytte deler av nisjen til den nært beslektede nordkaperen. Sistnevnte er kritisk truet i Nordvest-Atlanten og omtrent utryddet i Nordøst-Atlanteren. Basert på de lave bestandstall i Nordvest-Atlanteren synes det usannsynlig, at det vil skje en desidert gjenetablering av nordkaper i overskuelig fremtid i Norskehavsområdet. Arten har imidlertid stor vandringskapasitet og ser ut til å endre fordeling som respons på klimadrevne endringer i fordelingen av Calanus finnmarchicus. Da Norskehavet antakelig vil forbli ett av tyngdepunktene for denne planktonarten, kan en vis fremtidig forekomst av beitende nordkapere ikke utelukkes.
En annen art, som så langt ikke har respondert positivt på fredning er klappmyssen. Reduksjon i utbredelse og stabilitet i havisdekket spiller sannsynligvis en rolle i dette og vil sannsynligvis fortsette å påvirke klappmyssen negativt fremover. På grunn av forventede naturlige og menneskeskapte påvirkninger vil alle sjøpattedyrartene i utredningsområdet sannsynligvis oppleve fremtidige endringer i tilgjengelighet på byttedyr og andre viktige egenskaper ved leveområdene sine. Disse påvirkningene omfatter klimaendringer, endringer i høstingsmønster og øvrig menneskelig aktivitetsmønster i og nært området. Utvikling av nye marine næringer hører med i dette bilde. Militæraktiviteten i området er også økende, blant annet i form av aktiv overvåking av nye stillegående ubåter med militære sonarer. Noen sjøpattedyrarter reagerer kraftig på disse signalene, men også støy fra skipsfart og seismikk kan påvirke atferden til flere arter. De samlede fremtidige bestandseffekter av forventede miljøendringer er imidlertid veldig vanskelige å kvantifisere.
6.6 - Oppsummering og kunnskapsbehov
Basert på tilgjengelig kunnskap kan utredningsområdet forventes å være særlig viktig for de regionalt stasjonære artene som klappmyss, nebbhval og spekkhogger. Disse artene beiter i store deler av området året rundt og får unger i eller nært utredningsområdet, som dermed også får status som oppvekstområde. Nebbhvalen er den av disse artene det finnes minst kunnskap om. Det finnes ingen absolutte bestandsestimater for hele Norskehavet og forståelsen av bestandsstruktur, vandringer og habitatbruk er kun basert på fangstfordelingsdata fra tiden før 1973. Moderne metoder som satellittmerking og overvåking med lyttebøyer kunne forbedre kunnskapsstatus betraktelig for denne arten. Det er kjent, at nebbhval er ekstremt sårbar overfor akustiske forstyrrelser fra militære mellomfrekvente sonarer, men lite er kjent om artens reaksjoner på andre typer av undervannsstøy. Det finnes heller ingen oppdaterte studier av biologiske parametere eller data på diett, miljøgiftinnhold og generell helsetilstand til denne arten i Norskehavet. En del av dette er vanskelig å studere uten forskningsfangst, men noe informasjon kan innhentes fra strandete dyr eller ved hjelp av satellittmerking.
Telletoktene for vågehval har gitt god kunnskap om tallrikheten av de vanligst forekommende hvalartene, men det er fortsatt stor usikkerhet om sesongmessig variasjon i forekomst, da vågehvaltoktene bare utføres om sommeren. Bedre sesongmessig forståelse av hvalenes habitatbruk vil være viktig for å minimere effekter av menneskelig aktivitet i området. På grunn av veldig få observasjoner er bestandsutviklingen hos blåhval, seihval og grønlandshval fortsatt usikker. Nordkaperen ser så langt ikke ut til å ha gjeninnvandret til Norskehavet og viser negativ utviklingstrend i sitt nåværende habitat i Nordvest-Atlanteren. Det er imidlertid registrert flere individer i det nærliggende Irmingerhavet, som også tidligere var et viktig fangstområde. Modellering indikerer imidlertid at Norskehavet har et stor potensiale som beitehabitat for denne arten.
Nordøstatlantisk klappmyss har ikke vist noen tegn på gjenreising av bestanden etter den dramatiske fangstdrevne reduksjon fra 1945 frem til 1980 tallet. Både økt predasjon og endrede miljøforhold kan være medvirkende faktorer til dette. Bedre kunnskap om diett i de viktigste beiteområder samt effekter av endret isutbredelse er antakelig viktige brikker for å forstå denne bestandssituasjonen. Økologiske interaksjoner med andre predatorer kan også være relevant. Dette gjelder blant annet de andre dypdykkende sjøpattedyrene i området som nebbhval og spermhval. Med sin store kroppsstørrelse utgjør spermhvalene i Norskehavet en betydelig biomasse og dykketidskorrigerte bestandsestimater samt bedre data på diett og beiteatferd vil være viktig for å få best mulig forståelse av deres økologiske betydning. Oppdatert kunnskap om nivåer og effekter av menneskeskapte miljøgifter samt gifter produsert av alger er også viktige brikker i forståelsen av bestandssituasjonen til flere sjøpattedyrarter. Spekkhoggere er kjent for å kunne angripe og spise andre sjøpattedyr. Faren for spekkhoggerangrep kan også fordrive visse sjøpattedyrarter fra gunstige beiteområder. Undersøkelse og overvåking av spekkhoggerbestandenes størrelse, vandringer og beitemønster kan derfor være en nøkkel til å forstå endringer i forekomst og utbredelse av flere andre sjøpattedyrarter.
Nyere akustiske og visuelle registreringer av nordkaper nært Norskehavet samt identifiseringen av Norskehavet som et gunstig habitat for denne arten gjør, at man bør ta høyde for mulig forekomst av beitende individer av denne kritisk truede arten i utredningsområdet. Overvåking med lyttebøyer vil være spesielt nyttig for å få bedre oversikt over forekomsten av sjeldne og vanskelig observerbare arter som nordkaper, grønlandshval, narhval og arter av nebbhvalfamilien.
7 - Bentisk-pelagisk kobling
7.1 - Innledning
Organismer som lever i de frie vannmassene blir betegnet som pelagiske mens de organismer som lever ved eller på bunn eller i bunnsediment blir betegnet som bentiske. Der er ingen tydelig skillelinje mellom pelagiske og bentiske arter og heller ingen tydelig skille mellom pelagiske og bentiske økosystem. Mange bunnlevende arter, som korall og krepsdyr, er pelagiske som larver og ved tidlige livsstadier men blir bunnlevende (bentiske) som adulte (voksne). Andre arter vandrer mellom den bentiske og pelagiske verden hver dag. Bentisk-pelagisk kobling innefatter alle de prosesser som kobler de pelagiske og bentiske økosystemene sammen (Figur 7.1). Det kan være ved at de flytter energi, organismer eller næringsemner mellom de to sonene. Den bentiske-pelagiske koblingen spiller en viktig rolle når det kommer til å styre produksjon, struktur (sammensetning) og stabilitet i bentiske så vel som pelagiske økosystem (Graf 1992, Griffiths 2017). Den bentiske-pelagiske koblingen er generelt ansett å være sterkest og derfor også viktigst i grunne sokkelhav og ved kysten men er blitt identifisert som viktig på flere forskjellige plasser i det dype Atlanterhavet (Billet mfl. 1983, Lampitt mfl. 1985, Graf 1989) og Stillehavet (Pile og Young 2006).
Figur 7.1 Hovedprosesser som knytter sammen pelagialen med bunn og omvendt, så kalt bentisk-pelagisk kobling.
7.2 - Målinger fra De nordiske hav
Det finnes noen studier fra De Nordiske hav av de hovedprosesser som knytter sammen pelagialen med bunn og bunn med pelagialen i.e. 1) vertikal migrasjon, 2) trofiske interaksjoner, 3) den biogeokjemiske syklusen og 4) oseanografiske prosesser, men studiene er få, svært begrenset romlig sett, kun rettet mot en prosess av gangen og gir ikke på noen som helst måte en oversikt over situasjonen i Norskehavet i sin helhet. Heller, så gir de oss et innblikk i prosesser som er blitt identifisert som viktig for enkelt tidspunkter og enkelt lokaliteter. Vår nåværende kunnskap om vertikal migrasjon er blitt summert tidligere i rapporten, i.e. mesodyreplankton i kapittel 4.4, makrodyreplankton i kapittel 4.5 og fisk i kapittel 5. Det samme gjelder for den biogeokjemiske syklusen og trofiske interaksjoner mellom planteplankton, dyreplankton og fisk (kapittel 4 og 5). Trofiske interaksjoner mellom bunndyr og pelagiske organismer i det dype Norskehavet er nesten helt ukjent. Det om er imidlertid helt klart er at med veldig få unntak er de dyr som lever på bunn av Norskehavet totalt avhengig av mat som blir produsert av planteplankton i de øvre vannlag med hjelp av sollys. Hvordan dette blir transportert fra den fotiske sonen nedover i vannsøylen og regulerer biomasse og sekundær produksjon ved bunn er derfor et veldig viktig tema. I von Bogungen mfl. (1995) og Wassmann mfl. (1991) blir det presentert data fra målinger av vertikal fluks og eksport av organisk materiale fra den fotiske sonen til bunnen på noen utvalgte lokaliteter i Norskehavet under perioden 1986 til 1993. Det finnes også en nyere publikasjon av Drits mfl. (2020) som presenterer data fra en stasjon i Lofotenbassenget i 2017-2018. Disse relativt grundige studiene viser relativt lik gjennomsnittlig årlig ny og gammel produksjon i overflatevannet i Norskehavet (øst i undersøkelsesområdet) sammenlignet med Grønlandshavet (vest i undersøkelsesområdet) men at det er store forskjeller i hvordan våroppblomstringen utvikler seg i de to havene. Dette får følgeeffekter både på den vertikale eksporten av partikulært organisk materiale fra den fotiske sonen (sammensetning og tidspunkt) men også på ny produksjon. I Norskehavet vil et stort beitetrykk forhindre akkumulasjon av store mengder planteplankton i den fotiske sonen, noe som gjør at det de fleste år ikke er blitt observert en tidlig og stor sedimentasjon av kiselalger om våren. Istedet blir kiselalgene nyttiggjort i den pelagiske sonen av planteplanktonbeitere. Man sier at i Norskehavet er den vertikale eksport av organisk materiale mot bunn kontrollert av biologiske prosesser og maksimal sedimentasjon ved bunn skjer som oftest under sen sommer og tidlig høst når beitetrykket har avtatt. I Lofotenbassenget ser man for eksempel en kraftig forhøyet sedimentasjon av organisk karbon i august og september bestående i hovedsak av fekaliepartikler og pteropoder, men også med innslag av planktoniske foraminiferer. Dette er i motsetning til situasjonen i Grønlandshavet hvor den vertikale eksport av organisk materiale mot bunn er kontrollert fremfor alt av hydrodynamiske prosesser som issmelting, pakkisens driftsmønster og vertikal omblanding. Der er det observert forhøyet sedimentasjon tidlig om våren, og etter våroppblomstringen vil det i grunnere områder forekomme aggregat av kiselalger på havbunn (Graf mfl. 1995). Dette materiale kan raskt omsettes av bunndyr. Også i grunnere områder av Norskehavet, som Vøringplatået på 1000-1500 m dyp, vil ferskt organisk materiale med et høyt innhold av klorofyll a, raskt kunne nå bunnen. Slike pulser med mat vil raskt bli utnyttet av de dyr som lever på og i sedimentene og mineraliseringsrater er derfor sterkt knyttet til den vertikale eksporten av organisk materiale. Kun 0.5% av sedimentert organisk materiale vil bli begravd i sedimentene (observasjoner fra 1000-1500 m dyp på Vøringplatået, Graf mfl. 1995). Materialet kan også være refraktært, i.e. brukt i flere omganger i den pelagiske miljø og derfor av lavere ernæringsmessig kvalitet. Dette ser ut til å være typisk for dypere havbunn, f.eks. på stasjon M (Bathmann mfl. 1987) og på 3000 m dyp i Lofotenbassenget (Graf mfl. 1995). Men også det refraktære organiske materiale vil bli utnyttet av bunndyr som lever i et miljø der tilgangen på mat jevnt over er lav, og dyrene som lever der kan derfor ofte ta opp næring fra et veldig vidt spekter; alt fra små organiske molekyler (DOC = dissolved organic carbon), bakterier, marin snø (døde planteplankton og zooplankton rester, fekalier, etc.) og levende zooplankton (se for eksempel Mueller mfl. 2014).
Figur 7.2 Plasser i Norskehavet hvor det er blitt målt sedimentasjon av organisk materiale til bunn. Kart publisert i von Bodungen mfl. 1995 (venstre), Wassmann mfl. 1991 (mitt) og Drits et al. 2020 (høyre).
Det vil også skje en omfordeling og horisontal transport av organisk materiale ved resuspensjon (Graf mfl. 1995, Drits mfl. 2020). Dette kan føre til lokalt høyere sedimentasjonsrater av organiske partikler på bunn enn det man finner høyere opp i vannsøylen (Drits mfl. 2020). Det er grunn til å tro at oseanografiske prosesser som leder til resuspensjon er av stor betydning for strukturering av bunndyrssamfunnene i De nordiske hav. For eksempel så har man funnet tydelige indikasjoner på at tette samfunn av filtrerende bunndyr som korall, svamp og sekkedyr ofte opptrer i slike resuspensjonsområder. Indre bølger drevet av tidevann kan fanges ved skrånende havbunn og lage turbulens nedover i vannsøylen og/eller virvle opp partikler med næringsrikt materiale fra bunn (se Hanz et al. 2020) og øke tilgangen på mat for filtrerende bunndyr. Slike oseanografiske prosesser ser ut å være viktig for fordelingen av tette samfunn av svamp og korall blant annet på Schultz banken og ved Mohn´s Treasure (Meyer et al. 2019, Ramirez-Llodra et al. 2020). Slike sammenhenger er også vist ved dype områder med tette samfunn av glassvamp Stillehavet (Pile og Young 2006).
7.3 - Oppsummering og kunnskapsbehov
Dagens kunnskap om koblinger mellom pelagialen og bentos i De nordiske hav, såkalt bentisk-pelagisk kobling, er svært mangelfull. Vi har innblikk i prosesser som er blitt identifisert som viktig for enkelttidspunkter, enkeltarter og enkeltlokaliteter, men ikke tilstrekkelig kunnskap om situasjonen i De nordiske hav i sin helhet. Det er ikke heller mulig å forutsi den fremtidige situasjonen for prosesser som regulerer den bentisk-pelagiske koblingen.
8 - Litteraturliste
8.1 - Fysisk oseanografi
Anderson M, Orvik KA, LaCasce JH, Koszalka I, Mauritsen C (2011) Variability of the Norwegian Atlantic Current and associated eddy field from surface drifters. J Geophys Res, Vol 16, C08032, doi: 10.1029/2011JC007078.
Bosse A, Fer I (2019) Mean structure and seasonality of the Norwegian Atlantic Front Current along the Mohn Ridge from repeated glider transects. Geophysical Research Letters, 46(22), 13170-13179.
Deser C, Walsh JE, Timlin MS (2000) Arctic Sea Icea Variability in the Context of Recent Atmospheric Circulation Trends. Journal of Climate 13:3, 617-633.
Fossum P, Mork KA, Tverberg V, Boitsov S, Heldal HE, Bagøien E, Rønning J mfl. (2012) Beskrivelse av miljø og levende marine ressurser i havområdene ved Jan Mayen. Rapport fra Havforskningen, 16: 119 pp.
Hanssen-Bauer I, Førland EJ, Haddeland I, Hisdal H, Mayer S, Nesje A, ... Ådlandsvik B (2015) Klima i Norge 2100 Kunnskapsgrunnlag for klimatilpasning oppdatert i 2015. NCCS report, NCCS, Oslo, Norway, 203.
Hanz U, Roberts EM, Duineveld G, Davies A, van Haren H, Rapp HT, ...Mienis F (2021) Long‐term Observations Reveal Environmental Conditions and Food Supply Mechanisms at an Arctic Deep‐Sea Sponge Ground. Journal of Geophysical Research: Oceans, 126(3), e2020JC016776.
Helland-Hansen B, Nansen F (1909) The Norwegian Sea: its physical oceanography based upon the Norwegian researches 1900-1904. Det Mallingske Bogtrykkeri.
Gabrielsen GW, Bekke B, Alsos IG, Hansen JR (1997) Natur og kulturmiljø på Jan Mayen - med en vurdering av verneverdi, kunnskapsbehov og forvaltning. Meddelelser nr 144. Norsk Polarinstitutt. 129 pp.
Koszalka I, LaCasce J H, Andersson M, Orvik KA, Mauritzen C (2011) Surface circulation in the Nordic Seas from clustered drifters. Deep Sea Research Part I: Oceanographic Research Papers, 58(4), 468-485.
Kvingedal B (2005) Sea-Ice Extent and Variability in the Nordic Seas, 1967-2002. I: Drange, H., Dokken, T., Furevik, T., Gerdes, R. & Berger, W. (red.) b. 158 The Nordic Seas: An Integrated Perspective, Oceanography, Climatology, Biogeochemistry, and Modeling, s. 39-50, American Geophysical Uniom, Washington, DC, USA.
Mork KA, Drinkwater KF, Jónsson S, Valdimarsson H, Ostrowski M (2014) Water mass exchanges between the Norwegian and Iceland seas over the Jan Mayen Ridge using in-situ current measurements. Journal of Marine Systems, 139, 227-240.
Naveira Garabato AC, Oliver KI, Watson AJ, Messias MJ (2004) Turbulent diapycnal mixing in the Nordic seas. Journal of Geophysical Research: Oceans, 109(C12).
Nilsen JEØ, Falck E (2006) Variations of mixed layer properties in the Norwegian Sea for the period 1948–1999. Progress in Oceanography, 70(1), 58-90.
Onarheim IH, Eldevik T, Smedsrud LH, Stroeve J C (2018) Seasonal and regional manifestation of Arctic sea ice loss. Journal of Climate, 31(12), 4917-4932.
Orvik KA, Niiler PP (2002) Major pathways of the Atlantic water in the northern North Atlantic and Nordic Seas toward Arctic, Geophys. Res. Lett., 29(19), 1896, doi:10.1029/2002GL015002
Padman L, Erofeeva S (2004) A barotropic inverse tidal model for the Arctic Ocean. Geophysical Research Letters, 31(2).
Portner HO, Roberts DC, Mason-Delmotte V, Zhai P, Tognor M... Weyer N (2019) IPCC (2019) IPCC Special Report on the Ocean and Cryosphere in a Changing Climate.
Raj RP, Johannessen JA, Eldevik T, Nilsen JØ, Halo I (2016) Quantifying mesoscale eddies in the Lofoten Basin. Journal of Geophysical Research: Oceans, 121(7), 4503-4521.
Roberts EM, Mienis F, Rapp HT, Hanz U, Meyer HK, Davies AJ (2018) Oceanographic setting and short-timescale environmental variability at an Arctic seamount sponge ground. Deep-Sea Research Part I 138: 98-113.
Stensland A, Baumberger T, Mork KA, Lilley MD, Thorseth IH, Pedersen RB (2019) 3He along the ultraslow spreading AMOR in the Norwegian-Greenland Seas. Deep Sea Research Part I: Oceanographic Research Papers, 147, 1-11.
Søiland H, Rossby T (2013) On the structure of the Lofoten Basin Eddy. Journal of Geophysical Research: Oceans, 118(9), 4201-4212.
van Aken HM, Budeus G, Hähnel M (1995) The anatomy of the Atctic frontal zone in the Greenland Sea. Journal of Geophysical Research 100, 15,999-16,014.
Voet G, Quadfasel D, Mork KA, Søiland H (2010) The mid-depth circulation of the Nordic Seas derived from profiling float observations. Tellus A: Dynamic Meteorology and Oceanography, 62(4), 516-529.
8.2 - Kjemisk oseanografi
Chierici M, Skjelvan I, Skotte G, Borg Pedersen ÅA (2020) Havforsuring i Norskehavet-rapport fra Miljødirektoratet og Overvåkingsgruppen (HI), www. miljodirektoratet.no
Chierici M, Vernet M, Fransson A, Børsheim KY (2019) Net community production and carbon exchange from winter to summer in the Atlantic water inflow to the Arctic Ocean, Frontiers in Marine Science, section Global Change and the Future Ocean, doi: 10.3389/fmars.2019.00528
Chierici M, Skjelvan I, Norli M, Lødemel HH, Lunde LF, Børsheim KY, Sørensen K, Yakushev E, Renner A, Kutti T, Bellerby R, King AL, Lauvset KS, Johannessen R (2016) Overvåking av havforsuring i norske farvann i 2015, Rapport, Miljødirektoratet, M-354|2016.
Chierici M, Skjelvan I, Bellerby R, NorliM, Lunde Fonnes L, Lødemel HH, Børsheim KY, Lauvset SK, Johannessen T, Sørensen K, Yakushev E (2014) Overvåking av havsførsuring i norske farvann, Rapport, Miljødirektoratet, 218-2014.
Chierici M, Drange H, Anderson LG, Johannessen T (1999) Inorganic Carbon Fluxes through the boundaries of the Greenland Sea Basin based on in situ observations and water transport estimates. Journal of Marine Systems, 22, 295-309.
Dickson AG, Sabine CL Christian JR (2007) Guide to Best Practices for Ocean CO2 Measurements. PICES Special Publication 3, 191 pp
Jones E, Chierici M, Skjelvan I, Norli M, Frigstad H, Børsheim KY, Lødemel HH, Kutti T, King AL, Sørensen K, Lauvset SK, Jackson-Misje K, Apelthun LB, de Lange T, Johannsessen T, Mourgues C, Bellerby R (2020) Monitoring ocean acidification in Norwegian seas in 2019, Rapport, Miljødirektoratet, M-1735|2020.
Olafsson J, Olafsdottir SR, Takahashi T, Danielsen M, Arnarson TS (2021). Enhancement of the North Atlantic CO2 sink by Arctic waters. Biogeosciences, 18, 1689–1701, https://doi.org/10.5194/bg-18-1689-2021
Olsen A, Omar AM, Jeansson E, Anderson LG, Bellerby RGJ (2010) Nordic seas transit time distributions and anthropogenic CO 2 . Journal of Geophysical Research: Oceans. C05005.
Olsen A, Key RM, van Heuven S, Lauvset SK, Velo A, Lin X, Schirnick C, Kozyr A, Tanhua T, Hoppema M, Jutterström S, Steinfeldt R, Jeansson E, Ishii M, Pérez FF, Suzuki T (2016) The Global Ocean Data Analysis Project version 2 (GLODAPv2) – an internally consistent data product for the world ocean. Earth System Science Data 8, 297–323.
Pierrot D, Lewis E, Wallace D (2006). MS Excel program developed for CO2 system calculations. ORNL/CDIAC-105a. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, US Department of Energy, Oak Ridge, Tennessee 10.
Portner HO, Roberts DC, Mason-Delmotte V, Zhai P, Tognor M... Weyer N (2019) IPCC (2019) IPCC Special Report on the Ocean and Cryosphere in a Changing Climate.
Sabine C, Feely R, Gruber N, Key R (2004) The Oceanic Sink for Anthropogenic CO2. Science, 305(5682):367-7, DOI:10.1126/science.1097403
Skjelvan I, Jeansson E, Chierici M, Omar A, Olsen A, Lauvset S, Johannessen T (2014) Havforsuring og opptak av antropogent karbon i de Nordiske hav, 1981-2013, Rapport, Miljødirektoratet, M244-2014.
8.3 - Plankton
Aarbakke ONS (2007) Bio-acoustic investigations of sound scattering layers overlaying hydrothermal vents in the Arctic Ocean. Master of Science thesis in Marine Biology, University of Bergen, Department of Biology, 2007. Pp. i-viii, + pp. 1-97.
Aksnes DL, Blindheim J (1996) Circulation patterns in the North Atlantic and possible impact on population dynamics of Calanus finmarchicus. Ophelia 44: 7-28.
Azam F, Fenchel T, Field JG, Gray JS, Meyerreil, LA, Thingstad F (1983) The Ecological Role Of Water-Column Microbes In The Sea. Marine Ecology Progress Series 10: 257-263.
Batten SD, Clark R, Flinkman J, Hays GC, John E, John AWG, Jonas T, Lindley JA, Stevens DP, Walne A (2003) CPR sampling: the technical background, materials and methods, consistency and comparability. Prog. Oceanogr 58: 193–215.
Beaugrand G, Reid PC, Ibañez F, Lindley JA, Edwards M (2002) Reorganization of North Atlantic Marine Copepod Biodiversity and Climate. Science 296: 1692-1694.
Behrenfeld MJ, Falkowski PG (1997) Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnology and Oceanography, 42: 1-20.
Berge G (1958). The primary production in the Norwegian Sea in June 1954, measured by an adapted 14C technique. - Rapp. Cons. Explor. Mer 144: 85-91.
Berge J, Nahrgang J (2013) The Atlantic spiny lumpsucker Eumicrotremus spinosus: life history traits and the seemingly unlikely interaction with the pelagic amphipod Themisto libellula. Polish Polar Research 34(3): 279-287. doi: 10.2478/popore-2013-0013.
Bergh Ø, Børsheim KY, Bratbak G & Heldal M (1989) High abundance of viruses found in aquatic environments. Nature 340: 467-468.
Berkes F (1973) Production and comparative ecology of euphausiids in the Gulf of St. Lawrence. PhD. Thesis, Marine Sciences Cent. McGill University, Montreal. 188 p.
Bindoff NL, Cheung WWL, Kairo JG, Arístegui J, Guinder VA, Hallberg R, Hilmi NJM, Jiao N, Karim Md S, Levin L, O’Donoghue S, Purca Cuicapusa SR, Rinkevich B, Suga T, Tagliabue A & Williamson P (2019) Changing Ocean, Marine Ecosystems, and Dependent Communities. In Pörtner H-O, Roberts DC, Masson-Delmotte V, Zhai P, Tignor M, Poloczanska E, et al. (Eds.) IPCC Special Report on the Ocean and Cryosphere in a Changing Climate. Intergovernmental Panel on Climate Change, Switzerland, pp. 477-587.
Bjørke H, Gjøsæter H (1998) Cephalopods in the Norwegian Sea. I: The Norwegian Sea Ecosystem (Ed. HR Skjoldal). pp 371-394.
Bonnet D, Richardson A, Harris R, Hirst A, Beaugrand G, Edwards M, Ceballos S, Diekman, R, Lopez-Urrutia A, Valdes L, Carlotti F, Molinero JC, Weikert H, Greve W, Luci D, Albaina A, Yahia ND, Umani SF, Miranda A, dos Santos A, Cook K, Robinson S, de Puelles MLF (2005) An overview of Calanus helgolandicus ecology in European waters. Progress in Oceanography 65: 1–53.
Boyd PW, Claustre H, Levy M, Siegel DA, Weber T (2019) Multi-faceted particle pumps drive carbon sequestration in the ocean. Nature 568: 327-335.
Broms C, Melle W (2007) Seasonal development of Calanus finmarchicus in relation to phytoplankton bloom dynamics in the Norwegian Sea. Deep-Sea Research II 54: 2760–2775.
Broms C, Melle W, Kaartvedt S (2009) Oceanic distribution and life cycle of Calanus species in the Norwegian Sea and adjacent waters. Deep-Sea Research II 56: 1910–1921.
Burd BJ, Thomson RE (1994) Hydrothermal venting at Endeavour Ridge: Effect on zooplankton biomass throughout the water column. Deep-Sea Research I 41:140723.
Burd BJ, Thomson RE (1995) Distribution of zooplankton associated with the Endeavour Ridge hydrothermal plume. Journal of Plankton Research 17: 96597.
Burd BJ, Thomson RE, Calvert SE (2002). Isotopic composition of hydrothermal epiplume zooplankton: Evidence of enhanced carbon recycling in the water column. Deep-Sea Research I 49:1877900.
Børsheim KY (2000) Bacterial production rates and concentrations of organic carbon at the end of the growing season in the Greenland Sea. Aquatic Microbial Ecology 21: 115-123.
Børsheim KY (2017) Bacterial and primary production in the Greenland Sea. Journal of Marine Systems 176: 54-63.
Børsheim KY, Milutinovic S, Drinkwater KF (2014) TOC and satellite-sensed chlorophyll and primary production at the Arctic Front in the Nordic Seas. Journal of Marine Systems 139: 373-382.
Conover RJ (1988) Comparative life histories in the genera Calanus and Neocalanus in high latitudes of the northern hemisphere. Hydrobiologia 167/168: 127-142.
Dale T, Bagøien E, Melle W, Kaartvedt S (1999a) Can predator avoidance explain varying overwintering depth of Calanus in different oceanic water masses? Marine Ecology Progress Series 179: 113–121.
Dale T, Rey F, Heimdal BR (1999b) Seasonal development of phytoplankton at a high latitude oceanic site. Sarsia 84: 419-435.
Dalpadado P, Ellertsen B, Melle W, Skjoldal HR (1998) Summer distribution patterns and biomass estimates of macrozooplankton and micronekton in the Nordic seas. Sarsia, 83: 103-116.
Dalpadado P, Ellertsen B, Melle W, Dommasnes A (2000) Food and feeding conditions of Norwegian spring spawning herring Clupea harengus through its feeding migration. ICES Journal of Marine Science 57: 843–857.
Davison PC, Checkley Jr DM, Koslow JA, Barlow J (2013) Carbon export mediated by mesopelagic fishes in the northeast Pacific Ocean. Prog. Oceanogr. 116: 14–30.
Diel S, Tande K (1992) Does the spawning of Calanus finmarchicus in high latitudes follow a reproducible pattern? Marine Biology, 113: 21–31.
Doall MH, Colin SP, Strickler JR, Yen J (1998). Locating a mate in 3D: the case of Temora longicornis. Phil. Trans. R. Soc. Lond. B 353: 681–689.
Dommasnes A, Melle W, Dalpadado P, Ellertsen B (2004) Herring as a major consumer in the Norwegian Sea. ICES Journal of Marine Science 6: 739–751.
Falkenhaug T, Hobæk A, Willassen E (2016) Inventory of marine Copepoda and Cladocera (Crustacea) in Norway: COPCLAD. Poster presentation at the Scientific Session "Zooplankton diversity in the oceans by integrative morphological and molecular techniques", ICES/PICES 6th Zooplankton Production Symposium, 9-13 May 2016, Bergen.
Falkenhaug T, Hobæk A (2021) COPCLAD_Marine Copepoda and Cladocera in Norway_ Dataset - DS-TF100221_10Feb2021. dx.doi.org/10.5883/DS-TF100221.
Fields DM, Shaeffer DS, Weissburg MJ (2002) Mechanical and neural responses from the mechanosensory hairs on the antennule of Gaussia princeps. Marine Ecology Progress Series 227: 173–186.
Fossum P, Mork KA, Tverberg V, Boitsov S, Heldal HE, Bagøien E, Rønning J, Slotte A, Vollen T, Wienerroither R, Dahle G, Buhl-Mortensen P, Øien N, Øigård TA, Fauchald P (2012) Beskrivelse av miljø og levende marine ressurser i havområdene ved Jan Mayen. Rapport fra Havforskningen Nr 16-2012. Pp. 1-119.
Fuentes V, Alurralde G, Meyer B, mfl. (2016) Glacial melting: an overlooked threat to Antarctic krill. Sci. Rep. 6. doi:10/f8prwb
Gadd, GM (2010) Metals, minerals and microbes: geomicrobiology and bioremediation. Microbiology-Sgm, 156: 609-643.
Halldal P (1953) Phytoplankton investigations from weathership M in the Norwegian Sea, 1948–1949. Det Norske Videnskaps Akademi. Hvalrådets skrifter 38: 1–91.
Halvorsen E, Tande KS, Edvardsen A, Slagstad D, Pedersen OP (2003) Habitat selection of overwintering Calanus finmarchicus in the NE Norwegian Sea and shelf waters off Northern Norway in 2000-02. Fisheries Oceanography 12: 339-351.
Hassel A, Endresen B, Martinussen MB, Gjertsen K, Knutsen T, Johannessen ME (2020) Håndbok for forskningsgruppe Plankton. Prøvetaking og analyse. Prosedyrer for prøvetaking og pre-analyse av dyre- og planteplankton på forskningsfartøy og i laboratorium på land. Versjon 6.0 (desember 2020), Havforskningsinstituttet.
Havermans C, Auel H, Hagen W, Held C, Ensor NS, Tarling GA (2019) Chapter Two: Predatory zooplankton on the move: Themisto amphipods in high-latitude marine pelagic food webs. Advances in Marine Biology, 82, pp. 51-92. https://doi.org/10.1016/bs.amb.2019.02.002.
Hensen V (1887) Ueber die Bestimmung des Planktons oder des im Meere treibende Materials an Pflanzen und Thieren. Ber. Komm. Wiss. Unters. Dt. Meere 5: 1-109
Herndl GJ, Agogue H, Baltar F, Reinthaler T, Sintes E, Varela MM (2008) Regulation of aquatic microbial processes: the 'microbial loop' of the sunlit surface waters and the dark ocean dissected. Aquatic Microbial Ecology 53(1): 59-68
Hirai J, Jones DOB (2012) The temporal and spatial distribution of krill (Meganyctiphanes norvegica) at the deep seabed of the Faroe–Shetland Channel, UK: A potential mechanism for rapid carbon flux to deep sea communities, Marine Biology Research, 8:1, 48-60, DOI: 10.1080/17451000.2011.594891.
Hirche H-J (1991) Distribution of dominant calanoid copepod species in the Greenland Sea during late fall. Polar Biology, 11: 251-362.
Hirche H-J (1996) Diapause in the marien copepod, Calanus finmarchicus - a review. Ophelia 44: 129-143.
Hirche H-J (1997) Life cycle of the copepod Calanus hyperboreus in the Greenland Sea. Marine Biology 128: 607–618.
Hjøllo SS, Huse G, Skogen MD, Melle W (2012) Modelling secondary production in the Norwegian Sea with a fully coupled physical/primary production/individual-based Calanus finmarchicus model system. Marine Biology Research 8: 508-526.
Ingvaldsen RB, Bucklin A, Fauchald P, Gjøsæter H, Haug T, Jørgensen LL, Knutsen T, Naustvoll LJ, Wiebe P (2016) Cruise report SI_ARCTIC/Arctic Ecosystem survey R/V Helmer Hanssen, 19 August-7 September 2014. Toktrapport/Havforsknings-instituttet/ISSN 1503 6294/Nr. 11-2016. Pp. 1-53, + Tables A1-A8.
ICES (2020) Working Group on the Integrated Assessments of the Norwegian Sea (WGINOR; outputs from 2019 meeting). ICES Scientific Reports. 2:29. 46 pp. http://doi.org/10.17895/ices.pub.5996.
Jaschnov WA (1970) Distribution of Calanus species in the seas of the Northern Hemisphere. International Revue der gesamten Hydrobiologie und Hydrographie 55: 179-212.
Jonasdottir SH, Visser AW, Richardson K, Heath MR (2015) Seasonal copepod lipid pump promotes carbon sequestration in the deep North Atlantic. Proceedings of the National Academy of Sciences of the United States of America, 112: 12122-12126.
Jørgensen SL, Hannisdal B, Lanzen A, Baumberger T, Flesland K, Fonseca R, Øvreås L, Steen IH, Thorseth IH, Pedersen RB, Schleper C (2012) Correlating microbial community profiles with geochemical data in highly stratified sediments from the Arctic Mid-Ocean Ridge. Proceedings of the National Academy of Sciences of the United States of America, 109, E2846-E2855.
Kaartvedt S (2008) Photoperiod may constrain the effect of global warming in arctic marine systems. J. Plankton Res. 30: 1203–1206. doi:10.1093/plankt/fbn075
Kjesbu OS, Sundby S, Sandø AB, Alix M, Tiedemann M, mfl. (Manuskript innsendt til PNAS) Highly mixed impacts of near-future climate change on stock productivity in the North East Atlantic.
Kiørboe T (2011) What makes copepods so successful? Journal of Plankton Research 33: 677-685.
Kiørboe T, Bagøien E (2005) Motility patterns and mate encounter rates in planktonic copepods. Limnology & Oceanography 50:1999-2007.
Klevjer T, Melle W, Knutsen T, Strand E, Korneliussen R, Dupont N, Salvanes AGV, Wiebe PH (2019) Micronekton biomass distribution, improved estimates across four north Atlantic basins. Deep Sea Res. Part II Top. Stud. Oceanogr. 104691. doi:10/ggpws7
Klevjer T, Melle W, Knutsen T, Aksnes DL (2020) Vertical distribution and migration of mesopelagic scatterers in four north Atlantic basins. Deep-Sea Research Part II: Topical Studies in Oceanography 180
Knutsen T, Serigstad B (2001) Potential implications on the pelagic fish and zooplankton community of artificially induced deep-water releases of oil and gas during DeepSpill_2000— an innovative acoustic approach. Fisken. Havet. 14: 1-37. http://hdl.handle.net/11250/113901.
Langøy H, Nøttestad L, Skaret G, Broms C, Fernö A (2012) Overlap in distribution and diets of Atlantic mackerel (Scomber scombrus), Norwegian spring-spawning herring (Clupea harengus) and blue whiting (Micromesistius poutassou) in the Norwegian Sea during late summer. Marine Biology Research 8: 442–460.
Lass S, Tarling GA, Virtue P, Matthews JLB, Mayzaud P, Buchholz F (2001) On the food of northern krill Meganyctiphanes norvegica in relation to its vertical distribution. Mar Ecol Prog Ser 214: 177-200. DOI: 10.3354/meps214177.
Lotze HK, Tittensor DP, Bryndum-Buchholz A, Eddy TD, Cheung WWL, Galbraith ED, Barange M, Barrier N, Bianchi D, Blanchard JL, Bopp L, Büchner M, Bulman CM, Carozza DA, Christensen V, Coll M, Dunne JP, Fulton EA, Jennings S, Jones MC, Mackinson S, Maury O, Niiranen S, Oliveros-Ramos R, Roy T, Fernandes JA, Schewe J, Shin Y-J, Silva TAM, Steenbeek J, StockCA, Verley P, Volkholz J, Walker ND, Worm B (2019) Global ensemble projections reveal trophic amplification of ocean biomass declines with climate change. Proceedings of the National Academy of Sciences of the USA 116: 12907-12912.
Macnaughton MO, Thormar J, Berge J (2007) Sympagic amphipods in the arctic pack ice: redescriptions of Eusirus holmii Hansen, 1887 and Pleusymtes karstensi (Barnard, 1959). Polar Biology 30 (8): 1013-1025.
MacDonald R (1927) Food and habits of Meganyctiphanes norvegica. J. mar. biol. Ass. U.K. 14: 753784.
Mauchline J (1960) The biology of the Euphausiid Crustacean Meganyctiphanes norvegica (M. Sars). Proc. R. Soc. Edinb. B. 67: 141179.
Mauchline J, Fisher LR (1969) The biology of euphausiids. Adv. mar. Biol. 7, 1454.
Mauchline J (1980) The biology of mysids and euphausiids. Adv. mar. Biol. 18, 1681.
Mecklenburg C, Lynghammar A, Johanesen E, Byrkjedal I, Dolgov AV, Kaamushko OV, Meckelburg TA, Møller PR, Steinke D. Weinerroither RM, Christiansen JS (2018) Marine Fishes of the Arctic Region. CAFF Monitoring Series Report 28.
Melle W, Ellertsen B, Skjoldal HR (2004) Zooplankton: The link to higher trophic levels. In: Skjoldal HR (ed.), The Norwegian Sea Ecosystem. Tapir Academic Press, Trondheim, pp. 137-202.
Melle W, Runge J, Head E, Plourde S, Castellani C, Licandro P, Pierson J, Jonasdottir S, Johnson C, Broms C, Debes H, Falkenhaug T, Gaard E, Gislason A, Heath M, Niehoff B, Nielsen TG, Pepin P, Stenevik EK, Chust G (2014) The North Atlantic Ocean as habitat for Calanus finmarchicus: Environmental factors and life history traits. Progress in Oceanography, 129: 244-284.
Molinos JG, Halpern BS, Schoeman DS, Brown CJ, Kiessling W, Moore PJ, Pandolfi JM, Poloczanska ES, Richardson AJ, Burrows MT (2016) Climate velocity and the future global redistribution of marine biodiversity. Nature Climate Change 6: 83-88
Møller EF, Nielsen TG, Richardson K (2006) The zooplankton community in the Greenland Sea: Composition and role in carbon turnover. Deep-Sea Research Part I-Oceanographic Research Papers, 53: 76-93.
Müller O, Wilson B, Paulsen ML, Ruminska S, Armo HR, Bratbak G, Øvreas L (2018) Spatiotemporal Dynamics of Ammonia-Oxidizing Thaumarchaeota in Distinct Arctic Water Masses. Frontiers in Microbiology, 9:24.
Naustvoll LJ, Melle W, Klevjer T, Drinkwater KF, Strand E, Knutsen T (2020) Dynamics of phytoplankton species composition, biomass and nutrients in the North Atlantic during spring and summer - A trans-Atlantic study. Deep-Sea Research Part I - Topical Studies in Oceanography, 180: 104890.
Niehoff B, Hirche H-J (2000) The reproduction of Calanus finmarchicus in the Norwegian Sea in spring. Sarsia 85: 15–22.
Niehoff B, Klenke U, Hirche H-J, Irigoien X, Head R, Harris R (1999) A high frequency time series at Weathership M, Norwegian Sea, during the 1997 spring bloom: the reproductive biology. Marine Ecology
Progress Series 176: 81–92.
Niemann H, Lösekann T, de Beer D mfl. (2006) Novel microbial communities of the Haakon Mosby mud volcano and their role as a methane sink. Nature 443, 854-858.
Norheim E, Klevjer T, Aksnes N (2016) Evidence for light-controlled migration amplitude of a sound scattering layer in the Norwegian Sea. Mar. Ecol. Prog. Ser. 551: 45–52. doi:10.3354/meps11731
Olsen RE, Knutsen T, Melle W (2006) Nye marine fórressurser (NYMAR). Internrapport, Havforskningsinstituttet, 74 pp.
Olsen BR, Troedsson C, Hadziavdic K, Pedersen RB, Rapp HT (2014) A molecular gut content study of Themisto abyssorum (Amphipoda) from Arctic hydrothermal vent and cold seep systems. Molecular Ecology 23: 3877–3889. doi: 10.1111/mec.12511
Olsen BR, Troedsson C., Hadziavdic K, Pedersen RB, Rapp HT (2015) The influence of vent systems on pelagic eukaryotic micro-organism composition in the Nordic Seas. Polar Biol 38: 547–558. doi 10.1007/s00300-014-1621-8.
Paasche E (1960) Phytoplankton distribution in the Norwegian Sea in June, 1954, related to hydrography and compared with primary production data. Fiskeridirektoratets Skrifter Serie Havundersøkelser 12, 1–77.
Paffenhöfer GA, Strickler JR, Alcaraz M (1982) Suspension-feeding by herbivorous calanoid copepods. A cinematographic study. Marine Biology 67: 193–199.
Pakhomov EA, Froneman PW (1999) Macroplankton/micronekton dynamics in the vicinity of the Prince Edward islands (Southern Ocean). Mar. Biol. 134: 501–515.
Pedersen RB, Thorseth IH, Hellevang B mfl. (2005) Two vent fields discovered at the ultraslow spreading Arctic ridge system. Eos Transactions American Geophysical Union, 86. Fall Meeting Supplement. Abstract OS21C-01.
Pedersen RB, Rapp HT, Thorseth IH mfl. (2010a) Discovery of a black smoker vent field and vent fauna at the Arctic Mid-Ocean Ridge. Nature Communications, 1, 126.
Pedersen RB, Thorseth IH, Nygard TE, Lilley MD, Kelly DS (2010b) Hydrothermal activity at the Arctic mid-ocean ridges. In: Diversity of Hydrothermal Systems on Slow Spreading Ocean Ridges, Geophysical Monograph Series, vol. 188 (eds Rona PA, Devey CW, Dyment J, Murton BJ), pp. 67–89. AGU, Washington, District of Columbia.
Poloczanska ES, Burrows MT, Brown CJ, Molinos JG, Halpern BS, Hoegh-Guldberg O, Kappel CV, Moore PJ, Richardson AJ, Schoeman DS, Sydeman WJ (2016). Responses of Marine Organisms to Climate Change across Oceans. Frontiers in Marine Science 3.
Rey F, Noji T, Miller LA (2000) Seasonal phytoplankton development and new production in the central Greenland Sea. Sarsia, 85: 329-344.
Richardson AJ, Walne AW, John AWG, Jonas TD, Lindley JA, Sims DW, Stevens D, Witt M (2006) Using continuous plankton recorder data. Progress in Oceanography 68: 27–74.
Sameoto DD (1980) Quantitative Measurements of Euphausiids Using a 120-kHz Sounder and Their in situ Orientation. Canadian Journal of Fisheries and Aquatic Sciences, 37: 693-702.
Schander C, Rapp HT, Kongsrud JA, Bakken T, Berge J, Cochrane S, Oug E, Byrkjedal I, Cedhagen T, Fosshagen A, Gebruk A, Larsen K, Nygren A, Obst M, Pleijel F, Stöhr S, Todt C, Warén A, Handler-Jacobsen S, Kuening R, Levin L, Mikkelsen NT, Petersen KK, Thorseth IH, Pedersen RB (2010) The fauna of the hydrothermal vents on the Mohn Ridge (North Atlantic). Mar Biol Res 6(2): 155–171.
Schmidt K (2010) Food and Feeding in Northern Krill (Meganyctiphanes norvegica Sars). Advances in marine biology, 57. 127-71. 10.1016/B978-0-12-381308-4.00005-4.
Schmoker C, Hernández-León S, Calbet A (2013) Microzooplankton grazing in the oceans: impacts, data variability, knowledge gaps and future directions. Journal of Plankton Research 34: 691–706
Semenova TN (1974) Diurnal vertical migration of Parathemisto japonica Bov. (Hyperiidea) in the Sea of Japan. Oceanology 14: 272–276.
Sieburth J McN, Smetacek V, Lenz J (1978) Pelagic Ecosystem Structure: Heterotrophic Compartments of the Plankton and Their Relationship to Plankton Size Fractions. Limnology and Oceanography, 23(6.): 1256-1263.
Skjoldal HR, Dalpadado P, Dommasnes A (2004) Food webs and trophic interactions. In: Skjoldal, HR (ed), The Norwegian Sea Ecosystem. Tapir Academic Press, Trondheim, pp. 447-506.
Skreslet S (2004) Jan Mayen Island ecology. Its relation to the Arctic Mediterranean Ecosystem. In: Skreslet S (ed.), Jan Mayen Island in scientific focus. NATO Science Series, IV, Earth and Environmental Sciences – Volume 45. Kluwer Academic Publishers, pp. 101-112.
Stenevik EK, Melle W, Gaard E, Gislason A, Broms CTÅ, Prokopchup I, Ellertsen B (2007) Egg production of Calanus finmarchicus – a basin-scale study. Deep-Sea Research Part II 54: 2672–2685.
Storesund JE, Øvreas L (2013) Diversity of Planctomycetes in iron-hydroxide deposits from the Arctic Mid Ocean Ridge (AMOR) and description of Bythopirellula goksoyri gen. nov., sp nov., a novel Planctomycete from deep sea iron-hydroxide deposits. Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology 104: 569-584.
Strand E, Bagøien E, Edwards M, Broms C, Klevjer T (2020 A) Spatial distributions and seasonality of four Calanus species in the Northeast Atlantic. Progress in Oceanography. 185, 102344.
Strand E, Klevjer T, Knutsen T, Melle W (2020 B) Ecology of mesozooplankton across four North Atlantic Basins. Deep-Sea research Part II, 180: 104844
Strickler JR (1985) Feeding currents in calanoid copepods: two new hypotheses. In. Laverack, M. S. (ed.) Physiological adaptations of marine animals. Symp. Soc. Experimental Biology 89: 459–485.
Sundby S (2000) Recruitment to Atlantic cod stocks in relation to temperature and advection of copepod populations. Sarsia 85: 277–298.
Suttle CA (2005) Viruses in the sea. Nature 437: 356-361.
Sweetman A, Levin LL, Rapp HT, Schander C (2013) Deceased Faunal trophic structure at hydrothermal vents on the southern Mohn’s Ridge, Arctic Ocean. Marine Ecology Progress Series, 473: 115-131.
Tandberg AH, Rapp HT, Schander C, Vader W, Sweetman AK, Berge J (2011) Exitomelita sigynae gen. et sp. nov.: a new amphipod from the Arctic Loki Castle vent field with potential gill ectosymbionts. Polar Biology, 35: 705–716.
Tarling GA, Matthews JBL, Sabarowski R, Buchholz F. (1998) Vertical migratory behaviour of the euphausiids, Meganyctiphanes norvegica, and its dispersion in the Kattegat Channel. Hydrobiologia 375/376: 331-41.
Thomson RE, Burd BJ, Dolling AG, Lee Gordons R, Jamieson GS (1992) The deep scattering layer associated with the endeavour ridge hydrothermal plume. Deep Sea Research Part A. Oceanographic Research Papers 39 (1): 55-73.
Tsuda A, Miller CB (1998) Mate-finding behaviour in Calanus marshallae Frost. Phil Trans R Soc Lond B, 353: 713–720.
Turner JT (2015) Zooplankton fecal pellets, marine snow, phytodetritus and the ocean’s biological pump. Prog. Oceanogr. 130: 205–248. doi:10.1016/j.pocean.2014.08.005
UNESCO (1968) Standardization of zooplankton sampling methods at sea. In Zooplankton Sampling, Part II. Monographs on Oceanographic Methodology 2. Unesco Press, UNESCO, Paris, pp. 147-168.
Uric T, Lanzen A, Stokke R, Pedersen RB, Bayer C, Thorseth IH, Schleper C, Steen IH, Øvrea, L (2014) Microbial community structure and functioning in marine sediments associated with diffuse hydrothermal venting assessed by integrated meta-omics. Environmental Microbiology 16: 2699-2710.
Wenneck TdL, Falkenhaug T, Bergstad OA (2008) Strategies, methods, and technologies adopted on the R.V. G.O. Sars MAR-ECO expedition to the Mid-Atlantic Ridge in 2004. Deep Sea Res. II, 55: 6–28. doi: 10.1016/j.dsr2.2007.09.017
Weisshappel JB (2000) Distribution and diversity of the hyperbenthic amphipod family Eusiridae in the different seas around the Greenland-Iceland-Faeroe-Ridge. Sarsia 85: 227-236.
Wiborg KF (1954) Investigations on zooplankton in coastal and offshore waters of western and northwestern Norway. Fiskeridirektoratets Skrifter Serie Havundersøkelser 11: 1-246.
Wiborg KF (1955) Zooplankton in relation to hydrography in the Norwegian Sea. Fiskeridirektoratets Skrifter Serie Havundersøkelser 11: 1-66.
Wiebe PH, Burt KH, Boyd SH, Morton AW (1976) A multiple opening/closing net and environmental sensing system for sampling zooplankton. Journal of Marine Research 34: 313-326.
Wiebe PH, Copley N, van Dover C, Tamse, A., Manrique F (1988) Deep-water zooplankton of the Guayamas Basin hydrothermal vent field. Deep-Sea Research 35: 9851013.
Wiebe PH, Morton AW, Bradley AM, Backus RH, Craddock JE, Barber V, Cowles TJ, Flierl GR (1985) New development in the MOCNESS, an apparatus for sampling zooplankton and micronekton. Marine Biology 87: 313-323.
Williams PJ leB, Thomas DN, CReynolds CS (2002) Phytoplankton Productivity: Carbon Assimilation in Marine and Freshwater Ecosystems. Blackwell Science Ltd pp. 386
Østvedt OJ (1955). Zooplankton investigations from weathership M in the Norwegian Sea, 1948-1949. Hvalrådets skrifter
8.4 - Fisk
Albert OT, Nilssen M, Stene A, Gundersen AC, Nedreaas KH (2001) Maturity classes and spawning behaviour of Greenland halibut (Reinhardtius hippoglossoides). Fisheries Research Vol. 51 (2-3): 217-228.
Astthorsson OS, Gislason A (1997) On the food of capelin in the subarctic waters north of Iceland. Sarsia 82(2): 81-86.
Bachiller E, Skaret G, Nøttestad L, Slotte A (2016) Feeding Ecology of Northeast Atlantic Mackerel, Norwegian Spring-Spawning Herring and Blue Whiting in the Norwegian Sea. PlosOne 11(2).
Bergstad OA, Bjelland O, Gordon, JDM (1999) Fish communities on the slope of the eastern Norwegian Sea. Sarsia 84: 67-78.
Bjelland O, Bergstad OA, Skjæraasen JE, Meland K (2000) Trophic ecology of deep-water fishes associated with the continental slope of the eastern Norwegian Sea. Sarsia 85: 101-117.
Bjelland O, Holst JC (2004) Other fish species and fish communities. In: Skjoldal HR (red.) The Norwegian Sea Ecosystem. Tapir Academic Press, Trondheim. 357-370.
Campana SE, Joyce W, Marks L, Hurley P, Natanson LJ, Kohler NE, Jensen CF, Mello JJ, Pratt HL, Myklevoll S, Harley S (2008) The rise and fall (again) of the porbeagle shark population in the Northwest Atlantic. In: Sharks of the Open Ocean: Biology, Fisheries & Conservation. Camhi, M.D., Pikitch, E.K. & Babcock, E.A. (eds.). Fish and Aquatic Resources Series 13. Singapore: Blackwell Publishing Ltd.
Dahl E, Laubier L, Sibuet M, Strömberg JO (1976) Some quantitative results on benthic communities of the deep Norwegian Sea. Astarte 9 (01-79): 279-297.
Fordham S, Fowler SL, Coelho R, Goldman KJ, Francis M (2006) Squalus acanthias. In: IUCN 2013. IUCN Red List of Threatened Species. Version 2013.2. Available via http://www.iucnredlist.org. Accessed 9 April 2014
Fossum P, Mork KA, Tverberg V, Boitsov S, Heldal HE, Bagøien E, Rønning J, Slotte A, Vollen T, Wienerroither R, Dahle G, Buhhl-Mortensen P, Øien N, Øigård TA, Fauchald P (2012) Beskrivelse av miljø og levende marine ressurser i havområdene ved Jan Mayen. Rapport fra Havforskningen Nr 16-2012. 119 pp.
Hallfreðsson EH, Primicerio R (2017) Deep-sea fish communities at the eastern slope of the Norwegian Sea. Conference presentation. ESSAS Tromssø 2017.
Haedrich RL, Merrett NR (1988) Summary atlas of deep-living demersal fishes in the North Atlantic Basin. Journal of Natural History 22:1325-1362.
Hawkins AD, Picciulin M (2019) The importance of underwater sounds to gadoid fishes. The Journal of the Acoustical Society of America 146(5): 3536-3551.
Henriksen S, Hilmo O (2015) Norsk rødliste for arter. Artsdatabanken, Norge
Holm M, Hansen LP, Holst JC, Jacobsen JA (2004) Atlantic salmon (Salmo salar L.). In: Skjoldal HR (red.) (2004) The Norwegian Sea Ecosystem. Tapir Academic Press, Trondheim. 559 pp.
ICES (2018) Report of the Working Group on Elasmobranch Fishes (WGEF), 19–28 June 2018, Lisbon, Portugal. ICES CM 2018/ACOM:16. 1306 pp.
Lynghammar A, Christiansen JS, Mecklenburg CW, Karamushko OV, Møller PR, Gallucci VF (2013) Species richness and distribution of chondrichthyan fishes in the Arctic Ocean and adjacent seas. Biodiversity 14(1): 57-66.
McPhie RP, Campana SE (2009) Reproductive characteristics and population decline of four species of skate (Rajidae) off the eastern coast of Canada. Journal of Fish Biology 75:223-246.
Mecklenburg CW, Lynghammar A, Johannesen E, Byrkjedal I, Christiansen JS, Dolgov AV, Karamushko OV, Mecklenburg TA, Møller PR, Steinke D, Wienerroither RM (2018a) Marine Fishes of the Arctic Region vol 1. CAFF Monitoring Series Report 28: 1-454.
Mecklenburg CW, Lynghammar A, Johannesen E, Byrkjedal I, Christiansen JS, Dolgov AV, Karamushko OV, Mecklenburg TA, Møller PR, Steinke D., Wienerroither RM (2018b) Marine Fishes of the Arctic Region vol 2. CAFF Monitoring Series Report 28: 455-731.
Michalsen K, Nedreaas, KH (1998) Food and feeding of Greenland halibut (Reinhardtius hippoglossoides, Walbaum) in the Barents Sea and East Greenland waters. Sarsia 83: 401-407.
Nielsen J, Hedeholm RB, Heinemeier J, Bushnell PG, Christiansen JS, Olsen J, Ramsey CB, Brill RW, Simon M, Steffensen KF, Steffensen JF (2016) Eye lens radiocarbon reveals centuries of longevity in the Greenland shark (Somniosus microcephalus). Science 353(6300): 702-704.
Olafsdottir D, MacKenzie BR, Chosson-P V, Ingimundardottir T (2016) Dietary Evidence of Mesopelagic and Pelagic Foraging by Atlantic Bluefin Tuna (Thunnus thynnus L.) during Autumn Migrations to the Iceland Basin. Frontiers in Marine Science 3(18).
Pianka ER (1970) On r-and K-selection. American Naturalist 104: 592–597.
Porteiro FM, Sutton T, Byrkjedal I, Orlov AM, Heino M, Menezes, G, Bergstad, OA (2017) Fishes of the Northern Mid-Atlantic Ridge Collected During the MAR-ECO Cruise in June-July 2004: An Annotated Checklist. Nova Southeastern University Oeanography Faculty Reports 1-1-2017. ISBN: 978-989-99834-0-3. 128 pp.
Skjoldal HR (red.) (2004) The Norwegian Sea Ecosystem. Tapir Academic Press, Trondheim. 559 pp.
Skjæraasen, JE, Bergstad, OA (2001) Notes on the distribution and length composition of Raja lintea, R. fyllae, R. hyperborea and Bathyraja spinicauda (Pisces: Rajidae) in the deep northeastern North Sea and on the slope of the eastern Norwegian Sea. ICES Journal of Marine Science 58: 21-28.
Stevens JDR, Bonfil R, Dulvy NK, Walker PA (2000) The effect of fishing on sharks, rays, and chimaeras (Chondrichthyans), and the implications for marine ecosystems. ICES Journal of Marine Science 57: 476-494.
Williams T (2007) Cartilaginous fishes along the North-Norwegian coast. Distributions and densities with regard to fishing and sea temperature. Master thesis in International Fisheries Management. Norwegian College of Fishery Science. University of Tromsø, 62 pp.
Ådlandsvik, B, Gundersen, AC, Nedreaas, KH, Stene, A, Albert, OT (2004) Modelling the advection and diffusion of eggs and larvae of Greenland halibut (Reinhardtius hippoglossoides) in the north-east Arctic Fisheries oceanograpgy 3: 403-415.
8.5 - Sjøpattedyr
Accardo CM, Ganley LC, Brown MW, Duley PA, George JC, Reeves RR, Heide-Jørgensen MP, Tynan CT, Mayo CA (2018) Sightings of a bowhead whale (Balaena mysticetus) in the Gulf of Maine and its interactions with other baleen whales J CETACEAN RES MANAGE 19: 23–30
Ahonen H, Stafford KM, De Steur L, Lydersen C, Wiig Ø, Kovacs K 2017 The underwater soundscape in western Fram Strait:Breeding ground of Spitsbergen’s endangered bowhead whales. Marine Pollution Bulletin 123 (2017) 97–112
Ahonen H, Stafford KM, Lydersen C, de Steur L , Kovacs KM (2019) A multi-year study of narwhal occurrence in the western Fram Strait—detected via passive acoustic monitoring Polar Research 2019, 38, 3468, http://dxdoiorg/1033265/polarv383468
Allen RC, Keay I (2006) Bowhead whales in the Eastern Arctic, 1611–1911: Population reconstruction with historical whaling records Environment and History 12:89–113
Alter SE, Rosenbaum HC, Postma LD, Whitridge P, Gaines C, Weber D, Egan MG, Lindsay M, Amato G, Dueck L, Brownell JrRL, Heide-Jørgensen M-P8 , Laidre KL, Caccone G ,Brittany Log Hancock BL (2012) Gene flow on ice: the role of sea ice and whaling in shaping Holarctic genetic diversity and population differentiation in bowhead whales (Balaena mysticetus) Ecology and Evolution2: 2895–2911
Alter ES, Meyer M, Post K, Czechovski P, Gravlund P, Gaines C, Rosenbaum HC, Kaschner K, Turvey ST, Van der Plicht J, Shapiro B, Hofreiter M (2015) Climate impacts on transocean dispersal and habitat in gray whales from the Pleistocene to 2100 Molecular Ecology (2015) 24, 1510–1522
Andersen JM, Wiersma YF, Stenson GB, Hammill MO, Rosing-Asvid A, Skern-Maurizen M (2013) Habitat selection by hooded seals (Cystophora cristata) in the Northwest Atlantic Ocean – ICES Journal of Marine Science, 70: 173–185
Andvik C, Jourdain E, Ruus A, Lyche JL, Karoliussen R, Borgå K (2020) Preying on seals pushes killer whales from Norway above pollution defects thresholds Scientific Reports 10:11888 | https://doiorg/101038/s41598-020-68659-y
Aniceto AS, Pedersen G, Primicerio R, Biuw M, Lindstrøm U, Camus L (2020) Arctic Marine Data Collection Using Oceanic Gliders: Providing Ecological Context to Cetacean Vocalizations 7(November), 1–11 https://doiorg/103389/fmars2020585754
Azzara A J, von Zharen WM, Newcomb JJ (2013) Mixed-methods analytic approach for determining potential impacts of vessel noise on sperm whale click behavior J Acoust Soc Am 134, 4566–4574 doi: 101121/14828819
Bachmann L, Cabrera AA, Heide-Jørgensen MP, Shpak OV, Lydersen C, Wiig Ø, Kovacs KM, (2021) Mitogenomics and the genetic differentiation of contemporary Balaena mysticetus (Cetacea) from Svalbard, Zoological Journal of the Linnean Society, Volume 191: 1192–1203, https://doiorg/101093/zoolinnean/zlaa082
Baines M, Reichelt M (2014) Upwellings, canyons and whales: An important winter habitat for balaenopterid whales off Mauritania, northwest Africa Cetacean Res 14: 57–67
Baird AB, Bickham JW (2021)The stocks of bowheads2021 Editor(s): George, JC and JGM Thewissen JGM, The Bowhead Whale, Academic Presspp 19-29
Ballard K, Kovacs K (1995) The acoustic repertoire of hooded seals (Cystophora cristata) Canadian Journal of Zoology 73:1362–1374 https://doiorg/101139/z95-159
Baumgartner MF, Cole T, Campbell R, Teegarden G, Durbin E (2003) Associations between North Atlantic right whales and their prey, Calanus finmarchicus, over diel and tidal time scales Mar Ecol Prog Ser 264: 155–166
Benjaminsen T, Christensen I (1979) The natural history of the bottlenose whale, Hyperoodon ampullatus (Forster) In HE Winn, and BL Olla (eds), Behavior of Marine Animals Volume 3, 143–164 New York: Plenum https://doiorg/101007/978-1-4684-2985-5_5
Benti B, Miller PJO Biuw M, Curé C (2021) Indication that the behavioural responses of humpback whales to killer whale sounds are influenced by trophic relationships Vol 660: 217–232https://doiorg/103354/meps13592
Blanchet MA, Lydersen C, Ims RA, Lowther AD, Kovacs KM (2014) Harbour seal Phoca vitulina movement patterns in the high-Arctic archipelago of Svalbard, Norway Aquat Biol 21: 167−181
Boertmann EW, Kyhn A, Witting L, Heide-Jørgensen MP (2015) A hidden getaway for bowhead whales in the Greenland Sea Polar Biology 38: 1315-1319
Boertmann D, Blockley D, Mosbech A (2020) Greenland Sea – an updated strategic environmental impact assessment of petroleum activities Scientific report from DCE – Danish Cente for Environment and Energy No 375, 380 pp http://dce2audk/pub/SR375pdf
Boon JP, Van der Meer J, Allchin CR, Law RJ, Klungsoyr J, Leonards PEG, Spliid H, Storr-Hansen E, Mckenzie C og Wells DE (1997) Concentration dependent changes of PCB patterns in fish-eating mammals: structural evidence for induction of cytochrome P450 Arch Environ Contam Toxicol, 33, pp 298-311
Bockstoce JR, Burns JJ (1993) Commercial whaling in the North Pacific sector In The bowhead whale (eds J J Burns, J J Montague & C J Cowles), pp 563 – 577 Lawrence, Kansas: Society for Marine Mammalogy
Bourque J, Dietz R, Sonne C, St Leger J mfl. (2018) Feeding habits of a new Arctic predator: insight from full-depth blubber fatty acid signatures of Greenland, Faroe Islands, Denmark, and managed-care killer whales Orcinus orca Mar Ecol Prog Ser 603:1-12
Broms C, Melle W, Kårtvedt S (2009) Oceanic distribution and life cycle of Calanus species in the Norwegian Sea and adjacent waters Deep-Sea Research II 56:1910–1921
Burek KA, Gulland FMD, O’Hara TM (2008) Effects of climate change on marine mammal health Ecological Applications 18, S126–134
Burns JJ, Gavin A (1980) Recent records of hooded seals, Cystophora cristata Erxleben, from the western Beaufort Sea Arctic 33: 326-329
Carlström JJ, Denkinger P, Feddersen, Øien N (1997) Record of a new northern range of Sowerby’s beaked whale (Mesoplodon bidens) Polar Biol 17:459–461
Chambault P, Albertsen CM, Patterson TA, Hansen RG, Tervo O, Laidre KL, Heide-Jørgensen MP (2018) Sea surface temperature predicts the movements of an Arctic cetacean: the bowhead whale Scientific Reports, 8(1), [9658] https://doiorg/101038/s41598-018-27966-1
Christensen I (1973) Age determination, age distribution and growth of bottlenose whales, Hyperoodon ampullatus (Forster), in the Labrador Sea Norwegian Journal of Zoology 21:331-340
Christensen I, Haug T, Øien N (1992) Seasonal distribution, exploitation and present abundance of stocks of large baleen whales (Mysticeti) and sperm whales (Physeter Macrocepahlus) in Norwegian and adjacent waters ICES J MarSci 49: 341-355
Cooke JG (2018) b Balaenoptera acutorostrata The IUCN Red List of Threatened Species 2018: eT2474A50348265 https://dxdoiorg/102305/IUCNUK2018-2RLTST2474A50348265en Downloaded on 31 May 2021
Cooke JG (2018) c Balaenoptera musculus (errata version published in 2019) The IUCN Red List of Threatened Species 2018: eT2477A156923585 https://dxdoiorg/102305/IUCNUK2018-2RLTST2477A156923585en Downloaded on 31 May 2021
Cooke JG, Reeves R (2018) Balaena mysticetus The IUCN Red List of Threatened Species 2018: eT2467A50347659 https://dxdoiorg/102305/IUCNUK2018-1RLTST2467A50347659en Downloaded on 31 May 2021
Cooke JG (2020) Eubalaena glacialis (errata version published in 2020) The IUCN Red List of Threatened Species 2020eT41712A178589687 https://dxdoiorg/102305/IUCNUK2020-2RLTST41712A178589687en Downloaded on 03 June 2021
Craddock JE, Polloni PT, Hayward B mfl. (2009) Food habits of Atlantic white-sided dolphins (Lagenorhynchus acutas) off the coast of New England Fisheries Bulletin, 107:384-394
Croll DA, Clark CW, Acevedo A, Tershy B, Flores S, Gedamke J, Urban J (2002) “Bioacoustics: Only male fin whales sing loud songs,” Nature London 417, 809
Cziko PA, Munger LM, Santos NR, Terhune JM (2020) Weddell seals produce ultrasonic vocalizations The Journal of the Acoustical Society of America 148, 378 doi: 101121/100002867
Davis GE, Baumgartner MF, Bonnell JM, Bell J, Berchok C, Thornton JB, Brault S, Buchanan G, Charif RA, Cholewiak D, Clark CW (2017) Long-term passive acoustic recordings track the changing distribution of North Atlantic right whales (Eubalaena glacialis) from 2004 to 2014Scientific Reports 7(1):13460, https://doiorg/101038/s41598-017-13359-3
Davis DA, Mondo K, Stern E, Annor AK, Murch SJ, Coyne TM, mfl. (2019) Cyanobacterial neurotoxin BMAA and brain pathology in stranded dolphins PLoS ONE 14(3): e0213346 https://doi org/101371/journalpone0213346
Debey L, Pyenson ND (2013) Osteological correlates and phylogenetic analysis of deepdiving in living and extinct pinnipeds: What good are big eyes?Marine Mammal Science 29:48-83
De Boer MN, Jones D, Jones H (2017) Ocean wanderers: extralimital encounters with bowhead whales (Balaena mysticetus) in temperate European shallow waters Aquat Mamm 43: 279−288
De Boer M, Janinhoff N, Nijs G, Verdaat H (2019) Encouraging encounters: unusual aggregations of bowhead whales Balaena mysticetus in the western Fram Strait Endangered Species Research Vol 39: 51–62 https://doiorg/103354/esr00948
Dehnhardt G, Mauck B, Hanke W, Bleckmann H (2001) Hydrodynamic Trail-Following in Harbor Seals (Phoca vitulina) SCIENCE 293: 102-104
Descamps S, Aars J, Fuglei E, Kovacs KM, Lydersen C, Pavlova O, Pedersen ÅØ, Ravolainen V, Strøm H (2017) Climate change impacts on wildlife in a High Arctic archipelago – Svalbard, Norway Global Change Biology 23, 490–502, doi: 101111/gcb13381
Desforges JP, Hall A, McConnell B, Rosing-Asvid A, Barber JL, Brownlow A, De Guise S, Eullaers I, Jepson PD, Letcher RJ, Levi M, Ross PS, Samarra F, Vikingson G, Sonne, Dietz R (2018) Predicting global killer whale population collapse from PCB pollution Science 361:1373–1376
Desforges JP, Hall A, McConnell B mfl. (2019) Response to L Witting: PCBs still a major risk for global killer whale populations Mar Mam Sci 2019;35:1201–1206
Dietz R, Letcher RJ, Desforges J-P, Eulaers I, Sonne C mfl. (2019) Current state of knowledge on biological effects from contaminants on arctic wildlife and fish Science of the total Environment: 696 (December) https://doiorg/101016/jscitotenv2019133792
Doniol-Valcroze T, Gosselin J-F, Pike DG, Lawson JW, Asselin NC, Hedges K, Ferguson SH (2020) Distribution and Abundance of the Eastern Canada–West Greenland Bowhead Whale Population Based on the 2013 High Arctic Cetacean Survey NAMMCO Scientific Publications 11 https://doiorg/107557/35315
Doucette GJ, Mikulski CM, King KL, Roth PB, Wang Z, Leandro LF, DeGrasse SL, White KD, De Biase D, Gillett RM (2012) Endangered North Atlantic right whales (Eubalaena glacialis) experience repeated, concurrent exposure to multiple environmental neurotoxins produced by marine algae Environ Res 112, 67e76
Duarte CM, Chapuis L, Collin SP mfl. (2021) The soundscape of the Anthropocene ocean Science 371: 583
Dyke AS, Savelle JM (2001) Holocene History of the Bering Sea Bowhead Whale (Balaena mysticetus) in Its Beaufort Sea Summer Grounds off Southwestern Victoria Island, Western Canadian Arctic Quaternary Research 55, 371–379
Enoksen S, Haug T, Lindstrøm U, Nilssen KT (2016) Recent summer diet of hooded Cystophora cristata and harp Pagophilus groenlandicus seals in the drift ice of the Greenland Sea Polar Biol 40: 931-937
Evans K, Hindell M, Robertson K, Lockyer C, Rice D (2002) Factors affecting the precision of age determination of sperm whales (Physeter macrocephalus) J Cetacean Res Manage, 4, 193-201
Fall J, Skern-Mauritzen M (2014) White-beaked dolphin distribution and association with prey in the Barents Sea. Marine Biology Research, 10(10), 957–971
Falk-Petersen S, Pavlov V, Berge J, Cottier F, Kovacs K, Lydersen C (2015) At the rainbow’s end: high productivity fueled by winter upwelling along an Arctic shelf Polar Biology 38: 5-11
Fernández A, Edwards J F, Rodriguez F, Espinosa De Los Monteros A, Herraez P, Castro P, Jaber JR, Martin V, Arbelo M (2005) Gas and fat embolic syndrome involving a mass stranding of beaked whales (family Ziphiidae) exposed to anthropogenic sonar signals Vet Pathol 42:446–457
Fernandez R, Pierce GJ, Macleod C mfl. (2014) Strandings of northern bottlenose whales,Hyperoodon ampullatus, in the north-east Atlantic: seasonality and diet Journal of the Marine Biological Association of the United Kingdom, 2014doi:101017/S002531541300180X
Finley KJ, Miller GW, Davis RA, Greene CR (1990) Reactions of belugas, Delphinapterus leucas, and narwhals, Monodon monoceros, to icebreaking ships in the Canadian high arctic Can Bull Fish Aquat Sci 224, 97–117
Fire SE, Browning JA, Durden WN, Stolen MK (2020) Comparison of during-bloom and inter-bloom brevetoxin and saxitoxin concentrations in Indian River Lagoon bottlenose dolphins, 2002–2011 Aquatic Toxicology Volume 218, January 2020, 105371
Folkow LP, Mårtensson PE, Blix AS (1996) Annual distribution of hooded seals (Cystophora cristata) in the Greenland and Norwegian seas Polar Biol 16: 179–
Folkow LP, Blix AS (1999) Diving behaviour of hooded seals (Cystophora cristata) in the Greenland and Norwegian Seas Polar Biol 22: 61–74
Folkow LP, Nordøy ES, Blix AS (2010) Remarkable development of diving performance and migrations of hooded seals (Cystophora cristata) during their first year of life Polar Biol 33, 433-441
Folkow LP, Nordøy ES, Blix AS (2004) Distribution and diving behaviour of harp seals (Pagophilus groenlandicus) from the Greenland Sea stock Polar Biol 27, 281–298 https://doiorg/101007/s00300-004-0591-7
Foote AD, Newton J, Piertney SB, Willerslev E, Gilbert MTP (2009) Ecological, morphological and genetic divergence of sympatric North Atlantic killer whale populations Molecular Ecology 18(24):5207± 17 PMID: 20050301
Foote AD mfl (2013) Ancient DNA reveals that bowhead whale lineages survived Late Pleistocene climate change and habitat shifts Nat Commun 4:1677 doi: 101038/ncomms2714
Fossum P, Mork KA, Tverberg V mfl. (2012) Beskrivelse av miljø og levende marine ressurser i havområdene ved Jan Mayen Rapport fra Havforskningen 16: 119 pp
Foskolos I, Aguilar de Soto N, Madsen PT, Johnson M (2019) Deep-diving pilot whales make cheap, but powerful, echolocation clicks with 50 µL of air Scientific Reports Vol 9, October 31, 2019 doi:101038/s41598-019-51619-6
Friedlaender AS, Bowers MT, Cade D, Hazen EL, Stimpert A, Allen AN, Calambokidis J, Fahlbusch J, Segre P, Visser F, Southall B, Goldbogen, JA (2020) The advantages of diving deep: Fin whales quadruple their energy intake when targeting deep krill patches Functional Ecology 34:497–506 DOI: 101111/1365-243513471
Garde E, Heide-Jørgensen MP, Hansen SH, Nachman G, Forchammer M (2007) Age specific growth and remarkable longevity in narwhals (Monodon Monoceros) from West Greenland as estimated by aspartic acid racemization Journal of Mammalogy, 88(1):49–58
George JC, Bada J, Zeh J, Scott L, Brown SE, O’Hara T, Suydam R (1999) Age and growth estimates of bowhead whales (Balaena mysticetus) via aspartic acid racemization Can J Zool77, 571–580
George JC, Druckenmiller ML, Laidre KL, Suydam R, Person B (2015) Bowhead whale body condition and links to summer sea ice and upwelling in the Beaufort Sea Progress in Oceanography 136 (2015) 250–262
George JC, Horstmann,L, Fortune S, Sformo T L, Elsner R, Follmann E (2021) Thermoregulation and energetics 2021 Editor(s): George, JC and JGM Thewissen JGM, The Bowhead Whale,Academic Presspp 57-75,ISBN 9780128189696,https://doiorg/101016/B978-0-12-818969-600005-4
Givens GH, Edmondson SL, George JC, Tudor B, DeLong RA, Suydam R (2016) Horvitz-Thompson whale abundance estimation adjusting for uncertain recapture, temporal availability variation, and intermittent effort Envirometrics (wileyonlinelibrarycom; DOI: 101002/env2379)
Gobler CJ Doherty OM, Hattenrath-Lehmann TK, Griffith A W, Kanga Y, Litaker RW (2017) Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans PNAS 114: 4975–4980
Goldbogen JA, Calambokidis J, Oleson E, Potvin J, Pyenson ND, Schorr G, Shadwick RE (2011) Mechanics, hydrodynamics and energetics of blue whale lunge feeding: efficiency dependence on krill density J Exp Biol 214, 131-146
Goldbogen JA, Calambokidis J, Croll D, McKenna MF, Potvin J, Pyenson ND, Schorr G, Shadwick RE, Tershy BR (2012) Scaling of lunge feeding performance in rorqual whales: mass-specific energy expenditure increases with body size and progressively limits diving capacity Funct Ecol 26, 216-226
Goldbogen JA, Cade DE, Wisniewska DM, Potvin J, Segre PS, Savoca MS, Hazen EL, Czapanskiy MF, Kahane-Rapport SR, DeRuiter SL, Gero S, Tønnesen P, Gough WT, Hanson MB, Holt MM, Jensen FH, Simon M, Stimpert K, Arranz P, Johnston DW, Nowacek DP, Parks SE, Visser F, Friedlaender AS, Tyack PL, Madsen PT og ND Pyenson (2019) Why whales are big but not bigger: Physiological drivers and ecological limits in the age of ocean giants Science 366 (6471), 1367-1372
Gowan TA, Ortega-Ortiz JG, Hostetler JA, Hamilton PK, Knowlton AR, Jackson KA George RC, Taylor CR, Naessig PJ (2019) Temporal and demographic variation in partial migration of the North Atlantic right whale Scientific Reports 9:353 |DOI:101038/s41598-018-36723-3
Guilpin M, Lesage V, McQuinn I, Brosset P, Doniol-Valcroze T, Jeanniard-du-Dot T, Winkler G (2020) Repeated Vessel Interactions and Climate- or Fishery-Driven Changes in Prey Density Limit Energy Acquisition by Foraging Blue Whales Front Mar Sci 7:626doi: 103389/fmars202000626
Gullestad P, Aglen A, Bjordal A, Blom G, Johansen S, Krog J, Misund OA, Røttingen I (2014) Changing attitudes 1970–2012: evolution of the Norwegian management framework to prevent overfishing and to secure long-term sustainability – ICES Journal of Marine Science, 71:173–182
Hammill MO, Stenson GB (2000) Estimated prey consumption by harp seals (Phoca groenlandica), hooded seals (Cystophora cristata), grey seals (Halichoerus grypus) and harbour seals (Phoca vitulina) in Atlantic Canada J Northwest Atl Fish Sci 26:1–23
Hamilton CD, Lydersen C, Aars J mfl. (2021) Marine mammal hotspots in the Greenland and Barents Seas Marine Ecology Progress Series Vol 659: 3–28
Harris RE, Miller GW, Richardson J (2001) Seal responses to airgun sounds during summer seismic surveys in the Alaskan Beaufort Sea Marine Mammal Science 17(4):795-812
Harwood LA, Kingsley MCSK (2013) Trends in the Offshore Distribution and Relative Abundance of Beaufort Sea Belugas, 1982 – 85 vs 2007 – 09 ARCTIC 3:247-256
Haug T, Nilssen KT, Lindblom L (2004) Feeding habits of harp and hooded seals in drift ice waters along the east coast of Greenland in summer and winter Polar Res 23:35–42
Haug T, Nilssen KT, Lindblom L, Lindstrøm U (2007) Diets of hooded seals in coastal and drift ice waters along the east coast of Greenland Mar Biol Res 3:123–133
Hauksson E, Víkingsson GA, Halldorsson SD, Olafsdottir D, Sigurjónsson J (2011) Preliminary report on biological parameters for NA minke whales in Icelandic waters Report of the International Whaling Commission 63: 1–45
Hatch LT, Clark CW, Van Parijs SM, Frankel A, Ponirakis D (2012) Quantifying loss of communication space for right whales in and around a US National Marine Sanctuary Conservation Biology 26: 21–28
Heath HR, Rasmussen J, Ahmed Y mfl. (2008) Spatial demography of Calanus finmarchicus in the Irminger Sea Progress in Oceanography 76:39–88
Heide-Jørgensen MP, Laidre KL, Quakenbush L, Citta JJ (2012) The Northwest Passage opens for bowhead whales Biol Lett 8, 270–273 doi:101098/rsbl20110731
Heide-Jørgensen MP, Hansen RG, Westdal K, Reeves R, Mosbech A (2013) Narwhals and seismic exploration: Is seismic noise increasing the risk of ice entrapments? Biological Conservation 158: 50–54
Heide-Jørgensen M, Blackwell SB, mfl. (2020) Some like it cold: Temperature-dependent habitat selection by narwhals Ecology and Evolution 2020;10:8073–8090
Heide-Jørgensen MP, Hansen RG, Shpak OV (2021) Chapter 5 - Distribution, migrations, and ecology of the Atlantic and the Okhotsk Sea Populations, Editor(s): JC George, JGM Thewissen, The Bowhead Whale, Academic Press, pp 57-75
Hendrix AM, Lefebvre KA, Quakenbush L, Bryan A, Stimmelmayr R, Sheffield G, Wisswaesser G, Willis ML, Bowers EK, Kendrick P, Frame E, Burbacher T, Marcinek DJ (2021) Ice seals as sentinels for algal toxin presence in the Pacific Arctic and subarctic marine ecosystems Mar Mam Sci ;1–17
Henriksen S, Hilmo O (2015) Norsk rodliste for arter 2015 Artsdatabanken, Norge
Hoelzel AR, Sarigol F, Gridley T, Elwen SH (2021) Natal origin of Namibian grey whale implies new distance record for in-water migration Biol Lett172021013620210136 http://doiorg/101098/rsbl20210136
Holt MM, Noren DP, Emmons CK (2011) Effects of noise levels and call types on the source levels of killer whale calls J Acoust Soc Am 130: 3100–3106
Houghton LE, Ramirez-Martinez N, Mikkelsen B Víkingsson G, Gunnlaugsson T, Øien N, Hammond PS (2020) Oceanic Drivers of Sei Whale Distribution in the North Atlantic NAMMCO Scientific Publications 11 https://doiorg/107557/35211
Huijser LAE, Bérubé M, Cabrera AA, Prieto R, Silva MA, Robbins J, Kanda N, Pastene LA, Goto M,Yoshida H, Víkingsson, GA, Palsbøll PJ (2018) Population structure of North Atlantic and North Pacific sei whales (Balaenoptera borealis) inferred from mitochondrial control region DNA sequences and microsatellite genotypes Conservation Genetics19:1007–1024 https://doiorg/101007/s10592-018-1076-5
ICES 2019 ICES/NAFO/NAMMCO Working Group on Harp and Hooded Seals (WGHARP)ICES Scientific Reports 1:72 193 pp http://doiorg/1017895/icespub5617
Ingebrigtsen A (1929 )Whales caught in the North Atlantic and other seas Rapp P-v Reun ConsPermintExplorMer 56 (2),1-26
Isojunno S, Miller PJO (2018) Movement and biosonar behavior during prey encounters indicate that male sperm whales switch foraging strategy with depth Frontiers in Ecology and Evolution, 6(NOV), 1–15 https://doiorg/103389/fevo201800200
IWC (2018) Report of the Sub-Committee on the Revised Management Procedure Journal of Cetacean Research and Management 19(Suppl): 115-153
Jakobsen K-O, Marx M, Øien N (2003) Two-way Trans-Atlantic migration of a North Atlantic Right Whale (Eubalaena glacialis) Marine Mammal Science 20:161-166
Jepson, PD mfl (2016) PCB pollution continues to impact populations of orcas and other dolphins in European waters Sci Rep 6, 18573; doi: 101038/srep18573 (2016)
Jensen S-K, Lacaze J-P, Hermann G, Kershaw J, Brownlow, Turner A, Hall A (2015) Detection and effects of harmful algal toxins in Scottish harbour seals and potential links to population decline Toxicon 97:1-14
Jonsgård A (1977) Tables showing the catch of small whales (including minke whales) caught by Norwegians in the period 1938–75, and large whales caught in different North Atlantic waters in the period 1868–1975 Reports of the International Whaling Commission, 27, 413–426
Jourdain E, Vongraven D, Bisther A, Karoliussen R (2017) First longitudinal study of seal-feeding killer whales (Orcinus orca) in Norwegian coastal waters PLoS ONE 12(6): e0180099 https://doiorg/101371/journal pone0180099
Jourdain E, Ugarte F, Vikingsson G, Samarra F, Ferguson S, Lawson J, Vongraven D, Desportes G (2019) North Atlantic killer whale Orcinus orca populations: a review of current knowledge and threats to conservation Mammal Review 49 (2019)
Kavanagh AS, Nykänen M, Hunt W Richardson N, Jessopp MJ (2019) Seismic surveys reduce cetacean sightings across a large marine ecosystem Scientific Reports 9:19164 | https://doiorg/101038/s41598-019-55500-4
Konráðsson A, Gunnlaugsson Þ (1990) A note on earplug readings from fin whales caught off Iceland, in particular, the use of photographs Reports of the International Whaling Commission, 40, 375-378
Koski WR, Funk DW, Ireland DS, Lyons C, Christie K, Macrander AM, Blackwell SB (2009) An Update on Feeding by Bowhead Whales near an Offshore Seismic Survey in the Central Beaufort Sea IWC SC, pp 61
Kovacs KM, Lydersen C, Vacquiè-Garcia J, Shpak O, Glazov D, Heide-Jørgensen MP (2020) The endangered Spitsbergen bowhead whales’ secrets revealed after hundreds of years in hiding Biol Lett 16: 20200148 http://dxdoiorg/101098/rsbl20200148
Kvadsheim PH, Sevaldsen EM, Folkow L, Blix AS (2010) Behavioural and Physiological Responses of Hooded Seals (Cystophora cristata) to 1 to 7 kHz Sonar Signals Aquatic Mammals 36:239-247 ISSN 0167-5427 doi: 101578/AM3632010239
Kvadsheim PH, Forland TN, De Jong K, Nyqwist D, Grimsbø E, Sivle LD (2020) Effekter av støyforurensing på havmiljø- kunnskapsstatus og forvaltningsrådgiving FFI report 2020/01015 (http://rapporterffino/rapporter/2020/01015pdf)
Kyhn LA, Wisniewska DM, Beedholm K, Tougaard J, Simon M, Mosbech A, Madsen PT (2019) Basin-wide contributions to the underwater soundscape by multiple seismic surveys with implications for marine mammals in Baffin Bay, Greenland Marine Pollution Bulletin 138: 474–490
Laidre KL, Heide-Jørgensen MP, Nielsen TG (2007) Role of the bowhead whale as a predator in West Greenland Marine Ecology Progress Series Vol 346: 285–297
Laidre KL, Heide-Jørgensen MP (2012) Spring partitioning of Disko Bay, West Greenland, by Arctic and Subarctic baleen whales, ICES Journal of Marine Science 69: 1226–1233, https://doiorg/101093/icesjms/fss095
Lefort KJ, Garroway CJ, Ferguson SH (2020) Killer whale abundance and predicted narwhal consumption in the Canadian Arctic Global Change Biology, 26(8), 4276-4283 doi:101111/gcb15152
Lefebvre KA, Quakenbush L, Frame E, Huntington KB, Sheffield G, Stimmelmayr R, Bryan A, Kendrick P, Ziel H, Goldstein T (2016) Prevalence of algal toxins in Alaskan marine mammals foraging in a changing arctic and subarctic environment Harmful Algae 55:13–24
Leonard DM, Øien NI (2020) a Estimated Abundances of Cetaceans Species in the Northeast Atlantic from Two Multiyear Surveys Conducted by Norwegian Vessels between 2002–2013 NAMMCO Scientific Publications 11 https://doiorg/ 107557/34695
Leonard DM, Øien NI (2020) b Estimated Abundances of Cetacean Species in the Northeast Atlantic from Norwegian Shipboard Surveys Conducted in 2014–2018 NAMMCO Scientific Publications 11 https://doiorg/ 107557/34694
Louis M mfl. (2020) Influence of past climate change on phylogeography and demographic history of narwhals, Monodon monoceros Proc R Soc B 287: 20192964 http://dxdoiorg/101098/rspb20192964
Lydersen C, Freitas C, Wiig Ø, Bachmann L, Heide-Jørgensen MP, Swift R, Kovacs KM (2012) Lost highway not forgotten: satellite tracking of a bowhead whale (Balaena mysticetus) from the Critically Endangered Spitsbergen Stock Arctic 65: 76–86 (doi:1014430/arctic4167)
Lydersen C, Vacquie‑Garcia J, Heide‑Jorgensen M P, Øien N4, Guinet C, Kovacs KM (2020) Autumn movements of fin whales (Balaenoptera physalus) from Svalbard, Norway, revealed by satellite tracking Scientific Reports 10:16966 https://doiorg/101038/s41598-020-73996-z
McCordic J, Todd S, Stevick, P (2014) Differential rates of killer whale attacks on humpback whales in the North Atlantic as determined by scarification Journal of the Marine Biological Association of the United Kingdom, 94(6), 1311-1315 doi:101017/S0025315413001008
Magnusdottir EE, Lim R (2019) Subarctic singers: Humpback whale (Megaptera novaeangliae) song structure and progression from an Icelandic feeding ground during winter PLoS ONE 14(1): e0210057 https://doiorg/ 101371/journalpone0210057
Marriott S, Cowan E, Cohen J, Hallock RM (2013) Somatosensation, Echolocation, and Underwater Sniffing: Adaptations Allow Mammals Without Traditional Olfactory Capabilities to Forage for Food Underwater ZOOLOGICAL SCIENCE 30: 69–75
Martin AR, Clarke MR (1986) The diet of sperm whales (Physeter macrocaphalus) captured between Iceland and Greenland J Mar Biol Assoc, 66:779-790
Matthews CJD, Breed GA, LeBlanc B, Ferguson SH (2020) a Killer whale presence drives bowhead whale selection for sea ice in Arctic seascapes of fear Proc Natl Acad Sci 117 (12), 6590-6598
Matthews LP, Fournet MEH, Gabriele C, Klinck H, Parks SE (2020) b Acoustically advertising male harbour seals in southeast Alaska do not make biologically relevant acoustic adjustments in the presence of vessel noise Biol Lett 16: 20190795 http://dxdoiorg/101098/rsbl2019079
McKenzie CH, Bates SS, Martin JL, Haigh N, Howland KL, Lewis NL, Locke A, Pe˜na AG, Poulin M, Rochon A, Rourke WA, Scarratt MG, Starr M, Wells T (2021) Three decades of Canadian marine harmful algal events: Phytoplankton and phycotoxins of concern to human and ecosystem health Harmful Algae 102:101852
McKinney M, Iverson S, Fisk A, Sonne C, Riget F, Letcher R, Arts M, Born E, Rosing-Asvid A, Dietz R (2013) Global change effects on the long-term feeding ecology and contaminant exposures of East Greenland polar bears Glob Chang Biol 19:2360–2372 doi:101111/gcb12241
Mellinger DK, Nieukirk S, Klinck K, Klinck H, Dziak RP, Clapham PJ, Brandsdottir B (2011) Confirmation of right whales near a nineteenth-century whaling ground east of southern Greenland Biology Letters 7: 411-413
Merkel, B, Lydersen, C, Yoccoz, NG, Kovacs, KM (2013) The World’s Northernmost Harbour Seal Population – How Many Are There? PLoS One, 8(7) htt
Miller PJO, Kvadsheim PH, Lam FPA, Tyack PL, Cure C, DeRuiter SL, Kleivane L, Sivle LD, Van Ijsselmuide SP, Visser F, Wensveen PJ, Von Benda-Beckmann AM, Martín López LM, Narazaki T, Hooker SK (2015) First indications that northern bottlenose whales are sensitive to behavioural disturbance from anthropogenic noise Royal Society Open Science 2(6): 140484 doi: 101098/rsos140484
Monsarrat S, Pennino MG, Smith TD, Reeves RR, Meynard CN, Kaplan DM, Rodrigues ASL (2015) Historical summer distribution of the endangered North Atlantic right whale (Eubalaena glacialis): a hypothesis based on the environmental niche of a congeneric species Diversity and Distributions 21:925–937
Monsarrat S, Pennino MG, Smith TD, Reeves RR, Meynard CN, Kaplan DM, Rodrigues ASL (2016) A spatially explicit estimate of the prewhaling abundance of the endangered North Atlantic right whale Conservation Biology, Volume 30:783–791
NAMMCO-North Atlantic Marine Mammal Commission (2019) a Report of the Ad hoc Working Group on Narwhal in East Greenland September 2019, Copenhagen, Denmark Available at https://nammcono/topics/sc-wg-reports/
NAMMCO-North Atlantic Marine Mammal Commission (2019) b Report of the Scientific Committee 26th Meeting, October 29 – November 1, Tórshavn Faroe Islands, Available at https://nammcono/topics/scientific-committee-reports/
NAMMCO (2021): https://nammcono/topics/common-minke-whale/, accessed 25 May 2021
Nordøy E, Folkow L, Potelov V, Blix A (2008) Seasonal distribution and dive behaviour of harp seals (Pagophilus groenlandicus) of the White Sea–Barents Sea stock Polar Biology 31(9):1119-1135
Nøttestad L, Krafft BA, Anthonypillai V, Bernasconi M, Langård L, Mørk HL, Fernö A (2015) Recent changes in distribution and relative abundance of cetaceans in the Norwegian Sea and their relationship with potential prey Front Ecol Evol 2:83
Okolodkov YB (2005) The global distributional patterns of toxic, bloom dinoflagellates recorded from the Eurasian Arctic Harmful Algae 4:351–369
Olesiuk PF, Bigg MA, Ellis GM (1990) Life history and population dynamics of Resident killer whales (Orcinus orca) in the coastal waters of British Columbia and Washington State Report of the International Whaling Commission, 12: 209–243
Olsen E, Sunde J (2002) Age determination of minke whales (Balaenoptera acutorostrata) using the aspartic acid racemization technique Sarsia 87:1–8
Perry EA, Terhune JM (1999) Variation of harp seal (Pagophilus groenlandicus) underwater vocalisations among three breeding locations Journal of Zoology, 247, 181e186
Pettis HM, Pace RM, Schick RS, Hamilton PK (2020) North Atlantic Right Whale Consortium 2019 Annual Report Card
Pike DG, Víkingsson GA, Gunnlaugsson T, Øien N (2009) A note on the distribution and abundance of blue whales (Balaenoptera musculus) in the Central and Northeast North Atlantic NAMMCO Sci Publ 7, 19–29doi: 107557/32703
Polanowski AM, Robbins J, Chandler D, Jarman SN (2014) Epigenetic estimation of age in humpback whales Molecular Ecology Resources 14:976–987 doi:101111/1755-099812247
Pomerleau C, Ferguson SH, Walkusz W (2011) Stomach contents of bowhead whales (Balaena mysticetus) from four locations in the Canadian Arctic Polar Biol 34:615–620
Pomeroy P (2011) Reproductive cycles of marine mammals Animal Reproduction Science 124:184–193
Record NR, Runge JA, Pendleton DE mfl. (2019) Rapid Climate-Driven Circulation Changes Threaten Conservation of Endangered North Atlantic Right Whales Oceanography 32:162-169
Reeves RR, Mitchell E, Whitehead H (1993) Status of the northern bottlenose whale, Hyperoodon ampullatus Canadian Field-Naturalist, 107:490–508
Reichmut C, Holt MM, Mulsow J, Sills J, Southall J (2013) Comparative assessment of amphibious hearing in pinnipeds J Comp Physiol A 199:491–507
Rice DW (1998) Marine Mammals of the World Systematics and Distribution, Society for Marine Mammalogy Special Publication 4
Riekkola L, Zerbini AN, Andrews O mfl. (2018) Application of a multi-disciplinary approach to reveal population structure and Southern Ocean feeding grounds of humpback whales Ecol Indicat, 89 (2018), pp 455-465
Robertson F, Koski WR, Trites AW (2016) Behavioral responses affect distribution analyses of bowhead whales in the vicinity of seismic operations Marine Ecology Progress Series Volume: 549 Pages: 243-262DOI: 103354/meps11665
Rolland, R Parks, S Hunt, K mfl. (2012) Evidence that ship noise increases stress in right whales Proc Biol Sci 2012 Jun 22; 279(1737): 2363–2368
Rosa C, Zeh J, George JC, Botta O, Zauscher M, Bada J, O'Hara TM (2013) Age estimates based on aspartic acid racemization for bowhead whales (Balaena mysticetus) harvested in 1998–2000 and the relationship between racemization rate and body temperature Marine Mammal Science 29:424-445
Rotander A, Kärrman A, Van Bavel B, Polder A, Rigét F, Auðunsson GA, Vikingsson G, Bloch D, Dam M (2012) Increasing levels of long-chain perfluorocarboxylic acids (PFCAs) in Arctic and North Atlantic marine mammals, 1984–2009 Chemosphere 86:278-285
Santos MB, Pierce GJ, Boyle PR, Reid RJ, Ross HM, Patterson IAP, Kinze CC, Tougård S, Lick R, Piatkowski U, Hernandez GV (1999) Stomach contents of sperm whales Physeter macrocephalus stranded in the North Sea 1990–1996 Marine Ecology Progress Series 183: 281-294
Santos MB, Pierce GJ, Garcia Hartmann M, Smeenk C, Addink MJ, Kuiken T, Reid RJ, Patterson IAP, Lordan C, Rogan E, Mente E (2002) Additional notes on stomach contents of sperm whales Physeter macrocephalus stranded in the NE Atlantic Journal of the Marine Biological Association of the UK 82: 501-507
Scheinin AP, Kerem D, MacLeod CD, Gazo M, Chicote CA, Castellote M (2011) Gray whale (Eschrichtius robustus) in the Mediterranean Sea: anomalous even or early sign of climate-driven distribution change? Marine Biodiversity Records 4, e28
Schusterman RJ, David Kastak D, Levenson DH, Reichmuth CJ, Southall BL (2000) Why pinnipeds don’t echolocate The Journal of the Acoustical Society of America 107, 2256 (2000); doi: 101121/1428506
Silva MA, Prieto R, Jonsen I, Baumgartner MF, Santos RS (2013) North Atlantic Blue and Fin Whales Suspend Their Spring Migration to Forage in Middle Latitudes: Building up Energy Reserves for the Journey? PLoS ONE 8(10): e76507 doi: 101371/journalpone 0076507
Simon M, Johnson M, Tyack P, Madsen PT (2009) Behaviour and kinematics of continuous ram filtration in bowhead whales (Balaena mysticetus) Proc R Soc B (2009) 276, 3819–3828 doi:101098/rspb20091135
Simon M, Stafford K, Beedholm K, Lee CM, Madsen PT (2010) Singing behavior of fin whales in the Davis Strait with implications for mating, migration and foraging The Journal of the Acoustical Society of America 128, 3200 (2010); doi: 101121/13495946 doiorg/101121/13495946
Sivle LD, Kvadsheim PH, Fahlman A, Lam FPA, Tyack PL, Miller PJO (2012) Changes in dive behavior during naval sonar exposure in killer whales, long-finned pilot whales, and sperm whales Frontiers in Physiology 3: 400 doi: 103389/fphys201200400
Sivle LD, Kvadsheim PH, Cure C, Isojunno S, Wensveen PJ, Lam FPA, Visser F, Kleivane L, Tyack PL, Harris CM, Miller PJO (2015) Severity of Expert-Identified Behavioural Responses of Humpback Whale, Minke Whale, and Northern Bottlenose Whale to Naval Sonar Aquatic Mammals 41:469-502 doi: 101578/am4142015469
Skaug HJ, Øien N, Schweder T, Bøthun G (2004) Abundance of minke whales (Balaenoptera acutorostrata) in the Northeast Atlantic: variability in time and space Canadian Journal of Fisheries and Aquatic Science, 61, 870–886 https://doiorg/101139/f04-020
Solvang HK, Skaug HJ, Øien NI (2015) Abundance estimates of common minke whales in the Northeast Atlantic based on survey data collected over the period 2008–2013 Paper SC/66a/RMP8 presented to the Scientific Committee of the International Whaling Commission Unpublished
Solvang H, Yanagihara H, Øien N, Haug T (2017) Temporal and geographical variation in body condition of common minke whales (Balaenoptera acutorostrata acutorostrata) in the Northeast Atlantic Polar Biol (2017) 40:667–68
Stanistreet JE, Nowacek DP, Baumann-Pickering S, Bell JT mfl. (2017) Using passive acoustic monitoring todocument the distribution of beaked whale species in thewestern North Atlantic Ocean. Can J Fish Aquat Sci 74: 2098−2109
Stevick PT, Allen J, Berube M, Clapham PJ, Katona SK, Larsen F, Lien J, Mattila DK, Palsboll PJ, Robbins J, Sigurjonsson J, Smith TD, Øien N, Hammond PS (2003) Segregation of migration by feeding ground origin in North Atlantic humpback whales (Megaptera novaeangliae). Journal of Zoology, 259, 231- 237. https://doi.org/10.1017/S0952836902003151
Stirling I (1973) Vocalization in the ringed seal (Phoca hispida). J. Fish. Res. Board.Can. 30: 1592-1594.
Storrie L, Lydersen C, Andersen M, Wynn RB, Kovacs KM (2018) Determining the species assemblage and habitat use of cetaceans in the Svalbard Archipelago, based on observations from 2002 to 2014. Polar Research 37.doi.org/10.1080/ 17518369.2018.1463065
Tartu S, Fisk A, Götsch A, Kovacs K, Lydersen C, Routti H (2020) First assessment of pollutant exposure in two balaenopterid whale populations sampled in the Svalbard Archipelago, Norway. Science of the Total Environment 718. 137327. https://doi.org/10.1016/j.scitotenv.2020.137327
Taylor BL, Baird R, Barlow J, Dawson SM, Ford J, Mead JG, Notarbartolo di Sciara G, Wade P, Pitman RL (2008) a Physeter macrocephalus. The IUCN Red List of Threatened Species 2008:e.T41755A10554884.
Taylor BL, Baird R, Barlow J, Dawson SM, Ford J, Mead JG, Notarbartolo di Sciara G, Wade P, Pitman RL (2008) b Hyperoodon ampullatus. The IUCN Red List of Threatened Species 2008: e.T10707A3208523.
Thomas PO, Reeves RR, Brownell RL (2016) Status of the world's baleen whales Mar. Mammal Sci., 32 , pp. 682-734, 10.1111/mms.12281
Vacquié-Garcia J, Lydersen C, Marques TA, Aars J, Ahonen H, Skern-Mauritzen M, Øien N, Kovacs KM (2017) a. Late summer distribution and abundance of ice-associated whales in the Norwegian High Arctic. Endangered Species Research 32: 59-70.
Vacquié-Garcia J, Lydersen C, Biuw M, Haug T, Fedak MA, Kovacs KM (2017) b Hooded seal Cystophora cristata foraging areas in the Northeast Atlantic Ocean Investigated using three complementary methods. PLoS ONE 12(12): e0187889. https://doi.org/10.1371/journal. pone.0187889
Van Parijs SM, Kovacs KM, Lydersen C (2001) Spatial and temporal distribution of vocalizing male bearded seals: implications for male mating strategies. Behaviour, 138, 905e922.
Van Parijs SM, Corkeron PJ, Harvey J, Hayes SA, Mellinger DK, Rouget PA, Thompson PM, Wahlberg M, Kovacs KM (2003) Patterns in the vocalisations of male harbor seals. Journal of the Acoustical Society of America, 113, 3403e3410
VanWormer E, Mazet J, Hall A, Gill V, Boveng P, London J mfl. (2019) Viral emergence in marine mammals in the North Pacific may be linked to Arctic sea ice reduction. Sci. Rep. 9:15569.
Víkingsson GA, Pike DG, Valdimarsson H, Schleimer A, Gunnlaugsson T, Silva T, ElvarssonB.Þ., Mikkelsen, B., Øien,N., Desportes,G, Bogason,V og Hammond,P.S.2015 Distribution, abundance, and feeding ecology of baleen whales in Icelandic waters: have recent environmental changes had an effect? Front. Ecol. Evol. 3:6. doi: 10.3389/fevo.2015.00006
Waring G, Josephson E, Maze-Foley K, Rosel, P (2016) US Atlantic and Gulf of Mexico Marine Mammal Stock Assessments – 2015. NOAA Technical Memorandum NMFS-NE 238.
Watkins WA, Daher MA, Fristrup KM, Howald TJ, Di Sciara GN (1993) Sperm Whales Tagged with Transponders and Tracked Underwater by Sonar. Marine Mammal Science. 1:55–67. https://doi.org/10. 1111/j.1748-7692.1993.tb00426.
Watt CA, Heide-Jørgensen MP, Ferguson SH (2013) How adaptable are narwhal? A comparison of foraging patterns among the world’s three narwhal populations. Ecosphere 4:71
Watt CA, Orr JR, Heide-Jørgensen MP, Nielsen NH, Ferguson SH (2015) Differences in dive behavior among the world’s three narwhal (Monodon monoceros) populations correspond with dietary differences. Marine Ecology Progress Series, 525, 273–285. https://doi. org/10.3354/meps1 1202
Wensveen P.J, Isojunno S, Hansen RR mfl.l (2019) Northern bottlenose whales in a pristine environment respond strongly to close and distant navy sonar signals.Proc. R. Soc. B 286: 20182592.http://dx.doi.org/10.1098/rspb.2018.2592
Wenzel FW, Broms F, López-Suárez P, Lopes K,Veiga N, Yeoman K, Simão M, Rodrigues D, Allen J, Fernald TW, Stevick PT, Jones L, Jann B, Bouveret C, Berrow S, Corkeron P (2020) Humpback Whales (Megaptera novaeangliae) in the Cape Verde Islands: Migratory Patterns, Resightings, and Abundance. Aquatic Mammals 46:21-31, DOI 10.1578/AM.46.1.2020.21
Windsland K, Lindstrøm U, Nilssen KT, Haug T (2007) Relative abundance and size composition of prey in the common minke whale diet in selected areas of the northeast Atlantic during 2000–04. Journal of Cetacean Research and. Management, 9, 167–178.
Wiig Ø, Bachmann L, Øien N, Kovacs K, Lydersen C (2010) Observations of bowhead whales (Balaena mysticetus) in the Svalbard area 1940–2009. Polar Biol. 33:979–984.DOI 10.1007/s00300-010-0776-1
Witting L (2019) Rebuttal against Desforges m.fl. 2018. No evidence for a global killer whale population collapse. Marine Mammal Science 35: 1197-1200.
Øien N (2009) Distribution and abundance of large whales in Norwegian and adjacent waters based on ship surveys 1995-2001. NAMMCO Sci. Publ. 7:31-47.
Øien N, Hartvedt S (2011) Northern bottlenose whales Hyperoodon ampullatus in Norwegian and adjacent waters. Paper presented to the IWC Scientific Committee, May 2011 SC/63/SM1. IWC, Cambridge
Øigård TA, Haug T, Nilssen KT (2014) Current status of hooded seals in the Greenland Sea.Victims of climate change and predation? Biol Conserv 172: 29–36
8.6 - Bentisk-Pelagisk kobling
Barthel KG (1990) Relationships of Food Uptake and Body Components of Calanus finmarchicus, C. glacialis and C. hyperboreus to Particulate Matter and Water Characteristics
in Fram Strait. Polar Biology 10:343-350
Bathmann UV, Noji TT, Voss M, Peinert R (1987) Copepod fecal pellets: abundance, sedimentation and content at a permanent station in the Norwegian Sea in May-June 1986. Marine Ecology Progress Series 38: 45-51.
Billett DSM, Lampitt RS, Rice AL, Mantoura RFC (1983) Seasonal sedimentation of phytoplankton to the deep-sea benthos. Nature 302: 520-522.
Drits AV, Klyuvitkin AA, Kravchishina MD. mfl. (2020) Fluxes of Sedimentary Material in the Lofoten Basin of the Norwegian Sea: Seasonal Dynamics and the Role of Zooplankton. Oceanology 60:501-517.
Graf G (1992). Benthic-pelagic coupling: a benthic view. Oceanography and Marine Biology: An Annual Review 30:149-190.
Graf G, Gerlach SA, Linke P, Queisser W, Ritzrau W, Scheltz A, Thomsen L, Witte U (1995) Benthic-pelagic coupling in the Greenland-Norwegian Sea and its effect on the geological record Geol Rundsch 84:49-58.
Griffiths JR, Kadin M, Nascimento F,…Winder M (2017)The importance of benthic-pelagic coupling for marine ecosystem functioning in a changing world. Global Change Biology 23(6) doi: 10.1111/gcb.13642
Hanz U, Roberts EM, Duineveld G, Davies A, van Haren H, Rapp HT, et al. (2021). Long-term observations reveal environmental conditions and food supply mechanisms at an Arctic deep-sea sponge ground. Journal of Geophysical Research: Oceans, 126, e2020JC016776. https://doi.org/10.1029/2020JC016776
Lampitt RS (1985) Evidence for the seasonal deposition of detritus to the deep-sea floor and its subsequent resuspension. Deep-Sea Research A 32: 885-897.
Meyer HK, Roberts EM, Rapp HT, Davies AJ (2019) Spatial patterns of arctic sponge ground fauna and demersal fish are detectable in autonomous underwater vehicle (AUV) imagery. Deep-Sea Research I 153: 103137.
Mueller CE, Larsson AI, Veuger B, Middelburg JJ, van Oevelen D (2014) Opportunistic feeding on various organic food sources bu the cold-water coral Lophelia pertusa. Biogeosciences 11:123-133.
Pile AJ, Young CM (2006) The natural diet of a hexactinellid sponge: Benthic–pelagic coupling in a deep-sea microbial food web. Deep-Sea Research I 53: 1148-1156.
Ramirez-Llodra E, Hilario A, Paulsen E, Costa CV, Bakken T, Johnsen G and Rapp HT (2020) Benthic Communities on the Mohn’s Treasure Mound: Implications for Management of Seabed Mining in the Arctic Mid-Ocean Ridge. Front. Mar. Sci. 7:490. doi: 10.3389/fmars.2020.00490
von Bodungen B, Antia A, Bauerfeind E, Haupt O, mfl. (1995) Pelagic processes and vertical flux of particles: an overview of a long-term comparative study in the Norwegian Sea and Greenland Sea. Geol Rundsch (1995) 84:11-27
Wassmann P, Peinert R, Smetacek V (1991) Patterns of production and sedimentation in the boreal and polar Northeast Atlantic, Polar Research, 10:209-228, DOI: 10.3402/polar.v10i1.6740
9 - Vedlegg
Tabell 3.1 Oversikt over statistiske parametre til strømmålingene av daglige verdier (unntatt der det er spesifisert) i utvalgte dyp. Tabellen viser instrumenttype, måledyp, middelfart og standardavvik, middelhastighet og stabilitet, definert som middelhastighet dividert på middelfart, maksimum hastighet (fart og retning), halvdaglig tidevannskomponent M2, maksimum fart fra timeverdier. Fra Mork mfl., 2012. Posisjonene til riggene JM-1 og JM-2 er vist i Figur 6 mens riggen Lyngvi ligger i Islandshavet utenfor utredningsområdet.